
Deconvolution-free Subcellular Imaging with Axially Swept Light Sheet Microscopy
Upload
independentCategory
view
3download
0
Dit
Sa
b
a
ARRAA
KHTTSNP
1
oe
pAPtx
f
0h
Plant Science 196 (2012) 53– 66
Contents lists available at SciVerse ScienceDirect
Plant Science
jo u rn al hom epa ge: www.elsev ier .com/ locate /p lantsc i
ifferential subcellular targeting of recombinant human �1-proteinase inhibitornfluences yield, biological activity and in planta stability of the protein inransgenic tomato plants
hweta Jhaa, Saurabh Agarwala, Indraneel Sanyala, G.K. Jainb, D.V. Amlaa,∗
Plant Transgenic Lab, CSIR-National Botanical Research Institute, Rana Pratap Marg, P.O. Box 436, Lucknow 226 001, IndiaDivision of Pharmacokinetics & Metabolism, CSIR-Central Drug Research Institute, Chattar Manzil, Post Box No. 173, Lucknow 226 001, India
r t i c l e i n f o
rticle history:eceived 29 May 2012eceived in revised form 6 July 2012ccepted 9 July 2012vailable online 28 July 2012
eywords:uman �1-proteinase inhibitorherapeutic recombinant proteinransgenic tomato plantsubcellular localization-glycosylationrotein stability
a b s t r a c t
The response of protein accumulation site on yield, biological activity and in planta stability of therapeuticrecombinant human �1-proteinase inhibitor (�1-PI) was analyzed via targeting to different subcellularlocations, like endoplasmic reticulum (ER), apoplast, vacuole and cytosol in leaves of transgenic tomatoplants. In situ localization of the recombinant �1-PI protein in transgenic plant cells was monitored byimmunohistochemical staining. Maximum accumulation of recombinant �1-PI in T0 and T1 transgenictomato plants was achieved from 1.5 to 3.2% of total soluble protein (TSP) by retention in ER lumen,followed by vacuole and apoplast, whereas cytosolic targeting resulted into degradation of the protein.The plant-derived recombinant �1-PI showed biological activity for elastase inhibition, as monitored byresidual porcine pancreatic elastase (PPE) activity assay and band-shift assay. Recombinant �1-PI waspurified from transgenic tomato plants with high yield, homogeneity and biological activity. Purifiedprotein appeared as a single band of ∼48–50 kDa on SDS-PAGE with pI value ranging between 5.1 and5.3. Results of mass spectrometry and optical spectroscopy of purified recombinant �1-PI revealed thestructural integrity of the recombinant protein comparable to native serum �1-PI. Enzymatic deglyco-sylation and lectin-binding assays with the purified recombinant �1-PI showed compartment-specificN-glycosylation of the protein targeted to ER, apoplast and vacuole. Conformational studies based onurea-induced denaturation and circular dichroism (CD) spectroscopy revealed relatively lower stability
of the recombinant �1-PI protein, compared to its serum counterpart. Pharmacokinetic evaluation ofplant derived recombinant and human plasma-purified �1-PI in rat, by intravenous route, revealed sig-nificantly faster plasma clearance and lower area under curve (AUC) of recombinant protein. Our datasuggested significance of protein sorting sequences and feasibility to use transgenic plants for the produc-tion of stable, glycosylated and biologically active recombinant �1-PI for further therapeutic applications.© 2012 Elsevier Ireland Ltd. All rights reserved.
. Introduction
Plants offer an attractive alternative platform for the productionf recombinant therapeutic and industrial proteins owing to cost-ffective economical feasibility, free from contamination of human
Abbreviations: �1-PI, �1-proteinase inhibitor; ER, endoplasmic reticulum; PPE,orcine pancreatic elastase; CaMV35S, cauliflower mosaic virus 35S promoter; AMV,lfalfa mosaic virus; UTR, untranslated region; TIC, translation initiation context;R1a, pathogenesis related protein 1a; SPS-NTPP, sweet potato sporamine A – N-erminal propeptide; PNGase F, peptide N-glycosidase F; CD, circular dichroism; xyl,ylose; fuc, fucose.∗ Corresponding author. Tel.: +91 522 2297954/2297955;
ax: +91 522 2205836/2205839; mobile: +91 9935392989.E-mail addresses: [email protected], [email protected] (D.V. Amla).
168-9452/$ – see front matter © 2012 Elsevier Ireland Ltd. All rights reserved.ttp://dx.doi.org/10.1016/j.plantsci.2012.07.004
and animal pathogens, scalability and capability to perform com-plex post-translational modifications including glycosylation [1,2].High-level expression of stable heterologous proteins in plantsdepends on the proteolytic degradation, post-translational pro-cessing and accumulation of foreign protein during the life cycle oftransgenic plants [3–5]. Therefore, different strategies are requiredto increase the expression, yield and structural stability of recombi-nant proteins including stacking and accumulation of recombinantforeign proteins into various subcellular locations in the plant cells[6].
Human �1-proteinase inhibitor (�1-PI), also known as �1-
antitrypsin belongs to serpin super-family and is a majorcomponent of human plasma that protects elasticity of lungs byinhibiting the activity of serine proteases, particularly neutrophilelastase [7]. Single amino acid substitution (Glu 342 → Lys) in the
5 cienc
mi[iawscehsnptoi[
tppdafpmwgglobp
2
2
Pe[sotPdapBtmethngt(fowTts
4 S. Jha et al. / Plant S
obile domain of the protein results into �1-PI deficiency, lead-ng to potentially lethal pulmonary emphysema and liver cirrhosis8]. Intravenous augmentation of purified �1-PI from human serums the only available clinical treatment, which is in great demandnd always in limited supply along with high risk of contaminationith blood-borne pathogens [9]. Diverse alternative expression
ystems including microbes, yeast, transgenic animals and plantell cultures have been considered over the period to achieve over-xpression of glycosylated biologically active recombinant �1-PI,owever, none of them could fulfill the requirements of clinicallyafe, biologically active and economical production of recombi-ant �1-PI for therapeutic applications [10]. Genetically engineeredlants offer unique feasibility and versatility of large scale produc-ion for tissue and organ-specific accumulation of diverse typesf therapeutic and industrial recombinant proteins like antibod-es, vaccine antigens, enzymes and growth factors in plant tissues11–13].
Reports have illustrated the positive impact of sequesteringhe heterologous therapeutic and industrial proteins into ER oflant cells for higher accumulation, proper folding and proteolyticrotection of the recombinant proteins [11,14,15] whereas otherestinations like apoplast, vacuoles and peroxisomes have beenttributed for optimal glycosylation and stability [1,6,16]. There-ore appropriate subcellular targeting of recombinant proteins intolant cells seems to influence the extent of post-translationalodifications, integrity and the final yield. In the present study,e have expressed the human serum �1-PI protein in trans-
enic tomato plants from the modified and codon-optimizedene, and targeted the recombinant protein to different subcel-ular locations to assess the impact of subcellular environmentn accumulation, final yield, in planta stability, glycosylation andiological activity of the foreign protein in the transgenic tomatolants.
. Materials and methods
.1. Plant expression vectors and transformation of tomato plants
The modified and codon-optimized 1182 bp sequence of �1-I gene (GenBank accession no. EF638826) designed for optimumxpression in dicot plants has been used in the present study17]. The native transit peptide sequence of human �1-PI wasubstituted with 90 bp modified PR1a signal peptide sequencef tobacco (GenBank accession no. EF638827) in the pPWK vec-or for targeting of the recombinant protein to apoplast, whileR1a in conjunction with KDEL motif at 3′ end was used toevelop pPAK vector for ER retention of the protein (Fig. 1a, b,nd e). The 114 bp modified transit peptide sequence of sweetotato sporamine with N-terminus propeptide (SPS-NTPP) (Gen-ank accession no. EF638829) without 3′ KDEL motif was usedo develop pSWK vector for vacuolar targeting (Fig. 1f), while
odified gene without any flanking sequences either on 5′ or 3′
nd was used to develop pWSP vector for cytosolic accumula-ion of the protein. The 75 bp native signal peptide sequence ofuman �1-PI gene was also codon-optimized (GenBank accessiono. EF638828) and incorporated 5′ upstream of modified �1-PIene along with or without ER-retention motif KDEL, and the resul-ant constructs were designated as pNAK and pNWK, respectivelyFig. 1d). Agrobacterium tumefaciens strain LBA4404 was trans-ormed with these six vectors and used for nuclear transformation
f tomato (Solanum lycopersium var. PED) using leaf-disc methodith some specific modifications [18,19]. The kanamycin-resistant0 plantlets were developed under culture room conditions andhen transferred to glasshouse for growth, flower development andeed setting.
e 196 (2012) 53– 66
2.2. Molecular characterization of transgenic tomato plants
Genomic DNA was isolated from 100 mg of young leaf tis-sues of transgenic tomato plants using GenElute plant genomicDNA miniprep kit according to manufacturer’s instructions (Sigma,USA). PCR analysis and Southern hybridization with genomic DNAwere performed for detection of �1-PI gene [20]. Total RNA wasisolated from 100 mg fresh leaves of transgenic tomato plants in1 ml TRI-reagent for analysis of �1-PI transcript. The first strand ofcDNA was synthesized with enhanced Avian HS RT-PCR kit accord-ing to manufacturer’s instructions (Sigma, USA). Northern blottingwas performed using ∼30 �g total RNA resolved on 1.2% denatur-ing formaldehyde agarose gel, transferred onto Zeta Probe GT nylonmembrane (Bio-Rad, USA) and hybridized at 42 ◦C for 16–18 h with� [32P] dCTP labeled full-length �1-PI gene probe [20]. Relativequantification of �1-PI transcript(s) was performed by real-timeRT-PCR using 100 ng of cDNA template. The quantitative data wererepresented as mean values and standard error of three indepen-dent experiments with three replicates of T0 transgenic plantsdeveloped with different vectors.
