
Differential time courses and specificity of amygdala activity in posttraumatic stress disorder...
10
Differential Time Courses and Specificity of Amygdala Activity in Posttraumatic Stress Disorder Subjects and Normal Control Subjects Xenia Protopopescu, Hong Pan, Oliver Tuescher, Marylene Cloitre, Martin Goldstein, Wolfgang Engelien, Jane Epstein, Yihong Yang, Jack Gorman, Joseph LeDoux, David Silbersweig, and Emily Stern Background: Previous neuroimaging studies have demonstrated exaggerated amygdala responses to negative stimuli in posttrau- matic stress disorder (PTSD). The time course of this amygdala response is largely unstudied and is relevant to questions of habituation and sensitization in PTSD exposure therapy. Methods: We applied blood oxygen level dependent functional magnetic resonance imaging and statistical parametric mapping to study amygdala responses to trauma-related and nontrauma-related emotional words in sexual/physical abuse PTSD and normal control subjects. We examined the time course of this response by separate analysis of early and late epochs. Results: PTSD versus normal control subjects have a relatively increased initial amygdala response to trauma-related negative, but not nontrauma-related negative, versus neutral stimuli. Patients also fail to show the normal patterns of sensitization and habituation to different categories of negative stimuli. These findings correlate with measured PTSD symptom severity. Conclusions: Our results demonstrate differential time courses and specificity of amygdala response to emotional and control stimuli in PTSD and normal control subjects. This has implications for pathophysiologic models of PTSD and treatment response. The results also extend previous neuroimaging studies demonstrating relatively increased amygdala response in PTSD and expand these results to a largely female patient population probed with emotionally valenced words. Key Words: PTSD, amygdala, fMRI, time course, neuroimaging, emotion P osttraumatic stress disorder (PTSD) occurs when an indi- vidual experiences a traumatic event; reacts with intense fear, helplessness, or horror; and develops symptoms including reexperiencing (nightmares, flashbacks), avoidance, numbing, and hyperarousal that persist for at least a month (American Psychiatric Association 2000). Posttraumatic stress disorder is a common disorder with a lifetime prevalence of about 8% in the general population (American Psychiatric Asso- ciation 2000). In addition to assault, accident, and combat-related trauma, terrorism-related trauma has recently focused additional attention on this disorder (LeDoux and Gorman 2001). The amygdala is of interest to the study of PTSD because of its involvement in emotional memory, emotional/behavioral mani- festations of fear, and autonomic response (LeDoux 1996, 2000). Neurocircuitry models of PTSD have hypothesized an overactive amygdala with inadequate top-down governance over the amyg- dala by medial prefrontal cortex (mPFC; specifically, the rostral anterior cingulate cortex) and the hippocampus (Golier and Yehuda 2002; Rauch et al 2003b; Shin et al 2004; Vermetten and Bremner 2002). Posttraumatic stress disorder patients, compared with normal control subjects, have shown increased cardiac, skin conductance, and electromyogram responses to loud tones (Orr et al 1995; Shalev et al 1992). A disturbance in memory function is one of the core symptoms in PTSD (Golier and Yehuda 2002; McNally 1997). Functional neuroimaging studies in normal subjects have found selective activation of the amygdala in processing negative emotional stimuli, including words (Breiter et al 1996; Isenberg et al 1999; Morris et al 1996; Phelps et al 2001), and in fear conditioning (Buchel et al 1998; LaBar et al 1998). Numerous studies have found increased amygdala activity to traumatic stimuli in PTSD patients (Liberzon et al 1999; Rauch et al 1996, 2000; Shin et al 1997). Based on animal studies showing that mPFC lesions disrupt emotional regulation (Morgan et al 1993; Quirk et al 2000), this may represent an intrinsic lower threshold of amygdala response to fearful stimuli and/or a dysfunction of the mPFC, including the anterior cingulate, with a failure to inhibit amygdala activation (Pitman et al 2001; Villarreal and King 2001). A study demonstrating exaggerated amygdala response to masked fearful faces in PTSD supports an intrinsic lower thresh- old of amygdala response dissociated from medial frontal acti- vation (Rauch et al 2000). However, a number of studies that have successfully demonstrated alterations in hippocampal and prefrontal activation in PTSD patients have not found amygdala hyperresponsivity (Bremner et al 1999a, 1999b, 2003b). Further characterization of the amygdala response in PTSD subjects and normal control subjects is essential to gain an improved understanding of the neurobiology of PTSD. Such a characterization can also help test models of PTSD pathophysi- ology, understand mechanisms of treatment, and explain inter- study differences. Amygdala habituation to emotional stimuli is well documented in normal control subjects (Fischer et al 2003; Phelps et al 2001; Wright et al 2001). Posttraumatic stress disorder patients may differ from normal control subjects in temporal dynamics as well as magnitude of amygdala response. Current questions include the time course of the amygdala response, the specificity of the amygdala response to given classes of stimuli, the extent to which the amygdala response is generalizable to different PTSD patient populations, and the degree to which the From the Functional Neuroimaging Laboratory (XP, HP, OT, MG, WE, JE, YY, DS, ES), Weill Medical College of Cornell University; The Rockefeller Uni- versity Laboratory of Neuroendocrinology (XP); New York University School of Medicine (MC); Mount Sinai School of Medicine (JG); and New York University (JL), New York, New York. Address reprint requests to Dr. Emily Stern, Functional Neuroimaging Lab- oratory, Department of Psychiatry, Box 140, Room 1302, Weill Medical College of Cornell University, 1300 York Avenue, New York, NY 10021; E-mail: [email protected]. Received June 28, 2004; revised December 1, 2004; accepted December 13, 2004. BIOL PSYCHIATRY 2005;57:464 – 473 0006-3223/05/$30.00 doi:10.1016/j.biopsych.2004.12.026 © 2005 Society of Biological Psychiatry
-
Upload
transnationalsupport -
Category
Documents
-
view
1 -
download
0
Transcript of Differential time courses and specificity of amygdala activity in posttraumatic stress disorder...

DANXWa
BmaMscRntCiat
Ke
Pin(dacta
ifNadaYBwc
F
A
R
0d
ifferential Time Courses and Specificity of Amygdalactivity in Posttraumatic Stress Disorder Subjects andormal Control Subjects
enia Protopopescu, Hong Pan, Oliver Tuescher, Marylene Cloitre, Martin Goldstein,olfgang Engelien, Jane Epstein, Yihong Yang, Jack Gorman, Joseph LeDoux, David Silbersweig,
nd Emily Stern
ackground: Previous neuroimaging studies have demonstrated exaggerated amygdala responses to negative stimuli in posttrau-atic stress disorder (PTSD). The time course of this amygdala response is largely unstudied and is relevant to questions of habituationnd sensitization in PTSD exposure therapy.ethods: We applied blood oxygen level dependent functional magnetic resonance imaging and statistical parametric mapping to
tudy amygdala responses to trauma-related and nontrauma-related emotional words in sexual/physical abuse PTSD and normalontrol subjects. We examined the time course of this response by separate analysis of early and late epochs.esults: PTSD versus normal control subjects have a relatively increased initial amygdala response to trauma-related negative, butot nontrauma-related negative, versus neutral stimuli. Patients also fail to show the normal patterns of sensitization and habituation
o different categories of negative stimuli. These findings correlate with measured PTSD symptom severity.onclusions: Our results demonstrate differential time courses and specificity of amygdala response to emotional and control stimuli
n PTSD and normal control subjects. This has implications for pathophysiologic models of PTSD and treatment response. The resultslso extend previous neuroimaging studies demonstrating relatively increased amygdala response in PTSD and expand these results
o a largely female patient population probed with emotionally valenced words.
ey Words: PTSD, amygdala, fMRI, time course, neuroimaging,motion
osttraumatic stress disorder (PTSD) occurs when an indi-vidual experiences a traumatic event; reacts with intensefear, helplessness, or horror; and develops symptoms
ncluding reexperiencing (nightmares, flashbacks), avoidance,umbing, and hyperarousal that persist for at least a monthAmerican Psychiatric Association 2000). Posttraumatic stressisorder is a common disorder with a lifetime prevalence ofbout 8% in the general population (American Psychiatric Asso-iation 2000). In addition to assault, accident, and combat-relatedrauma, terrorism-related trauma has recently focused additionalttention on this disorder (LeDoux and Gorman 2001).
The amygdala is of interest to the study of PTSD because of itsnvolvement in emotional memory, emotional/behavioral mani-estations of fear, and autonomic response (LeDoux 1996, 2000).eurocircuitry models of PTSD have hypothesized an overactivemygdala with inadequate top-down governance over the amyg-ala by medial prefrontal cortex (mPFC; specifically, the rostralnterior cingulate cortex) and the hippocampus (Golier andehuda 2002; Rauch et al 2003b; Shin et al 2004; Vermetten andremner 2002). Posttraumatic stress disorder patients, comparedith normal control subjects, have shown increased cardiac, skin
onductance, and electromyogram responses to loud tones (Orr
rom the Functional Neuroimaging Laboratory (XP, HP, OT, MG, WE, JE, YY,DS, ES), Weill Medical College of Cornell University; The Rockefeller Uni-versity Laboratory of Neuroendocrinology (XP); New York UniversitySchool of Medicine (MC); Mount Sinai School of Medicine (JG); and NewYork University (JL), New York, New York.
ddress reprint requests to Dr. Emily Stern, Functional Neuroimaging Lab-oratory, Department of Psychiatry, Box 140, Room 1302, Weill MedicalCollege of Cornell University, 1300 York Avenue, New York, NY 10021;E-mail: [email protected].
eceived June 28, 2004; revised December 1, 2004; accepted December 13,
2004.006-3223/05/$30.00oi:10.1016/j.biopsych.2004.12.026
et al 1995; Shalev et al 1992). A disturbance in memory functionis one of the core symptoms in PTSD (Golier and Yehuda 2002;McNally 1997).
