
Diffusion of the Nearctic leafhopper Scaphoideus titanus Ball in Europe: a consequence of human...
11
ORIGINAL RESEARCH Diffusion of the Nearctic leafhopper Scaphoideus titanus Ball in Europe: a consequence of human trading activity Sabrina Bertin Carmela R. Guglielmino Nisrine Karam Ludvik M. Gomulski Anna R. Malacrida Giuliano Gasperi Received: 21 July 2006 / Accepted: 22 December 2006 / Published online: 23 January 2007 ȑ Springer Science+Business Media B.V. 2007 Abstract Scaphoideus titanus Ball is a Nearctic leaf- hopper that was introduced for the first time in Europe probably at the beginning of the 20th century. In Europe, this species is a specialist on cultivated grapevines and is of great economic importance as the vector of Flavescence dore ´ e (FD), a Grapevine Yel- lows disease caused by Candidatus Phytoplasma vitis. The Random Amplified Polymorphic DNA (RAPD) technique was employed to obtain genetic information about the diffusion and the structure of S. titanus populations. Two American and 14 European popula- tions were analysed. A total of 188 reproducible bands, obtained from three arbitrary primers, were considered to assess the amount and the pattern of genetic varia- tion within and among leafhopper populations. American populations showed high levels of intra- population polymorphism and dissimilarity and appeared to be the most isolated of all the tested samples. The results confirm the historical role of American samples as the sources for the more recently founded European populations. RAPD analyses revealed a weak genetic structure of European samples that could probably be explained invoking the human role in their diffusion. The non-natural spreading of S. titanus across Europe is in fact attributable to the exchange of grapevine canes and grafts carrying eggs that the insect laid under the bark to overwinter. Keywords Diffusion Á Genetic relatedness Á Grapevine Á Polymorphism Á RAPD-PCR Á Scaphoideus titanus Introduction Scaphoideus titanus Ball (Schvester et al. 1962, 1969) is a Nearctic leafhopper belonging to the family Cicad- ellidae, subfamily Deltocephalinae. It is a hemime- tabolous and phloem-feeding insect, which is of great economic importance as vector of Flavescence dore ´e (FD), a persistent disease of cultivated grapevines. The FD, considered the most threatening among Grapevine Yellows (GY) diseases in Europe (Boudon-Padieu 2003), is caused by Candidatus Phytoplasma vitis (16Sr-V group) (Firrao et al. 2005), which is a phloem- restricted bacterium. In Europe, S. titanus is a specialist for cultivated grapes (Vitis vinifera) (Vidano 1964), while in the ori- ginal area it is also widespread on other Vitis spp. and highly abundant on wild Vitis riparia (Vidano 1966; Maixner et al. 1993). This leafhopper is univoltine: the eggs are laid under the bark of 2-year-old wood, where they overwinter (Vidano 1964). In the original area, the species was found in the USA and South Canada (Barnett 1976) and it was probably introduced into Europe as eggs, uninten- tionally transported on imported grapevine canes from North America (Boudon-Padieu 1999). Scaphoideus titanus was reported for the first time in 1958 in vine- S. Bertin Á N. Karam Á L. M. Gomulski Á A. R. Malacrida Á G. Gasperi (&) Dipartimento di Biologia Animale, Universita ` di Pavia, Piazza Botta 9, 27100 Pavia, Italy e-mail: [email protected] C. R. Guglielmino Dipartimento di Genetica e Microbiologia, Universita ` di Pavia, Via Ferrata 1, 27100 Pavia, Italy 123 Genetica (2007) 131:275–285 DOI 10.1007/s10709-006-9137-y
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Diffusion of the Nearctic leafhopper Scaphoideus titanus Ball in Europe: a consequence of human...

ORIGINAL RESEARCH
Diffusion of the Nearctic leafhopper Scaphoideus titanus Ballin Europe: a consequence of human trading activity
Sabrina Bertin Æ Carmela R. Guglielmino ÆNisrine Karam Æ Ludvik M. Gomulski ÆAnna R. Malacrida Æ Giuliano Gasperi
Received: 21 July 2006 / Accepted: 22 December 2006 / Published online: 23 January 2007� Springer Science+Business Media B.V. 2007
Abstract Scaphoideus titanus Ball is a Nearctic leaf-
hopper that was introduced for the first time in Europe
probably at the beginning of the 20th century. In
Europe, this species is a specialist on cultivated
grapevines and is of great economic importance as the
vector of Flavescence doree (FD), a Grapevine Yel-
lows disease caused by Candidatus Phytoplasma vitis.
The Random Amplified Polymorphic DNA (RAPD)
technique was employed to obtain genetic information
about the diffusion and the structure of S. titanus
populations. Two American and 14 European popula-
tions were analysed. A total of 188 reproducible bands,
obtained from three arbitrary primers, were considered
to assess the amount and the pattern of genetic varia-
tion within and among leafhopper populations.
American populations showed high levels of intra-
population polymorphism and dissimilarity and
appeared to be the most isolated of all the tested
samples. The results confirm the historical role of
American samples as the sources for the more recently
founded European populations. RAPD analyses
revealed a weak genetic structure of European samples
that could probably be explained invoking the human
role in their diffusion. The non-natural spreading of
S. titanus across Europe is in fact attributable to the
exchange of grapevine canes and grafts carrying eggs
that the insect laid under the bark to overwinter.
Keywords Diffusion � Genetic relatedness �Grapevine � Polymorphism � RAPD-PCR �Scaphoideus titanus
Introduction
Scaphoideus titanus Ball (Schvester et al. 1962, 1969) is
a Nearctic leafhopper belonging to the family Cicad-
ellidae, subfamily Deltocephalinae. It is a hemime-
tabolous and phloem-feeding insect, which is of great
economic importance as vector of Flavescence doree
(FD), a persistent disease of cultivated grapevines. The
FD, considered the most threatening among Grapevine
Yellows (GY) diseases in Europe (Boudon-Padieu
2003), is caused by Candidatus Phytoplasma vitis
(16Sr-V group) (Firrao et al. 2005), which is a phloem-
restricted bacterium.
In Europe, S. titanus is a specialist for cultivated
grapes (Vitis vinifera) (Vidano 1964), while in the ori-
ginal area it is also widespread on other Vitis spp. and
highly abundant on wild Vitis riparia (Vidano 1966;
Maixner et al. 1993). This leafhopper is univoltine: the
eggs are laid under the bark of 2-year-old wood, where
they overwinter (Vidano 1964).
