
TN 19: Gradient Elution In Ion Chromatography - Thermo Fisher
Upload
independentCategory
view
5download
0
ORIGINAL PAPER
Differential Binding to Glycotopes Among the Layers of ThreeMammalian Retinal Neurons by Man-Containing N-linkedGlycan, Ta (Galb1–3GalNAca1-), Tn (GalNAca1-Ser/Thr)and Ib/IIb (Galb1–3/4GlcNAcb-) Reactive Lectins
Wei-Chi Wu Æ Chi-Chun Lai Æ Jia-Hau Liu ÆTanuja Singh Æ Lien-Ming Li Æ Willy J. Peumans ÆEls J. M. Van Damme Æ Albert M. Wu
Accepted: 22 February 2006 / Published online: 23 May 2006
� Springer Science+Business Media, Inc. 2006
Abstract Carbohydrate structures between retinal neu-
rons and retinal pigment epithelium (RPE) play an
important role in maintaining the integrity of retinal
adhesion to underlying RPE, and in retinal detachment
pathogenesis. Since relevant knowledge is still in the
primary stage, glycotopes on the adult retina of mongrel
canines (dog), micropigs and Sprague-Dawley rats were
examined by lectino-histochemistry, using a panel of 16
different lectins. Paraffin sections of eyes were stained
with biotinylated lectins, and visualized by streptavidin-
peroxidase and diaminobenzidine staining. Mapping the
affinity profiles, it is concluded that: (i) all sections of
the retina reacted well with Morniga M, suggesting that
N-linked glycans are present in all layers of the retina;
(ii) no detectable human blood group ABH active gly-
cotopes were found among retinal layers; (iii) outer and
inner segments contained glycoconjugates rich in ligands
reacting with Ta (Galb1–3GalNAca1-Ser/Thr) and Tn
(GalNAca1-Ser/Thr) specific lectins; (iv) cone cells of
retina specifically bound peanut agglutinin (PNA), which
recognizes Ta residues and could be used as a specific
marker for these photoreceptors; (v) the retinas of rat,
dog and pig, had a similar binding profile but with dif-
ferent intensity; (vi) each retinal layer had its own
binding characteristic. This information may provide
useful background knowledge for normal retinal physi-
ology and miscellaneous retinal diseases, including reti-
nal detachment (RD) and age-related macular
degeneration (ARMD).
Keywords Carbohydrate Æ Disaccharide structural
units Æ Lectins Æ Retina
Introduction
The retina is a light-sensitive membrane lining the
internal surface of the eye that transduces light intensity
and color into electrical signals. The neurons of the
retina are divided into three layers: (i) the most external
(the outer layer) is the photoreceptor cell layer, which
includes the outer (OS) and inner (IS) segments and the
layer of photoreceptor cell bodies (outer nuclear layer,
ONL); (ii) the layer of intermediate neurons (inner
nuclear layer, INL); and (iii) the layer of ganglion cells
(GCL). The axons of ganglion cells converge to form the
nerve fiber layer (NFL) and exit the eye as the optic
nerve. The synapses are confined to two synaptic, or
plexiform layers—the outer (OPL) and inner (IPL)
plexiform layers. The photoreceptor layer of the retina is
intimately apposed to the retinal pigment epithelium
(RPE). The outer limiting membrane (OLM) is formed
by junctional complexes between cell membranes of the
W.-C. Wu Æ C.-C. Lai Æ J.-H. Liu Æ L.-M. Li
Department of Ophthalmology, Chang Gung Memorial Hospital,
Tao-yuan, Taiwan
T. Singh Æ A. M. Wu (&)
Glyco-Immunochemistry Research Laboratory, Institute of
Molecular and Cellular Biology, Chang-Gung University, 333,
Tao-yuan, Kwei-san, Taiwan
e-mail: [email protected]
W. J. Peumans Æ E. J. M. Van Damme
Department of Molecular Biotechnology, Faculty of Bioscience
Engineering, Ghent University, Coupure Links 653, 9000 Gent,
Belgium
Neurochem Res (2006) 31:619–628
DOI 10.1007/s11064-006-9060-8
123

major glial cells, the Muller cells, and the inner seg-
ment of photoreceptor cells. The inner limiting
membrane (ILM) consists of a basement membrane,
which is actually a surface modification of the vitreous
body, and the expanded vitreal processes of Muller cells
[1, 2].
