
Differential effects of dopamine D2 and D3 receptor antagonists in regard to dopamine release, in...
17
J Neural Transm [GenSect] (1994) 98:39-55 Journal of Neural Transmission Springer-Verlag 1994 Printed in Austria Differential effects of dopamine D2 and D3 receptor antagonists in regard to dopamine release, in vivo receptor displacement and behaviour N. Waters 1, L. Liifberg 1, S. Haadsma-Svensson 1'2,K. Svensson ~, C. Sonesson 1, and A. Carlsson ~ 1 Department of Pharmacology, University of G6teborg, G6teborg, Sweden 2Upjohn Laboratories, Medicinal Chemistry Research, The Upjohn Company, Kalamazoo, MI, USA Accepted May 25, 1994 Summary. To establish possible functional differences between the dopamine D2 and D3 receptor we investigated the relation between the ability, for a set of nine mixed dopamine D2 and D3 receptor antagonists, to displace N, N- dipropyl-2-amino-5,6-dihydroxy tetralin (DP-5,6-ADTN) from striatal binding sites and the subsequent behavioural consequences in vivo. Dopamine D2 re- ceptor preferring antagonists are powerful displacers of DP-5,6-ADTN from the striatum. Maximal displacement is followed by strong hypomotility. Dis- placement of the agonist by the D3 preferring antagonist U99194A is only partial and results in synergistic increases in locomotor activity. Superimposing haloperidol upon GBR12909 leads to a synergistic increase in striatal dialysate dopamine concentrations. This effect is absent when combining GBR12909 with the putative D3 antagonist U99194A. These data give support for the hypothesis that the dopamine D3 receptor is functionally relevant at the post- synaptic level. Here, in contrast to the D2 receptor, it is proposed to exert an inhibitory influence on psychomotor functions. Keywords: Dopamine, D2, D3, receptors, antagonists, displacement, release Introduction The variety of effects of dopamine receptor antagonists have been explained by combination of two generally accepted hypotheses of receptor subdivision. One hypothesis dividing the dopamine receptors into D 1 and D2 receptors (for review see Kebabian and Calne, 1979). The other dividing dopamine receptors into presynaptic autoreceptors and postsynaptic receptors respectively (Carls- son, 1975). The presynaptic autoreceptors are thought to be of the D2 subclass
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Differential effects of dopamine D2 and D3 receptor antagonists in regard to dopamine release, in...

J Neural Transm [GenSect] (1994) 98:39-55 Journal of
Neural Transmission
�9 Springer-Verlag 1994 Printed in Austria
Differential effects of dopamine D2 and D3 receptor antagonists in regard to dopamine release, in vivo receptor displacement
and behaviour
N. Waters 1, L. Liifberg 1, S. Haadsma-Svensson 1'2, K. Svensson ~, C. Sonesson 1, and A. Carlsson ~
1 Department of Pharmacology, University of G6teborg, G6teborg, Sweden 2 Upjohn Laboratories, Medicinal Chemistry Research, The Upjohn Company,
Kalamazoo, MI, USA
Accepted May 25, 1994
Summary. To establish possible functional differences between the dopamine D2 and D3 receptor we investigated the relation between the ability, for a set of nine mixed dopamine D2 and D3 receptor antagonists, to displace N, N- dipropyl-2-amino-5,6-dihydroxy tetralin (DP-5,6-ADTN) from striatal binding sites and the subsequent behavioural consequences in vivo. Dopamine D2 re- ceptor preferring antagonists are powerful displacers of DP-5,6-ADTN from the striatum. Maximal displacement is followed by strong hypomotility. Dis- placement of the agonist by the D3 preferring antagonist U99194A is only partial and results in synergistic increases in locomotor activity. Superimposing haloperidol upon GBR12909 leads to a synergistic increase in striatal dialysate dopamine concentrations. This effect is absent when combining GBR12909 with the putative D3 antagonist U99194A. These data give support for the hypothesis that the dopamine D3 receptor is functionally relevant at the post- synaptic level. Here, in contrast to the D2 receptor, it is proposed to exert an inhibitory influence on psychomotor functions.
Keywords: Dopamine, D2, D3, receptors, antagonists, displacement, release
Introduction
The variety of effects of dopamine receptor antagonists have been explained by combination of two generally accepted hypotheses of receptor subdivision. One hypothesis dividing the dopamine receptors into D 1 and D2 receptors (for review see Kebabian and Calne, 1979). The other dividing dopamine receptors into presynaptic autoreceptors and postsynaptic receptors respectively (Carls- son, 1975). The presynaptic autoreceptors are thought to be of the D2 subclass

40 N. Waters et al.
(Di Chiara etal., 1977) while both D1 and D2 receptors are present at the postsynaptic level. The clinically observed effects of antipsychotic drugs are believed to be dependent on their ability to displace dopamine from dopamine receptors (Carlssson, 1978). However, it is difficult to explain all the observed behavioural and biochemical effects of dopamine D2 receptor antagonists based on one D2 receptor, although located at different anatomical positions. In accordance more complicated hypotheses of dopamine D2 receptor subdivision have been put forward (Cools and Van Rossum, 1976, 1980; Ljungberg and Ungerstedt, 1978; Seeman, 1980) but these are not as widely accepted nor as easily tested as the postsynaptic D1 and D2 dicotomy, or the D2 pre- and postsynaptic, hypotheses. More recently, the introduction of molecular biolog- ical techniques has shown that the dopamine receptors are a more heterogeneous group of receptors than previously anticipated. At present dopamine receptors are considered to consist of two groups, the "D1 family" (D1, D5 or by another nomenclature Dla Dlb) and the "D2 family" (D2, D3, D4) (for reviews see Civelli et al., 1991; Sibley and Monsma Jr., 1992). These findings have prompted the search for receptor subtype selective ligands and for functional correlations to these subtypes.
Much recent interest has focused on the dopamine D3 receptor (Sokoloff et al., 1990). The D3 receptor differs in its in vitro binding properties and regional distribution from the D1 and D2 receptors. Dopamine D3 mRNA has been found to be expressed predominantly in limbic dopaminergic projection areas associated with cognitive, emotional and endocrine functions but also in the substantia nigra and the ventral tegmental area (Bouthenet et al., 1991), although the occurrence of D3 mRNA and translation product both in human and rat substantia nigra is debated (Landwehrmeyer etal., 1993 a, b). More recently the existence of D3 mRNA colocalized with D2 and D 1 mRNA in striatal cells have also been reported (Surmeier etal., 1992, 1993), although the relation between mRNA and the expression of protein is unclear. Taken together, these studies seem to agree in that the D3 receptor is mainly located in dopamine projection areas in the ventral striatum. These observations have led to spec- ulations about its involvement in various psychiatric disorders (Snyder, 1990; Strange, 1991; Sokoloff etal., 1992; Schwartz etal., 1993).