In the original area, the species was found in the
USA and South Canada (Barnett 1976) and it was
probably introduced into Europe as eggs, uninten-
tionally transported on imported grapevine canes from
North America (Boudon-Padieu 1999). Scaphoideus
titanus was reported for the first time in 1958 in vine-
S. Bertin � N. Karam � L. M. Gomulski �A. R. Malacrida � G. Gasperi (&)Dipartimento di Biologia Animale, Universita di Pavia,Piazza Botta 9, 27100 Pavia, Italye-mail: [email protected]
C. R. GuglielminoDipartimento di Genetica e Microbiologia, Universita diPavia, Via Ferrata 1, 27100 Pavia, Italy
123
Genetica (2007) 131:275–285
DOI 10.1007/s10709-006-9137-y

yards in South France (Bonfils and Schvester 1960) and
in 1960s it diffused towards Italy (1963 in Liguria,
Vidano 1964) and Western and Southern Switzerland
(Baggiolini et al. 1968; Clerc et al. 1997). In Italy it has
spread to almost every northern region where grape-
vines are cultivated (Alma 2002). In 1987 S. titanus was
found in Croatia and Slovenia (Gabrijel 1987; Seljak
2002) and in 2003 in Serbia (Duduk et al. 2003). More
recently the species was detected in 1995 in Spain
(Lavina et al. 1995; Batlle et al. 1997) and in 1999 in
North Portugal (Quartau et al. 2001). In most of these
areas FD became an epidemic disease and spread as
the leafhopper dispersed.
As vector of FD, S. titanus was the target of studies
to analyse the modality of phytoplasma acquisition and
transmission (Schvester et al. 1969; Alma et al. 1997;
Bressan et al. 2005a, 2006). The unique sources for
vector acquisition in the field appear to be FD-infected
grapevines, since no other source plants have been
identified for the natural acquisition of FD phytoplas-
ma by S. titanus (Bressan et al. 2006). The leafhopper
vector is involved in vine-to-vine phytoplasma trans-
mission and the incidence of FD disease may increase
exponentially over successive years (Bressan et al.
2006). All developmental feeding stages of S. titanus
may acquire FD phytoplasma, although a long latent
period associated with the multiplication and persis-
tence of the phytoplasma in the vector’s body, and
flight ability, make adults the most important life stage
involved in the spread of FD (Bressan et al. 2006).
Acquisition efficiency of FD phytoplasma by the leaf-
hopper vector is influenced by factors such as the sus-
ceptibility of grapevine cultivars, the developmental
nymph instar and probably the period of growing
season (Bressan et al. 2005b).
Vector movement and dispersal represent important
components of incidence and spread of phytoplasma
disease (Hoy et al. 1992; McClure 1980). The inability
of the leafhopper to disperse over long distances has
been documented (Lessio and Alma 2004, 2006): the
adults do not spread significantly outside the vineyard
and their movements within the vineyard appear to be
strongly influenced by the planting layout.
There is no question that human activities have
played the major role in the long-distance dispersal and
in introducing this vector into previously unoccupied
areas (Weintraub and Beanland 2006). However the
frequency and the patterns of long-range movements is
largely unknown. DNA markers may provide effective
tools to infer movements within and between leaf-
hopper populations and to identify possible sources of
introduction.
Given the dearth of molecular data for S. titanus, we
have used the random amplified polymorphic DNA
technique (RAPD, Williams et al. 1990; Welsh and
McClelland 1990), which amplifies DNA fragments
from any genome, to derive suitable genetic markers
for S. titanus. The obvious advantage of the RAPD
approach is the relatively easy assay of a large number
of marker loci across the entire genome (Lynch and
Milligan 1994). The limitations associated with RAPD
markers include variable reproducibility unless reac-
tion conditions are stringently controlled (Baruffi et al.
1995; Sebastiani et al. 2001), and a dominant mode of
inheritance (Black 1993). The dominant nature of
RAPDs precludes the direct estimation of allele fre-
quencies and can bias calculations of genetic diversity
and population differentiation (Lynch and Millingan
1994). However, different estimates for measuring
genetic similarity and/or dissimilarity have been
developed (Wang 2004; Ritland 2005; Kosman and
Leonard 2005). These estimates provide a rough eval-
uation of the genetic similarity at the different levels of
population hierarchy and are relevant as descriptive
tools.
This study regards 16 populations of S. titanus from
North America and from Europe and its principal aims
were to (i) assess the level of genetic variation of the
species; (ii) evaluate the level of population differen-
tiation and structuring; (iii) determine if there is a
relationship between the level of genetic variation and
the spread of the species during the last 50 years.
Materials and methods
Sample collection
Fifteen out of the 16 samples of S. titanus were field-
caught as adults during the summer of 2001 from the
following locations: 2 in USA (Valois and Geneva
Ontario, NY State), 1 in France (Ponteves), 6 in Italy
(Chatillon, Chambave, San Colombano, Corniglia,
Vignale, Treiso), 2 in Switzerland (Piano Magadino
and Castelrotto), 3 in Slovenia (Goriska Brda, Izola,
Nova Gorica), and 1 in Spain (Penedes). An additional
sample from France was obtained from eggs collected
in Moroges during the winter of 2005. The adults hat-
ched from these eggs were obtained as ethanol pre-
served specimens (Table 1; Fig. 1).
Also all the other samples were obtained as ethanol
preserved specimens and DNA was extracted from
each single individual of both sexes, according to the
method of Baruffi et al. (1995).
276 Genetica (2007) 131:275–285
123

RAPD-PCR amplification
DNA from 15–18 individuals per population was
amplified using three oligonucleotide random primers
of 24–26 bp in length (Baruffi et al. 1995; Sebastiani
et al. 2001)
494N (5¢-AGCGCTGTGAGAAAGATGAAA-
GAT-3¢);
D17 (5¢-CGAAACAAGCGTGCATGAGCCCG-
AA-3¢);
NP4 (5¢-CTAATGCAGGAGTCGCATAAGG-
GAGA-3¢).
Primer selection was based on the clarity and con-
sistency of the resulting banding patterns.
Amplification reactions were performed in a final
volume of 10 ll, containing 10 mM Tris–HCl (pH 8.3),
50 mM KCl, 2 mM MgCl2, 200 lM dNTPs, 30 lM
primer, 0.1% gelatin, 3–7 ng of template DNA and 0.2
units of Taq DNA polymerase (Roche).
Amplifications were performed using a program-
mable thermocycler (PTC-100 MJ-Research Inc.) using
the following protocol: an initial denaturing step at
94�C for 90 s and 45 cycles of denaturation at 94�C for
30 s, annealing at 45�C for 1 min, extension at 75�C
for 2 min. The samples were finally incubated at 75�C
for 10 min and 60�C for 10 min and stored at 4�C.
Aliquots of 5 ll of the PCR products were loaded
together with a DNA size marker (50-bp ladder,
Amersham) onto a 2.5% (w/v) agarose TBE gel (1%
SeaKem GTG Agarose plus 1.5% NuSieve GTG
Agarose). Electrophoresis was carried out at 5.5 V/cm
for 4 h; the gels were stained using 0.5 lg/ml (w/v) of
ethidium bromide and the bands were visualised using
UV light.
Duplicate PCR reactions on each individual tem-
plate DNA were performed to ensure the reproduc-
ibility of the individual DNA bands. The
reproducibility of this method has already been tested
in other organisms (Baruffi et al. 1995; Sebastiani et al.