2.3. Immunohistochemical localization of recombinant ˛1-PI
Immunolocalization of recombinant �1-PI in young leaf tissuesof transgenic tomato plants was performed by fixing the leaf pieces(�1 mm2) in 4% (w/v) paraformaldehyde for 60 min at low vacuum(5 Hg inch−1), followed by washing with 1× microtubule stabiliz-ing solution (15 g PIPES, 1.9 g EGTA, 1.32 g MgSO4·7H2O, 5 g KOH,pH 6.8–7.0). Leaf tissues were cut into thin sections (�10 �m),washed with PBST, incubated in blocking solution (2% BSA) at37 ◦C followed by incubation with rabbit anti �1-PI primary anti-body (1:300 dilution) overnight at 4 ◦C, and subsequently withanti-rabbit secondary antibody (1:100 dilution) coupled with Cy-3fluorescence dye for 2 h (Sigma, USA). The same leaf sections weretreated with unlabeled anti-rabbit secondary antibody and incu-bated overnight with rabbit anti-calreticulin (ER-specific marker)primary antibody (1:300 dilution) at 4 ◦C, followed by incubationwith FITC-conjugated anti-rabbit secondary antibody (1:1000 dilu-tion) (Sigma, USA). Stained sections were mounted in fluoromountand fluorescence images were captured on LSM 510 META con-focal laser scanning microscope (Zeiss, Germany) with LSM METAsoftware, using 543 nm helium–neon and 488 nm argon ion lasersources, to record the fluorescence at 560 nm and 525 nm for Cy-3and FITC signals, respectively.
2.4. Quantitative estimation of recombinant ˛1-PI protein
Cell free plant extracts were prepared from leaves of 12-weekold transgenic tomato plants and quantification of recombinant �1-PI protein in the crude extracts or purified protein samples wasperformed by direct antigen coating-enzyme linked immunosor-bent assay (DAC-ELISA) using the commercial anti-human �1-PIantibody (Sigma, USA) as described earlier [17]. Expression levelswere quantified on a linear standard curve plotted with pure humanserum �1-PI protein (Sigma, USA).
2.5. Western immunoblotting and two-dimensional SDS-PAGE
A 50 �g of cell free plant extract or 100 ng of purified recom-binant �1-PI protein sample was fractionated on 12% SDS-PAGEfollowed by staining with silver salts according to the standardprocedure [20]. Electrophoresed protein samples were transferred
onto Immobilon polyvinylidene difluoride (PVDF) membrane(Millipore, USA) for Western blotting using the commercial anti-human �1-PI antibodies [17]. Recombinant �1-PI protein samplespurified from the transgenic tomato plants were analyzed by
S. Jha et al. / Plant Science 196 (2012) 53– 66 55
Fig. 1. Schematic representation of T-DNA region having the modified synthetic �1-PI gene with different modified and codon optimized signal peptide sequences at 5′ end.(a) With KDEL or (b) without KDEL sequence at 3′ end as ER retention motif. (c) Multiple cloning site (MCS) having sequences for four restriction enzymes and optimal TICsequence having purines at −3 and +4 sites around ATG initiator codon. (d) 5′ modified native signal sequence of human �1-PI with KDEL motif at 3′ end (a) to developpNAK vector for ER retention, and without KDEL motif (b) for development of the vector pNWK for apoplast targeting. (e) 5′ modified PR1a signal sequence of Nicotianatabacum with KDEL motif at 3′ end (a) to develop pPAK vector for ER retention, and without KDEL motif (b) for development of the vector pPWK for apoplast targeting. (f)5′ modified sporamine-A signal sequence of Ipomoea batatas with N-terminal propeptide (SPS-NTPP) and without KDEL motif at 3′ end (b) to develop the vector pSWK forv equenn ficatioa ation c
tInr3nE(aeebiDwi
2i
era
acuolar targeting; whereas modified �1-PI gene without any 5′ and 3′ flanking sative nucleotide sequences of signal peptides are shown in capital letters and modifter the initiation codon ATG in (d) and (f) to provide an optimum translation initi
wo-dimensional (2-DE) gel electrophoresis as described by [21].soelectric focusing was carried out with 1 �g of purified recombi-ant protein using IPG ReadyStrip (7 cm, pH 3–10; Bio-Rad, USA)ehydrated in buffer containing 8 M urea, 2% CHAPS, 0.5% Bio-Lyte/10 ampholyte, 25 mM dithiothreitol (DTT) and 0.002% bromophe-ol blue (BPB). Isoelectric focusing was performed at 23 ◦C in thettan IPGphor 3 system (Amersham Biosciences, USA) in five steps0:30 h at 250 V, 0:15 h at 450 V, 0:15 h at 750 V, 0:30 h at 2000 Vnd 2:00 h at 8000 V), followed by equilibration for 15 min in SDSquilibration buffer (50 mM Tris–HCl, pH 8.8, 6 M urea, 30% glyc-rol, 2% SDS, 20 mM DTT and 0.002% BPB) and for 15 min in the sameuffer containing 125 mM iodoacetamide, without DTT. Separation
n the second dimension was carried out on 10% SDS-PAGE. All 2-E gels were run in replicates for each sample; one was stainedith SYPRO-Ruby (Sigma, USA) and another was used for Western
mmunoblotting with �1-PI specific antibodies.
.6. Biological activity of recombinant ˛1-PI protein for elastasenhibition
The biological activity of recombinant �1-PI in cell free plantxtracts and of purified protein samples was determined byesidual porcine pancreatic elastase (PPE) inhibition activityssay using N-succinyl-Ala-Ala-Ala-p-nitroanilide (Sigma, USA) as
ces led to development of pWSP vector for cytosolic targeting of the protein. Thens incorporated are shown in small letters. An extra GCT codon for alanine is addedontext (TIC) sequence harboring G at position +4.
chromogenic substrate. The detailed procedure has been describedearlier [17]. Pure human serum �1-PI was used as a standard toquantify the biologically active recombinant �1-PI.
2.7. Band shift assay
Purified human serum or recombinant �1-PI protein sampleswere incubated with 100 pmol of PPE in equimolar ratio for 20 minat 37 ◦C. Aliquots of the samples were electrophoresed on 12%SDS-PAGE gel followed by staining and documentation for shift inmobility of bands compared to untreated samples.
2.8. Stoichiometry of elastase inhibition
Stoichiometry of inhibition (SI) of elastase enzyme by �1-PIprotease inhibitor (human serum and ER-targeted recombinant �1-PI) was determined by incubating 100 pmol of PPE with varyingamounts of native or recombinant �1-PI (I/E molar ratio of 0, 0.5,1.0, 1.5, 2.0) in the standard assay buffer (20 mM Tris–HCl, pH 8.0,
150 mM NaCl, 0.01% Tween-80) at 25 ◦C. The amount of remainingactive elastase enzyme was measured by taking aliquots at differ-ent time intervals. The activities were expressed as fractions of PPEactivity without �1-PI. The fraction of active enzyme in each sample
5 cienc
wfi
2p
peg(MtsiT5afiiaUs0uent(f
2
aiosie
2˛
pUto
2
CibawBsaxwip
6 S. Jha et al. / Plant S
as plotted against the ratio of [�1-PI]/[PPE] and the results weretted with linear regression to determine the SI value.
.9. Purification of recombinant ˛1-PI from transgenic tomatolants
A 100 g fresh leaf tissue from the promising transgenic tomatolants developed with different vectors and showing maximumxpression of recombinant �1-PI was homogenized in liquid nitro-en and resuspended in five volumes of homogenization buffer20 mM Tris–HCl, pH 7.4, 150 mM NaCl, 10 mM EDTA, 14 mM �-
E and 0.05% Triton X-100). The crude homogenate was filteredhrough nylon mesh, centrifuged at 12,000 rpm for 20 min andoluble proteins in the supernatant were precipitated by addingncreasing amounts of ammonium sulphate up to 50% saturation.he remaining proteins in the supernatant were precipitated with0–95% saturation of ammonium sulphate, resuspended, dialyzedgainst equilibration buffer (0.1 M Tris–HCl, pH 8.0, 0.5 M NaCl) andltered through 0.22 �m syringe filter (Whatman, CA) prior to load-
ng onto immunoaffinity column prepared by coupling of rabbitnti �1-PI antibody to CNBr-activated Sepharose 4B matrix (Sigma,SA). The column was pre-equilibrated and the clarified protein
amples were loaded onto the antibody column at a flow rate of.5 ml min−1. The column was washed with equilibration bufferntil A280 of the effluent reached to zero. The bound antigen wasluted with 0.1 M Na2CO3, pH 11.2, 0.5 M NaCl and immediatelyeutralized with 1 M Tris–HCl, pH 6.8. The eluted fractions con-aining �1-PI protein were pooled, dialyzed against storage buffer50 mM Tris–HCl, pH 7.5, 75 mM NaCl), concentrated and used forurther biochemical investigations.
.10. Mass spectrometric analysis
Protein spots were excised from SYPRO-Ruby stained 2-DE gelsnd ‘in-gel’ digested with trypsin [22]. Identification and character-zation of purified protein samples and molecular weight analysisf intact protein was performed with 4800 MALDI-TOF/TOF masspectrometer (Applied Biosystems, USA) in linear high mass pos-tive and reflector positive ion mode, respectively, as describedarlier [23].
.11. Enzymatic deglycosylation of plant derived recombinant1-PI
The purified human serum and recombinant �1-PI protein sam-les were treated with peptide N-glycosidase F (PNGase F, NEB,SA) of Flavobacterium meningosepticum according to manufac-
urer’s instructions, and the shift in molecular mass due to removalf glycan chains from the protein was analyzed on SDS-PAGE.
.12. N-Glycan analysis by lectin and specific antibody binding
Binding of glycan chains of recombinant �1-PI withoncanavalin-A (Con-A, from Canavalia ensiformis) was exam-
ned on Con-A Sepharose 4B column (Sigma, USA) followedy Western immunoblotting with �1-PI specific antibodies, orffinoblotting with biotin-labeled Con-A followed by incubationith AP-conjugated streptavidin (Sigma, USA) and detection withCIP/NBT. Binding of recombinant �1-PI to DSA-lectin (from Daturatramonium) was investigated using DIG glycan differentiation kit
ccording to manufacturer’s instructions (Roche, Germany). Anti-yl and anti-fuc rabbit antibodies (Agrisera antibodies, Sweden)ere used for detection of �-1,2 xylose and �-1,3 fucose residuesn N-glycan chains of different glycoforms of recombinant �1-PIrotein.
e 196 (2012) 53– 66
2.13. Urea-induced equilibrium unfolding transition
The conformational stability of purified human serum proteinand ER-targeted recombinant �1-PI was measured as a function ofurea induced equilibrium unfolding by fluorescence spectroscopyand circular dichroism (CD) spectroscopy. Aliquotes of purified pro-teins (4 �M for fluorescence and 3 �M for CD measurements) wereallowed to equilibrate in equilibration buffer (10 mM phosphatepH 6.5, 50 mM NaCl, 1 mM EDTA, 1 mM �-ME) containing differ-ent concentrations of urea (0–9 M) for 8 h at 25 ◦C. The changesin intrinsic tryptophan fluorescence were measured using PerkinElmer LS-55 luminescence spectrometer, following excitation at280 nm and emission at 360 nm at slit width of 5 nm. The CD signalswere recorded at 25 ◦C with JASCO J-800 spectropolarimeter cali-brated with ammonium (+)-10-camphor sulfonate, by continuouswavelength scans from 200 to 250 nm at scan speed of 20 nm/minusing 0.1 cm path length cell. Average of two scans per samplewas calculated for measurement of CD ellipticity (�). Experimentaldata of the fluorescence measurements were fitted to a two-stateunfolding model, and the free energy of stabilization in the absenceof denaturant, �G(H2O), was determined as described earlier [24].