Although the distribution of the D3 receptors in the rat brain has been extensively studied, little data suggesting possible functional roles for this re- ceptor subtype has emerged. However, the dopamine receptor antagonists ( + ) - UH232, ( + ) - AJ76 (Svensson et al., 1986) and ( - ) - DS121 (Sonesson et al., 1993) have all 3-5 times higher affinity for the D3 than the D2 receptor in in vitro binding studies (Sokoloff et al., 1990; Sonesson etal., 1993). Unlike classical neuroleptics, these three compounds produce a dopamine dependent increase in locomotor activity and, like the classical neuroleptics, they induce increases in the presynaptic release of dopamine (Waters et al., 1993 a; Sonesson et al., 1993). These data suggest that preferential antagonism of the D3 receptor may lead to behavioural activation in the rat and that the D3 receptor could

Differential effects of dopamine D2 and D3 receptor antagonists 41
be involved in the control of rat psychomotor acitivity. More recently, the finding that the 20-fold D3 over D2 preferring putative antagonist U99194A increases locomotor activity without any concomitant effects on dopamine release has led to the hypothesis that the D3 receptor is mainly functionally relevant at the postsynaptic level. Here, in contrast to the D2 receptor, it would then serve an inhibitory role on psychomotor functions (Waters etal., 1993 b).
The aim of the present study was to further investigate the possible functional role of the D3 receptor and to test the hypothesis cited above. To do this we evaluated the relation between the ability, for a set of dopamine D2 and D3 antagonists, to displace N,N-dipropyl-2-amino-5,6-dihydroxy tetralin (DP-5,6- ADTN) from striatal binding sites (Feenstra etal., 1983a, b; Carlsson and L6fberg, 1985) and the subsequent behavioural consequences. Furthermore we employed brain microdialysis (Zetterstr6m et al., 1983; Westerink et al., 1987) to monitor presynaptic effects on dopamine release of the different D2 and D3 antagonists. We used a series of dopamine receptor antagonists ranging from the more D2 preferring ligands haloperidol, raclopride and sulpiride (Sokoloff etal., 1990) through ( + ) - U H 2 3 2 , ( + ) - A J 7 6 and ( - ) -DS121 (Sonesson etal., 1993) which have a certain D3 receptor preference, to the 20-fold D3 receptor preferring compound U99194A (Waters etal., 1993 b).
Materials and methods
Subjects
Animals used in the present study were male rats (200-250 g) of the Sprague-Dawley strain (Beekay, Sollentuna, Sweden). The animals were kept five per cage under controlled en- vironmental conditions (temperature 21-22 ~ and humidity 55-65%, 5 a.m.-7 p.m./7 p.m.- 5 a.m. light/dark cycle) and were allowed to rest at least one week after arrival until used in the experiments. During this period they had free access to food (Ewos-R34 pellets) and water.
In vivo brain dialysis
Brain microdialysis experiments were performed in awake and freely moving rats as de- scribed by Waters et al. (1993 a). Briefly, male rats were anaesthetised with a 1:2 vol: vol mixture of xylazine (13 mg/kg i.p., Bayer Leverkusen) and ketamine (67 mg/kg i.p., Parke- Davis) and mounted into a Kopf stereotaxic instrument. The scull was exposed and burr holes were drilled. Co-ordiantes were calculated relative to bregma (striatum: A + 1, L - 2.6, D-6.3) according to Paxinos and Watson (1986). The probes were then slowly lowered down through the burr hole into the brain tissue until they reached their final position. The probes were cemented in this position with phosphatine dental cement (Svedia dental industri AB, Sweden). The rats were then housed individually in spherical cages for 48 hours before the dialysis experiments. On the day of experiment the rats were connected to a micro perfusion pump (CMA/100, Carnegie Medicine, Sweden). The pump was set to a perfusion speed of 2 ~tl/min and 40 ~tl samples were collected every 20 min. 30 gl of each sample was injected into the chromatograph. The perfusion medium was a Ringer solution containing in mmol/l: NaC1; 140, CaC12; 1.2, KC1; 3.0, MgC12; 1.0 and ascorbic acid; 0.04 according to Moghaddam and Bunney (1989).

42 N. Waters etal.
In vivo DP-5,6-ADTN displacement and motor activity measurements
The procedures applied in these experiments are essentially the same as described by Feenstra et al. (1983 a) but altered according to Carlsson and L6Poerg (1985). The rats were injected with DP-5,6-ADTN (0.25 btmol/kg s.c.). Sixty rain later the test drug was injected. After 5 rain the rats were placed in the motility meter boxes. The locomotor activity was recorded for 30min. After an additional 5 min the rats were killed by decapitation, their brains rapidly taken out and the striatum and cerebellum dissected for further analysis of their content of DP-5,6-ADTN and DOPAC. The "specific" binding to striatal binding sites were calculated by subtracting the DP-5,6-ADTN content of the cerebellum from that of the striatum. The motility measurements were carried out using a set of eight photocell animal motility meters (Digiscan activity monitor RXYZM(16)TAO, Omnitech Electronics, Inc. Columbus, Ohio, USA). The motility meters were kept in sound- and light-proof boxes equipped with semi-transparent mirrors allowing observation of the animals during the course of the experiments. For a more detailed discussion of the properties of DP-5,6- ADTN and other dopamine receptor ligands as tools for in vivo binding see Feenstra (1984). The procedures applied in these experiments were approved by the Animal Ethics Committee in G6teborg, Sweden.
Statistics
The statistical analysis of the results from the dialysis experiments were performed using the raw data collected in each experiment (n = 4-6 for each data point). The comparisons between dialysate dopamine levels at various time points after initiation of the experiments were performed using an ANOVA for repeated measures. For graphical clarity only sig- nificant differences between the effects of GBR12909 and the combination of GBR12909 and different dopamine receptor antagonists are depicted in Fig. 1. Data are expressed as % of baseline levels (three consecutive samples with < 7% variation before administration of GBR12909). The data collected in the DP-5,6-ADTN displacement and locomotor activity experiments displayed normal distribution, i.e. means and medians of each treatment group were similar, and these data were subjected to analysis of variance (ANOVA) followed
3000
2000 0 ~9
1000
0 '0 ' ' 0 6 120 180 ~ Time (min)
a b
Fig. 1. Temporal pattern of striatal dialysate levels of dopamine after treatment with a. GBR12909, 20 mg/kg s.c., in combination with administration of haloperidol, 1.3 gmol/kg s.c. (A), or (+)-UH232, 50btmol/kg s.c. (O), or (+)-AJ76, 50gmol/kg s.c. (O), or UH99194A, 50 gmol/kg s.c. (D), at point b. Data are expressed as % of baseline controls.
*P < 0.05 vs GBR12909 only at point b (60min)

Differential effects of dopamine D2 and D3 receptor antagonists 43
by Fischer's protected least significance difference (Fischer's PLSD). The relation between DP-5,6-ADTN displacement and locomotor activity were treated by regression analysis, fitting the data to a function of the first order, according to the method of least squares. Probability levels (P) less than 0.05 were considered significant.
