
Frequent detection of human cytomegalovirus in neuroblastoma: A novel therapeutic target?
Upload
independentCategory
view
1download
0
Di�erentiation of neuroblastoma cells by phorbol esters and insulin-likegrowth factor 1 is associated with induction of retinoic acid receptor b geneexpression
German Perez-Juste1 and Ana Aranda*,1
1Instituto de Investigaciones BiomeÂdicas, Consejo Superior de Investigaciones Cientõ®cas and Universidad AutoÂnoma de Madrid,Arturo Duperier 4, Madrid 28029, Spain
The retinoic acid (RA) receptor b isoform (RARb) playsan important role in RA-induced di�erentiation of humanneuroblastoma. In this study we show that insulin-likegrowth factor 1 (IGF-1) and tetradecanoyl phorbolacetate (TPA) induce RARb gene expression inneuroblastoma SH-SY5Y cells. IGF-1 and TPA causeda marked induction of RARb2 promoter activity and hada synergistic e�ect with RA that also upregulatestranscription. The e�ect of RA is mediated by two RAresponsive elements (RAREs), whereas the IGF-1 andTPA actions are independent of the RAREs and map tosequences that overlap the TATA box. These resultssuggest that the signaling pathways stimulated by TPAand IGF-1 could modify the components assembled atthe core RARb2 promoter and activate transcription.Expression of RasVal12 mimics the e�ect of IGF-1 andTPA on the promoter, and a dominant negative Rasmutant abrogates activation. A dominant negative Rafalso blocks activation showing that the Ras-Raf pathwaymediates stimulation of the RARb2 promoter. Ourresults show that neuronal di�erentiation induced bynon-retinoid agents that activate Ras is accompanied byincreased transcription of the RARb gene.
Keywords: neuroblastoma; retinoic acid; RARb; TPA;IGF-1; Ras
Introduction
Neuroblastoma is a common childhood tumor ofneuroectodermal origin with a high rate of spontaneousregression. These tumor cells provide an excellent modelsystem to analyse the mechanisms that revert tumor-igenicity and induce neuronal di�erentiation. Neurobla-toma cell lines can be induced to di�erentiate by severalagents (Thiele, 1991). Di�erentiation of neuroblastomacells, which is characterized by the appearance of longneurites and changes in the expression of several markergenes, is accompanied by arrest of tumor cells growth.Among the agents that induce neuroblastoma celldi�erentiation retinoic acid (RA) appears to be quitepotent (Haussler et al., 1983). This has led to clinicaltrials of retinoids to treat neuroblastoma and otherneoplastic diseases (Gudas et al., 1994; Villablanca et al.,1995). Phorbol esters, which activate protein kinase C
and act as tumor promoters in other cell types alsopromote di�erentiation of neuroblastoma cells as judgedby morphology and expression of neuronal markers(Bjelfman et al., 1990; Pahlman et al., 1991). On the otherhand, insulin-like growth factor 1 (IGF-1), which is amitogen for some undi�erentiated neuroblastoma cells,shifts from promoting cell division to potentiatingmaturation during neuroblastoma cell di�erentiation(Pahlman et al., 1991; Sumantran and Feldman, 1993).The actions of IGF-1 are mediated by tyrosine kinasereceptors. The Ras proto-oncogene is one of theintracellular e�ectors of tyrosine kinase receptors.Ligand activation of these receptors leads to activationof Ras and of the serine/threonine kinase Raf which actsdownstream of Ras in the signaling pathway (LeRoith etal., 1995).
The e�ects of RA are mediated by nuclear receptors(RARs) which belong to the subfamily of nonsteroidhormone receptors (Mangelsdorf and Evans, 1995).These receptors normally act as ligand-inducibletranscription factors by binding to cis-acting respon-sive elements (RA responsive elements or RAREs)which are normally located in the regulatory region oftarget genes. RAREs typically consist of at least twocopies of the consensus sequence PuGG/TTCA, that canbe con®gured into a variety of motifs. Data fromseveral laboratories have shown that the most potentRAREs are con®gured as direct repeats (DRs)separated by 2 or 5 nucleotides (DR2 and DR5,respectively) (Glass, 1994). As opposed to receptors forsteroid hormones that bind to DNA as homodimers,RAR preferentially binds DNA as a heterodimer withthe retinoid X receptor (RXR). Since DRs areinherently assymetric, heterodimers may bind to themwith two distinct polarities. Indeed, it has now beenestablished that on the RARE, RXR occupies the 5'half-site and the heterodimeric partner (e.g. RAR)occupies the 3' half-site (Zechel et al., 1994). Thisparticular arrangement permits RAR, upon ligandbinding, to undergo conformational changes whichfacilitate the recruitment of a coactivator complex.This multisubunit complex contains factors withhistone acetylase activity, which allows chromatinremodeling and creates a permissive state forpromoter activation (Torchia et al., 1998).
RARs are encoded by three di�erent genes (a, band g) which give rise to di�erent isoforms (Glass,1994). While RARa is ubiquitously expressed, RARband RARg expression is more restricted (Ruberte etal., 1991). Although some degree of redundancyamong the di�erent isoforms has been observed inknockout mice (Kastner et al., 1995), the di�erent
*Correspondence: A ArandaReceived 18 January 1999; revised 14 April 1999; accepted 14 April1999
Oncogene (1999) 18, 5393 ± 5402ã 1999 Stockton Press All rights reserved 0950 ± 9232/99 $15.00
http://www.stockton-press.co.uk/onc

receptor isoforms might have distinct biologicalfunctions. It has been described that neuroblastomaprimary tumor tissue and cell lines express RARa,and to a lesser degree RARg. RARb is expressed ata very low level, but is strongly induced by treatmentwith RA (Lovat et al., 1993; Redfern et al., 1994;Carpentier et al., 1997). In neuroblastoma cellstransfected with RARs, only RARb transfectantsexhibited marked growth inhibition and neuritogenice�ects (Cheung et al., 1996). Therefore, the inductionof RARb is thought to play an important role in themorphological di�erentiation as well as in the arrestof neuroblastoma cell growth. The important role ofthis isoform has been con®rmed in recent studies inwhich the e�ects of receptor-selective retinoids onneuroblastoma cells have been analysed (Giannini etal., 1997; Lovat et al., 1997; Irving et al., 1998).
The promoter of the RARb2 isoform is stronglyactivated by RA in undi�erentiated embryonalcarcinoma (EC) cells. The human promoter containsa strong DR5-type RARE (bRARE) located atnucleotides 737 to 753 with respect to transcriptioninitiation (De The et al., 1990; Sucov et al., 1990).More recent studies have suggested the existence of aweaker RARE at positions 767 to 783 (Sanguedolceet al., 1997). The close proximity of the RAREs to theTATA box and the Initiator (Inr) sequence rather thanthe arrangement of these elements appears to be crucialto establish productive interactions of the receptorheterodimers with neighboring components of the basaltranscriptional machinery (Sanguedolce et al., 1997).Additional regulatory elements have been also de-scribed to be involved in RARb2 promoter activation.Thus, a putative tetradecanoyl phorbol acetate (TPA)-responsive element (TRE) at position 778 to 784overlapping the distal RARE, and a cAMP-responsiveelement (CRE) at position 792 to 799 adjacent to
that element have been described to be involved in RA-dependent RARb2 promoter activation in EC cells(Kruyt et al., 1992; Dey et al., 1994).
In this study we report that a common feature ofRA, TPA and IGF-1-induced di�erentiation ofneuroblastoma cells is induction of RARb genetranscription. IGF-1 and TPA induce RARb tran-scripts, as well as RARb2 promoter activation in theneuroblastoma cell line SH-SY5Y. These agents alsopotentiate RA-dependent induction of the RARb gene.The e�ect of TPA is PKC and Ras-dependent, whereasIGF-1 stimulates promoter activity by a mechanismindependent of PKC but dependent of the Ras/MAPKsignaling pathway. A full induction by RA requiresboth RAREs present in the promoter, but stimulationby TPA and IGF-1 is independent of the RAREs andalso of the putative TRE. Furthermore, both agentsinduced activity of a construct which contains only the737 to +14 promoter fragment. These sequences donot contain binding sites for known transcriptionfactors and are composed essentially of the TATAbox and the Inr. Deletion of the Inr does not impairregulation by IGF-1 or TPA, suggesting that an e�ecton components of the basal transcriptional machineryis responsible for stimulation by these factors. Inagreement with this hypothesis, the amount ofcomplexes bound to this sequence increased in IGF-1- and TPA-treated SH-SY5Y cells.