2.14. Thermostability analysis
To monitor the thermostability of ER-targeted recombinant �1-PI and human serum �1-PI, the purified protein samples (5 �g ml−1)were incubated at 54 ◦C followed by measurement of remaininginhibitory activity against elastase at periodic intervals. Half-life ofthermal denaturation was calculated from fitting the experimentaldata into a single exponential decay.
2.15. Pharmacokinetic evaluation of recombinant ˛1-PI
The pharmacokinetic parameters of ER-targeted recombinant�1-PI in male Sprague-Dawley rats weighing 200–220 g (n = 3)were compared with human plasma-derived native �1-PI, by intra-venous administration into lateral tail vein. Aliquots of 250 �l bloodwas collected from retro-orbital plexus of rats under mild etheranesthesia into heparinized microfuge tubes at intervals of 0.5,15, 30, 60, 90, 120, 180 and 240 min post-administration. Plasmawas harvested by centrifugation at 15,700 × g for 10 min, stored at−70 ± 1 ◦C and used for the quantitative estimation of �1-PI con-centration by ELISA as described earlier [17].
3. Results
3.1. Construction of plant expression vectors and development oftransgenic tomato plants
Six plant expression vectors were developed harboring the1.2 kb modified synthetic �1-PI gene with different transit peptidesequences at 5′-end in the backbone of pBI121 for targeting andaccumulation of the recombinant protein to different subcellularlocations in transgenic tomato plants (Fig. 1d–f). The modified �1-PI gene is expressed under the control of CaMV35S double enhancerconstitutive promoter along with 38 bp AMV 5′-UTR sequence, amultiple cloning site (MCS) and translation initiation context (TIC)at 5′ end of the gene (Fig. 1c). The binary vector containing the mod-ified synthetic �1-PI gene with PR1a transit peptide sequence at 5′
end and ER retention motif – KDEL at 3′ end (pPAK) was used forretention of the recombinant protein in ER while the same constructwithout KDEL sequence (pPWK) was used for apoplast targeting
(Fig. 1a, b, and e). The plant-preferred codon-optimized native tran-sit peptide of human �1-PI gene with or without KDEL (pNAK andpNWK constructs) were used for targeting the recombinant �1-PIto ER and secretory pathway, respectively (Fig. 1a, b, and d). The
S. Jha et al. / Plant Science
Fig. 2. Assessment of �1-PI transcript in T0 transgenic tomato plants developedwith different vectors. (a-i) Northern blot hybridized with �1-PI specific radiola-beled probe. Lane 1, negative control; lane 2–13, transgenic plants developed withdifferent vectors: lane 2–4, pPAK; lane 5–7, pPWK; lane 8–10, pSWK and lane 11–13,pWSP vectors. (a-ii) The corresponding RNA gel showing clear intact bands of rRNAisolated from transgenic plants developed with respective vectors as in (a-i). About30 �g of total RNA was used in all the samples. (b) Relative quantitation of �1-PItranscript by real time RT-PCR of transgenic plants developed with different vec-tors in reference to the vector pPAK. Tomato ˇ-actin gene was taken as endogenouscie
bau5w�t
3
dpaiwstwspSstaipsaplwetv
ontrol. Relative values are represented as mean value ± standard error of threendependent experiments with three replicates of transgenic plants developed withach vector.
inary vector substituted with SPS-NTPP transit peptide sequencet 5′ end and without KDEL motif at 3′ end (pSWK) was used for vac-olar targeting (Fig. 1b and f) whereas the construct without any′ transit peptide signal sequence or 3′ ER retention motif (pWSP)as used for expression of recombinant �1-PI into cytosol. These1-PI gene vectors were used for Agrobacterium-mediated nuclear
ransformation of tomato for developing stable transgenic lines.
.2. Molecular analysis of stable transgenic tomato plants
Several independent stable T0 transgenic tomato plants wereeveloped with each vector following kanamycin selection (30 withPAK, 18 with pNAK, 30 with pPWK, 21 with pNWK, 27 with pSWKnd 24 with pWSP vectors, respectively). The integration of mod-fied �1-PI transgene in the genome of transgenic tomato plants
as confirmed by PCR and Southern hybridization while expres-ion of �1-PI transcript was confirmed by RT-PCR. Results showedhe amplification of anticipated fragment of 1182 bp for �1-PI geneith specific set of primers in PCR and RT-PCR assays and no
uch amplification was observed in untransformed control tomatolants under identical assay conditions (Supplementary Fig. S1).outhern hybridization of genomic DNA from T0 transgenic plantshowed single copy insertion of �1-PI transgene in most of theransgenic plants except few plants showing higher copy number,nd the results also suggested random integration of the transgenen the genome of transgenic tomato plants (data not shown). PCRositive transgenic tomato plants were analyzed for �1-PI tran-cript by Northern blot hybridization which showed the presence of
unique band of the expected size (1182 bp) in all the T0 transgeniclants developed with different vectors (Fig. 2a). The transcript
evel of the modified �1-PI gene in promising transgenic plants
as quantified by real-time RT-PCR using tomato ˇ-actin gene asndogenous control. The relative values obtained from quantita-ive RT-PCR revealed steady-state level of mRNA and no significantariation in the amount of �1-PI transcript expressed in transgenic
196 (2012) 53– 66 57
plants developed with different vectors (Fig. 2b). The specificityof reaction products were verified by agarose gel electrophoresisand melt-curve analysis, revealing anticipated amplicon of 125 bpfor �1-PI gene and 194 bp for ˇ-actin gene. These results indicatedactive transcription of modified �1-PI gene, and excluded any sub-stantial contribution of transgene integration site and transcriptstability for differential pattern of recombinant �1-PI protein accu-mulation in various subcellular locations.
3.3. Subcellular localization of recombinant ˛1-PI in leaves oftransgenic plants
The efficiency of various transit peptide sequences used in thisstudy for subcellular targeting of recombinant �1-PI was monitoredby immunohistochemical localization of the recombinant proteinin young leaf cells of transgenic tomato plants developed withdifferent vectors using laser confocal microscopy. Leaf sectionsfrom different transgenic plants of tomato were fixed, permeabi-lized and incubated successively with rabbit anti-�1-PI antibodyand a secondary anti-rabbit antibody conjugated to either Cy-3or FITC fluorescent dyes. Leaf sections from non-transgenic plantwere used as negative control, which exhibited no fluorescencesignal except very faint auto-fluorescence of the cell wall (Fig. 3a).A typical ER localized fluorescence pattern was observed in trans-genic plants developed with pPAK vector (Fig. 3b, first panel) whichreflected maximum accumulation and retention of recombinantprotein in ER. A similar pattern of fluorescence was observed forthe calreticulin protein, an ER-resident chaperone, specific markerand internal control for ER co-localization (Fig. 3b, second panel).Intense immunostaining of apoplast was noticed in leaf cells oftransgenic plants developed with pPWK vector, indicating efficientsorting and maximum accumulation of recombinant �1-PI in cellwall and intercellular spaces (Fig. 3c). Transgenic tomato plantsdeveloped with pWSP vector without flanking sequences on bothends exhibited accumulation of recombinant �1-PI protein in thecytosol (Fig. 3d) while the plants transformed with pSWK constructshowed maximum accumulation of recombinant �1-PI in vacuoles,which were co-localized with acidic compartments of the cells viaspecific staining with lyso-tracker dye as internal control (Fig. 3eand f). These observations illustrate efficient sorting and accumula-tion of the recombinant �1-PI protein in corresponding subcellularlocations in the leaves of transgenic tomato plants developed withdifferent chimeric vectors.
3.4. Expression of recombinant ˛1-PI protein in transgenictomato plants
PCR-positive transgenic tomato plants were analyzed for quan-titative expression of �1-PI protein by DAC-ELISA and Westernimmunoblotting using the pure human serum �1-PI protein asthe standard. Apparent quantitative differences in expression ofrecombinant �1-PI were observed in cell free leaf extracts of indi-vidual T0 transgenic tomato plants developed with six differentplant transformation vectors. The quantity of recombinant �1-PIprotein accumulated in T0 population of transgenic plants devel-oped with corresponding vectors ranged from 0.44 to 1.55% of TSPfor pPAK, 0.15 to 1.1% of TSP for pNAK (both for ER), 0.026 to 1.3%of TSP for pPWK, 0.01 to 1.7% of TSP for pNWK (both for apoplast),0.14 to 1.8% of TSP for pSWK (for vacuole) and 0.12 to 0.43% of TSPfor pWSP (for cytosol), and average values obtained for expressionof recombinant �1-PI are shown in Fig. 4a. The comparative anal-ysis of T0 transgenic population revealed that recombinant �1-PI
protein targeted to ER showed 1.5–4.0 fold increased accumula-tion in comparison to protein targeted to vacuole, apoplast andcytosol, as validated statistically significant by Student’s t-test at0.01% level of significance. Western immunoblotting performed
58 S. Jha et al. / Plant Science 196 (2012) 53– 66
Fig. 3. Immunohistochemical localization of recombinant �1-PI targeted to different subcellular locations in transgenic tomato plant cells using confocal laser microscopy.(a) Leaf section of untransformed control plant, showing faint autofluorescence of cell wall (shown with arrow). (b) Co-localization of recombinant �1-PI targeted to ER inplants developed with pPAK vector, with ER-resident chaperone calreticulin; recombinant �1-PI (showing red fluorescence, panel 1) was immunodetected by Cy-3 labeledantibody while calreticulin (showing green fluorescence, panel 2) was immunodetected by FITC-conjugated antibody. (c) Recombinant �1-PI localized into apoplast in plantsdeveloped with pPWK vector. (d) Cytosolic localization of recombinant �1-PI in plants developed with pWSP vector. (e) Recombinant �1-PI localized into vacuoles in plantsd s staino eachs = 10 �
wslppnc
viTsosTsdpell
eveloped with pSWK vector, showing co-localization with acidic vacuolar proteinf recombinant �1-PI protein is shown with arrows in (b)–(e). The middle panel intructure, and the last panel shows the overlay image of first two images. Scale bar
ith cell-free extracts of transgenic plants showed a prominentingle band of ∼50.0 kDa in apoplast-targeted protein, and slightlyower molecular weight of ∼48.0 kDa in ER and vacuole targetedrotein, as compared to ∼52 kDa of purified human serum �1-PIrotein, while the expression of cytosol-targeted �1-PI protein wasearly undetectable. No such band was detected in untransformedontrol tomato plants (Fig. 4b).
