
Photorespiration in C 4 grasses remains slow under drought conditions

Dgr
NAa
b
c
d
e
f
g
a
ARRA
KAHMPSW
1
an
WT
n(e(
v
0h
Environmental and Experimental Botany 87 (2013) 69– 78
Contents lists available at SciVerse ScienceDirect
Environmental and Experimental Botany
journa l h omepa g e: www.elsev ier .com/ locate /envexpbot
ifferential tolerance to combined salinity and O2 deficiency in the halophyticrasses Puccinellia ciliata and Thinopyrum ponticum: The importance of K+
etention in roots
.L. Teaklea,b,1, N. Bazihizinaa,b, S. Shabalac, T.D. Colmerb,d,∗, E.G. Barrett-Lennarda,b,e,. Rodrigo-Morenoc,g, A.E. Läuchlib,f
State Centre of Excellence for Ecohydrology, The University of Western Australia, 35 Stirling Highway, Crawley, WA 6009, AustraliaSchool of Plant Biology, The University of Western Australia, 35 Stirling Highway, Crawley, WA 6019, AustraliaSchool of Agricultural Sciences and Tasmanian Institute of Agricultural Research, Private Bag 54, University of Tasmania, Hobart TAS 7001, AustraliaThe UWA Institute of Agriculture, The University of Western Australia, 35 Stirling Highway, Crawley, WA 6009, AustraliaDepartment of Agriculture and Food Western Australia, 3 Baron-Hay Court, South Perth, WA 6151, AustraliaDepartment of Land, Air and Water Resources, University of California, 1 Shields Ave, Davis CA 95616-8627, USAUniversitat Autònoma de Barcelona, Facultat de Biociències. Edifici C, 08193 Bellaterra, Barcelona, Spain
r t i c l e i n f o
rticle history:eceived 21 June 2012eceived in revised form 21 August 2012ccepted 19 September 2012
eywords:dventitious rootsalophytesembrane potential
otassiumodium,
a b s t r a c t
Saline environments of terrestrial halophytes are often prone to waterlogging, yet the effects on halo-phytes of combined salinity and waterlogging have rarely been studied. Either salinity or hypoxia (lowO2) alone can interfere with K+ homeostasis, therefore the combination of salinity or hypoxia is expectedto impact significantly on K+ retention in roots. We studied mechanisms of tolerance to the interactionof salinity with hypoxia in Puccinellia ciliata and Thinopyrum ponticum, halophytic grasses that differ inwaterlogging tolerance. Plants were exposed to aerated and stagnant saline (250 mM NaCl) treatmentswith low (0.25 mM) and high (4 mM) K+ levels; growth, net ion fluxes and tissue ion concentrations weredetermined. P. ciliata was more tolerant than T. ponticum to stagnant-saline treatment, producing twicethe biomass of adventitious roots, which accumulated high levels of Na+, and had lower shoot Na+. After24 h of saline hypoxic treatment, MIFE measurements revealed a net uptake of K+ (∼40 nmol m−2 s−1) for
+ −2 −1
aterlogging P. ciliata, but a net loss of K (∼20 nmol m s ) for the more waterlogging sensitive T. ponticum. NaClalone induced K+ efflux from roots of both species, with channel blocker tests implicating GORK-likechannels. P. ciliata had constitutively a more negative root cell membrane potential than T. ponticum(−150 versus −115 mV). Tolerance to salinity and hypoxia in P. ciliata is related to increased productionof adventitious roots, regulation of shoot K+/Na+, and a superior ability to maintain negative membranepotential in root cells, resulting in greater retention of K+.. Introduction
Salinity is an environmental constraint that affects plant growthnd development in a range of landscapes, for both irrigated andon-irrigated agricultural land (Ghassemi et al., 1995). Species
∗ Corresponding author at: School of Plant Biology (M084), The University ofestern Australia, 35 Stirling Highway, Crawley, WA 6009, Australia.
el.: +61 8 6488 1993; fax: +61 8 6488 1108.E-mail addresses: [email protected] (N.L. Teakle),
[email protected] (N. Bazihizina), [email protected]. Shabala), [email protected] (T.D. Colmer),[email protected] (E.G. Barrett-Lennard), [email protected]. Rodrigo-Moreno), [email protected] (A.E. Läuchli).
1 Present address: Natasha Ayers, Graduate Research School, Edith Cowan Uni-ersity, 270 Joondalup Dr, Joondalup, WA 6027, Australia.
098-8472/$ – see front matter © 2012 Elsevier B.V. All rights reserved.ttp://dx.doi.org/10.1016/j.envexpbot.2012.09.006
© 2012 Elsevier B.V. All rights reserved.
that thrive in these environments and are adapted to salinityare called halophytes. The physiology of halophytes has beenstudied extensively, and their salinity tolerance is primarily gov-erned by controlled uptake and compartmentalization of Na+,K+, and Cl− (Flowers and Colmer, 2008; Shabala and Mackay,2011). Many of the habitats occupied by terrestrial halophytesare not only saline, but also prone to waterlogging (Colmer andFlowers, 2008). Waterlogging causes soils to become hypoxic, dueto biological consumption of O2 without its effective replacement(Ponnamperuma, 1984). While the growth effects of combinedsalinity and waterlogging have been studied for some halophytes(see Colmer and Flowers, 2008), the mechanisms of tolerance are
poorly understood. Furthermore, the adverse interactive effects ofthese stresses must be considered in the design of revegetation pro-grammes for salt-affected lands (Barrett-Lennard, 2003; Bennettet al., 2009).
7 nd Experimental Botany 87 (2013) 69– 78
eThla(gtJais
wtgsalettmtpgep
miotglr(aittwegowPwhithcba
rtpieaogo
Table 1Nutrient solution (full-strength) used for growing Puccinellia ciliata and Thinopyrumponticum in hydroponics. For stagnant treatments, agar was added to the same solu-tions at either 0.1% (Experiments 1–3) or 0.05% (Experiment 4). For all solutions, pHwas adjusted to 6.3 with NaOH.
Final pot concentration (mM)
4 mM K+
(‘normal’)0.25 mM K(‘low’)
MacronutrientsCaSO4 4 4Na2O3Si 0.1 0.1KH2PO4 0.2 0.2MgSO4 0.4 0.4NH4NO3 0.625 0.625FeNa3EDTA 0.05 0.05KNO3 3.75 0NaNO3 0 3.75NaCl 0 or 250 0 or 246.4
MicronutrientsKCl 0.05 0.05H3BO3 0.025 0.025MnSO4 0.002 0.002ZnSO4 0.002 0.002CuSO4 0.0005 0.0005Na2MoO4 0.0005 0.0005NiSO4 0.001 0.001
0 N.L. Teakle et al. / Environmental a
Some halophytes show differential waterlogging tolerance, forxample the perennial pasture grasses Puccinellia ciliata (Bor.) andhinopyrum ponticum (Podp.) Z.-W. Liu and R.R.-C.Wang. P. ciliataas high waterlogging tolerance, whereas T. ponticum has much
ower waterlogging tolerance (Bennett et al., 2009). These grassesre often planted on saline agricultural land as fodder for livestockBarrett-Lennard, 2003). Differential stress tolerance of these tworasses was considered to be primarily caused by differences in theolerance to waterlogging under saline conditions (Jenkins, 2007;enkins et al., 2010). However, the underlying physiological mech-nisms of the differential waterlogging-salinity tolerances and thempact on salt uptake and sequestration are not yet fully under-tood.
Most studies aimed at plant responses to combined salinity andaterlogging have been conducted on agricultural crops, showing
hat shoot Na+ concentrations of plants grown under waterlog-ing in saline conditions increased above those of plants in aeratedaline zones by as much as 6-fold (Barrett-Lennard, 2003). Even
short-term root exposure to hypoxia is known to trigger a pro-onged (up to several days) increase in Na+ uptake for barley (Brauert al., 1987). Studies on wheat by Trought and Drew (1980) showedhat nutrient deficiency (including K+) was induced by growinghe plants in N2-flushed nutrient solution; but no salinity treat-
ent was included in this study. More recently, the addition ofhe Nax1 and Nax2 genes to the wheat cultivar Westonia did notrevent a large decrease in leaf K+/Na+ after 10 days of waterlog-ing plus 150 mM NaCl (James et al., 2011). Little is known on theffect of combined salinity and O2 deficiency on K+/Na+ regulation,articularly for halophytes.
