
Excavations at 41LK67 a Prehistoric Site in the Choke Canyon ...
Upload
khangminh22Category
view
2download
0
Studies -
Studies on the choke disease of grasses and. 14e-
causal organism, Enichloe typhina (Fr.) Tut.
A thesis presented by
Josephat James Njoroge, B.Sc.
In part fulfilment of the requirements for the
Ph.D. degree of the
University of London
December 1961
From: The Department of Botany and Plant Technology, Imperial College of Science and Technology, London, S.W.7.

-1.-
ABSTRACT
In cocksfoot, attempts to transmit Eachloe ty-Qhina. through the vegetative plant and flowers were unsuccessful, but on cut ends of flowering stems, spores germinated and hyphae grew down the stems. Conidia were liberated when a moist air current was passed over conidial stromata. The maxinni number of ascospores was discharged between 6 and 10 p.m.
The formation on infected plants of looping stromata, several stromata on the same tiller, and "double stromata" (one true and one false) are described. Weight and length of mans, and weight and germination of seed from diseased plants were reduced. Cocksfoot, among three host species examined, had most fungus-free tillers.
Cultivated on several liquid media, E.typhina grew best on corn meal/glucose peptone. Vigour of growth of different 4srIl^tes on this medium and on their hosts were unrelated. Production of coniala was best in glucose-peptone agar medium. Of three phosphate sources tested, K2HPO4 gave the greatest yield.
The pathogen was not deficient for growth factors, except for two isolates which were partially thiamine deficient, one .at 25°C. and the other at 10 and 8°C. The fungus grew well vegetatively with carbon sources of different complexity but yield of mycelium was poor with lactose, pectin and cellulose. Of the nitrogen courses tested, glutanic acid gave the greatest vegetative growth. Organic N-cources were better than inorganic. No evidence for nitrogen fixation was obtained. Rate of growth ny,a ultimate yield were separately influenced by pmperature. The greatest yield of mycelium was obtained at 18 C. Light had no effect on mycelial yield. Attempts to obtain the perithecial stage in artificial culture failed.
Conidia germinated best at 20°C. after 24 hours incubation in free water. Asoospores germinated best at 18° C. at 100% R.H. Stored under various conditions, conic9ia survived for less than 30 days and ascospores about 55 days.

-2 —
CONTENTS. Page
I Introduction. 5
II Review of Literature. 7
III Pathological studies. 27
A.Inoculation experiments. 27 (i) Introduction 27
1959 Experiments ilj Materials and general methods 28 iii); Inoculation through vegetative tillers 29 iv) Inoculation through tiller buds 30
1960/61 Experiments v) Inoculation through leaves 32 vi); Inoculation through flowers 32 vii)) Inoculation through flowering stems 37
B.Spore liberation and trapping. 44
C.Effect of the disease on the host. 52 i) Effect on choked tiller 52 ii) Effect on leaf sheath 55 iii) Effect on culms and seed 58
D.Distribution of mycelium in infected plants. 65
IV Physiological studies 71
A.Introduction. 71
B.Materials and general methods. 71 i) Materials 71
Methods 72
a Media 72 b Culture vessels 73
Methods of sterilization 73 d Stock solutions 7L. e Maintenance of stock cultures 75 f Method of inoculation 75 g Incubation 76 h Method of assessing growth 76

- 3 -
CONTENTS cont'd. Page
C. Preliminary experiments 78 i) Growth of E.typhina on different media 78 ii) Standardization of inocula and source 84 iii) Influence of phosphate and pH 87
D. Experiments with growth factors. 90
(i) Response of E.typhina to thiamine, biotin and yeast extract 90
(ii) Response of isolates of E.typhina from different grasses to thiamine in the culture medium. 102
(110 Investigatibn of thiamine requirement of E.typhlap isolated from lomerata, in cultivation at low temperature. 107
E. Carbon nutrition: Utilization of different sources of carbon. 113
F. Nitrogen nutrition. 120
i) Utilization of different sources of nitrogen 120 (ii) Growth of E.typhina in a medium lacking a
nitrogen source. 128
G. Influence of physical environmental factors. 132
i) Temperature 132 ii) Light and dark 137
H. Attempts to obtain the perithecial stage in 140 artificial culture.
i) Introduction 140 ii) On rice medium 140 iii) On cocksfoot stems 143 iv). On malt extract gelatine 144
I. Germination and viability of spores. 146 i) Effect of temperature on germination 147 ii), Effect of humidity on germination 153 iii), Longevity of conidia 156 iv) Longevity of ascospores 159

CONTENTS cont'd.
aim
V General discussion. 162
VI Summary. 183
VII Acknowledgements. 190
VIII Literature cited. 191

INTRODUCTION
Grass is recognized as an important crop in ley farming.
To be efficient, this system of farming needs a continuous supply
of grass seed. With increasing demand for seed, diseases which
reduce its yield or quality have assumed importance. One such
disease is choke, Epichloe typhine (Pers, ex which has
caused concern in Britain to growers of seed and plant breeders
(Kirby, 1958). At the Welsh Plant Breedirg institute, cocksfoot
(Dactylis glomerate) plants collected for breeding were fotrd to be
infected and rendered useless (Sampson, 1955). A batch of fescue
seed (Festuca rubra) imported to the United States of America from
Hungary was similarly found to be infected. When grown to maturity,
many cuims were choked off and many plants could not be used for
breeding (Wernham, 1942). More recently, the National Agrtcultural
Advisory Service has carried out a. survey of the disease on
cocksfoot plants grown for seed and found that the disease is
generally distributed throughout England and Wales and may cause
some reduction of yield of seed,
The disease is systemic with mycelium perennating in the
vegetative parts of the plant (Sampson, 1955). It may be present
for a season or more before giving rise to external symptoms.
This condition was described by Sampson (1955) as 'latent'. So

far, most attempts to transmit the disease by conidia and asco-
spores have failed. The role of conidia and ascospores in the life-
cycle of E.typhina has been investigated by the writer.
There is little irEormation in the literature on the
physiology of E.typhina. Although Vladimirskaya (1928) reported
that the optimum growth temperature was between 16 and 19°C., the
fungus has been grown at 25°C. by Lilly and Barnett (1949) and
Kirby (1958). As a first stage in studying the physiology of a
fungus, it is necessary to determine the conditions which affect its
growth. Experiments have therefore been carried out with a view
to establishing the basic nutritional requirements of E.typhina.
Endophytes of Loliva and Festuca which have shown
similarity to E.typhina in that they are systemic and are trans-
mitted by seed have been described by Sampson, (1954; 1951) and
Neill, (1942). A Lolium endophyte was isolated in the course of
the present study. An attempt has been mAde to compare it to
isolates of Epichloe typhina in culture in the hope of finding a
clue to its identity.

REVIEW QF LITERATURE
The choke disease of grasses is no newcomer to British
agriculture. The pathogen, Epichloe typhina, has long been known
here and over twenty grass hosts are known (Western and Cavett,
1959). The fungus was first recorded by Persoon in 1798 under
the name Sphaeria,typhina. It was transferred to the genus
Epichloe by Tulasne. In 1865, de Bary refuted the view that it
was only an epiphyte by showing the connection between mycelium
inside the plant andthe fruiting organs.
The disease stifles the development of the panicle,
"choking" it while it is still enclosed in the leaf-sheath and
generally preventing its emergence. Its effect is, however, not
the same on all grasses. In some, for example cocksfoot
( ractylis glomerata), the disease entirely inhibits the emergence-
of infected panicles from the sheath, so that no seed is formed.
In red fescue (Festuca rubra) in which infected panicles are not
prevented from emerging, all grades of infection may be found.
For instance, panicles may, by rapid growth entirely outpace in-
fection travelling up the stem and remain fertile. On the other
hand, where infection has continuously kept pace with host
development, infection is found in all parts of the flower, in-
cluding the seed (Butler and.Jomes, 1955).

The fungal stroma, which is formed above the third or the
fourth node of the culm, is seen at about the time of flowering of
the grass host. The stroma is cylindrical, 1-10 cm. long and
2-5 mm. in diameter, its size varying with the grass host. The
lamina of the leaf upon which the stroma forms may extend beyond
it. The stromq is first white, becoming deep yellow with the
formation of the perithecia. It dries and flakes off in the late
summer or may be attacked by other fungi. Sometimes the stroma
occurs only on part of the panicle (Benedict, 1929; Sampson, 1955).
World Distribution.
The disease is widely distributed throughout Northern
Europe and accounts of outbreaks of varying severity have been
noted from countries in this region. In 1925 it was unuswirly
prevalent in Denmark (Gram and Thompsen, 1927). It was noted in
Sweden by Erikson (1912) and on fodder grasses in the same country
by Wahlin (1949). There are reports of choke from Esthot►ia, where
it occurred in an epidemic form (Lepik, 1951) and Russia (Eakin,
1924; Vladinirskayal 1928). In Germany, Nolz (1924) reported the
poisoning of geese feeding on infected grass. In Great Britain
the disease is of general distribution. It has been reported from
Vales (Sampson, 1°35); Scotland (Dennis and Foister, 1942) and on
temporary pastures in Kent (Carruthers, 1905). The National

Agricultural Advisory Service surveys of choke, concerned only
with. Aberystwyth strains of cocksfoot indicate that the disease
occurs with eval intensity in all provinces of England and Wales.
Outside Europe the disease has little significance,. It
is widely distributed in North America but is of less economic
importance than in Europe (Benedict, 1929). The disease is un-
known in New Zealand (Neill, 1942).
The host.
The following list of host species shows that the disease
attacks many genera, all belonging to the Gramineae. With the
exception of a report of the disease on wheat and rye (Vladinirskaya,
1928) all the host species are non-cereal grasses. The disease
has not been found on Lolium species.
Table 1.
Species of Gramineae on which choke has been found
(Kirby,- :1959).
SPECIES AUTHORITY
Agrovron caninum V1ndimirskaya, 1928
A. junceum
Agrostis vulgaris (=A.tenuis) Vladimirskayal 1928; Ingold, 1948
A. alba
A. stolonifera

-10-
Table 1. (Cont
SPECIES
Aira. oaespitosa
A• flex-wpm.
Alopecurus pratensis
A. geniculatua
Anthoxanthum odoratum
AUTHORITY
Dennis & Foister, 1942
Vladimirskaya
; Sampson, 1929
tl
Brachypoditm sylvaticum Vladimirskaya; Sampson
B. pinnatum
31romus spp. Eriksson, /930
Calamagrostis arundinaceae Vladimiratkaya
O. baltica.
C. lelleriana ft
Daotylis glomerata Sampson; Ingold
Festuca ovina Sampson: Vladimirskaya
F. rubra
Glyceria nervata Benedict 1929
Hot cus 1Rratun Sampson; Vladimirskaya
H. mollis Sampson; Vladimirskaya; Ingold
Koeleria cristata Sampson
Minium spp. Eriksson
Holinia caerulea Dennis & Foister

-11-
Table 1. (Cont d.)
SPECIES
AUTHORITY
Phleum pratensis
Vladimirskaya; Sampson
Poaannua
Vladimirskaya
Poa bulbosa
Sampson; Tulasne, 1865
P. nemoralis Vladimirskaya; Tulasne
P. pratensis
Vladimirskaya
P. trivialis
Sampson
Schedononis spp. Eriksson
Triticum vulgare
Vladimirskaya
Secale cereale
Vlaaimirskaya
Only a few of these species are of any economic importance
(Western and Cavett, 1959).
The fungus.
Sampson (1953) showed that E.typhina mycelium could be
detected by removing strips from the pith of infected plants and
staining them with cotton blue or gentian. violet. The hyphae are
1-2',L} in diameter, sparsely septate and run for long distances
without branching. The fungus develops most of the year as an
intercellular parasite, occasionally emerging between epidermal cells.
Fungal hyphae can be demonstrated in any season in longitudinal

sections of tiller buds, leaf-b1Ddes, leaf-sheaths, root-stocks,
creeping stems and bulbils. In F.rubra and H.mollis, Sampson
(1955) also found mycelium in the stolons, rhizomes and in the thick
scale leaves of 7.bulbosa. Epichloe typhina mycelium has not
been found in the roots of infected plants.
Sampson (1955) describes the fructification of the fungus.
The development of the conidial stroma coincides with the period of
panicle exsertion in healthy individuals. In preparation for
fruiting, the mycelium grows out from the leaves of the young
tillers in early spring and during the first week in May (in Wales).
A delicate felt of mycelium may be seen covering the youngest leaf
of potentially fertile tillers. It appears first on the early
grasses such as .tlopecurus pratensis, later on Dactylis glomerata
and Festuca rubra; and last of all on Fhleum pratense which is a
late flowering species. A section of the tillers shows that the a
leaves are united by/ complete weft of mycelium which soon develops
into a stromatic sheath, trapping the young infloresence and causing
its degeneration.
In Glyceria nervata the stromn can be seen in the covering
leaf sheath two days before it emerges. Elongation of the inter-
node finally freen the stroma and mnrks the end of the growth of
that particular culm. Internodes hidden in the terminal leaf do
not develop (Benedict, 1929).

-15-
The stroma at first produces conidia which are born singly
on narrow, flask-shaped conidiophores. The coniain are hyaline,
cylindrical or narrow-oval, measuring 5-9 by 1-2 A (retch, 1957).
The conidia germinate readily (Brefeld, 1891). Benedict (1929)
showed that they were fully formed and capable of germination as
soon as the stroma emerged from the leaf-sheath.
The conidial stage lasts 5-4 weeks after which time the
stomata increase in thickness, change to yellow and then orange,
and develop numerous immersed perithecia with papillate ostioles.
At first the perithecia are free but later become confluent. The
perithecia are oval, 0.5 to 0.6 mm. high and 0.25 mm. in diameter
and with an obtuse apex. The asci are cylindrical, very long and
capitate; they contain eight hyaline, filiform, many-celled
ascospores 1.5 - 2.0 gin diameter and septate at intervals of
8-12 ;4. They do not divide into spore parts (Petch, 1957). The
length of ascospores differs with host species and ranges from
190,4 in Dactylis glomerata to 52 )4 in !grostis tenuis (Ingold,
1948).
Western and Cavett (1959) secured the production of
conidia by passing a current of air containing atomized water over
the stromata. The counts of conidia were most difficult to make

owing to their small size and similarity to other bodies trapped
on the slide. Their results derived from the trapping of spores
in the field were completely confirmed by those based on
observations on the maturation of stromata and healthy panicles,
which extended from the first appearance of the disease and the
emergence of the flowering head up to seed harvesting time when
ascospore production was at its peak, From the results atained,
it appeared that if stigma infection occurred at all, it could
only be by means of the conidia and not by the ascospores, since
these were not produced in any nunber until well after the
pollination period (Western and Cavett, 1959).
The discharge of ascospores from perithecia has been
described by a_ numberof workers (Sowerby, 1803; Tulasne, IBM;
Ingold, 1948). In describing this phenomenon, Sowerby states
that the spores are continually being extrvded through ostioles
tith a sort of spontaneous motion, looking in the sun like sparks
of fire". Ingold (1948) has made a detailed study of their
discharge. If spore discharge is not too rapid, their extrusion
from a single ascus can be seen. The eight ascospores are dis-
charged in less than a second. When activity is high, the rate of
discharge from a single stroma may reach 2,000 to 10,000 spores per
minute. There tends to be a daily periodicity with a minimum in

the morning and amAxitum in the afternoon or evening. The rate
fluctuates considerably over short intervals but when the trans-
piration stream is cut, spore liberation falls rapidly.
The fungus in culture.
E.typhinahas been isolated by taking long strips of pith
with sterile forceps and laying them on nutrient media (Sampson,
1935). Kirby (1958) isolated the fungus from leaves and growing
apices. Be removed a piece of leaf 1-2 in. long, from an infected
plant and immersed it in 0.1 percent mercuric chloride solution for
4 minutes. After washing in sterile water he laid it on nutrient
agar on which the fungus developed in 8-12 days. Growth was also
obtained from apices dissected outs sterilized and laid on agar.
It was not found possible to isolate the fungus from infected seed.
The fungus was grown on potato dextrose agar and other
media and on each a circular colony of closely-knit mycelium was
obtained (Kirby, 1958). Viewed through the agar, a. yellow
colouration was seen. Isolates from different host species showed
morphological differences. Those from Fhleum pratense and Thctylis
glon,wata were more compect and raised from the agar than those
from igrostis stolonifera and Bblcus. mollis. Isolates from the last
two host species showed a more diffuse, low-growing habit. These
characteristics could not be shown to have any significance with

regard to the host specificity of strains of the fungus until
cross-inoculation experiments could be performed. The cultures
produced_ conidia profusely but no perithecia were formed.
The vitamin requirement of E.tvphinahas been investigated
by Lilly and Bnrnett (1949). The response of the fungus to the
addition of thiamine, pyridoxine, inositol and biotin was tested.
The addition of thiamine to the medium led to increased growth.
Comparison of the growth rates of aifferent Isolates of Estyphina.
Sampson (1955) stated that there was definite evidence
that the disease is not biologically identical upon all hosts.
Kirby (1958) observed that there was a difference in the amount of
choking of different species, i.e. in the amount of fungal material
formed on the plant. He designed an experiment to see if this
difference in behaviour could be related to the difference in the
growth rates of the isolates. He found that the growth curves
for the isolates within each species were very similar, and the
mean growth curves of isolates from different species showed only
small. differences. However, the period of initial lag as shown by
the growth curve was greater for the isolates from H.mollis and
A.stolonifera than for those from Etszatense and I4glomerata.
There was no relationship between the growth of the fungus in the
plant and its rate of growth on artificial media. The isolates

from clones which had produced unaffected or partially affected
panicles gave growth curves very similar to isolates from plants
with typical stromata.
Similar differences in cultural characteristics are
found in the smut fungi (Fischer and Holton, 1957), In these
fungi, races differing in rate and type of growth, colour and other
features may also 410w aifferent pathogenic speciPa4zationt
Whether or not the differences in cultural characteristics of the
isolates of p.typhina. indicate any differences in pathogenie
specialization.an not yet be determined,
The transmission of the disease.
Seed transmission has been investigated by Sampson (1935)
and Werth= (1942) but has been established with certainty only in
F.rnbxl., though, according to Sampson (1955), it probably occurs in
F.ovina and 'D bulbosa Sampson (1955) showed that the fungus
enters seed from inside the stem in F,rUbra and not from the outside.
Wernham (1942) demonstrated the seed-borne nature of the disease for
P.rubra seed imported from Hungary for planting in airfields, fair-
ways and playing surfaces. Some of the plants raised from the
imported seed showed the symptoms of choke and these were burned,
The possibility of seed contnnination as a means of trans-
mission in cocksfoot was studied by Western and Cavettp (1959).

-18-
They threshed seed from sheaves to which a considerable amount of
perithecial material had been added but recovered no viable
ascospores from the seed. No E.typhina mycelium was observed in
the seedlings raised from this seed. They also washed seed
harvested from a very heavily infected field. They obtained
asesspores from the dry and mature seed but these spores were dead.
In each case enough seed was saved for planting in the field but
no stromata developed at any time during the next three years.
Western and Cavett (1959) further tested the possibility
of seed transmission by the use of an organo-mercurial fungicide
and warm water disinfection of seed harvested from a very heavily
infected cocksfoot crop. Drills of the various treatments and
controls were sawn and kept under observation for 5 years but no
E.typhina developed in any treatment or in the controls. They
also grew seed from partially emerged heads and searched the
resulting plants for mycelium but with negative results. From
the results of these experiments and observations in the fields
they concluded that transmission by this means was improbable in
cocksfoot.
To test the possibility of seedling infection, Western
and Cavett (1959) attempted to induce penetration following
germination of the seed. They germinated seeds on active cultures

of E.typhina and used other methods to bring inoculun into close
contact with the young coleoptiles and other parts of the
germinating embryo. No plant from these experiments produced
stromata in the field during the following three years, but a
microscopical examination of inoculated seedlings showed that a
vigorous growth of hyphae occurred over the basal parts of the
young shoot and also on the root, but no actual invasion of the
host tissues was observed. From these results, and others from
similar experiments, they concluded that the seedling was unlikely
to be the source of initial infection.
Proof that the disease is transmitted by vegetative
propagation of the host was obtained repeatedly by Sampson (1933.)
She showed that this took place in Dactylis glomerata, Phleun
pratense, Alopecurus 2ratensis„ Jgrostis stolonifera, Festuca rubra,
Koeleria cristata, Holcuslana.tin and Festuca ovina. The most
striking example of this method of distribution was that of the
single plant of D.glomerata which was successfully broken up to
form 287 root divisions, only one of which failed to produce the
disease in the following season. Unfortunately, this plant was
not grown for a second year since the "latent condition" of the
disease was not realized at the time and, therefore, it was not
known whether the plant had really escaped infection,

-20-
Effect of the disease on the host.
The effect of choke on the number of vegetative tillers
was investigated by the National Agricultural Advisory Service in
surveying cocksfoot crops for choke (Large, 1952, 1954). It
appeared that plants with choked tillers tended to show a. more
luxurious growth than those without choke but whether this was due
to increase in length and vigour or number of tillers was not
established. The number of vegetativetillemplotted against the
percentage of choked tillers showed no relationship. Their
number was so greatly affected by the management of the crops, as
regards date of sowing, autumn burning off, grazing etc., that the
effect of choke if any, on the production of herbage was almost
completely masked. The survey also indicated that the percentage
of choked tillers in a field may be a fair measure of the reduction
caused by choke in the number of seed heads to be harvested.
Whatever the level of choke, it was found that the mean total
number of tillers, (choked flowering), per foot run of drill
remained approximately constant. There was therefore, little or
no compensatory effect tending to increase the number of flowering
heads when some tillers were choked and little or no systemic
effect tending to reduce the number. In the absence of evidence
to show that the amount of seed per head was reduced, it was con-

-21-
eluded that the reduction in yield was proportional to the number of
choked tillers in the crop.
The incidence and significance of infection of
Agrostis tonuis by Epichloe typhina has been studied by Bradshaw
(1959). L. comparison of the growth of infected and non-infected
plants showed that infection, in preventing infloresence production,
caused a considerable increase in tiller density and vegetative
height without any other deleterious effects. These effects are
of advantage to plants growing under conditions where vegetative
growth is important, He concluded that infection of Agrostis spp.
by Epichloe typhina must be considered to be not an example of
parasitism but of a curious form of symbiosis.
Development of panicles and infloresences.
In Festuca rubra mycelium has been found in all parts of
the flower in infected plants. This mycelium penetrates the
rachilla, glunes and pales, the filaments and anthers of the
stamens as well as the various tissues of the ovary including the
branches of the style (Sampson, 1933). Sometimes the stamens may
be so damaged that pollen does not form, or ovaries may be so
riddled with mycelium as to be rendered abortive. On the other
hand, some plants may give a yield of viable seed almost equal to
that of normal uninfected plants.