Independent T0 transgenic plants developed with differentectors were self-pollinated and their seeds were analyzed fornheritance and segregation of transgene. T1 seeds of promising0 transgenic tomato plants were germinated on antibiotic-upplemented medium and investigated for segregation patternf nptII gene in T1 progeny. The results of chi-square (�2) analy-is showed typical 3:1 Mendelian segregation ratio except for few0 plants like PAK-25, SWK-8, and PWK-6. Constitutive expres-ion of heterologous recombinant �1-PI in transgenic tomato plantsid not show any morphological alterations and all the transgenic
lants were phenotypically normal, healthy and fertile, bearingqual number of fruits similar to control plants. The expressionevel of recombinant �1-PI was consistently improved and stabi-ized in T1 progeny with maximum average yield of recombinanted with Lyso-tracker dye (f), and shown as punctuate structures. The localization block is the DIC (differential interference contrast) image showing actual cellularm.
�1-PI to 3.05 ± 0.89% of TSP in T1 plants developed for ER reten-tion, followed by 1.89 ± 0.65% of TSP for vacuole, 1.40 ± 0.48% of TSPfor apoplast and 0.45 ± 0.08% of TSP for cytosol-targeted protein,respectively (data not shown). On the basis of segregation patternand performance of T0 plants, four promising stable T1 transgeniclines expressing higher levels of �1-PI, designated as PAK-31, SWK-1, PWK-26 and WSP-14 were selected and used for the purificationand further characterization of recombinant �1-PI.
3.5. Biological activity of plant expressed recombinant ˛1-PI
Biological activity of recombinant �1-PI in cell-free leaf extractsof transgenic tomato was determined for inhibition of elastaseenzyme. T0 population of transgenic plants developed with dif-ferent vectors like pPAK, pNAK, pPWK, pNWK, pSWK and pWSPshowed expression of biologically active recombinant �1-PI pro-tein with average values of 0.39 ± 0.17%, 0.34 ± 0.25%, 0.33 ± 0.22%,
0.30 ± 0.24%, 0.38 ± 0.23%, and 0.065 ± 0.04% of TSP respectively(Fig. 4a) which was improved to about 1.69 ± 0.46% for ER,0.63 ± 0.43% for apoplast and 0.72 ± 0.37% TSP for vacuolar forms inT1 progeny of the promising T0 plants, except for cytosolic form of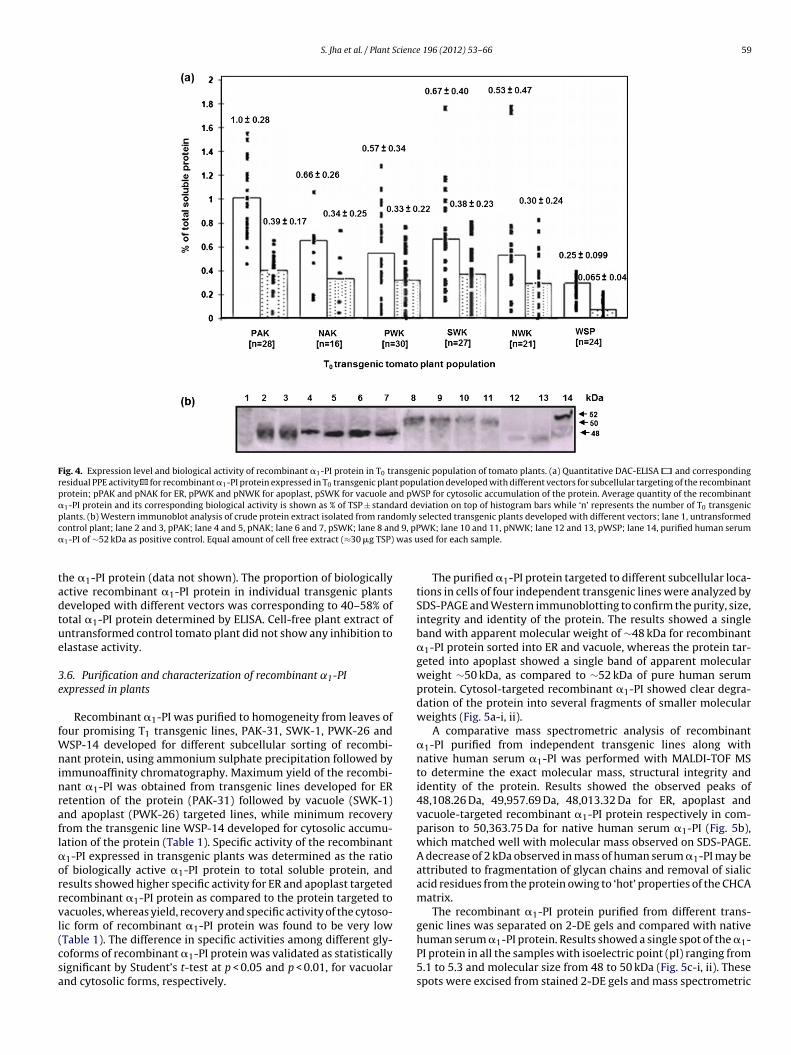
S. Jha et al. / Plant Science 196 (2012) 53– 66 59
Fig. 4. Expression level and biological activity of recombinant �1-PI protein in T0 transgenic population of tomato plants. (a) Quantitative DAC-ELISA and correspondingresidual PPE activity for recombinant �1-PI protein expressed in T0 transgenic plant population developed with different vectors for subcellular targeting of the recombinantprotein; pPAK and pNAK for ER, pPWK and pNWK for apoplast, pSWK for vacuole and pWSP for cytosolic accumulation of the protein. Average quantity of the recombinant�1-PI protein and its corresponding biological activity is shown as % of TSP ± standard deviation on top of histogram bars while ‘n’ represents the number of T0 transgenicplants. (b) Western immunoblot analysis of crude protein extract isolated from randomly selected transgenic plants developed with different vectors; lane 1, untransformedc d 9, pP� was u
tadtue
3e
fWninrafl�orrvl(csa
ontrol plant; lane 2 and 3, pPAK; lane 4 and 5, pNAK; lane 6 and 7, pSWK; lane 8 an1-PI of ∼52 kDa as positive control. Equal amount of cell free extract (≈30 �g TSP)
he �1-PI protein (data not shown). The proportion of biologicallyctive recombinant �1-PI protein in individual transgenic plantseveloped with different vectors was corresponding to 40–58% ofotal �1-PI protein determined by ELISA. Cell-free plant extract ofntransformed control tomato plant did not show any inhibition tolastase activity.
.6. Purification and characterization of recombinant ˛1-PIxpressed in plants
Recombinant �1-PI was purified to homogeneity from leaves ofour promising T1 transgenic lines, PAK-31, SWK-1, PWK-26 and
SP-14 developed for different subcellular sorting of recombi-ant protein, using ammonium sulphate precipitation followed by
mmunoaffinity chromatography. Maximum yield of the recombi-ant �1-PI was obtained from transgenic lines developed for ERetention of the protein (PAK-31) followed by vacuole (SWK-1)nd apoplast (PWK-26) targeted lines, while minimum recoveryrom the transgenic line WSP-14 developed for cytosolic accumu-ation of the protein (Table 1). Specific activity of the recombinant
1-PI expressed in transgenic plants was determined as the ratiof biologically active �1-PI protein to total soluble protein, andesults showed higher specific activity for ER and apoplast targetedecombinant �1-PI protein as compared to the protein targeted toacuoles, whereas yield, recovery and specific activity of the cytoso-ic form of recombinant �1-PI protein was found to be very low
Table 1). The difference in specific activities among different gly-oforms of recombinant �1-PI protein was validated as statisticallyignificant by Student’s t-test at p < 0.05 and p < 0.01, for vacuolarnd cytosolic forms, respectively.WK; lane 10 and 11, pNWK; lane 12 and 13, pWSP; lane 14, purified human serumsed for each sample.
The purified �1-PI protein targeted to different subcellular loca-tions in cells of four independent transgenic lines were analyzed bySDS-PAGE and Western immunoblotting to confirm the purity, size,integrity and identity of the protein. The results showed a singleband with apparent molecular weight of ∼48 kDa for recombinant�1-PI protein sorted into ER and vacuole, whereas the protein tar-geted into apoplast showed a single band of apparent molecularweight ∼50 kDa, as compared to ∼52 kDa of pure human serumprotein. Cytosol-targeted recombinant �1-PI showed clear degra-dation of the protein into several fragments of smaller molecularweights (Fig. 5a-i, ii).
A comparative mass spectrometric analysis of recombinant�1-PI purified from independent transgenic lines along withnative human serum �1-PI was performed with MALDI-TOF MSto determine the exact molecular mass, structural integrity andidentity of the protein. Results showed the observed peaks of48,108.26 Da, 49,957.69 Da, 48,013.32 Da for ER, apoplast andvacuole-targeted recombinant �1-PI protein respectively in com-parison to 50,363.75 Da for native human serum �1-PI (Fig. 5b),which matched well with molecular mass observed on SDS-PAGE.A decrease of 2 kDa observed in mass of human serum �1-PI may beattributed to fragmentation of glycan chains and removal of sialicacid residues from the protein owing to ‘hot’ properties of the CHCAmatrix.