Halophytic Puccinellia grasses often inhabit wet, saline environ-ents. In an early study on Puccinellia peisonis, a species that occurs
n a salt marsh region that is frequently flooded in the border areaf Austria–Hungary, Na+ was found to be highly excluded fromhe shoots while K+ levels in the shoot were still high for plantsrown in their natural habitat (Stelzer and Lauchli, 1978). This high-ights the importance of maintaining an optimal cytosolic K+/Na+
atio, an important feature of salinity tolerance in some plantsMaathuis and Amtmann, 1999; Shabala and Cuin, 2008; Munnsnd Tester, 2008). In a more recent study on Puccinellia tenuiflora,t was demonstrated that restricting unidirectional Na+ influx intohe root with a strong selectivity for K+ over Na+ likely contributedo salt tolerance, but the possible interaction with waterloggingas not part of this study (Wang et al., 2009). In contrast, Jenkins
t al. (2010) compared the interaction of salinity with waterlog-ing on both P. ciliata and T. ponticum, with the emphasis beingn Na+ and K+ concentrations in the leaves. The combination ofaterlogging together with salinity increased shoot dry matter in
. ciliata by 150%, but decreased it in T. ponticum by 90% (comparedith salinity alone); the growth responses were associated withigher Na+ exclusion and improved maintenance of K+/Na+ ratios
n the shoot of P. ciliata compared to T. ponticum. Greater produc-ion of adventitious roots may be a key plant response to root-zoneypoxia and may partly explain the better shoot K+/Na+ ratio in P.iliata (Jenkins, 2007); however K+/Na+ selectivity of roots has noteen measured for P. ciliata or T. ponticum under combined salinitynd waterlogging.
The low energy status under hypoxic or anoxic conditionsesults in a substantial depolarization of plasma membrane poten-ial in root cells (Buwalda et al., 1988). This can impair ion transportrocesses through voltage-gated uptake channels, directly affect-
ng uptake of most essential cations (e.g. K+, NH4+ or Mg2+) that
nter the root via this pathway (Colmer and Greenway, 2011). Also,
noxia-induced H+ pump inhibition will have a detrimental effectn secondary active transport processes such as uptake of inor-anic anions (e.g. HPO42− or NO3−), high-affinity cation uptake,
r Na+ extrusion via Na+/H+ exchangers (Felle, 2005). It is not
MES buffer (2-[N-morpholino]ethanesulfonic acid)
2.5 2.5
surprising, therefore, that both K+ uptake and exclusion of Na+
can be inhibited under root O2 deficiency (Barrett-Lennard, 2003;Colmer and Flowers, 2008).
The importance of K+/Na+ regulation for salt tolerance has beenmostly discussed in the context of non-halophyte crop species(Greenway and Munns, 1980; Munns and Tester, 2008; Shabalaand Cuin, 2008; Plett and Møller, 2010; Kronzucker and Britto,2011) with only two recent reviews on halophytes (Flowers andColmer, 2008; Shabala and Mackay, 2011). Hordeum marinum-wheat amphiploids had higher leaf K+/Na+ than the wheat parentsunder saline conditions (Munns et al., 2011), but this study did notmeasure ion concentrations under combined salinity and hypoxia.Stelar hypoxia after growing plants in O2 deficient conditions islikely to also affect the K+/Na+-selectivity of transport to the shoot(reviewed by Colmer and Greenway, 2011), however their discus-sion focused only on non-saline treatments. The role of membranevoltage in the maintenance of the optimal cytosolic K+/Na+ ratio,and the mechanisms of K+/Na+ selectivity, should also be evaluatedin halophytic species subjected to combined salinity and water-logging stress. In the present paper, we focus on the responses ofthe halophytic plants P. ciliata and T. ponticum to the interactionof salinity with hypoxia and K+ deficiency. Our working hypothesisis that tolerance to salinity and O2 deficiency in P. ciliata is relatedto its superior ability to maintain negative membrane potential inroot cells, resulting in more efficient K+ retention in root tissuesand thus high K+/Na+ in shoots.
2. Material and methods
2.1. Plant material and growth conditions
Seeds of P. ciliata (cv. Menemen) and T. ponticum (cv. Tyrell)were washed in 0.04% NaHClO, rinsed thoroughly in deionized (DI)water and then imbibed in aerated 0.5 mM CaSO4 in darkness for 3 h
before being transferred to mesh over aerated 10%-strength nutri-ent solution (see Table 1 for composition of full strength solution).After 3 d, seedlings were transferred to 25%-strength nutrient solu-tion and exposed to light. Seven days after imbibition seedlings
nd Ex
waiTiEanrrpw
2s
idamstihdtAaaaN
ivbCssroaho2p
2s
ttTdptE(0(NPih
N.L. Teakle et al. / Environmental a
ere transferred to 50% nutrient solution, still on mesh. Two weeksfter imbibition, seedlings were transplanted into pots contain-ng 4.5 L of 100%-strength nutrient solution (‘normal’ K+ solution,able 1). Nutrient solutions were aerated (unless otherwise spec-fied), changed weekly and topped up with DI water as required.ach pot initially contained 8 plants held by polyethylene foamnd there were four replicate pots per species/treatment combi-ation. Pot bases and lids were wrapped in aluminum foil to keepoots in darkness. Plants were grown in a controlled environmentoom set to 25/20 ◦C and relative humidity 60/80% (day/night), 12 hhotoperiod and PAR at plant level of 400–500 �mol m−2 s−1. Potsere arranged randomly and rotated weekly.
.2. Comparison between species under aerated and stagnantaline treatments (Experiment 1)
Salinity treatments were imposed on half the plants 28 d aftermbibition by increasing the salt concentration by 50 mM NaCl peray until the final concentration of 250 mM was reached. This gave
Na+/Ca2+ ratio of approximately 60:1, similar to seawater andarine-derived soil salinity such as salt lakes, where Puccinellia
pecies naturally grow. Stagnant treatments began the day afterhe final salt concentrations were reached (33 d after imbibition). Tompose stagnant treatments, a hypoxic pre-treatment was given toalf the pots in each salinity treatment by bubbling with N2 until theissolved O2 level was approximately 10% of air-saturated solution,o avoid subsequent ‘anoxic shock’ (Gibbs and Greenway, 2003).fter 24 h, solutions were changed to a stagnant deoxygenated (i.e.noxic) 0.1% (w/v) agar solution. Final treatments were imposed for
further 15 d (21 d total treatment time), and consisted of (1) aer-ted, 0.2 mM NaCl; (2) stagnant, 0.2 mM NaCl; (3) aerated, 250 mMaCl; and (4) stagnant, 250 mM NaCl.
Plants were harvested 0 and 21 d after salt treatments weremposed (28 and 49 d after imbibition, respectively). For each har-est, shoots were separated from roots, rinsed in DI water andlotted dry. Roots were rinsed 3 times, for 10 s each time, in 4 mMaSO4 plus mannitol iso-osmotic to 250 mM NaCl. Roots were theneparated into seminal and adventitious roots. Dead leaves wereeparated from shoots and a young fully expanded leaf was alsoemoved. Fresh mass of shoot and root samples was recorded andven-dried for 3 d at 70 ◦C for dry mass (DM) measurements. Na+
nd K+ concentrations were measured in dried tissue samples thatad been ground to a fine powder and 100 mg extracted with 10 mlf 0.5 M HNO3 by shaking for 48 h in darkness at 30 ◦C (Munns et al.,010). Diluted extracts were analysed for Na+ and K+ using a flamehotometer (Jenway PFP7, Essex, UK).