Functional neuroimaging studies in normal subjects havefound selective activation of the amygdala in processing negativeemotional stimuli, including words (Breiter et al 1996; Isenberg etal 1999; Morris et al 1996; Phelps et al 2001), and in fearconditioning (Buchel et al 1998; LaBar et al 1998). Numerousstudies have found increased amygdala activity to traumaticstimuli in PTSD patients (Liberzon et al 1999; Rauch et al 1996,2000; Shin et al 1997). Based on animal studies showing thatmPFC lesions disrupt emotional regulation (Morgan et al 1993;Quirk et al 2000), this may represent an intrinsic lower thresholdof amygdala response to fearful stimuli and/or a dysfunction ofthe mPFC, including the anterior cingulate, with a failure toinhibit amygdala activation (Pitman et al 2001; Villarreal and King2001). A study demonstrating exaggerated amygdala response tomasked fearful faces in PTSD supports an intrinsic lower thresh-old of amygdala response dissociated from medial frontal acti-vation (Rauch et al 2000). However, a number of studies thathave successfully demonstrated alterations in hippocampal andprefrontal activation in PTSD patients have not found amygdalahyperresponsivity (Bremner et al 1999a, 1999b, 2003b).
Further characterization of the amygdala response in PTSDsubjects and normal control subjects is essential to gain animproved understanding of the neurobiology of PTSD. Such acharacterization can also help test models of PTSD pathophysi-ology, understand mechanisms of treatment, and explain inter-study differences. Amygdala habituation to emotional stimuli iswell documented in normal control subjects (Fischer et al 2003;Phelps et al 2001; Wright et al 2001). Posttraumatic stress disorderpatients may differ from normal control subjects in temporaldynamics as well as magnitude of amygdala response. Currentquestions include the time course of the amygdala response, thespecificity of the amygdala response to given classes of stimuli,the extent to which the amygdala response is generalizable to
different PTSD patient populations, and the degree to which theBIOL PSYCHIATRY 2005;57:464–473© 2005 Society of Biological Psychiatry

as
tl(t(tcipawesns
M
S
2sfahcfs7pbH
1dsTwsstasds
md(D1die(((eV1C
X. Protopopescu et al BIOL PSYCHIATRY 2005;57:464–473 465
mygdala response correlates with overall PTSD severity andpecific PTSD symptoms.
We previously demonstrated increased amygdala response tohreatening words and have been utilizing the specificity ofinguistic stimuli to study limbic function in psychiatric disordersIsenberg et al 1999). Here, we developed a linguistic emotionalask in conjunction with blood oxygenation level dependentBOLD) functional magnetic resonance imaging (fMRI) to inves-igate the neurocircuitry underlying PTSD. We examined the timeourse of amygdala response by dividing our 24-minute studynto early and late epochs. Using trauma-relevant negative,anic-relevant negative (negative control condition), positive,nd neutral words, we tested the hypothesis that PTSD subjectsould show differential amygdala activation to these differentmotional word categories over time and specifically demon-trate initial increased amygdala activity to trauma-related versuseutral words, possibly in the context of a “primed” trauma-pecific limbic response.
ethods and Materials
ubjectsParticipants consisted of 11 sexual/physical assault PTSD and
1 normal control subjects. Data sets were excluded if there wasignificant loss of echo-planar imaging (EPI) signal in the orbito-rontal cortex, ghosting artifact, or head motion. Subjects werelso excluded for failing to respond to the task and for medicalistory misinformation. Subjects included in the final analysisonsisted of 9 PTSD subjects (mean age � 35, range � 20–55; 7emale subjects and 2 male subjects) and 14 normal controlubjects (mean age � 27, range � 22–42; 7 female subjects andmale subjects). All subjects gave informed consent prior to
articipation in the study, which was part of a protocol approvedy the Institutional Review Board at New York-Presbyterianospital/Weill Medical College of Cornell University.All subjects were right-handed, native English speakers, aged
8 to 55. Posttraumatic stress disorder subjects all had a primaryiagnosis of PTSD by DSM-IV criteria, eight of nine subjects hadexual assault PTSD, and one subject had physical assault PTSD.wo subjects had secondary diagnoses of social phobia, one ofhich also had secondary diagnoses of specific phobia, obses-
ive-compulsive disorder, and dysthymia. One subject had aecondary diagnosis of major depressive disorder. Only one ofhe nine PTSD subjects had ever taken psychiatric medication,nd that subject was medication-free for over 1 year prior tocanning. No subjects had any history of substance abuse orependence, and urine toxicology was performed prior tocanning.
Normal and PTSD subject characterization used the followingeasures: Structured Clinical Interview for DSM-IV Axis I Disor-ers (SCID-I) (First et al 1997), Beck Depression Inventory (BDI)Beck et al 1981), State-Trait Anxiety Inventory (STAI/TRAI), andissociative Experience Scale (DES) (Bernstein and Putnam986). Posttraumatic stress disorder subject characterization ad-itionally used a variety of measures including: Structured Clin-cal Interview for DSM-IV Personality Disorders (SCID-II) (Spitzert al 1994), State Trait Anger Expression Inventory (STAXI)Spielberger et al 1983), PTSD Symptom Scale-Self ReportPSS-SR) (Foa 1995), COPE, and Anxiety Sensitivity Index (ASI)Reiss et al 1986). For PTSD subjects, a trauma history was firststablished using the Sexual Assault and Adult Interpersonaliolence (SAAIV), Childhood Interpersonal Violence Before Age8 (CIVI), and Adult Non-Interpersonal Violence scales. The
linician Administered PTSD Scale (CAPS) (Blake et al 1995) wasused to establish a diagnosis of PTSD. To meet criteria fordiagnosis of PTSD, the subject must have had the followingsymptoms: for symptom B cluster (reexperiencing symptoms), atleast one symptom with a frequency rating of 1 and intensityrating of 2; for symptom C cluster (avoidance and numbingsymptoms), at least three symptoms; and for symptom D cluster(startle response/hyperarousal), at least two symptoms. TheCAPS scores of our PTSD subjects ranged from 37 to 91 with amean of 60.
StimuliStimuli consisted of 48 negative/anxiety (24 negative/PTSD,
24 negative/panic), 48 neutral, and 48 positive/safety words,balanced across categories for frequency, length, and part ofspeech (nouns and adjectives/verbs). Posttraumatic stress disor-der words were designed to be relevant to physical/sexualtrauma; panic words were designed to be relevant to panic attacksymptoms and somatic/illness-related anxiety (a negative controlcondition, as well as explicit probes for panic disorder patientsalso studied as part of a larger project); and positive words weredesigned to be counter-anxiety and evocative of safety, relax-ation, and reward, as defined by the literature and clinicalexperience. These word types were rated for suitability by apanel of three experienced clinicians. They were based on asimilar list of words that had been piloted on 34 normal subjects,who rated the three word types (positive, negative, and neutral)as significantly different in valence (p � .001) and rated positiveand negative words as not significantly different in intensity (p �.2). Examples are as follows: negative/PTSD–rape, assault, force;negative/panic–frantic, death, cancer; neutral–bookcase, clari-net, rotate; positive/rewarding–safe, gentle, delighted.
The three valences of words were presented within a blockdesign (six words per block, eight blocks per valence), withblocks balanced to control for order and time effects. Posttrau-matic stress disorder words and panic words were presented forfour blocks each (each representing half of the total negativeword blocks). Each word appeared for 2 seconds, followed by aninterstimulus interval jittered around an average of 2.8 seconds,for a total of 28.8 seconds per block. Blocks were presented infour study epochs containing six blocks each (Figure 1). Eachblock was followed by 24 seconds of rest, with each epoch as awhole preceded and followed by 2 additional 12-second restperiods. The entire word paradigm took approximately 24minutes. Stimulus presentation and response collection wereperformed within the Integrated Functional Imaging System–Stand Alone (IFIS-SA; www.MRIDevices.com/Funct/IFIS.asp)/E-Prime environment. During presentation of stimuli, subjects wereinstructed to read each word silently and to then immediatelypress a button under their right index finger. During rest periods,they were instructed to look at a dash at the center of the screenand to have their minds either blank or floating freely.
Subjects also completed an instructed fear-conditioning par-adigm in the scanner (discussed elsewhere). Approximately halfof the subjects completed this before and half completed thisafter the word paradigm. After the emotional word paradigm,subjects were removed from the scanner and given an incidentalmemory test utilizing a list of words consisting of the 144 stimuliseen during scanning (targets) randomly interspersed with 72other words (distracters to control for false alarms); dividedequally into negative (PTSD and panic), neutral, and positivecategories; and balanced for the same qualities as the targets.They were instructed to read each word and to indicate those
that they believed they had seen in the scanner. Followingwww.elsevier.com/locate/biopsych

cv
I
SmpCslmu1�mr2sr
I
Dwavaesds(
aicabaginsnrte
F e.