The retinal interphotoreceptor matrix (IPM), which
lies between the apical surfaces of the neural retina and
the RPE, surrounds the ellipsoid portion of the inner and
outer segments of photoreceptor cells. The IPM is
important in mediating the interaction between the pig-
ment epithelium and the neural retina. Carbohydrate
structures in the IPM may play an important role in the
maintenance of the integrity of retinal adhesion to its
underlying RPE structure [3]. However, related infor-
mation is still limited, especially the functional role and
distribution of glycotopes in the retina. We examined the
expression of glycotopes of the mature retina of mongrel
canine (dog), micropig and Sprague-Dawley rat by
lectino-histochemistry, using a panel of 16 different
lectins with mannose-containing N-linked, Ta (Galb1–
3GalNAca1-Ser/Thr)/Tn (GalNAca1-Ser/Thr), I/II
(Galb1–3/4GlcNAc), E (Gala1–4Gal), B (Gala1–3Gal),
A/F (GalNAca1–3Gal/GalNAc) and H (LFuca1–2Gal)
carbohydrate specificities [4, 5]. In order to exclude the
possibility that differences in binding pattern affinity
were due to non-specific interactions, a series of poly-
valent glycotopes occurring in natural glycoproteins were
used to inhibit the interactions of lectin-retinal glycans.
Materials and methods
Animals
Twenty Sprague-Dawley rats (weight range: 150–200 g),
ten mongrel canines (age: 3-year-old, weight: 20 kg) and
ten Yucatan micropigs (age range, 4–6 months old;
weight range, 9–14 kg) were used in this study. The
animals were handled in accordance with the ARVO
statement for the Use of Animal in Ophthalmic and
Vision Research. The rats, dogs, and pigs were
anesthetized with intramuscular injections of an equal
volume mixture of 2-(2,6-xylidino)-5,6-dihydro-4H-1,3-
thiazine-hydrochloride, methylparaben (Rompun; Bayer
AG, Leverkusen, Germany) and 50 mg/ml Ketamine
hydrochloride (Ketalar; Parke-Davis, Morris Plains, NJ)
of 1.5 ml/kg, 0.5 ml/kg, and 1 ml/kg, respectively. The
eyeballs were excised from the anesthetized animals.
After removal of the eyeballs, the animals were sacri-
ficed with intravenous injection of sodium chloride
(Taiwan Biotech, Tao-yuan, Taiwan).
Lectins
Morniga M was purified from Morus nigra bark by a
combination of affinity chromatography and ion exchange
chromatography as previously described [6]. Artocarpus
integrifolia agglutinin (Jacalin; L-7775), Ricinus communis
agglutinin 1 (RCA1 or RCA120; L-7886), Ricin (RCA60;
L-8508), Vicia villosa B4 (VVL-B4; L-7638), Agaricus
bisporus agglutinin (ABA; L-5640), Arachis hypogea
(PNA), wheat germ agglutinin (WGA; L-1005), Griffonia
simplicifolia lectin-I isolectin A4 (GSI-A4; L-1509),
Anguilla anguilla agglutinin (AAA) from fresh water eel,
were purchased from Sigma (St. Louis, MO, USA). Ama-
ranthus caudatus lectin (ACL; B-1255), Erythrina crista-
galli lectin (ECL; B-1145) and Griffonia simplicifolia
lectin-I isolectin B4 (GSI-B4; B-1205) were purchased
from Vector Laboratories (Burlingame, CA, USA). Macl-
ura pomifera lectin (MPL) was prepared as previously
described [7]. Abrus precatorious agglutinin (APA) and
abrin-a were kindly given by Drs. L.P. Chow and J.Y. Lin,
Institute of Biochemistry, College of Medicine, National
Taiwan University, Taipei, Taiwan. Biotinylation of lectins
were performed as described previously [8].