.3. K+/Na+ selectivity under variable external K+ with stagnantaline treatments (Experiment 2)
Plants were grown as above, except 24 d after imbibition halfhe plants were transferred to 0.25 mM K+ nutrient solution, andhe other half remained at 4 mM K+. For details of the solutions seeable 1. NaCl was added to all pots 28 d after imbibition in 50 mMaily increments until the final concentration of 250 mM. A hypoxicre-treatment was given 24 h later to half the pots and stagnantreatments were imposed the following day (see description forxperiment 1). Final treatments were imposed for a further 15 d21 d total salt treatment, 26 d low K+), and consisted of (1) aerated,.25 mM K+; (2) stagnant, 0.25 mM K+; (3) aerated, 4 mM K+; and4) stagnant, 4 mM K+. Note all four treatments contained 250 mM
aCl. Solutions were changed every two days to avoid K+ depletion.lants were sampled 0 and 16 d after stagnant treatments weremposed (34 and 50 d after imbibition, respectively). Plants werearvested and measurements taken as in Experiment 1.perimental Botany 87 (2013) 69– 78 71
2.4. Changes in net uptake of K+ due to hypoxic and salinetreatments (Experiment 3)
Plants were grown as in Experiment 1. Salinity was imposedon half the pots 28 d after imbibition by adding 50 mM NaCl dailyuntil the final concentration of 250 mM was reached. Two dayslater all plants were transferred to 0.25 mM K+ solutions (with 0.2or 250 mM NaCl) so that changes in K+ uptake or loss would bedetectable. After 2 d ‘pre-treatment’ in low K+ solutions, all plantswere transferred from the 4.5 L pots to individual 100 mL vials(containing the same solution), which were either bubbled withhumidified air (aerated treatment) or nitrogen (hypoxic treatment)for a total of 24 h. Vials with no plants and bubbled with humid-ified air or nitrogen were also included as evaporation controls.Final treatments consisted of (1) aerated, 0.2 mM NaCl; (2) aerated,250 mM NaCl; (3) hypoxic, 0.2 mM NaCl; and (4) hypoxic, 250 mMNaCl. Note all solutions still contained 0.25 mM K+. Plants were har-vested at 0, 1, 3, 6, 12 and 24 h after hypoxic treatment was imposed.At each time point, 1 mL of solution was removed for measuring K+
uptake or loss from roots and all vials (containing one plant andthe respective solution) were weighed to estimate transpirationlosses. Data from the first hour were excluded from the analysisas K+ losses occurred due to handling of the plants. Solutions werechanged after 6 h and 12 h to avoid K+ depletion. Plants were har-vested 0 and 24 h after the hypoxic treatment commenced for massand ion concentrations as in Experiment 1. K+ was measured inthe solutions from each time point using a flame photometer. K+
uptake or loss was calculated from the changes in K+ of the externalsolution between each time point, relative to the concentration at1 h, accounting for loss of water due to transpiration and nutrientsolution changes.
2.5. Use of MIFE to measure ion fluxes and membrane potential(Experiment 4)
P. ciliata and T. ponticum were grown as described for Experi-ment 1. All plants were initially exposed to aerated 4 mM K+ and,2 d after step-up to 250 mM NaCl, K+ concentrations in the mediawere reduced to 0.25 mM K+ (i.e. ‘low’ K+; Table 1). Net fluxes ofK+ and H+ were measured three days later using the non-invasiveMIFE system (University of Tasmania, Hobart, Australia). Details onthe fabrication and calibration of the microelectrodes have beendescribed previously (Shabala and Newman, 2000; Shabala andShabala, 2002). Net fluxes were measured on adventitious rootsof P. ciliata and T. ponticum (about 200 mm long), essentially asdescribed for barley roots by Pang et al. (2006). Briefly, plants wereselected and placed in a measuring chamber with 60 mL of basalsolution unbuffered (BSM). The BSM had a pH of 5.5 and contained0.1 mM CaCl2 and 0.25 mM KCl, plus either 0.2 or 250 mM NaClaccording to treatment. The low Ca2+ level was chosen to avoidblocking effects of bath Ca2+ upon conductance of non-selective(NSCC; a major route of Na+ transport in plant roots; Demidchik andTester, 2002) and K+—selective outward rectifying (Shabala et al.,2006a) channels. Adventitious roots were then immobilized in thehorizontal position and left for 1.5 h. The chamber was then putonto the microscope stage in the Faraday cage. K+ and H+ selectivemicroelectrodes were positioned close to (50 �m above) the rootsurface. Net H+ and K+ were measured at 2 cm from the root tip byslow (10 s half-cycle) movement of microelectrodes between twopositions, 50 and 100 �m from the tissue, by a computer drivenhydraulic micromanipulator (WR88, Narishige, Tokyo, Japan). Therecorded difference in electrochemical potential between two pos-
itions was then converted into the net ion flux by the specialisedMIFEFLUX software (University of Tasmania, Hobart, Australia) tak-ing into account diffusion profiles, as described elsewhere (Shabalaet al., 2006b).
7 nd Ex
sNbTsiffoiaBga
2
bdKba(t2
2
egti2wLwaasamr
2
(aIndum
3
3
awtDAh
2 N.L. Teakle et al. / Environmental a
Steady state fluxes were measured for 10 min to ensure theirtability and then 60 mL of BSM with 0.1% agar pre-bubbled with2 for 6 h (with 0.2 or 250 mM NaCl) was added to 60 mL of theath solution to give a hypoxic treatment (cf. Pang et al., 2006).his resulted in the final concentration of 0.05% agar in the batholution with O2 concentration of around 20 �mol L−1. As measur-ng chambers could not be aerated due to strict technical limitationor non-stirred layer conditions, the term ‘normoxia’ will be usedor control conditions in the measuring chamber with normal levelsf O2, and the term ‘hypoxic’ will refer to conditions in the measur-ng chamber with 0.05% agar. The unstirred layer conditions tookpproximately 2 min to be regained after the addition of agar to theSM; this period was removed from calculations and appears as aap in the figures. K+ and H+ net fluxes were then monitored fornother 20 min or until stable values were obtained.
.5.1. Pharmacological experimentsGiven the qualitative similarity of ion flux responses to hypoxia
etween the species, pharmacological experiments were con-ucted only on T. ponticum. Two channel blockers – TEA+, a putative+ channel blocker, and Gd3+, a non-selective cation channel (NSCC)locker – were used. These inhibitors were mixed with BSM tochieve the final concentration of 20 mM TEACl or 100 �M GdCl3Demidchik and Tester, 2002; Shabala et al., 2006a) and then addedo roots exposed to 250 mM NaCl followed by K+ flux recordings for0–30 min.
.5.2. Membrane potential measurementsMembrane potentials were measured on epidermal cells in
xcised segments of adventitious roots in P. ciliata and T. ponticumrown in aerated control or saline conditions using the MIFE sys-em according to Chen et al. (2007). Excised root segments weremmobilized in plexigass with 5 mL of BSM with either 0.2 or50 mM NaCl for 24 h before the measurements. Cells were impaledith conventional KCl-filled Ag/AgCl microelectrodes (Shabala and
ew, 2002; Cuin and Shabala, 2005) and membrane potentialsere recorded for 2 min. Subsequently 5 mL of BSM with 0.1%
gar pre-bubbled with N2 for 6 h (with 0.2 or 250 mM NaCl) wasdded resulting in the final concentration of 0.05% agar in the batholution. Measurements were continued for another 20 min afterddition of the agar solution. Five individual plants for each treat-ent were measured with up to 4 readings from each individual
oot.
.6. Statistical analyses
Statistical analyses were performed using Genstat 10th EditionVSN International Ltd. Hemel Hempstead, UK) and graphs cre-ted using SigmaPlot for Windows Version 11.0 (Systat Softwarenc). Residuals were analysed for normality and homogeneity ando transformations were necessary. Two-way ANOVA was used toetermine statistical differences with a significance value of P < 0.05nless otherwise stated. Significant differences between treatmenteans were determined using least significant difference (LSD0.05).
. Results
.1. Biomass responses to stagnant and saline treatments
Shoot and root DM responses of P. ciliata and T. ponticum toerated and stagnant saline treatments were determined. P. ciliataas more tolerant than T. ponticum to all treatments, in particular
o combined stagnant saline (250 mM NaCl) treatment, with shootM 88% of control for P. ciliata and only 20% for T. ponticum (Table 2).dventitious roots as a proportion of total root mass were twice asigh for P. ciliata as T. ponticum under the stagnant saline treatment.
perimental Botany 87 (2013) 69– 78
The shoots of T. ponticum consisted of 13.5% dead leaves, comparedto only 2.8% for P. ciliata with stagnant saline treatment. Roots of P.ciliata were more affected by the treatments than shoots, with theroot:shoot ratio decreasing in response to all treatments (Table 2).
3.2. Ionic relations in plant tissues
Concentrations of Na+ and K+ were measured in shoots, semi-nal and adventitious roots. In both the aerated and stagnant salinetreatments, T. ponticum had approximately twice the shoot Na+
concentration of P. ciliata (Table 3). However, P. ciliata accumulatedmore Na+ in roots than T. ponticum, and both species accumulatedabout 500 �mol g−1 DM more Na+ in adventitious roots, relative toseminal roots, under combined stagnant-saline treatment. Surpris-ingly, there were few differences between the species in shoot orroot K+ concentrations, therefore K+/Na+ ratios followed the trendsdetermined by differences in Na+ concentrations.