-22 —
In the very young ovary, Sampson (1933) found that the
mycelium grows between the cells of the integuments and nucellus
but later encroaches on the embryo sac. In half-developed ovaries
and in mature grains the mycelium forms a. loose and irregular net-
work outside the aleurone layer, round the embryo and it may
penetrate the endosperm tissue. The results obtained from
examination of ovaries confirmed the conclusions based on the
examination of pith scrap.,12:u, that the panicles exserted by
infected plants of cocksfoot do not normally carry mycelium.
Benedict (1929) studying the conidial stroma, of
E.typhina under green-house conditions found that the fungus could
be made to appear in the conidial form in mid-winter. He
concluded that the time of appearance of the stroma was dependent
upon the stage of development of the host ratherlthealupon a
seasonal condition. He suggested that besides the host parasite
relationship, there were probably other contributing factors,
physiological or otherwise, which determined the conidial phase.
The development of infloresences in relation to flowering
has been studied by Kirby (1958). In infected plants, he found
that the vegetative shoot apex was always normal but malformation
due to the disease appeared soon after floral initiation, followed
by the emergence of the fungal hyphae which bound the panicle and

-25—
leaves into a stroma. By manipulating the environment, he found
that the flowering behaviour of the grasses was associated with
the change from vegetative to floral condition. The development
of infloresences of Phleum pratense, Dactylis Flomerata, Holcus
mollis and Agrostis stolonifera infected with E.typhina was studied.
All the species tested flowered much earlier than normally if
given a long day treatment in early spring (March and April).
Seasonal appearance of the disease.
Sampson (1933) made observations on the seasonal
appearance of the fungus on cocksfoot. A population of 863 plants
derived from various sources was observed during 1927 and 1928.
Of these, 565 showed the disease both years, 22 were visibly
infected in 1928 but not in 1927, while 25 were recorded as infected
in 1927 but outwardly showed no infection in 1928. The degree
of infection as indicated by the percentage of fertile tillers
carrying perithecia gave a range of variation from 0.7 to 100 percent.
From observations on single plants she concluded that there was a
tendency towards complete infection in course of time in cocksfoot.
The endophytes of Lolium spp. and New Zealand Festuca spp.
Interest in the endophytic fungus of Loliun dates back to
1898 when Vogl recorded mycelium in the seed ofli.terruleentum.

Freeman (1905) gave a detailed description of the progress of the
mycelium in the young ovule, the fruit and the embryos He also
expressed belief in the existence of strains lacking the fungus.
Hanig (1907) grew infected and healthy plants for several
generations and found that the disease was usually seed-borne.
Infected plants usunlly produced infected progeny but occasionally
seed from infected plants did not contain the fungus and these
could give rise to fungus-free populations. Further experiments
confirmed the existence of fungus-free races of L.perenne and
indicated that they may arise from fungus-infected plants. No
knowledge as to how and when members of the genus Loliun become
infected by this fungus was obtained.
Sampson (1934, 1937) has reported two endophytes of
Loliun. She cultured the first on egg medium after attempts to
isolate it on other media had failed. The growth consisted of
more or less spherical cushions which finally joined together to
form a raised mass about 2 mm. in diameter, the surface of which
was covered with short, erect aerial hyphae. lifter initial
isolation on egg medium, this fungus was grown on a variety of
media including potato discs, agar prepared with Ithopis solution,
egg, liver extract, potato dextrose, oatmeal and gelatine con-
taining egg.

Sampson (1937) also described the second endophyte of
Lolium. This fungus did not fructify in the plant and caused no
pathological symptoms. It was transmitted by vegetative
propagation and by seed. To this extent it resembled the first
Lolium fungus. However its nycelial characteristics, the fact
that it did not form a thick zone of mycelium in the seed and that
it grew readily on all kinds of media, distinguished it from the
first endophyte.
Mycelium was usually present in abundance in pith
scrapings made from plants carrying the second endophyte but it
stained faintly with cotton blue.. This fungus was easily
cultured by placing strips of pith from infected plants on nutrient
agar, on which white, rather fluffy aerial mycelium was produced.
On natural media such as oatmeal agar, the fungus provided good
crops of microconidia. The first Lolium endophyte produced no
spores.
Investigation of the Lolium endophyte had been undertaken
in the hope of finding a link between this organism and Epichloe
typhina but culture work brought to light points of difference
rattier than mnrks of similarity.
Neill (1942) found that some New Zealand strains of
Festuca arundinacea and F.elatior normally contained an endophyte

-26-.
whose transmission through seed was similar to that of Epichloe
typhina. The fungus was found in leaves in the form of inter-
cellular hyphae 1.5 - 2.5;14 in diameter. It formed a network
over the aleurone layer in seed and proliferated in the pith where
many hyphae were coarse and vesicular. Plants containing the
fungus were healthy and no sign of it was seen on the outside.
The fungus was cultured from surface-sterilized seed.
It could also be obtained in culture from seedlings grown from in-
fected seed as well as from the pith of young flowering stems. On
most media, the endophyte appeared as a white, sub-lamons mat which
spread slowly. The conidia were hyaline, born singly on sterigmata,
which measured 12 - 25 by 1.5 7.; at the base, and arose
perpendicularly at intervals from trailing aerial hyphae. The
conidia were somewhat irregularly elliptical and measured 6 .0* 8
by 2 - 3 :1.1. In dried cultures the conidia tended to be curved,
with attenuated extremities and attained a length of 11 u. The
conidial apparatus was similar to that for Epichloe typhina, but
the conidia were rather larger. The New Zealand Festuca
vndophyte did not produce asci or prevent flowering. Endophyte-
free plants were obtained from seed infected with this fungus when
it was stored for twelve'or more months before being germinated.

III PATHOLOGICAL STUDIES.
A. INOCULATION MCPERDIENTS.
(i) Introduction
The role of conidia and ascospores in the transmission of
choke has been investigated by Sampson (1955), Benedict (1929),
Vladimirskaya (1928) and Western and Cavett (1959). Although
most attempts by these workers to transmit the disease by means of
spores have failed, Vladimirskaya (1928) claimed successful
inoculation of tiller buds, but she did not say whether this
resulted in the production of stromata. Sampson (1955) has
pointed out that apparently healthy plants sometimes contain
Epichloe typhina mycelium. It is possible that Vladimirskaya was
working with plants containing such mycelium. The only clear
demonstration of the production of stromata by artificially
inoculated plants is that by Western and Cavett (1959). They
inoculated cocksfoot (Lactylis glomerata) flowering stems with
conidia and ascospores which they applied to the cut ends under
conditions of high humidity.
Using a large number of plants, attempts have been made,
in the course of this investigation to obtain infection artificially
by introducing spores and mycelium through vegetative and floral
parts.

-28-
1959 Experiments
(ii) Materials and general methods.
Most of the inoculation experiments were carried out at
Silwood Park, on cocksfoot plants raised from seed which was sown
in pots in the greenhouse in April 1959. In June, some of the
seedlings were transplanted into other pots in the greenhouse.
Others were planted at 2 ft. by 2 ft. intervals in the Walled
Garden.
A culture of Epichloe typhina was obtained by laying
strips of pith from choked cocksfoot plants on potato dextrose agar
slopes. The strips were first surface-sterilized in 0.1 per cent.
mercuric chloride solution for 5-4 minutes and then washed in
sterile water before laying them on the medium (Bisby, 1953).
Cultures on potato dextrose agar sporulated after 7 days
incubation in a 23°C. incubator room. When a spore suspension
was required, a few ml. of sterile water were added to such a
slope culture and the surface of the agar was rubbed gently with
a sterile inoculating needle to loosen the spores.
Most of the inoculated plants were kept under observation
for two years to see if any stromata would develop. In some
plants, pith scrapings were made five months after inoculation .
The scrapings were stained with cotton blue in lactophenol at room
temperature and searched for mycelium.

-29-
(iii) Inoculation through vegetative tillers.
a) Spraying cut back stems with spore suspensions
To test the possibility of infection through vegetative
tillers, 48 stems from four of the potted cocksfoot plants were cut
back, leaving about 4 inches of stubble. The cut ends were
sprayed with conidial suspensions from cultures, by means of an
atomizer. These stems were inoculated in August 1959. In July
1960, 44 more stems from cocksfoot plants raised from seed in 1959,
were inoculated by this method, with ascospores obtained from
choked tillers of cocksfoot. The inoculated plants were kept in
the inoculation chamber for 24 hours after inoculation. The
chamber consisted of a cage, standing in a shallow layer of water
and covered on all sides and the top with a polythene sheet, so that
the plants were in a virtually saturated atmosphere for this period.
b) Injecting spore suspensions into stems by means of a hypodermic syringe.
An attempt was made to introduce spores into the pith by
means of a "Summit" hypodermic syringe and needle (1 inch; 16 gauge).
Eight one-month old cocksfoot seedlings which had been transplanted to
the Walled Garden were inoculated by this method. Eight other
plants in an adjacent row were left uninoculated to provide controls.
The plants were first inoculated on 27.6.59. Inoculation of the
same plants was repeated every fourteen days for 8 weeks.

-50.4
(iv) Inoculation through tiller buds.
To test the possibility of infection through tiller buds)
ascospores were washed into the leaf-sheaths of cocksfoot plants
in August 1959. The ascospores were collected by the method
described by Ingold (1948) from choked cocksfoot tillers and were
washed off the slides into the leaf-sheaths by means of a wash-
bottle. Ascospores tended to stick to the slides) but if they
were collected in large numberS so that they formed several layers
on the slide) some were easily transferred to the leaf sheaths by
the water jet. Twenty plants were inoculated in this manner.
Results.
Table
Methods employed to inocul7Ae Dactylis glomerata plants with
Epichloe typhina in 1959/60:' time of inoculation and number
of stems or plants inoculated.
Method of inoculation Tine Number of Number of Stems. , plants.
Spraying vegetative tillers with
conidial suspension August,1959 b- ascospores July$1960
Injecting conidial suspensions into vege- tative tillers with the June - hypodermic syringe July$1959
8
Washing ascospores into leaf sheaths July11959
20

-51-
The plants in all these experiments were observed over
periods up to two years but no stromata were seem Pith
scrapings were made from some of the inoculated plants but
no mycelium was revealed. All the plants produced healthy flowers.
No stromata were observed on healthy cocksfoot plants grown along-
side infected plants of the same species for Lvo years.
The results of the above experiments are in agreement
with those of Western and Cavett (1959) who carried out many
similar experiments. They sprayed cocksfoot plants with spore
suspensions at 5-day intervals from June onwards. They also grew
seedlings and plants to various vegetative stages in pots, into
which were inserted stromata of E.typhina (conidial and
ascigerous) in tubes of water. These plants were kept for 6 days
at high h-midity during which spoke discharge was heavy. Further,
tiller buds were inoculated with spores and mycelium from pure
cultures. Both surface and hypodermic inoculations were made..
All these plants were grown in the field for 5 years but in none
were stromata produced.
1960-1961 Experiments
In 1960 and 1961, the possibility of infection of cocks-
foot plants through leaves, flowers and flowering stems was
investigated by the writer.

-52-
Inoculation through leaves.
Four leaves were obtained from 5 months old cocksfoot
plants grown in pots. From these leaves eight 1" long pieces
were cut, sterilized in 0.1,5mercuric chloride solution and then
washed in sterile water. The pieces of leaves were then placed in
pairs on moist filter paper in sterile Petri dishes. Four of
these pieces mere inoculated in August 1961 with a suspension,
, a mainly of conidial prepared from a glucoseaeegnxton slope culture.
Two drops were placed on each piece of leaf, one on either end.
The other four pieces were left uninooulated. All were incubated
at 20°C. in the anrk.
The leaf tissues were exc.-lined for growth and penetration
of the fungus after 24 hours, for the first two pieces and after
48 hours for the other two. Before examining the pieces of
leaves were cleared by the method described by McMeekin (1960).
The pieces of leaves were first immersed in 95% ethyl alcohol to
remove chlorophyll and wax. They were further cleared with cold,
5% KOH, stained in cotton blue and mounted in lactophenol. No
hyphal penetration of the leaf tissue was detected.
(vi) Inoculation through flowers
The development of the conidial strona in the host plant
coincides with the period of panicle exsertion in healthy

-55-
Laviduals (Sampson, 1955). The flowers would therefore appear
to be a possible path for the entry of the fungus into a healthy
plant. Diehl (1950) in his work on the related fungi, the
Balansiae, successfully inoculated stigmas of Conohruz echinatus L.
with conidia of Balansia obtecta. The parasitism of Epichloe
typhina is sinilar in many respects to that of the smuts in which
flower infection occurs with considerable frequency (Western and
Cavett, 1959). The possibility of seed contamination or of
flower infection as a means of transmission was therefore tested.
Two methods were employed to inoculate cocksfoot
flowers:
1. Inflorescences were dusted with conidial stromata obtained
from choked cocksfoot plants in the field. After inoculation,
each tiller was staked and labelled. Pnrlier, it had been
Observed in the laboratory that if a conidial stroma was rubbed
gently on a slide a deposit of conidia was obtained. Eighty-one
cocksfoot flowers at different stages of development were inoculated
by this method. Inoculation was carried out from June onwards in
1960.
2. Some infloresences were inoculated by the partial vacuum
method (Moore, 1956) (see Fig. 1). Az described by Cherewick and
Popp (1950), apiece of sponge rubber was inserted in the rubber
stopper which secured the panicles in the inoculating cylinder.

IS*
A
-34-
(After Moore, 1936). The apparatus used for partial- vacuum inoculation of flowers.
The apparatus comprises:- A. A support. The lower portion of the support Is made of iron
rod to which are welded an iron step to aid in pushing the rod into the ground and an old apparatus clamp to support the inoculum flask.The upper portion of the support is made of wood.
B. A flask of inoculum. C. An inoculating chamber made from a large test tube.
The lower end is fitted with a rubber bung as described in the text. It is connected through a length of rubber tube and a pinch cock with the inoculum flask A length of small bore rubber tubing connects the top of the chamber to the vacuum pump D.

-55-
Tnis prevented damage to the necks of the flower-stalks. After
inserting the flower heads into the cylinder, the conidial
suspension was drawn into the cylinder by way of a small rubber
tube attached to the conical flask B which served as a reservoir.
L hand pump D, provided the necessary suction. 7:hen the
incloresences enclosed in the cylinder were completely submerged
in the spore suspension, the pinch cock in the tube supplying the
inoculum was closed and twelve rapid strokes of the pump were
given to create a partial vacuum. Fifty-three cocksfoot
infloresences were inoculated this way.
After the seed had set, the inoculated seed heads were
collected and stored in paper packets. Some of the seed was
later sown in pots for observation. Other seed was examined for
mycelium by the embryo test method described by Simonds (1946).
Grains were soaked overnight in a 1 °sodium hydroxide
Solution. This caused the seed to swell and rupture. The
embryos floated off and were separated by means of an inoculating
wire loop into a snnll beaker where they were washed with water.
They were then boiled in lactophenol until they became transparent.
This took about 50 minutes. The embryos were then stained in
cold cotton blue in lactophenol for 3-4 minutes, rinsed in lacto-
phenol to remove excess stain and placed on glass slides for

_56...
examination under the microscope. No mycelium was detected in
any of the seed or seedlings.
The plants raised from the seed from plants inoculated
in this way showed no infection.
Discussion.
These results are in agreement with those of Western and
Cavett (1959) who tested the possibility of flower infection or
seed contamination but without success. In addition to the
vacuum method (Noore, 1956) they appliedl conidia and ascospores
directly to the stigmas by various methods. Some flowers were
emasculated and bagged, inoculated with conidia when the stigmas
were exserted and pollinated the next day. Others were pollinated
and later inoculated while some were dusted with conidia on a dry
brush before stigmas were exserted. They placed humidity tubes
containing saturated cotton wool over the inoculated heads to
prevent premature drying out of the stigmas. Samples of material
inoculated in these ways were examined under the microscope to
trace the development or fate of the conidia deposited on the
stigmas. They found that hyphae grew superficially on stigmas but
no penetration of the ovule was observed. Seeds from these
experiments were drilled in the field but no Epichloe typhina
developed in the three succeeding harvest years.

Incculation through flowering stems.
a) Introduction
Since 1..typhinia mycelium travels rapidly up the pith of
the flowering stem before stroaata formation, the possibility of
its passage in the reverse 9irection was investigated by Western
and Cavett (1959). They found that under conditions of high
humidity conidia and ascospores germinated on cut surfaces of
cocksfoot and the resulting hyphae entered the pith cavity and
grew down the stem. Hyphae grew in 6 of 28 stems inoculated with
conidia. They also carried out inoculations on a field scale by
arranging jars of ripe perithecial stromata along drills of
seeding cocksfoot, some of which were cut back as in normal
harvesting, leaving about 6 inches of stubble. Cut surfaces were
lightly brushed with stromata. Adjoining rows were untreated and
used as controls. These inoculations were made in July 1953 and
in September the whole area was cut back as a normal cultural
measure. In 1954, they obtained infection in the inoculated rows
ranging from 0 4-- 10%. The distribution of stromata in the crop
was similar to that found in a. natural attack, i.e. single or
small groups of 2-5 stromata, well scattered along the rows.
This result was confirmed in 1957 on potted plants after
unsuccessful attempts to do so in plots.

-38-
Inoculations have here been carried out in 1960 with
view to confirming the entry of the fungus through flowering stems
of plants growing in pots and in drills in the 'Walled Garden.
b) Materials and method.
Flowering stems of cocksfoot plants grown from seed were
cut at differing levels (5-10 inches) and freshly collected dry
ascospores were applied to the cut surfaces. Inoculation was
first carried out on potted plants in a conservatory in July 1960.
Ascospores were collected by the method described by Ingold (1948)
on clean slides on which they formed a thick cover. A srinll
brush was used to transfer them to the cut surfaces of the stems.
After inoculation the plants were left in the inoculation chamber
for 48 hours. Uninoculated plants which were used as control
were also cut back and left in the inoculation chamber for 48 hours.
c) Results
The first plant which was inoculated had 14 stems, 7 of
which were examined by taking pith scrapings. Of these 6 were
found to contain mycelium 15 days after inoculation.
Examination 3 days after infection showed that the asco-
spores had germinated on the cut surface but there was not much
penetration. A hypha 574A long was observed.

After 9 days the ascospores were found. to have produced
conidia on the cut surface. One stem was cut near ground level
and pith scrapings made at 1 inch intervals. Mycelium was found
4 inches below the cut end and beyond a node through which the
mycelium must have passed. The node was sectioned using the
freezing microtome and mycelium was found on either side of it.
Another stem was cut from the same plant 15 days after
inoculation. Mycelium was traced 664 inches below the cut end.
Again in this stem mycelium had passed through a node which was
sectioned and mycelium found on both sides. In the =inoculated
plant 6 out of 12 stems were searched for mycelium but none was
found. These plants were grown further to see if any stromata
would develop. Two more potted plants were cut back in the
conservatory, one was inoculated with conidia from culture and
the other left uninoculated and used as control. Mycelium was
found in 5 out of 12 stems of the inoculated plant. None was
found in the uninoculated plant.
Four plants with a total of 89 stems were inoculated in
August 1960 under field conditions. Two of these were inoculated
with conidia while the other two were inoculated with ascospores.
An adjacent row was left uninoculated and used as control. No
stromata were produced on these plants in 1961•

masses of asccspores.
X 640.
Plate. I. E.typhina ascospores on a cut D.glomerata stem after inoculation.

Cornidia.
X 640.
Plate 2. Etyphina. ascospores germinating and producing conidia on a cut D.giomerata. stem after inoculation.

hyphal branch
hypha
X 640
Piate.3. E. typhina hypha growing down the pith of D. glomerate. stem after inoculation.

Discussion.
The results of these experiments were interesting in
that they were the only ones in which growth and penetration of the
pith of healthy flowering plants by hyphae resulting from artificially
placed spores was obServed. In this respect, the results are in
agreement with those of Western and Cavett (1959), However, none
of the plants produced any stromata. The potted plants had
become pot-;bound, They all made poor growth and no flowers were
produced by the uninoculated ones, These plants could not be
looked after properly because the writer had to leave Silwood to
work at South Kensington,
On the other hand, the control and the inoculated plants
in the Walled Garden grew luxuriantly and flowered heavilyr but
produced no stromata. Western and Cavett (1959) obtained stromata
from artificially inoculated plants in the field in 1954, but
attempts to do so in 1957 were unsuccessful, The most likely
explanation for the failure of the inoculations in the field
described here would seem to be that the environmental conditions
obtaining at the time they were made were not favourable for the
germination of the ascospores.

B. SPORE LIBERATION AND TRAPPING.
(1) Introduction
Ingold (1948) studied the water relations of spore dis-
charge in Epichloe typhina. He found that ascospore discharge
was not limited to dnup periods when the strona was wetted by
rain or dew. Instead the fungus, during periods of active dis-
charge, obtained its water from the host cells and, therefore,
indirectly from the transpiration stream of the grass. When the
present investigation was started in 1959, there was no
information in the literature as to how conidia are liberated.
This process was therefore investigated.
Ingold (1948) found that the form of E.typhina on
Dactylis glomerata produced long, threadlike ascospores which
showed no tendency to break into smaller units. The units in the
spore deposit measured 190 x 2A. On the other hand, from
specimens of the fungus on Holcus mollis the units measured
57 x 2j. Ingold attributed this to the breaking of the ascospores
into snarler The process appeared to be carried still
further in the form on Agrostis ternl:'1, since the spore deposit
from this strain was composed of units measuring 52 x 274. Each
measurement was the average of 100 spore units, chosen at random
from the spore deposit. Ingold (1948) suggested that it would be

interesting to know if these differences existed in other
localities and if the forms on other grasses showed further
variations. To this end, ascospores from three host species have
been measured.
(ii) Materials and methods.
Dactylis glomerata, Agrostis stolonifera, Holcus lanatus
and Holcus monis plants showing conidial stromata were taken from
the field and planted in pots in the conservatory at Silwood Park.
The following methods were used to release conidia from these
plants:
First, an attempt was made to collect the conidia from
stromata by a modification of the method described by Ingold
(1948). If the mature conidia fell freely from the stroma, they
would collect upon a slide. Each diseased shoot used for an
experiment consisted of a length of stem carrying a stroma. The
choked tiller was severed from the plant by a cut made below water
to avoid the interruption of the transpiration stream by n-sr
blocks in the vessels. The shoot was then transferred under
water into 4 one-inch diameter specimen tube through one of two
holes previously made in the cork stopper (see Fig. 2). The
specimen tube was supported obliquely so that the stroma .lay above
a clean slide on which any shed spores would fall. Although

-46-
Fig. 2. The apparatus used for collecting ascospores
from grass shoots carrying Epichloe typhina. stromata.
A. Specimen tube supported by a block of wood Hi B. Cork stopper; C.stem of grass; D. stroma;
E. flag leaf; F. slide supported on a block of wood,G.

stroma
slide
plasticinc
Plate.4. A modification of the apparatus shown in Fig.2. The specimen tube is supported by a lump of plasticinc in a sandwich box.

left for 48 hours on the bench in the laboratory, no conidia were
obtained by this methods
Since severed diseased shoots failed to discharge conidia,
an attempt was made to secure their production from living, choked
tillers which were still attached to the plants
The choked tiller was arranged in an inoculating chamber
(see Figs 1), which was clamped horizontally to a retort stands
A slide was expOsed under the stroma for 24 hours and afterwards
scanned under the microscopes No conidia were obtained on clean
or vaselined slidess
As a third method, a. modification of the Hirst spore
trap (Hirst, 1952), was used to collect conidia from unsevered,
living shoots carrying conidial stromata.
The choked tiller was enclosed in an inoculating chamber
(see. Fig. 1). The narrow end of the chamber was connected to the
spore trap by a. piece of rubber tube which was joined to the trap
at the orifice, through which air was sucked into the spore trap.
A glass tube, about 6 inches long, was inserted into a hole in the
rubber stopper which closed the wider end of the inoculating
chamber (see Fig. 1). was sucked into the trap having passed
over the conidial stroma in this tube. Suction pressUre was
provided by a motor operated ',limp which drew the air through the

-49-
trap at the rate of 10 litres per minute. Air was moistened by
passing it through water before it entered the chamber containing
the stromata. The conidia were collected on stationary,
vaselined slides. It was not possible to count the spores
trapped on the slide because of their smell size and the presence
of foreign bodies. However, more conidia were collected when the
air passed over the stromata was moist than when dry.
These results agree with those of Western and Cavett
(1959) who found that more of these could be recovered on trap
slides when stronnta were subjected to a. current of air carrying
atomized water than when a similar current of dry air was used.
Western and Cavett (1959) trapped spores in a heavily and uniformly
infected field near York, using a modification of the apparatus
described by Hyde and Williams (1945).
For all purposes, where ascospores were required during
this investigation, the first method described above, in which the
discharged spores fell on a glass slide (see Fig. 2) was used.
For each host species, 100 ascospores chosen at random from the
appropriate spore deposit were measured and the average size
determined. The average measurements of ascospores are shown in
Table 5.