Drugs
Drugs used were cis-(+)-(1S,2R)-5-methoxy-l-methyl-2-(n-propylamino) tetralin, (+)- A J76 HC1 (Upjohn, Kalamazoo, MI, USA), (+)-UH232 cis-(+ )-(1S,2R)-5-methoxy- 1- methyl-2-(n-dipropylamino)tetralin (Upjohn, Kalamazoo, MI, USA), raclopride tartrate (ASTRA Arcus, S6dertfilje, Sweden), remoxipride (ASTRA Arcus, S6dert/ilje, Sweden), haloperidol (Janssen Pharma, Helsingborg, Sweden), sulpiride (RBI, Natick, MA, USA), GBR12909 (Gist Brocade, Haarlem, The Netherlands). cis-(+)-(1S,2R)-5-methoxy-1- methyl-2-(n-dimethylamino)tetralin, (+)-AJ 118 (Johansson et al., 1987). (S-(-)-3-(3-Cy- anophenyl)- N-n-propyl piperidine), ( - )-DS121 HC1 (Sonesson et al., 1993), N, N-dipropyl- 2-amino-5,6-dihydroxy tetralin HC1, DP-5,6-ADTN, and U99194A were synthesised in our own laboratory. All other chemicals used were of analytical grade and obtained from standard commercial sources. The drugs were dissolved in physiological saline, in addition (+)-A J76 was heated and sonicated to dissolve. Haloperidol was dissolved in a few drops of glacial acetic acid and made up to volume in 5.5% glucose solution. Drugs were injected subcutaneously (s.c.) in the flank.
Results
In vivo brain dialysis
In each experiment GBR12909 (20mg/kg s.c. Fig. 1) induced a rapid increase in dialysate dopamine levels that reached a maximum at about 400% of baseline levels (5.2 • 0.5nM) 60rain after the injection that was coinciding with an increase in spontaneous locomotion as judged by gross behavioural observa- tions. When haloperidol (1.3~tmol/kg s.c. Fig. 1) were superimposed upon GBR12909 a rapid synergistic increase in dopamine to ~ 2500% of baseline levels was observed. Behavioural observations of the rats revealed that they displayed hypomotility and when tested they showed catalepsy ("freezing" in an awkward position with both forepaws on a 6 cm high block of wood for more than 20 seconds).
(+) -UH232 or (+)-AJ76 (501~mol/kg s.c. Fig. 1), superimposed upon GBR12909, produced a synergistic increase to about 1300% of controls. At these doses (+) -UH232 or (+)-AJ76, by themselves, increase dopamine to
200% and 300% respectively (Waters etal., 1993 a). No hypomotility was observed after treatment with these two agents. Combination of GBR12909 with the putative D3 antagonist U99194A (50 gmol/kg s.c. Fig. 1) did not lead to any significant increase in dopamine release as compared to GBR12909 only. However, the rats receiving the combination of GBR12909 with U99194A were actively exploring their environment and displayed a clear increase in locomotor activity and stereotyped sniffing as compared to treatment with GBR12909 only. The increase in behavioural activities lasted throughout the experimental session. Similar data was obtained from nucleus accumbens dialysates (not shown).

44 N. Waters et al.
Displacement of DP-5,6-ADTN and locomotor activity measurements
DP-5,6-ADTN at the dose used in these experiments increases locomotor ac- tivity to 205 4- 12%, n = 45, of non treated controls during a 30 minute session. At the end of this session the striatum contains 34.7 + 1.41 pmol DP-5,6-ADTN/ g wet tissue, n -- 44. The levels in the cerebellum, 6.1 + 0.35pmol/g, n = 44, are regarded as nonspecific binding to brain tissue and subtracted from the striatal levels to obtain the "specific binding". None of the dopamine receptor antagonists tested in this study changed the binding in the cerebellum signifi- cantly.
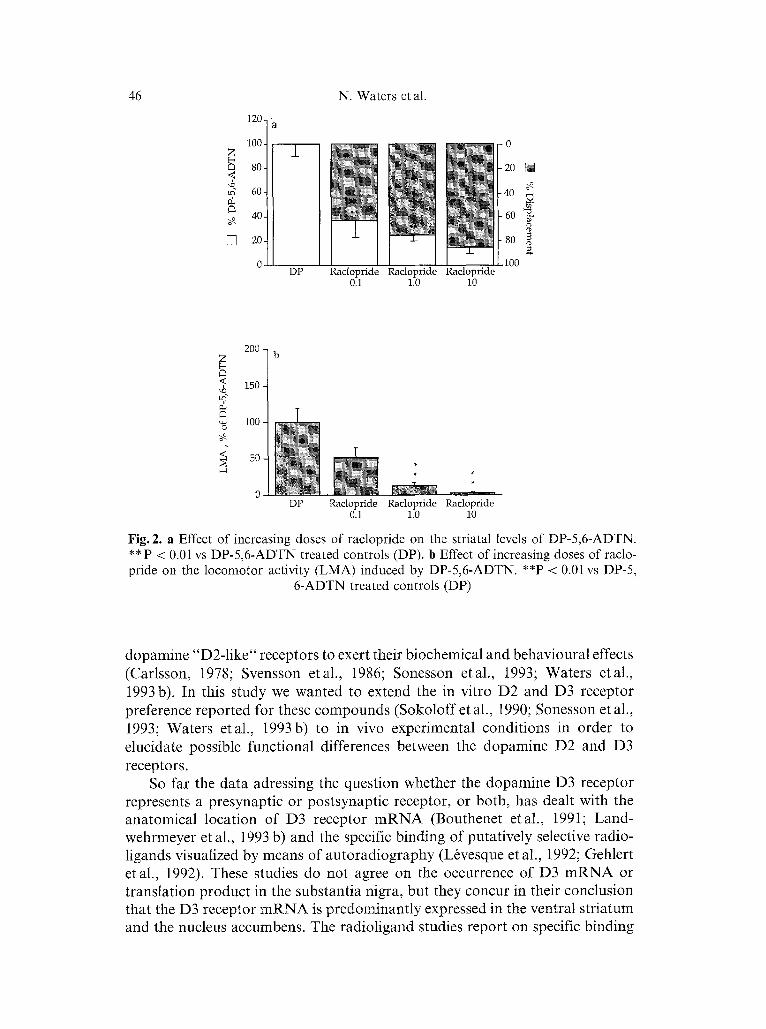
Increasing the doses of haloperidol (0.2-1.3 Bmol/kg s.c. Table 1) reduced the striatal levels of DP-5,6-ADTN maximally by about 85 %. The displacement of DP-5,6-ADTN was followed by a dose dependent decline in locomotor activity (Table 1) and at the higher doses catalepsy was noted. Raclopride displayed similar properties, i.e. a dose dependent decline in striatal DP-5,6- ADTN (Fig. 2 a, Table 1) to a maximal displacement of 85% was accompanied by a decline in motor activity (Fig. 2b), and at the highest doses catalepsy occurred.