The recombinant �1-PI protein purified from different trans-genic lines was separated on 2-DE gels and compared with native
human serum �1-PI protein. Results showed a single spot of the �1-PI protein in all the samples with isoelectric point (pI) ranging from5.1 to 5.3 and molecular size from 48 to 50 kDa (Fig. 5c-i, ii). Thesespots were excised from stained 2-DE gels and mass spectrometric
60 S. Jha et al. / Plant Science 196 (2012) 53– 66
Table 1Comparative expression profile and molecular characteristics of recombinant �1-PI targeted to different subcellular locations in transgenic tomato plants.a
Transgenic line Signal peptide Site foraccumulation
Molecular mass(Da)b
Nature ofglycosylationc
Protein content (mg)d Yield (%)h Specific activityi
Total solubleproteine
Recombinant�1-PIf
Biologicallyactive �1-PIg
PAK 31 PR1a/+KDEL ER 48,108.26 High mannose 2.93 2.61 ± 0.21 1.72 ± 0.12 17 0.59 ± 0.042SWK 1 SPS/−KDEL Vacuole 48,013.32 Complex
(Paucimannosidic)1.8 1.4 ± 0.11 0.74 ± 0.05 14 0.41 ± 0.028*
PWK 26 PR1a/−KDEL Apoplast 49,957.69 Complex 3.5 2.67 ± 0.18 1.99 ± 0.19 11 0.57 ± 0.056ns
WSP 14 −SP/−KDEL Cytosol Fragments ofdifferent masses
No glycosylation 3.1 0.14 ± 0.02 0.074 ± 0.013 2.58 0.024 ± 0.004**
a Purified from 100 g leaves of transgenic tomato plants.b Determined by MALDI-TOF MS in linear high-mass positive ion mode.c Analyzed by lectin-binding assays.d Determined after final purification step (immunoaffinity chromatography).e Estimated by Bradford assay.f Estimated by DAC-ELISA using anti human �1-PI antibody.g Estimated by residual PPE activity assay.h Recovery of recombinant �1-PI protein after final purification step, considering 100% yield in the crude extract, as determined by DAC-ELISA.i Calculated by taking ratio of biologically active �1-PI protein to total soluble protein and can be defined as the amount of �1-PI that inhibit PPE per mg of total protein
under standard assay conditions.nsnon-significant.
* p < 0.05.** p < 0.01.
Fig. 5. Purification and characterization of plant-expressed recombinant �1-PI protein. (a) Immunoaffinity purified recombinant �1-PI targeted into different subcellularlocations; (i) silver stained SDS-PAGE: lane 1, pre-stained molecular weight marker; lane 2, recombinant �1-PI targeted into cytosol; lane 3 and 4, vacuole; lane 5 and 6,apoplast; lane 7 and 8, ER; lane 9 and 10, pure human serum �1-PI protein as positive control; lane 11, untransformed plant as a negative control; (ii) Western immunoblot ofthe same gel developed with �1-PI specific antibody. Equal amount of the purified protein (100 ng) was used for each sample. (b) Determination of molecular masses of intactpurified recombinant �1-PI protein(s) by MALDI-TOF MS in linear mode; (i) human serum �1-PI, (ii) ER-targeted form, (iii) apoplast-targeted form, (iv) vacuole-targetedform of recombinant �1-PI. BSA was used as internal control. (c) Two-dimensional (2-DE) PAGE of differentially targeted purified recombinant �1-PI; (i) SYPRO-ruby stained2-DE gels: human serum �1-PI, ER, apoplast and vacuole-targeted �1-PI (from top to bottom); (ii) Western blot of the corresponding gels (i) developed with �1-PI specificantibody.

cience
aoms(d�vwir
auteeew
Eopm�o[hih(
3
pmstlemiyon
nbDawhpRttt�2sCwtop
S. Jha et al. / Plant S
nalysis of tryptic fragments generated by ‘in-gel’ trypsin digestionf recombinant �1-PI was performed by MALDI-TOF MS (peptideass fingerprinting) and MS/MS (peptide sequencing) which
howed identical peptide profile with that of native human �1-PISupplementary Fig. S2). The recombinant protein(s) purified fromifferent subcellular compartments was significantly identified as1-PI with a MOWSE score of 212, 91, 130 and 408 for ER, apoplast,acuole-targeted and native human protein respectively at p < 0.05ith high sequence coverage. These results suggested complete
ntact N-terminal sequences, pI and identity of plant-expressedecombinant �1-PI similar to serum �1-PI.
The ability of plant-purified recombinant �1-PI protein to bindnd form an irreversible stable complex with elastase was doc-mented by band-shift assay. Results showed a shift in mobilityo higher molecular weight on SDS-PAGE corresponding to thexpected covalent inhibitor-protease complex after incubation inquimolar ratio with PPE. Recombinant �1-PI was also cleaved bylastase and resulted into anticipated product of lower moleculareight in all the PPE treated samples (Fig. 6a).
Time course inhibition of elastase activity by human serum andR-targeted recombinant �1-PI showed complete inhibition (>99%)f enzyme activity within 10 min at equimolar ratio of inhibitor torotease (data not shown). Stoichiometry of inhibition was deter-ined by incubating the serum and plant-derived recombinant
1-PI protein in various molar ratios with constant concentrationf PPE. A plot of residual PPE activity versus �1-PI/PPE molar ratiosI/E] showed a straight line with linear regression (r2) of 0.897 foruman �1-PI and 0.917 for recombinant �1-PI at p < 0.01. The x-
ntercept resulted in the SI value, corresponding to 1.3 and 1.4 foruman serum �1-PI and recombinant �1-PI protein, respectivelyFig. 6b).
.7. Glycosylation analysis of plant-expressed recombinant ˛1-PI
The presence of glycan chains in recombinant �1-PI proteinurified from different transgenic lines was confirmed by enzy-atic deglycosylation with PNGase F, followed by SDS-PAGE which
howed significant shift in mobility of the recombinant protein(s)o lower molecular weight ∼45 kDa (Fig. 7a-i), that is nearly equiva-ent to theoretical mass of non-glycosylated �1-PI or Escherichia colixpressed �1-PI. The residual bands of higher molecular weightay be due to incomplete deglycosylation by PNGase F. The shift
n molecular mass was also confirmed by mass spectrometric anal-sis of deglycosylated �1-PI protein, showing observed mass peakf 45,011.49 Da, comparable to 44,328.35 Da for E. coli expressedon-glycosylated �1-PI (Fig. 7a-ii).
The glycosylation profile of the differentially targeted recombi-ant �1-PI protein(s) in transgenic tomato plants was determinedy its binding affinity to different types of lectins such as Con-A andSA agglutinin, which binds specifically to inner trimannosyl corend complex N-glycan chains, respectively. The recombinant �1-PIas retained on the Con-A column and the eluted protein showedigh immunoreactivity with anti-�1-PI antibodies, indicating theresence of glycan chains in the plant expressed �1-PI (Fig. 7b).esults of affinoblotting with biotin-labeled Con-A showed struc-ural heterogeneity in glycan chains of recombinant �1-PI targetedo different subcellular organelles (Fig. 7c). Affinoblotting of pro-ein samples with DIG-labeled DSA lectin showed that recombinant1-PI accumulated in ER did not bind to DSA lectin (Fig. 7d, lane), as compared to apoplast and vacuole-targeted �1-PI whichhowed strong affinity for the DSA lectin (Fig. 7d, lane 3 and 4).arboxypeptidase Y, which has high mannose type of N-glycans
as used as negative control (Fig. 7d, lane 6), whereas asialofe-uin and pure human serum �1-PI comprising of complex typesf N-glycosylation showed affinity to DSA lectin and served as theositive controls (Fig. 7d, lane 1 and 5).
196 (2012) 53– 66 61
The nature and type of glycan residues in the complex type ofN-glycan chains in different glycoforms of recombinant �1-PI wasfurther determined by immunoblotting with anti-xylose and anti-fucose antibodies that specifically cross-react against xylose andfucose residues at �-1,2 and �-1,3 positions of the glycan chains,respectively. Results of immunoblotting showed the presence ofthese residues only in vacuolar and apoplastic forms, and their com-plete absence in ER-targeted form of the recombinant �1-PI protein(Fig. 7e and f). Horseradish peroxidase (HRP), a 44 kDa model gly-coprotein consisting of both the plant specific glyco-epitopes wasused as the positive control whereas human serum �1-PI devoid ofboth of these residues served as the negative control (Fig. 7c and f).
3.8. Structural analysis and stability of recombinant ˛1-PI
The structural integrity and conformational stability of the ER-targeted from of recombinant �1-PI was determined by opticalspectroscopy and urea-induced equilibrium unfolding, respec-tively, and compared with human serum �1-PI. CD spectroscopicanalyses were performed in the far UV/amide region (200–250 nm)to characterize the secondary structure of the recombinant �1-PI protein. The CD spectra obtained for the recombinant �1-PIproteins were comparable to that of pure human serum �1-PI innative folded state in 0 M urea, and in completely unfolded statein 9 M urea (Fig. 8a), which confirmed the structural conformationof plant-derived recombinant �1-PI, similar to human �1-PI. TheCD spectra of native human �1-PI as well as of recombinant �1-PI showed the presence of substantial �-helical conformation inthe far-UV region, with prominent peaks at 222 nm and 208 nm.In order to assess the conformational stability of recombinant �1-PI protein, urea-induced unfolding was observed with changesin intrinsic tryptophan fluorescence intensity at 360 nm by fluo-rescence spectroscopy (Fig. 8c). The values for midpoints of theunfolding transition (Cm) of the recombinant �1-PI and humanserum �1-PI proteins in urea at 25 ◦C were 1.4 M and 4.7 M, respec-tively. Urea denaturation curves were used to determine the freeenergy of stabilization in the absence of denaturant, �G(H2O), bylinear extrapolation of the �G values to zero denaturant concen-tration. The values of �G(H2O) for recombinant and serum �1-PIprotein were 1.89 and 4.8 kcal mol−1, respectively. Far-UV CD anal-ysis of recombinant �1-PI demonstrated the impact of increasingurea concentrations on the CD ellipticity at 222 nm for both plant-derived and human �1-PI (Fig. 8b). The urea unfolding transitionsmonitored by the fluorescence signal were well fitted to the two-state unfolding model, whereas CD signal showed multi-phasicunfolding pattern for both the proteins which followed three-stateunfolding reaction with the formation of one stable intermediate.
Thermal stability of the ER-targeted form of the plant-derivedrecombinant �1-PI and its serum counterpart was analyzed at 54 ◦Cwhich showed that recombinant �1-PI had completely lost its bio-logical activity after 30 min of incubation at 54 ◦C with half-life ofheat denaturation of 11 min, whereas human serum �1-PI showedsignificantly enhanced resistance to thermal inactivation and lostonly 20% of its activity after 60 min incubation at 54 ◦C (Fig. 8d).