3.3. K+/Na+ selectivity under low external K+ and changes in netuptake of K+
P. ciliata and T. ponticum were grown under ‘low’ external K+ todetermine if the species differed in K+/Na+ selectivity in responseto stagnant saline treatment. The ‘low’ K+ treatment consisted of0.25 mM K+, which was not sufficiently low to affect the growth ofthe plants relative to 4 mM K+ (data not shown). For both species,K+/Na+ was affected by both the level of K+ supply and by stagnanttreatment. In shoots, the K+/Na+ ratio decreased by ∼25–60% with0.25 mM external K+ (compared with 4 mM K+) and by ∼50–70%with stagnant compared with aerated treatments (Table 4A). RootK+/Na+ ratios decreased mainly due to low external K+, and thedecrease was greater for P. ciliata, which had about half the K+ con-centration of T. ponticum in both seminal and adventitious roots inresponse to stagnant saline treatment at low external K+ (Table 4B).
To increase the sensitivity of measuring changes in K+ uptake,short-term (0–24 h) uptake measurements were taken for hypoxicand saline-treated plants. Data from the first hour were removedas K+ losses occurred due to transferring the plants from 4.5 L potsto the 100 mL vials. For P. ciliata, there were net K+ losses dueto salinity treatment (aerated and hypoxic) over the 24 h periodof approximately 5–60 �mol g−1 root FM (Fig. 1A). There was nosignificant difference in K+ uptake or loss between stagnant andaerated treatments. In contrast, T. ponticum had significantly higherlosses of K+ from stagnant compared to aerated roots, with morethan 100 �mol g−1 root FM of K+ loss from roots in hypoxic salinetreatment after 24 h (Fig. 1B). By calculating the rates of K+ loss inthe first 12 h compared to the last 12 h of treatment, the detrimen-tal effect of the hypoxic saline treatment on T. ponticum comparedwith P. ciliata was even more evident. For P. ciliata, the rates of K+
uptake increased, or were the same, in the last 12 h of non-salinehypoxic treatment relative to the first 12 h (Fig. 1C). Yet for T. pon-ticum, the rate of K+ loss was up to 2-fold higher in the second 12 hof hypoxic treatment (Fig. 1D). Rates of loss were particularly highfor T. ponticum plants under hypoxic saline conditions, with morethan 2 nmol K+ g−1 root FM s−1 effluxed from roots between 12 and24 h of hypoxic treatment, which was twice the rate of efflux fromP. ciliata.
3.4. K+ and H+ net fluxes in adventitious roots under low externalK+
Net K+ uptakes were measured in adventitious roots of both
P. ciliata and T. ponticum under control (0.2 mM NaCl) ‘normoxic’and ‘hypoxic’ conditions. Under non-saline conditions, the onset ofhypoxia had no effect on net K+ fluxes for both species (Fig. 2A). Inthe short-term measurements, there was no difference in K+ efflux
N.L. Teakle et al. / Environmental and Experimental Botany 87 (2013) 69– 78 73
Table 2Shoot and root dry mass responses of Puccinellia ciliata and Thinopyrum ponticum to saline and stagnant treatments (Experiment 1). 28 d after imbibition NaCl was added in50 mM daily increments until the final concentration of 250 mM. A hypoxic pre-treatment was given 24 h later and stagnant treatments were imposed the following day.Plants were harvested after 21 d treatment. Values are means (±SE, n = 4).
Treatment Species Shoot DMa Root DMa Root/Shoot drymass ratio
Dead leaves (%total shoot mass)
Adventitious roots(% total root mass)
g/plantControl:Aerated0.2 mM NaCl
P. ciliataT. ponticum
2.03 ± 0.64.60 ± 0.8
0.9 ± 0.21.6 ± 0.2
0.50 ± 0.050.40 ± 0.04
00
8.5 ± 1.35.9 ± 1.3
Dry mass % of controlStagnant0.2 mM NaCl
P. ciliataT. ponticum
105 ± 9.464.7 ± 13.6
68.8 ± 19.535.2 ± 7.7
0.34 ± 0.090.47 ± 0.1
1.6 ± 0.53.0 ± 0.7
12.2 ± 4.413.2 ± 1.2
Aerated250 mM NaCl
P. ciliataT. ponticum
85.8 ± 13.231.1 ± 6.2
44.9 ± 17.653.6 ± 11.9
0.29 ± 0.060.99 ± 0.2
05.8 ± 1.1
3.1 ± 1.94.0 ± 1.6
Stagnant250 mM NaCl
P. ciliataT. ponticum
88.5 ± 15.820.4 ± 4.7
36.1 ± 12.923.1 ± 9.0
0.31 ± 0.040.51 ± 0.08
2.8 ± 0.813.5 ± 4.3
21.5 ± 3.29.8 ± 2.1
LSD b 22 28 0.53 4.8 7
valuesot DM
aaaaomrst
iutHbfs
3b
tFan(e
TSaw
0.05
a values are g/plant for control treatment only for shoot and root DM, treatment
b LSD values are for treatment × species interaction (% of control for shoot and ro
fter 20 min hypoxia for saline-treated plants. However, over 24nd 48 h, differences between species emerged (Fig. 3). For P. cili-ta under saline conditions there was a shift in K+ flux kinetics,nd compared to the initial K+ efflux measured 2 h after the onsetf hypoxia, a net K+ influx of approximately 40 nmol m−2 s−1 waseasured after 24 and 48 h hypoxia (Fig. 3B). Interestingly, this
ecovery of K+ uptake was not observed in P. ciliata under normoxicaline conditions (Fig. 3A). For T. ponticum in hypoxic saline condi-ions there was a net loss of K+ of about 20 nmol m−2 s−1 (Fig. 3B).
In contrast to K+ flux kinetics, 20 min of hypoxia caused a signif-cant reduction in net H+ effluxes compared with those measurednder normoxic conditions (Fig. 2C and D). Moreover under con-rol (0.2 mM NaCl) normoxic conditions, significantly higher net+ efflux rates were measured in P. ciliata compared to T. ponticumefore and after the onset of hypoxia (Fig. 2C). By contrast, no dif-erences were found in the net H+ efflux rate between species underaline conditions before and after the onset of hypoxia (Fig. 2D).
.5. Net K+ fluxes in adventitious roots as mediated by channellockers
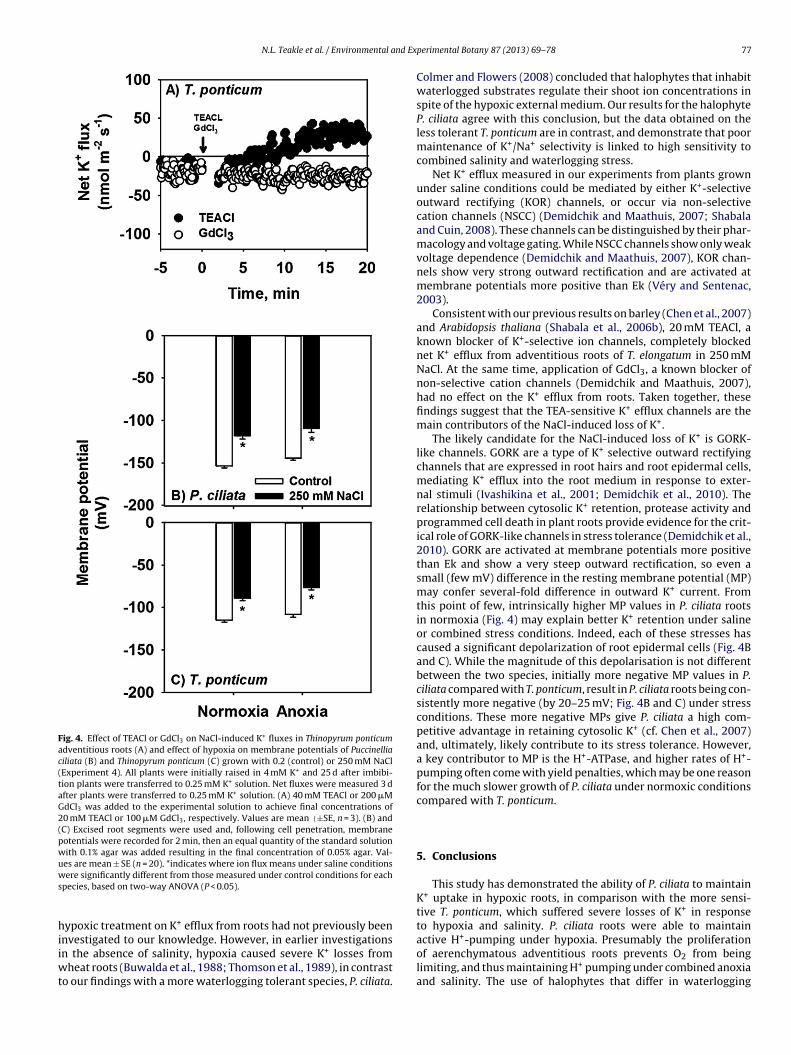
Channel blockers were applied to T. ponticum adventitious rootso determine the cause of the NaCl-induced K+ efflux measured inig. 2. 20 mM of TEA completely blocked net K+ efflux from the
dventitious roots of T. ponticum grown with 250 mM NaCl, andet K+ influxes were measured about 5 min after the treatmentFig. 4A). Conversely, applying GdCl3 had no effect on the net K+fflux from roots.