(+)-UH232, (+)-AJ76 and the novel dopamine receptor antagonist ( - ) - DS121 (Table 1, Fig. 3 a) were similar in their effects. They all maximally de- creased the striatal levels of DP-5,6-ADTN by approximately 65-70% (Table 1, Fig. 3 a). At this level an asymptote was reached for these three compounds. When DP-5,6-ADTN is administered to the animals, it produces a decline in striatal metabolism of DA, measured as DOPAC (Feenstra etal., 1983 a). In this set of experiments DP-5,6-ADTN reduced the levels of DOPAC from 5.74• 0.16nmol/g, n = 21, to 3.61-4-0.1nmol/g, n = 4 3 . (-)-DS121 (100, 200 ~mol/kg s.c.) reversed this reduction (Fig. 3 b), indicating antagonism of dopamine autoreceptors. (+)-UH232, (+)-A J76 and ( - )-DS 121 reduced the locomotor activity to ~ 80-65% of that produced by DP-5,6-ADTN. Inter- estingly, this reduction is significantly less than that produced by haloperidol and raclopride (Table 1). Moreover, no catalepsy was apparent under the in- fluence of either (+)-UH232, (+)-AJ76 or (-)-DS121.
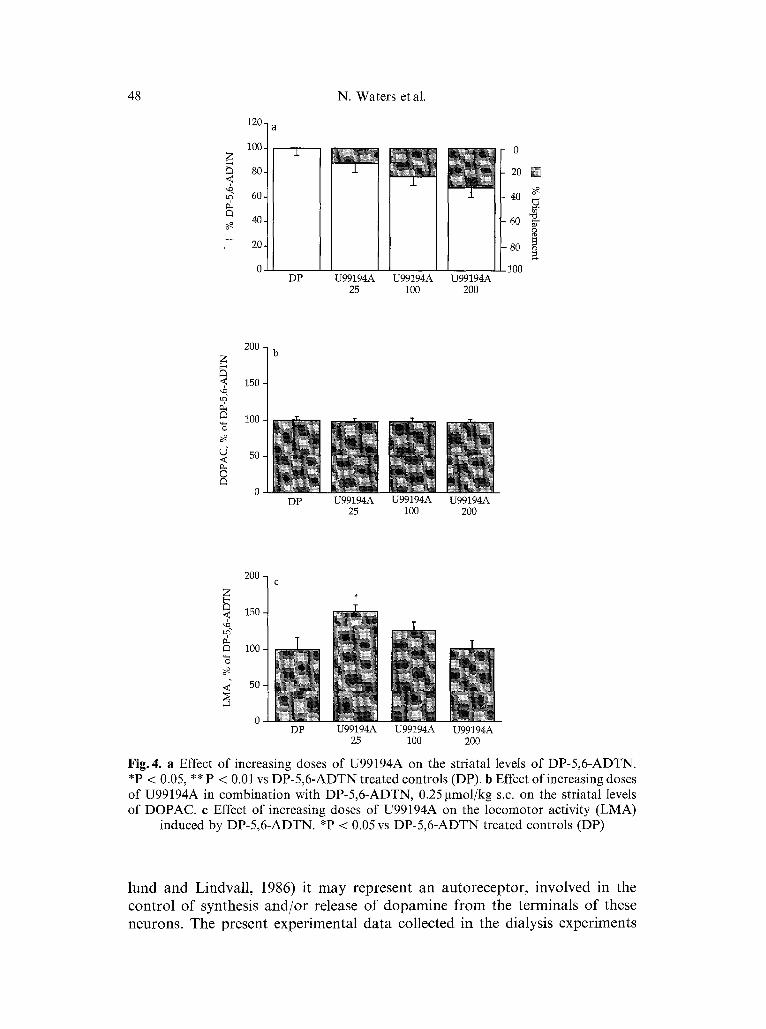
U99194A, the putative D3 receptor antagonist, was much less efficacious in displacing the agonist ligand DP-5,6-ADTN from its striatal binding sites. A maximal effect of 35% displacement was reached at the highest dose tested (200~tmol/kg s.c. Fig. 4 a). At this dose no change in locomotor activity or increase in DOPAC (Fig. 4 b) as compared to only DP-5,6-ADTN treated an- imals was measured. At the lower doses (25 and 100~tmol/kg s.c.), when U99194A displaced only ~ 10-20% of the striatal DP-5,6-ADTN binding sites, an increase in locomotor activity was observed as compared to DP-5,6-ADTN alone. This increase was significant at 25 gmol/kg s.c. (Fig. 4c).
To establish the relation between the D2/D3 receptor antagonist induced displacement of DP-5,6-ADTN and the subsequent effects on motor activity we included the "atypical dopamine antagonists" remoxipride (100, 200 gmol/

Differential effects of dopamine D2 and D3 receptor antagonists 45
Table 1. Effect of mixed dopamine D2/D3 receptor antagonists administered s.c., on the levels of DP-5, 6-ADTN 0.25 gmol/kg s.c. in the striatum, and on locomotor activity (LMA). Data are expressed as % reduction of DP-5, 6-ADTN treated controls (%-displacement) and % ofDP-5, 6-ADTN treated controls (LMA). *P < 0.05 vs displacement by haloperidol
1.3 gmol/kg s.c. and raclopride 10 gmol/kg s.c., n.t . not tested
Dose DP-5, 6-ADTN LMA n = (gmol/kg s.c.) %-displacement % of ctrl
Haloperidol 0.2 56 + 26 n.t. 4 0.4 75 + 11 n.t. 4 1.3 824-6 74- 1 5
Raclopride 0.1 634- 5 51 4- 14 5 1.0 75 + 1 13 • 4 5
10 84+ 11 34- 1 10
(+)-AJl l8 3.2 39 4- 7 62 4- 15 4 13 604-8 11 4-2 4 50 78 4- 3 5 4- 2 4
(+)-AJ76 25 38 4- 10 128 4- 1 4 50 59 + 3 54 + 6 4
200 65 4- 6* 20 4- 10 4
(+)-UH232 12.5 56 4- 7 78 4- 17 4 50 66 • 5 24 4- 7 4
200 69 4- 4* 22 + 13 4
(-)-DS121 25 45 4- 2 67 4- 14 4 100 66 4- 6 31 4- 9 4 200 67 4- 11" 35 4- 10 4
Remoxipride 100 48 4- 6 91 4- 9 8 200 43 • 11" 76 4- 9 8
Sulpiride 100 37 4- 5* 67 4- 14 8
U99194A 25 12 4- 6 152 • 9 6 100 23 • 6 126 + 12 6 200 32 4- 8* 101 4- 11 6
kg s.c.) and sulpiride (100 ~tmol/kg s.c.) in this investigation (Table 1, Fig. 5). In addi t ion experimental da ta on the cataleptogenic analog of ( + ) - U H 2 3 2 , i.e.