2001). The faintest bands and bands that differed in
replicates were not considered; several bands were not
taken into account because they could not be distin-
guished clearly from neighbouring bands. A negative
control without genomic DNA was included in each
series of PCR amplifications to ensure that there was
no contamination.
Detection of Flavescence doree phytoplasma
in vector populations
The DNAs from the same individuals used for RAPD-
PCR analysis were also checked for the possible pres-
ence of Flavescence doree phytoplasma DNA (Can-
didatus Phytoplasma Vitis 16Sr-V). Diagnostic PCRs
were performed using the following primers, that were
designed on the partial sequence of Flavescence doree
phytoplasma strain FD70 16S rRNA gene (Accession
Number AY197643; Lee et al. 2004):
FDf141 forward (5¢-GGAAACGGTTGCTAA-
GACTGGATA-3¢);
Table 1 Field collected samples of Scaphoideus titanus
Country Location Coordinates
USA (NY state) Valois 42�32¢N 76�52¢EGeneva Ontario 42�86¢N 76�98¢E
France Moroges 46�44¢N 04�40¢EPonteves 43�33¢N 06�01¢E
Italy Chatillon 45�44¢N 07�36¢EChambave 45�44¢N 07�32¢ESan Colombano 45�10¢N 09�29¢ECorniglia 44�07¢N 09�42¢EVignale 44�54¢N 08�24¢ETreiso 44�41¢N 08�05¢E
Switzerland Piano Magadino 46�08¢N 08�51¢ECastelrotto 45�59¢N 08�50¢E
Slovenia Goriska Brda 46�03¢N 13�30¢EIsola 45�32¢N 13�39¢ENova Gorica 45�56¢N 13�39¢E
Spain Penedes 41�20¢N 01�41¢E
12 - Castelrotto
U.S.A 1 - Valois
3 - Moroges4 - Ponteves
France
Italy
5 - Chatillon6 - Chambave
9 - Vignale8 - Corniglia7 - San Colombano
10 - Treiso
Switzerland
Slovenia
16 - PenedesSpain
(New York State)
14
11 - Piano Magadino
13 - Goriska Brda14 - Izola15 - Nova Gorica
810 95 6 7
1112
13153
4
New York State
1
2
16
2 - Geneva OntarioFig. 1 Geographic locationsof the sampling sites ofScaphoideus titanus analyzedin this study
Genetica (2007) 131:275–285 277
123

FDr1391 reverse (5¢-TCGGGTATTGCTAAC
TTTCGTGGT-3¢).
This primer pair, which yields a product of 1250 bp,
was used in direct PCR assays. Reaction products were
diluted 1:100 with sterile water and used as templates
in nested PCR, driven by R16(V)F1/R1 primer pair
that is specific for group V phytoplasmas (Lee et al.
1994; amplicon size 1100 bp). DNA amplifications
were performed in a total reaction volume of 25 ll; the
reaction mixture contained as template 1 ll of
extracted DNA or the diluted first amplification
product, 20 mM Tris–HCl (pH 8.0), 50 mM KCl,
1.5 mM MgCl2, 200 lM dNTPs, 1 lM of each primer
and 1 unit of Taq DNA polymerase (Invitrogen). The
conditions for amplification consisted of a pre-dena-
turation step at 94�C for 120 s, 35 cycles of denatur-
ation at 94�C for 60 s, annealing at 63�C in direct or
50�C in nested PCR for 90 s, extension at 72�C for
120 s and a final extension step at 72�C for 10 min.
A positive control (DNA extracted from an indi-
vidual of S. titanus known to be infected by FD) and a
negative control (without DNA) were included in each
PCR reaction.
Amplification products were analysed by electro-
phoresis on a 1% (w/v) agarose TAE gel stained using
0.5 lg/ml (w/v) of ethidium bromide and visualised
under UV light.
RAPD data analysis
For statistical analysis of RAPD data, the amount
and distribution of variability of the RAPD profiles
obtained from 283 individuals using the three primers
(NP4, D17, 494N) were considered. Under the
assumption that variation in banding patterns repre-
sents allelic segregation at homologous and inde-
pendent dominant loci, each locus was treated as a
two-allele system corresponding to the presence/ab-
sence of the amplified band. At each locus, the plus
allele (presence of a band) dominates over the null
allele (unamplified with PCR); therefore individuals
with a given amplified band are homozygous or het-
erozygous for a dominant allele whereas individuals
without that amplified band are homozygous reces-
sive. Each leafhopper was scored at each locus as 1
(present) or 0 (absent) across all polymorphic loci to
create a binary matrix. A locus was considered
polymorphic if the frequency of the most common
allele did not exceed 0.99. Bands with the same
molecular weight were assumed to represent homol-
ogous loci.
The presence/absence data were used to obtain
estimates of genetic relatedness between pairs of
individuals. Two estimates of pairwise relatedness have
been considered: ‘‘Dissimilarity index’’ (D), evaluated
according with Nei and Li (1985), and the ‘‘Similarity
estimator’’ (SM, Wang 2004). SM estimator has been
found to have consistently a small mean squared
deviation (MSD) over wide types of data (number of
loci, allele frequencies, sample size) (Wang 2004). The
two indices, calculated by the RAPDPLOT program
(Black 1995) and the MER (V3) program (Wang
2004), are respectively based on phenotypic data
(D) and on estimated allele frequencies (SM).
The presence/absence data were directly subjected
to analysis of molecular variance (AMOVA; Excoffier
et al. 1992), in ARLEQUIN Ver. 2.000 software
(Schneider et al. 2000), in order to describe the distri-
bution of genetic variability in the populations hierar-
chy. The significance of each variance component
was tested using a non-parametric permutation test
(Excoffier et al. 1992), consisting of 1023 random
permutations.
Allele frequencies were estimated using the
RAPDBIOS 2.0 program (Black 1995), assuming that
genotypes are in Hardy–Weinberg equilibrium and
applying the Lynch and Milligan (1994) correction.
On the basis of the derived gene frequencies, the
following analyses were performed. The degree of
polymorphism detected by each primer for the total
sampled leafhoppers were calculated using the pro-
gram BIOSYS-2 (modified from Swofford and
Selander 1981). Cavalli-Sforza and Edwards’s (1967)
Chord measures (DCE) were calculated between pairs
of populations, using the RAPDDIST program
(Black 1995). As this distance measure is correlated
to the pairwise fixation index (FST) (Cavalli-Sforza
et al. 1994), the effect of geographical distance on
genetic differentiation of populations was tested: we
regressed DCE/(1 – DCE) between pairs of popula-
tions to the natural logarithm (ln) of the geographical
distance between the pairs (Rousset 1997). The sig-
nificance of this relationship was assessed using the
Mantel test (Mantel 1967) with 100,000 random per-
mutations, in the ISOLDE routine within GENEPOP
3.4 program (Raymond and Rousset 1995). The
RAPDDIST program was also used to estimate the
Nei’s (1978) unbiased genetic distances among pop-
ulations. A consensus tree was derived from 1000
bootstrap resamplings (PHYLIP 3.57C package; Fel-
senstein 1995). A principal coordinate analysis
(PCoA; Gower 1966) was performed on the allele
frequencies to visualize the geometric relationship
among leafhopper populations in a two-dimensional
graph. PCoA analysis was conducted using the
NTSYS-PC program (Rohlf 1993).