466 BIOL PSYCHIATRY 2005;57:464–473 X. Protopopescu et al
w
ompletion of that task, subjects were also asked to rate thealence and intensity of each word on scales of �3 to �3.
mage AcquisitionAll image data were acquired with a research-dedicated GE
igna 3 Tesla MRI scanner (maximum gradient strength 40mT/m,aximum gradient slew rate 150T/m/s) (General Electric Com-any, Waukesha, Wisconsin) at the Weill Medical College ofornell University. Blood oxygen level dependent fMRI mea-ures hemodynamic and oxygenation changes associated withocalized neuronal activity in the brain. After shimming toaximize homogeneity, a series of 3T fMRI scans was collectedsing gradient echo echo-planar imaging (repetition time [TR] �200; echo time [TE] � 30; flip angle � 70o; field of view [FOV]240 mm; 15 slices; 5 mm thickness with 1 mm interslice space;atrix � 64 x 64), with a modified z-shimming algorithm to
educe susceptibility artifact at the base of the brain (Gu et al002). T1-weighted anatomical images were acquired using apoiled gradient recalled acquisition (SPGR) sequence with aesolution of .9375 x .9375 x 1 mm3.
mage Processing and Data AnalysisStatistical Parametric Mapping (SPM99) software (Wellcome
epartment of Imaging Neuroscience, London, United Kingdom)as used for processing and preprocessing of the data (Friston etl 1995a, 1995b). To build the statistical model, a whole-brain,oxel-by-voxel multiple linear regression model was employedt the single subject level. The resulting set of voxel values forach contrast constituted a statistical parametric map of the ttatistic (SPM{t}), which was then transformed to the unit normalistribution, SPM{Z} (Friston et al 1995b). All coordinates pre-ented here are in Montreal Neurological Institute (MNI) spaceMNI average 152 T1 brain).
For group analyses, we used a random effects model, whichccounts for intersubject variability and allows population-basednferences to be drawn (Friston et al 1999). For each subject, theontrast image for each condition (i.e., the condition effectsgainst the resting state) was generated, and these were com-ined in a series of linear contrasts to assess group effects. Welso ran the analysis using the nine PTSD patients and nineender-matched normal control subjects. The conditions ofnterest were the four word types: positive, neutral, PTSD/egative, and panic/negative. In addition, the analysis was runeparately using age, gender, or paradigm order as covariates ofo interest in an analysis of variance (ANOVA) setting. Theesults were analyzed from the first two study epochs (early) andhe last two study epochs (late) separately to examine time
igure 1. Schematic figure of the neuropsychological paradigm architectur
ffects. Group and condition comparisons of interest generated
ww.elsevier.com/locate/biopsych
statistical parametric maps (SPMs) of the t statistic (SPM{t}), whichwas again transformed to a unit normal distribution (SPM{Z}).
Because we had a region-specific hypothesis for the amyg-dala, a region-of-interest analysis was performed. For this pur-pose, to ensure complete coverage of all amygdala subnuclei, acombined mask of the amygdala (as defined by AutomatedAnatomical Labeling [AAL] areas 41 and 42 [Tzourio-Mazoyer et al2002]) and adjacent periamygdaloid cortex (Brodmann area 28)was created in MNI space. Reflecting hypotheses, an initial t mapthreshold of p � .01 uncorrected was used (p values presentedare uncorrected unless specified). As the AAL-Brodmann area 28combined mask extends somewhat anteriorly and posteriorly tothe amygdala proper, only those points falling within the maskand meeting the additional criteria of falling between �6 and �3in the y-direction were considered as amygdala cluster peaks.Within the amygdala, a priori regions of interest were specifiedusing the small volume correction (SVC) function to a surround-ing sphere of radius 4 mm (approximately .25 cc) in SPM andwere considered significant if the corresponding voxelwise pvalue was less than .05 corrected (Friston 1997; Worsley et al1996). Significant differences in group, condition, and interactioneffects were assessed in these regions within the context of aGeneral Linear Model and Gaussian Random Field Theory. Acorrelation analysis was also performed to determine the associ-ation between activity in the amygdala and a clinical measure ofPTSD severity (CAPS total). A correlation analysis was also per-formed to determine the association between activity in the amyg-dala and the clinical measure thought to be of greatest relevance tothe experimental paradigm, subjective rating of psychological dis-tress experienced at trauma reminders (CAPS B4).
Results
Between-Group Effects of ConditionAs hypothesized and consistent with previous studies in PTSD
subjects, PTSD versus normal subjects show increased left amyg-dala BOLD response to trauma-relevant negative (PTSD) wordsversus neutral words p � .006 (Table 1). Of note, here thisdifference was demonstrated in the early, but not late, half of thestudy. Posttraumatic stress disorder versus normal subjects dem-onstrate differential time courses of left amygdala BOLD re-sponse to PTSD words controlled for neutral words, p � .005(Table 1, Figures 2 and 3). The [(PTSD/early versus Neutral/early) versus (PTSD/late versus Neutral/late)] contrast, abbrevi-ated [(PTe vs. NUe) vs. (PTl vs. NUl)], represents the change inresponse to PTSD words, controlled for neutral words, from thefirst half of the study to the second half of the study.
A direct comparison of early versus late PTSD words in
normal subjects shows a sensitization pattern, with left amygdala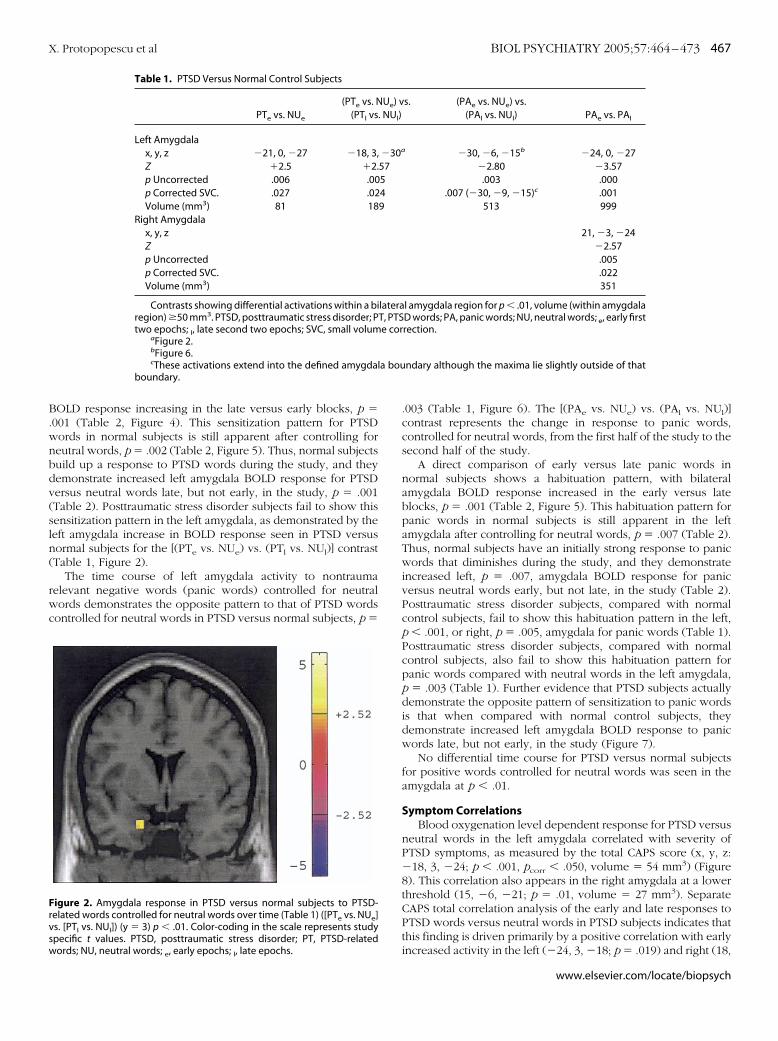
B.wnbdv(sln(
rwc
a bou
Frvsw
X. Protopopescu et al BIOL PSYCHIATRY 2005;57:464–473 467
OLD response increasing in the late versus early blocks, p �001 (Table 2, Figure 4). This sensitization pattern for PTSDords in normal subjects is still apparent after controlling foreutral words, p � .002 (Table 2, Figure 5). Thus, normal subjectsuild up a response to PTSD words during the study, and theyemonstrate increased left amygdala BOLD response for PTSDersus neutral words late, but not early, in the study, p � .001Table 2). Posttraumatic stress disorder subjects fail to show thisensitization pattern in the left amygdala, as demonstrated by theeft amygdala increase in BOLD response seen in PTSD versusormal subjects for the [(PTe vs. NUe) vs. (PTl vs. NUl)] contrastTable 1, Figure 2).