Immunohistochemistry
Immediately after dissection, the eyeballs were fixed
overnight at 4�C with 4% paraformaldehyde in phosphate-
buffered saline (PBS), then dehydrated and embedded in
paraffin. The retinal sections (5 lm thick) were placed on
slides that had been coated with 1% gelatin and 0.1%
chromium potassium sulfate (Sigma) in distilled water to
promote adhesion of the sections to the glass surface.
Paraffin sections of retinas were soaked in xylene to re-
move paraffin, washed with PBS and incubated in 3%
H2O2 for 10 min to block endogenous peroxidase activity.
Samples were then blocked with 1% goat serum and 1%
bovine serum albumin for 30 min after washing in PBS.
Samples were then incubated with a panel of biotinylated
lectins with a concentration of 1 lg/ml for 60 min in a
dark, humidified chamber. Control sections were prepared
by either incubating in buffer solution without lectins or
preincubating the lectins with their corresponding specific
inhibiting sugars at a concentration of 10 lg/ml. The bio-
tinylated lectins and their inhibitory sugars used in this
study are listed in Table 1. After washing with PBS,
samples were incubated with 2 lg/ml peroxidase-labeled
streptavidin (Sigma) for 1 h. The lectin binding sites were
then visualized by adding diaminobenzidine as a chromo-
gen. Hematoxylin and eosin-stained sections served as
controls to assess the histologic quality of the specimens.
The slides were dehydrated, cover slipped and examined
for the location of brown reaction product. Photos were
620 Neurochem Res (2006) 31:619–628
123

taken and printed under the same conditions so that direct
comparisons of binding intensity could be made.
The intensity of lectin binding was quantitatively graded
as: ), negative; +, weak; ++, moderate; and +++, strong.
Results
Sections stained with hematoxylin and eosin (Figs. 1–3)
revealed well-preserved retina in the retinal specimen. In
most of the eyes the retina had become detached during
processing. With some variations, the lectin-binding profile
of all specimens was similar even in different species of
mammalian retinal neurons. Table 2 summarizes the
results of lectin-carbohydrate binding in the three mam-
malian retinal neurons.
Interaction of retinal glycans with a lectin exhibiting
an exclusive specificity towards mannose-containing
N-linked glycans—Morus nigra lectin (Morniga M)
(Figs. 4–6)
In rat retina, Morniga M bound strongly to GCL, moder-
ately to RPE, photoreceptor OS and IS, ONL, OPL, INL,
IPL, NFL, and blood vessel walls. Morniga M bound,
weakly to ILM. In the retina of dog and pig, the stain
showed strong binding to the OS of photoreceptor cell
layer, ganglion cells, and blood vessel walls, and moderate
binding to RPE, photoreceptor IS, ONL, OPL, INL, IPL,
NFL, and ILM. There was no obvious difference in the
binding pattern of Morniga M among the retina of rat, dog,
and pig, implying the presence of N-linked glycans in all
layers of the retina.
Interaction of retinal glycans with GalNAca1-Ser/Thr
(Tn) specific lectin—Vicia villosa isolectin B4
(VVL-B4) (Figs. 7–9)
In the rat retina, VVL-B4 bound moderately to the regions
of photoreceptor OS and IS. Photoreceptor OS were in-
tensely labeled in the retinas of dog and pig. Photoreceptor
IS were moderately stained in the retina of dog, and heavily
stained in pig retina. There was no binding at other retinal
layers.