able 3hoot and root concentrations (�mol g−1 DM) of Na+ and K+ of Puccinellia ciliata and Thfter imbibition NaCl was added in 50 mM daily increments until the final concentration
ere imposed the following day. Plants were harvested after 21 d treatment. Values are m
Treatment Species Ion concentrations (�mol g−1 DM)
Shoot Na+ Shoot K+ S
Aerated0.2 mM NaCl
P. ciliataT. ponticum
120 ± 51110 ± 39
937 ± 801066 ± 90
33
Stagnant0.2 mM NaCl
P. ciliataT. ponticum
72 ± 3679 ± 32
655 ± 46424 ± 35
18
Aerated250 mM NaCl
P. ciliataT. ponticum
519 ± 981045 ± 138
849 ± 69877 ± 111
11
Stagnant250 mM NaCl
P. ciliataT. ponticum
690 ± 541052 ± 62
463 ± 49420 ± 18
11
LSD0.05 214 294 1
are expressed as % of control).
3.6. Membrane potentials in adventitious roots under salinityand hypoxia
Differences in membrane potential could affect K+ retention,therefore we measured the membrane potential of epidermal cellsin excised segments of adventitious roots from P. ciliata and T.ponticum. P. ciliata constitutively had a more negative membranepotential than T. ponticum, with −150 mV compared with −115 mVunder control condition (Fig. 4B and C). For both species, treatmentwith 250 mM NaCl caused depolarisation, with membrane poten-tials about 25% less negative. Under non-saline conditions, hypoxiadid not affect membrane potentials. However, for T. ponticum undersaline conditions, hypoxia did cause a small significant decrease(14% compared with normoxic saline, P < 0.05) in membrane poten-tial to about −75 mV (Fig. 4C). By contrast, the membrane potentialfor P. ciliata under hypoxic saline conditions was −110 mV, andthis was similar to the membrane potential under aerated salineconditions (Fig. 4A).
4. Discussion
The superior tolerance of P. ciliata to combined salt and water-logging stress, as compared with T. ponticum, was shown in thisstudy to be associated with maintenance of tissue K+ concentration
in roots, low shoot Na+ (and thus high K+/Na+), production of adven-titious roots and a constitutive more negative membrane potential.Compared to T. ponticum, shoot and root dry mass of P. ciliata wasless affected by combined stagnant saline treatment, with shootinopyrum ponticum after 21 d saline and stagnant treatments (Experiment 1). 28 dof 250 mM. A hypoxic pre-treatment was given 24 h later and stagnant treatmentseans (±SE, n = 4).
eminal Na+ Seminal K+ Adventitious Na+ Adventitious K+
98 ± 384 ± 1.6
1044 ± 1171086 ± 16
370 ± 16238 ± 37
1121 ± 1291115 ± 118
59 ± 110 ± 42
721 ± 28807 ± 45
359 ± 32215 ± 7.1
1262 ± 7.71239 ± 85
434 ± 42185 ± 27
475 ± 70684 ± 69
1998 ± 7.41921 ± 144
500 ± 62779 ± 89
422 ± 87084 ± 53
379 ± 19329 ± 44
2021 ± 911532 ± 48
598 ± 44511 ± 71
31 178 219 291

74 N.L. Teakle et al. / Environmental and Experimental Botany 87 (2013) 69– 78
Table 4Changes in shoot (A) and root (B) K+ and Na+ concentrations for saline-treated plants growing in low (0.25 mM) and standard (4 mM) K+ nutrient solutions under aeratedand stagnant saline (250 mM NaCl) treatments (Experiment 2). All plants were initially raised in 4 mM K+. 24 d after imbibition ‘low K+’ plants were transferred to 0.25 mMK+ nutrient solution. 28 d after imbibition NaCl was added to all pots in 50 mM daily increments until the final concentration of 250 mM. A hypoxic pre-treatment was given24 h later and stagnant treatments were imposed the following day. Plants were harvested after 21 d treatment and concentrations expressed as �mol g−1 DM. Values aremeans (±SE, n = 4). Root Na+ concentrations did not change with different external K+ concentrations, therefore data not shown.
A
Treatment Species Shoot ion concentrations (�mol g−1 DM) K+/Na+
Na+ K+
4.0 mM K+
aeratedP. ciliataT. ponticum
683 ± 110782 ± 65
880 ± 61801 ± 29
1.5 ± 0.41.0 ± 0.09
4.0 mM K+
stagnantP. ciliataT. ponticum
900 ± 501079 ± 58
473 ± 67432 ± 28
0.51 ± 0.080.41 ± 0.04
0.25 mM K+
aeratedP. ciliataT. ponticum
968 ± 107943 ± 77
529 ± 29702 ± 41
0.56 ± 0.040.76 ± 0.09
0.25 mM K+
stagnantP. ciliataT. ponticum
838 ± 301194 ± 99
243 ± 27262 ± 14
0.29 ± 0.040.23 ± 0.03
LSD0.05 234 119 0.4
B
Treatment Species Root ion concentrations (�mol g−1 DM) Seminal K+/Na+ Adventitious K+/Na+
Seminal K+ Adventitious K+
4.0 mM K+
aeratedP. ciliataT. ponticum
391 ± 21587 ± 45
621 ± 91907 ± 54
0.31 ± 0.040.54 ± 0.05
0.30 ± 0.020.47 ± 0.007
4.0 mM K+
stagnantP. ciliataT. ponticum
270 ± 27172 ± 50
442 ± 81362 ± 77
0.22 ± 0.010.15 ± 0.04
0.19 ± 0.030.26 ± 0.05
0.25 mM K+
aeratedP. ciliataT. ponticum
67 ± 11175 ± 8.8
111 ± 10582 ± 40
0.05 ± 0.0080.14 ± 0.01
0.06 ± 0.0070.30 ± 0.006
0.25 mM K+ P. ciliata 93 ± 2.4 166 ± 14.2 0.07 ±0.02 0.21 ± 0.07
dbstptrttw
toAa(t2rwampestt1hti
stagnant T. ponticum 161 ± 11.2
LSD0.05 83
ry mass 88% of control (Table 2). Similar findings were reportedy Jenkins et al. (2010), although the difference between the twopecies was even more than in the present study. A higher propor-ion of dead leaf material was measured in T. ponticum (Table 2),resumably due to a high concentration of Na+ in shoots. T. pon-icum was also unable to prevent rapid loss of K+ from roots inesponse to hypoxia and this is likely a major factor contributingo the higher sensitivity of this species. The role of K+/Na+ selec-ivity of adventitious roots in tolerance to combined salinity andaterlogging tolerance will be discussed.