278 Genetica (2007) 131:275–285
123

Results
RAPD patterns
The amplification of 283 individual DNAs from 16
populations with the three arbitrary primers (494N,
D17, NP4) resulted in a total of 188 interpretable and
reproducible bands. The RAPD patterns obtained
from the amplification of male and female genomic
DNA did not show differences. As assessed in all the
leafhopper individuals assayed, the fragment sizes
varied from 120 to 1300 bp. An average of 62.67 ± 4.04
(SD) bands were amplified per primer. The percentage
of polymorphism detected by each primer ranged be-
tween 33.33 ± 11.35 for NP4 to 44.19 ± 5.13 for 494N.
Out of the 188 bands, only four (NP4 – 345 bp, NP4
– 610 bp, D17 – 660 bp, 494N – 430 bp) were mono-
morphic over all individuals. Moreover, the 494N
– 410 bp band appears to be specific for the two Amer-
ican populations (Geneva Ontario and Valois) (Fig. 2).
Detection of Flavescence doree phytoplasma
in vector populations
Three individuals, all Italian, of S. titanus out of the 283
tested (1.06%) for the 16 populations, resulted FD
phytoplasma infected (2 individuals from Vignale and
1 individual from San Colombano populations). Com-
paring RAPD patterns of these three individuals with
RAPD patterns of the remaining individuals, no bands
discriminating between FD positive and non-positive
leafhoppers was observed. Therefore the presence of
FD DNA did not affect the resulting RAPD patterns
and the subsequent population analysis of S. titanus.
RAPD variability within populations
As shown in Table 2, considering all three primers,
a similar mean number of fragments per individual was
scored within each population (xi), with an average
value of 22.03 ± 0.916.
High intra-population heterogeneity levels for
RAPD variation were observed for all the geographic
samples. An average of 93.00 ± 3.374% of fragments
scored for each population were polymorphic. The two
estimators of intra-population differentiation, i.e. the
dissimilarity index D and the similarity index
SM, which are negatively correlated (non-parametric
Wilcoxon’s Signed-Ranks Test: P < 0.001), testify a
high level of variability.
All the polymorphism and differentiation estimates
indicate that the American populations, Valois and
Geneva Ontario are the most polymorphic. The
European populations were rather heterogeneous
within each country, both in terms of polymorphism
and intra-population differentiation.
Population structure of S. titanus
Phenotypic AMOVA analysis was performed in order
to test the significance of the partitioning of genetic
variance between the group of American populations
and the group including the fourteen European popu-
lations (Table 3A). A significant 12.77% of total vari-
ance was attributable to differences between the two
groups. When only the European populations were
considered (Table 3B), the value of the variance
component among the groups, corresponding to the
five European countries, was very low (1.86%) and not
significant (P = 0.165). In both analyses most of the
variance was significantly attributable to within popu-
lation variability (71.99 and 81.30%, respectively).
Pairwise Cavalli-Sforza and Edwards’s genetic dis-
tances (DCE, 1967) were derived in order to estimate
the genetic relationships between the American and
European populations and within and among Euro-
pean countries. In general (Table 4) no very high DCE
estimates were observed in all the comparisons (from
0.055 to 0.143). As expected, the largest DCE values
occurred with all the comparisons involving the two
American populations, especially with French (DCE
range: 0.125–0.143) and Spanish samples (DCE range:
0.137–0.142). In Europe, low values of differentiation
may be found in the comparisons between countries,
whilst higher or comparable values may be observed
within each country. For example, between Italian and
Slovenian samples DCE values ranged from 0.055 to
0.106, while DCE values vary from 0.068 to 0.109 within
Fig. 2 494N RAPD patterns in individuals from two populationsof S. titanus: San Colombano (Italy) and Valois (USA). A specificfragment (494N – 410 bp) is present only in American speci-mens; a monomorphic fragment (494N – 430 bp) is present inboth populations. M = 50 bp ladder
Genetica (2007) 131:275–285 279
123

Ta
ble
2R
AP
Dp
oly
mo
rph
ism
an
dd
iffe
ren
tia
tio
ne
stim
ate
sin
wil
dp
op
ula
tio