Dactylis glomerata
Holcus lanatus
Agrostis stolonifera.
161.7 x 2.0 222.4 - 115.1
52.5 x 2.0 95.5 - 38.5
10.7 x 2.0 54.0 - 5.4
Table 5.
Average measurements of E.typhina ascospores from
different host species.
Average size (in microns)
Host species
I Mean Range of lengths*
(* breadth uniformly 2 A)
The differences in the sizes of the ascospores obtained
at Silweod were of the same magnitude as those found by Ingold
(1948) in material collected at Sevenoakes, Kent. It would appear
that the process of ascospores breaking into part-spores, to which
Ingold (1948) referred, was taken even further in E.typhina on
fgrostis stolonifera,
Periodicity of ascospore discharge.
Ingold (1948) noted a periodicity of ascospore discharge
in Dactylis glomerata, with a minimum in the morning and a maximum
in the late afternoon or evening. This was re-investigated for
ascospores from D.glomerata, H.lanatus and A.stolonifera.
The apparatus used in this experiment consisted of a
circular slide mounted on a clock. The surface of the slide was

marked into 12 divisions so that each division passed a fixed point
above the clock in one hour. A choked tiller in a specimen tube
(see Fig. 2) was suspended above the edge of the slide by means
of a clamp and retort stand. A plastic disc about 6 inches square,
with a small hole near one side was placed between the slide and
the stroma so that ascospores fell through the smi'll hole. This
ensured that the ascospores were deposited in a narrow band on the
slide. The disc was clamped to another retort stand. The
apparatus was enclosed in a polythene covered box to minimize air
movement. These experiments were carried out in late June and
July, 1960 at room temperature. The ascospores were deposited in
a layer arround the edge of the slide surface in the course of
12 hours,after which time the slide was removed and examined. It
was found that with the stromata from the three host species tested,
the heaviest deposit occurred between 6 and 10 p.m. The ascospore
deposit on the disc was too thick for a count to be made.

C. THE EFFECT. OF THE DISEASE ON THE HOST
(i) Effect on choked tiller.
Kirby (1958) described the "flag leaf" symptom of choke.
He observed the symptom on some diseased tillers in which the
stroma did not appear, but instead, the culm terminated in a "flag
leaf". He found this symptom on plants in controlled environment
rooms and in the field. As a result of his observations, he
concluded that the symptom was caused by the fungus killing the
flowering apex at an early stage, thereby preventing the formation
of the stroma. In the course of the current investigation, other
previously undescribed effects of choke on diseased tillers have
been observed.
Several tillers of Dactylis glomerata in which the strong
formed a loop protruding from the leaf-sheath were observed on
diseased plants in the Walled Garden and in the field. Some of
these were brought into the laboratory for closer examination.
The protruding loop appeared to form because the portion of the
internode below the stroma continued to elongate after the top of
the infloresence had been trapped and held tightly by the mycelium
in the stroma (see Plate 6). In a few tillers showing this
symptom, the pressure from the growing internode appeared to be so
great that the looping stroma sometimes snapped. This symptom was

str omo .
? l o t ~ . 5. C h o ke d cocksf oot t ill er s sh owing no r rn al st r mo to.

Platcz.6. Coeksfoot tiller
in the f orm f I
·nfeet ed with E. t sh owing strom ata--:"'--L..~_ _
p pr t ru di ng from th leaf sheaths.

-55-
also observed on Holcus mollis and Agrostis stolonifera, plants.
Other tillers were seen which besides the usual stroma, carried
several others on shoots arising from tiller buds from the same
stem (see Plate 7).
(ii) Effect on leaf-sheath.
Some tillers observed in the field, showed what appeared
to be "double stromata". There was one in a typical position,
at the top of the culn, and another one on the leaf sheath
arising from the second node below this stroma (see Plate 8).
However, the second "strona" was found to be a patch on the leaf-
sheath which had become infected and discoloured. Scrapings made
from the inside of this patch showed Epichloe typhina mycelium but
the part of the stem next to this patch showed no mycelium
externally. The stroma at the top of the culm was found to be a
normal one.
Two explanations are possible for the existence of the
diseased patch on the leaf-sheath below the true stronn. The
first is that the infection was from an external source. However,
attempts to inoculate plants through leaf-sheaths have repeatedly
failed to produce infection. Also, good growth of the fungus
externally is unknown, other than in association with floral
initiation. Kirby (1958) states that the formation of a strona is

stroma
stroma
stroma
Plate.7. A cocksfoot tiller infected with E.typhina. showing subsidiary
stromata on tiller buds.

..........~-.:.......... C ock sfo o t shoots inf ec t d withE.t yp hina. showing ·d o u b l ~ stromata:'
5 : tru troma.P : di as d pat ch on Iczaf- shea t h.
'--'__ s
,-_P
---_ P

-58-
always associated with floral initiation. The most likely
explanation seems to be that the diseased patch on the leaf-sheath
was formed in the early stages of floral initiation, when the
flower initials were in contact with this portion of the leaf -
sheath. The subsequent elongation of the internode freed the
diseased culm from the sheath, thus preventing the formation of
the strona at the point where the diseased patch was found.
(iii) Effect on calms and seed.
The adverse influence of Epichloe typhina on the host is
obvious when as in Dactylis glomerata panicles are destroyed and
little or no seed is obtained. However, its influence on a grass
like Festuca rubra is not so clearly defined (Sampson, 1933).
Sampson studied a number of populations of F.rubral infected and
healthy, and concluded that the adverse influence of E.typhina
may show itself in several ways. The disease may lead to a high
proportion of barren plants and an abnornnlly wide variation in
the number of panicles produced by fertile plants. In one
population, she found that the number of panicles per plant varied
from 0 to 700 while in another the range was 47 to 796. Exsertion
of panicles may be spread over an unusually long period and the
first panicles to be exserted are more likely to appear healthy
than later ones. Sampson (1955) also showed that the disease

causes reduction in seed production. This was shown in the
weight of seed produced, the proportion of "heavy" seed and the
germination (Tables 4 and 5). The tern "heavy seed" was used to
describe seed other than "chaff" or "light seed" in the samples.
The results of certain of Sampson's germination tests
are presented in tables 4 and 5,. These were carried out by the
Continental Method on Copenhagen tanks. Only one set of 100
seeds of each sample was tested and the seed was two years old at
the time the germination tests were carried out.
It nay be concluded from Sampsonts results that, in
F.rubra, the disease has an adverse effect, not only on the
quantity, but also on the quality of the seed produced from
infected plants.
In the present study, the effect of the fungus on the
size and weight of panicles and on the weight and germination of
seed of Dactylis glomerata, Festuca rubra and Holcus mollis was
investigated.
"Healthy" panicles were collected in the field from
plants producing "healthy" flowers only. Infected panicles were
obtained from plants showing 10% dhoked tillers. The choice of
this percentage was arbitrary, but it was used to ensure that the
diseased panicles were more or less uniform in that they came from
plants showing the sAne level of choke. The panicles were cut

Origin of seed
Weight of seed obtained from 10 panicles in gm.
Number of seedlings obtained from 10 panicles
1.11
Av.of 8 lots Range
159
Av.of 8 lots Range
0.28 (0.10 - 41 (0 - 108‘ 0.60)
0.11 (0.04 - 16 (0 - 97) 0.14)
Table 5.
I 'Healthy plant I Infected plants:
(a) Panicles not visibly infected
b) Panicles visibly Iinfected I
-60-
Table 4.
The might and the germination of seed from plants
of Festuca rubra infected by Epichloe typhina
(data of Sappson,1955).
The percentage of "heavy" seed and the germination
of samples from plants of Festuca rubra infected by
Epichloe typhina. (data of Sappson,1955)
Origin of seed No. or-heavy seed Germination fo
Healthy plant 69.2 70
Infected plants:
a) Panicles not visible infected: 6 lots 154 (0.2-51.9) 26 (0 - 77)
b) Panicles visibly infected (1) 58. 31
(2) 0.5 OOP

-61—
from the tillers as soon as they were dried and as near as
possible to the bottom pedicel. Random samples of 10 diseased
and 10 "healthy" seed heads each were procured from each host
species. Each seed head was weighed (fresh weight) and measured
from apex to basal pedicel. From each of the samples, 200 seed
were obtained by threshing, weighed (fresh weight) and subjected
to a duplicated germination test. The germination tests were
carried out on Copenhagen tanks at laboratory temperature, The
results are shown in tables 6 and 7,

-62-
Table 6.
The weight and lengths of panicles from visibly
infected and apparently non-infected plants of
Daetylis glonerata, Festuca rubra and Holcus mollis.
;Origin of panicles Fresh wt. of 110 panicles (g.)+ i
Av. length (cm.) from tip to basal pedicel. (10 panicles)*
Dactvlis glonerata i Range
a) visibly infected plant b) apparently non-infected
plant
2.01
4.67
10.3 ( 6.2 - 16.1)
18.2 (14.2 - 25.9)
Festuca rubra
0.40
0.81
10.6 (7,5 - 14.5)
11.1 (9.1 - 14.9)
a) visibly infected plant
b) apparently non-infected plant
Holcus mollis
0.58
0.90
5.9 (5.0 - 7.5)
7.4 (5.7 - 8.5)
a) visibly infected plant
b) apparently non-infected plant
▪ Representing the total weight of 10 panicles. The panicles
were not weighed individually because a sensitive balance was
not available at Silwood.
* Significant differences between means,
Dactylis glonerata
Festuca rubra
Holcus mollis
P=0.07 P=0.05
P = 0.01 P 2 0.05
P = 0.01 P=0.05
10.5 cm. 7.5 cm.
5.8 cm. 4.4 cm.
2.5 cm. 1.8 cm.

-63-
Table 7.
The weight and germination of seed from visibly
infected and apparently non-infected plants.
Origin of seed Fresh wt. of 200 seed in g.
Germination g_ Test I Test II Mean
Eactylis glomer-Aa
0.15
0.18
12
54
31
49
21.5
41.5
a) visibly infected plant
b) apparently non-infected plant
Festuca rubra
0.10
0.15
9
54
12
59
10.5
56.5
a) visibly infected plant
b) apparently non-infected plant
Holcus mollis
0.08
0.12
4
5
5
6
4.5
5.5
a) visibly infected plant
b) apparently non-infected plant
Before drawing any conclusions from these results
(Tables 6 and 7), it must be pointed out that the panicles from
visibly non-infected plants of D.glomerata alone could be regarded
as truly healthy. Sampson (1933) and Western and Cavett (1959)
have pointed out that healthy cocksfoot panicles never contain
mycelium. On the other hand, F.rubra panicles showing no

-64—
symptons of choke could contain mycelium of EpichIoe typhina
(Sampson, 1955). There is no similar information for ji.mollis
and, therefore, the possibility that panicles from this species
showing no symptoms of choke could contain mycelium cannot be
ignored. The fact that visibly non-infected panicles from
F.rubra and H. mollis could not be guaranteed to have been free of
the fungus detracts from the usefulness of the results obtained
for the two species and shown in Tables 6 and 7.
It would therefore appear that, on Dsglomerata, the
disease reduces not only the length and weight of the culm but
also the weight and germination of seeds The same conclusion
would hold for F.rubra and Hemollis if the panicles obtained from
visibly non-infected plants of these species in fact contained no
mycelium. Since this was not ascertained, the depressing effect
of the disease on these two species could have been due to the
degree of infection rather than the clear-cut presence or absence
of choke. Western and Cavett (1959) state that the proportion of
infected tillers on a diseased plant tends to increase in course of
time. Whether the reduced germination of seed from infected
F.rubra and H..relis plants was due to the attack of the disease
on the seed per se or whether it resulted from the general
weakening of the panicles was not determined.

-65
D. DISTRIBUTION OF MYCELIUM IN TILLERS
OF INFECTED PLANTS.
Sampson (1935) has pointed out that apparently healthy
tillers on infected grasses could contain mycelium of Epichloe
typhina. Western and Cavett (1959) state that choked cocksfoot
tillers always have abundant mycelium in the pith of the stem but
that tillers producing healthy panicles never do. The distribution
of mycelium in visibly infected plants, i.e. plants showing at
least one tiller carrying a stroma, was investigated by the writer.
Plants with at least one tiller carrying a stroma were
taken from the field and brought to the laboratory. Mycelium was
easily detected in the pith of the stems by slitting then
longitudinally with a sharp knife and removing some of the pith
tissue by scraping with a needle. The pith scraping was placed on
a slide and stained with cotton blue. In most cases, one scraping
was enough to demonstrate the presence of mycelium. However, when
no nyceliun was detected, other scrapings were made at one inch
intervals along the stem. al the tillers of every plant were
scraped at least once.
The results for D.glomerata and H.nollis are shown in
Tables 8 and 9. In addition, five F.rubra plants, with a total of
321 stems, all of which were searched for mycelium, showed that 520
stems contained the fungus. Only one was apparently free of it.

Table 8.
The distribution of E.typhina mycelium
in 9 infected D.glomerata plants.
Number of tillers
Plant Flowering* Showing stromata.
Showing no stromata but containing mycelium.
Not showing Total. mycelium or stromata,
1 5 21 2 15 41
2 0 4 5 21 28
5 6 5 2 9 22
4 6 52 6 22 66
5 19 2 2 17 40
6 12 25 1 9 47
7 2 7 5 4 16
8 0 11 27 12 50
9 24 15 1 5 45
Total 74 122 47 110 555
% of total 20.9 54.6 13.5 51.2
( * no flowering tiller contained mycelium)

-67-
Table 9.
The distribution of E.typhina mycelium in 5 infected H.mollis plants.
Nunber A of tillers
Plant Flowering Showing / stromata
Showing no stronata but containing mycelium
Not showing mycelium or stromata
Total
1 0 12 17 3 • 52
2 0 25 16 0 43
3 7 1 13 0 21
4 1 2 10 1 14
5 1 6 5 0 10
Total 9 46 1 61 4 120
% of Total 7.50 58.55 i 50.85 5.55

-68-
Since the ages of these plants and the period during
which they were infected were unknown, the figures for different
plants could not be compared. However, the proportion of tillers
not showing mycelium or stromata was high in ILglomerataz. In
F.rubra and H.mollis it was very low. It is therefore concluded
that when F.rubra and H.mollis plants are attacked by the fungus,
the mycelium tends to permeate all the tillers on the plant.
Observations made on plots of infected F.rubra and H.mollis plants
provide circumstantial evidence in support of this contention.
Whereas diseased plants of these two species produced a high
proportion of partirJ1y choked tillers, only once was one of the
latter nein on D.glomerata (see Plates Sand 10).

Plate. 9. Flowering shoots of Holcus mollis infected by E.typhina .
Showing partially choked flowers.

Plate.10. A flowering shoot of Dactylis glomerata, infected by
.typhina Showing a partially choked flower.

IV. PHYSIOLOGICAL STUDIES
A. Introduction
Vladimirskaya (1928) made a study of Epichloe typhina in
culture. She grew the fungus on a variety of media, both natural
and synthetic, and obtained good growth on potato, rice, wheat ears,
agar agar and malt extract gelatine. On the last two media, she
reported that she obtained perithecia but these formed no asci or
ascospores. Lilly and Barnett (1949) investigated the vitAnin
requirement of the fungus and found it to be heterotrophic for
thiamine. Vladimirskaya (1928) found the optimum growth temperature
for E.typhina to be between 16 and 19°C. However, Lilly and
Barnett (1949) and Kirby (1958) grew the fungus at 25°O., but they
did not state why they chose this temperature. It is generally true
that a knowledge of the basic nutritional requirements of a parasitic
fungus leads to a better unaerstanding of its parasitism and viru-
lence. With this in view, a study of the basic nutritional
requirements of E.typhina and of the physical factors affecting its
growth was undertaken.
B. Mr÷erials and General Methods
(i) Materials.
The cultures of E.typhina used in the following
experiments were isolated from diseased grasses at Silwood Park.

-72-
Choked tillers obtained from the field were cut into 2-inch long
pieces which were surface-sterilized in 0.1 per cent, mercuric
chloride for 3-4 minutes and then washed in sterile water. A
piece of stem was removed and cut open with a sharp scalpel
sterilized by dipping in alcohol and flaming. Pith scrapings
which were removed with a sterile needle were placed on potato
dextrose agar slopes. The slopes were incubated at 25°C.„ and
colonies appeared within a few days. This method was employed to
obtain cultures of isolates from Dactylisglomerata, Festuca rubra,
Holcus mollis and Agrostis stolonifera. Except where otherwise
stated, only the D.glomerata isolate was used in these experiments.
Cultures of Phycomyces blakesleeanus and Aspergillus niger which
were used in certain experiments, were obtained from the culture
collection of the Botany Depgrtment of Imperial College, South
Kensington.
(ii) Methods.
a.) Media
All chemicals used in the preparation of media were of
lAnalarl grade. Each chemical substance was dissolved separately
before being incorporated into the medium. 25 ml. of medium per
flask was used. Variations in this procedure are mentioned at
appropriate places in the'text. For studies concerned with fruiting,

-75-
the fungus was grown on a rice medium prepared from 10 g, rice and
25 ml. distilled pater. Long-grain Patna rice was used in
experimental work on fruiting.
b) Culture vessels
For most cultural work, 150 nl. conical flasks were used.
Plugs were made of cotton wool but in certain growth factor
experiments, aluminium foil caps were used instead of plugs because
pieces of cotton wool falling into the medium might have added growth
factors. All the vessels which had already been cleaned with water
and detergents in the laboratory were rinsed three tines with warm
tap water and three times with distilled water and allowed to drain
before use. Pipettes were first soaked in cleaning acid (50 g. of
Qr03; 100 ml. distilled water and 1,100 ml. corc. H2SO4) for
24 hours before being rinsed thoroughly in tap water and then in
distilled water. Agar slopes for the maintenance of stock cultures
were prepared in McCartney tubes. All the glassware used in these
experiments was of the Pyrex brand.
c) Methods of sterilization
Unless it is otherwise stated, all the media and the culture
vessels were sterilized by autoclaving for 20 minutes at 15 pounds
per square inch steam pressure. Before placing the culture vessels
in the autoclave, cotton wool plugs were covered with aluminium foil

-74-
caps to prevent the penetration of any condensed water. Pipettes
were wrapped in brown paper. Between the inoculation of one
flask and another, the inoculating pipette was sterilized by
heating over a spirit lamp.
d) Stock solutions
A minor elements solution was prepared and kept in the
refrigerator at 2-4°C. This solution was used for most liquid
cultures at the rate of 1 ml. per 100 ml. of medium. The
composition of this solution was as follows:
PeS04*7H20 0.1245 g.
ZnS04.7H20 0.1100 g.
CuS04'5H20 0.0195 g.
MnSO4.4H20 0.0205 g.
Na2Mo04.21120 0.0250 g.
These substances were dissolved separately and then combined, The
resulting solution was made up to 500 ml. and acidified with a.
trace of H2SO4. A stock solution of thiamine was prepared by
dissolving .00125 g. of the substance in a little distilled water
and making up to 10 ml. with distilled water acidified with 0.1 ml.
of N/1 HC1. The solution obtained, which contained 125 microgram
per ml. was stored in the refrigerator at 2-4°C. Suitably- diluted:
it was used in a number of experiments on growth factor requirement

•-75-
to give a final concentration of thiamine of 100 microgram per
litre of medium. This was obtained by diluting the stock thiamine
solution 50 times. This gave a solution containing 2.5 microgram
per ml. One ml. of this solution was incorporated into each
culture vessel containing 25 ml. of medium.
e) Maintenance of stock cultures
Stock cultures were grown on slopes made from 10 ml. of
glucose peptone agar medium. They were incubated at 18°C. and were
sub-cultured every 14 days. More cultures were stored under liquid
paraffin in a locker in the laboratory.
f) Method of inoculation
All cultures were inoculated from a bulb pipette ihich
was made in the laboratory. A piece of glass tithing about 1 ft.
long was drawn out to give two pipettes of equn3 length after
cutting. A cotton wool plug was placed just inside the wider end
of the pipette to prevent the entry of contaminants from the rubber
bulb into which this end was inserted.
The cultures from which the inoculum was obtained were
1-5 weeks old. 'When inoculum was required, 10 ml. of sterile
water was pipetted into a McCartney tube containing an actively
sporulating culture. A spore suspension was obtained by shaking the
tube vigorously. Rach culture vessel was inoculated with two drops

-76--
of the spore suspension. In growth factor experiments, the spore
suspension was decanted into a clean sterile tube innediately after
shaking, to minimize the solution in the spore suspension of growth
factors from the medium. Inoculation was carried out in the
inoculating room which was sterilized with ultraviolet light for
20 minutes before use.
g) Incubation
Incubation conditions varied with the experiment and are
described at the appropriate places in the text. In most of the
experiments the cultures were incubated in the constant temperature
room at 23°C. with artificial illumination. Unless it is other-
wise stated, all treatments were replicated four times.
h Method of assessing growth
To determine the weight of mycelium, aluminium foil cups
were prepared by moulding 2-inch square pieces of the foil on the
base of a one-inch diameter specimen tube. The cups were
numbered on the bottom with a 1Chinagraphl pencil. A piece of
muslin 2-3 inch square was placed in each cup which was then heated
in the oven at 70°C. for at least 2 hours to constant weight.
The weights of cups plus muslin were determined_ on an analytical
balance. Mycelium was collected by filtering the growth from
culture vessels through the muslin which had previously been

-77--
arranged in funnels for the purpose. The culture flask was rinsed
thoroughly, using a wash-bottle to dislodge bits of mycelium which
adhered to the sides, and the washings added to the growth on the
muslin. After squeezing off excess water, the muslin containing
the growth was transferred from the funnel to the appropriate cup.
The dry weight of the mycelium was determined after drying in the
oven at 70°C. for 24 hours.