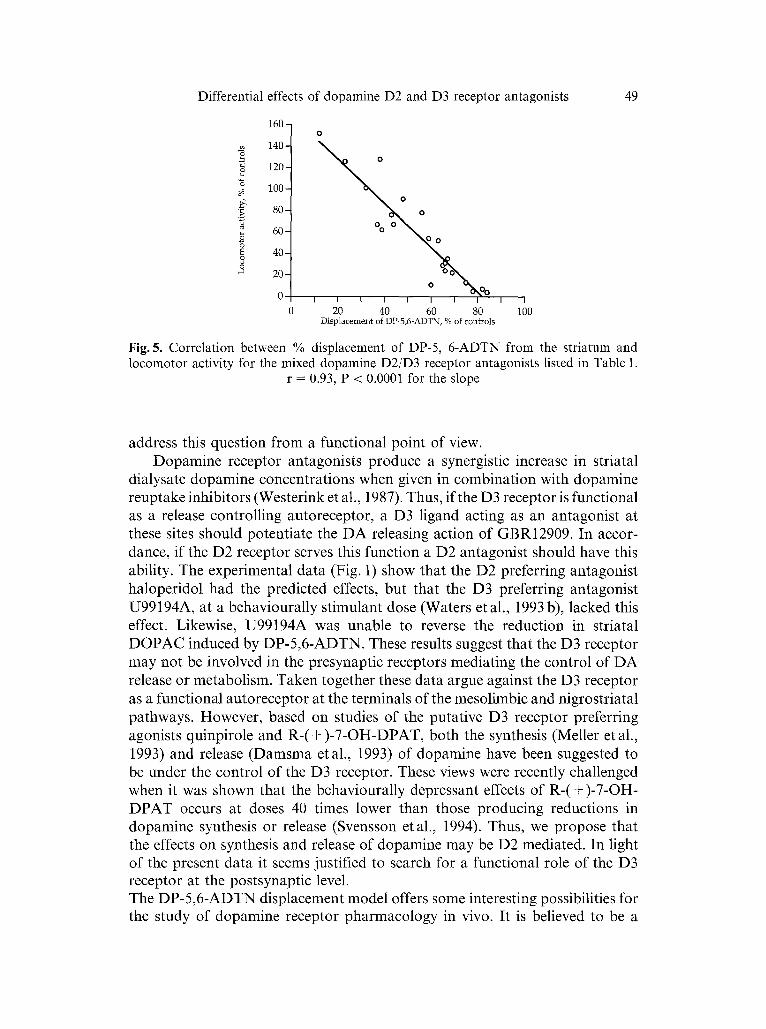
( + ) - A J118, (Svensson et al., 1986), were included in the calculations (Table 1, Fig. 5). A linear relat ion was found between the displacement of DP-5 ,6 -ADTN
and m o t o r activity (r = 0.93, P < 0.0001, Fig. 5).
Discuss ion
The compounds studied in this investigation represent a broad spectrum of dopamine receptor antagonis ts which are though t to interact pr imari ly with
46
Z
<
[ ]
120- a
i00- T
80-
60-
40-
20-
0 DP
N. Waters et al.
20 []
40
60
100 Raclopride Raclopride Raclopride
0.1 1.0 10
200 - Z
< 150 m,
,.6 100
#
DP Raclopride Raclopride Raclopride 0.1 1.0 10
Fig. 2. a Effect of increasing doses of raclopride on the striatal levels of DP-5,6-ADTN. ** P < 0.01 vs DP-5,6-ADTN treated controls (DP). b Effect of increasing doses of raclo- pride on the locomotor activity (LMA) induced by DP-5,6-ADTN. **P < 0.01 vs DP-5,
6-ADTN treated controls (DP)
dopamine "D2-1ike" receptors to exert their biochemical and behavioural effects (Carlsson, 1978; Svensson etal., 1986; Sonesson etal., 1993; Waters etal., 1993 b). In this study we wanted to extend the in vitro D2 and D3 receptor preference reported for these compounds (Sokoloff et al., 1990; Sonesson et al., 1993; Waters etal., 1993b) to in vivo experimental conditions in order to elucidate possible functional differences between the dopamine D2 and D3 receptors.
So far the data adressing the question whether the dopamine D3 receptor represents a presynaptic or postsynaptic receptor, or both, has dealt with the anatomical location of D3 receptor m R N A (Bouthenet etal., 1991; Land- wehrmeyer et al., 1993 b) and the specific binding of putatively selective radio- ligands visualized by means of autoradiography (L6vesque et al., 1992; Gehlert etal., 1992). These studies do not agree on the occurrence of D3 m R N A or translation product in the substantia nigra, but they concur in their conclusion that the D3 receptor rnRNA is predominantly expressed in the ventral striatum and the nucleus accumbens. The radioligand studies report on specific binding

Differential effects of dopamine D2 and D3 receptor antagonists
,6
[]
120.
100,
80-
60-
40-
20-
a
// // D P (-)-DS121 (-)-DS121 (-)-DS121
25 100 2OO
- 0
-20 [ ]
40 ~
60 ~_
80
100
47
Z C3 <
m,
U < �9
200
150
I00
50
D P (-)-DS121 (-)-DS121 (-)-DS121 25 100 200
200 -
z
150 - <, m,
~3 I00 -
50-
0 D P (-)-DS121 (-)-DS121 (-)-DS121
25 100 200
Fig. 3. a Effect of increasing doses of ( - ) -DS121 on the striatal levels of DP-5,6-ADTN. *** P < 0.001 vs DP-5,6-ADTN treated controls (DP). b Effect of increasing doses of ( - ) - DS121 in combination with DP-5,6-ADTN, 0.25gmol/kg s.c. on the striatal levels of DOPAC. ** P < 0.01 vs DP-5,6-ADTN treated controls (DP). e Effect of increasing doses of ( - ) -DS121 on the locomotor activity (LMA) induced by DP-5,6-ADTN. ** P < 0.01 vs
DP-5,6-ADTN treated controls (DP)
mainly in the ventral striatum, that coincide with m R N A , but also in the substantia nigra and the ventral tegmental area. If the dopamine D 3 receptor is expressed in the nigral dopaminergic cells that project to the striatum (Bj6rk-
48 N. Waters et al.
<
[ ]
120. a
loo.
80-
60.
40-
20.
0 DP
| / l M U99194A U99194A U99194A
25 100 200
0
20
o~ 40
60 e0
�9 80 ~
100
z
<
m,
C3
c3 <
�9
200 -
150 -
100 -
50 / DP U99194A U99194A U99194A
25 100 200
200
z
< 150
m,
C3 100
50
0 DP U99194A U99194A U99194A
25 100 200
Fig.4. a Effect of increasing doses of U99194A on the striatal levels of DP-5,6-ADTN. *P < 0.05, ** P < 0.01 vs DP-5,6-ADTN treated controls (DP). b Effect of increasing doses of U99194A in combination with DP-5,6-ADTN, 0.25 gtmol/kg s.c. on the striatal levels of DOPAC. e Effect of increasing doses of U99194A on the locomotor activity (LMA)
induced by DP-5,6-ADTN. *P < 0.05 vs DP-5,6-ADTN treated controls (DP)
lund and Lindvall, 1986) it may represent an autoreceptor, involved in the control o f synthesis and/or release o f dopamine from the terminals o f these neurons. The present experimental data collected in the dialysis experiments
Differential effects of dopamine D2 and D3 receptor antagonists 49
160 -
140 -
120 -
"8 100 -
�9 ~ 80-
6 0 -
4 0 -
2 0 -
0
O
0 O0 0
I I t I I I I I
20 40 60 80 Displacement of DP-5,6-ADTN, % of controls
I I
100
Fig. 5. Correlation between % displacement of DP-5, 6-ADTN from the striatum and locomotor activity for the mixed dopamine D2/D3 receptor antagonists listed in Table 1.