Proliferation of adventitious roots can be an important responseo waterlogging (Jackson and Drew, 1984); however little is knownn the function of these roots when also exposed to salinity.dventitious roots of wetland species form aerenchyma and oftenlso a barrier to radial O2 loss, which enhances internal aerationArmstrong, 1979; Colmer, 2003). The porosity of P. ciliata adven-itious roots has previously been found to be 27.5% in hypoxic50 mM NaCl nutrient solution (Jenkins, 2007), indicating theseoots will likely receive substantial internal O2 when in saline,aterlogged substrates. In the present study, P. ciliata had extensive
dventitious root production in response to stagnant saline treat-ent, forming 21% of the total root mass, which was twice that of T.
onticum (Table 2). We found that P. ciliata accumulated high lev-ls of Na+ (2021 �mol g−1 DM) in adventitious roots under stagnantaline treatment; 25% more than T. ponticum, which also had twicehe shoot Na+ of P. ciliata (Table 2). For P. ciliata, this was equivalento 22% of total plant Na+ being accumulated in adventitious roots,
0% in seminal roots and 68% transported to the shoot. Both speciesad much higher Na+ concentrations in adventitious comparedo seminal roots, suggesting a possible role for adventitious rootsn accumulating Na+ to reduce shoot uptake. The stimulation of380 ± 26 0.15 ± 0.01 0.24 ± 0.01
232 0.08 0.1
adventitious roots in P. ciliata due to combined salinity and water-logging is an interesting phenomenon, given that for other species,even halophytes, salinity normally impairs adventitious root for-mation (Colmer and Flowers, 2008). In a recent study, salt levelsof 200 and 600 mM NaCl delayed the formation and decreased thenumber of adventitious roots produced by an inland population ofSuaeda salsa (Song et al., 2011). The response of P. ciliata to increaseproduction of highly porous, Na+ accumulating, adventitious rootscould be a key trait contributing to the exceptional tolerance of thisspecies to combined salinity and waterlogging.
Adventitious root functioning in O2 deficient substrates, in par-ticular for nutrient uptake, is superior to seminal roots (Colmerand Greenway, 2011). For both P. ciliata and T. ponticum, K+ con-centrations were higher in adventitious compared to seminal roots(Tables 3 and 4), demonstrating these roots with more aerenchyma(Jenkins, 2007) are better able to maintain nutrient uptake. Similarfindings were reported for wheat, where net K+ uptake rates understagnant conditions were higher in adventitious compared to semi-nal roots (Wiengweera and Greenway, 2004). Similarly, Buwaldaet al. (1988) reported higher K+ concentrations in adventitiousversus seminal roots, regardless of the roots being excised or intacton whole plants. Presumably higher porosity in adventitious rootsmaintains respiration and thus ATP for critical ion transport pro-cesses (Colmer and Greenway, 2011). Although beyond the scopeof the present work, future studies could examine if differencesexist in H+ extrusion between seminal and adventitious roots. Themechanisms for maintaining root K+ transport in response to com-
bined salinity and waterlogging is discussed below for P. ciliata incomparison with T. ponticum.The differences in loss of K+ from the roots could explain thedifferences in K+/Na+ selectivity between the two species in our

N.L. Teakle et al. / Environmental and Experimental Botany 87 (2013) 69– 78 75
Fig. 1. K+ uptake or loss from roots of Puccinellia ciliata and Thinopyrum ponticum in response to saline and hypoxic treatments (Experiment 3). All plants were initially raisedin 4 mM K+. 28 d after imbibition NaCl was added to saline pots in 50 mM daily increments until the final concentration of 250 mM. 2 d later all plants were transferred to0.25 mM K+ nutrient solution and hypoxic treatments were imposed after a further 2 d for a total of 24 h. Plants were harvested at 0, 1, 3, 6, 12 and 24 h after hypoxic treatmentwas imposed (A and B). Data shown are calculated from the changes in K+ of the external solution between each time point, relative to the concentration at 1 h, accountingfor loss of water due to transpiration and changes in nutrient solution. Rates of K+ uptake or loss from roots were calculated from the change in concentration of the solutionbetween 1 to 12 and 12 to 24 h after hypoxic treatments began (C and D). LSD not shown for P. ciliata (C) as there was no significant difference between treatments or time.D e plan(
sts2ittKsTttsetil2s
ti
ata from the first hour were removed as K+ losses were found due to handling of th±SE, n = 4).
tudy. It has been proposed by several researchers that the main-enance of a high cytosolic K+/Na+ ratio is a key determinant ofalt tolerance (Maathuis and Amtmann, 1999; Shabala and Cuin,008). Our low K+ treatment of 0.25 mM was not low enough to
nduce growth differences, but the decrease in shoot K+ caused byhe stagnant treatment at low external K+ was greater for T. pon-icum than P. ciliata (Table 4). Low external K+ decreased the root+/Na+ ratios to a greater extent in P. ciliata (by almost one-third) inaline conditions, possibly indicating a greater K+ requirement than. ponticum for optimal growth. Our results demonstrate an impor-ant link between K+ deficiency and response to salinity, similaro previous studies. For example, Degl’Innocenti et al. (2009), in atudy on barley, found that K+ deficiency increased the negativeffects on photosynthesis induced by salt. In maize, a combina-ion of K+ deficiency and salt stress inhibited growth more than thendividual stresses, with antioxidative enzymes and antioxidantsower than at the individual K+ deficiency or salt stress (Gong et al.,011). These studies suggest that K+ deficiency might contribute to
alt-induced oxidative stress.The interaction between K+ deficiency and stagnant saline-reated plants is not well understood, and to our knowledge thiss the first study to have directly measured the effect of combined
ts. Solutions were changed after 6 and 12 h to avoid K+ depletion. Values are means
salinity and hypoxia on K+ uptake by roots. K+ deficiency inducedby salinity is likely to be more severe when waterlogging is com-bined with salinity, as demonstrated by lower shoot K+/Na+ ratiosin hypoxic saline compared to aerated saline for sensitive speciessuch as maize (Drew et al., 1988) and wheat (Barrett-Lennard et al.,1999; James et al., 2011). In the present study of two halophytes,the saline hypoxic treatment caused significant K+ loss from rootsfor the more waterlogging sensitive T. ponticum, whereas in P. cili-ata there was no difference between aerated and hypoxic salinetreatment after 12 h (Fig. 1). MIFE measurements showed a netuptake of K+ under saline hypoxic treatment in P. ciliata and a netK+ loss for T. ponticum (Fig. 4). Given that 24 h is sufficient timefor expression and operation of high affinity K+ transporters, themeasured differences in root fluxes between P. ciliata and T. pon-ticum are likely to be physiologically significant and contribute tothe lower shoot K+/Na+ values in T. ponticum. Salt-induced K+ effluxfrom root cells can contribute greatly to the calculated K+/Na+ ratiovalues. For example, in barley cultivars differing in salt tolerance
there was a negative correlation between the magnitude of K+efflux and salt tolerance (Chen et al., 2007). Similar results wereobtained with wheat (Cuin et al., 2008) and lucerne (Smethurstet al., 2005), but the possible impact of a combined salinity and

76 N.L. Teakle et al. / Environmental and Experimental Botany 87 (2013) 69– 78
Fig. 2. Hypoxia-induced transient K+ (A and B) and H+ (C and D) fluxes measured from adventitious roots of Puccinellia ciliata and Thinopyrum ponticum exposed to aerated‘low K+’ solutions with 0.2 or 250 mM NaCl (Experiment 4). All plants were initially raised in 4 mM K+ and 25 d after imbibition plants were transferred to 0.25 mM K+ solution.Net fluxes were measured 3 d after plants were transferred to 0.25 mM K+ solution. Agar was added to the chamber (final concentration 0.05%) to impose hypoxia. Valuesshown in the inserts are the average value for 0–2 min prior and 20–22 min after the onset of hypoxia. Values are means (±SE, n = 6). *indicates where ion flux means after20–22 min hypoxic treatment were significantly different (P < 0.05) based on two-way ANOVA from those measured before hypoxic treatment commenced for each species.
Fig. 3. Steady state K+ net fluxes of Puccinellia ciliata and Thinopyrum ponticum exposed to normoxic (A) or hypoxic (B) ‘low K+’ solutions with 250 mM NaCl (Experiment 4).All plants were initially raised in 4 mM K+ and 25 d after imbibition plants were transferred to 0.25 mM K+ solution. Net fluxes were measured 3 d after plants were transferredto 0.25 mM K+ solution. Steady state K+ net fluxes were measured after 2, 24 and 48 h K+ fluxes (all positive) in control plants exposed to 0.2 mM NaCl are not shown as thelarge declines in K+ net uptake were related to sharp declines in K+ concentrations in the BSM after 24 h, due to uptake and consequent reduction in K+ concentration of thesolutions. Values are means (±SE, n = 4). * indicates where ion flux means were significantly different between species based on two-way ANOVA (P < 0.05).