-78-
C. FREL114INARY EXPERIMENTS.
(i) Growth of E.typhina on different media
E.typhina has been successfully cultured on complex media
by Vladimirskaya (1928) and Kirby (1958). The results obtained by
Kirby have already been described on page 16. He compared the
E.typhina isolates from D.glonerata, P.pratense, A.stolonifera and
H.mollis in culture and found that there was no relation between
the growth of the fungus on the plant and its growth rate on
artificial media.
An experiment was conducted to see if the isolates used
in the present investigation show sinilar differences. An isolate
of the endophytic fungus from LoIiun perenne was included to see if
it would show siniJarity to the E.typhina isolates.
To determine the best medium on which to compare the
isolates, 5 media were tested. These were Conn's)corn meal/
glucose peptone, Czapek's, glucose peptone, and malt, all of which
were prepared in liquid form. The composition of these media was
as described by Ainsworth and Bisby (1943).
To eliminate bacterial growth in the corn meal/glucose
peptone medium, an extract of corn meal was first prepared under
cold conditions. The appropriate quantity of corn meal was weighed
and soaked overnight in distilled water at 4°C. It was then stirred

-79.
for 4 hours at laboratory temperature and then filtered through
muslin. The solution obtained was centrifuged at 6000 r.p.m.
This technique gave a maize meal extract free from solids.
Glucose and peptone were then incorporated.
The culture vessels used in this experiment were
horizontally disposed 10 oz, medical flats each containing 30 ml,
of medium. Rich treatment was duplicated. The inoculuri consisted
of a piece of growing colony from an actively sporulating cultUre of
the El.gIomerata isolate. Incubation was for 7 days at 23°C. in the light
The mean results of this experiment are shown in the
histogram (see Fig. 3). The corn meal/glucose peptone medium gave
the greatest weight, and therefore was chosen for comparing the
growth of the different isolates.
The isolates of E.typhina Prom Dftglomerata, F,rubra,
A.stolonifera and the endophyte from Lolium perenne were compared.
The Loliun endophyte was isolated from pith scrapings made from
stems of the host obtained in the field at Silwood. The method cr
isolation was similar to that used for E.typhina isolates (see page 72).
L.perenne did not show any disease externally and many stems had
been scraped for mycelium without success, before a patch of plants
was discovered in which the stems showed mycelium in pith scrapings.
Several isolations were made and the sane kind of mycelium was

40
-80--
-30
-20
10
—0 glucose Conns Corn meal Malt peptone glucose peptone
Fig. 3. Growth of E.typhina. on different media. Two replications incubated seven days at 23°C In the light.
7, /'. ,*>
Czapcks

-81-
. obtained consistently. The mean results of the
comparison of dry weight growth rates of the different isolates are
shown in the graph (Fig. 4).
All the Epichloe typhina isolates produced a white,
floating mat of growth whose surface showed brain like folds. No
morphological differences were observed and all the isolates
produced conidia.
The isolate from L.perenne gave a good growth which was
grey in colour. No spores were observed. Exarlination of the
mycelium under the microscope showed that some cells were swollen
at one end, gibing the hyphae a beaded appearance. The hyphae were
5.5 M thick but in the swollen parts they attained a diameter of
6.6 m.
Discussion.
E.typhina is entirely a parasite of the Gramineae and as
such, it was not surprising that it should grow best on the medium
containing maize. meal. In infected D.glomerata plants, the cilms
exhibiting the external symptoms of the disease, with very few
exceptions, were always completely choked. At Silwood, partially
emerged heads were more common on F.rubra than on D.glomerata.
This may indicate that the amount of fungal material produced in the
host plant, is greater on choked shoots of D.glomerata than of F,rubra.

r, i
—82—
i • N ..•...• .....i
i ..., ,-Al . I ------- •,,, \
. ‘
I s . • ‘,N,
i g ' \ ,\
I• i •
..!1-4. •• 1 .., 1•,... N .•
t„. _ \•.. .04 Ig ...• • ... • i •••• ...
• ....NO- - -.p.c.' ,..—• I ..., 40;.......: ... -, ......-t-.
IC EY.
Dactylis glomcrata. Agrostis stolonifera.
--Festuca rubra. perenne.
500"
400.
OF M
YC
ELIU
M.
300'
200'
ce
100.
0 7 I'4 21 1 2 TIME IN DAYS.
Fig. 4. The growth (Dry weight)of different isolates of E.typhina and the Lolium percnne endophyte in culture
(Two replicates incubated at 236C in the light)

Howeler the resUltS it Vigi 4 show thatt on the mediUt tested, the
Diglomerata isolate grew no faster than the PaUbra ones It is
therefore concluded that there .s no simple relation between the
gro wth of the fungus on the plant and its rate of growth in
weight on corn mealiglUdobe peptone medilam. Preliminary
experiments previously carried out by the writer gave similar
results and these agree with those of Kirby (1958),
The isolate from Lolium perenrs produced no spores and
therefore its identity remains unknown*

(ii) Standardization of inocula and source.
Introduction
The age, history and kind of inoculum may influence the
rate and amount of growth and other functions of the fungus (Lilly
and Rirnett, 1951). To ensure consistent results therefore, an
experiment vas designed to standardize the inoculum and its source.
Materials and Methods
Agar slopes of nnit, corn meal, pea extract, Czapekts,
Coon's and glucose peptone media were prepared in McCartney tubes.
Each tube was inoculated with 6 drops of spore suspension from an
actively sporulating culture of E.typhina. The inoculun was spread
over the medium surface with the inoculating pipette. The
cultures were incubated at 18°C. under constant illumination,
After 14 days incubation, spore suspensions were prepared by adding
10 ml. of sterile water to the cultures and shaking the tubes
vigorously. Haemocytometer counts were made and the number of
spores per ml. of the spore suspension prepared from each medium
was calculated. Haemocytometer counts are shown in the histogram
(Fig. 5).
Glucose peptone agar gave the best spore production and
consequently was used for the source of inoculun for all the sub-
sequent experiments. The number of spores per drop of spore

CHICK PEA AGAR (13.6 x106/m1)
COON'S AGAR (4.4x 106/m0
GLUCOSE PEPTONE AGAR (444 x 106 /ml,)
CZAPEKS AGAR Q3.16 x 106/m1:)
PEA EXTRACT AGAR(76x141)
AM ALT AGAR (37.2 x 106/m1)
O 10 20 310 40 50 (.1x10 6 ) spores/ml.of spore suspension
(0ml.prepared from each slope culture)
E.typhina spore production in different media. (Incubated 14 days at I8°C in the light)

-86,-
suspension delivered by the pipette described on page /Yyms
determined by calculation. One drop of the spore suspension
prepared as above from glucose peptone agar medium was found to
contain 1.12 x 106 spores.

Influence of phosphate and pH,
The three salts of orthophosphoric acid, i.e. KH2PO4,
:2HPO4 and K5PO4 furnish utilizable phosphate but their effects on
the acidity of the medium are quite different. It was not known
how E.typhina in culture would react to different pH and therefore
the effect of the different phosphates on the fungus in culture and
the accompanying pH changes were investigated.
The composition of the basal medium used in this
experiment was as follows!
Glucose (w/v)
Casein hydrolysate 0.4%
MgS0407H20 0.05%
Thiamine 50 microgram per 100 ml.
Minor elements
Each phosphate was separately incorporated into the basal
medium before autoclaving at the rate of 0.1% w/v. A control
treatment consisting of the basal medium only was also prepared.
Incubation was at 18°C, in the light, The mean dry weight yields
of mycelium after 15 days incubation at 18°C, in the light are shown
in Table 9,

Table 9.
The effect of different phosphates on the
growth of Ep3ohloe typhina.
(4 replicates, incubated 15 days at 18°C. under constant illumination)
Treatment Mean dry vet. of mycelium (mg.)*
Initial pH
_.c,....,
Final pH pH Range
KH2PO4 125.0 3,8 4.2 5,7 - 4.4
K2HPO4 150.7 7.1 7.6 7,1 - 7.7
K5PO4 121.6 7,5 8,1 7.5 - 844
Control 105,7 5.7 5.7 5.5 - 5,8
* Significant differences between means,
P = 0,01 14,4 mg,
P = 0.05 10.2 mg.
All the treatments gave better growth than the control
but K2HP04 gave significantly better growth than the rest (at P =
0.01). It was concluded that K2HPO4 was the best phosphate source.
In i11 subsequently described experiments in which the medium
contained a potassium phosphate K2HPO4 was usedr except in the
growth factor experiments which were conducted before this result
was available.

-89-
The growth obtained in the control treatment deserves
further comment. In this treatment it is assumed that the fungus
obtained phosphorous from casein hydrolysate, casein being a
phosphoprotein. However, there vas no compound that could supply
potassium, which is also essential for fungal growth. Steinberg
(1946) found that when Aspergillus niger was grown in media
containing sub-optimal amounts of potassium, sodium gave increased
yield. This element (sodium) acted by biological substitution.
Bacto-vitamin free casein hydrolysate which was used in the
experiment under discussion contains 58% sodium chloride (Blifco
manual, 1955), and in addition a small quantity of sodium had been
added to the medium in the form of sodium molybqate, which was a
component of the minor elements solution. This nay explain the
relatively good growth in the control treatment.

-90-
D. Experiments vr3.th growth factors
(i) Response of E.typhina to thiamine, biotin and
yeast extract.
Introduction
The growth and reproduction of many fungi are dependent
upon the presence in the medium of certain growth factors. These
are often the members of the vitamin B-complex (Lilly and PArnett,
1949). The effects of vitamins in the medium are usually measured
by the resulting vegetative growth. Lilly and Barnett (1949)
tested the response of E.typhina to the addition of thiamine,
pyridoxine, inositol and biotin to the medium but only thiamine
led to increased growth. They also stated that different isolates
of any one species of fungus may have different vitamin deficiencies.
The response of the D.glomerata isolate of E.typhina to
thiamine, biotin and yeast extract was investigated.
Materials and Methods
The basal nutrient solution used in this experiment was
composed of:
Glucose
Casein hydrolysate (vitamin free) 0.4%
IQi2PO4 0.1%
MgSO4.7H20 0.05%

-91-
After dissolving those substances, the resulting solution
was boiled with 0,5% activated charcoal to remove growth factors.
When cooled, the charcoal was filtered off using glass wool,
Minor elements were then added and the volume adjusted before
25 ml, of medium was dispensed into each of the appropriate flasks.
These were plugged with cotton wool and autoclaved,
The growth factor solutions used in this experiment
were: Thiamine: 12.5 microgram per ml.
Biotin : 1.25 microgram per ml,
Yeast extract: 0.1%)yeastrel solution
These solutions were autoclaved separately and afterwards
aseptically combined with the basal medium.
The following treatments were prepared in quadruplicate:
a) 25 ml. of basal medium + 1 ml. thiamine + 1 ml. sterile water.
b) 11 n n tt n + 1 ml. biotin + 1 ml. sterile water.
o) tt n n tt ft + 1 ml. thiamine + 1 ml. biotin.
d) tt ft it It f► + 2 ml. sterile water.
e) It II ft il If + 2 ml. yeastrel.
After inoculation the cultures were incubated at 18°C in
the light.
Table 10 shows the response of the isolate of E.typhina
from D.glomerata to the g=ezh factors as assessed by determining
the dry weight yield of mycelium after 55 days incubation at 18°C

-92-
under constant illumination,
Table 10.
Response of E.typhina (ex p.glomerata) to thiamine,
biotin and yeast extract.
(4 replicates, incubated 55 days at 18°C. under
constant illumination)
Treatment Mean dry wt. of mycelium (pg.)
Thinmine 142,4
Biotin 155.4
Thiamine Biotin 155.2
Yeast extract 144.6
Control 144.5
Significant differences between means.
P = 0.01 17.1
P = 0.05 12.5
It is evident from the data in Table 10 that the
addition of growth factors gave no, significant increase in growth.
Biotin appeared to have a slightly inhibitory effect although this
was not significant at the P = 0.05 level. However there was
significant difference (at P = 0.01) between the biotin and the
thin ne plus biotin treatments.

-95-
To test whether biotin had a slightly depressing effect
on the growth of this isolate, a further experiment was designed.
This experiment consisted of three treatments prepared as
described in the preceding experiment and in quadruplicate.
a) 25 ml. basal. medium + 1 ml.biotin 1 ml. sterile water.
b) 25 ml. It 91 • 2 ml. yeast extract solution.
c) 25 ml. tt PI • 2 ml. sterile water.
The results obtained in this experiment are shown in
Table 11.
Table 11.
Response of Estyphina (ex D.glomerata) to biotin and
yeast extract.
(4 replicates, incubated 15 days at 1800 in the light)
Treatment Mean dry wt. mycelium (mg.)
Biotin 126.7
Yeast extract 125.2
Control 120.1
Significant differences between means,
P = 0.01 12.2
P = 0.05 8.5

-94-
From the data in Table 11, it is quite clear that biotin
led neither to increase nor reduction of the growth of E.typhina.
It was therefore concluded that the previous small apparent
effect of biotin was fortuitous.
The data In Tables 10 and 11 indicate that the
D.glomerata isolate of E.typhina used in these experiments was
self-sufficient for thiamine and biotin and that yeast extract did
not improve its growth in the conditions under which it was cultivated.
However, the need for a given vitamin in some
instances depends upon the composition or pH of the medium or upon
the temperature of incubation (Lilly and Barnett, 1949). As
these workers (Lilly and Barnett, 1949) had found their isolate of
E.typhina to be thiamine deficient, it was decided to test the
thiamine requirement of the D.glomerata isolate using their medium
and methods.
The basal solution consisted of:
Glucose 2.5%
Casein hydrolysate (vitamin free) 0.2%
Pumaric acid 0.152%
The solution obtained when these substances were
dissolved in distilled water, was boiled with 0.5% activated
charcoal. After cooling and filtering off the charcoal, the

-95-
following mineral salts were incorporated into the medium.
KH2PO4 0.1%
m004.71120 0.05%
&ISO4.71120 0.00002%
FeS04'7H20 0.00002%
MnS04.4H20 0.00001%
The pH of the medium was adjusted to 6.1 and the thinnine
added at 100 microgram per litre of the medium, before autoclaving
for 15 minutes at 15 lbs. per sq. in. steam pressure. The pH
after autoclaving was 5.8. The cultures were prepared in
quadruplicate. The treatments comprised media with and without
thiamine at each of three temperatures, with sufficient cultures
to permit two harvesting dates. Miter inoculation, they were
incubated at 10 and 18°C. in the light and at 25°C. in the (lark.
Lilly and Barnett (1949) state that an incubation period of at
least 1 or 2 months is required to distinguish between partial and
total growth factor deficiencies. For this reason, the cultures
were harvested after 14 and 51 days. The data are shown in
Table 12.

-96-
Table 12.
The response of E.typhina (ex D.glomerata) to thiamine
in a medium as used by Lilly and Rnrnett (1949).
(4 replicates, incubated at 10 and 18°C . in the
light and at 25°C. in the rinrk)
Mean dry weight of Mycelium
Days of incubation Treatment 1 Temperature of incubation °C.
10 18 25
14 Thiamine 147.4 165.8 172.7
Control 154.2 166.5 172.7
Thiamine 168.5 161.5 161.9 5/ Control 16962 162.1 161.2
Significant differences between means,
after 14 days incubation: P = 0.01 9.2
P = 0.05 6.7
after 51 days incubation: P = 0.01 7.1
P = 0.05 5.1
As the data 2, Table 12 show, thiamine led to a smn,31,
highly significalli; increase in the growth of Estyphina_
(ex D.glomerata) after 14 days incubation at 10°C. (significant
at P = 0.01). Apart from this, the addition of thiamine had little
effect,

-97-
There are two possible explanations for the discrepancy
between these results and those obtained by Lilly and Barnett (1949)
a) that the media used in these experiment were not
entirely free of thiamine, or
b) that the strain used in this investigation was
self-sufficient for thiamine.
To test the first of these possibilities, a. further
experiment was designed.
In this experiment, the growth of the E.tyohina isolate
under study was investigated in a medium with and without thiamine,
using additionally, a thiamine deficient fungus, Phycomyces
blakesleeanus, to test that the medium was in fact thiamine free.
The basal medium had the composition:-
Glucose 2.5%
NH4H2PO4
0.2%
KH2PO4 0.1%
MgSO4.7H20 0.05%
An inorganic N-source was employed to reduce the
likelihood of inadvertently adaing growth factors of exogenous
origin. Minor elements were added after boiling with activated
charcoal as in the previous experiment. Treatments with and
without thiamine were preparedlin triplicate for each of the two

fungi. Thiamine was added at the rate of 100 microgram per litre
of medium. The pH was adjusted before autoclaving to 5.9.
After autoclaving the pH was 5.6, The cultures were incubate&
for 9 days at 25°C. in the dark. Table 15 shows the results
obtained in this experiment.
Table 15.
Response of E.typhina (ex LI.glomerata) and Poblakesleeanus
to thiamine.
(3 replicates incubated 9 days at 25°C. in the dark)
Purgus Treatment Wan dry weight of mycelium (mg.)
E.typhina + Thiamine 176.4
Control 172.2
Potlakesleeanus + Thiamine 65.5
Control 2.7
Significant differences between means,
E.t-vphina P = 0.01 52.5
P = 0.05 19.6
P.b1akesleeanup P = 0.01 10.1
P = 0.05 6.1
It is clear from the data on Table 15 that there couIel
have been virtually no thiamine in the basal medium. The slight

growth obtained in the control treatment inoculated with
P.blakesleeanus was probably because snfill quantities of thiamine
night have been added with the inoculun. No sporangiophores
fcrned in the basal medium whereas these were produced abundantly
the treatment to which thiamine had been added (see Plates 11
rnd 12). The conclusion was therefore drawn that the cocksfoot
:,solate of E.typhimumder study mRs not thiamine deficient,
except possibly at 10°C., where, as shown in Table 12, growth
vitbout exogenous thiamine was slightly, but significantly less
than witla thiamine.

WITH THIAMINE. WITHOUT THIAMINE.
Plate. II. The effect of thiamine on the growth and
sporulation of p. blak es le ea nu s.

WITH THIAMINE. WITHOUT THIAMINE.
Plate.12. The growth of E.typhina (exD.glomerata) in the medium containing thiamine and in the control.

(ii) Response of isolates of E.typhina from different
grasses to thiamine in the culture medium.
Using a medium of the same composition as that used in
the last experiment, and following the sane method ae preparation,
the response of the isolates of E.typhina from Dactylis glonerata,
Festuca rubra„ Holcus nollis and Agrostis stolonifera to thiamine
was studied in artificial culture at 25°C. Treatment with
ayconyces blakesleeanus mere also included to serve as indicators
that the basal medium was in fact free from thiamine. The
treatments were prepared in quadruplicate. The pH before and
after autoclaving vas 5.9 and 5.6, respectively. The dry weight
yields of mycelium after 10 days incubation in anrkness are
tabulated below (Table 14).

-103—
Table 14.
Response of E.typhina isolates from D.glomerata, F.rubra,
B.mollis and A.stolonifera to thiamine, using
R.blakesleeanus as control.
(4 replicates, incubated 10 days at 25°C. in the dark)
Mean dry n wt. of celium •
Fungus . Host species Treatments
Thiamine Control
E.typhina D.glomerata 189.8 189.8
F.rubra 200.7 198.3
H.mollis 144.9 150.9
A.stolonifera 132.1 128.7
P.blakesleeanus 155.9 7.8
Significant differences between means:
E.typhina P = 0.01 16.1
P = 0.05 12.3
Fcblakesleeanus P = 0.01 25.0
P = 0.05 16.6
Only the Holcus mollis isolate gave a significant, albeit
son11, increase in growth due to the addition of thiamine (P = 0.05).
The Festuca rubra and Agrostis stolonifera isolates showed slight
growth increases but these were not significant. It was therefore

-104--
concluded that, like the isolate from D.glomerata, the F.rubra and
A.stolonifera isolates were self-sufficient for thiamine. The
results obtained for ELglomerata isolate confirmed those obtained
previously (see Tables 10, 12 and 15). An experiment was
designed to further test whether E.typhina from H.mollis was
slightly thiamine deficient.
This experiment was set up exactly as the above, except
that only P.blakesleeanus and the H.mollis isolate of E.typhina
were used. The pH of the nutrient solution before and after
autoclaving vas 5.8 and 5.5, respectively. The dry weight yields
of E.typhina (ex H,mollis) after 5, 7 and 12 days incubation at
25°C. in darkness are shown in Figure 6. The corresponding
figures for P.blakesleeanus after 7 days incubation were:
control 6.5 mg.; thiamine 69.1 mg.
The results obtained in this experiment confirmed those
of the preceding experiment in respect of the H.mollis isolate of
E.typhina, revealing a slight partial thiamine deficiency, as
shown by the slower rate of growth in the thiamine-free medium
than in the presence of added thinnine. Addition of thiamine to
the medium led to increased amounts of growth on the 5th and 7th
days but not on the 12th day. The increase was, however, only
significant on the 5th day (P = 0.05).

-105-
140
120 ay
1100 / D / 71 / W / O / 80 / ›. M o /
/ U. 0 60
t9
• 40
cC —Thiamine. —Control.
• 2 0 i Significant difference between means. §dc
124.-0.0 I 12.1 593 I5.3 13,10.05 SO 39-I 10.1
0 2 4 6 es 10 12 INCUBATION. (PAYS)
Fig.6. The growth of E.typhina (ex H.mollis) in a basal medium with and without thiamine.
(Four replicates at25ccb in the dark)

-1 Ce -
It therefore appears that the H.mollis isolate of
E.typhina suffers initially from a thierilne deficiency, but that
after about 5-7 days growth it presumably synthesizes enough
to meet its needs adequately.

-107-
(iii) Investigation of thiamine requirement of E.typhina
isolated from D.glomerata, in cultivation at low
temperatures.
It was shown in an experiment in this series (see Table 12)
that thiamine led to a small but significant increase in the growth
of E.typhina isolated from D.glomerata when incubated at 10°C. for
14 days. A deficiency for riboflavin conditioned by temperature
was reported by Mitchell and Houlahan (1946) for a mutant of
Neurospora, with which growth was poor or lacking at temperatures
above 25°C. unless riboflavin was added. Below 25°C. the fungus
was able to synthesize riboflavin. The partial deficidncy of
Sclerotinia camelliae for inositol was influenced by temperature,
particularly in the supraoptimal range (Bnrnett and Lilly, 1948).
The influence of temperature on the thiamine deficiency of
E.typhina isolated from D.glomerata was therefore studied.
The basal nutrient solution used in this experiment, and
the method of preparation was according to Lilly and BrIrnett (1949)
as described on page V.1.. The basal medium contained glucose 2.5%,
casein hydrolysate (vitamin-free) 0.2%1 fumaric acid 0.132% and
mineral salts (see page 95). The cultures were incubated at 8 and
18°C. in the light. Sufficient cultures were prepared to permit 4
to be harvested from each treatment on the 7th, 14th and 21st days.
The results are shown in Figure 7 and Table 15.