The time course of left amygdala activity to nontraumaelevant negative words (panic words) controlled for neutralords demonstrates the opposite pattern to that of PTSD words
ontrolled for neutral words in PTSD versus normal subjects, p �
Table 1. PTSD Versus Normal Control Subjects
PTe vs. NUe
(PTe vs. N(PTl vs
Left Amygdalax, y, z �21, 0, �27 �18, 3,Z �2.5 �2.p Uncorrected .006 .00p Corrected SVC. .027 .02Volume (mm3) 81 18
Right Amygdalax, y, zZp Uncorrectedp Corrected SVC.Volume (mm3)
Contrasts showing differential activations within a biregion) �50 mm3. PTSD, posttraumatic stress disorder; Ptwo epochs; l, late second two epochs; SVC, small volum
aFigure 2.bFigure 6.cThese activations extend into the defined amygdal
boundary.
igure 2. Amygdala response in PTSD versus normal subjects to PTSD-elated words controlled for neutral words over time (Table 1) ([PTe vs. NUe]s. [PTl vs. NUl]) (y � 3) p � .01. Color-coding in the scale represents studypecific t values. PTSD, posttraumatic stress disorder; PT, PTSD-related
ords; NU, neutral words; e, early epochs; l, late epochs..003 (Table 1, Figure 6). The [(PAe vs. NUe) vs. (PAl vs. NUl)]contrast represents the change in response to panic words,controlled for neutral words, from the first half of the study to thesecond half of the study.
A direct comparison of early versus late panic words innormal subjects shows a habituation pattern, with bilateralamygdala BOLD response increased in the early versus lateblocks, p � .001 (Table 2, Figure 5). This habituation pattern forpanic words in normal subjects is still apparent in the leftamygdala after controlling for neutral words, p � .007 (Table 2).Thus, normal subjects have an initially strong response to panicwords that diminishes during the study, and they demonstrateincreased left, p � .007, amygdala BOLD response for panicversus neutral words early, but not late, in the study (Table 2).Posttraumatic stress disorder subjects, compared with normalcontrol subjects, fail to show this habituation pattern in the left,p � .001, or right, p � .005, amygdala for panic words (Table 1).Posttraumatic stress disorder subjects, compared with normalcontrol subjects, also fail to show this habituation pattern forpanic words compared with neutral words in the left amygdala,p � .003 (Table 1). Further evidence that PTSD subjects actuallydemonstrate the opposite pattern of sensitization to panic wordsis that when compared with normal control subjects, theydemonstrate increased left amygdala BOLD response to panicwords late, but not early, in the study (Figure 7).
No differential time course for PTSD versus normal subjectsfor positive words controlled for neutral words was seen in theamygdala at p � .01.
Symptom CorrelationsBlood oxygenation level dependent response for PTSD versus
neutral words in the left amygdala correlated with severity ofPTSD symptoms, as measured by the total CAPS score (x, y, z:�18, 3, �24; p � .001, pcorr � .050, volume � 54 mm3) (Figure8). This correlation also appears in the right amygdala at a lowerthreshold (15, �6, �21; p � .01, volume � 27 mm3). SeparateCAPS total correlation analysis of the early and late responses toPTSD words versus neutral words in PTSD subjects indicates thatthis finding is driven primarily by a positive correlation with early
s. (PAe vs. NUe) vs.(PAl vs. NUl) PAe vs. PAl
�30, �6, �15b �24, 0, �27�2.80 �3.57.003 .000
.007 (�30, �9, �15)c .001513 999
21, �3, �24�2.57.005.022351
l amygdala region for p � .01, volume (within amygdalaD words; PA, panic words; NU, neutral words; e, early firstrection.
ndary although the maxima lie slightly outside of that
Ue) v. NUl)
�30a
57549
lateraT, PTSe cor
increased activity in the left (�24, 3, �18; p � .019) and right (18,
www.elsevier.com/locate/biopsych
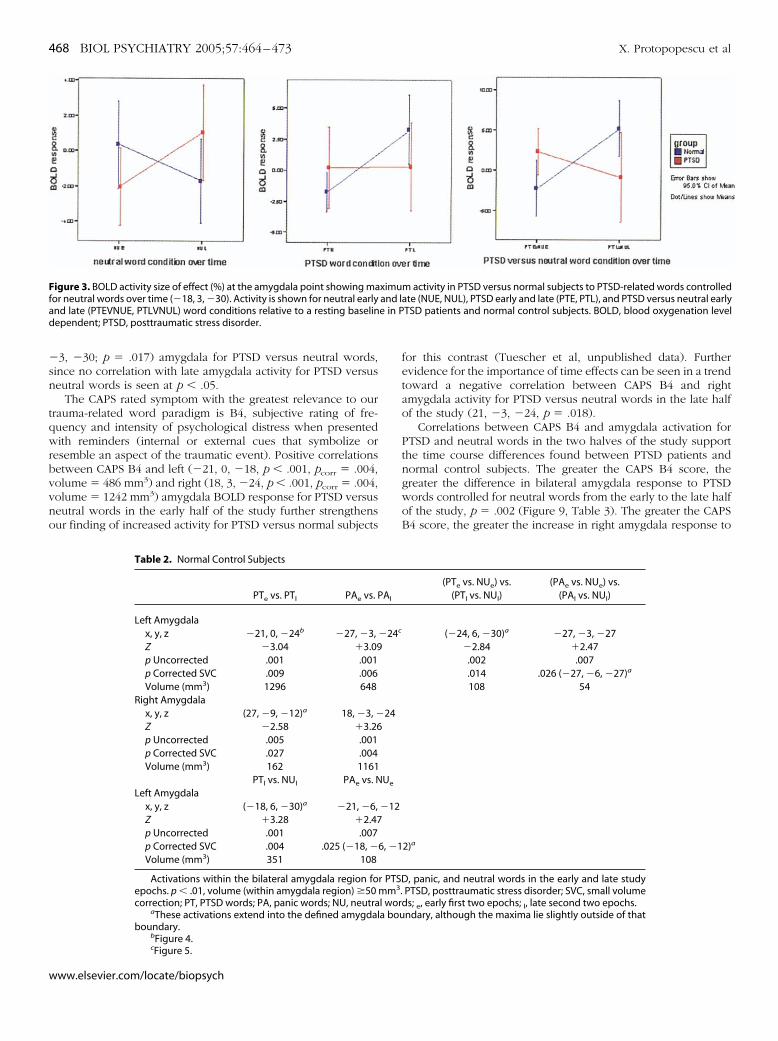
�sn
tqwrbvvno
Ffa e in Pd
468 BIOL PSYCHIATRY 2005;57:464–473 X. Protopopescu et al
w
3, �30; p � .017) amygdala for PTSD versus neutral words,ince no correlation with late amygdala activity for PTSD versuseutral words is seen at p � .05.
The CAPS rated symptom with the greatest relevance to ourrauma-related word paradigm is B4, subjective rating of fre-uency and intensity of psychological distress when presentedith reminders (internal or external cues that symbolize or
esemble an aspect of the traumatic event). Positive correlationsetween CAPS B4 and left (�21, 0, �18, p � .001, pcorr � .004,olume � 486 mm3) and right (18, 3, �24, p � .001, pcorr � .004,olume � 1242 mm3) amygdala BOLD response for PTSD versuseutral words in the early half of the study further strengthensur finding of increased activity for PTSD versus normal subjects
igure 3. BOLD activity size of effect (%) at the amygdala point showing maor neutral words over time (�18, 3, �30). Activity is shown for neutral earlynd late (PTEVNUE, PTLVNUL) word conditions relative to a resting baselinependent; PTSD, posttraumatic stress disorder.
Table 2. Normal Control Subjects
PTe vs. PTl PAe vs. P
Left Amygdalax, y, z �21, 0, �24b �27, �3,Z �3.04 �3.09p Uncorrected .001 .001p Corrected SVC .009 .006Volume (mm3) 1296 648
Right Amygdalax, y, z (27, �9, �12)a 18, �3, �Z �2.58 �3.26p Uncorrected .005 .001p Corrected SVC .027 .004Volume (mm3) 162 1161
PTl vs. NUl PAe vs. NLeft Amygdala
x, y, z (�18, 6, �30)a �21, �6,Z �3.28 �2.47p Uncorrected .001 .007p Corrected SVC .004 .025 (�18, �Volume (mm3) 351 108
Activations within the bilateral amygdala region foepochs. p � .01, volume (within amygdala region) �50correction; PT, PTSD words; PA, panic words; NU, neutra
aThese activations extend into the defined amygdalboundary.
bFigure 4.
cFigure 5.ww.elsevier.com/locate/biopsych
for this contrast (Tuescher et al, unpublished data). Furtherevidence for the importance of time effects can be seen in a trendtoward a negative correlation between CAPS B4 and rightamygdala activity for PTSD versus neutral words in the late halfof the study (21, �3, �24, p � .018).