Results
Induction of RARb2 mRNA by RA in SH-SY5Y
We have analysed induction of RARb transcripts byRA in neuroblastoma SH-SY5Y cells, a clonalderivative of SK-N-SH cells which consists solely of
Figure 1 RA induces RARb mRNA transcripts in SH-SY5Y cells. Northern blot analysis were carried out with 30 mg of RNAfrom cells incubated with (a) 1 mM RA for the time periods indicated or (b) with increasing concentrations of RA for 48 h.Representative autoradiograms are shown in the lower panels, and quantitation of RARb mRNA levels obtained in twoindependent experiments each performed with duplicate cultures are illustrated in the upper panels. The values are mean+s.d. andhave been corrected by the amount of RNA applied
Induction of RARb by TPA and IGF-1G Perez-Juste and A Aranda
5394
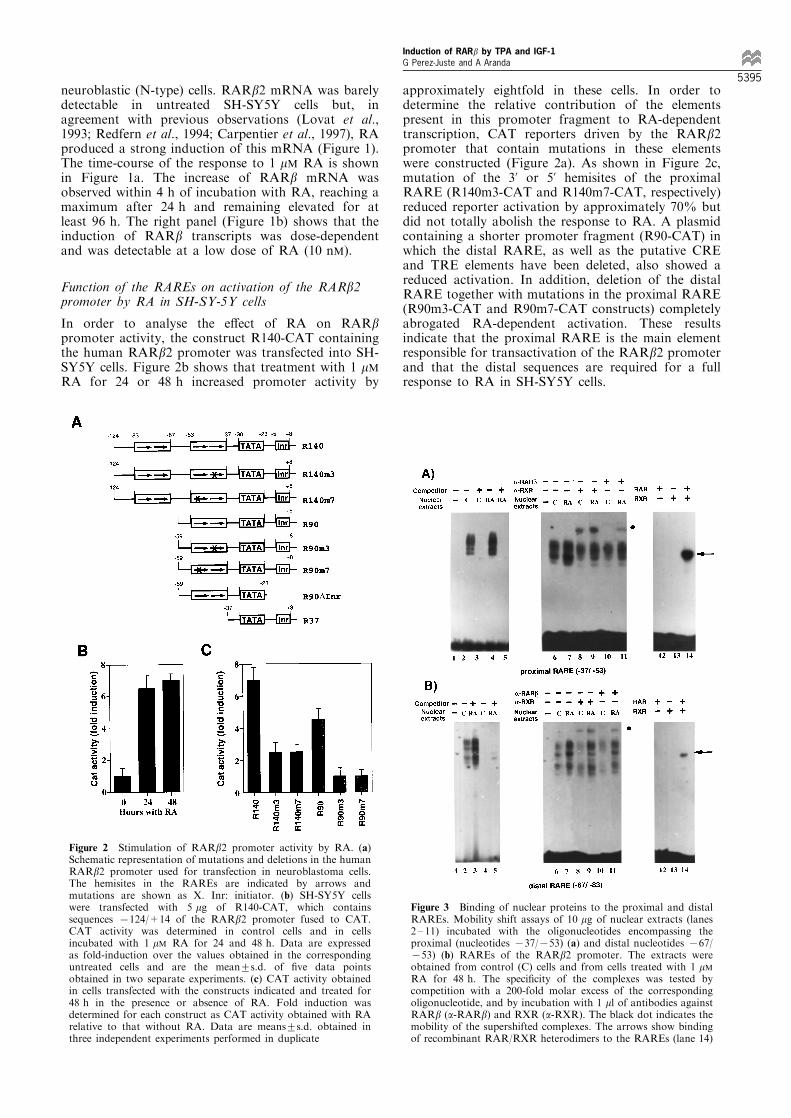
neuroblastic (N-type) cells. RARb2 mRNA was barelydetectable in untreated SH-SY5Y cells but, inagreement with previous observations (Lovat et al.,1993; Redfern et al., 1994; Carpentier et al., 1997), RAproduced a strong induction of this mRNA (Figure 1).The time-course of the response to 1 mM RA is shownin Figure 1a. The increase of RARb mRNA wasobserved within 4 h of incubation with RA, reaching amaximum after 24 h and remaining elevated for atleast 96 h. The right panel (Figure 1b) shows that theinduction of RARb transcripts was dose-dependentand was detectable at a low dose of RA (10 nM).
Function of the RAREs on activation of the RARb2promoter by RA in SH-SY-5Y cells
In order to analyse the e�ect of RA on RARbpromoter activity, the construct R140-CAT containingthe human RARb2 promoter was transfected into SH-SY5Y cells. Figure 2b shows that treatment with 1 mMRA for 24 or 48 h increased promoter activity by
approximately eightfold in these cells. In order todetermine the relative contribution of the elementspresent in this promoter fragment to RA-dependenttranscription, CAT reporters driven by the RARb2promoter that contain mutations in these elementswere constructed (Figure 2a). As shown in Figure 2c,mutation of the 3' or 5' hemisites of the proximalRARE (R140m3-CAT and R140m7-CAT, respectively)reduced reporter activation by approximately 70% butdid not totally abolish the response to RA. A plasmidcontaining a shorter promoter fragment (R90-CAT) inwhich the distal RARE, as well as the putative CREand TRE elements have been deleted, also showed areduced activation. In addition, deletion of the distalRARE together with mutations in the proximal RARE(R90m3-CAT and R90m7-CAT constructs) completelyabrogated RA-dependent activation. These resultsindicate that the proximal RARE is the main elementresponsible for transactivation of the RARb2 promoterand that the distal sequences are required for a fullresponse to RA in SH-SY5Y cells.
Figure 2 Stimulation of RARb2 promoter activity by RA. (a)Schematic representation of mutations and deletions in the humanRARb2 promoter used for transfection in neuroblastoma cells.The hemisites in the RAREs are indicated by arrows andmutations are shown as X. Inr: initiator. (b) SH-SY5Y cellswere transfected with 5 mg of R140-CAT, which containssequences 7124/+14 of the RARb2 promoter fused to CAT.CAT activity was determined in control cells and in cellsincubated with 1 mM RA for 24 and 48 h. Data are expressedas fold-induction over the values obtained in the correspondinguntreated cells and are the mean+s.d. of ®ve data pointsobtained in two separate experiments. (c) CAT activity obtainedin cells transfected with the constructs indicated and treated for48 h in the presence or absence of RA. Fold induction wasdetermined for each construct as CAT activity obtained with RArelative to that without RA. Data are means+s.d. obtained inthree independent experiments performed in duplicate
Figure 3 Binding of nuclear proteins to the proximal and distalRAREs. Mobility shift assays of 10 mg of nuclear extracts (lanes2 ± 11) incubated with the oligonucleotides encompassing theproximal (nucleotides 737/753) (a) and distal nucleotides 767/753) (b) RAREs of the RARb2 promoter. The extracts wereobtained from control (C) cells and from cells treated with 1 mMRA for 48 h. The speci®city of the complexes was tested bycompetition with a 200-fold molar excess of the correspondingoligonucleotide, and by incubation with 1 ml of antibodies againstRARb (a-RARb) and RXR (a-RXR). The black dot indicates themobility of the supershifted complexes. The arrows show bindingof recombinant RAR/RXR heterodimers to the RAREs (lane 14)
Induction of RARb by TPA and IGF-1G Perez-Juste and A Aranda
5395
RARE-binding activity in vitro
The induction of RARb gene expression should resultin increased binding of RXR/RAR heterodimers to theRAREs upon RA treatment. To analyse in vitrooccupancy of these elements, gel retardation assayswere performed with nuclear extracts from SH-SY5Ycells using 32P-labeled distal and proximal RAREs asprobes. As seen in Figure 3, extracts from bothuntreated cells and cells treated with 1 mM RA for48 h produced the appearance of several retardedbands with both probes. These bands were speci®csince they were competed in the presence of an excessof the corresponding unlabeled oligonucleotide (lanes1 ± 5). Similarly, lanes 12 ± 14 show that both RAREsare bound by recombinant RXR/RAR heterodimers.The presence of RARb and RXR in the retardedcomplexes was analysed by incubation with antibodiesagainst both receptors. Lanes 8 and 9 demonstrate thatthe complexes formed with extracts from untreated andRA-treated cells were partially super-shifted in thepresence of an anti-RXR antibody (a-RXR), demon-strating that the binding activity contains this receptorboth before and after RA treatment. In contrast, ananti-RARb antibody (a-RARb) did not cause super-shift of the complexes from control cells (lane 10). Thisindicates the absence of this receptor isoform insigni®cant amounts previous to RA induction.Stimulation of RARb gene expression by RAproduces RARb binding to the RAREs as demon-strated by the appearance of a super-shifted band witha-RARb (lane 8).