ns
of
Sca
ph
oid
eus
tita
nu
s
Po
pu
lati
on
sN
um
be
ro
fin
div
idu
als
Me
an
no
.o
ffr
ag
me
nts
sco
red
pe
rin
div
idu
al
(xi)
To
tal
no
.o
ffr
ag
me
nts
sco
red
pe
rp
op
ula
tio
n(x
p)*
Pe
rce
nta
ge
of
po
lym
orp
hic
ba
nd
s(P
)**
Dis
sim
ila
rity
ind
ex
(D±
SD
)
Sim
ila
rity
est
ima
tor
(SM
±S
D)
US
AV
alo
is1
82
1.6
1±
1.2
95
10
45
4.7
9(9
9.0
4)
0.6
2±
0.1
78
-0.1
1±
0.1
77
Ge
ne
va
On
tari
o1
82
1.2
2±
1.1
34
95
50
.00
(98
.95
)0
.64
±0
.18
00
.06
±0
.15
9F
ran
ceM
oro
ge
s1
82
3.2
2±
1.7
34
71
32
.45
(85
.91
)0
.32
±0
.15
90
.31
±0
.15
8P
on
tev
es
18
21
.56
±1
.30
27
63
7.2
4(9
2.1
2)
0.3
5±
0.1
14
0.2
6±
0.1
66
Ita
lyC
ha
till
on
18
22
.61
±1
.25
47
33
6.1
7(9
3.1
5)
0.4
1±
0.1
25
0.1
6±
0.2
01
Ch
am
ba
ve
16
22
.56
±1
.28
67
73
7.7
7(9
2.2
1)
0.3
8±
0.1
18
0.1
9±
0.1
46
Sa
nC
olo
mb
an
o1
82
2.8
9±
1.1
45
81
40
.43
(93
.83
)0
.43
±0
.14
40
.10
±0
.18
3C
orn
igli
a1
82
0.2
7±
1.2
65
81
39
.89
(92
.59
)0
.41
±0
.12
30
.24
±0
.16
4V
ign
ale
18
21
.83
±1
.18
97
23
5.6
4(9
3.0
6)
0.3
7±
0.1
13
0.2
3±
0.1
29
Tre
iso
18
21
.11
±1
.20
47
73
8.8
3(9
4.8
1)
0.4
3±
0.1
29
0.1
6±
0.1
61
Sw
itze
rla
nd
Pia
no
Ma
ga
din
o1
52
3.0
6±
1.2
65
89
45
.21
(95
.51
)0
.44
±0
.14
80
.03
±0
.15
2C
ast
elr
ott
o1
82
3.1
1±
1.2
45
65
31
.38
(90
.77
)0
.32
±0
.12
80
.32
±0
.21
5S
lov
en
iaG
ori
ska
Brd
a1
82
2.5
6±
1.1
79
88
44
.15
(94
.32
)0
.43
±0
.13
30
.10
±0
.19
0Iz
ola
18
21
.17
±1
.23
36
93
2.9
8(8
9.8
6)
0.3
5±
0.1
05
0.2
8±
0.1
30
No
va
Go
rica
18
21
.00
±1
.19
87
53
7.2
3(9
3.3
3)
0.4
2±
0.1
33
0.1
9±
0.1
72
Sp
ain
Pe
ne
de
s1
82
2.7
2±
1.2
24
61
28
.72
(88
.52
)0
.34
±0
.11
10
.29
±0
.16
0
Intr
asp
ecifi
ca
ver
ag
e±
SD
17
.69
±0
.87
32
2.0
3±
0.9
16
78
.37
±1
1.1
53
38
.93
±6
.87
1(9
3.0
0±
3.3
74
)0
.42
±0
.09
30
.17
±0
.11
7
*O
ve
rth
e1
88
fra
gm
en
tssc
ore
da
cro
ssth
ep
op
ula
tio
ns
**
Po
lym
orp
his
mv
alu
es
ov
er
the
18
8fr
ag
me
nts
sco
red
acr
oss
the
po
pu
lati
on
s;in
pa
ren
the
sis
po
lym
orp
his
mv
alu
es
rela
ted
toth
efr
ag
me
nts
sco
red
for
ea
chp
op
ula
tio
n
280 Genetica (2007) 131:275–285
123

Italian group and from 0.066 to 0.092 within Slovenian
group. Between Slovenian and Swiss samples DCE
values ranged from 0.059 to 0.109, while the value
between the two Swiss populations is 0.096.
It is clear from these data that there is absence of
isolation by distance in Europe. In fact, the correlation
between the matrix of DCE/(1 – DCE) and the natural
logarithm of the pairwise geographical distances is
significant only if American populations are included
(Spearman Rank correlation coefficient r = 0.011;
Mantel P = 0.005).
A good description of the complex pattern of pop-
ulation relationships is shown in the Neighbour Joining
(NJ) consensus tree derived from Nei’s genetic dis-
tances (Fig. 3) and in the Principal Component Anal-
ysis (PCoA) (Fig. 4), both performed on RAPD-
derived allelic frequency data. In the NJ consensus
tree, obtained from 1000 bootstrap resamplings of the
data set, after the first statistically significant separation
of the two American populations, the only one division
supported by more than 50% of the resamplings sep-
arates the Italian sample of Vignale from the Spanish
sample of Penedes. The other branches separate a
heterogeneous mix of European samples.
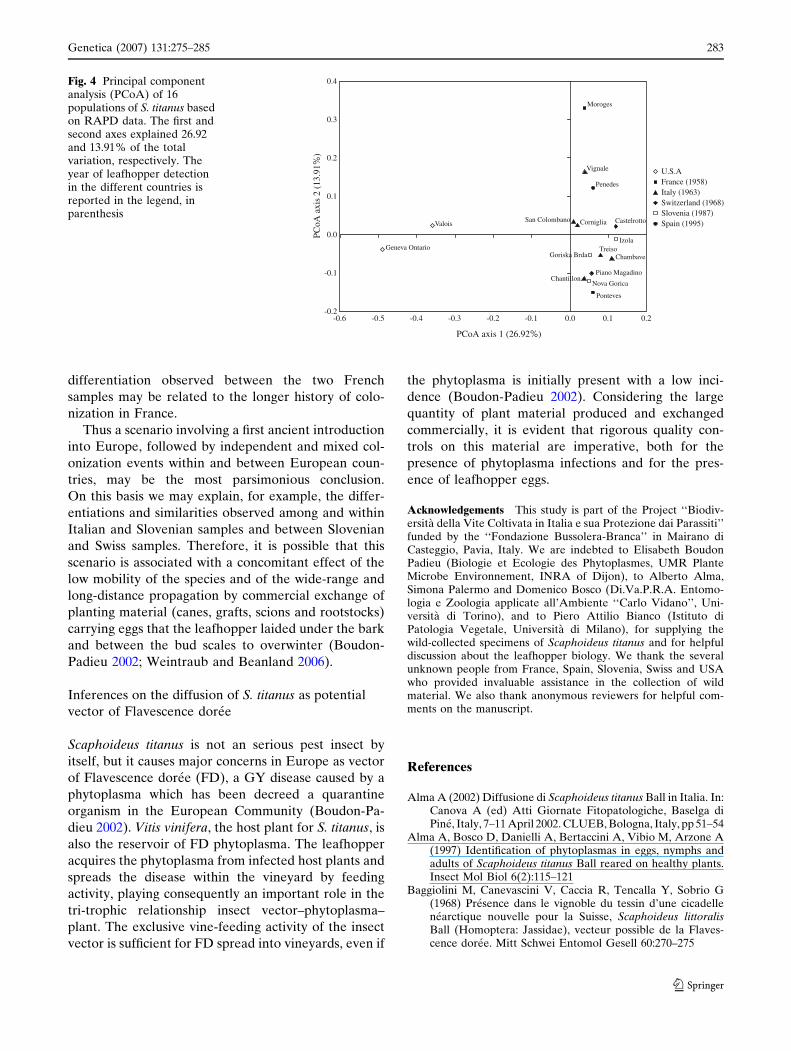
The PCoA plot (Fig. 4) confirms the major separation
of American from European samples along the first PCo
axis (27% of variation). The European populations are
scattered along the second PCo axis (14% of variation)
without any apparent connection with the country of
origin and the historical chronology of S. titanus invasion.
Along this axis the two French populations from Moroges
and Ponteves have the extreme values.
Discussion
We have used RAPD markers as a descriptive tool for
inferences about DNA variability and population
structure of the leafhopper S. titanus, a species for
which little genetic knowledge is available.
The degree of polymorphism revealed by these
markers is extensive: more than 85% of fragments
scored per population are polymorphic, demonstrating
that there is a vast reservoir of hidden nuclear DNA
polymorphism in this species. On the basis of this
variability, the RAPD technique appears to be ade-
quate to reveal the level of intra- and inter-population
divergence in order to provide insights into the pattern
of leafhopper colonization.