3.9. Pharmacokinetic evaluation of ˛1-PI protein
Purified ER targeted glycoform of recombinant �1-PI and serumpurified native �1-PI were administrated intravenously into inde-pendent sets of male Sprague-Dawley rats and the blood sampleswere collected initially at 15 min intervals up to 4 h, continued upto three days. We have neither observed any adverse affect on the
feeding regimen and behavioral response in the treated rats com-pared to non-administered control rats nor any allergic reaction ormortality was noticed in treated rats after 3 days of incubation. Theresults showed the plasma elimination half-life of plant expressed
62 S. Jha et al. / Plant Science 196 (2012) 53– 66
Fig. 6. Inhibitory activity of human and recombinant �1-PI protein towards elastase enzyme. (a) Band shift assay for native and recombinant �1-PI forming an inhibitorycomplex with PPE on SDS-PAGE; lane 1, native human serum �1-PI protein; lane 2, pure human �1-PI incubated with PPE at molar ratio [I]/[E] of 1:1; lane 3, 5 and 7,purified recombinant �1-PI targeted to ER, apoplast and vacuoles, respectively; lane 4, 6 and 8, corresponding recombinant �1-PI protein samples incubated with PPE atequal molar ratio; lane 9, pure PPE. Arrows indicate the �1-PI–PPE covalent complex, intact (native or recombinant) �1-PI and cleaved �1-PI, from top to bottom respectively.(b) Stoichiometry of inhibition of PPE activity by purified native human serum (—�—) and ER-targeted recombinant �1-PI (. . .�. . .), represented as plot of final remainingP 0 pmd
r��t�
4
taHtnm[itswmissfe
PE activities v/s [I]/[E] ratio. The concentration of elastase was held constant at 10etermined as SI.
ecombinant �1-PI to be 0.94 ± 0.52 h whereas the half-life of serum1-PI was 2.00 ± 0.68 h. Negligible concentration of recombinant1-PI was detected (<5%) in plasma after 3 h (180 min) of adminis-
ration, whereas about 40% of the protein content of native serum1-PI was retained in the rat plasma after 3 h incubation (Fig. 9).
. Discussion
Expression of recombinant therapeutic proteins in plant sys-em offers a safe, scalable and low-cost platform, besides thedditional advantage of post-translational modifications [13,25].owever, the expression level and stability of recombinant pro-
ein in plants are influenced by several factors like transgene copyumber, chromosomal location, mRNA stability, cis-regulatory ele-ents and final site of protein accumulation in plant cells or tissues
3]. We have achieved high-level expression of recombinant �1-PIn leaves of transgenic tomato plants with incorporation of optimalranslation initiation context (TAAACAATGG) and a 38 bp 5′-UTRequence of viral origin upstream of modified �1-PI gene alongith substitution of the native signal peptide of human �1-PI withodified signal sequences for sorting of the recombinant protein
nto different subcellular locations. Our results for expression of
table recombinant �1-PI protein in transgenic tomato are muchtraight forward and in accordance to earlier reports in plantsor over-expression of various recombinant proteins using differ-nt strategies [1,4,26]. The codon-optimization of the candidateol. Results were fitted with linear regression (p < 0.01); x-intercept of this line was
gene according to dicot-preferred codon usage, introduction ofvarious regulatory elements and subcellular targeting have beenanticipated to enhance transcription and translation efficiencies,yield and stability of the recombinant proteins in plant system[13,19].
The expression of recombinant �1-PI protein have been demon-strated in rice cell culture using Amy 3D sugar-regulated genecassette [27,28], in transgenic tobacco (transient expression)using chemically-inducible viral amplicon system [29,30] and intransplastomic tobacco [31]. However, yield, quality and glyco-sylation profile of the recombinant �1-PI was low and poor, thatposes severe limitations for its possible therapeutic application. Wedid not observe any microheterogeneity in the size of the recom-binant �1-PI protein expressed in transgenic tomato plants andonly one single band of ∼48 or 50 kDa for different glycoforms wasobtained, except for cytosolic form where more than one bandswere seen due to degradation. Moreover our results have shownthat depending on the site of accumulation of recombinant �1-PI intransgenic tomato leaf cells, the ratio of biologically active proteinfor elastase inhibition ranged between 40 and 58% of the total �1-PIprotein quantified by ELISA. This advocates the advantage of stabletransgenic plants for high-level expression of biologically active
human therapeutic proteins, including �1-PI, over the plant sus-pension culture or transient expression systems where extensiveoptimization of different parameters are required for cost effectiveproduction and quality of the protein [32,33].
S. Jha et al. / Plant Science 196 (2012) 53– 66 63
Fig. 7. Glycosylation analysis of recombinant �1-PI protein accumulated into different subcellular locations in transgenic tomato plants. (a) Enzymatic deglycosylation ofpurified recombinant �1-PI; (i) SDS-PAGE of purified �1-PI protein samples after deglycosylation by PNGase F. Lane 1, human serum �1-PI; lane 2, PNGase F treated human�1-PI; lane 3, 5 and 7, purified recombinant �1-PI targeted into ER, apoplast and vacuoles respectively; lane 4, 6 and 8, PNGase F treated corresponding �1-PI protein samples;lane 9, E. coli expressed unglycosylated �1-PI. Arrow indicates molecular weight of deglycosylated �1-PI (∼45 kDa); (ii) determination of molecular mass of deglycosylated�1-PI by MALDI-TOF/TOF in linear mode: enzymatically deglycosylated recombinant �1-PI (left panel) and E. coli expressed non-glycosylated �1-PI (right panel). (b) Westernimmunoblot of eluted recombinant �1-PI after Con-A affinity chromatography, developed with �1-PI specific antibody; lane 1, 2 and 3, purified recombinant �1-PI proteintargeted to ER, apoplast and vacuoles, respectively; lane 4, pure human serum �1-PI protein. (c) Affinoblotting of plant expressed recombinant �1-PI exhibiting cross-reactionwith biotin conjugated Con-A. Lane 1, non-glycosylated E. coli expressed �1-PI as a negative control; lane 2, 3 and 4, immunoaffinity purified recombinant �1-PI targetedto vacuoles, apoplast and ER, respectively; lane 5, pure human serum �1-PI protein. (d) Detection of type of N-glycans associated with recombinant �1-PI by affinoblottingwith DIG-labeled DSA lectin. Lane 1, asialofetuin as a positive control for glycoproteins containing complex-type N-glycans; lane 2, 3 and 4, purified recombinant �1-PI fromE arboxI ne 1,2l trol.
pdpttolcsiagsbtlibtrdTlwiad
R, apoplast and vacuoles, respectively; lane 5, pure human �1-PI protein; lane 6, cmmunoblotting with antibodies specific to �-1,2 xylose (e) and �-1,3 fucose (f); laane 4, HRP as positive control; lane 5, pure human serum �1-PI as the negative con
The sorting of foreign proteins into different subcellular com-artments of the plant cell have been shown to play a key role inetermining the nature of glycosylation, yield and stability of therotein owing to the different biochemical microenvironment ofhese compartments [34,35]. Considering the complexity of pro-ein maturation processes in plant cells and unpredictable naturef transgenic host plants, we have evaluated different subcellularocations for optimum yield, stability, glycosylation and biologi-al activity of the recombinant �1-PI protein. Immunolocalizationtudies have shown efficient sorting of the recombinant �1-PInto the respective subcellular destination, suggesting functionalitynd efficacy of modified signal peptide sequences in heterolo-ous environment of transgenic tomato plant cells. Our resultshowed the maximum accumulation and recovery of the recom-inant �1-PI from transgenic plants developed for ER retention ofhe protein, followed by vacuole and apoplast-targeted transgenicines. Although quantitative RT-PCR analyses showed almost sim-lar �1-PI transcript levels in the four groups of transgenic plantut the final yield of �1-PI protein in different subcellular loca-ions revealed significant differences. The average accumulation ofecombinant �1-PI protein in leaves of transgenic tomato plantseveloped with different vectors ranged between 1.5 and 3.2% ofSP except for cytosol targeted transgenic lines where significantow yield with apparent degradation of the recombinant protein
as evident. This may be attributed to negative redox potentialn cytosolic milieu, absence of correct folding and glycosylation,nd the existence of ubiquitin-proteasome pathway for proteolyticegradation. However, expression of heterologous proteins with
ypeptidase Y as a negative control containing high-mannose type N-glycans. (e, f) and 3, ER, vacuolar and apoplastic glycoforms of recombinant �1-PI respectively;
C-terminal purification tag or anchorage into the cytosolic face ofER in transgenic tobacco plants have been documented as promis-ing strategies for cytosolic accumulation of recombinant proteins[16,36].
The relative higher yield of recombinant �1-PI up to ∼3.29% ofTSP in T1 transgenic tomato plants developed with pPAK vectorfor ER retention may be due to several factors acting in synergywith codon-optimized gene along with KDEL as ER retention sig-nal [12,35]. Targeting of foreign proteins with C-terminal KDEL orHDEL tetrapeptide has been demonstrated for higher accumulationin plants cells [37,38]. The ER lumen seems to provide protec-tive oxidizing environment and molecular chaperones for correctprotein folding, low abundance of proteolytic enzymes and largespace for protein accumulation. Despite several advantages withER accumulation, it is not always considered a suitable destina-tion for all the foreign proteins because a number of therapeuticproteins require late post-translational modifications like complexglycosylation for stability and biological activity which occur down-stream of the ER along the secretory pathway [26], whereas certainforeign proteins have shown structural heterogeneity due to prote-olysis by ER-resident proteases [39]. Protein targeting to apoplastmay help to prevent unintended proteolytic processing, correctmaturation and glycosylation of the recombinant protein [6,40].Recombinant �1-PI targeted to apoplast has revealed considerable
yield and specific activity. Likewise, substitution of native signalpeptide with sporamine signal sequence has shown accumulationof recombinant �1-PI into vacuoles with comparable yield as for ER-retension, but with lower specific activity, that may be due to high
64 S. Jha et al. / Plant Science 196 (2012) 53– 66
Fig. 8. Structural conformation and stability of ER-targeted recombinant �1-PI and human serum �1-PI for urea denaturation and heat treatment. (a) Characterization ofsecondary structure of human serum �1-PI (—�—) and recombinant �1-PI (—♦—) by circular dichroism. CD spectra of the native folded protein at 0 M urea (solid line) andcompletely unfolded protein at 9 M urea concentration (dotted line) obtained by continuous wavelength scans from 200 to 250 nm, at a scan speed of 20 nm/min using0.1 cm path length cell. Protein concentration used was 3.0 �M and averages of two scans/sample were calculated and plotted as ellipticity (�, mdeg) v/s wavelength (�, nm).(b) Urea-induced equilibrium unfolding transitions of human serum �1-PI (. . .�. . .) and recombinant �1-PI (. . .♦. . .), as monitored by changes in CD ellipticity at 222 nmobtained from the far-UV CD curves at increasing concentration of urea (0–9 M). (c) Increase in tryptophan fluorescence emission intensity at 360 nm (�ex = 280 nm, excitationand emission slit widths = 5 nm) of human serum (. . .�. . .) and recombinant protein (. . .♦. . .). The protein concentration was 3.0 and 4.0 �M for (b) and (c), respectively.The data are represented as the relative percentage of ellipticity or fluorescence, taking the value observed for native protein as 100%. (d) Thermal stability of human serum� ining
c ental d
pl
dPtfpcttchpwteepa
1-PI (—�—) and ER-targeted recombinant �1-PI (—♦—) at 54 ◦C measured as remaoncentration for each sample was 5 �g ml−1. Lines were drawn by fitting experim
roteolytic content and acidic environment of lytic vacuoles ineaves, in contrast to protein storage vacuoles in seeds [41].