Interaction of retinal glycans with Galb1–3GalNAca1-
(Ta) and Tn specific lectins
Artocarpus integrifolia agglutinin (Jacalin) (Figs. 10–12)
The OS of the photoreceptor layer were moderately stained
by Jacalin in the retinas of rat and dog, and strongly stained
in the pig retina. The IS were weakly stained by Jacalin in
the three mammalian retinas. There was weak Jacalin
Table 1 Summary of the lectins used in this study [4, 5, 9, 10]
No. Lectins Sources Carbohydrate specificity
(expressed by mammalian
disaccharides)
Polyvalent glycotopes as inhibitors
1 Morniga M Morus nigra mII oligomannosyl residues >>Tri-Man oligomer > Penta-Man
oligomer ‡ Mana1–2,3,or 6 Man
Asialo bovine a1-acid gp (mII)
2 VVL-B4 Vicia villosa isolectin-B4 Tn mainly Asialo OSM (Tn)
3 Jacalin Artocarpus integrifolia T > Tn >>> I(II) Asialo PSM (Tn, Ta, Ah, H) and Asialo human
a1-acid gp (mII)
4 MPL Maclura pomifera T > Tn Asialo PSM (Tn, Ta, Ah, H)
5 ABA Agaricus bisporus T, Tn > I/II > L Asialo PSM (Tn, Ta, Ah, H) and Asialo human
a1-acid gp (mII)
6 ACL Amaranthus caudatus T, Tn > I/II Asialo PSM (Tn, Ta, Ah, H)
7 PNA Arachis hypogea T >>> I/II Asialo fetuin (T, II)
8 Ricin Ricinus communis toxin T > I/II > E > B > Tn Asialo fetuin (T, II)
9 APA Abrus precatorius T > I/II > E > B > Tn Asialo fetuin (T, II)
10 WGA Triticum vulgaris I/II Asialo human a1-acid gp (mII)
11 ECL Erythrina cristagalli mI/II, A and mII clusters,
F > A > mII >> IIAsialo human a1-acid gp (mII)
12 RCA1 Ricinus communis II > I > B > T >> Tn Asialo human a1-acid gp (mII)
13 Abrin-a Abrus precatorius toxin-a E > I/II, L Asialo Bird nest gp (II, E, Ta, Fa)
14 GSI-A4 Griffonia simplicifolia-A4 F > A > and Tn clusters > E > B > I > T >> L Cyst Beach phenol insoluble (B)
15 GSI-B4 Griffonia simplicifolia-B4 B > E Cyst Beach phenol insoluble (B)
16 AAA Anguilla anguilla Ah, Bh, H Hog gastric mucin #4 (Ah, H)
Carbohydrate specificity of lectins as expressed by mammalian disaccharide structural units – Fp, Forssman pentasaccharide (GalNAca1–