N.L. Teakle et al. / Environmental and Ex
Fig. 4. Effect of TEACl or GdCl3 on NaCl-induced K+ fluxes in Thinopyrum ponticumadventitious roots (A) and effect of hypoxia on membrane potentials of Puccinelliaciliata (B) and Thinopyrum ponticum (C) grown with 0.2 (control) or 250 mM NaCl(Experiment 4). All plants were initially raised in 4 mM K+ and 25 d after imbibi-tion plants were transferred to 0.25 mM K+ solution. Net fluxes were measured 3 dafter plants were transferred to 0.25 mM K+ solution. (A) 40 mM TEACl or 200 �MGdCl3 was added to the experimental solution to achieve final concentrations of20 mM TEACl or 100 �M GdCl3, respectively. Values are mean (±SE, n = 3). (B) and(C) Excised root segments were used and, following cell penetration, membranepotentials were recorded for 2 min, then an equal quantity of the standard solutionwith 0.1% agar was added resulting in the final concentration of 0.05% agar. Val-ues are mean ± SE (n = 20). *indicates where ion flux means under saline conditionsws
hiiwt
active H+-pumping under hypoxia. Presumably the proliferation
ere significantly different from those measured under control conditions for eachpecies, based on two-way ANOVA (P < 0.05).
ypoxic treatment on K+ efflux from roots had not previously beennvestigated to our knowledge. However, in earlier investigations
n the absence of salinity, hypoxia caused severe K+ losses fromheat roots (Buwalda et al., 1988; Thomson et al., 1989), in contrasto our findings with a more waterlogging tolerant species, P. ciliata.
perimental Botany 87 (2013) 69– 78 77
Colmer and Flowers (2008) concluded that halophytes that inhabitwaterlogged substrates regulate their shoot ion concentrations inspite of the hypoxic external medium. Our results for the halophyteP. ciliata agree with this conclusion, but the data obtained on theless tolerant T. ponticum are in contrast, and demonstrate that poormaintenance of K+/Na+ selectivity is linked to high sensitivity tocombined salinity and waterlogging stress.
Net K+ efflux measured in our experiments from plants grownunder saline conditions could be mediated by either K+-selectiveoutward rectifying (KOR) channels, or occur via non-selectivecation channels (NSCC) (Demidchik and Maathuis, 2007; Shabalaand Cuin, 2008). These channels can be distinguished by their phar-macology and voltage gating. While NSCC channels show only weakvoltage dependence (Demidchik and Maathuis, 2007), KOR chan-nels show very strong outward rectification and are activated atmembrane potentials more positive than Ek (Véry and Sentenac,2003).
Consistent with our previous results on barley (Chen et al., 2007)and Arabidopsis thaliana (Shabala et al., 2006b), 20 mM TEACl, aknown blocker of K+-selective ion channels, completely blockednet K+ efflux from adventitious roots of T. elongatum in 250 mMNaCl. At the same time, application of GdCl3, a known blocker ofnon-selective cation channels (Demidchik and Maathuis, 2007),had no effect on the K+ efflux from roots. Taken together, thesefindings suggest that the TEA-sensitive K+ efflux channels are themain contributors of the NaCl-induced loss of K+.
The likely candidate for the NaCl-induced loss of K+ is GORK-like channels. GORK are a type of K+ selective outward rectifyingchannels that are expressed in root hairs and root epidermal cells,mediating K+ efflux into the root medium in response to exter-nal stimuli (Ivashikina et al., 2001; Demidchik et al., 2010). Therelationship between cytosolic K+ retention, protease activity andprogrammed cell death in plant roots provide evidence for the crit-ical role of GORK-like channels in stress tolerance (Demidchik et al.,2010). GORK are activated at membrane potentials more positivethan Ek and show a very steep outward rectification, so even asmall (few mV) difference in the resting membrane potential (MP)may confer several-fold difference in outward K+ current. Fromthis point of few, intrinsically higher MP values in P. ciliata rootsin normoxia (Fig. 4) may explain better K+ retention under salineor combined stress conditions. Indeed, each of these stresses hascaused a significant depolarization of root epidermal cells (Fig. 4Band C). While the magnitude of this depolarisation is not differentbetween the two species, initially more negative MP values in P.ciliata compared with T. ponticum, result in P. ciliata roots being con-sistently more negative (by 20–25 mV; Fig. 4B and C) under stressconditions. These more negative MPs give P. ciliata a high com-petitive advantage in retaining cytosolic K+ (cf. Chen et al., 2007)and, ultimately, likely contribute to its stress tolerance. However,a key contributor to MP is the H+-ATPase, and higher rates of H+-pumping often come with yield penalties, which may be one reasonfor the much slower growth of P. ciliata under normoxic conditionscompared with T. ponticum.
5. Conclusions
This study has demonstrated the ability of P. ciliata to maintainK+ uptake in hypoxic roots, in comparison with the more sensi-tive T. ponticum, which suffered severe losses of K+ in responseto hypoxia and salinity. P. ciliata roots were able to maintain
of aerenchymatous adventitious roots prevents O2 from beinglimiting, and thus maintaining H+ pumping under combined anoxiaand salinity. The use of halophytes that differ in waterlogging

7 nd Ex
tt
A
CePDaf(p
R
A
B
B
B
B
B
C
C
C
C
C
C
D
D
D
D
D
FF
G
G
G
8 N.L. Teakle et al. / Environmental a
olerance has revealed the importance of K+ retention by roots forolerance to combined salinity and hypoxia.
cknowledgements
Funding was provided to Natasha Teakle by the WA Stateentre of Excellence in Ecohydrology and the University of West-rn Australia (UWA) Faculty of Natural and Agricultural Sciencesump Priming Grant. Andre Läuchli received funding from theepartment of Food and Agriculture Distinguished Visitor Programnd the UWA Visiting Professor Program. Sergey Shabala grate-ully acknowledges the support of the Australian Research CouncilDP1094663 and LP100200456). Ana Rodrigo-Moreno was sup-orted by the Spanish government (BES-2008-005096).
eferences
rmstrong, W., 1979. Aeration in higher plants. Advances in Botanical Research 7,225–332.
arrett-Lennard, E.G., 2003. The interaction between waterlogging and salinity inhigher plants: causes, consequences and implications. Plant and Soil 253, 35–54.
arrett-Lennard, E.G., Ratingen, P., van Mathie, M.H., 1999. The developing pattern ofdamage in wheat (Triticum aestivum L.) due to the combined stresses of salinityand hypoxia: experiments under controlled conditions suggest a methodologyfor plant selection. Australian Journal of Agricultural Research 50, 129–136.
ennett, S.J., Barrett-Lennard, E.G., Colmer, T.D., 2009. Salinity and waterlogging asconstraints to saltland pasture production: a review. Agriculture Ecosystemsand Environment 129, 349–360.
rauer, D., Everett Leggett, J., Egli, D.B., 1987. Changes in K Rb, and Na transport toshoots after anoxia. Plant Physiology 83, 219–224.
uwalda, F., Thomson, C.J., Steigner, W., Barrett-Lennard, E.G., Gibbs, J., Greenway,H., 1988. Hypoxia induces membrane depolarization and potassium loss fromwheat but does not increase permeability to sorbitol. Journal of ExperimentalBotany 39, 1169–1183.
hen, Z.H., Zhoum, M.X., Newman, I.A., Mendham, N.J., Zhang, G.P., Shabala, S., 2007.Potassium and sodium relations in salinised barley tissues as a basis of differ-ential salt tolerance. Functional Plant Biology 34, 150–162.
olmer, T.D., 2003. Long distance transport of gases in plants: a perspective on inter-nal aeration and radial oxygen loss from roots. Plant, Cell & Environment 26,17–36.
olmer, T.D., Flowers, T.J., 2008. Flooding tolerance in halophytes. New Phytologist179, 964–974.
olmer, T.D., Greenway, H., 2011. Ion transport in seminal and adventitiousroots of cereals during O2 deficiency. Journal of Experimental Botany 62,39–57.
uin, T.A., Betts, S.A., Chalmandrier, R., Shabala, S., 2008. A root’s ability to retainK+ correlates with salt tolerance in wheat. Journal of Experimental Botany 59,2697–2706.
uin, T.A., Shabala, S., 2005. Exogenously supplied compatible solutes rapidlyameliorate NaCl-induced potassium efflux from barley roots. Plant and CellPhysiology 46, 1924–1933.
egl’Innocenti, E., Hafsi, C., Guidi, L., Navari-Izzo, F., 2009. The effect of salinity onphotosynthetic activity in potassium-deficient barley species. Journal of PlantPhysiology 166, 1968–1981.
emidchik, V., Tester, M., 2002. Sodium fluxes through nonselective cation channelsin the plasma membrane of protoplasts from Arabidopsis roots. Plant Physiology128, 379–387.