Genetic structure and inferences on dispersal
of S. titanus
Among the North-American genus Scaphoideus, S. tit-
anus is so far the only species which has become
established in Europe, probably through the introduc-
tion of planting material initially in South-Western
France (Boudon-Padieu 2002). In agreement with the
Nearctic origin of the species, the two American pop-
ulations analysed in this study result clearly differen-
tiated from the European populations and were shown
to possess local private alleles and the highest levels of
intra-population genetic variability. In its home range
S. titanus has been found to be associated not only with
V. vinifera but also with different native Vitis species
(Vidano 1966; Maixner et al. 1993). This capacity to
adapt to different host plants may increase the dis-
persion of the species, favouring the maintenance of
high population size and high genetic variability (Nei
et al. 1975). The wide reservoir of genetic variation in
the leafhopper native populations may constitute the
background of adaptive processes during colonization.
Upon its arrival in Europe, this species settled pro-
gressively since the 1960’s in several countries from
West to East, into a climatic area with cold winter and
relatively long summer. In these areas, S. titanus
Table 3 Analysis of molecular variance (AMOVA) among Scaphoideus titanus samples
Source of variation d.f. SSD Variance components % Variation P-value
(A) between European and American samplesEuropean versus American 1 153.03 1.70 12.77 < 0.008Among populations within the two groups 14 636.52 2.03 15.24 < 0.001Within populations 267 2562.00 9.59 71.99 < 0.001Total 282 3351.55 13.33
(B) among five groups of European populations: France, Italy, Switzerland, Slovenia and SpainAmong five European countries 4 210.92 0.21 1.86 = 0.165Among populations within countries 9 388.68 1.92 16.84 < 0.001Within populations 233 2163.17 9.28 81.30 < 0.001Total 246 2762.77 11.42
d.f., degrees of freedom; SSD, sum of squared deviation; % of variation, percentage of total variance contributed by each component;P-value, probability value obtained by significant test (1023 permutations).
Genetica (2007) 131:275–285 281
123

appears to be well adapted to V. vinifera and it seems
that its biological cycle could adjust to all climates
compatible with vineyard cultivation (Boudon-Padieu
2002).
A question arises about the modality of diffusion in
Europe of this species, which is characterized by a very
restricted range of mobility. It is known that S. titanus
does not disperse significantly beyond 24 m from the
vineyard, suggesting that the leafhopper can colonise
by its own means only nearby vineyards (Lessio and
Alma 2004, 2006). This characteristic, together with its
univoltine reproductive behaviour, would suggest
a population structure reflecting the geographical
distance and/or the chronology of the first records of
the species in Europe. On the contrary, our RAPD
data show an almost complete lack of isolation by
distance from France (which probably was the first
colonized country in Europe) to Slovenia and Spain
(which are the most recent colonized among our con-
sidered samples), reflecting a complex genetic frag-
mentation among populations, both within and
between countries. It seems likely that such scattered
pattern in the distribution of genetic diversity derives
from independent introduction events of S. titanus
within and between the different European areas. TheTa
ble
4P
air
wis
eC
av
all
i-S
forz
aa
nd
Ed
wa
rds’
s(1
96
7)
Ch
ord
me
asu
res
(DC
E)
am
on
gth
e1
6p
op
ula
tio
ns
of
Sca
ph
oid
eus
tita
nu
s
12
34
56
78
91
01
11
21
31
41
51
6
US
AV
alo
is1
–G
en
ev
aO
nta
rio
20
.09
6–
Fra
nce
Mo
rog
es
30
.13
20
.12
5–
Po
nte
ve
s4
0.1
43
0.1
34
0.0
92
–It
aly
Ch
ati
llo
n5
0.1
40
0.1
27
0.0
95
0.0
73
–C
ha
mb
av
e6
0.1
28
0.1
00
0.1
11
0.1
15
0.0
95
–S
an
Co
lom
ba
no
70
.12
60
.13
50
.08
50
.10
20
.08
20
.10
0–
Co
rnig
lia
80
.12
30
.11
50
.08
30
.07
50
.06
80
.09
90
.06
8–
Vig
na
le9
0.1
37
0.1
27
0.0
77
0.0
84
0.0
76
0.0
99
0.0
86
0.0
83
–T
reis
o1
00
.12
20
.13
00
.11
40
.08
00
.07
40
.10
90
.10
00
.08
60
.08
4–
Sw
itze
rla
nd
Pia
no
Ma
ga
din
o1
10
.12
90
.12
80
.10
90
.06
90
.06
60
.08
50
.08
60
.07
90
.07
10
.07
7–
Ca
ste
lro
tto
12
0.1
29
0.1
27
0.1
12
0.1
29
0.1
16
0.0
76
0.1
34
0.1
35
0.1
05
0.1
20
0.0
96
–S
lov
en
iaG
ori
ska
Brd
a1
30
.12
40
.12
30
.10
20
.07
20
.05
80
.08
90
.07
30
.07
10
.07
80
.06
50
.05
90
.10
6–
Izo
la1
40
.12
20
.11
00
.11
00
.09
60
.08
10
.07
70
.10
00
.09
70
.09
70
.10
10
.07
80
.08
90
.08
5–
No
va
Go
rica
15
0.1
10
0.1
29
0.1
15
0.0
92
0.0
86
0.1
06
0.0
93
0.0
83
0.0
94
0.0
55
0.0
73
0.1
09
0.0
66
0.0
92
–S
pa
inP
en
ed
es
16
0.1
42
0.1
37
0.0
75
0.1
04
0.0
85
0.1
20
0.0
68
0.0
82
0.0
89
0.0
97
0.1
00
0.1
01
0.0
74
0.1
14
0.1
01
–
539
1000
Chambave
Castelrotto
Izola
Piano Magadino
Ponteves
Chatillon
Goriska Brda
Vignale
Penedes
Corniglia
San Colombano
Treiso
Nova Gorica
Geneva Ontario
Valois
Moroges
Fig. 3 Neighbour Joining consensus tree based on Nei’s geneticdistances among 16 populations of S. titanus. Only bootstrapvalues greater than 50% (out of 1000 replications) are reported
282 Genetica (2007) 131:275–285
123
differentiation observed between the two French
samples may be related to the longer history of colo-
nization in France.
Thus a scenario involving a first ancient introduction
into Europe, followed by independent and mixed col-
onization events within and between European coun-
tries, may be the most parsimonious conclusion.
On this basis we may explain, for example, the differ-
entiations and similarities observed among and within
Italian and Slovenian samples and between Slovenian
and Swiss samples. Therefore, it is possible that this
scenario is associated with a concomitant effect of the
low mobility of the species and of the wide-range and
long-distance propagation by commercial exchange of
planting material (canes, grafts, scions and rootstocks)
carrying eggs that the leafhopper laided under the bark
and between the bud scales to overwinter (Boudon-
Padieu 2002; Weintraub and Beanland 2006).