r = 0.93, P < 0.0001 for the slope
address this question from a functional point of view. Dopamine receptor antagonists produce a synergistic increase in striatal
dialysate dopamine concentrations when given in combination with dopamine reuptake inhibitors (Westerink et al., 1987). Thus, if the D3 receptor is functional as a release controlling autoreceptor, a D3 ligand acting as an antagonist at these sites should potentiate the DA releasing action of GBR12909. In accor- dance, if the D2 receptor serves this function a D2 antagonist should have this ability. The experimental data (Fig. 1) show that the D2 preferring antagonist haloperidol had the predicted effects, but that the D3 preferring antagonist U99194A, at a behaviourally stimulant dose (Waters et al., 1993 b), lacked this effect. Likewise, U99194A was unable to reverse the reduction in striatal DOPAC induced by DP-5,6-ADTN. These results suggest that the D3 receptor may not be involved in the presynaptic receptors mediating the control of DA release or metabolism. Taken together these data argue against the D3 receptor as a functional autoreceptor at the terminals of the mesolimbic and nigrostriatal pathways. However, based on studies of the putative D3 receptor preferring agonists quinpirole and R-(+)-7-OH-DPAT, both the synthesis (Meller et al., 1993) and release (Damsma etal., 1993) of dopamine have been suggested to be under the control of the D3 receptor. These views were recently challenged when it was shown that the behaviourally depressant effects of R-(+)-7-OH- DPAT occurs at doses 40 times lower than those producing reductions in dopamine synthesis or release (Svensson etal., 1994). Thus, we propose that the effects on synthesis and release of dopamine may be D2 mediated. In light of the present data it seems justified to search for a functional role of the D3 receptor at the postsynaptic level. The DP-5,6-ADTN displacement model offers some interesting possibilities for the study of dopamine receptor pharmacology in vivo. It is believed to be a

50 N. Waters etal.
model mainly for the study of drug interactions of postsynaptic D2-type receptor sites within the dopaminergic system (Feenstra et al., 1983 a; Hjorth and Carls- son 1988; Hajos et al., 1988). This model also gives the opportunity to monitor the behavioural effects of the interaction between the D2-type receptor agonist DP-5,6-ADTN and any test substance at the same time. However, the possibility to determine the specificity of the receptor(s) involved in the interactions is limited to the D2-family, and the interpretation of the data in terms of specific receptor subtypes so far relies on previous in vitro radioligand binding char- acterisation of the test compounds. Moreover, the finding that the lobules 9 and 10 of the cerebellar cortex contains D3 receptor mRNA (Bouthenet et al., 1991) warrants further caution in the interpretation of the "specific binding" of DP-5,6-ADTN to the striatum since we use the cerebellum as a control for non-specific binding (see Method section and for further discussion see Carlsson and L6fberg, 1985). Thus, we are at risk to underestimate the contribution of the D3 receptor in the striatal binding. However, the models in this study are not designed for proper quantification of the various dopamine D2 receptor subtypes. The qualitative validity for this model is, however, substantiated by the strong linear correlation between the displacement of DP-5,6-ADTN from the striatum and the concomitant behavioural effects by the dopamine D2 type antagonists (Fig. 5).
The dopamine D2 preferring (Sokoloff etal., 1990) receptor antagonists haloperidol and raclopride are very similar in their displacement of DP-5,6- ADTN. Their displacement of the agonist ligand is accompanied by a strong reduction in locomotor activity and subsequent catalepsy. They are both able to displace DP-5,6-ADTN to ,~ 85%, which is suggested to be the maximal level possible in this model (Feenstra et al., 1983 a). The intercept (at 82%) with the x-axis in Fig. 5 strongly supports this notion. Thus, preferential binding to, and displacement of an agonist from, D2 receptors leads to hypomotility and subsequently to catalepsy. The behaviourally stimulatory and somewhat D3 preferring dopamine antag- onists (+)-UH232, (+) -A J76 (Svensson et al., 1986) and ( - )-DS 121 (Sonesson etal., 1993) all seem to reach a displacement asymptote at about 65%. This is significantly lower than haloperidol and raclopride. In addition, the reduction in locomotor activity is less than that produced by haloperidol and raclopride. These data suggest that the classical (i.e. cataleptogenic) DA antagonists can reach a population of DP-5,6-ADTN binding sites that is not available for (+)- UH232, (+)-AJ76 or (-)-DS121. Thus the displaceable receptor populations do not seem to be fully identical for these two classes of drugs. The reason for this difference is not clear. Interestingly, (+) -A J118 a structural analog of( + )- UH232 and (+)-A J76 that has a "classical" dopamine D2 receptor antagonist profile, consisting of increases in the synthesis of dopamine, hypolocomotion and catalepsy (Svensson etal., 1986) displaces ~ 80% of the DP-5,6-ADTN from its binding sites in the striatum, which is similar to haloperidol and raclopride (Table 1). Thus, one could speculate that these additional binding

Differential effects of dopamine D2 and D3 receptor antagonists 51
sites, not reached by (+)-UH232, (+)-AJ76 or (-)-DS121, are involved in the cataleptogenic properties of the classical neuroleptics as well as of (+)- A J118.The existence of a population of [125I]iodosulpiride displaceable sites in the striatum, other than dopamine D2 or D3, were suggested recently (Schotte etal., 1992). It is also tempting to associate the present findings in the rat to receptor occupancy studies with [11C]raclopride in man. It has been shown that an occupancy of 82 + 4% of the D2 receptors is needed to produce EPS while an occupancy of 74 + 4% by neuroleptics does not procedure EPS (Farde et al., 1992). Furthermore, the almost maximal displacement of DP-5,6-ADTN from postsynaptic dopamine receptors needed to produce severe motor dys- functions in this study, resembles the nearly total reduction in dopamine avail- ability in 6-hydroxydopamine depleted rats required to produce motor dis- turbances similar to catalepsy (Zigmond etal., 1990). Thus, compounds like (+)-UH232 and (-)-DS121 that, for some hitherto unidentified reason, are unable to totally displace an agonist ligand from striatal dopamine receptor binding sites in the rat, may prove beneficial in terms of reduced neurological side effects, in comparison to classical neuroleptics, when administered to man. In this context it is interesting to note that antipsychotics that are clinically classified as disinhibitory and less prone to produce parkinsonian side effects seem to have a slightly higher preference for the D3 receptor, in vitro, than their classical counterparts (Schwartz et al., 1993).