Correlations between CAPS B4 and amygdala activation forPTSD and neutral words in the two halves of the study supportthe time course differences found between PTSD patients andnormal control subjects. The greater the CAPS B4 score, thegreater the difference in bilateral amygdala response to PTSDwords controlled for neutral words from the early to the late halfof the study, p � .002 (Figure 9, Table 3). The greater the CAPSB4 score, the greater the increase in right amygdala response to
m activity in PTSD versus normal subjects to PTSD-related words controlledate (NUE, NUL), PTSD early and late (PTE, PTL), and PTSD versus neutral earlyTSD patients and normal control subjects. BOLD, blood oxygenation level
(PTe vs. NUe) vs.(PTl vs. NUl)
(PAe vs. NUe) vs.(PAl vs. NUl)
(�24, 6, �30)a �27, �3, �27�2.84 �2.47.002 .007.014 .026 (�27, �6, �27)a
108 54
2)a
, panic, and neutral words in the early and late studyPTSD, posttraumatic stress disorder; SVC, small volume
ds; e, early first two epochs; l, late second two epochs.ndary, although the maxima lie slightly outside of that
ximuand l
Al
�24c
24
Ue
�12
6, �1
r PTSDmm3.l wora bou
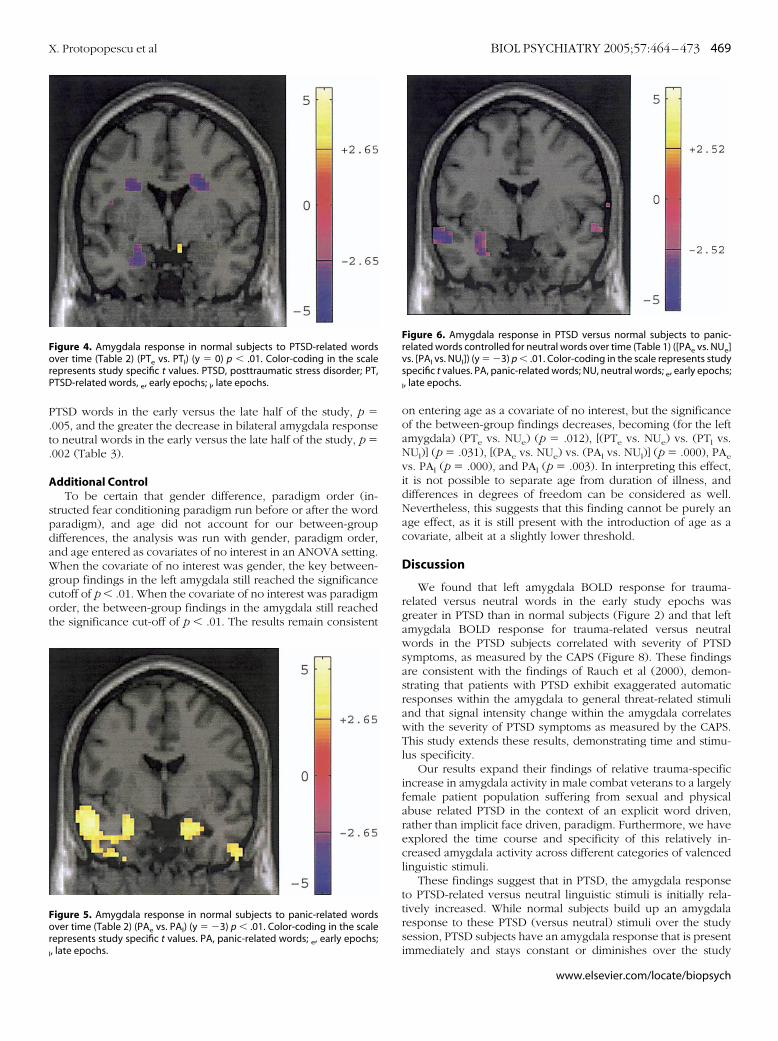
P.t.
A
spdaWgcot
Forl
ForP
X. Protopopescu et al BIOL PSYCHIATRY 2005;57:464–473 469
TSD words in the early versus the late half of the study, p �005, and the greater the decrease in bilateral amygdala responseo neutral words in the early versus the late half of the study, p �002 (Table 3).
dditional ControlTo be certain that gender difference, paradigm order (in-
tructed fear conditioning paradigm run before or after the wordaradigm), and age did not account for our between-groupifferences, the analysis was run with gender, paradigm order,nd age entered as covariates of no interest in an ANOVA setting.hen the covariate of no interest was gender, the key between-
roup findings in the left amygdala still reached the significanceutoff of p � .01. When the covariate of no interest was paradigmrder, the between-group findings in the amygdala still reachedhe significance cut-off of p � .01. The results remain consistent
igure 5. Amygdala response in normal subjects to panic-related wordsver time (Table 2) (PAe vs. PAl) (y � �3) p � .01. Color-coding in the scale
epresents study specific t values. PA, panic-related words; , early epochs;
igure 4. Amygdala response in normal subjects to PTSD-related wordsver time (Table 2) (PTe vs. PTl) (y � 0) p � .01. Color-coding in the scale
epresents study specific t values. PTSD, posttraumatic stress disorder; PT,TSD-related words, e, early epochs; l, late epochs.
e
, late epochs.
on entering age as a covariate of no interest, but the significanceof the between-group findings decreases, becoming (for the leftamygdala) (PTe vs. NUe) (p � .012), [(PTe vs. NUe) vs. (PTl vs.NUl)] (p � .031), [(PAe vs. NUe) vs. (PAl vs. NUl)] (p � .000), PAe
vs. PAl (p � .000), and PAl (p � .003). In interpreting this effect,it is not possible to separate age from duration of illness, anddifferences in degrees of freedom can be considered as well.Nevertheless, this suggests that this finding cannot be purely anage effect, as it is still present with the introduction of age as acovariate, albeit at a slightly lower threshold.
Discussion
We found that left amygdala BOLD response for trauma-related versus neutral words in the early study epochs wasgreater in PTSD than in normal subjects (Figure 2) and that leftamygdala BOLD response for trauma-related versus neutralwords in the PTSD subjects correlated with severity of PTSDsymptoms, as measured by the CAPS (Figure 8). These findingsare consistent with the findings of Rauch et al (2000), demon-strating that patients with PTSD exhibit exaggerated automaticresponses within the amygdala to general threat-related stimuliand that signal intensity change within the amygdala correlateswith the severity of PTSD symptoms as measured by the CAPS.This study extends these results, demonstrating time and stimu-lus specificity.
Our results expand their findings of relative trauma-specificincrease in amygdala activity in male combat veterans to a largelyfemale patient population suffering from sexual and physicalabuse related PTSD in the context of an explicit word driven,rather than implicit face driven, paradigm. Furthermore, we haveexplored the time course and specificity of this relatively in-creased amygdala activity across different categories of valencedlinguistic stimuli.
These findings suggest that in PTSD, the amygdala responseto PTSD-related versus neutral linguistic stimuli is initially rela-tively increased. While normal subjects build up an amygdalaresponse to these PTSD (versus neutral) stimuli over the studysession, PTSD subjects have an amygdala response that is present
Figure 6. Amygdala response in PTSD versus normal subjects to panic-related words controlled for neutral words over time (Table 1) ([PAe vs. NUe]vs. [PAl vs. NUl]) (y � �3) p � .01. Color-coding in the scale represents studyspecific t values. PA, panic-related words; NU, neutral words; e, early epochs;l, late epochs.
immediately and stays constant or diminishes over the study
www.elsevier.com/locate/biopsych
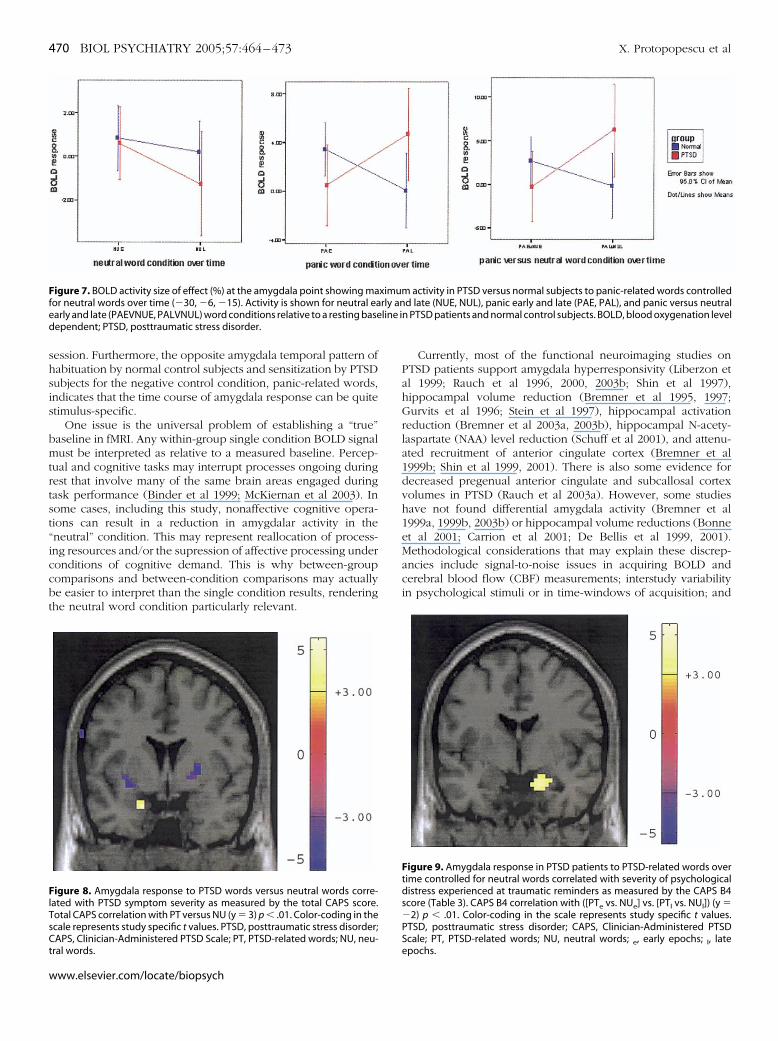
shsis
bmtrtst“iccbt
Ffe line id
FlTsCt
470 BIOL PSYCHIATRY 2005;57:464–473 X. Protopopescu et al
w
ession. Furthermore, the opposite amygdala temporal pattern ofabituation by normal control subjects and sensitization by PTSDubjects for the negative control condition, panic-related words,ndicates that the time course of amygdala response can be quitetimulus-specific.