Inferences on the diffusion of S. titanus as potential
vector of Flavescence doree
Scaphoideus titanus is not an serious pest insect by
itself, but it causes major concerns in Europe as vector
of Flavescence doree (FD), a GY disease caused by a
phytoplasma which has been decreed a quarantine
organism in the European Community (Boudon-Pa-
dieu 2002). Vitis vinifera, the host plant for S. titanus, is
also the reservoir of FD phytoplasma. The leafhopper
acquires the phytoplasma from infected host plants and
spreads the disease within the vineyard by feeding
activity, playing consequently an important role in the
tri-trophic relationship insect vector–phytoplasma–
plant. The exclusive vine-feeding activity of the insect
vector is sufficient for FD spread into vineyards, even if
the phytoplasma is initially present with a low inci-
dence (Boudon-Padieu 2002). Considering the large
quantity of plant material produced and exchanged
commercially, it is evident that rigorous quality con-
trols on this material are imperative, both for the
presence of phytoplasma infections and for the pres-
ence of leafhopper eggs.
Acknowledgements This study is part of the Project ‘‘Biodiv-ersita della Vite Coltivata in Italia e sua Protezione dai Parassiti’’funded by the ‘‘Fondazione Bussolera-Branca’’ in Mairano diCasteggio, Pavia, Italy. We are indebted to Elisabeth BoudonPadieu (Biologie et Ecologie des Phytoplasmes, UMR PlanteMicrobe Environnement, INRA of Dijon), to Alberto Alma,Simona Palermo and Domenico Bosco (Di.Va.P.R.A. Entomo-logia e Zoologia applicate all’Ambiente ‘‘Carlo Vidano’’, Uni-versita di Torino), and to Piero Attilio Bianco (Istituto diPatologia Vegetale, Universita di Milano), for supplying thewild-collected specimens of Scaphoideus titanus and for helpfuldiscussion about the leafhopper biology. We thank the severalunknown people from France, Spain, Slovenia, Swiss and USAwho provided invaluable assistance in the collection of wildmaterial. We also thank anonymous reviewers for helpful com-ments on the manuscript.
References
Alma A (2002) Diffusione di Scaphoideus titanus Ball in Italia. In:Canova A (ed) Atti Giornate Fitopatologiche, Baselga diPine, Italy, 7–11 April 2002. CLUEB, Bologna, Italy, pp 51–54
Alma A, Bosco D, Danielli A, Bertaccini A, Vibio M, Arzone A(1997) Identification of phytoplasmas in eggs, nymphs andadults of Scaphoideus titanus Ball reared on healthy plants.Insect Mol Biol 6(2):115–121
Baggiolini M, Canevascini V, Caccia R, Tencalla Y, Sobrio G(1968) Presence dans le vignoble du tessin d’une cicadellenearctique nouvelle pour la Suisse, Scaphoideus littoralisBall (Homoptera: Jassidae), vecteur possible de la Flaves-cence doree. Mitt Schwei Entomol Gesell 60:270–275
-0.2
-0.1
0.0
0.1
0.2
0.3
0.4
-0.6 -0.5 -0.4 -0.3 -0.2 -0.1 0.0 0.1 0.2
Moroges
Vignale
Penedes
Castelrotto
Izola
CornigliaSan ColombanoValois
Geneva Ontario
Ponteves
Nova GoricaChantillon
ChambaveGoriska BrdaTreiso
Piano Magadino
PCoA
axi
s 2
(13.
91%
)
PCoA axis 1 (26.92%)
U.S.AFrance (1958)Italy (1963)Switzerland (1968)Slovenia (1987)Spain (1995)
Fig. 4 Principal componentanalysis (PCoA) of 16populations of S. titanus basedon RAPD data. The first andsecond axes explained 26.92and 13.91% of the totalvariation, respectively. Theyear of leafhopper detectionin the different countries isreported in the legend, inparenthesis
Genetica (2007) 131:275–285 283
123

Barnett DE (1976) A revision of the Nearctic species of thegenus Scaphoideus (Homoptera: Cicadellidae). Trans AmEntomol Soc 102:485–593
Baruffi L, Damiani G, Guglielmino CR, Bandi C, Malacrida AR,Gasperi G (1995) Polymorphism within and between pop-ulations of Ceratitis capitata: comparison between RAPDand multilocus enzyme electrophoresis data. Heredity74:425–437
Batlle A, Lavina A, Clair D, Larrue J, Kuszala C, Boudon-Padieu E (1997) Detection of Flavescence doree in grape-vine in Northern Spain. Vitis 36(4):211–212
Black WC IV (1993) PCR with arbitrary primers: approach withcare. Insect Mol Biol 2:1–6
Black WC IV (1995) FORTRAN programs for the analysis ofRAPD-PCR markers in populations. Colorado State Uni-versity, Fort Collins, CO
Bonfils J, Schvester D (1960) The leafhoppers (Homoptera:Auchenorrhynchas) and their relationship with vineyards inSouth-Western France. Ann Epiphyt 11(3):325–336
Boudon-Padieu E (1999) Grapevine phytoplasmas. In: Firstinternet conference on phytopathogenic Mollicutes, 24–29May 1999. http://web.uniud.it/phytoplasma//conf.html
Boudon-Padieu E (2002) Flavescence doree of the grapevine:knowledge and new developments in epidemiology, etiologyand diagnosis. In: Canova A (ed) Atti Giornate Fitopato-logiche, Baselga di Pine, Italy, 7–11 April 2002. CLUEB,Bologna, Italy, pp 15–34
Boudon-Padieu E (2003) The situation of grapevine yellows andcurrent research directions: distribution, diversity, vectors,diffusion and control. In: Proceedings of the 14th ICVGConference, Locorotondo, Italy, 12–17 September 2003
Bressan A, Larrue J, Boudon-Padieu E (2006) Patterns ofphytoplasma-infected and infective Scaphoideus titanusleafhoppers in vineyards with high incidence of Flavescencedoree. Entomol Exp Appl 119:61–69
Bressan A, Girolami V, Boudon-Padieu E (2005a) Reducedfitness of Scaphoideus titanus exposed to Flavescence doreephytoplasma. Entomol Exp Appl 115:283–290
Bressan A, Spiazzi S, Girolami V, Boudon-Padieu E (2005b)Acquisition efficiency of Flavescence doree phytoplasma byScaphoideus titanus Ball from infected tolerant or suscep-tible grapevine cultivars or experimental host plants. Vitis44:143–146
Cavalli-Sforza LL, A.Edwards WF (1967) Phylogenetic analysis:models and estimation procedures. Evolution 21:550–570
Cavalli-Sforza LL, Menozzi P, Piazza A (1994) The history andgeography of human genes. Princeton University Press,Princeton, NJ
Clerc L, Linder C, Gunthart H (1997) Premiere observation enSuisse romande de la cicadelle Scaphoideus titanus Ball(Homoptera: Jassidae), vecteur de la Flavescence doree dela vigne. Rev Suisse Vitic Arboric Hortic 29(4):245–247