We found that the putative D3 receptor antagonist U99194A only partially displaced DP-5,6-ADTN at behaviourally stimulant doses (Waters etal., 1993 b). If it is appropriate to extrapolate the in vitro binding to the in vivo situation the population displaced by this compound should consist to a large part of D3 receptor sites. This suggests that the D3 receptors are less abundant than the D2 receptors in the area studied here. This is supported by the esti- mation that the D3 receptor represents about 10-20% (L6vesque etal., 1992) 13% (Gehlert et al., 1992) or 10% (Boundy et al., 1993) of the total "D2"-type protein in the rat striatum. At low levels of displacement by U99194A (10- 20%) we observed a relative increase in DP-5,6-ADTN-induced locomotor activity. This effect on behaviour is similar to the increase in locomotor activity observed when U99194A is superimposed upon either apomorphine, d-am- phetamine (Waters etal., 1993b) or GBR12909 (see results), and may be ex- plained similarly; a small population of postsynaptic dopamine receptors, that under these experimental conditions are stimulated by the agonist DP-5,6- ADTN, are involved in the suppression of locomotor activity. When these receptors are occupied by an antagonist, a more pronounced effect of the parallel stimulation of postsynaptic excitatory dopamine receptors is allowed and ex- pressed as a relative increase in locomotor activity. These postsynaptic inhibitory receptors presumably belong to the D3 type. Similar effects on behaviour have been reported when sulpiride is superimposed upon pergolide in dopamine depleted rats (Rubinstein etal., 1988). Furthermore, it has been proposed that the behaviourally depressant effects by DP-5,6-ADTN is dependent on post-

52 N. Waters et al.
synaptic D2 receptors in the striatum (Scheel-KriJger, 1986). Previous data interpreted in favour of such postsynaptic behaviourally inhibitory "D2 like" receptors have recently been reviewed by Sffthle (1992).
In summary, we find that D2 receptor preferring antagonists are powerful displacers of DP-5,6-ADTN from its binding sites in the striatum and that maximal displacement is followed by strong hypomotility and catalepsy. Dis- placement of the agonist by the D3 preferring antagonist U99194A is at best only partial and results in synergistic increases in locomotor activity. These are the extreme points in the strong correlation between displacement of DP-5,6- ADTN and locomotor activity by mixed D2/D3 antagonists shown here. Pro- voking the release of dopamine by superimposing haloperidol upon GBR12909 leads to a synergistic increase in striatal dialysate dopamine concentrations. This effect is absent when combining GBR12909 with the putative D3 antagonist U99194A. Taken together these data give support for the suggestion that the dopamine D3 receptor is functionally relevant at the postsynaptic level (Waters etal., 1993b). Here, in contrast to the D2 receptor, it is proposed to exert an inhibitory influence on psychomotor functions.
Acknowledgements
The financial support provided by the Upjohn Company and the Medical Faculty, Uni- versity of G6teborg is gratefully acknowledged. We gratefully acknowledge S. Ahl for the construction and building of the motility meter boxes and T. Kling-Petersen for the com- puterisation of the same. Dr. T. Magnusson is acknowledged for valuable and stimulating discussions about the statistical evaluation of the data.
References
Bj6rklund A, Lindvall O (1986) Catecholaminergic brain stem regulatory systems. In: Field J (ed) Handbook of physiology. The nervous system IV. Am Physiol Soc, Washington DC, pp 155-235
Boundy VA, Luedtke RR, Gallitano AL, Smith JE, Filtz TM, Kallen RG, Molinoff PB (1993) Expression, and characterization of the rat D3 dopamine receptor: pharma- cologic properties and development of antibodies. J Pharmacol Exp Ther 264: 1002- 1011
Bouthenet ML, Souil E, Martres MP, Sokoloff P, Schwartz JC (1991) Localization of dopamine D3 receptor mRNA in the rat brain using in situ hybridization histochemistry comparison with dopamine D2 receptor mRNA. Brain Res 564:203-219
Carlsson A (1975) Receptor-mediated control of dopamine metabolism. In: Usdin E, Bunney Jr WE (ed) Pre- and postsynaptic receptors. Marcel Dekker, New York, pp 49-65
Carlsson A (1978) Mechanism of action of neuroleptic drugs. In: Lipton MA, DiMascio A, Killam KF (eds) Psychopharmacology: a generation of progress. Raven Press, New York, pp 1057-1070
Carlsson A, L6fberg L (1985) In vivo displacement by 3-PPP enantiomers of N,N-Dipropyl- 5,6-ADTN from dopamine receptor-binding sites in rat striatum. J Neural Transm 64: 173-185
Civelli O, Bunzow JR, Grandy DK, Zhou Q-Y, Van Tol HHM (1991) Molecular biology of the dopamine receptors. Eur J Pharmacol 207:277-286
Cools AR, Van Rossum JM (1976) Excitation-mediating and inhibition-mediating dopa-

Differential effects of dopamine D2 and D3 receptor antagonists 53
mine-receptors: a new concept towards a better understanding of electrophysiological, biochemical, pharmacological, functional and clinical data. Psychopharmacologia (Berl) 45:243-254
Cools AR, van Rossum JM (1980) Multiple receptors for brain dopamine in behavior regulation: concept of dopamine-E and dopamine-I receptors. Life Sci 27:1237-1253
Damsma G, Bottema T, Westerink BHC, Tepper PG, Dijkstra D, Pugsley TA, MacKenzie RG, Heffner TG, Wikstr6m H (1993) Pharmacological aspects of R-(+ )-7-OH-DPAT, a putative dopamine D 3 receptor ligand. Eur J Pharmacol 249:R9-R10
Di Chiara G, Porceddu ML, Spano PF, Gessa GL (1977) Haloperidol increases and apomorphine decreases striatal dopamine metabolism after destruction of striatal do- pamine-sensitive adenylate cyclase by kainic acid. Brain Res 130:374-382
Farde L, Nordstr6m A-L, Wiesel F-A, Pauli S, Halldin C, Sedvall G (1992) Positron emission tomographic analysis of central D 1 and Da dopamine receptor occupancy in patients treated with classical neuroleptics and clozapine. Relation to extrapyramidal side effects. Arch Gen Psychiatry 49:538-544
Feenstra M, Rollema H, Mulder T, Westerink BHC, Horn AS (1983 a) In vivo dopamine receptor binding with a non-radioactively labeled agonist, dipropyl-5,6-ADTN. Life Sci 32:1313 1323
Feenstra MGP, Rollema H, Mulder TBA, De Vries JB, Horn AS (1983 b) In vivo dopamine receptor agonist binding in rat brain: relation with pharmacological effects. Eur J Pharmacol 90:433-436
Gehlert DR, Gackenheimer SL, Seeman P, Schaus J (1992) Autoradiographic localization of [3H]quinpirole binding to dopamine D2 and D 3 receptors in rat brain. Eur J Phar- macol 211:189-194
Hajos M, Hjorth S, Svensson K, Carlsson A (1988) In vivo dopamine (DA) receptor binding and behavioural effects of putative DA autoreceptor antagonists (+)-A J76 and (+)- UH232 in rats with a unilateral nigral 6-OH-DA lesion. Exp Brain Res 70:577-584
Hjorth S, Carlsson A (1988) In vivo receptor binding, neurochemical and functional studies with the dopamine D-1 receptor antagonist SCH 23390. J Neural Transm 72:83-97
Johansson AM, Nilsson JLG, Karl6n A, Hacksell U, Sanchez D, Svensson K, Hjorth S, Carlsson A, Sundell S, Kenne L (1987) C1- and C3-Methyl-substituted derivatives of 7-hydroxy-2-(di-n-propylamino)tetralin: activities at central dopamine receptors. J Med Chem 30:1827-1837
Kebabian JW, Calne DB (1979) Multiple receptors for dopamine. Nature 277:93-96 Landwehrmeyer B, Mengod G, Palacios JM (1993 a) Dopamine D3 receptor mRNA and
binding sites in human brain. Mol Brain Res 18:187-192 Landwehrmeyer B, Mengod G, Palacios JM (1993 b) Differential visualization of Dopamine
D2 and D3 receptor sites in rat brain. A comparative study using in situ hybridization histochemistry and ligand binding autoradiography. Eur J Neurosci 5:145-153
L6vesque D, Diaz J, Pilon C, Martres M-P-, Giros B, Souil E, Schott D, Morgat J-L, Schwartz J-C, Sokoloff P (1992) Identification, characterization, and localization of the dopamine D 3 receptor in rat brain using 7-[3H]hydroxy-N,N-di-n-propyl-2-ami - notetralin. Proc Natl Acad Sci USA 89:8155-8159
Ljungberg T, Ungerstedt U (1978) Classification of neuroleptic drugs according to their ability to inhibit apomorphine-induced locomotion and gnawing: evidence for two different mechanisms of action. Psychopharmacology 56:239-247
Meller E, Bohmaker K, Goldstein M, Bashman DA (1993) Evidence that striatal synthesis- inhibiting autoreceptors are dopamine D3 receptors. Eur J Pharmacol 249:R5-R6
Moghaddam B, Bunney BS (1989) Ionic composition of microdialysis perfusing solution alters the pharmacological responsiveness and basal outflow of striatal dopamine. J Neurochem 53:652-654
Rubinstein M, Gershanik O, Stefano FJE (1988) Postsynaptic bimodal effect of sulpiride