Mean dry weight of mycelium (mg.)
Temperature of incubation.
Treatment Days of incubation
C. 14 21
Thiamine 9.3 61.2 112.8 8
Control 8.6 42.5 73.9
Thiamine 126.3 196.2 213..8 18
Control 131..4 198.2 221.4
Significant differences between means:
at 8 C. P = 0.01 11.8 48.2 53.8
P = 0.05 7.8 31.8 34.9
at 18°C. P = 0.01 73..1 31.3 20.9
P = 0.05 46.9 20.7 13.9
-108-
Table 15.
The growth of E.typhina (ex D.glomerata) in a medium with
and without thiamine, incubated at 8 and 18°C. in the
light.
(4 replicates harvested after 7, 14 and 21 days incubation)
It is evident from the Graph (Figure 7) that the isolate
was self-sufficient for thiamine at 18°C., there being no
significant Oifferences between the control and the treatment with
thiamine at any time during the period of incubation (21 days).
However, at 8°C., the addition of thiamine to the medium resulted.
in increased growth on the14th and 21st days although the increase

.....-0 18°C.CONTROL.
... - - --- 0 18°C. THIAMINE. 2 20
200
180
160
e-I ;140
8°C. THIAMINE.
80-
- .0 8°C. CONTROL.
7
TIME IN
I4
DAYS,
— THIAMINE.
--- CONTROL.
21
)- 40- cc 0
20-
-109-
FIG. 7. RESPONSE OF E.TYPHINA. ISOLATED FROM D.GLOMERATA. TO THIAMINE IN THE CULTURE MEDIUM. INCUBATED AT eAND I8°C. IN THE LIGHT.

Days of incubation Treatment
Thiamine
Control
162.5
147.6
144.7
1/14.6 59.6
21.9
5.0
5.8
86.5
70.7
Mean dry weight of mycelium (mg.)
55 42 7 14 20
Significant differences between means:
P = 0.01
P = 0.05 1.8 j 10.8
1.2 11 7.1
41.5
27.4
19.2 26.5
12.7 17.5
was statistically significant only on the 21st day of incubation
(P = 0.05).
This experiment was repeated at 8°C. and the period of
incubation extended to 42 days. The results are shown in
Table 16 and Figure 8.
Table 16.
The growth of E.typhina (ex D.glomerata) in a medium
with and without thiamine, incubated at 8°C. in the
light.
(4 replicates harvested after 7, 14, 20, 55 and 42 days
incubation)
Again thiamine led to a srin31 increase in the rate of
growth but this was significant only on the 14th day of incubation
(P = 0.01). The results of the two experiments indicate that the
isolate of E.typhima from D.glomerata has a partial thiamine

0 14 21 2 8 35 42 49
-11.1-
6, M
TIME IN DAYS. Fig. 8. The response of E.typhina. isolated from D.glomcrata.
to thiamine in the culture medium incubated at 86C In the light.

-112-
deficiency at low temperatures,, manifested as a smalls temporary
retardation of grouth in the absence of exogenous thiamine.

-315-
E. Carbon nutrition: Utilization of
different sources of carbon.
Introduction
Carbon occupies a unique position among the essential
elements required by fungi. Almost half of the dry weight of
fungus cells consists of carbon (Lilly and Rnrnett, 1951).
Protoplasm, enzymes, the cell wall and the reserve nutrients
stored within the cells are compounds of carbon. Carbon compounds
are equally important in fungal nutrition and fungi obtain energy
by oxidizing organic compounds. The utilization of carbon
compounds by fungi depends on the composition, structure and con-
figuration of the compounds. An experiment was designed to study
the utilization of different carbon compounds by Epichloe typhina.
Only the isolate from Dactylis glomerata was used in this and the
subsequent experiments.
Materials and method.
In this experiment, a number of organic compounds were
separately added to a. basal medium supplying all the necessary
nutrients save the carbon-plus-energy source. The medium was made
up of: C - source 1% (weight/volume)
(NH4) 21T04 0.1%
MgSO4.7H20 O.05%
X01 0.00
Minor elements

-114-
In order to sterilize the c-source solution separately,
the medium was prepared in two parts. After the c-source solution
was prepared and its pH adjusted to 4.5 (with NaOH or HCl), 5 ml.
portions were dispensed into the appropriate flasks. The basal
medium was the sane for all treatments and was prepared in bulk.
After autoclaving the solutions, 20 ml. of basal medium of an
appropriate concentration wereadded aseptically to each flask con-
taining a. carbon source solution. The 5 ml. portions of the 0-
source solution dispensed into each flask, contained an amounlit Cf
c-source equivalent to 1% w/v in the final volume of the medium
(25 ml.). Likewise, the 20 ml. of the basal medium added con-
tained amounts of constituents equivalent to 25 ml. of the medium.
In order to study the rate at which growth proceeded with the
different 0-sources, the experiment was designed:. to allow three
harvesting dates for each c-source, four cultures from each treat-
ment being harvested at each. Incubation was at 25°C. under
constant illunination.
The following carbon-energy sources were tested:
Honosaccharides:
Pentose L-arabinose
Hexoses DI-glucose, DI-fructose and L-galactose
Disaccharides: lactose and sucrose
Polysaccharides: cellulose, starch and pectin
The results are shown in Figures 9 and 10 and Table 17.

-115-
100
90-
801
60
WE
IGH
T OF
MY
CE
LIU
M
40'
50
30
20
I0
0 7 I4
2 1 TIME IN DAYS.
Fiq.9• Utilization of different carbon scources by Etyphina.
(Four replicates incubated at 23° C in the light)

-116-
100 .1 •
90
kAcr,ft NI" vse
80
do 70
z
60
•
r I
40
20
DR
Y W
EIG
HT
OF M
YC
ELI
UM
PECTIN
7 14 21 TIME IN DAYS.
F19.10. Utilization of different carbon sources by .typhina., (Four replicates, incubated at 23°C in the light).

-117-
Table 17.
Utilization of different carbon sources by E.typhina.
Each figure in the table represents the mean of
4 cultures (12 for glucose). Incubation was at 25°C. in the light.
Mean dry weight of my celiun
carbon source Days of incubation
7 14 21 11.
Ir-Arabinose 75.2 98.1 88.5
D-Glucose 105.5 96.4 89,9
D.Fructose 61.8 95.6 84,5
L-Galactose 100.1 85.6 78.6
Lactose 6,9 5.9 6.1
Sucrose 110.8 94.2 84.7
Cellulose 29.1 15.0 11.9
Starch 82.9 88.0 87.5
Pectin 2.9 5.1 5.6
Control no growth ,no growth no growth
Significant differences between means:
P = 0.01
P = 0.05
na 8.2
::: 6.6
4.9

-118-
Periodic visual observations were made on the cultures.
It was found that glucose, sucrose, galactose, starch and fructose
gave rise to a. thick, white sheet of mycelium covering the whole
surface of the mediun. After 7 days incubation, conidia were
produced profusely in all except the treatment containing fructose
in which conidia were observed after 14 days incubation.
After the first week of incubation, arabinose and
cellulose gave a patchy growth, consisting of very thin,
scattered pieces of mycelium on the surface of the medium. A
thin layer of slimy, transparent, submerged growth was found on the
bottom of the flasks. However, during the 2nd and 5rd weeks of
inceation„ the nutrient solutions containing these two c-sources
sustained a thicker growth, which covered the whole surface of the
medium. Conidia were produced profusely after the 2nd week of
incubation.
Lactose and pectin gave a little, thin and submerged
growth throughout the period of incubation. After 14 days
incubation, a few conidia were seen on mycelium obtained from the
cultures containing pectin. No conidia were seen at any time in
the cultures containing lactose.
On the basis of these results, the c-sources tested in
this experiment fall roughly into three groups. The first group,
comprising glucose, sucrose and galactose consists of the compounds

in which the fungus attained maximum growth in the first week
after which autolysis set in. Arabinose and fructose fall into a
second group on which the fungus attained_maximun growth after 2
weeks incubation. With all 5 of these sources, the maximum
yield was clearly similar (95.6 w 11048 mg.). Starch would seen
to lie between the first and the secondgroup. The amount of
growth when starch was used. as the carbon source in the nutrient
solution, remained substantially uniform after the first week of
incubation until the end of the experiment (5 weeks). The
third group of carbon sources consists of pectin, lactose and
celluloce, The medium sustained little growth when these sub-
stances were used as sources of carbon. They are therefore con-
sidered to be poor sources of carbon for the fungus.

420-
7, Nitrogen nutrition
(i) Utilization of different sources of nitrogen,
Nitrogen is an essential element for fungi and is used
for functional as well as structural ptrposes (Lilly and wirnett,
1951). Not all nitrogen sources are equally suitable for all
fungi, Many fungi can utilize nitrate-N as a sole N-source, but
these must be able to reduce the nitrogen to the oxidation level
of nnmonia, Fungi lacking the appropriate reductase systems
will need already reduced N-sources, for example NH4.1.
salts,
Others are deficient in synthetic powers and require amino acids,
peptides and mixtures of these such as peptone, According to the
Ns-source used, there are often characteristic pH changes in the
medium during growth,
An experiment was designed to investigate the
utilization of different N-sources by Estyphina and the accompanying
pH changes in the medium. In order to obtain a basis for com-
parison of the utility of the nitrogen presented to the fungus in
different chemical combinations, the sane amount of nitrogen was
presented in each treatment, The final concentration of nitrogen
was calculated so as to give l/50th ' gram atomic weight of
nitrogen per litre of mediumf The N-sources solution was
prepared double strength in each case and the pH adjusted to 4.5

before combining with the basal mediums which had the final
composition:-
glucose i% (weight/volume)
K21 P04 0.1%
MgSO4.7H20 0.05%
Minor elements
The basal medium was also prepared double strength and
Mluted by an equal volume of the N-source solution before auto-
claving, to give the above final concentration. An extra flask of
medium in each series served for initial pH determination, After
inoculations the cultures were incubated at 2500. in continuous
illumination. Four cultures were harvested from each treatment at
weekly intervals for a month. The dry weight yields of mycelia
and final hydrogen ion concentrations are presented in Table 18 and
Figures 11 and 12.

Final pH
Dry wt. of Myc. ug.
7 r.e
7.7.6 7.8
7,7
7.9 115,3
3.5 1 4.1 2.5 1 5,1
N..souroe Initial pH
Number of days of incubation
21 28 Dry Isiainal of UV. mg,
pH Dry wt, of Ilya, tg
Pine, pH
15.2 5.9 19,8 4.1
65.5 8.7 62.2 8.8
82.1 2.2 81.4 2.4
81,2 2,4 78.7 2,6
140.2 8,7 155.2 8,T
112.0 8,2 115.5 8.7
114,9 7,6 107.6 7.8
102.9 7.8 85.9 8,4
105.5 8.1 102.7 8,5
5.1 8.6 5.8 6.2
14 Dry wtainal of Hyd, pH mg.
10,5
76,9
84,1
85,7
7,8
Control
KN 03
NH 01 4 (NH4)2 04
L-Glutamio acid'_
L-Asparagine
Casein hydrolysate
Peptone (14.5%N)
Gelatine
6.8
6.9
7,0
6.5
4.9
7.1
6.6
6.7
6.9
10.2
95.5
98,2
101.0
155.5
126.9
123.2
137.9
8.6 143.7
7.9 112.8
4.6
8.5
2,4
2.4
8.8
7,8
8.1
8.4
4,1
8.7
2.0
2.7
Significant aft. between means:
P = 0.01 P =. 0.05 !
T-.ble 18
Utilization of d!fferent nitrogen sources by E.typhina and the associa±ed pH changes in the culture media.
Each figure in the table reprooen-bs the mean of 4 cultures (12 for KNO3). Incubation runS at 23°C. in the light.
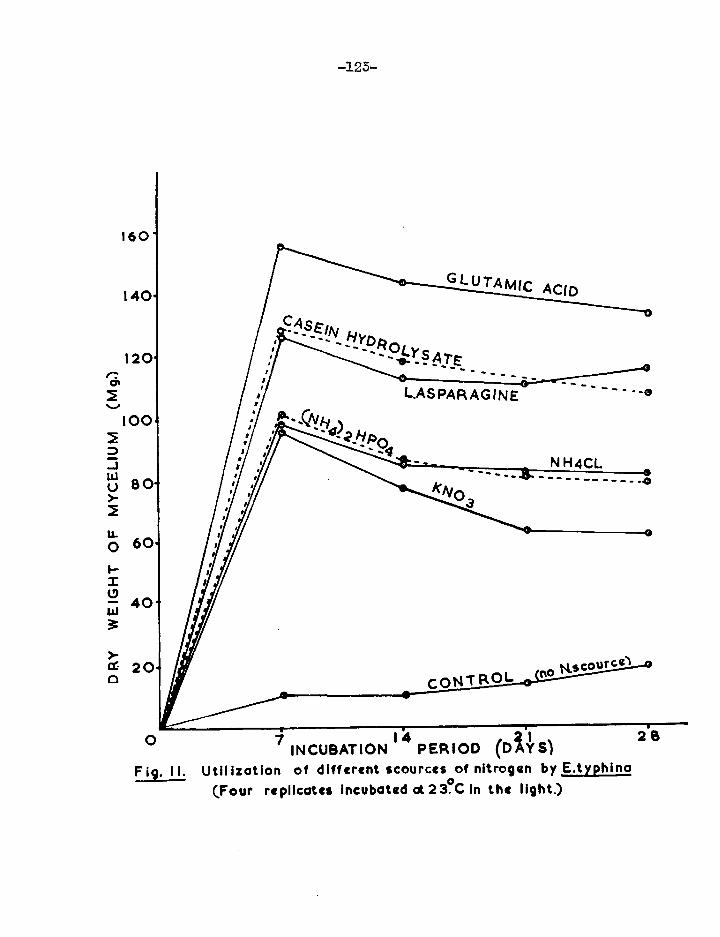
160
140.
120. e-: o% M
100‘
D .7 J W U 80'
M
0 60•
F-
U 172 40.
3
cr 20• ci
-125-
GLUTAMIC ACID
ASEIN HYDRO
LYsATE I
LASPARAGINE 0
......,___________,„--..4
, • CAIly‘Qz
A 1 .
_ „ -----41- NH4CL
c, N.scourcel CONTROL
0 7 I 4 1 u 2B INCUBATION PERIOD (DAYS)
Fig. 11. Utilization of different scources of nitrogen by E.typhina
(Four replicates Incubated ot23°.0 in the light.)

160"
140-
e4 a& M 120- 1 D .71 WI00 U
2
U. 0
80-
0
20-
-124-
0 7 .14 21 INCUBATION PERIOD (DAYS)
Fig.I2. Utilization of different sources of nitrogen byE.typhina. Tour replicates incubated at 23°C. In the light)
23

-125-
Observations were made on the growth produced by E.typhina
on different nitrogen sources and control. The following sorts
of growth were observed.
Control: A thin sheet of slimy, submerged growth.
KNO5: A rather thin mat, covering the whole surface of the
medium. After 21 days incubation some nycelial growth
took place along the sides of the culture flasks.
NH4C1: A thin, white, floating sheet. Uncovered patches of
medium were seen even after 28 days incubation. There
was no growth along the sides of the flasks,
HP04: The surface of the medium wa6 almost but not quite
covered by a thin sheet of mycelium which attained the
height of 5 nn. above the surface of the medium and along
the sides of the culture flasks.
L-Glutanic acid: Avery thick, white sheet of mycelium whose surface
was convoluted. There was about 10 mm. of growth above
the surface of the medium, along the sides of the flasks.
L-Asparagine: A thick, white, floating mat, smooth on top and
covering the whole surface of the medium was observed.
After 21 days incubation, there was growth along the sides
of the flasks which attained a height of 12 mm. above the
surface of the medium, after 28 days incubation,

-126-
Casein hydrolysate: Athick, white mycelium, completely covering
the surface of the medium and rising along the sides of
the flasks to a height of 8 mm,
?Peptone: A thick, white sheet covering the whole surface of the
medium. Growth along the sides of the flasks attained
a height of 8 =4
Gelatine: A thick, white sheet, showing slight convolutions on the
surface. About 15 mu. of growth along the sides of the
flasks and above the surface of the medium was recorded.
On the basis of their effectiveness as sources of
nitrogen, these compounds (see Table 18) could be divided into two
grou ps. 1111 the organic compounds gave a much better growth than
the inorganic and are therefore placed in the first group. The
second group consists of the inorganic compounds, of which KNO5
supported least growth, although it was still fairly good.
All the substances tested. gave a good growth in the first
7 days of incubation, after which there was a slow but steady
decline. The fungus grew best in the cultures containing
glutamic acid and therefore this substance was used as a nitrogen
source in the subsequent experiments. The slight but progressive
growth of E.typhina in the control treatment which lacked a. nitrogen
source, was the subject of a further experiment.

The direction of pH drift associated with each N-source
was related to the type of ion utilized by the fungus. When
cations were utilized, for example, the ammonium ion in NH4C1 and
(NH4)2HPO4'
an equivalent number of anions was left in the
nutrient solution. Since solutions are electrically neutral
(Lilly and Bnrnett, 1951) an equivalent number of both cations
and anions must be present. Thus, when the Na4 ions were
removed from the solutions, an equivalent number of cations {Hf)
was formed from the water, leading to an increase in acidity.
Conversely, removal of nitrate ions from the medium which oontined
KNOi would lead to alkaline conditions arising. The moderately
steep alkaline drift when glutamic acid was utilized may represent
the consequence of the removal of this acid from the medium.

Nitrogen nutrition
(ii) The growth of E.typhina in a medium lacking
a nitrogen source.
In a previous experiment (see Table 16) E.typhinagrew
slightly but progressively in the nutrient solution ostensibly
lacking a nitrogen source. This suggested two possibilities.
Firstly, that traces of nitrogen got into the basal medium as an
impurity in one of the constituent substances. Secondly, that
the fungus was able to obtain usable nitrogen from the atmosphere.
The first of these possibilities was ruled out because the growth
of the fungus increased substantially with time. An impurity
present from the beginning would have soon been exploited to give
an amount of growth which would not have continued to increase
throughout the period of incubation. Concerning the second
possibility, either the fungus could fix atmospheric nitrogen or it
was able to absorb traces of volatile nitrogen compounds present
in the air in the laboratory.
bugger and Davis (1916) showed that Aspergillus niger
did not fix nitrogen. Using an
isolate of A.niger as control, the possibility that E.typhina could
fix atmospheric nitrogen was studied. To this end, a medium of
the following composition was prepared from analar compounds.

-129—
Glucose 1% (weight/Volume)
K2HP04 0.1%
MgSO4.7H20 0.05%
Minor elements
Twenty flasks of nutrient solution were prepared for each
fungus so as to allow five harvesting dates, each with four
cultures. After autoclaving and inoculating each series with a
spore suspension of the appropriate fungus, the cultures were in-
cubated at 25°C. under constant illumination. The results are
shown in Table 19 and Figure 15.
Table 19
The growth of E.typilina and A.niger in a medium lacking
a nitrogen source.
(4 replicates, incubated at 25°C. in the light)
Dry weight of mycelium (mg.) Fungus Days of incubation
14 28 42 56 73.
E.typhina 7.7 9.5 12.9 17.9 17.5
A.niger 5.7 7.'7 8.6 15.1 16.2
Significant differences between means:
P = 0.01 9.2 8.0 8.7 12.2 14.8
P = 0.05 6.3. 5.5 5.8 8.0 9.8

lee
16.
14-
2.
-130-
. 0 14 25 42 5'6 7 .0 INCUBATION PERIOD. (DAYS)
Fig. 13. The growth of E.typhina. and A.niger. in a medium locking a nitrogen source.
(Four replicates incubated at 23°C In the light)

-151—
The total nitrogen content of cultivated fungi has been
found to range from 2.27 to 5.15 per cent. of dry weight 1958).
(Cochrane,. Even if it were assumed that nitrogen formed
5% of the dry weight of E.typhina the maximum growth of the fungus
in 25 ml. of medium (179 mg. in 56 days) would be equivalent to only
0.895 mg. of N. Duggar and Davis (1916) found the bacterium
Azotobacter vinlandii to fix about 40 mg. of N in 100 ml. of
medium after 28 days incubation. Clearly E.typhina if it fixes
nitrogen at all, does not belong to this class.
E.typhina grew slightly better than A.niger throughout
the priod of incubation although there vas no significant
difference between the two. It is possible that the two fungi
obtained nitrogen from impurities in the laboratory atmosphere which
is bound to be contaminated with various types of chemical fumes,
products of combustion of natural gas from laboratory burners and
tobacco smoke (Foster, 1949). The greater rate of growth in both
fungi in the first two weeks as compared with the second two was
probably the result of rapid utilization by the colonies of traces
of nitrogen present as impurities in the chemicals used in preparing
the media, and the inocula themselves. However, only the use of
labelled nitrogen (N15) would convincingly establish whether or not
E.typhina was able to fix atmospheric nitrogen. Nitrogen fixation
has been established in Phoma easy:Irina by Tove, et. al. (1949) by
this method.