One issue is the universal problem of establishing a “true”aseline in fMRI. Any within-group single condition BOLD signalust be interpreted as relative to a measured baseline. Percep-
ual and cognitive tasks may interrupt processes ongoing duringest that involve many of the same brain areas engaged duringask performance (Binder et al 1999; McKiernan et al 2003). Inome cases, including this study, nonaffective cognitive opera-ions can result in a reduction in amygdalar activity in theneutral” condition. This may represent reallocation of process-ng resources and/or the supression of affective processing underonditions of cognitive demand. This is why between-groupomparisons and between-condition comparisons may actuallye easier to interpret than the single condition results, renderinghe neutral word condition particularly relevant.
igure 7. BOLD activity size of effect (%) at the amygdala point showing maor neutral words over time (�30, �6, �15). Activity is shown for neutral eaarly and late (PAEVNUE, PALVNUL) word conditions relative to a resting baseependent; PTSD, posttraumatic stress disorder.
igure 8. Amygdala response to PTSD words versus neutral words corre-ated with PTSD symptom severity as measured by the total CAPS score.otal CAPS correlation with PT versus NU (y � 3) p � .01. Color-coding in thecale represents study specific t values. PTSD, posttraumatic stress disorder;APS, Clinician-Administered PTSD Scale; PT, PTSD-related words; NU, neu-
ral words.
ww.elsevier.com/locate/biopsych
Currently, most of the functional neuroimaging studies onPTSD patients support amygdala hyperresponsivity (Liberzon etal 1999; Rauch et al 1996, 2000, 2003b; Shin et al 1997),hippocampal volume reduction (Bremner et al 1995, 1997;Gurvits et al 1996; Stein et al 1997), hippocampal activationreduction (Bremner et al 2003a, 2003b), hippocampal N-acety-laspartate (NAA) level reduction (Schuff et al 2001), and attenu-ated recruitment of anterior cingulate cortex (Bremner et al1999b; Shin et al 1999, 2001). There is also some evidence fordecreased pregenual anterior cingulate and subcallosal cortexvolumes in PTSD (Rauch et al 2003a). However, some studieshave not found differential amygdala activity (Bremner et al1999a, 1999b, 2003b) or hippocampal volume reductions (Bonneet al 2001; Carrion et al 2001; De Bellis et al 1999, 2001).Methodological considerations that may explain these discrep-ancies include signal-to-noise issues in acquiring BOLD andcerebral blood flow (CBF) measurements; interstudy variabilityin psychological stimuli or in time-windows of acquisition; and
activity in PTSD versus normal subjects to panic-related words controlledd late (NUE, NUL), panic early and late (PAE, PAL), and panic versus neutral
n PTSD patients and normal control subjects. BOLD, blood oxygenation level
Figure 9. Amygdala response in PTSD patients to PTSD-related words overtime controlled for neutral words correlated with severity of psychologicaldistress experienced at traumatic reminders as measured by the CAPS B4score (Table 3). CAPS B4 correlation with ([PTe vs. NUe] vs. [PTl vs. NUl]) (y ��2) p � .01. Color-coding in the scale represents study specific t values.PTSD, posttraumatic stress disorder; CAPS, Clinician-Administered PTSDScale; PT, PTSD-related words; NU, neutral words; , early epochs; , late
ximumrly an
e l
epochs.

Ptatm
nrcaet
intiTi(1mfRww(L
m(lslc(wa
T
L
R
vgcpe
t
X. Protopopescu et al BIOL PSYCHIATRY 2005;57:464–473 471
TSD population heterogeneity with respect to instigatingrauma, psychiatric comorbidity, substance abuse, age, gender,nd other individual differences. In addition, as imaging resolu-ion improves, distinguishing activity in amygdalar subnucleiay resolve some interstudy differences.Our results, showing differential activity between PTSD and
ormal subjects in the early and late sessions considered sepa-ately but not for the average of all sessions, indicate that carefulonsideration of time course effects needs to be taken intoccount and may shed light on the lack of demonstrated differ-nces between psychiatric patient populations and normal con-rol subjects in some studies.
The amygdala is central to most models for a neural circuitryn PTSD because it permits assessment of the fear-producingature of an event, is critical to effectively lay down memoryraces related to a potential threat, and is able to influence anndividual’s neuroendocrine, autonomic, and motor responses.here is strong evidence that the amygdala and hippocampus are
nvolved in differentiable aspects of learning and/or memoryBechara et al 1995; Eichenbaum and Cohen 2001; LeDoux996), but information processed within the amygdala alsoodulates synaptic activity and function within the hippocampal
ormation (Ikegaya et al 1996a, 1996b; Packard et al 1994;oozendaal and McGaugh 1997; Thomas et al 1984). The mostell-studied amygdala-mediated behavior is fear conditioning, inhich an animal learns to associate a tone with a shock
Eichenbaum and Cohen 2001; Fanselow and LeDoux 1999;eDoux 2000).
Gender is an important consideration, as males and femalesay have differential responses depending on the modality used
semantic vs. visual stimuli) and gender may have an effect onaterality of responses (Cahill 2003). Due to our decision to studyexual and physical abuse PTSD, our patient population wasargely female subjects. This should be kept in mind whenomparing with PTSD studies largely consisting of male subjectse.g., combat veterans), although our main findings are presentith introduction of gender as a nuisance covariable. Age is
able 3. PTSD Subjects Symptom Correlations
(PTe vs. NUe) vs.(PTl vs. NUl) PTe vs. PTl NUe vs. NUl
eft Amygdalax, y, z �24, 3, �18b �12, 0, �18Z �2.87 �2.85p Uncorrected .002 .002p Corrected SVC .013 (�21, 3, �18)a .019Volume (mm3) 270 162
ight Amygdalax, y, z 18, �3, �24 18, �3, �24 15, �3, �18Z �2.95 �2.54 �3.24p Uncorrected .002 .005 .001p Corrected SVC .012 (18, �6, �24)a .035 .008Volume (mm3) 648 108 972
Correlations Between CAPS B4 and temporal changes in amygdala acti-ation for PTSD and neutral words. p � .01, volume (within amygdala re-ion) �50 mm3. PTSD, posttraumatic stress disorder; SVC, small volumeorrection; CAPS, Clinician-Administered PTSD Scale; PT, PTSD words; PA,anic words; NU, neutral words; e, early first two epochs; l, late second twopochs.
aThese activations extend into the defined amygdala boundary, al-hough the maxima lie slightly outside of that boundary.
bFigure 9.
nother important consideration. One caveat is that although
none of the subjects were elderly and while average ages rangedfrom late 20s to mid 30s, the study groups are not entirelybalanced for age. Age can be a difficult variable to control for, asit may correlate with duration of illness. However, our keyfindings in this hypothesized region do not appear to be simplyan effect of age, as they are present, although to a slightly lowerthreshold, with age entered as a nuisance covariate.
In the interpretation of these findings, a limitation is theabsence of a trauma-exposure–matched control group. How-ever, the strong correlations between PTSD symptoms asmeasured by the CAPS scale and amygdala activations in ourconditions of interests strongly indicate that our results arerelevant to PTSD and not simply trauma exposure. Further-more, other studies from the literature support the finding oftrauma stimulus specific amygdala activation in PTSD versustrauma-exposed PTSD-negative control subjects (Shin et al1997).
Although our data highlight clear differences in the timecourse of amygdala response to differential emotional wordstimuli in PTSD and normal control subjects, any explanationfor the behavior of the amygdala in the normal controlsubjects, sensitization to PTSD words and habituation to panicwords, is speculative. One hypothesis is that the majority ofpanic words used in the study (death, illness, suffocate, etc.)are instantly evocative of negative associations, while a num-ber of the PTSD words used in the study (semen, intercourse,fondle, etc.) may require the context of the other words seenover time (rape, murder, assault, etc.) to become negativelyevocative in normal control subjects. Regardless of the reasonfor this pattern of amygdala activations in normal controlsubjects, it serves as an interesting baseline against whichdifferential stimulus-specific activations can be seen in PTSDpatients. While the initial relative increase of amygdala activityin PTSD patients to PTSD words (versus neutral words) fitswith the current literature, the sensitization to panic wordsindicates that rather than being hyperresponsive to negativestimuli in general, the amygdala in PTSD patients may beinitially increased to trauma-relevant versus neutral stimuliand actually exhibit a delayed response, in comparison withnormal control subjects, to nontrauma-related negative stim-uli.