Duduk S, Botti S, Ivanovic M, Dukic N, Bertaccini A (2003)Molecular characterization of a Flavescence doree phytopl-asma infecting grapevine in Serbia. In: Proceedings of the14th ICVG Conference, Locorotondo, Italy, 12–17 Septem-ber 2003
Excoffier L, Smouse PE, Quattro JM (1992) Analysis ofmolecular variance inferred from metric distances amongDNA haplotypes: application to human mitochondrial DNArestriction data. Genetics 131:479–491
Felsenstein J (1995) PHYLIP: Phylogeny inference package,Version 3.57 C. University of Washington, Seattle, WA
Firrao G, Gibb K, Streten C (2005) Short taxonomic guide to thegenus ‘Candidatus Phytoplasma’. J Plant Pathol 87(4):249–263
Gabrijel S (1987) Scaphoideus titanus Ball (= S. littoralis Ball), novistetnik vinove loze u Jugoslaviji. Zastita Bija 38(4):349–357
Gower JC (1966) Some distance properties of latent root andvector methods used in multivariate analysis. Biometrika53:325–338
Hoy CW, Heady SE, Koch TA (1992) Species composition,phenology and possible origins of leafhoppers (Cicadelli-dae) in Ohio vegetable crops. J Econ Entomol 85:2336–2343
Kosman E, Leonard KJ (2005) Similarity coefficients formolecular markers in studies of genetic relationshipbetween individuals for haploid, diploid and polyploidspecies. Mol Ecol 14(2):415–424
Lavina A, Batlle A, Larrue J, Daire X, Clair D, Boudon-PadieuE (1995) First report of grapevine bois noir phytoplasma inSpain. Plant Dis 79(10):1075
Lee IM, Gundersen DE, Hammond RW, Davis RE (1994) Useof Mycoplasma-like Organism (MLO) group-specific oligo-nucleotide primers for nested-PCR assays to detect mixed-MLO infections in a single host plant. Phytopathology84:559–566
Lee IM, Martini M, Marcone C, Zhu SF (2004) Classification ofphytoplasma strains in the elm yellows group (16SrV) andproposal of ‘Candidatus Phytoplasma ulmi’ for the phytopl-asma associated with elm yellows. Int J Syst Evol Microbiol54:337–347
Lessio F, Alma A (2004) Dispersal patterns and chromaticresponse of Scaphoideus titanus Ball (Homoptera: Cicadel-lidae), vector of the phytoplasma agent of grapevineFlavescence doree. Agric For Entomol 6:121–127
Lessio F, Alma A (2006) Spatial distribution of nymphs ofScaphoideus titanus (Homoptera: Cicadellidae) in grapesand evaluation of sequential sampling plans. J Econ Ento-mol 99:578–582
Lynch M, Milligan B (1994) Analysis of population geneticstructure with RAPD markers. Mol Ecol 3:91–99
Maixner M, Pearson RC, Boudon-Padieu E, Caudwell A (1993)Scaphoideus titanus, a possible vector of Grapevine Yellowsin New York. Plant Dis 77:408–413
Mantel N (1967) The detection of disease clustering anda generalised regression approach. Cancer Res 27:209–220
McClure MS (1980) Spatial and seasonal distribution of leaf-hopper vectors of peach X-disease in Connecticut. EnvironEntomol 9:668–672
Nei M, Li WH (1985) Mathematical model for studying geneticvariation in terms of restriction endonucleases. Proc NatlAcad Sci USA 76:5269–5273
Nei M (1975) Molecular population genetics and evolution.North Holland, Amsterdam
Nei M (1978) Estimation of average heterozygosity and geneticdistance from a small number of individuals. Genetics89:583–590
Quartau, JA, Guimaraes JM, Andre G (2001) On the occurrencein Portugal of the Nearctic Scaphoideus titanus Ball(Homoptera: Cicadellidae), the natural vector of the grape-vine ‘‘Flavescence doree’’ (FD). In: Carlo Lozzia (ed)IOBC/wprs Bulletin, 24(7):273–276. Proceedings of themeeting of the Working group ‘‘Integrated Control inViticulture’’, Ponte de Lima, Portugal, 3–7 March 2000
Raymond M, Rousset F (1995) GENEPOP: population geneticssoftware for exact tests and ecumenicism. J Hered 86:248–249
Ritland K (2005) Multilocus estimation of pairwise relatednesswith dominant markers. Mol Ecol 14(10):3157–3165
Rohlf FJ (1993) NTSYS-PC. Numerical taxonomy and multi-variate analysis system, Version 1.80. Applied BiostatisticsInc, New York, NY
284 Genetica (2007) 131:275–285
123

Rousset F (1997) Genetic differentiation and estimation of geneflow from F-statistics under isolation by distance. Genetics145:1219–1228
Schneider S, Roessli D, Excoffier L (2000) Arlequin ver. 2.000:A software for population genetics data analysis. Geneticsand Biometry Laboratory, University of Geneva, Switzer-land
Schvester D, Moutous G, Carle P (1962) Scaphoideus littoralisBall (Homoptera: Jassidae), cicadelle vectrice de la Flaves-cence doree de la vigne. Rev Zool Agric Appl 91:118–131
Schvester D, Carle P, Moutous G (1969) Nouvelles donnees surla transmission de la flavescence doree de la vigne parScaphoideus littoralis Ball. Ann Zool Ecol Anim 1969:445–465
Sebastiani F, Meiswinkel R, Gomulski LM, Guglielmino CR,Mellor PS, Malacrida AR, Gasperi G (2001) Moleculardifferentiation of the Old World Culicoides imicola speciescomplex (Diptera, Ceratopogonidae), inferred using ran-dom amplified polymorphic DNA markers. Mol Ecol10:1773–1786
Seljak G (2002) Non-European Auchenorrhyncha (Hemiptera)and their geographic distribution in Slovenia. Acta EntomolSlov 10:97–101
Swofford DL, Selander RB (1981) BIOSYS-1: a FORTRANprogram for the comprehensive analysis of electronicdata in population genetics and systematics. J Hered72:281–283
Vidano C (1964) Scoperta in Italia dello Scaphoideus littoralisBall cicalina Americana collegata alla Flavescence doreedella vite. L’Italia Agric 101:1031–1049
Vidano C (1966) Scoperta della ecologia ampelofila del Cica-dellide Scaphoideus littoralis Ball nella regione nearticaoriginaria. Ann Fac Sci Agrarie Stud Torino 3:297–302
Wang J (2004) Estimating pairwise relatedness from dominantgenetic markers. Mol Ecol 13:3169–3178
Weitraub PG, Beanland L (2006) Insect vectors of phytoplasmas.Ann Rev Entomol 51:91–111
Welsh J, McClelland M (1990) Fingerprinting genomes usingPCR with arbitrary primers. Nucleic Acids Res 18:7213–7218
Williams JGK, Kubelik AR, Livak KJ, Rafalski JA, Tingey SV(1990) DNA polymorphisms amplified by arbitrary primersare useful as genetic markers. Nucleic Acids Res 18:6531–6535
Genetica (2007) 131:275–285 285
123




