TPA and IGF-1 cooperate with RA to induce RARbgene expression
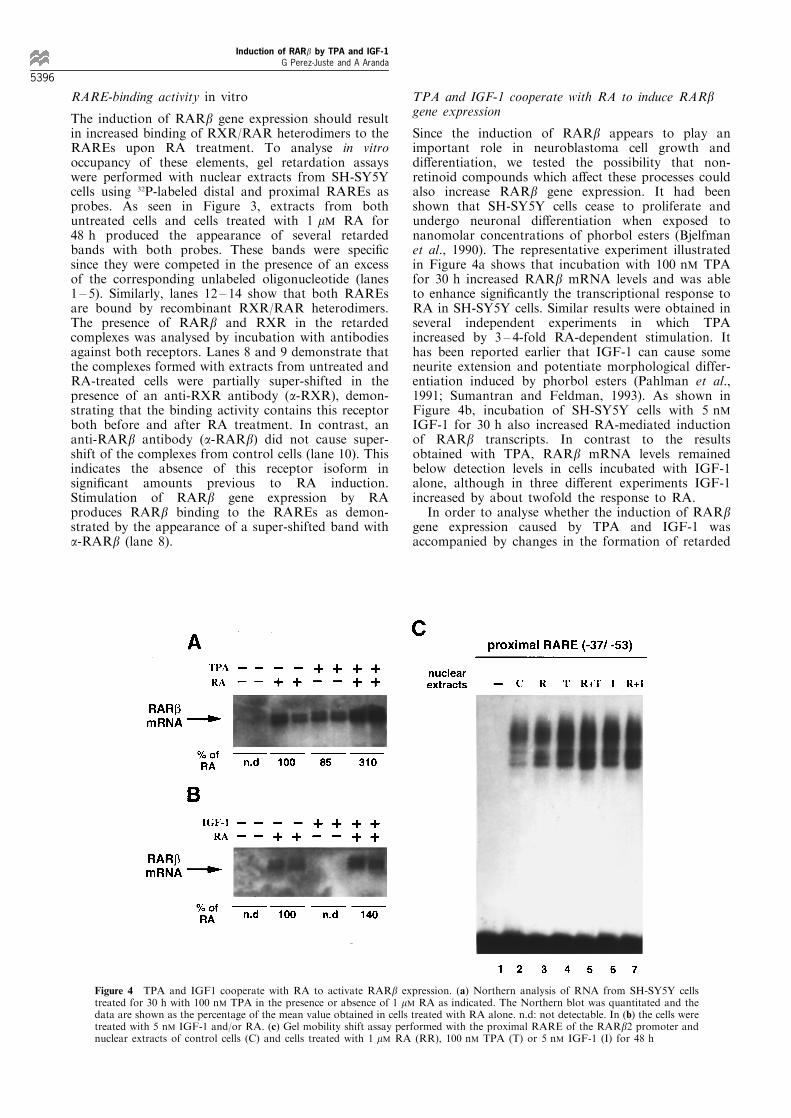
Since the induction of RARb appears to play animportant role in neuroblastoma cell growth anddi�erentiation, we tested the possibility that non-retinoid compounds which a�ect these processes couldalso increase RARb gene expression. It had beenshown that SH-SY5Y cells cease to proliferate andundergo neuronal di�erentiation when exposed tonanomolar concentrations of phorbol esters (Bjelfmanet al., 1990). The representative experiment illustratedin Figure 4a shows that incubation with 100 nM TPAfor 30 h increased RARb mRNA levels and was ableto enhance signi®cantly the transcriptional response toRA in SH-SY5Y cells. Similar results were obtained inseveral independent experiments in which TPAincreased by 3 ± 4-fold RA-dependent stimulation. Ithas been reported earlier that IGF-1 can cause someneurite extension and potentiate morphological differ-entiation induced by phorbol esters (Pahlman et al.,1991; Sumantran and Feldman, 1993). As shown inFigure 4b, incubation of SH-SY5Y cells with 5 nMIGF-1 for 30 h also increased RA-mediated inductionof RARb transcripts. In contrast to the resultsobtained with TPA, RARb mRNA levels remainedbelow detection levels in cells incubated with IGF-1alone, although in three di�erent experiments IGF-1increased by about twofold the response to RA.
In order to analyse whether the induction of RARbgene expression caused by TPA and IGF-1 wasaccompanied by changes in the formation of retarded
Figure 4 TPA and IGF1 cooperate with RA to activate RARb expression. (a) Northern analysis of RNA from SH-SY5Y cellstreated for 30 h with 100 nM TPA in the presence or absence of 1 mM RA as indicated. The Northern blot was quantitated and thedata are shown as the percentage of the mean value obtained in cells treated with RA alone. n.d: not detectable. In (b) the cells weretreated with 5 nM IGF-1 and/or RA. (c) Gel mobility shift assay performed with the proximal RARE of the RARb2 promoter andnuclear extracts of control cells (C) and cells treated with 1 mM RA (RR), 100 nM TPA (T) or 5 nM IGF-1 (I) for 48 h
Induction of RARb by TPA and IGF-1G Perez-Juste and A Aranda
5396
complexes with the proximal RARE, gel retardationassays were performed with nuclear extracts from SH-SY5Y cells treated with these agents in the presenceand absence of RA. Figure 4c shows that treatmentwith TPA or IGF-1 did not cause the appearance ofnew retarded bands but augmented the intensity of thecomplexes. Furthermore, both compounds potentiatedthe e�ect of RA. Similar results were obtained with thedistal RARE (not illustrated).