Time courses of response to trauma-related and generalnegative stimuli have clinical relevance. Exposure therapy,which requires the patient to focus on and describe the detailsof a traumatic experience in a therapeutic manner, is the mostwell-established treatment for PTSD with many positive clin-ical outcome studies (Rothbaum et al 2000; Rothbaum andSchwartz 2002). However, there is still some debate over theefficacy of exposure therapy (Tarrier et al 1999).
Exposure methods share the common feature of confron-tation with frightening, yet realistically safe, stimuli thatcontinues until the anxiety is reduced (Rothbaum andSchwartz 2002). Habituation, a decreased response to thesame stimulus with repeated presentation, is one of thesimplest mechanisms accounting for this reduction in anxiety(Rothbaum and Schwartz 2002). It is essential for successfulexposure therapy that patients remain in the exposure situa-tion long enough for their anxiety to decrease (Astin andRothbaum 2000). Short exposures may further sensitize thepatient, making fear worse by leaving it unchecked (Roth-baum and Mellman 2001).
Thus, key issues in exposure therapy are the time courses
of habituation and sensitization to trauma-related stimuli inwww.elsevier.com/locate/biopsych

Pctotmbsrdsrad
eutarTia
“WG
BS
A
A
B
B
B
B
B
B
B
B
B
B
472 BIOL PSYCHIATRY 2005;57:464–473 X. Protopopescu et al
w
TSD subjects. We have demonstrated differential timeourses of amygdala response to different categories of emo-ionally valenced words in PTSD and normal control subjectsver a 24-minute study. This time window is comparable tohat employed in a single-exposure therapy session. Our dataay be relevant to mechanisms underlying exposure therapyy indicating that the amygdala of PTSD patients does notensitize to trauma-relevant words in this time period butather has a constant to diminishing response. We have alsoemonstrated correlations between the severity of a PTSDymptom, CAPS B4, and these time courses of amygdalaesponse. Such differential temporal patterns of amygdalactivation may provide information of relevance to therapeuticecision making.
A clearer understanding of the neurocircuitry of PTSD ismerging, as results accumulate from neuroimaging studiestilizing assorted psychological stimuli in different popula-ions of PTSD patients. This study extends reports of amygdalactivation to trauma-related stimuli by providing informationegarding temporal dynamics and specificity of response.hese features are an important consideration in understand-
ng the neurocircuitry of PTSD and have pathophysiologicalnd therapeutic implications.
This work was supported by NIMH Grant 5-P50MH58911,Center for Neural Systems of Fear and Anxiety”; the DeWittallace Fund of the New York Community Trust; and NIH MSTPrant GM07739 (XP).
We thank Paul Toth, Dan Zimmerman, Jude Allen, Josefinoorja, Hong Gu, Hilary Hochberg, James Murrough, and Erikatrohmayer for their help on this project.
merican Psychiatric Association (2000): Diagnostic Criteria from DSM-IV-TR.Washington, DC: American Psychiatric Association.
stin MC, Rothbaum BO (2000): Exposure therapy for the treatment ofposttraumatic stress disorder. NC-PTSD Clin Q 9:49 –54.
echara A, Tranel D, Damasio H, Adolphs R, Rockland C, Damasio AR (1995):Double dissociation of conditioning and declarative knowledge relativeto the amygdala and hippocampus in humans. Science 269:1115–1118.
eck AT, Ward CH, Mendelsohn M, Mock J, Erbaugh J (1981): An inventory formeasuring depression. Arch Gen Psychiatry 4:561–571.
ernstein EM, Putnam FW (1986): Development, reliability and validity of adissociation scale. J Nerv Ment Dis 174:727–735.
inder JR, Frost JA, Hammeke TA, Bellgowan PS, Rao SM, Cox RW (1999):Conceptual processing during the conscious resting state. A functionalMRI study. J Cogn Neurosci 11:80 –95.
lake DD, Weather FW, Nagy LM, Kaloupek DG, Gusman FD, Charney DS, et al(1995): The development of a clinician-administered PTSD scale.J Trauma Stress 8:75–90.
onne O, Brandes D, Gilboa A, Gomori JM, Shenton ME, Pitman RK, et al(2001): Longitudinal MRI study of hippocampal volume in trauma survi-vors with PTSD. Am J Psychiatry 158:1248 –1251.
reiter HC, Etcoff NL, Whalen PJ, Kennedy WA, Rauch SL, Buckner RL, et al(1996): Response and habituation of the human amygdala during visualprocessing of facial expression. Neuron 17:875– 887.
remner JD, Narayan M, Staib LH, Southwick SM, McGlashan T, Charney DS(1999a): Neural correlates of memories of childhood sexual abuse inwomen with and without posttraumatic stress disorder. Am J Psychiatry156:1787–1795.
remner JD, Randall P, Scott TM, Bronen RA, Seibyl JP, Southwick SM, et al(1995): MRI-based measurement of hippocampal volume in patientswith combat-related posttraumatic stress disorder [see comment]. Am JPsychiatry 152:973–981.
remner JD, Randall P, Vermetten E, Staib L, Bronen RA, Mazure C, et al(1997): Magnetic resonance imaging-based measurement of hippocam-pal volume in posttraumatic stress disorder related to childhood physi-
cal and sexual abuse–a preliminary report. Biol Psychiatry 41:23–32.ww.elsevier.com/locate/biopsych
Bremner JD, Staib LH, Kaloupek D, Southwick SM, Soufer R, Charney DS(1999b): Neural correlates of exposure to traumatic pictures and soundin Vietnam combat veterans with and without posttraumatic stress dis-order: A positron emission tomography study. Biol Psychiatry 45:806 –816.
Bremner JD, Vythilingam M, Vermetten E, Southwick SM, McGlashan T,Nazeer A, et al (2003a): MRI and PET study of deficits in hippocampalstructure and function in women with childhood sexual abuse and post-traumatic stress disorder. Am J Psychiatry 160:924 –932.
Bremner JD, Vythilingam M, Vermetten E, Southwick SM, McGlashan T, StaibLH, et al (2003b): Neural correlates of declarative memory for emotion-ally valenced words in women with posttraumatic stress disorder relatedto early childhood sexual abuse. Biol Psychiatry 53:879 – 889.
Buchel C, Morris J, Dolan RJ, Friston KJ (1998): Brain systems mediatingaversive conditioning: An event-related fMRI study. Neuron 20:947–957.
Cahill L (2003): Sex-related influences on the neurobiology of emotionallyinfluenced memory. Ann N Y Acad Sci 985:163–173.
Carrion VG, Weems CF, Eliez S, Patwardhan A, Brown W, Ray RD, et al (2001):Attenuation of frontal asymmetry in pediatric posttraumatic stress dis-order. Biol Psychiatry 50:943–951.
De Bellis MD, Hall J, Boring AM, Frustaci K, Moritz G (2001): A pilot longitudi-nal study of hippocampal volumes in pediatric maltreatment-relatedposttraumatic stress disorder. Biol Psychiatry 50:305–309.
De Bellis MD, Keshavan MS, Clark DB, Casey BJ, Giedd JN, Boring AM, et al(1999): A.E. Bennett Research Award. Developmental traumatology. PartII: Brain development [see comment]. Biol Psychiatry 45:1271–1284.
EichenbaumH,CohenNJ(2001): From Conditioning to Conscious Recollection: Mem-ory Systems of the Brain. Upper Saddle River, NJ: Oxford University.
Fanselow MS, LeDoux JE (1999): Why we think plasticity underlying Pavlov-ian fear conditioning occurs in the basolateral amygdala. Neuron 23:229 –232.
First MB, Spitzer RL, Gibbon M, Williams JBW (1997): Structured Clinical Inter-view for DSM-IV (SCID). Washington, DC: American Psychiatric Press.
Fischer H, Wright CI, Whalen PJ, McInerney SC, Shin LM, Rauch SL (2003):Brain habituation during repeated exposure to fearful and neutral faces:A functional MRI study. Brain Res Bull 59:387–392.
Foa E (1995): The Post Traumatic Diagnostic Scale Manual. Minneapolis: NCS.Friston KJ (1997): Testing for anatomically specified regional effects. Hum
Brain Mapp 5:133–136.Friston KJ, Frith CD, Turner R, Frackowiak RS (1995a): Characterizing evoked
hemodynamics with fMRI. Neuroimage 2:157–165.Friston KJ, Holmes AP, Price CJ, Buchel C, Worsley KJ (1999): Multisubject
fMRI studies and conjunction analyses. Neuroimage 10:385–396.Friston KJ, Holmes AP, Worsley KJ, Poline JP, Frith CD, Frackowiak RSJ
(1995b): Statistical parametric maps in functional imaging: A generallinear approach. Hum Brain Mapp 2:189 –210.
Golier J, Yehuda R (2002): Neuropsychological processes in post-traumaticstress disorder. Psychiatr Clin North Am 25:295–315.
Gu H, Feng H, Zhan W, Xu S, Silbersweig DA, Stern E, et al (2002): Single-shotinterleaved z-shim EPI with optimized compensation for signal lossesdue to susceptibility-induced field inhomogeneity at 3 T. Neuroimage17:1358 –1364.