3.3a
G. INFLUENCE OF PHYSICAL ENVIRONMENTAL FACTORS.
(1) Temperature
The temperature response of Epichlpe ,typ.hina was ins-
vestigated by Vladimirskaya (1928) who concluded that the optimum
growth temperature was between 16 and 19°C. and the minimum
between 2 and 9°C. She did not establish the maximums Lilly and
Barnett (1949) and Kirby (1958) grew the fungus at 25°C. An
experiment was therefore designed to investigate this important
factor in the growth of B.typhinaft
The following liquid medium was uSeds-
Glucose 1% (wt./01,)
L-Glutamie acid 0.56%
KePO4 04%
MgSO4.7H20 0.05%
Minor elements
The vrtght of glutamie acid 'was ealdUated so as to give
lhoth gram Attic. Weight of nitrogen per litre of medium, as used
successfully in the above described experiment on utilization of
different nitrogen sources (see pagel2A) The glutamic acid
solution was prepared separately at double strength. Its pH was
adjusted to 4.5 before it was combined with the rest of the medium,
which had also been prepared double strength. Inoubation was at

— 133-
8, 18, 200 25 and 30°C., all in the dark. AB the incubators at
8 and 18°C. were illuminated, the cultures to be incubated there
were first wrapped with black paper. To avoid interference with
gaseous exchange, the wrapping papers were formed into chimneys
about 2" high above the mouths of the flasks. The chimneys so
formed. were covered with loose cotton wool plugs. The dry weight
yields of mycelium were determined in quadruplicated sample
cultures taken at weekly intervals. The results of this experiment
are shown in Table 20 and Figure 14.
Table 20
The growth of E.typhina at different temperature$.
(Each figure in the table represents the mean of
4 replicates, incubated in the dark)
Mean dry wt. of mycelium in mg.
....20.erature °C. Days of incubation
7 14 23.
8 4.4 27.7 112.5
18 48.1 164.7 154.4
20 122.8 151.9 143.9
25 145.1 142.1 151.1
30 20.1 62.5 98.9
Significant differences between means,
P = 0.01 20.6 56.8 19.6
P=0.05 14.9 26.6 14.0

140- 25
oC
160.
— 134- —,
1 14 TIME. (DAYS).
Fig. 14. The growth of E.typhina at different temperatures.
(Four replicates. incubated in the dark)
211

-di 135
At 8°C., there was little growth after 7 days incubation
but after 14 days there was a thin and scattered. growth. By the
21st day, the fungus had formed a thin sheet of mycelium over the
surface of the median and a few conidia were seen.
Growth was fast in the first 7 days of incubation at 20
and 25°C., comprising a thick white sheet of mycelium on the
surface of the medium. Conidia were produced. profusely after
7 days incubation. Maxi= growth was attained after 14 days at
20C, and after 7 days at 25°C.
Although at 18°C* growth was slower during the first
7 days than at 20 and 25°C., more was obtained after 14 and 21
days of incubation than at any other temperature. After 21
days, the growth at 18°C. consisted of &thick, white sheet of
mycelium with a yellowish tinge along the edges. Conidia were
produced after 7 days incubation.
At 30C., growth was -very sparse and. only a. few conidia
were seen after the first week of inoubation• There was an in-
crease in the growth rate in the next two weeks but this 'iv-es still
slower than the fastest rates in the other treatments. Through-
out the period of incubation, growth at this temperature was in
the form of small round scattered colonies.
It was vIonclucled from these results that the myeelium of

— 136 —
E.typhina could assume similar, high growth rates over a wide
range of temperatures (note the maximum slope of the growth curves
at 8, 18, 20 and 25°C.), but that it assumed these high rates
earliest at 25 and 2000., and more slowly at 18 and 8°C. However,
within the 5 weeks duration of the experiment the greatest yield.
was at 18°C., although it was not significantly higher than the
yield at 20°C. The treatment at 8°C. had apparently not reached
its maxim, yield at the end of the experiment. The results thus
show that growth rate and ultimate quantity of mycelial growth are influenced separately by temperature.

— 137 —
(ii) Light and dark.
In most fungi, light is reported to affect reproductive
rather than vegetative growth. However the growth of Penicilliun
aL.uctixia and a species of Briarea was depressed by strong, diffuse
daylight (Lilly and Barnett, 1951). These writers also state
that scattered observations indicate that the depressing effect of
strong light may be rather common.
The effect of light on the growth of E.typhina was
studied on a glucose/glutamic acid medium prepared as in the above
experiment (see page 132). A series of cultures was incubated at
18oC. under constant illimination while another series were
wrapped in black pzpol' . as described in the previous experiment.
The dry weight yields of mycelium were determined in quadruplicated
sample cultures at intervals of a few days.
The results of this experiment are shown in Table 21 and
Figure 15. In both treatments, the growth consisted of a white,
thick mat, covering the whole surface of the medium and producing
oonidia after 5 days incubation. There was little difference in
the growth of E.typhina in these two treatments. However, after
14 days incubation, by which time the colonies in both series were
losing weight, the yield from cultures in the Onrk was significantly
greater (P = 0.05) than in the light, though the difference was not

— 138 —
large. There were no signifiCant differences after 5) 7 and 12
days.
Table 21
The effect of lAght and dirk on the growth of E.typhina.
(4 replicates, incubated at 1000
Mean ht of celiura
Treatment Lays of incubation
7 12 14
Light 51.9 167.5 161.2 145.1
Dark 69.6 157.6 169.2 161.5
Significant differences between meant.
P = 0.01 56,9 49.7 21.2 17.1
P 0.05 24,4 32,8 13,9 11.7

20. /
170-
140-
e•120- vI
• • 100- • • •
•
/
/
•
— 139 —
02 4 6 is 10 INCUBATION. (DAYS) 12
14
Fig. 15. The growth of E.typhino in the light and in the dark. (Four replicates incubated at is%)
DARK.
LIGHT.

— 140 —
H. ATTEMPTS TO OBTAIN THE PERITHECIAL STAGE IN
ARTIFICIAL CULTURE.
(i) Introduction
Vladimirskaya (1928) cultivated E.typhina on a variety
of media and found that on fgar agar and malt extract gelatine the
fungus produced structures resembling perithecia, but no asci or
ascospores were formed. Koboyasi (1941) obtained fruiting in
Cordyceps militaris on a rice medium. Both C.militaris and
E.typhina belong to the Clavicipitales. The medium used by
Koboyasi 22_, cit. consisted of 10 g. rice grains and 25 ml.
distilled water in 100 ml. Erlenmeyer flasks. The medium was
sterilized by boiling twice.
Attempts were made in this study to obtain fruiting of
E.typhina in artificial culture.
(ii) On rice medium
Long grain Patna rice was used in this experiment. Ten
grams of rice were plac,7;d in each 150 ml. conical flask, soaked in
25 ml. distilled water for 1 hour and then autoclaved at 15 lbs.
for 20 mins. The method of inoculation was similar to that used
in the physiological expefiments which is described on page 75
The inoculuri consisted of 8 drops of spare suspension --.)r culture
flask. The drops were scattered over the surface of the rice.

— 141
The cultures were prepared in quadruplicate for each treatment and
were incubated under conditions as summarized in Table 22,
Table 22.
Methods of incubation used in attempts to Obtain the
perithedial stage of E.typhina on a. rice mediUm$
Place of incubation Temps °C$ Range Light conditions
Thermostatic incubator 15.0 low intensity )16fse. Thermostatic incubator 18.0 strong ,78f.o• Constant temp. room 25.0 low intensits 1630f 6 C o
Laboratory bench *25.4 (18,8-39.0)4- daylight
Laboratory bench *25.4 (18.8-59.0)+ dark
Greenhouse bench *24.2 (15.4-55.6)+ daylight
Cold frame (on roof) *15,7 ( 8.4-26,5) daylight
Roof (facing west) *15,5 ( 7,5-24.0) daylight
Roof (facing east) *14.6 ( 7.5-24.3) daylight
(*Mean temperature calculated and ranges obtained from readings
taken three times a day, at 10 am., 1.50 and 5 pm. from 15/3/61
to 29/5/61, inclusive.)
(+Occasional very high temperatures were recorded on the
laboratory and greenhouse benches owing to sunlight shining
directly upon the thermometers which were laying on the
benches.)

— 142
In those cultures incubated in the open on the roof,
fain water was prevented from entering through the cotton wool
plugs by covering them with inverted 4" x 1-" boiling tubes,
supported on the sides of the flasks by pieces of plasticine.
The cultures were inoculated on 16/2/61.
After 6 days incubation, there.was good growth in all
cultures except those incubated in the open on the roof. The
growth consisted of a. white mycelium covering the whole surface of
the rice substrate. On the 12th day after inoculation, most of
the cultures showed a yellow colouration in the rice. However,
those in the constant temperature room did not. Further
observations were made at intervals.
32 days: In the cultures at a constant 18°C. and those on the
laboratory bench in the light, mycelium had now begun to grow up
the sides of the flasks.
48 days: Growth of mycelium along the sides of the flasks was
observed in the cultures incubated in the 23°C. constant
temperature room and on the greenhouse bench.
78 days: The cultures on the greenhouse bench showed signs of
d.siee-0:-Uonit The undersides of the media in the cultures on the
roof showed yellow oolouration.
103 days: The rice had begun to shrivel in all the cultures
except those incubated at 18°C., those in the cold frame and those
on the roof.

- 143 -
On the 125th day, all the cultures were taken to the
laboratory for microscopic examination. All had produced
conidia but none bore perithecia or stromata. Mycelial growth
was nornnl.
The rice media were removed from the culture flasks for
examination. The yellow colour they displayed was distributed
throughout the rice in all the cultures, including those from the
constant temperature room in which the yellow colour was not
externally visible. In a similar experiment with C.militaris
Koboyasi (1941) found that a. yellowish colour was formed in the
rice medium after one months incubatIon.
(iii) On cocksfoot stems.
Fresh cocksfoot stems from healthy plants raised from
seed at Silwood were brought to South Kensington in a polythene
bag. The stems were cut into 1-inch lengths. Twenty of these
were put into each of 1C9 ml. conical flasks, to which 0.5 ml.
propylene oxide was added to cold-sterilize the material (Snyder
and Hansen, 1947). The necks of the flasks were then sealed with
cotton wool and made air-tight with two layers of aluminium foil.
After 24 hours at laboratory temperature, the aluminium foil caps
were removed to allow the propylene oxide to escape. 10 ml. of
sterile water were then added aseptically to each flask.

1/14
On 29/12/60, three such flasks were inoculated with an
B.ivphina spore suspension and incubated at 18°Ca in the light.
After 11 days incubation, a thin growth of white mycelium was seen,
No perithecia appeared during the 2 months that the cultures were
kept under observation.
On 28/2/61, three more similar cultures were prepared,
but again no perithecia appeared during the two months the cultures
were incubated.
(iv) On malt extract gelatine.
An attempt was made to secure the proauntion of perithecia
in Epichloe typhina cultured on malt extract gn-:ntine. The com-
prsition of the medium was as recorded by Ainsworth and Bisby (1955).
Malt extract (oxoid) 15 g.
gelatf_-,e (Michrome, bacteriological) 36 g.
distilled water 600 ml.
These constituents were mixed and left overnight. On the
following day, the solution was dispensed into 150 ml. conical
flasks and sterilized by steaming for 30 mins. on three consecutive
days. On the third day (29 /3/61) the cultures were each
inoculated with 8 drops of spore suspension per flask. The
cultures which had been prepared in quadruplicate were incubated
under the following conditions: (Table 23).

— 145 —
Table 25
Methods of incubation used. in attempts to obtain the
perithecial stage of E.typhina on nnJt extract gelatine.
Place of incubation Temp.oC. Range Light conditions
Thermostatic incubator
Constant temp. room
Laboratory bench
Greenhouse bench
Roof (facing west)
18.0 strongs,78f.c.
23.0 low intensity,30f.c.
*25.4 (18.8-59.0) daylight
*24.2 (15.4-55.6) daylight
*15.5 ( 7.5-24.0) daylight
(* see footnote to Table 22)
The cultures were observed over a. period of 60 days.
A white mycelia' growth started on all cultures after 6 days in-
cubation. After 15 days, the cultures on both laboratory and
greenhouse benches showed slight folds in the mycelial mat. Most
of the cultures shored yellowing of the mycelium beneath the nnrgin
of the colonies. Those incubated in the constant temperature
room did not. No perithecia formed in any of the cultures.

— 146 —
1. GERMINATION AND VIABILITY OF SPORES.
Benedict (1929) showed that E.typhina conidia
germinated in 15-18 hours at 18 to 21°C. in distilled or tap
water. At 10oC. only a few conidia germinated after 24 to 50
hours incubation. Kirby (1958) obtained 90% germination of
conidin in tap water after 12-14 hours at 18°C.
In the present investigation, experiments were carried
out to study the germination and longevity of E.typhina conidia
and ascospores.
Attempts to germinate conidia scraped from fresh
stromata were abandoned owing to the inevitable contnnination with
foreign matter. Conidia were therefore obtained from fresh
cultures flooded with sterile water as described earlier (see
page 75) Ascospores were collected from ascigerous stromata by
Ingoldts (1948) method (see page 4V7
Slides on which spores were to be germinated were washed
in a cleaning acid solution (see page? 5f for 24 hours, after
which they were rinsed thoroughly sevdral tines with distilled
water and then stored in alcohol. When required they were dried
between clean sheets of blotting paper. Two drops of spore sus-
pension of a concentration of 1 x 106 spores per ml. were plaemd on each slide.

147 --
(i) Effect of temperature on germination.
To study the effect of temperature on their germination,
conidia and ascospores of Eetyphina mere germinated at 8, 18, 20
and 25°C. The incubators for the first two temperatures were
illuminated ones, the latter two were not. The slides bearing
conidia suspended in distilled water and those bearing thin
deposits of evenly distributed ascospores were supported on V»
shaped glass rods over moist filter papers in clean, sterile Petri
dishes, Two sets of slides bearing conidia and one set bearing
deposits of ascospores were incubated, The two sets of slides
with conidia enabled the percentage germination to be determined
after 18 and 24 hours,
The data for percentage germination of conidia (Table
24> were calculated Prom counts of germinating and non-germinating
coniMe in 20 high power fields of the microscope on each slides
10 from each drop of spore suspension, For each treatment the
total number of conidia observed lay between 665 and 1,270.
Before counting, the coni.dlo were stabled. in cotton blue in
lactophenol,
The data for percentage germination of ascospores
(Table 20 were calculated from counts of germinating and non-
germinating ascospores in 20 low power fields of the microscope

,- 4g r-
whichCh were chosen at random on the slide, or each treatment the
total number of ascospores observed lay between 554 and 574.
Germinating ascospores were divided into three groups aCcording to
the type of germinationt.
n) those producing germ tubes only.,
b) those producing conidia only.
e) those producing germ tubes and. Conidia.
/abe
Percentage germination ofBstnahina conidia incubated at different temperatures in distilled water.
'(Two sets of ,slides incubated/ one set obserVed after each time interval)
Time interval hr.) Temperature -in degrees C.
8 18 ,20 25
18
2,1
0
9,6
•69...6
87,0
82,..2 ,
92.7
81.1
§-14 ,.,

— 149 —
Table 25
Percentage germination of E.typhina gscospores, in
moist atmosphere at different temperatures (after 24 hours incubation)
Type of germination Temperature in degrees C.
8 18 20 25
Germ tubes only 0 60.2 25.5 0
Conidia only 0 6.5 15.6 79.4
Germ tubes + conidia 0 6,6 6.0 0.1
Total germinated 0 1 75.1 44.9 79.5
The germination of conidia was best at 20°C. They
germinated poorly at 8°C. even .after 24 hours incubation (see
Table 24). There vas l!ttle difference in the percentage
germination obtained at 20 and 25°C. but at 18°C. the germination
percentage was slightly lower. The effect of temperature on the
germination of conidia was similar to that obtained by Benedict
(1929)and Kirby (1958).
The germination of ascospores to produce germ tubes day
was best at 18°C. while production of conidia only was best at
25°C. (Table 25)
The production of conidia by germinating ascospores (see

- 150 -
plates 15(a) and (b)) is not an unuausl phenomenon in the
Pyrenomycetes (personal communication with Dr. C. Booth of C.M.I.).
However, the phenomenon does not appear to have been noted before
in E.typhina. It was interesting that the percentage of asco-
spores producing conidia. increased with temperature in the range
18-25°C., while the percentage of those producing germ tubes only
or producing both germ tubes and conidia decreased. In
preliminary experiments in which ascospores were germinated in a
moist atmosphere at laboratory temperature, chains of conidia were
formed (see plate- 13) The conidia from those chains
measured 5.0 to 4.0 by 1.5 to 2.0 )1, which contrasts with conidia
obtained from cultures, which measured 5.0 to 6.0 by 2.0 to 5.0

I
PLATE. 130.(x160)
PLATE.13qx 160).
Conidia produced singly on germinating ascospores ofE.typhina
obtained from infected D.glomerata. plants.
(Ascospores germinating on slides in moist atmospher4, at laboratory temperature)

•
PLATE. I3c.(x 160)
Conidio produced in chains on germinating ascosporcs of E.typhina obtained from infected D.glomerato plants.
Ascospores germinating on a slide in moist atmosphere at laboratory temperature.

- 153 -
(ii) Effect of humtdity on germination
The germination of E.typhina conidia and ascospores was
studied at different humidities at 20°C. To obtain the different
humidities, saturated salt solutions were prepared according to
the method described by Wexler and Hasegawa (1954).
(NH4)2HSO4 R.H. 60.0
KNOB R.H. 95.2%
K2SO4 R.H. 97.2%
Distilled H2O R.H. 100%
Saturated solutions of these salts were prepared in
screw-capped jars, 5 cm. high and 7 cm. diameter. Glass
cylinders 1 cm. high and Ion. diameter were placed upright in the bottom of the jars. One drop of a suspension of conidia was
placed on each clean, sterile coverslip 12 min. diameter. These
were left 1 to 2 hours at 25°C. to dry, after which they were in-
verted and laid on top of the cylinders in the screw-capped jars.
The jars were then sealed and placed in an incubator at 2000. in
the dark.
The same general method was used to study the germination
of ascospores. To collect the ascospores, cleanl .sterile cover-
Slips 12 mm. diameter were placed on slides under ar-izero:us
stromata (see Fig. 2). In the afternoons, when spore discharge is

- 154 -
heavy, a sufficient number of ascospores fell on the coverslips in
a few seconds. The coverslips were then inverted and placed on
glass cylinders in screw-capped jars as described above. With
both conidia and r."ooepores a. further humidity treatment comprised
spores suspended in free water.
The results of the-germination of conidia and ascospores
under different humidity conditions are shown in Tables 26 and 27,
respectively. The data for percentage germination were calculated
from counts of germinating and non-germinating spores in 20 high
and low power fields of the microscope for conidia and ascospores,
respectively. These fields were chosen at random on each cover-
slip. The conidia were stained in cotton blue in lactophenol.
The total numbers of conidia. Observed in each treatment lay between
905 and 1,270; the corresponding numbers of ascospores were 415
and 558.
Table 26.
Effect of humidity on percentage of E.typhina conidia
germinating at 20°C. in the dark.
(One set of coverslips bearing conidia observed after
each time interval)
Time interval (hr.) % Relative humidity. - 80.6 ! 95.2 97.2 100 Free water
18 0 0 1.4 24.6 67.1 24 0 i 0 12.9 28.6 70.5

— 155 —
Table 27
Effect of humidity on percentage of E.ty_phina ascospores
germinating at 20°C. in the dark.
Type of germination %Relative humidity
80.6 95.2 97.2 100 Free water
Germ tubes only 0 0 0 5.8 15.6
Conidia only 0 0 0 27.9 0
Conidia + germ tubes 0 0 0 6.0 0
Total germinated 0 CI 0 59.7 15.6
The data in Table 26 show that the germination of conidia
was best in free distilled water. At 100% R.H., the percentage of
germinating conidia was considerably less than in free water. On
the other hand, the percentage of germinating ascospores at 100%
R.H. was higher than in free water. These were the only two
treatments in which ascospores germinated.

- 156 -
(iii) Longevity of conid*o.
The longevity of Estyphina conidia was tested under
different temperature and humidity conditions. Two drops of a
suspension of conidia from a slope culture were placed on each of
a series of clean, sterile slides. The slides were then left far
1-2 hours at 23°C. to dry.
To investigate the effect of temperature on the
longevity of conidia, such slides were kept at constant relative
humidities in the range 53.8 - 58.5% above saturated solutions of
Na2 Cr2 07.2H20 (Wexler and Hasegawa, 1955), in plastic boxes
(17.5 x 11.5 x 6.0 cm.) sealed with a water-proof 'Parafilm'
tape and stored at 8, 18 and 25°C. in the dark. Wexler and
Hasegawa's data show that the relative humidity above saturated
Na2Cr207.2H20 is affected by temperature. By interpolation from
their data, at 8°C. the R.H. would be 58.5%, at 18°C. 55.0.
From the same data of Wexler and Hasegame.(1955) the relative
humidity ,at 25°C. was 53.8%.
The longevity of conidia at a constant temperature of
18°C. was studied at relative humidities of 34 and 75%, obtained
by the use of saturated solutions of MgC12.2H20 and NaCl,
respectively, (Wexler and Hasegawa, 1955). Slides were removed
from these series of treatments at intervals as shown in Tables

-157 -
28 and 29. A drop of sterile, distilled water was placed on
each dried film of conidia cind the slides incubated for 24 hours
at 20°C. in the dnrk. On each slide, germinating and non-
germinating conidia were counted in 20 high power fields of the
microscope. The fields were selected at random, 10 from each
drop of coniain suspension. From these counts the percentage
germinations were calculated (Tables 28 and 29).
Table 28.
Longevity of.E.typhina conidia in the ark at different
temperatures, at a relative humidity of 55.8 - 58.5%*
(Percentage germination after 24 hours incubation at 20°0. in water)
Storage interval (days)
Storage temperature degrees C.
8 18 25
5 22.9 16.2 14.2
6 4.7 8.5 6.6
15 5.7 4.2 1.8
20 2.8 0 1.2
50 0 0 0
(* 58.5% R.H. at 8°C.; 55.0 R•H,1 at 18°C.; 554 at 25°C.