emidchik, V., Cuin, T.A., Svistunenko, D., Smith, S.J., Miller, A.J., Shabala, S.,Sokolik, A., Yurin, V., 2010. Arabidopsis root K+-efflux conductance acti-vated by hydroxyl radicals: single-channel properties, genetic basis andinvolvement in stress-induced cell death. Journal of Cell Science 123,1468–1479.
emidchik, V., Maathuis, F.J.M., 2007. Physiological roles of nonselective cationchannels in plants: from salt stress to signalling and development. New Phy-tologist 175, 387–404.
rew, M.C., Guenther, J., Lauchli, A., 1988. The combined effects of salinity androot anoxia on growth and net Na+ and K+-accumulation in Zea mays grownin solution culture. Annals of Botany 61, 41–53.
elle, H.H., 2005. pH regulation in anoxic plants. Annals of Botany 96, 519–532.lowers, T.J., Colmer, T.D., 2008. Salinity tolerance in halophytes. New Phytologist
179, 945–963.hassemi, F., Jakeman, A.J., Nix, H.A., 1995. Salinisation of Land and Water Resources:
Human Causes, Extent, Management and Case Studies. University of New SouthWales Press, Sydney.
ibbs, J., Greenway, H., 2003. Mechanisms of anoxia tolerance in plants. I. Growth,
survival and anaerobic catabolism. Functional Plant Biology 30, 1–47.ong, X., Chao, L., Zhou, M., Hong, M., Luo, L., Wang, L., Ying, W., Jingwei, C., Songjie,G., Fashui, H., 2011. Oxidative damages of maize seedlings caused by exposureto a combination of potassium deficiency and salt stress. Plant and Soil 340,443–452.
perimental Botany 87 (2013) 69– 78
Greenway, H., Munns, R., 1980. Mechanisms of salt tolerance in non-halophytes.Annual Review of Plant Physiology 31, 149–190.
Ivashikina, N., Becker, D., Ache, P., Meyerhoff, O., Felle, H.H., Hedrich, R., 2001. K+
channel profile and electrical properties of Arabidopsis root hairs. FEBS Letters508, 463–469.
Jackson, M.B., Drew, M.C., 1984. Effects of flooding on growth and metabolism ofherbaceous plants. In: Kozlowski, T.T. (Ed.), Flooding and Plant Growth. Aca-demic Press, New York, pp. 47–128.
James, R.A., Blake, C., Byrt, C.S., Munns, R., 2011. Major genes for Na+ exclusion,Nax1 and Nax2 (wheat HKT1;4 and HKT1;5), decrease Na+ accumulation in breadwheat leaves under saline and waterlogged conditions. Journal of ExperimentalBotany, http://dx.doi.org/10.1093/jxb/err003.
Jenkins S., 2007. Ecophysiological principles governing the zonation of puccinel-lia (Puccinellia ciliata) and tall wheatgrass (Thinopyrum ponticum) on salinewaterlogged land in south-western Australia, School of Earth and GeographicalSciences. PhD Thesis, University of Western Australia, Perth.
Jenkins, S., Barrett-Lennard, E.G., Rengel, Z., 2010. Impacts of waterlogging and salin-ity on puccinellia (Puccinellia ciliata) and tall wheatgrass (Thinopyrum ponticum):zonation on saltland with a shallow water-table, plant growth, and Na+ and K+
concentrations in the leaves. Plant and Soil 329, 91–104.Kronzucker, H.J., Britto, D.T., 2011. Sodium transport in plants: a critical review. New
Phytologist 189, 54–81.Maathuis, F.J.M., Amtmann, A., 1999. K+ nutrition and Na+ toxicity: the basis of
cellular K+/Na+ ratios. Annals of Botany 84, 123–133.Munns, R., James, R., Islam, A., Colmer, T.D., 2011. Hordeum marinum-wheat
amphiploids maintain higher leaf K+:Na+ and suffer less leaf injury than wheatparents in saline conditions. Plant and Soil 348, 365–377.
Munns, R., Tester, M., 2008. Mechanisms of salinity tolerance. Annual Review ofPlant Biology 59, 651–681.
Munns, R., Wallace, P.A., Teakle, N.L., Colmer, T.D., 2010. Measuring soluble ion con-centrations (Na+, K+, Cl−) in salt-treated plants. In: Sunkar, R. (Ed.), Plant StressTolerance: Methods and Protocols. Springer, New York, 371–382.
Pang, J.Y., Newman, I.A.N., Mendham, N., Zhou, M., Shabala, S., 2006. Microelectrodeion and O2 fluxes measurements reveal differential sensitivity of barley roottissues to hypoxia. Plant, Cell & Environment 29, 1107–1121.
Plett, D.C., Møller, I.S., 2010. Na+ transport in plants: what we know and would liketo know. Plant, Cell & Environment 33, 612–626.
Ponnamperuma, F.N., 1984. Effects of flooding on soils. In: Kozlowski, T.T. (Ed.),Flooding and Plant Growth. Academic Press, New York, pp. 9–45.
Shabala, L., Ross, T., McMeekin, T., Shabala, S., 2006a. Non-invasive microelectrodeion flux measurements to study adaptive responses of microorganisms to theenvironment. FEMS Microbiology Reviews 30, 472–486.
Shabala, S., Cuin, T., 2008. Potassium transport and plant salt tolerance. PhysiologiaPlantarum 133, 651–669.
Shabala, S., Demidchik, V., Shabala, L., Cuin, T.A., Smith, S.J., Miller, A.J., Davies, J.M.,Newman, I.A., 2006b. Extracellular Ca2+ ameliorates NaCl-induced K+ loss fromArabidopsis root and leaf cells by controlling plasma membrane K+-permeablechannels. Plant Physiology 141, 1653–1665.
Shabala, S., Mackay, A., 2011. Ion transport in halophytes. Advances in BotanicalResearch 57, 151–199.
Shabala, S., Newman, I., 2000. Salinity effects on the activity of plasma membraneH+ and Ca2+ transporters in bean leaf mesophyll Masking role of the cell wall.Annals of Botany 85, 681–686.
Shabala, S., Shabala, L., 2002. Kinetics of net H+ Ca2+, K+, Na+, NH4+, and Cl- fluxes
associated with post-chilling recovery of plasma membrane transporters in Zeamays leaf and root tissues. Physiologia Plantarum 114, 47–56.
Shabala, S.N., Lew, R.R., 2002. Turgor regulation in osmotically stressed Arabidop-sis epidermal root cells. Direct support for the role of inorganic ion uptake asrevealed by concurrent flux and cell turgor measurements. Plant Physiology 129,290–299.
Smethurst, C.F., Garnett, T., Shabala, S., 2005. Nutritional and chlorophyll fluores-cence responses of lucerne (Medicago sativa) to waterlogging and subsequentrecovery. Plant and Soil 270, 31–45.
Song, J., Shi, G., Gao, B., Fan, H., Wang, B., 2011. Waterlogging and salin-ity effects on two Suaeda salsa populations. Physiologia Plantarum 141,343–351.
Stelzer, R., Lauchli, A., 1978. Salt- and flooding tolerance of Puccinellia peisonis III: dis-tribution and localization of ions in the plant. Zeitschrift Fur Pflanzenphysiologie88, 437–448.
Thomson, C.J., Atwell, B.J., Greenway, H., 1989. Response of wheat seedlings to lowO2 concentrations in nutrient solution II K+/Na+ selectivity of root tissues ofdifferent age. Journal of Experimental Botany 40, 993–999.
Trought, M.C.T., Drew, M.C., 1980. The development of waterlogging damage inyoung wheat plants in anaerobic solution cultures. Journal of ExperimentalBotany 31, 1573–1585.
Véry, A.A., Sentenac, H., 2003. Molecular mechanisms and regulation of K+ transportin higher plants. Annual Review of Plant Biology 54, 575–603.
Wang, C.M., Zhang, J.L., Liu, X.S., Li, Z., Wu, G.Q., Cai, J.Y., Flowers, T.J., Wang, S.M.,2009. Puccinellia tenuiflora maintains a low Na+ level under salinity by limitingunidirectional Na+ influx resulting in a high selectivity for K+ over Na+. Plant,
Cell & Environment 32, 486–496.Wiengweera, A., Greenway, H., 2004. Performance of seminal and nodal roots ofwheat in stagnant solution: K+ and P uptake and effects of increasing O2 partialpressures around the shoot on nodal root elongation. Journal of ExperimentalBotany 55, 2121–2129.
Copyright © 2022 FDOKUMEN




