Activation of the RARb2 promoter by IGF-1 and TPA
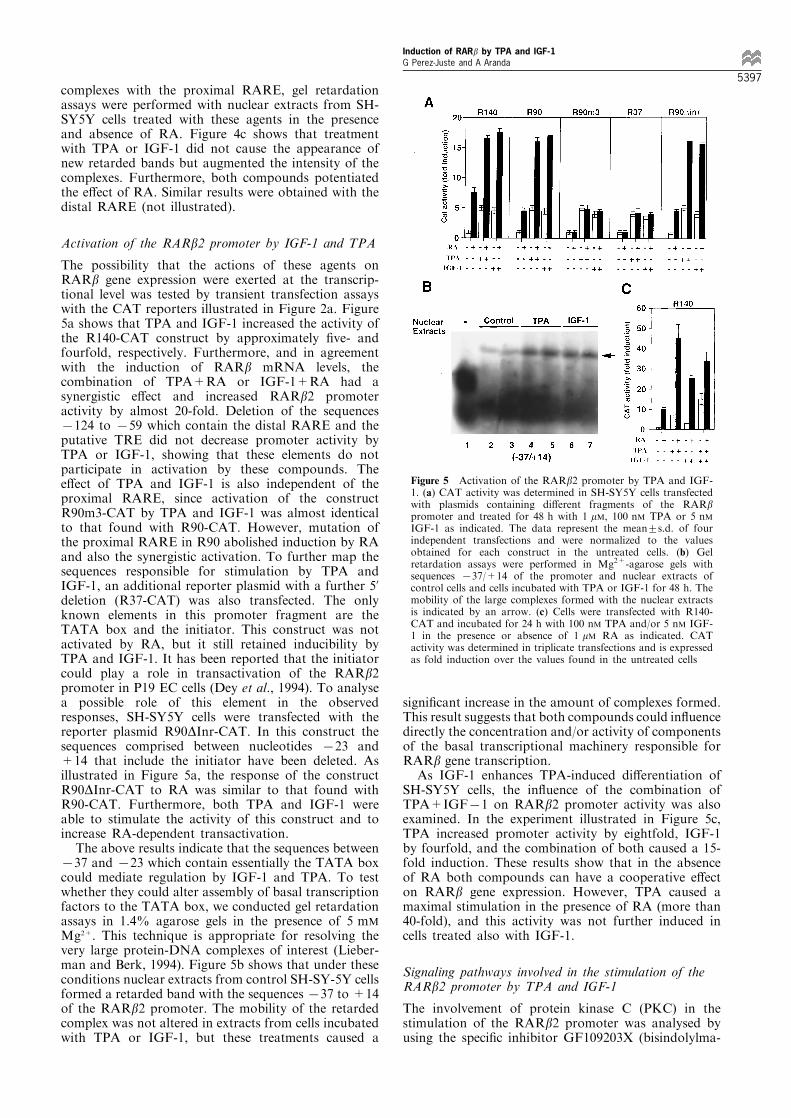
The possibility that the actions of these agents onRARb gene expression were exerted at the transcrip-tional level was tested by transient transfection assayswith the CAT reporters illustrated in Figure 2a. Figure5a shows that TPA and IGF-1 increased the activity ofthe R140-CAT construct by approximately ®ve- andfourfold, respectively. Furthermore, and in agreementwith the induction of RARb mRNA levels, thecombination of TPA+RA or IGF-1+RA had asynergistic e�ect and increased RARb2 promoteractivity by almost 20-fold. Deletion of the sequences7124 to 759 which contain the distal RARE and theputative TRE did not decrease promoter activity byTPA or IGF-1, showing that these elements do notparticipate in activation by these compounds. Thee�ect of TPA and IGF-1 is also independent of theproximal RARE, since activation of the constructR90m3-CAT by TPA and IGF-1 was almost identicalto that found with R90-CAT. However, mutation ofthe proximal RARE in R90 abolished induction by RAand also the synergistic activation. To further map thesequences responsible for stimulation by TPA andIGF-1, an additional reporter plasmid with a further 5'deletion (R37-CAT) was also transfected. The onlyknown elements in this promoter fragment are theTATA box and the initiator. This construct was notactivated by RA, but it still retained inducibility byTPA and IGF-1. It has been reported that the initiatorcould play a role in transactivation of the RARb2promoter in P19 EC cells (Dey et al., 1994). To analysea possible role of this element in the observedresponses, SH-SY5Y cells were transfected with thereporter plasmid R90DInr-CAT. In this construct thesequences comprised between nucleotides 723 and+14 that include the initiator have been deleted. Asillustrated in Figure 5a, the response of the constructR90DInr-CAT to RA was similar to that found withR90-CAT. Furthermore, both TPA and IGF-1 wereable to stimulate the activity of this construct and toincrease RA-dependent transactivation.
The above results indicate that the sequences between737 and 723 which contain essentially the TATA boxcould mediate regulation by IGF-1 and TPA. To testwhether they could alter assembly of basal transcriptionfactors to the TATA box, we conducted gel retardationassays in 1.4% agarose gels in the presence of 5 mM
Mg2+. This technique is appropriate for resolving thevery large protein-DNA complexes of interest (Lieber-man and Berk, 1994). Figure 5b shows that under theseconditions nuclear extracts from control SH-SY-5Y cellsformed a retarded band with the sequences 737 to +14of the RARb2 promoter. The mobility of the retardedcomplex was not altered in extracts from cells incubatedwith TPA or IGF-1, but these treatments caused a
signi®cant increase in the amount of complexes formed.This result suggests that both compounds could in¯uencedirectly the concentration and/or activity of componentsof the basal transcriptional machinery responsible forRARb gene transcription.
As IGF-1 enhances TPA-induced di�erentiation ofSH-SY5Y cells, the in¯uence of the combination ofTPA+IGF71 on RARb2 promoter activity was alsoexamined. In the experiment illustrated in Figure 5c,TPA increased promoter activity by eightfold, IGF-1by fourfold, and the combination of both caused a 15-fold induction. These results show that in the absenceof RA both compounds can have a cooperative e�ecton RARb gene expression. However, TPA caused amaximal stimulation in the presence of RA (more than40-fold), and this activity was not further induced incells treated also with IGF-1.
Signaling pathways involved in the stimulation of theRARb2 promoter by TPA and IGF-1
The involvement of protein kinase C (PKC) in thestimulation of the RARb2 promoter was analysed byusing the speci®c inhibitor GF109203X (bisindolylma-
Figure 5 Activation of the RARb2 promoter by TPA and IGF-1. (a) CAT activity was determined in SH-SY5Y cells transfectedwith plasmids containing di�erent fragments of the RARbpromoter and treated for 48 h with 1 mM, 100 nM TPA or 5 nMIGF-1 as indicated. The data represent the mean+s.d. of fourindependent transfections and were normalized to the valuesobtained for each construct in the untreated cells. (b) Gelretardation assays were performed in Mg2+-agarose gels withsequences 737/+14 of the promoter and nuclear extracts ofcontrol cells and cells incubated with TPA or IGF-1 for 48 h. Themobility of the large complexes formed with the nuclear extractsis indicated by an arrow. (c) Cells were transfected with R140-CAT and incubated for 24 h with 100 nM TPA and/or 5 nM IGF-1 in the presence or absence of 1 mM RA as indicated. CATactivity was determined in triplicate transfections and is expressedas fold induction over the values found in the untreated cells