54 N. Waters et al.
on locomotor activity induced by pergolide in catecholamine-depleted mice. Naunyn Schmiedebergs Arch Pharmacol 337:115-117
Scheel-Kriiger J (1986) The syndrome of sedation and yawning behaviour in the rat is dependent on postsynaptic dopamine D-2 receptors. Psychopharmacology $32
Schotte A, Janssen PFM, Gommeren W, Luyten WHLM, Leysen JE (1992) Autoradio- graphic evidence for the occlusion of rat brain dopamine D 3 receptors in vivo. Eur J Pharmacol 218:373-375
Schwartz J-C, Levesque D, Martres MP, Sokoloff P (1993) Dopamine D3 receptor: basic and clinical aspects. Clin Neuropharmacol 16:295-314
Seeman P (1980) Brain dopamine receptors. Pharmacol Rev 32:229-313 Sibley DR, Monsma Jr. FJ (1992) Molecular biology of dopamine receptors. TiPS 13:61-
69 Snyder SH (1990) The dopamine connection. Nature 347:121-122 Sokoloff P, Andrieux M, Besangon R, Pilon C, Martres M-P, Giros B, Schwartz J-C (1992)
Pharmacology of human dopamine D 3 receptor expressed in a mammalian cell line: comparison with D 2 receptor. Eur J Pharmacol 225:331-337
SokoloffP, Giros B, Martres M-P, Bouthenet M-L, Schwartz J-C (1990) Molecular cloning and characterization of a novel dopamine receptor (D3) as a target for neuroleptics. Nature 347:146-151
Sonesson C, Waters N, Svensson K, Carlsson A, Smith M-W, Piercey M-F, Mayor E, Wikstr6m H (1993) Substituted 3-phenyl piperidines: new centrally acting dopamine autoreceptor antagonists. J Med Chem 36:3188-3196
Strange PG (1991) Interesting times for dopamine receptors. TINS 14:43-45 Sffthle L (1992) Do autoreceptors mediate dopamine agonist-induced yawning and suppres-
sion of exploration? A critical review. Psychopharmacology 106:1-13 Surmeier D J, Eberwine J, Wilson C J, Cao Y, Stefani A, Kitai ST (1992) Dopamine receptor
subtypes colocalize in rat striatonigral neurons. Proc Natl Sci 89:10178-10182 Surmeier D J, Reiner A, Levine MS, Ariano MA (1993) Are neostriatal dopamine receptors
co-localized? TiNS 16:299-305 Svensson K, Carlsson A, Johansson AM, Arvidsson L-E, Nilsson JLG (1986) A homologous
series of N-alkylated Cis-(+)-(1S,2R)-5-Methoxy-l-methyl-2-aminotetralins: central dopamine receptor antagonists showing profiles ranging from classical antagonism to selectivity for autoreceptors. J Neural Transm 65:29-38
Svensson K, Carlsson A, Waters N (1994) Locomotor inhibition by the D3 ligand R-(+)- 7-OH-DPAT is independent of changes in dopamine release. J Neural Transm [Gen Sect] 95:71-74
Svensson K, Johansson AM, Magnusson T, Carlsson A (1986) (+)-AJ76 and (+)-UH232: central stimulants acting as preferential dopamine autoreceptor antagonists. Naunyn Schmiedebergs Arch Pharmacol 334:234-245
Waters N, Lagerkvist S, L6fberg L, Piercey M, Carlsson A (1993 a) The dopamine D3 receptor and autoreceptor preferring antagonists (+)-A J76 and (+)-UH232; a micro- dialysis study. Eur J Pharmacol 242:151-163
Waters N, Svensson K, Haadsma-Svensson SR, Smith MW, Carlsson A (1993b) The dopamine D3-receptor: a postsynaptic receptor inhibitory on rat locomotor activity. J Neural Transm [GenSect] 94:11-19
Westerink BHC, Damsma G, De Vries JB, Koning H (1987) Dopamine re-uptake inhibitors show inconsistent effects on the in vivo release ofdopamine as measured by intracerebral dialysis in the rat. Eur J Pharmacol 135:123-128
Westerink BHC, Damsma G, Rollema H, De Vries JB, Horn AS (1987) Scope and limi- tations of in vivo brain dialysis: a comparison of its application to various neurotrans- mitter systems. Life Sci 41:1763 1776
Zetterstr6m T, Sharp T, Marsden CA, Ungerstedt U (1983) In vivo measurement of

Differential effects of dopamine D2 and D3 receptor antagonists 55
dopamine and its metabolites by intracerebral dialysis: changes after d-amphetamine. J Neurochem 41:1769-1773
Zigmond M J, Abercrombie ED, Berger TW, Grace AA, Stricker EM (1990) Compensations after lesion of central dopaminergic neurons: some clinical and basic implications. TiNS 13:290-296
Authors' address: N. Waters, Department of Pharmacology, University of G6teborg, Medicinaregatan 7, S-41390 G6teborg, Sweden
Received February 14, 1994




