158 -
Table 29.
The longevity of E.typhina conidia in the dark, at
relative humidities of 54 and 75% at 18°C.
(Percentage germination after 24 hours incubation
at 20°C, in water).
Storage interval (days) R.H. 34% R.H. 75%
5 1067 16.3
6 5.2 9.2
15 104 464
20 0 0
50 0 0
These results (Tables 28 and 29) show that the viability
of conidia decreased: rapidly under all tested conditions of storages
no conidia surviving for 50 days.
The data in Table 24 show that 92.7% of a fresh sample of
conidia germinated after 24 hours incubation. However t&bles 28
and 29 show that within 5 days this proportion was reduced more
than fourfold. Storage tenperature in the range 8 to 25°C, had
little consistent effect on the rate of decline of viability.
The results suggest that viability-declined slightly less rapidly
at 75 and 55.8 - 58.5% R.H. than at 54%,

- 159 -
(iv) Longevity of ascospores
The longevity of ascospores was studied in ways similar
to those described above for conidia. Ascospores were collected
from discharging perithecia onto clean slides. As described on
page148three sorts of germination were recorded, viz., those
producing germ tubes only; those producing conidia only; and
those producing both conidia and ascospores. Germination
percentages were calculated from the counts of germinating and
non-germinating ascospores made in 20 fields selected at random on
each slide. The percentage germinations after 24 hours incubation
at 20°C. in moist atmosphere are shown in Tables 50, 31 and 32.
Table 50
The longevity of E.typhina ascospores at different
temperatures ,at a. constant relative humidity of
55.8 - 58.5%* in the dark.
(Percentage germination after 24 hours incubation at
20oC. in moist atmosphere.)
e of ermination Storage temperature degrees C.
tern tubes only onidia only
rm tubes + conidia
otal germination
8 18 Gb
Storage interval (days) 1
Storage interval (days)
Storage interval (days)
7 2l 55 7 21 55 7 21 55
51.9
9.8
12.1
55.8
0 50.6
15.9
51.5
0.3
0
0
0.3
29.6 2.3.
4.4
36.1
16.9 7.3 4.8
29.0
1.8 0 0
1.8
5.4 57.4
6.8
69.6
20.2 19.8
15.9
55.9
0.3 0
0
0.3
(* see footnote to Table 28)

-- 160.
Virtually all ascospores stored at temperatures of 8) 18
and 25°C. (Table 30) were no longer viable after 55 days. None of
those which did germinate after 55 days storage produced conidia.
Conidia were) however) much in evidence after 21 days storage at 8
and 25°C. though unexpectedly less so at 18°C.
Table 51.
Longevity of E.typhina ascospores stored at 54 and 75%
Relative Humidities at 18°C. in the dirk.
(Percentage germination after 24 hours incubation at 20°C.
in moist atmosphere)
Type of germination Storage Relative Humidity
54% 75%
Storage interval Storage interval (day's) (days)
7 21 55 7 21 55
Germ tubes only 9.8 0 0 23.8 15.5 0.9
Conidia only 7.0 1.2 0 4.3 21.5 0
Conidia + germ tubes 4.5 0 0 61.4 6.6 0
Total germinated 21.3 1.2 0 89.5 41.4 0.9
It was concluded from the data on Table 31 that the
longevity of ascospores was greater at 75 than at 54% R.H. It
would appear that the production of conidia by germinating asco-
spores was adversely affected by the lower R.H. (34%).
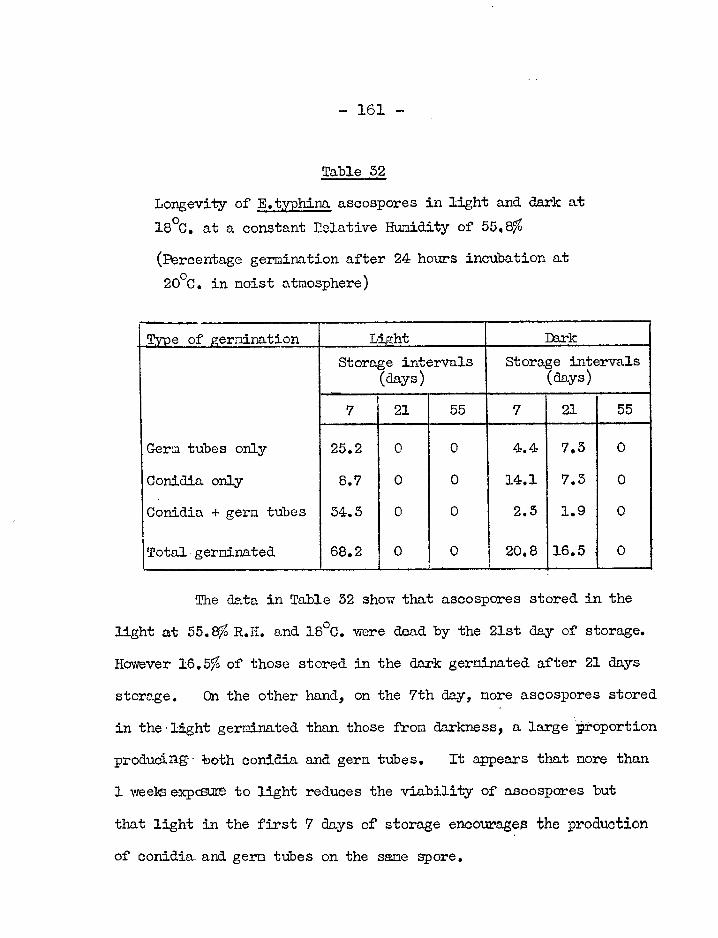
— 161 —
Table 52
Longevity of E.typhina ascospores in Tight and aRrk at 18°C. at a constant Relative Humidity of 55.8%
(Percentage germination after 24 hours incubation at o 20 C. in moist atmosphere)
Type of geruination Light Thrk
Storage intervals Storage intervals (days) (days)
7 21 55 7 21 55
Germ tubes only 25.2 0 0 4.4 7.5 0
Conidia only 8.7 0 0 14.1 7.5 0
Conidia + germ tubes 54.5 0 0 2.5 1.9 0
Total germinated 68.2 0 0 20.8 16.5. 0
The data in Table 52 show that ascospores stored in the
light at 55.8% R.H. and 18°C. were dead by the 21st day of storage.
However 16.5% of those stored in the dark germinated after 21 days
storage. On the other hand, on the 7th day, more ascospores stored
in the.lIght germinated than those from darkness, a large proportion
produc,ng both conidia and germ tubes. It appears that more than
1 weeksexpoant to light reduces the viability of ascospores but
that light in the first 7 cloys of storage encourages the production
of conidia and germ tubes on the same spore.

— 162 —
V. GENERAL DISCUSSION
The ecoLcmic importance of choke lies in damage to crops
grown primarily for seed production (Sampson and Western, 1954).
While the disease has been reported in some years and localities
to cause considerable damage to certain kinds of pastures, it
does not usually reduce the amount of herbage, and appears to do
little damage to pastures generally (Butler and Jones, 1955). The
disease is common on Dactylis glomerate crops grown for seed and
those intended for hay or pasture.
In attempting to check a disease of plants, it is
usually rocessary to know the cause of the disease, and if the
disease is infectious, the life history of the causal organism and
the circumstances which influence the establishment of parasitic
relations between it and the host (Butler and Jones, 1955). One
of the objects of the present study was to determine the means by
which healthy cocksfoot plants become infected with E.typhina.
Insofar as it was relevant to this, the germination and longevity
of conidia and ascospores of the pathogen were studied. Further,
rn In vitro study was undertaken to establish the basic nutritional
requirements of the fungus, and to attempt to secure the production
of its perithecia.

- 163 -
The most notable result from the inoculation
experiments carried out during this study was the almost complete
failure to transmit the disease through vegetative and floral
parts of the plant by spores and mycelia. The only experiment
in which the result indicated a possible route of infection was
the one in which spores were placed on the cut ends of flowering
stems of cocksfoot under humid conditions. Hyphae originating
from such spores travelled considerable distances down the stem.
For the greater part of the year, V.tvphina develops as
an intercellular parasite, living entirely within the host.
Sampson (1935) found mycelium in infected plants at all times of
the year. However prior to the formation of the conidial
stromata, the mycelium becomes abundant in and between leaf-sheaths,
binding them together in a solid mass, and preventing the exsertion
of the inflorescence. Kirby (1958) found that this vigorous
fungal growth was associated with floral initiation. He observed
this vigorous growth at different times of the year and in widely
varying environments, and concluded that it was not caused by
seasonal factors acting directly on the fungus. He suggested
that sone change in the plant, associated with the transition
from the vegetative to a flowering condition, encouraged the rapid
growth of the fungus.

- 164 -
It is an attractive hypothesis that the growth of
E.typhina. is partially inhibited in the vegetative plant and that
changes associated with flowering involve the disappearance of the
inhibitor, thereby malemg the plant tissue more favourable to the
development of the fungus. This might also explain why attempts
to inoculate vegetative plants failed. The fact that the fungus
grows readily in amide range of media. yet is most difficult to
establish in a. vegetative grass plant further supports the
possibility that conditions in the vegetative plant are inhibitory.
The fungus grew in inoculated, flowering stems (see page42), as
also reported by Western and Cavett (1959). This would imply
that the hypothetical inhibitor had disappeared.
That substances occurring naturally in plants can in-
fluence infection has been shown by various workers. Walker and
Link (1957) found that certain phenolic compounds occurring in
brown varieties of onion inhibited the growth of fungi pathogenic
to onions. Wurinnn (1950) states that the phenolic compounds in
the cell sap of brown onions contain fungicidal substances such as
catechol and protocatechuic acid. He stated that the germination
of Colletotricun circinans and Fusarium cepae is inhibited by these,
even in dilutions of 500 to 1,200 parts per million, respectively.
Defago (1940), who studied the influence of thiamine and hetero-

- 165 -
auxin on the growth of wheat parasites to ascertain whether certain
anomalies in the biology and parasitism of Tilletia tritici were
explicable on the basis of the production of auxogenic substances
by the host, found that the in vitro development of T.tritici and
Carcosporella herpotrichoides was delayed by indole-3-acetic acid
at 1 and 150 microgram per 25 ml., respectively.
The initiation of flowers has been investigated by
Leopold and Thimann (1949), who showed that the process was
associated with low auxin concentrations. They found that
flowering was initiated in ITTinte).c barley by applying a very low
concentration of 0(-naphthalene acetic acid (1 mg. per litre or
less). When the concentration of this substance- was 400 mg. per
litre, flower initiation was inhibited. Khudna and Hamner (1955)
found that flowering in Xanthiun pennlylvanicum was brought about
by a decrease in auxin content. It is generally agreed that
flowering is associatelvith a low auxin level (Kirby, 1958).
Since auxins do influence fungal development and also
do change in concentration in relation to flowering, the onset of
flowering could conceivably prove favourable to the growth of a
parasitic fungus.
E.typhina normally grows only in intercellular spaces,
becoming intracellular only at the time of flowering (Sampson,1955),

- 166 -
It is possible that within the cells of a vegetative plant, growth
is inhibited. It would be interesting to study the germination
of E.typhina spores in cell sap expressed from flowering and non-
flowering grasses.
The almost complete failure to transmit choke through
vegetative parts of the plant, and the successful growth and
establishment of the fungus on cut flowering stems, as reported
here on page 39, and also by Western and Cavett in 1959, could
be explained on a nutritional basis as well as by the above
hypothesis of an inhibitor. Kirby (1958) states that the
organization of the shoot apex of grasses changes greatly when
flowering is induced and that this must be a reflection of changed
patterns of metabolism. A critical, qualitative or quantitative
nutritional change associated with flowering might make the tissues
of the stem more favourable to the growth of the fungus.
The hypotheses erected to explain why the disease could
not be transmitted through the vegetative parts of the plant
(viz, inhibition or unfavourable nutritional conditions in the
vegetative plant) do not assist in understanding why infection
through the flowers themselves and their seed has so far failed in
cocksfoot. The fact that transmission through seed has been
established in F.rubra_ by Wernhau (1942) suggests that the forms of

- 167 -
E.typhina on D.glomerata and Fambra differ at least in the way in
which they are transmitted.
As a third_ possibility, it is suggested that the failure
to artificially infect healthy plants is a result of the existence
of distinct, highly host-specific strains of the fungus, to which
only particular host species, or indeed, strains of those host
species are susceptible. Diehl (1950) suggests that such
susceptibility of the host to a particular strain may explain why
in any one locality a particular species of the Balansiae (a.
group related to Epichlco) is found almost invariably upon one
host species only, although other grasses, known as hosts else-
where, may beErowing nearby.
Nhen a moist air current was experimentally passed over
conidial stronataa conidia mere detected in the air stream.
ascospores were liberated in large numbers from mature
perithecial stromata. Such spores being light could easily be
carried about in air currents. It therefore seems unlikely
that it was because spores failed to reach them that healthy
plants were not infected from adjacent diseased ones.
Natural infection by E.typhina reduced the length and
weight of culms, and the germination of seed. Such an effect on
the culms was perhaps not unexpected, since in addition to

-168-
supporting their own growth and development, the infected tillers
had to maintain the parasite within. Bradshaw (1959), studying
a population of .grostis sp. infected with E.typhina, found that
the total weight of the plant was slightly reduced by infection.
Diehl (1950) states that all the hosts infected with Balansia,
Atkinsonella and Balansiopsis, as well as E.typhina (all of which
belong to the sane sub-family Clavicipitoidae of the
Clavicipitaceae), tend to be dwarfed, and some are fasciated.
Internodes are shortened.
In infected plants of different species, the number of
tillers which contained mycelium varied considerably. Infected
plants of D.glomerata had a fairly high percentage of fungus-free
tillers. H.mollis and F.rubra had low percentages of fungus-
free tillers suggesting that when these two species are attacked
by the fungus, the mycelium tends to permeate all the tillers on
the plant. It is not known whether this is because there are
strains of the fungus at different virulence. Sampson (1955)
however, stated that choke is not biologically identical in all
host species. It wou-1 therefore be interesting to cross-
inoculete plants of different species through flowering stems.
Cultivated on different media,. E.typhina grew best on
corn meal/glucose peptone liquid in medium. There was no simple

— 169 —
relationship between the parasitic growth of different isolates,
as assessed by the number of completely choked tillers on their
host plants and their rates of saprophytic growth on corn meal/
glucose peptone medium, in terms of weight. The results
obtained in this experiment supported the contention that the
fungus is not biologically the same in all host species,
different isolates differing much in their rates of growth on this
culture medium.
When the three potassium salts of orthophosphoric acid
were separately incorporated in the same synthetic basal media,
E.typhina grew fairly well with all, despite the fact that the
initial pH ranged from 3.5 to 8,4.
It was demonstrated in experiments with growth factors
that:
a) the isolate of E.typhina from 110.glomerata had a partial
thiamine deficiency at low temperatures (8 and 10°C.).
b) the isolate of E.typhina from H.mollis was slightly thiamine
deficient at 25°C.
c)the addition of thinnine to media at 10, 18 and 25°C. did not
enhance the growth of E.typhina isolates from F.rubra and
A. stolonifera.
Clearly the isolates were different in their thiamine

- 170 -
requirements. This is not an unusual phenomenon in species of
fungi. TAlly and Barnett (1949) state that rlif'ferent isolates
of the ;game fungal species nay have different vitnnin deficiencies.
For example, Leonian and Lilly (1942) demonstrated this in ten
strains of Saccharomyces cerevisiae. Burkholder (1943) similarly
reported strains of yeast differing in their vitamin requirement.
According to Robbins and Kavanagh (1942)0 the
deficiency of a fungus for a specific vitamin may be absolute or
conditioned, In a conditioned deficiency, the synthesis of the
vitamin may be influenced by certain environmental conditions such
as temperature or composition, concentration and pH of the medium.
For example, Mitchell and HoUlahan (1946) reported a deficiency
for riboflavin in a mutant of Neurospora which was conditioned by
temperature while Rqrnett and Telly (1948) reported the snme in
Sclerotinia camelliae which is partially deficient for inositol.
Clearly the deficiency of the E.typhina isolate from
D.glomerata here studied was conditioned by temperature. Only
when cultivated at 8 and 1CbC., was the growth in cultures without
exogenous thiamine less than that in cultures with thiamine.
The isolates of E.typblna from F.rubra and A.stolonifera
showed no thiamine deficiency, total or partial, absolute or
condition. It is therefore assumed that these isolates were able

— 171 —
to synthesize enough thiamine for their need in the conditions
under which they were cultivated. However the isolate from
H.mollis showed a partial deficiency for thiamine at 25°C.
Just as the isolates here studied differed among
themselves in respect of their need for thiamine, so too all
differed from the isolate studied by Lilly and R-rnett (1949).
This latter was almost totally thiamine deficient.
Apart from Lilly and Barnett's (1949) study on E.typhina,
there is little information in the literature about its
physiology. Rather than study the behaviour of a series of
different isolates of E,typhina, it was decided to investigate in
more detail the influence of nutrition and physical factors on
the growth of the E.typhina isolate from D,glomerata onlyi
Experiments showed that Eityphina was able to utilize a
variety of carbon sources. Of the 9 carbon sources of different
complexity tested, the fungus grew well on six. The greatest dry
weight was produced with sucrose and the least growth with pectin.
Growth was also poor with lactose and cellulose.
Fructose and glucose are usually equally effective as
sources of carbon for fungi but this is not always the case.
E. yphina grew more slowly when fructose was used as sole carbon
source than when glucose was used. Maximum growth in weight was

— 172 —
attained with glucose after 7 days incubation but, with fructose
after 14 days. In a paper cited by Lilly and Rrnett (1951)
Diplodia macroapora, Phytophthora cactorum, P.erythroseptica and
P.fagopyri have been shown to differ in growth with these two
C-sources.
E.typhina clearly utilized all the N-sources here tested
both organic and inorganic. The maximum dry weight yields
Obtained from the media with organic sources of nitrogen ranged
from 126.9 —155.5 mg. and for media containing equivalent amounts
of inorganic sources of nitrogen, 95.5 - 101.0 mg.
Growth was poorest with KNO3. The moderately good
growth on the two media containing ammonium salts (NH4C1 and
(NH4)2HP04) were remarkably similar, there being no significant
differences in the weights of growth obtained, except after 7 days
incubation (P = 0.05). L-glutamic acid supported significantly
better (P = 0.01) growth than the rest of the tested N-sources.
The amcnint of nitrogen provided to the fungus in each
tested N-source was made the same. However the inorganic sources
of nitrogen were also sources of adeitional carbon in the culture
media. They also presented nitrogen in an already reduced form,
so that, unlike growth on nitrate, expenditure of energy on
reduction was not entailed. Growth with the organic sources may
have been greater for these reasons.

— 173 —
As was expected, KNOB and the two ammonium salts had
opposite effects on the ultimate reactions of the culture media.
As nitrate ions were consumed, the culture medium became more
alkaline while as ammonium ions were utilized the culture media
became more acid. All the organic sources of nitrogen tested
led to an increase in alkalinity of the media. With glutamic
acid, increase in alkalinity could have resulted from the removal
of the acid from the culture medium.
E.typhina was found to grow slightly and slowly in the
cultures which had not been supplied with a source of nitrogen.
After 28 days incubation, a growth of 19.8 pg. was recorded in
25 ml. of medium. This raised the question as to whether or not
the fungus was able to fix atmospheric nitrogen. Duggar and
Davis (1916) found that saprophytes such as 19pergillus niger,
Penicillium expansum and P.digitatum were able to fix about 1-2 mg.
of nitrogen per 50 ml. of medium, but considered such an amount of
N to be within the limits of experimental error. However they
found that Phoma betae which is cogeneric with the fungi of
Eppaceolls mycorrhizo, fixed 3.0 - 7.8 mg. N in 100 ml. of medium.
They considered that this was beyond the limits of experimental
error and that it was strong evidence for N fixation in this
species.

- 174 -
In a. further experiment described above, E.typhina was
cultivated in a medium lacking a nitrogen source, while A.nigerl
as representing a fungus incapable of fixing atmospheric
nitrogen (according to Duggar and Davis, 1916) was used as a
control. The maximum weight of growth obtained for E.typhina
after 56 days incubation was slightly less than that obtained.
previously after 28 days incubation. The growth of Aoniger was
less than that of E.typhina throughout the perioa of incubation
(71 days) but not significantly so. It is therefore conclude&
that there is no clear evidence of fixation of molecular nitrogen
by E.typhina. The snail amounts of growth in ostensibly nitrogen-
free media could possibly have been supported by traces of
nitrogenous impurity, or by the utilization of combined nitrogen
dissolving in the medium from the atmosphere, in which it was
present as a contaminant (e.g. vapours,. smoke eto.). Even
assuming that the weight of nitrogen comprised-2% of the dry
weight of E.typhina (Cochrane, 1958) (see Fage131), the maximum
yield in weight of the fungus obtained in this experiment would
contain only 0.89 mg, of nitrogen.
Although the abjlity to utilize gaseous-Vas the sole
source of nitrogen for cell. synthesis has been chimed form
variety of fungi, critical examination of the evidence indicates

- 175 -
that clear-cut N-fixation is at best a property of very few fungi,
these being mycorrhizal or near mycorrhizal types (Foster, 1949).
However cogent evidence for N-fixation by species of Mona has
been obtained by Ternetz (1907); Duggar and Davis (1916);
Rayner (1922); JWIles and Smith (1928) and Tove et.al. (1949).
Butler and Jones (1955) considered E.typhina as living
in symbiosis with grasses, insofar as it sometimes fails to
fructify even when the grass which it has systemically infected
has grown to maturity. Bradshaw (1958) who investigated the
incidence and significance of infection of Agrostis sp. by
Epichloe typhina, concluded that infection was not an example of
parasitism but a curious form of symbiosis. He found that the
tiller density vas significantly increased by infection even though
the total weight was slightly reduced.
It seems parnaoxical that a fungus should infect a plant
and sometimes permeate almost all tillers (see Page 68) without
seriously reducing the vigour of the plant. It is more difficult
to explain why more tillers are formed on infected plants, as
reported by Bradshaw (1958). One may tentatively speculate that
E.typhina in its hosts fixes atmospheric nitrogen and thereby
causes less harm to its host.
Techniques using labelled nitrogen (45), constitute the

- 176 -
only satisfactory test for N-fixation. By such means, nitrogen
fixation has been demonstrated in Phoma casuorinar by Tove et. al.
(1949). Foster (1949) states that this technique would provide
a. check on claims of fixation of very small amounts of nitrogen
by saprophytic fungi. Eftyphina is an endophyte, and, so far,
1$-fixation appears to have been demonstrated only in endophytic
fungi. An investigation by means of labelled nitrogen would
seem worthwhile.
Cultivated in a glucose/glutamic acid medium, E.typhina
here produced the greatest yield of mycelium at 18°C. However
the yield was not significantly higher than that obtained at 20°C.
(see Table 20 and Fig. 14). This result is in agreement with
that of Vladimirskaya (1928) who found the optimum temperature to
be between 16 and 19°C.
The growth rate of the fungus was higher at 20 and 25°C.
than at 18°C., as shown by the steeper slopes of the growth curves.
This suggests that the growth rates and the ultimate quantity of
mycelium were influenced separately by temperature.
There was little difference in the growth rate or the
quantity of mycelium produced when E,typhina was cultivated in a
glucose/glutanic acid medium in light and dark (see Table 21 and
Figure 15). After 14 days incubation, the mean pydelial yield

— 177 —
from the cultures incubated in darkness was significantly greater
= 0.05) than the yield from cultures in the light, but by then
the colonies had entered the autolytio phase. Further, the
difference was not a large one.
There are reports in the literature, of light depressing
growth of fungi, for example, Karlingia (Rhizophlyotis)roseal one
of the Chytridiales (Haskins and Weston, 1950), and Sclerotinia
fructigena (Hall, 1955), However there is no indication that
actively growing colonies of E.typhina do so faster in the dark
than in light. All attempts to secure the production of asco—
spores by E.typhina in artificial culture were unsuccessful. On
ricer pieces of cocksfoot stems and malt extract medial the fungus
produced copious, white, aerial mycelium and abundant conialn.
A yellow colouration was observed in most of the rice cultures
after 6 days incubation.
Ferithecial stromata of the allied fungus, Cordyceps
militarist were obtained by Koboyasi (1941) and Rnsith (1961)
who cultivated the fungus on rice media. Basith stated that,
in darkness and in red light, rice cultures remained wholly
vegetative, producing copious, white, aerial mycelium. In white
and green light C.militaris praliced perithecial stromata. Lilly
and BrIrnett (1951) state that sexual reproduction in the fungi,

- 3278 -
with all the necessary preliminary metabolic activities, is a
complex phenomenon. There are many factors which influence it.
Examples are temperature of incubation, pH and concentration of
medium, presence of vitnnins, micro elements, culture filtrates
and compounds such as glutaric acid which has been shewil to be
influentTn1 in certain cases. The addition of certain specific
sexual factors or hormones is known to be effective for certain
Phyconycetes. Aeration, and intensity and wavelength of
illumination may also be decisive.
Clearly the attempts made here to induce fruiting in
E.typhina do not exhaust the possibilities that night be investi-
gated.
The germination and longevity of E.typhina conidia and
ascospores was studied under different temperature and humidity
conditions.
The highest percentage germination of conidia was
obtained when they were incubated for 24 hours at 20°C. in
distilled water (see Tal-le 24), altl-Dugh there was little
difference between the percentage here and that obtained at 25°C.
The conidia nevertheless germinated well after 18 hours incubation
at all the temperatures tested except 8°C., where no germination
was observed. However when incubated for 24 hours at 8° C.,