Induction of RARb by TPA and IGF-1G Perez-Juste and A Aranda
5397
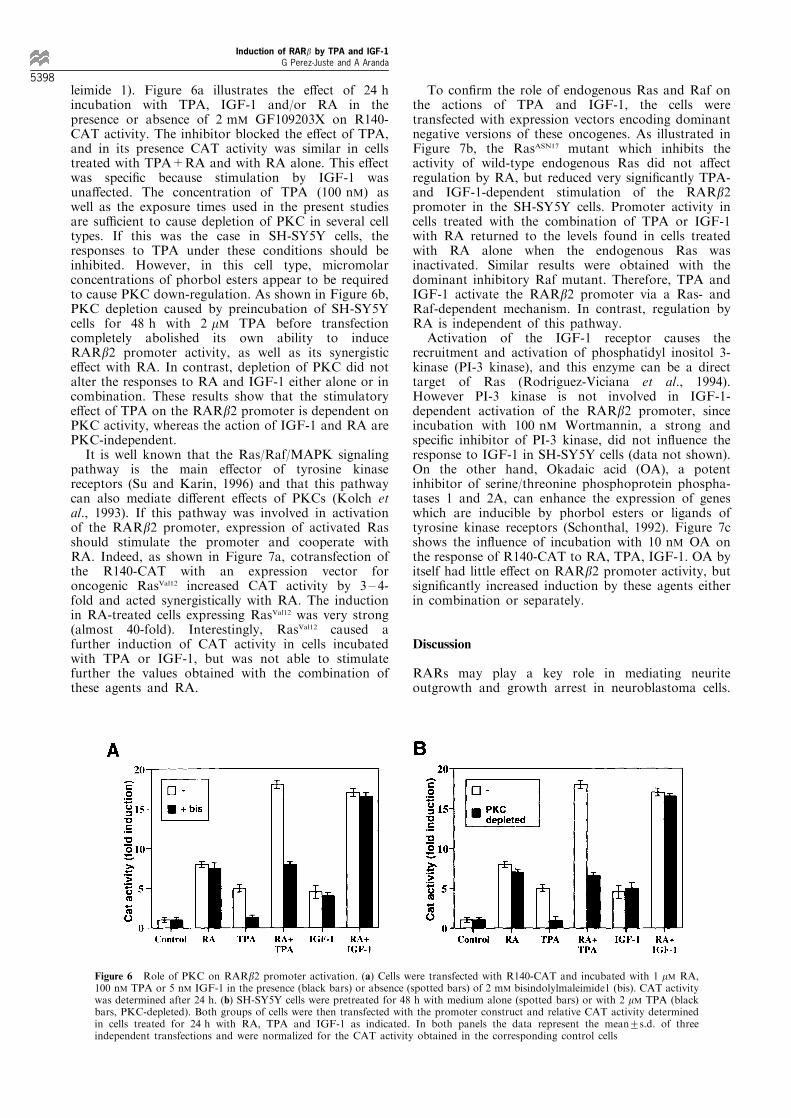
leimide 1). Figure 6a illustrates the e�ect of 24 hincubation with TPA, IGF-1 and/or RA in thepresence or absence of 2 mM GF109203X on R140-CAT activity. The inhibitor blocked the e�ect of TPA,and in its presence CAT activity was similar in cellstreated with TPA+RA and with RA alone. This e�ectwas speci®c because stimulation by IGF-1 wasuna�ected. The concentration of TPA (100 nM) aswell as the exposure times used in the present studiesare su�cient to cause depletion of PKC in several celltypes. If this was the case in SH-SY5Y cells, theresponses to TPA under these conditions should beinhibited. However, in this cell type, micromolarconcentrations of phorbol esters appear to be requiredto cause PKC down-regulation. As shown in Figure 6b,PKC depletion caused by preincubation of SH-SY5Ycells for 48 h with 2 mM TPA before transfectioncompletely abolished its own ability to induceRARb2 promoter activity, as well as its synergistice�ect with RA. In contrast, depletion of PKC did notalter the responses to RA and IGF-1 either alone or incombination. These results show that the stimulatorye�ect of TPA on the RARb2 promoter is dependent onPKC activity, whereas the action of IGF-1 and RA arePKC-independent.
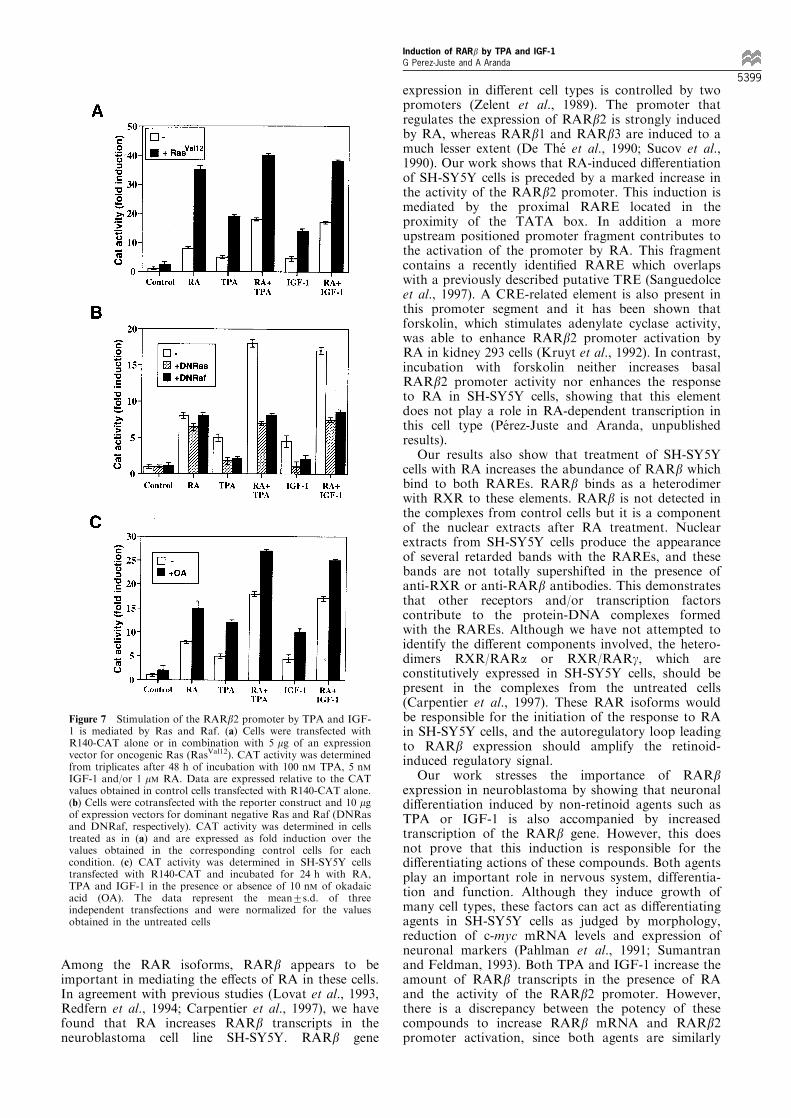
It is well known that the Ras/Raf/MAPK signalingpathway is the main e�ector of tyrosine kinasereceptors (Su and Karin, 1996) and that this pathwaycan also mediate di�erent e�ects of PKCs (Kolch etal., 1993). If this pathway was involved in activationof the RARb2 promoter, expression of activated Rasshould stimulate the promoter and cooperate withRA. Indeed, as shown in Figure 7a, cotransfection ofthe R140-CAT with an expression vector foroncogenic RasVal12 increased CAT activity by 3 ± 4-fold and acted synergistically with RA. The inductionin RA-treated cells expressing RasVal12 was very strong(almost 40-fold). Interestingly, RasVal12 caused afurther induction of CAT activity in cells incubatedwith TPA or IGF-1, but was not able to stimulatefurther the values obtained with the combination ofthese agents and RA.
To con®rm the role of endogenous Ras and Raf onthe actions of TPA and IGF-1, the cells weretransfected with expression vectors encoding dominantnegative versions of these oncogenes. As illustrated inFigure 7b, the RasASN17 mutant which inhibits theactivity of wild-type endogenous Ras did not a�ectregulation by RA, but reduced very signi®cantly TPA-and IGF-1-dependent stimulation of the RARb2promoter in the SH-SY5Y cells. Promoter activity incells treated with the combination of TPA or IGF-1with RA returned to the levels found in cells treatedwith RA alone when the endogenous Ras wasinactivated. Similar results were obtained with thedominant inhibitory Raf mutant. Therefore, TPA andIGF-1 activate the RARb2 promoter via a Ras- andRaf-dependent mechanism. In contrast, regulation byRA is independent of this pathway.
Activation of the IGF-1 receptor causes therecruitment and activation of phosphatidyl inositol 3-kinase (PI-3 kinase), and this enzyme can be a directtarget of Ras (Rodriguez-Viciana et al., 1994).However PI-3 kinase is not involved in IGF-1-dependent activation of the RARb2 promoter, sinceincubation with 100 nM Wortmannin, a strong andspeci®c inhibitor of PI-3 kinase, did not in¯uence theresponse to IGF-1 in SH-SY5Y cells (data not shown).On the other hand, Okadaic acid (OA), a potentinhibitor of serine/threonine phosphoprotein phospha-tases 1 and 2A, can enhance the expression of geneswhich are inducible by phorbol esters or ligands oftyrosine kinase receptors (Schonthal, 1992). Figure 7cshows the in¯uence of incubation with 10 nM OA onthe response of R140-CAT to RA, TPA, IGF-1. OA byitself had little e�ect on RARb2 promoter activity, butsigni®cantly increased induction by these agents eitherin combination or separately.