Gurvits TV, Shenton ME, Hokama H, Ohta H, Lasko NB, Gilbertson MW, et al(1996): Magnetic resonance imaging study of hippocampal volume inchronic, combat-related posttraumatic stress disorder. Biol Psychiatry40:1091–1099.
Ikegaya Y, Saito H, Abe K (1996a): The basomedial and basolateral amygda-loid nuclei contribute to the induction of long-term potentiation in thedentate gyrus in vivo. Eur J Neurosci 8:1833–1839.
Ikegaya Y, Saito H, Abe K (1996b): Dentate gyrus field potentials evoked bystimulation of the basolateral amygdaloid nucleus in anesthetized rats.Brain Res 718:53– 60.
Isenberg N, Silbersweig D, Engelien A, Emmerich S, Malavade K, Beattie B, etal (1999): Linguistic threat activates the human amygdala. Proc Natl AcadSci U S A 96:10456 –10459.
LaBar KS, Gatenby JC, Gore JC, LeDoux JE, Phelps EA (1998): Human amyg-dala activation during conditioned fear acquisition and extinction: Amixed-trial fMRI study. Neuron 20:937–945.
LeDoux J (1996): Emotional networks and motor control: A fearful view. ProgBrain Res 107:437– 446.
LeDoux JE (2000): Emotion circuits in the brain. Annu Rev Neurosci 23:155–184.LeDoux JE, Gorman JM (2001): A call to action: Overcoming anxiety through
active coping. Am J Psychiatry 158:1953–1955.

L
M
M
M
M
O
P
P
P
Q
R
R
R
R
R
R
R
R
R
X. Protopopescu et al BIOL PSYCHIATRY 2005;57:464–473 473
iberzon I, Taylor SF, Amdur R, Jung TD, Chamberlain KR, Minoshima S, et al(1999): Brain activation in PTSD in response to trauma-related stimuli.Biol Psychiatry 45:817– 826.
cKiernan KA, Kaufman JN, Kucera-Thompson J, Binder JR (2003): A para-metric manipulation of factors affecting task-induced deactivation infunctional neuroimaging. J Cogn Neurosci 15:394 – 408.
cNally RJ (1997): Implicit and explicit memory for trauma-related informa-tion in PTSD. Ann N Y Acad Sci 821:219 –224.
organ MA, Romanski LM, LeDoux JE (1993): Extinction of emotional learn-ing: Contribution of medial prefrontal cortex. Neurosci Lett 163:109 –113.
orris JS, Frith CD, Perrett DI, Rowland D, Young AW, Calder AJ, et al (1996):A differential neural response in the human amygdala to fearful andhappy facial expressions. Nature 383:812– 815.
rr SP, Lasko NB, Shalev AY, Pitman RK (1995): Physiologic responses to loudtones in Vietnam veterans with posttraumatic stress disorder. J AbnormPsychol 104:75– 82.
ackard MG, Cahill L, McGaugh JL (1994): Amygdala modulation of hip-pocampal-dependent and caudate nucleus-dependent memory pro-cesses. Proc Natl Acad Sci U S A 91:8477– 8481.
helps EA, O’Connor KJ, Gatenby JC, Gore JC, Grillon C, Davis M (2001):Activation of the left amygdala to a cognitive representation of fear. NatNeurosci 4:437– 441.
itman RK, Shin LM, Rauch SL (2001): Investigating the pathogenesis ofposttraumatic stress disorder with neuroimaging. J Clin Psychiatry62(suppl 17):47–54.
uirk GJ, Russo GK, Barron JL, Lebron K (2000): The role of ventromedialprefrontal cortex in the recovery of extinguished fear. J Neurosci 20:6225– 6231.
auch SL, Shin LM, Segal E, Pitman RK, Carson MA, McMullin K, et al (2003a):Selectively reduced regional cortical volumes in post-traumatic stressdisorder. Neuroreport 14:913–916.
auch SL, Shin LM, Wright CI (2003b): Neuroimaging studies of amygdalafunction in anxiety disorders. Ann N Y Acad Sci 985:389 – 410.
auch SL, van der Kolk BA, Fisler RE, Alpert NM, Orr SP, Savage CR, et al (1996):A symptom provocation study of posttraumatic stress disorder usingpositron emission tomography and script-driven imagery. Arch Gen Psy-chiatry 53:380 –387.
auch SL, Whalen PJ, Shin LM, McInerney SC, Macklin ML, Lasko NB, et al (2000):Exaggerated amygdala response to masked facial stimuli in posttraumaticstress disorder: A functional MRI study. Biol Psychiatry 47:769–776.
eiss S, Peterson RA, Gursky DM, McNally RJ (1986): Anxiety sensitivity, anxietyfrequency and the predictions of fearfulness. Behav Res Ther 24:1–8.
oozendaal B, McGaugh JL (1997): Basolateral amygdala lesions block thememory-enhancing effect of glucocorticoid administration in the dorsalhippocampus of rats. Eur J Neurosci 9:76 – 83.
othbaum BO, Meadows EA, Resick P, Foy DW (2000): Cognitive-behavioraltherapy. In: Keane T, editor. Effective Treatments for Posttraumatic StressDisorder: Practice Guidelines from the International Society for TraumaticStress Studies. New York: Guilford, 60 – 83.
othbaum BO, Mellman TA (2001): Dreams and exposure therapy in PTSD.J Trauma Stress 14:481– 490.
othbaum BO, Schwartz AC (2002): Exposure therapy for posttraumaticstress disorder. Am J Psychother 56:59 –75.
Schuff N, Neylan TC, Lenoci MA, Du AT, Weiss DS, Marmar CR, et al (2001):Decreased hippocampal N-acetylaspartate in the absence of atrophy inposttraumatic stress disorder. Biol Psychiatry 50:952–959.
Shalev AY, Orr SP, Peri T, Schreiber S, Pitman RK (1992): Physiologic re-sponses to loud tones in Israeli patients with posttraumatic stress disor-der. Arch Gen Psychiatry 49:870 – 875.
Shin LM, Kosslyn SM, McNally RJ, Alpert NM, Thompson WL, Rauch SL, et al(1997): Visual imagery and perception in posttraumatic stress disorder. Apositron emission tomographic investigation. Arch Gen Psychiatry 54:233–241.
Shin LM, McNally RJ, Kosslyn SM, Thompson WL, Rauch SL, Alpert NM, et al(1999): Regional cerebral blood flow during script-driven imagery inchildhood sexual abuse-related PTSD: A PET investigation. Am J Psychia-try 156:575–584.
Shin LM, Orr SP, Carson MA, Rauch SL, Macklin ML, Lasko NB, et al (2004):Regional cerebral blood flow in the amygdala and medial prefrontalcortex during traumatic imagery in male and female Vietnam veteranswith PTSD. Arch Gen Psychiatry 61:168 –176.
Shin LM, Whalen PJ, Pitman RK, Bush G, Macklin ML, Lasko NB, et al (2001): AnfMRI study of anterior cingulate function in posttraumatic stress disor-der. Biol Psychiatry 50:932–942.
Spielberger CD, Jacobs G, Russell S, Crane R (1983): Assessment of anger: TheState-Trait Anger Scale. In: Spielberger CD, editor. Advances in Personal-ity Assessment, vol 2. Hillsdale, NJ: Lawrence Erlbaum.
Spitzer RL, Williams JBW, Gibbon M, First MB (1994): Structured Clinical Inter-view for DSM-IV - Patient Edition. New York: New York State PsychiatricInstitute, Biometrics Research Department.
Stein MB, Koverola C, Hanna C, Torchia MG, McClarty B (1997): Hippocampalvolume in women victimized by childhood sexual abuse. Psychol Med27:951–959.
Tarrier N, Pilgrim H, Sommerfield C, Faragher B, Reynolds M, Graham E, et al(1999): A randomized trial of cognitive therapy and imaginal exposure inthe treatment of chronic posttraumatic stress disorder. J Consult ClinPsychol 67:13–18.
Thomas SR, Assaf SY, Iversen SD (1984): Amygdaloid complex modulatesneurotransmission from the entorhinal cortex to the dentate gyrus ofthe rat. Brain Res 307:363–365.
Tzourio-Mazoyer N, Landeau B, Papathanassiou D, Crivello F, Etard O, Del-croix N, et al (2002): Automated anatomical labeling of activations inSPM using a macroscopic anatomical parcellation of the MNI MRI single-subject brain. Neuroimage 15:273–289.
Vermetten E, Bremner JD (2002): Circuits and systems in stress. II. Applica-tions to neurobiology and treatment in posttraumatic stress disorder.Depress Anxiety 16:14 –38.
Villarreal G, King CY (2001): Brain imaging in posttraumatic stress disorder.Semin Clin Neuropsychiatry 6:131–145.
Worsley KJ, Marrett S, Neelin P, Vandal AC, Friston KJ, Evans AC (1996): Aunified statistical approach for determining significant signals in imagesof cerebral activation. Hum Brain Mapp 4:58 –73.
Wright CI, Fischer H, Whalen PJ, McInerney SC, Shin LM, Rauch SL (2001):Differential prefrontal cortex and amygdala habituation to repeatedly
presented emotional stimuli. Neuroreport 12:379 –383.www.elsevier.com/locate/biopsych




