179 -
9,0 of the conidia then germinated. The lowest relative
humidity permitting germination in a humid atmosphere as opposed
to in free water, lay between 97.2 and 95.2%. Germination at
Ng% somas however, still less than half that in free water.
Ascospores germinated in one or other of three ynyss
either producing germ tubes only, or producing conidia only or
producing both conidia and germ tubes.
Although germ tvbess conidia and both gerri tubes and
conidia were produced at 100% R.H., after 24 hours incubation at
20°C. (see Table 27) no ascospores germinated after 24 hours at
804, 93.2 and 97. cR.H. In free water, 15.6% of ascospores
were found to produce germ tubes but no other type of gernination
was Observed.
The direct production of conidia by germinating asco-
sporess.was previohnly observed on cut' flowering stems (see
Plate 2 ) and on slides (see Plate 13). However neither the
conditions for their production nor for their germination were
studied here. Their possible role in the life-cycle of the
fungus therefore remains unknown.
Men the longevity of con-T.dia was tested in the clerk at
8, 18 and 25°C., at relative humidities of 58.5, 55.8 and 55.8%
respectively, viability decreased rapidly and no conidia

- 180 -
survived after 20-30 days. The viability of conidia endured
longest at 8°C. and 58.5% R.H. However their viability was
greater at 75% R.H. than at 30 at 18°C, in the dnrk.
The decrease in the viability of ascospores stored
under various conditions of temperature, relative humidity and
light and dark (Tables 30, 31 and 32) was such that there were
virturOly no viable ascospores after 55 days.
Western and Cavett (1959) studied the viability of
ascospores and obtained results similar to those described above.
For three seasons they stored collections of perithecial stromata
under various conditions, under partial cover and in the open air.
They obtained sinilar results in all three years, namely: very
high germination capacity in July followed by a steady diminution
of viability until November when only 14% of the stronata yielded
any viable spores at all and these were few. It therefore
seemed unlikely that B.typhina overwinters in the form of asco-
spores.
Gottlieb (1950), reviewing spore germination in fungi,
stated that temperature influences the longevity of spores and
that a long period of senscence is correlated with low
temperatures, while high temperatures shorten this period. He
also stated that high humidity tends to shorten the priod of

- 181 -
viability of spores, but for a nunber of species, humidity has
not been observed to have an adverse effect. For example, the
spores of Actinomyces scabies were unaffected by moisture between
12 and 50°C. Gottlieb further stated that when ascospores are
enclosed in a fruiting body, such as a perithecium, they retain
their viability longer than when removed and stored on glass.
This situation might well exist with E.typhina. The viability
of ascospores on slides was here found to last only about
55 days, while those stored-by Western and Cavett (1959) in
perithecial strorinta, lasted about 120 days (July - November).
In view of the results obtained here on the longevity
of spores under various conditions of temperature, hun3dity and
light, and those Obtained by Western and Cavett (1959) for asco-
spores stored in perithecial stromata under various conditions,
it seems improbable that the fungus overwinters in the form of
spores. It is more likely that it perennates in the form of
mycelium within the living host.
E.typhina has been propagated in D.Flonerata only by
dividing infected plants (Sampson, 1955), and has been artificially
transmitted only through cut flowering stens. As reported here,
this did not lead to stroma production, but Western and Cavett
(1959) Succeeded in this respect. However it is still not clear

-a 182 —
how infection of D.glomerata takes place in the field where plants
may not be propagated through root divisions nor may expose their
internal tissues as a result of being cut back periodically, as
in seed-production fields.
The mode of transmission in other host species is
likewise largely unknown, although seed transmission has been
demonstrated_ in F.rubra (Wernham, 1942). Western and. Cavett
(1959) state that stromata of E.typhina appear to be relished by
insects and other soil animals and that this nay have a bearing on
the spread of the disease. Clearly further investigation on the
transmission of choke is warranted.

— 183 —
VI. SUMMARY.
1. Attempts to transmit Epichloe typhina through vegetative
tillers, tiller buds, leaves and flowers of healthy
cocksfoot plants were unsuccessful. The following
inoculation methods were employed:
a) spraying cut stems with spore suspensions,
b) injecting spores and bits of mycelium into stems,
c) washing ascospores into leaf-sheaths,
d) placing spore suspension drops on pieces of detached
leaves,
e) dusting flowers with conidial stromata from infected
cocksfoot plants,
f) the partial vacuum method (Moore, 1936).
2) Ascospores freshly obtained from choked cocksfoot plants
germinated and produced conidia in situ when placed on cut
ends of flowering stems of cocksfoot plants in a moist
atmosphere. Byphae from such ascospores were traced several
centimetres down the pith of the stems, beyond nodes through
which the hyphae must have grown. No fungus was observed in
control material. No stromata or flowers were produced on
any of these plants.

- 184 -
3. Myceliun was revealed in pith scrapings from flowering stens
of healthy cocksfoot plants which had been cut and inoculated with E.typhina from cultures. None was present in the
controls, No stromata or flowers were produced on any of
these plants.
4. No stromata mere produced by cocksfoot flowering stems
inoculated with conidia and ascospores under field conditions
in 1960. The plants bearing the inoculated stens flowered
heavily in 1961.
5, Only a few conidia were shed when a current of air was
passed over conidial stromata; more were obtained when the
current was moist.
6, The lengths of spores of E.typhina discharged from perithecia
on—D.glomerata, H.lanatus and A.stolonifera collected at
Silwood Park were 161.7, 52.5 and 10.7)i, respectively.
The breadth was unifermy 2m„
7. The maximum rate of disCharge of ascospores of E.typhina from
ascigerous stromata from D.glonerata, H.lanatus and
A.stolonifera occurred beLween 6 and 10 p.m.
8. On tillers of b.glemeratar Hemollis and A.stolonifera, the

- 185 -
stromata sometimes formed a loop protruding from the leaf-
sheath. Besides the usual stroma in a typical position,
tillers sometimes carried several others on shoots arising
from tiller buds on the same stem.
9. The formation of "double-stromata" is described. These
consisted of a true strona. at the top of the culm and a
diseased patch on the she4th of the leaf arising from the
second node below the true stroma.
10. The mean lengths of cuims from D.glomerata and H.mollis
plants visibly infected with E.tvohina were significantly
less (P = 0.05) than those from apparently non-infected
plants of the corresponding species, but were the same with
F.rubra. The fresh weights of panicles of infected plants
of D.glomerata, F.rubra or H.mollis were about half those of
the corresponding apparently non-infected hosts.
11. The fresh weights of seeds from visibly infected D.glomerata,
F.rubra and H.mollis plants were slightly less than those
from apparently non-infected plants. The percentage
germination of seed of D.glomerata and F.rubra from
apparently non-infected plants were considerably higher than
those of seed from visibly infected plants. In H.mollis the
difference was small.

— 186 —
12. No mycelium was seen in those stems of D.glonerata plants
infected with Eityphina whidh bore healthy flowers. The
proportion of tillers free from mycelium was higher in
infected plants of I4glomerata than in those of H.mollis or
in Which practically. all tillers contained the
fungus.
l3. Of a number of liquid dui-hire media, E.typhina erodUced the
greateSt yield of mycelihm in corn meal/glucose peptone.
14. There was no simple relationship between the growth of
EstYphina- isolates from D.glomerata, P.rhbra. and A.stolonifera,
in corn meal/glucose peptone medium and on the respective
hosts as assessed by the proportion of coMpletely Choked
tillers. An isolate of the endophYte from Loliun perenne
cultivated on the same mediUm prodhded no spores.
15, E,typhina produced more donidia on slope cultures of
glucose.-peptone agar than on malt, corn meal, pea extract,
Otapek s or Coon's agar.
16. E.typhina grew well in media containing KH2PO4„ K2BP04 or
K5 PO4; • Ky04 sustained the greatest yield.
17. The isolate of E.typhiiia from D.Romerata did not respond to

— 187 —
thiamine, biotin or yeast extract in culture media at 18 or
25°C. but at 8 and 10°C. thiamine significantly increased
its mycelial yield (P = 0.05).
18. Thiamine significantly increased the yield of mycelium of
Phycomyces blakesleeanus (P mg 0.01) but not of Epichloe
typhina (2ED.glomerata), when the two fungi were cultivated
in media of the same basic composition at 25°C. in the dark.
19. E.typhina grew well vegetatively in media containing sucrose,
glucose, galactose, arabinose, fructose and starch as C
sources. The yield of mycelium was poor when cellulose,
lactose or pectin was used.
20. Of the nitrogen sources tested, glutanic acid gave the
greatest yield of mycelium. Organic N-sources were better
than inorganic. There was a small but persistent growth
in the control treatment lacking a nitrogen source.
21. Both Epichloe typhina and Aspergillus niger grew slowly but
progressively in a medium ostensibly lacking a nitrogen
source, but the growth of A.niger was slightly inferior
throughout the period of incubation (71 anys).

-188-
22. The rate of growth and the ultimate mycelia' yield of
E.typhina are influenced separately by temperature in the
range 8-5000. Yield was greatest at 18°C. after 14 and
21 days incubation.
23. Light had little effect on the vegetative growth of E.typhina
although after 14 days incubation the yield in the dark was
very slightly greater (P = 0.05) than in the light.
24. Attempts to obtain the perithecial stage of E.typhina on rice
cocksfoot stems and malt extract gelatine mediawre un-
successful, though all supported copious vegetative growth.
25. After 24 hours incubation in distilled water, conidia of
E.typhina germinated most abundantly at 2000. At 80C. only
9.6% germinated. Germination at 100% R.H. was less than
half that observed in free water. A. few conidia germinated
after 24 hours incubation at 97.2% R.H. but none at 95.2%.
26. E.typhina ascospores germinated in one of the following three
Ways: germ tubes only,
conidia only,. or
conidia and gem tubes.

— 189 —
27. After 24 hours incubation in a moist atmosphere, germinating
E.typhina ascospores produced germ tubes best at 18°C.
while they formed conidia best at 25°C. Ascospores did not
germinate at 97.2;10 R.H. and less. Germination in all three
ways was best at 1000 R.H., except for the production of
germ tubes which was highest in free water. In free water,
no other type of germination was observed.
28. The viability of conidia endured longest at 8°C. in the dark
at 58.5% R.H. At 18°C. in the dark, it was longest at
75% R.H.
29. Virtually all ascospores did not germinate after storage in
the anrk at temperatures of 8$ 18 and 25°C, at humidities of
58.5, 55.8 and 53. for 55 days respectively# The longevity
of ascospores at 18°C. in the dark was greater at 75% R.H.
than at 54%.
30. Exposure to light for more than 1 week reduced the viability
of ascospores in comparison to those in darkness but in the
first 7 days of storage it appeared to encourage the
production of conidia and germ tubes on the sane spore.

— 190 —
VII ACKNOWLEDGEMENTS.
I wish to express my indebtedness to the late
Dr. C.C.V. Batts who suggested the problem and who gave me much
advice in the earlier part of this study.
I also wish to thank Dr. R.K.S. Wood and Dr. M.F.
Madelin for their guidance and unstinted help during the course
of this work and for their helpful criticism during the
preparation of the manuscript.
My thanks are also due to Mr. R. Adams for his
technical assistance, Mr. A. Horne for the photographic work and
Mr. E. Green for help in various ways.
I wish to acknowledge the financial support received
from the Government of Kenya, which enabled me to carry out this
work.

— 191
VIII. LITERATURE CITED
AINSWORTH, G.C. & BISBY, G.R. (1945). L dictionary of the fungi. pp. 359. The Imperial Mycological Institute, Kew, Surrey.
ANNONTIOUS. (1953). Difco manual of dehydrated culture media and reagents for microbiological and clinical laboratory procedures. 9th Edition. pp.350. Difco Laboratories Inc., Detroit 1, Michigan.
BARNETT, H.L. & LTLLY, V.G. (1948). The interrelated effects of vitamins, temperature and pH upon vegetative growth of 5clerotinia camelliae. Amer. J. Bet. 35: 297-302.
BASITH, M. (1961). An experimental study of the nutrition of Cordyceps militaris (Fr.) Link. and certain aspects of its life-cycle. D.I.C. Thesis, Imperial College of Science and Technology, London.
BENEDICT, D.M. (1929). A greenhouse study of the conidial stroma of Epichloe typhina. Pap. Mich. Acad. Sci.„ 9, 47-54.
BISBY, G.R. (1955). An introduction to the taxonomy and nomenclature of fungi. pp.143. Commonwealth Mycological Institute, Kew, Surrey.
BRADSHM, A.D. (1959). Population differentiation in Agrostis tenuis Sibth. II. The incidence and significance of infection by Epichloe typhina. New Phytol., 58, 510-315.
BREFELD, O. (1891). Mykologie, vol. 10. 191, Leipzig.
BUCKHOLDER, P.R. (1945). Vitamin deficiencies in yeasts. Amer. J. Bot. 50 206-211.
BUTLER: E.J. & JONES, S.G. (1955). Plant pathology. pp. 979. Macmillan & Co. Ltd., London.

- 192 -
CARRUTHERS, W. (1903). Annual report for 1905, of the consulting botanist. J.R. Agric. Soc., 64, 302-505.
TICK, W.J. & POPE', W. (1950). A modification of Moorets method of inoculating wheat and barley with Loose Smut. Phytopath. 40, 1054-1060.
COCHRANE, V.W. (1958). Physiology of fungi. pp. 558. John Wiley & Sons, Inc., New York.
DE GARY, A. (1887). Comparative morphology and biology of fungi, mycetozoa and bacteria. The Clarendon Press, Oxford.
IEF&GO, G. (1959). Influence de l'aneurine et de heteroauxine sur la croissance de trois parasites de Ble'. (The influence of aneurine and heteroauxin on the growth of three wheat parasites). Ber. Schweiz. bob. Gez.$.49, 413-414. (Abstr. in Rev. appl. Mycol., 649).
DENNIS, R.G.W. 8tFOISTER, C.E. (1942). List of diseases of economic plants recorded in Scotland, Trans. Brit. mycol. Soc., 25, 275-277.
DIEHL, W.W. (1950). Balansia and the Wlansiae in America. Agric. Monograph No. 4, pp.82, 11 pls. Washington D.0.1 U.S. Lep. Agric.
DUGGAR, B.M. & DAVIS, A.B. (1916). Studies in the physiology of the fungi. I. Nitrogen fixation. Ann. Mo. bot. Gan., 3, 413-437.
ERIKSSON* J. (1912). Fungoid diseases of Agricultural plants. pp.208. Bailliere, Tindn11. add Cox, 8, Hanrietta Street, Covent Garden, London.
FISCHER, G.W. & HOLTON, C.S. (1957). Biology and control of the smut fungi. Ronn10 Press Co.,. New York.
FOSTER* J.W. (1949). Chemical activities of fungi. Academic Press Inc., NeNTYork.

193 -
FOKIN A.D. (1924). Diseases and injuries of cultivated plants observed during the sumer of 1922 in the government of Vyatka. Fourth All Russian Entomo.Phytopath. Congress in Moscow 8.-14 Dec. 1922, 108. (Abstr. in Rev, appl. Mycol. 596,-7).
FREEMAN, E.M. (1903). The seed fungus of LoliumAremulentum. Phils. Trans., London, B. 2m, 1027.
CAUMANN, E. (1950). Principles of plant infection. pp. 543. Crossby Lockwood, London.
GOTTLIEB, D. (1950). The physiology of spore germination in fungi. Bot. Rev., 26 229-257.
GRAM, E. &THOMSEN, M. (1925). Oversigt over Sygdomme hos Landbrugets off; Havebrugets Kulturplanter i 1925. (Survey of the diseases of Agricultural and Horticultural cultivated plants in 1925). Tidsskr. for Planteavl.,
84448. (Abstr. in Rev. appl. Mycol., S!, 5554.)
nms, rm. (1933). An analysis of the factors controlling the growth forn of certain fungi, with especial reference to Sblerotinia (Monilinia) fruetigena. Ann. Bot., Land. 47, 543-578.
HANIG, E. (1907). Uberpilereies Loliun tenulentun. Bot. Ztg. 25-38.
R.H. a WESTON, W.H. Jr. (1950), Studies in the lower Chytridiales. I. Factors affecting pignentation, growth and metabolism of a strain of Karlingia (Rizophlyctis) rosea. Amer. J. Bot., 37: 759-750.
HIRST, J.M. (1952). An automatic volumetric spore trap, Ann. -Tpl. Biol., ZD 257-265.
HYDE, H.L. a WILLIAMS, D.A. (1945). Studies in atmospheric pollen. II. Diurnal variation in the incidence of grass pollen. New Phytol., ig, 85-94.
HASKINS,

-4 194 -
MOLD, Ca, (1948). The water relations of spore discharge in Epichloe. Trans. Brit, mycol. Soc,' 32, 277..4280.
JONES, W.N. & SMITH, M.L. (1928). On the fixation of atmospheric nitrogen by Fhoma radiciis callunae including a new method of investigating nitrogen fixation in micro- organisms. Brit. J. exp. Biol., 6, 167-189.
KHUDAIRI, L.K. & HAMNER, O.K. (1953). Effect of ethylene chlorohydrin on floral initiation in Xanthium. Bot, Gaz., 115, 289-291.
KIRBY, E.J.M. (1958). Host parasite relationship in the choke disease of grasses. Ph.D. Thesis, University of Nottingham.
KOBOYASI, Y. (1941). The genus Cordyceps and its allies. Sci. Rep. Tokyo. Bun. Dug, Sect. B5, 55-260.
LARGE, E.C. (1952). Surveys for choke (Epichloe typhina) in cocksfoot seed crops, 1951. Plant Path., 1, 25-28,
(1954). Surveys for choke (Epichloe typhina)in cocksfoot seed crops, 1951-53. Plant Path., 3, 6-11.
LTIONIAN, L.H. & LTLLY, (1942)-.- -The effects of vitamins on ten strains of Saccharomyces cerevisiae. Amer. J. Bot., 29, -45Q-464,
LEOPOLD, A.C. & THIMANN, K.V. (1949). The effect of auxin on flower initiation. Amer. J. Bot., 36, 542-547.
LEPIK, E. (1931). (Phytopathological notes 2-8). pith. Phytopath. Versuchsstat. Univ. Tartu. (Esthonia). 2, 14.
(Abstr. in Rev. appl. Mycol., 22, 24-25.)
LILLY, V.G. & BARNETT, B.L. (1949). Growth rates and vitamin deficiencies in various fungi. Proc. W. Va. Acad. Sci., 19, 8-1, 27-35.
(1951). Physiology of the fungi. pp. 464. McGrawHill Book Company Inc., New York,
London.

195 -
El. (1960). The role of the oospore of Feronospor& parasitica in downy mildew of crucifers. Phytopath., 95..97.
H.K. & HOULAHAN, M.B. (1946). Neurosporm. IV, 11 temperature-isensitive riboflavinloss mutant. Amer. J. Bet. E5 31-35.
(1924). (The toxicity of the /strangulation' fungus frequently parasitizing grasses). Deutsche Landw. Presse, 2.5 254. (astr. in Rev. apple Mycol., 1:5 5840
M.B. (1956). A uethod of inoculating wheat and barley with loose smut. Phytopath. gsz, 597-400.
J.C. (1942). The endophytes of Loliun and Festuca, N.Z. J. Sci. Tech., 25A, 185-193. (Abstr. in Rev. appl. Mycol., 22, 158-159.)
RERSOON„ C.H. (1798). Sphaarta typhina. Icons et descriptiones fungorum minus cognitorun, Lipsiae, p.21.
FETCH. T. (1957). British Hypocreales. Trans. Brit. mycol. Soc„ 22, 245-305.
RAYNER. M.C. (1922). Nitrogen fixation in EriCaceae. Bot. Gaz. 751 226-255.
ROBBINS, W.J. ez KAVANAGH, V. (1942). Vitamin deficiencies of filamentous fungi. Bot. Rev., 8,. 411-471.
SLOSON. K. (1955). The systemic infection of grasses by Epichloe typhina. Trans. Brit. mycol. Soc., 18, 50-47.
(1954). The presence and absence of an endophytiC fungus in Loliun tenulentun and 1....,..percane. Trans. Brit. m 7-3ycol. Soc. 19, 5345.
(1957). Further observations on the systemic infection of Loliun. Trans. Brit. nycol. Soc., 22,
McMEEKIN,
MITCHELL,
MOLZ, B.
MOORE,
NEILL,

-196 -
SLMPSON, K, &WESTERN, J.H. (1954). Diseases of British grasses and herbage legumes, pp. 132, Cambridge University Press,
SIMONDS, P.M, (1946). Detection of the loose smut fungi in embryos of barley and wheat. Sci. Agric., 26, 51-58.
SNYDER W.C. & HANSTN, H.N. (1947). Gaseous sterilization of biological materials for use as culture media. Fhytopath., 57, 569...371.
SOWERBY$ J, (1905). English fungi. London.
STEINBERG, R.A. (1946). Specificity of potassium and magnesium for the growth of Aspergillus niger, Amer. J. Bot.$ 35$ 210.-214.
TERNETZ, C. (1907). Ueber die assimilation des atmospharischen stickstoff durch pilze. Jahrb. NiSS. Botan., 44, 555.-
408:
TOVE„, S.R., NI55, B.F. & WILSON, P.W. (1949). Nitrogen fixation by the fungus, Phona easuarina, 49th meeting, Soc. Ala. Both. (Lbs. No. A-26, p.59).
TULLtSNE, L.R. & C. (1865). Selector fungorun Carpologia. III pp.201. Imperial Press, Paris.
VLADIMIRSKAYA, N.N. (1928). (Contribution to the biology of Epichloe typhina Tul.) La Defense des Plantes, Leningrad, 5, 5-4 535-347. (Abstr. in Rev, appl. Mycol.$ 8, 315-4.)
WAHLIN, B. (1949). Some important outbreaks of diseases and pests on agricultural crops in East Gothland, 1949). Vaxtskyddsnotiser, Vaxtskyddsanst., Stockh., 6, 11-15. (Abstr. in Rev. appl. Mycol., 246 249.)
WALKER, J.C. & LINK, K.P. (1955). Toxicity of phenolic compounds to certain onion bulb parasites. Bat. Gaz., 96, 468 '1.81.

- 197 -
VJERN1ILi,I, C.C. (1242). Epichloe typhina on imported fescue seed. Phytopath., 32, 1093.
VEST ERN„ J.H. & CAVETT, J.J. (1959). The choke disease of cocksfoot (ractylis gloperata) caused by Epichloe typhina (Pr.) Tul. Trans. Brit. nycol. Soc.) 42, 5, 298-507.
WEXLER, II. & MEG/15A, S. (1954). Relative humidity, temperature relationships of sore saturated salt solutions in the temperature range 0 to 5000. J. Re's. nat. Bur. 'Stand., 53,' 12-26.
VOGL, A. (1898). "Mehl and anderen• Mehlprodukte der Cerealien ux d Legumiiosen."Z.1,fahrungsmittel-Untersuchung, Hygiene u. Warenkunde, 12,- 25-9.
Copyright © 2022 FDOKUMEN




