Discussion
RARs may play a key role in mediating neuriteoutgrowth and growth arrest in neuroblastoma cells.
Figure 6 Role of PKC on RARb2 promoter activation. (a) Cells were transfected with R140-CAT and incubated with 1 mM RA,100 nM TPA or 5 nM IGF-1 in the presence (black bars) or absence (spotted bars) of 2 mM bisindolylmaleimide1 (bis). CAT activitywas determined after 24 h. (b) SH-SY5Y cells were pretreated for 48 h with medium alone (spotted bars) or with 2 mM TPA (blackbars, PKC-depleted). Both groups of cells were then transfected with the promoter construct and relative CAT activity determinedin cells treated for 24 h with RA, TPA and IGF-1 as indicated. In both panels the data represent the mean+s.d. of threeindependent transfections and were normalized for the CAT activity obtained in the corresponding control cells
Induction of RARb by TPA and IGF-1G Perez-Juste and A Aranda
5398
Among the RAR isoforms, RARb appears to beimportant in mediating the e�ects of RA in these cells.In agreement with previous studies (Lovat et al., 1993,Redfern et al., 1994; Carpentier et al., 1997), we havefound that RA increases RARb transcripts in theneuroblastoma cell line SH-SY5Y. RARb gene
expression in di�erent cell types is controlled by twopromoters (Zelent et al., 1989). The promoter thatregulates the expression of RARb2 is strongly inducedby RA, whereas RARb1 and RARb3 are induced to amuch lesser extent (De The et al., 1990; Sucov et al.,1990). Our work shows that RA-induced di�erentiationof SH-SY5Y cells is preceded by a marked increase inthe activity of the RARb2 promoter. This induction ismediated by the proximal RARE located in theproximity of the TATA box. In addition a moreupstream positioned promoter fragment contributes tothe activation of the promoter by RA. This fragmentcontains a recently identi®ed RARE which overlapswith a previously described putative TRE (Sanguedolceet al., 1997). A CRE-related element is also present inthis promoter segment and it has been shown thatforskolin, which stimulates adenylate cyclase activity,was able to enhance RARb2 promoter activation byRA in kidney 293 cells (Kruyt et al., 1992). In contrast,incubation with forskolin neither increases basalRARb2 promoter activity nor enhances the responseto RA in SH-SY5Y cells, showing that this elementdoes not play a role in RA-dependent transcription inthis cell type (Pe rez-Juste and Aranda, unpublishedresults).
Our results also show that treatment of SH-SY5Ycells with RA increases the abundance of RARb whichbind to both RAREs. RARb binds as a heterodimerwith RXR to these elements. RARb is not detected inthe complexes from control cells but it is a componentof the nuclear extracts after RA treatment. Nuclearextracts from SH-SY5Y cells produce the appearanceof several retarded bands with the RAREs, and thesebands are not totally supershifted in the presence ofanti-RXR or anti-RARb antibodies. This demonstratesthat other receptors and/or transcription factorscontribute to the protein-DNA complexes formedwith the RAREs. Although we have not attempted toidentify the di�erent components involved, the hetero-dimers RXR/RARa or RXR/RARg, which areconstitutively expressed in SH-SY5Y cells, should bepresent in the complexes from the untreated cells(Carpentier et al., 1997). These RAR isoforms wouldbe responsible for the initiation of the response to RAin SH-SY5Y cells, and the autoregulatory loop leadingto RARb expression should amplify the retinoid-induced regulatory signal.
Our work stresses the importance of RARbexpression in neuroblastoma by showing that neuronaldi�erentiation induced by non-retinoid agents such asTPA or IGF-1 is also accompanied by increasedtranscription of the RARb gene. However, this doesnot prove that this induction is responsible for thedi�erentiating actions of these compounds. Both agentsplay an important role in nervous system, differentia-tion and function. Although they induce growth ofmany cell types, these factors can act as di�erentiatingagents in SH-SY5Y cells as judged by morphology,reduction of c-myc mRNA levels and expression ofneuronal markers (Pahlman et al., 1991; Sumantranand Feldman, 1993). Both TPA and IGF-1 increase theamount of RARb transcripts in the presence of RAand the activity of the RARb2 promoter. However,there is a discrepancy between the potency of thesecompounds to increase RARb mRNA and RARb2promoter activation, since both agents are similarly
Figure 7 Stimulation of the RARb2 promoter by TPA and IGF-1 is mediated by Ras and Raf. (a) Cells were transfected withR140-CAT alone or in combination with 5 mg of an expressionvector for oncogenic Ras (RasVal12). CAT activity was determinedfrom triplicates after 48 h of incubation with 100 nM TPA, 5 nMIGF-1 and/or 1 mM RA. Data are expressed relative to the CATvalues obtained in control cells transfected with R140-CAT alone.(b) Cells were cotransfected with the reporter construct and 10 mgof expression vectors for dominant negative Ras and Raf (DNRasand DNRaf, respectively). CAT activity was determined in cellstreated as in (a) and are expressed as fold induction over thevalues obtained in the corresponding control cells for eachcondition. (c) CAT activity was determined in SH-SY5Y cellstransfected with R140-CAT and incubated for 24 h with RA,TPA and IGF-1 in the presence or absence of 10 nM of okadaicacid (OA). The data represent the mean+s.d. of threeindependent transfections and were normalized for the valuesobtained in the untreated cells
Induction of RARb by TPA and IGF-1G Perez-Juste and A Aranda
5399

e�ective to increase promoter activity, whereas TPAwas more e�ective than IGF-1 inducing RARb. Thereasons for this discrepancy are presently unclear. Thepresence of a TRE (a TPA-responsive element) in theRARb2 promoter suggested that the phorbol estercould stimulate RARb gene expression through thisDNA element. However, mapping of the elementsresponsible for promoter activation demonstrated thatthe TRE does not mediate stimulation by TPA andmight be a non-functional element. Moreover, both thedistal and proximal RAREs are dispensable for TPA-and IGF-1-dependent promoter activation. In factsequences comprised between 737 and +14 conferfull promoter responsiveness to both agents, aresponsiveness which is not a�ected either by deletionof the initiator-containing sequences 723 to +14.These results predict that proteins interacting in thevicinity of the TATA box, by inference components ofthe basal transcriptional machinery, play an importantregulatory role in activation by TPA and IGF-1. Thereare several indications that transcriptional responsescan be mediated through sequences centered aroundthe TATA box (Wefald et al., 1990; Leitman et al.,1992, Desmarais and Royal, 1996), although themechanisms underlying this type of regulation arepresently unclear. That IGF-1 and TPA could facilitatethe assembly of a functional preinitiation complex issuggested by the ®nding that these agents increase theamount of complexes bound to RARb2 promotersequences containing the TATA box. The TATA-binding protein (TBP) can associate with a variety ofTBP-associated factors (TAFs) and transcriptionalactivators (Hampsey and Reinberg, 1997). It is thenpossible that the signaling pathways stimulated by TPAand IGF-1 could modify the components assembled atthe core RARb2 promoter and activate transcription.Elucidation of the molecular basis for this regulationwill require a ®ne analysis of the molecules associatedwith the transcription initiation complex.
Our results show that PKC activity is required forTPA-dependent transcription of the RARb gene. It hasbeen shown that PKCa is the isoform expressed in SH-SY5Y cells, and that this enzyme remains functionallyactive after long-term treatment with concentrations ofTPA that induce neuronal di�erentiation. Very highconcentrations of TPA appear to be required to causecomplete down-regulation of PKCa in SH-SY5Y cells(Parrow et al., 1992) and, in agreement with thisobservation, preincubation with micromolar concentra-tions of TPA abrogates activation of the RARb2promoter by the phorbol ester. PKC is an importantpoint of intersection of di�erent signal transductionpathways since many cellular receptors for extracellularchemical messengers can activate PKC (Nishizuka,1995). Among them, receptors that have tyrosinekinase activity and activate Ras can also activatePKC (Pawson, 1992). However, PKC is not involved inRARb gene expression by IGF-1 in SH-SY5Y cells. Incontrast the Ras/Raf/MAPK pathway is involved inthe response to both IGF-1 and TPA. Our data showthat expression of activated Ras mimics the e�ect ofboth compounds and synergizes strongly with RA tostimulate the promoter. Furthermore, expression of adominant inhibitory mutant of Ras blocks theinduction of RARb2 promoter stimulation in responseto TPA and IGF-1, as well as the synergism with RA.