Our results from band-shift assay and SI experiments clearlyemonstrated the formation of stable complex of recombinant �1-I with the specific substrate elastase in equimolar ratio, similar tohat of plasma-derived �1-PI. These results strongly suggest correctolding, retention of inhibitory properties and the binding affinity oflant expressed recombinant �1-PI to elastase [42]. The CD spectraonfirmed the secondary structure and conformational integrity ofhe recombinant �1-PI identical to that of serum �1-PI, however,he recombinant protein showed relatively less thermostability inomparison to its human counterpart. The pharmacokinetic studiesave also shown faster blood clearance of recombinant �1-PI com-ared to human serum derived native protein in rat model systemhich may be attributed to association of sialic acid to the glycan
ermini in serum derived glycosylated �1-PI in comparison to plant
xpressed glycosylated recombinant �1-PI which lacks this moi-ty [43,44]. In several earlier reports, removal of sialic acid fromlasma-derived �1-PI has shown significant reduction in half-lifend rapid clearance from animal blood [45,46].inhibitory activity against elastase at different time intervals up to 60 min. Proteinata to a single exponential decay.
The purification of recombinant �1-PI from transgenic rice cellcultures involved a series of complex chromatography steps result-ing into significant loss of yield and increased cost [28]. In thepresent study, recombinant �1-PI has been purified to homogene-ity through immunoaffinity chromatography in a single step. Thepurified protein showed the biological activity and other biochem-ical properties similar to the human serum �1-PI, suggesting thatno physicochemical changes have occurred during the purifica-tion process. The plant purified protein showed the molecularmass ∼48–50 kDa as compared to ∼45 kDa for non-glycosylated�1-PI expressed in E. coli or chloroplast [23,31], suggesting theassociation of glycan chains with plant derived protein. The smallvariation in the molecular mass of plant expressed protein withits serum counterpart (∼52 kDa) may be attributed to differentialglycosylation particularly for terminal galactosylation and sialyla-tion [35,43]. The N-glycosylation of plant-expressed recombinant
�1-PI was determined by enzymatic deglycosylation and lectin orantibody binding assays. ER-targeted recombinant �1-PI showedhigh-mannose type of glycosylation; however, the N-glycansborne by vacuolar and apoplastic forms are of the complex-type,
S. Jha et al. / Plant Science
Fig. 9. Pharmacokinetics of native human serum �1-PI and plant derived ER-targeted recombinant �1-PI in rat model system. Graphical representation of theresidual concentration of native human �1-PI (-�-) and recombinant �1-PI (-�-) inta
awaedadbpaccmTrBwwitnicaMst
iirKriaiiso
[
[
[
[
[
[
[
[
[
[
[20] J. Sambrook, D.W. Russell, Molecular Cloning A Laboratory Manual, Cold Spring
he serum of rats at different time intervals. The �1-PI concentration after 5 min ofdministration was set as 100% �1-PI plasma concentration.
ssociated with �(1,2)-linked xylose and �(1,3)-linked fucosehich are typical characteristic of the plant N-glycans and are
bsent in mammals. These residues are constituents of glyco-pitopes, known to be important IgE binding carbohydrateeterminants of plant allergens [47] which may expose humanllergic patients to severe hypersensitivity reaction, as recentlyescribed for a chimeric mouse–human IgG1 monoclonal anti-ody used for colorectal cancer therapy [48]. However, severallant-derived glycoproteins have undergone clinical trials withoutdverse immunological effects [49]; moreover these glyco-epitopesan also elicit IgG production in humans, causing rapid immunelearance of glycoprotein from the blood circulation [50], whichay greatly compromise their effectiveness as therapeutic agents.
riguero et al. [51] have recently shown a faster serum clearanceate for plant derived monoclonal antibody against the hepatitis
surface antigen compared with the analogous murine antibody,hereas different glycoforms of plant derived antibody with andithout ER retention signal showed indistinguishable in vivo stabil-
ty. It would be interesting to compare the half-life of apoplastic andhe ER-derived glycoforms of the recombinant human �1-PI in theear future. The vacuolar targeting of recombinant �1-PI resulted
nto cleavage of glycan chains due to the presence of several gly-osidases and hydrolytic enzymes in vacuolar lumen of leaf cells,nd showed paucimannosidic type of N-glycans, consisting only thean3GlcNAc2 core (Man: mannose; GlcNAc: N-acetylglucosamine)
ubstituted with �-1,3 fucose and �-1,2 xylose which may arise dueo the removal of terminal GlcNAc residues in post-Golgi passage.
In summary, our results suggest significance of protein sort-ng into different subcellular organelles of plant cells, particularlyn leaf tissues and their impact on final yield and stability ofecombinant protein. Our results have clearly shown that theDEL-mediated ER retention of the recombinant �1-PI proteineflect several advantages including improved yield, specific activ-ty, structural integrity of the protein and prevention for theddition of immunogenic N-glycan residues. We are now attempt-ng to increase the stability of ER-targeted recombinant � -PI by
1ntroducing site-specific mutations in the molecule [23]. Futuretudies will be required to improve in vivo stability and half-lifef plant-expressed �1-PI protein via engineering the pathway for[
196 (2012) 53– 66 65
humanized glycosylation including sialylation [52] and silencing ofspecific xylosyl/fucosyltransferase [53,54].
Acknowledgements
We are grateful to Director, CSIR-NBRI, Lucknow for providinginfrastructural support, and Dr. Vinod Bhakuni, CSIR-CDRI, Luc-know for providing facility for optical spectroscopy. We thankfullyacknowledge Council of Scientific and Industrial Research (C.S.I.R.),New Delhi, India for providing funds and senior research fellow-ships to SJ and SA. This work was carried out under the CSIR-NBRIIn-house Project OLP 0031.
Appendix A. Supplementary data
Supplementary data associated with this article can befound, in the online version, at http://dx.doi.org/10.1016/j.plantsci.2012.07.004.
References
[1] S. Ma, Y. Huang, A. Davis, Z. Yin, Q. Mi, R. Menassa, J.E. Brandle, A.M. Jevnikar,Production of biologically active human interleukin-4 in transgenic tobaccoand potato, Plant Biotechnol. J. 3 (2005) 309–318.
[2] E. Stoger, J.K. Ma, R. Fischer, P. Christou, Sowing the seeds of success: pharma-ceutical proteins from plants, Curr. Opin. Biotechnol. 16 (2005) 167–173.
[3] P.M. Doran, Foreign protein degradation and instability in plants and tissuecultures, Trends Biotechnol. 24 (2006) 426–432.
[4] S.J. Streatfield, Approaches to achieve high-level heterologous protein produc-tion in plants, Plant Biotechnol. J. 5 (2007) 2–15.
[5] M. Benchabane, C. Saint-Jore Dupas, M. Bardor, L. Faye, D. Michaud, V. Gomord,Targeting and post-translational processing of human �1-antichymotrypsin inBY-2 tobacco cultured cells, Plant Biotechnol. J. 7 (2009) 146–160.
[6] M. Benchabane, C. Goulet, D. Rivard, L. Faye, V. Gomord, D. Michaud, Prevent-ing unintended proteolysis in plant protein biofactories, Plant Biotechnol. J. 6(2008) 633–648.
[7] Y. Wu, R.C. Foreman, The molecular genetics of alpha-1-antitrypsin deficiency,Bioessays 13 (1991) 163–169.
[8] D.A. Lomas, Molecular mousetraps, alpha-1-antitrypsin deficiency and the ser-pinopathies, Clin. Med. 5 (2005) 249–257.
[9] G.A. Heresi, J.K. Stoller, Augmentation therapy in alpha-1 antitrypsin deficiency,Expert Opin. Biol. Ther. 8 (2008) 515–526.
10] E. Karnaukhova, Y. Ophir, B. Golding, Recombinant human alpha-1 proteinaseinhibitor: towards therapeutic use, Amino Acids 30 (2006) 317–332.
11] S. Petruccelli, M.S. Otegui, F. Lareu, O.T. Dinh, A.C. Fitchette, A. Circosta, AKDEL-tagged monoclonal antibody is efficiently retained in the endoplasmicreticulum in leaves, but is both partially secreted and stored to protein storagevacuoles in seed, Plant Biotechnol. J. 4 (2006) 511–527.
12] R. Strasser, J. Stadlmann, M. Schähs, G. Stiegler, H. Quendler, L. Mach, J. Glössl, K.Weterings, M. Pabst, H. Steinkellner, Generation of glyco-engineered Nicotianabenthamiana for the production of monoclonal antibodies with a homogeneoushuman-like N-glycan structure, Plant Biotechnol. J. 6 (2008) 392–402.
13] S.R. Karg, P.T. Kallio, The production of biopharmaceuticals in plant systems,Biotechnol. Adv. 27 (2009) 879–894.
14] C.I. Wandelt, M.R.I. Khan, S. Craig, H.E. Schroeder, D. Spencer, T.J.V. Higgins,Vicilin with carboxy-terminal KDEL is retained in the endoplasmic reticulumand accumulates to high levels in the leaves of transgenic plants, Plant J. 2(1992) 181–192.
15] A. Vitale, E. Pedrazzini, Recombinant pharmaceuticals from plants: the plantendomembrane system as bioreactor, Mol. Interventions 5 (2005) 216–225.
16] C. Marusic, J. Nuttall, G. Buriani, C. Lico, R. Lombardi, S. Baschieri, E. Benvenuto, L.Frigerio, Expression, intracellular targeting and purification of HIV Nef variantsin tobacco cells, BMC Biotechnol. 7 (2007) 1–12.
17] S. Agarwal, R. Singh, I. Sanyal, D.V. Amla, Expression of modified gene encod-ing functional human alpha-1-antitrypsin protein in transgenic tomato plants,Transgenic Res. 17 (2008) 881–896.