3GalNAcb1–3Gala1–4Galb1–4Glc); F, GalNAca1–3GalNAc; A, GalNAca1–3Gal; Ah, GalNAca1–3(LFuca1–2)Gal; Tn, GalNAca1-Ser/Thr;
B, Gala1–3Gal; E, Gala1–4Gal; I/II, Galb1–3/4GlcNAcb1-; L, Galb1–4Glc; T, Galb1–3GalNAc; Ta, Galb1–3GalNAca1-; mII, multivalent II
Neurochem Res (2006) 31:619–628 621
123
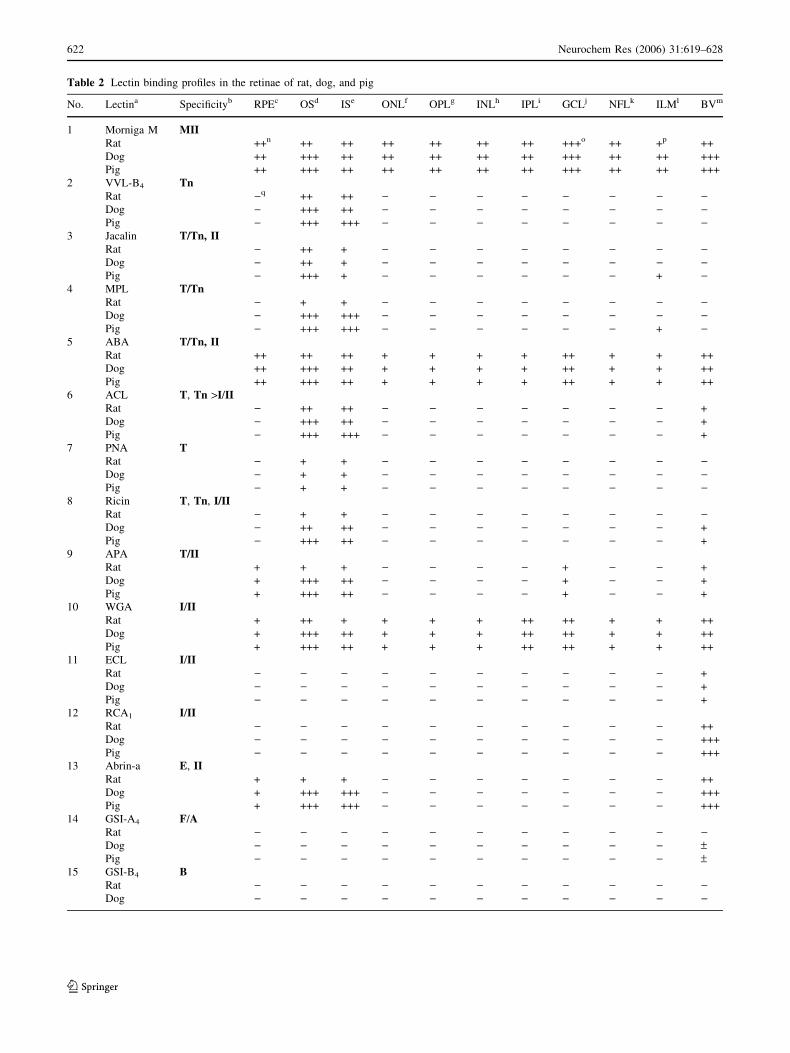
Table 2 Lectin binding profiles in the retinae of rat, dog, and pig
No. Lectina Specificityb RPEc OSd ISe ONLf OPLg INLh IPLi GCLj NFLk ILMl BVm
1 Morniga M MIIRat ++n ++ ++ ++ ++ ++ ++ +++o ++ +p ++
Dog ++ +++ ++ ++ ++ ++ ++ +++ ++ ++ +++
Pig ++ +++ ++ ++ ++ ++ ++ +++ ++ ++ +++
2 VVL-B4 TnRat )q ++ ++ ) ) ) ) ) ) ) )Dog ) +++ ++ ) ) ) ) ) ) ) )Pig ) +++ +++ ) ) ) ) ) ) ) )
3 Jacalin T/Tn, IIRat ) ++ + ) ) ) ) ) ) ) )Dog ) ++ + ) ) ) ) ) ) ) )Pig ) +++ + ) ) ) ) ) ) + )
4 MPL T/TnRat ) + + ) ) ) ) ) ) ) )Dog ) +++ +++ ) ) ) ) ) ) ) )Pig ) +++ +++ ) ) ) ) ) ) + )
5 ABA T/Tn, IIRat ++ ++ ++ + + + + ++ + + ++
Dog ++ +++ ++ + + + + ++ + + ++
Pig ++ +++ ++ + + + + ++ + + ++
6 ACL T, Tn >I/IIRat ) ++ ++ ) ) ) ) ) ) ) +
Dog ) +++ ++ ) ) ) ) ) ) ) +
Pig ) +++ +++ ) ) ) ) ) ) ) +
7 PNA TRat ) + + ) ) ) ) ) ) ) )Dog ) + + ) ) ) ) ) ) ) )Pig ) + + ) ) ) ) ) ) ) )
8 Ricin T, Tn, I/IIRat ) + + ) ) ) ) ) ) ) )Dog ) ++ ++ ) ) ) ) ) ) ) +
Pig ) +++ ++ ) ) ) ) ) ) ) +
9 APA T/IIRat + + + ) ) ) ) + ) ) +
Dog + +++ ++ ) ) ) ) + ) ) +
Pig + +++ ++ ) ) ) ) + ) ) +
10 WGA I/IIRat + ++ + + + + ++ ++ + + ++
Dog + +++ ++ + + + ++ ++ + + ++
Pig + +++ ++ + + + ++ ++ + + ++
11 ECL I/IIRat ) ) ) ) ) ) ) ) ) ) +
Dog ) ) ) ) ) ) ) ) ) ) +
Pig ) ) ) ) ) ) ) ) ) ) +
12 RCA1 I/IIRat ) ) ) ) ) ) ) ) ) ) ++
Dog ) ) ) ) ) ) ) ) ) ) +++
Pig ) ) ) ) ) ) ) ) ) ) +++
13 Abrin-a E, IIRat + + + ) ) ) ) ) ) ) ++
Dog + +++ +++ ) ) ) ) ) ) ) +++
Pig + +++ +++ ) ) ) ) ) ) ) +++
14 GSI-A4 F/ARat ) ) ) ) ) ) ) ) ) ) )Dog ) ) ) ) ) ) ) ) ) ) –Pig ) ) ) ) ) ) ) ) ) ) –
15 GSI-B4 BRat ) ) ) ) ) ) ) ) ) ) )Dog ) ) ) ) ) ) ) ) ) ) )
622 Neurochem Res (2006) 31:619–628
123

staining to ILM in the pig retina. There was no visible
binding in other layers of the retina.