These results show the requirement of functional Rasfor these responses. In addition a dominant inhibitoryRaf also inhibited the e�ect of IGF-1 and TPA,demonstrating that endogenous Raf kinase is alsorequired and showing that IGF-1 and TPA activateRARb gene expression through a Ras-Raf dependentsignal transduction in SH-SY5Y cells. The serine/threonine phosphatase inhibitor OA enhances stimula-tion by TPA and IGF-1 and also by RA. Di�erentnuclear receptors, including RARs can be phosphory-lated (Weigel, 1996) and phosphorylation of RARaand RARg by CDK7 and protein kinase A stimulatestransactivation by RA in a promoter contextdependent manner and plays a role in EC celldi�erentiation (Rochette-Egly et al., 1997; Taneja etal., 1997). Thus, there was the possibility that theactions of PKC and Ras on transactivation of theRARb2 promoter by RA in neuroblastoma cells couldbe mediated by receptor phosphorylation. However,the ®nding that IGF-1 and TPA can activate thepromoter in the absence of RAREs suggests that theiractions are mediated by phosphorylation of otherproteins. The identity of these molecules remains tobe established.
SH-SY5Y cells can be considered as representing aclonal strain of neuroblastoma at an immature stage ofdi�erentiation. Our results show that stimulation ofPKC or Ras/MAPK which elicit an arrest in cellgrowth and promote maturation in these cellsconverges with the actions of retinoids to activateRARb gene transcription providing an example ofcross talk among di�erent signaling pathways. Aninhibition of expression of N-myc in cells with thisoncogene ampli®ed (Thiele et al., 1985), or theinduction of TrkB, a tyrosine kinase receptor knownto be important in neuron survival and di�erentiation,may be some of the mechanisms by which RAsubsequently causes neuroblastoma cell maturation(Kaplan et al., 1993; Nakagawara et al., 1994).Retinoids have proven to be of clinical use in thetreatment of di�erent tumors. Our observations thatTPA and IGF-1 increase RA sensitivity can providevaluable directions for the development of newstrategies that can be exploited for the treatment ofneuroblastoma tumors.
Materials and methods
Plasmids
R140-CAT construct containing the fragment 7124 to +14of the human RARb2 promoter was obtained from thepreviously described R140-Luc (14). The promoter fragmentwas obtained by PCR with the oligonucleotides 5'-GGGAAGCTTGGATCCTGGGAGTTGGT-3' and 5'GCT-CTAGAGCTCACTTCCTACTAC-3' and subcloned in Hin-dIII and XbaI sites of pBLCAT8+ (which lacks the AP-1element present in the plasmid backbone), replacing thethymidine kinase (tk) promoter. The same strategy was usedto obtain R140m3 and R140m7 from the parental constructs(Vivanco-Ruiz et al., 1991). In these plasmids the 3' (m3) and5' (m7) half-sites of the proximal RARE have been mutated.In the construct R90-CAT the fragment from 759 to +14 ofthe RARb2 promoter, obtained by PCR with the oligonu-cleotides 5'-GGGAAGCTTGGGAAGGGTTCACCGAA-3'and 5'-GCTCTAGAGCTCACTTCCTACTAC-3', was simi-larly cloned into the HindIII/XbaI sites of pBLCAT8+. The
Induction of RARb by TPA and IGF-1G Perez-Juste and A Aranda
5400

mutant R90m7 was constructed using the R90m7 luciferaseconstruct as template and R90m3 was constructed using as 5'primer the mutated primer 5'-GGGAAGCTTGGGAA-GAACTCACCGAAA-3'. The plasmid R37-CAT containingnucleotides 737 to +14 of the promoter was also obtainedby PCR using as the 5' primer 5'-GGGAAGCTTGCATA-TATTAGGCAATTC-3'. In the construct R90DInr-CAT thatincludes the sequences from 757 to 723, the initiatorsequence was deleted using as the 3' primer 5'-CGCTCTA-GAGAATTGCCTAATATATGC-3'. Constitutive expressionvectors for RasVal12, RasAsn17, and a dominant negative Raflacking the catalytic domain (Cale s et al., 1998; Feig andCooper, 1988; Kolch et al., 1993) under control of the RousSarcoma virus (RSV) promoter have been previouslydescribed.
Cell culture, transient transfections and CAT assays
SH-SY5Y cells were cultured in RPMI containing 10% fetalcalf serum and plated 24 h prior to transfection into 60 mmdishes. The cells were then shifted to DMEM containing 10%AGX100 resin-charcoal treated newborn calf serum andtransfected with calcium phosphate with 5 mg of the reporterplasmid plus 1 mg of a luciferase internal control plasmid). Inco-transfection experiments the reporter plasmids weretransfected with expression vectors for the Ras or Rafmutants. In all cases the total amount of DNA amongdi�erent transfections was kept constant by addition of anempty noncoding expression vector. After 16 h of incubationthe precipitates were washed and the treatments with RA(1 mM), IGF-1 (5 nM) and TPA (100 nM) were administeredin RPMI containing 10% resin-charcoal treated new borncalf serum. After 48 h CAT activity was determined byincubation of the cell extracts with [14C] chloramphenicol.The unreacted and acetylated [14C] chloramphenicol wereseparated by thin layer chromatography and quanti®ed withan Instantimager. Each treatment with the ligands wasperformed at least in duplicate cultures that normallyexhibited less than 10% variation in CAT activity, and theexperiments were repeated at least two or three times withsimilar relative di�erences in regulated expression. The resultsare expressed as the mean+standard deviation (s.d.) of theCAT values obtained.
Gel retardation assays
Nuclear extracts of SH-SY5Y prepared following the methodof Andrews and Faller were used for gel retardation assayswith the oligonucleotides 5'-GGGTAG-GGTTCACC-GAAAGTTCACTCG-3' corresponding to the proximalRARE of the RARb2 promoter and 5'-TGG-TTGGGTCATTTGAAGGTTAGCAGC-3' correspondingto the distal RARE. For the binding reaction, the nuclearextracts (10 mg) or recombinant RARa and RXRa (200 ng)were incubated at room temperature for 15 min at 48C in a
bu�er (10 mM HEPES-(KOH) (pH 7.9), 80 mM KCl, 1 mM
dithiothreitol, 5% Ficoll) containing 3 mg poly (dl-dC) andthen for 30 min at room temperature with approximately30 000 d.p.m. of labeled double stranded oligonucleotide end-labeled with 32P-ATP. For competition experiments an excessof unlabeled double-stranded oligonucleotides were added tothe binding reactions. For supershift experiments, 1 ml ofspeci®c antibodies against RXR (which recognizes thedi�erent receptor isoforms) and RARb were added to thebinding reactions and incubated on ice for 30 min before theaddition of the labeled probe. Finally, DNA-proteincomplexes were resolved on 6% polyacrylamide gels in0.56 Tris-Borate-EDTA bu�er. The gels were then ®xed,dried and autoradiographed at 7708C.
For agarose gel mobility shift assays, 5 mg of nuclearextracts were incubated 15 min at 48C in 10 mM HEPES-(KOH) (pH 7.9), 80 mM KCl, 5 nM MgCl2, 1 mM dithio-threitol, 5% Ficoll, containing 3 mg poly (dG-dC) and thenfor 30 min at room temperature with approximately50 000 c.p.m. of labeled probe corresponding to thenucleotides 737 to +14 of the RARb2 promoter. DNA-protein complexes were resolved on 1.4% agarose gels in0.56 Tris-Borate-EDTA bu�er and 5 mM magnesiumacetate.