18] S. McCormick, J. Niedermeyer, A. Branson, H. Horsch, R. Fraley, Leaf disc trans-formation of cultivated tomato (L. esculentum) using Agrobacterium tumefaciens,Plant Cell Rep. 5 (1986) 81–84.
19] S. Agarwal, S. Jha, I. Sanyal, D.V. Amla, Effect of point mutations in transla-tion initiation context on the expression of recombinant human �1-proteinaseinhibitor in transgenic tomato plants, Plant Cell Rep. 28 (2009) 1791–1798.
Harbor Laboratory Press, Cold Spring Harbor, New York, 2001.21] A. Görg, W. Postel, J. Weser, S. Günther, J.R. Strahler, S.M. Hanash, Elimina-
tion of point streaking on silver-stained two-dimensional gels by addition ofiodoacetamide to the equilibration buffer, Electrophoresis 8 (1987) 122–124.

6 cienc
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
6 S. Jha et al. / Plant S
22] A. Shevchenko, H. Tomas, J. Havlis, J.V. Olsen, M. Mann, In-gel digestion formass spectrometric characterization of proteins and proteomes, Nat. Protoc. 1(2006) 2856–2860.
23] S. Agarwal, S. Jha, I. Sanyal, D.V. Amla, Expression and purification of recom-binant human �1-proteinase inhibitor and its single amino acid substitutedvariants in Escherichia coli for enhanced stability and biological activity, J.Biotechnol. 147 (2010) 64–72.
24] C.N. Pace, Determination and analysis of urea and guanidine hydrochloridedenaturation curves, Methods Enzymol. 131 (1986) 266–280.
25] J.K. Ma, P.M.W. Drake, P. Christou, The production of recombinant pharmaceu-tical proteins in plants, Nat. Rev. Genet. 4 (2003) 794–805.
26] L. Faye, A. Boulaflous, M. Benchabane, V. Gomord, D. Michaud, Protein modifica-tions in the plant secretory pathway: current status and practical implicationsin molecular pharming, Vaccine 23 (2005) 1770–1778.
27] M. Terashima, Y. Murai, M. Kawanura, S. Nakanishi, T. Stoltz, L. Chen, W. Drohan,R.L. Rodriguez, S. Katoh, Production of functional human �-1-antitrypsin byplant cell culture, Appl. Microbiol. Biotechnol. 52 (1999) 516–523.
28] J. Huang, T.D. Sutliff, L. Wu, S. Nandi, K. Benge, M. Terashima, A.H. Ralston,W. Drohan, N. Huang, R.L. Rodriguez, Expression and purification of functionalhuman �-1-antitrypsin from cultured plant cells, Biotechnol. Prog. 17 (2001)126–133.
29] M.R. Sudarshana, M.A. Plesha, S.L. Uratsu, B.W. Falk, A.M. Dandekar, T.-K. Huang,K.A. McDonald, A chemically inducible cucumber mosaic virus amplicon systemfor expression of heterologous proteins in plant tissues, Plant Biotechnol. J. 4(2006) 551–559.
30] T.-K. Huang, M.A. Plesha, K.A. McDonald, Semicontinuous bioreactor produc-tion of a recombinant human therapeutic protein using a chemically inducibleviral amplicon expression system in transgenic plant cell suspension cultures,Biotechnol. Bioeng. 106 (2010) 408–421.
31] M. Nadai, J. Bally, M. Vitel, C. Job, G. Tissot, J. Botterman, M. Dubald, High-level expression of active human alpha-1-antitrypsin in transgenic tobaccochloroplasts, Transgenic Res. 18 (2009) 173–178.
32] T.-K. Huang, K.A. McDonald, Bioreactor systems for in vitro production of foreignproteins using plant cell cultures, Biotechnol. Adv. 30 (2012) 398–409.
33] S.M.-H. Shih, P.M. Doran, Foreign protein production using plant cell and organcultures: advantages and limitations, Biotechnol. Adv. 27 (2009) 1036–1042.
34] N.S. Outchkourov, B. Rogelj, B. Strukelj, M.A. Jongsma, Expression of seaanemone equistatin in potato: effect of plant proteases on heterologous proteinproduction, Plant Physiol. 133 (2003) 379–390.
35] V. Gomord, C. Sourrouille, A.-C. Fitchette, M. Bardor, S. Pagny, P. Lerouge, L. Faye,Production and glycosylation of plant-made pharmaceuticals: the antibodiesas a challenge, Plant Biotechnol. J. 2 (2004) 83–100.
36] A. Barbante, S. Irons, C. Hawes, L. Frigerio, A. Vitale, E. Pedrazzini, Anchorage tothe cytosolic face of the endoplasmic reticulum membrane: a new strategy tostabilize a cytosolic recombinant antigen in plants, Plant Biotechnol. J. 6 (2008)560–575.
37] A. Schouten, J. Roosien, F.A. van Engelen, G.A. de Jong, A.W. Borst-Vrenssen,J.F. Zilverentant, D. Bosch, W.J. Stiekema, F.J. Gommers, A. Schots, J. Bakker,The C-terminal KDEL sequence increases the expression level of a single-chain
antibody designed to be targeted to both the cytosol and the secretory pathwayin transgenic tobacco, Plant Mol. Biol. 30 (1996) 781–793.38] L. Frigerio, A. Pastres, A. Prada, A. Vitale, Influence of KDEL on the fate of trimericor assembly-defective phaseolin: selective use of an alternative route to vac-uoles, Plant Cell 13 (2001) 1109–1126.
[
e 196 (2012) 53– 66
39] A. Schmitz, V. Herzog, Endoplasmic reticulum-associated degradation: excep-tions to the rule, Eur. J. Cell Biol. 83 (2004) 501–509.
40] M. Gils, R. Kandzia, S. Marillonnet, V. Klimyuk, Y. Gleba, High-yield produc-tion of authentic human growth hormone using a plant virus-based expressionsystem, Plant Biotechnol. J. 3 (2005) 613–620.
41] J. Yang, L.A. Barr, S.R. Fahnestock, Z.B. Liu, High yield recombinant silk-likeprotein production in transgenic plants through protein targeting, TransgenicRes. 14 (2005) 313–324.
42] J. Travis, M. Owen, P. George, R. Carrell, S. Rosenberg, R.A. Hallewell, P.J. Barr,Isolation and properties of recombinant DNA produced variants of human �-l-proteinase inhibitor, J. Biol. Chem. 260 (1985) 4384–4389.
43] V. Gomord, A.C. Fitchette, L. Menu-Bouaouiche, C. Saint-Jore Dupas, C. Plas-son, D. Michaud, L. Faye, Plant-specific glycosylation patterns in the context oftherapeutic protein production, Plant Biotechnol. J. 8 (2010) 564–587.
44] C. S-J-Dupas, L. Faye, V. Gomord, From planta to pharma with glycosylation inthe toolbox, Trends Biotechnol. 25 (2007) 317–323.
45] S.-D. Yu, J.C. Gan, The role of sialic acid and galactose residues in determiningthe survival of human plasma �1-antitrypsin in the blood circulation, Arch.Biochem. Biophys. 179 (1977) 477–485.
46] S.-D. Yu, J.C. Gan, Effects of progressive desialylation on the survival of humanplasma �1-antitrypsin in rat circulation, Int. J. Biochem. 9 (1978) 107–115.
47] R. Van Ree, M. Cabanes-Macheteau, J. Akkerdaas, J.P. Milazzo, C. Loutelier-Bourhis, C. Rayon, M. Villalba, S. Koppelman, R. Aalberse, R. Rodriguez, L.Faye, P. Lerouge, �(1,2)-xylose and �(1,3)-fucose residues have a strong con-tribution in IgE binding to plant glycoallergens, J. Biol. Chem. 275 (2000)11451–11458.
48] C.H. Chung, B. Mirakhur, E. Chan, Q.T. Le, J. Berlin, M. Morse, B.A. Murphy, S.M.Satinover, J. Hosen, D. Mauro, R.J. Slebos, Q. Zhou, D. Gold, T. Hatley, D.J. Hicklin,T.A. Platts-Mills, Cetuximab-induced anaphylaxis and IgE specific for galactose-alpha-1,3-galactose, N. Engl. J. Med. 358 (2008) 1109–1117.
49] Y. Shaaltiel, D. Bartfeld, S. Hashmueli, G. Baum, E. Brill-Almon, G. Galili, O.Dym, S.A. Boldin-Adamsky, I. Silman, J.L. Sussman, A.H. Futerman, D. Aviezer,Production of glucocerebrosidase with terminal mannose glycans for enzymereplacement therapy of Gaucher’s disease using a plant cell system, PlantBiotechnol. J. 5 (2007) 579–590.
50] M. Bardor, C. Faveeuw, A.-C. Fitchette, D. Gilbert, L. Galas, F. Trottein, L. Faye,P. Lerouge, Immunoreactivity in mammals of two typical plant glyco-epitopes,core alpha(1,3)-fucose and core xylose, Glycobiology 13 (2003) 427–434.
51] A. Triguero, G. Cabrera, M. Rodriguez, J. Soto, Y. Zamora, M. Perez, M.R. Wormald,J.A. Cremata, Differential N-glycosylation of a monoclonal antibody expressedin tobacco leaves with and without endoplasmic reticulum retention signalapparently induces similar in vivo stability in mice, Plant Biotechnol. J. 9 (2011)1120–1130.
52] A. Castilho, R. Strasser, J. Stadlmann, J. Grass, J. Jez, P. Gattinger, R. Kunert, H.Quendler, M. Pabst, R. Leonard, F. Altmann, H. Steinkellner, In planta proteinsialylation through overexpression of the respective mammalian pathway, J.Biol. Chem. 285 (2010) 15923–15930.
53] K.M. Cox, J.D. Sterling, J.T. Regan, J.R. Gasdaska, K.K. Frantz, C.G. Peele, A. Black,D. Passmore, C. Moldovan-Loomis, M. Srinivasan, S. Cuison, P.M. Cardarelli, L.F.
Dickey, Glycan optimization of a human monoclonal antibody in the aquaticplant Lemna minor, Nat. Biotechnol. 24 (2006) 1591–1597.54] K. Matsuo, T. Matsumura, Deletion of fucose residues in plant N-glycans byrepression of the GDP-mannose 4,6-dehydratase gene using virus-inducedgene silencing and RNA interference, Plant Biotechnol. J. (2010) 1–18.
Copyright © 2022 FDOKUMEN




