RNA extraction and hybridization
Total RNA was extracted from cell cultures with guanidinethiocyanate. The RNA (30 mg) was run in 1% formaldehyde-agarose gels and transferred to nylon-nitrocellulose mem-branes (Nytran) for Northern blot analysis. The RNA wasstained with 0.02% methylene blue, and the blots werehybridized with a cDNA probe for RARb labeled by randomoligonucleotide priming. Membranes were prehybridized for10 min at 658C with 7% SDS, 500 nM phosphate bu�erpH 5.2 and 1 mM EDTA. Hybridizations were carried out forat least 16 h at 658C, and then the ®lters were washed for30 min at 658C in the presence of 1% SDS. Quanti®cation ofmRNA levels was carried out by densitometric scan of theautoradiograms. The values obtained were always correctedfor the amount of RNA applied in each lane which wasdetermined by the densitometry of the methylene blue stainedmembranes.
AcknowledgementsWe thank H Stunnenberg for constructs containing theRARb 2 promoter, E Santos for the Ras and Raf vectors,and P Chambon for the receptor antibodies used in thisstudy. We also thank Jorge Martinez de la Mata for hisvaluable aid. This work has been supported by GrantPM97-0135 from the DGES and 08.1/0032 from theComunidad de Madrid. G Pe rez-Juste is a Fellow fromthe Comunidad de Madrid.
References
Bjelfman C, Meyerson G, Cartwright CA, Mellstrom K,Hammerling U and Pahlman S. (1990). Mol. Cell. Biol.,10, 361 ± 370.
Cale s C, Hancock JF, Marshall CJ and Hall A. (1988).Nature, 332, 548 ± 551.
Carpentier A, Balitrand N, Rochette-Egly C, Shroot B,Degos L and Chomienne C. (1997). Oncogene, 9, 1805 ±1813.
Cheung B, Hocker JE, Smith SA, Reichert U, Norris MD,Haber M, Steward BW and Marshall GM. (1996).Biochem. Biophys. Res. Commun., 229, 349 ± 354.
De The H, Vivanco Ruiz MdM, Tiollais P, Stunnenberg HGand Dejean A. (1990). Nature, 343, 177 ± 180.
Desmarais D and Royal A. (1996). J. Biol. Chem., 271,24976 ± 24981.
Dey A, Minucci and Ozato K. (1994). Mol. Cell. Biol., 14,8191 ± 8201.
Feig L and Cooper GM. (1998). Mol. Cell. Biol., 8, 3235 ±3243.
Giannini G, Dawson MI, Zhang X-k and Thiele CJ. (1997).J. Biol. Chem., 272, 26693 ± 26701.
Glass CK. (1994). Endocr. Rev., 15, 391 ± 407.Gudas LJ, Sporn MB and Roberts AB (ed). (1994). The
Retinoids: Cellular Biology and Biochemistry of theRetinoids. Raven Press: New York, pp. 443 ± 520.
Induction of RARb by TPA and IGF-1G Perez-Juste and A Aranda
5401

Hampsey M and Reinberg D. (1997). Curr. Biol., 7, R44 ± 46.Haussler M, Sidell N, Kelly M, Donaldson C, Altman A and
Mangelsdorf D. (1983). Proc. Natl. Acad. Sci. USA, 80,5525 ± 5529.
Iriving H, Lovat PE, Hewson QC, Malcolm AJ, Pearson ADand Redfern CP. (1998). Eur. J. Cancer, 34, 111 ± 117.
Kaplan DR, Matsumoto K, Lucarelli E and Thiele CJ.(1993). Neuron, 11, 321 ± 331.
Kastner P, Mark M and Chambon P. (1995). Cell, 83, 859 ±869.
Kolch W, Heidecker G, Kochs G, Hummel R, Vahidi H,Mischak H, Finkerzeller G, Marme D, Rapp UR. (1993).Nature, 364, 249 ± 252.
Kruyt FAE, Folkers G, Van den Brink CE and van der SaagPT. (1992). Nucl. Acid. Res., 20, 6393 ± 6399.
Leitman DC, Mackopw ER, Williams T, Baxter JD andWestBL. (1992). Mol. Cell. Biol., 12, 1352 ± 1356.
LeRoith D, Werner H, Beitner-Johnson D, Roberts Jr CT.(1995). Endocr. Rev., 16, 143 ± 163.
Lieberman PM and Berk AJ. (1994). Genes Dev., 8, 995 ±1006.
Lovat PE, Pearson ADJ, Malcolm A and Redfern CPF.(1993). Neurosci. Lett., 162, 109 ± 113.
Lovat PE, Irving H, Annichiarico-Petruzzelli M, BernassolaF, Malcolm AJ, Pearson AD, Melino G and Redfern CP.(1997). Eur. J. Cancer., 33, 2075 ± 2080.
Mangelsdorf DJ and Evans RM. (1995). Cell, 83, 841 ± 850.Nakagawara A, Azar CG, Scavarda NJ and Brodeur GM.
(1994). Mol. Cell. Biol., 14, 759 ± 767.Nishizuka Y. (1995). FASEB J., 9, 484 ± 496.Pahlman S, Meyerson G, Lindgren E, Schalling M and
Johansson I. (1991). Proc. Natl. Acad. Sci. USA, 88,9994 ± 9998.
Parrow V, Nanberg E, Heikkila J, Hammerling U andPahlman S. (1992). J. Cell. Physiol., 152, 536 ± 544.
Pawson T. (1992). Curr. Opin. Genet., 2, 4 ± 12.Redfern CPF, Lovat PE, Malcolm AJ and Pearson ADJ.
(1994). Biochem. J., 304, 147 ± 154.
Rochette-Egly C, Adam S, Rossignol M, Egly J-M andChambon P. (1997). Cell, 90, 97 ± 197.
RodrõÂ guez-Viciana P, Warne PH, Dhand R, VanhaesegroekB, Gout I, Fry MJ, Water®eld MD and Downward J.(1994). Nature, 370, 527 ± 532.
Ruberte E, Dolle P, Chambon P, Morriss-Kay G. (1991).Development, 111, 45 ± 60.
Schonthal A. (1992). New Biol., 4, 16 ± 21.Sanguedolce M, Leblanc BP, Betz JL and Stunnenberg HG.
(1997). EMBO J., 16, 2861 ± 2867.Su B and Karin M. (1996). Curr. Opin. Immunol., 8, 402 ±
411.Sucov HM, Murakami KK and Evans RM. (1990). Proc.
Natl. Acad. Sci. USA, 87, 5392 ± 5396.Sumantran VN and Feldman EL. (1993). Endocrinology,
132, 2017 ± 2023.Taneja R, Rochette-Egly C, Plassat J-L, Penna L, Gaub M-P
and Chambon P. (1997). EMBO J., 16, 6452 ± 6465.Thiele CJ. (1991). Cancer Metast. Rev., 10, 311 ± 319.Thiele C, Reynolds C and Israel M. (1985). Nature, 313,
404 ± 406.Torchia J, Glass CK and Rosenfeld MG. (1998). Curr. Opin.
Cell. Biol., 10, 373 ± 383.Villablanca JG, Khan AA, Avramis VI, Seeger RC, Mathay
KK, Ramsay NKC, Reynolds CP. (1995). J. Clin. Oncol.,13, 894 ± 901.
Vivanco-Ruiz MdM, Bugge TH, Hirschmann P andStunnerberg HG. (1991). EMBO J., 10, 3829 ± 3838.
Wefald FC, Devlin BH and Williams RS. (1990). Nature,344, 260 ± 262.
Weigel LN. (1996). Biochem. J., 319, 657 ± 667.Zechel C, Shen X-Q, Chen J-Y, Chambon P and Gronemeyer
H. (1994). EMBO J., 13, 1425 ± 1433.Zelent A, Krust A, Petkovich M, Kustner P and Chambon P.
(1989). Nature, 339, 714 ± 717.
Induction of RARb by TPA and IGF-1G Perez-Juste and A Aranda
5402
Copyright © 2022 FDOKUMEN




