Maclura pomifera lectin (MPL) (Figs. 13–15)
MPL bound weakly to the OS and IS of photoreceptor
layer of rat retina. However, in the retinas of dog and
pig, MPL showed strong binding at the photoreceptor OS
and IS. There was weak MPL staining at the ILM in the
pig retina. No reactions were found by MPL to other
retinal layers of rat, dog, or pig was found.
Agaricus bisporus agglutinin (ABA) (Figs. 16–18)
Photoreceptor OS and IS were moderately stained by
ABA in rat retina. In the retinas of dog and pig, pho-
toreceptor OS were heavily stained and photoreceptor IS
were moderately stained by ABA. Agaricus bisporus
Table 2 continued
No. Lectina Specificityb RPEc OSd ISe ONLf OPLg INLh IPLi GCLj NFLk ILMl BVm
Pig ) ) ) ) ) ) ) ) ) ) )16 AAA ABH
Rat ) ) ) ) ) ) ) ) ) ) )Dog ) ) ) ) ) ) ) ) ) ) )Pig ) ) ) ) ) ) ) ) ) ) )
a,bAbbreviations of lectins and glycotope are shown in Table 1cRPE=retinal pigment epithelium; dOS=outer segment; eIS=inner segment; fONL=outer nuclear layer; gOPL=outer plexiform layer; hINL=inner
nuclear layer; iIPL=inner plexiform layer; jGCL=ganglion cell layer; kNFL= nerve fiber layer; lILM=internal limiting layer; mBV=blood vessels
Lectin staining intensity: n++, moderate; o+++, strong binding; p+, weak; q), invisible
Figs. 1–15 (1–3) These photos reveal well-preserved retina in
hematoxylin and eosin stain retinal specimen in rat, dog, and pig.
(4–6) Morniga M, a mannose-specific lectin from mulberry tree
(Morus nigra), labels all layers of retina. There is no apparent
difference in the binding pattern of Morniga M among the retinas of
rat, dog, and pig. (7–9) Vicia villosa isolectin-B4 (VVL-B4) intensely
labels photoreceptor outer segments. Photoreceptor inner segments
react with VVL-B4 moderately in the retinas of rat and dog, and
intensely in the retina of pig. (10–12) The outer segments of
photoreceptor layer are moderately stained by Jacalin in the retina of
rat and pig, and strongly stained in pig retina. The inner segments are
weakly stained by Jacalin in the three mammalian retinas. The arrow
in figure 12 indicates weak Jacalin staining to internal limiting
membrane in pig retina. (13–15) Maclura pomifera lectin (MPL)
binds weakly to the outer segments and inner segments of
photoreceptors in rat retina and binds strongly to the outer segments
and inner segments of photoreceptor layer in dog and pig retina. The
internal limiting membrane of pig retina is also weakly stained by
MPL (arrow). Scale bar in figure 15 applies to all figures. Scale
bar=50 lm
Neurochem Res (2006) 31:619–628 623
123

agglutinin labeled RPE, ganglion cells, and blood vessels
moderately. It also weakly labeled cells of the ONL,
OPL, INL, IPL, NFL, and ILM. There was no clear
difference in the binding pattern of ABA among the
retina of rat, dog, and pig.
Amaranthus caudatus lectin (ACL) (Figs. 19–21)
ACL exhibited moderate staining in the OS and IS of the
photoreceptor layer in the rat retina. In the dog retina, ACL
showed strong binding to photoreceptor OS, and moderate
binding to photoreceptor IS. However, ACL bound
strongly to photoreceptor OS and IS in the retina of pig.
ACL also weakly stained the blood vessel wall of retina in
these mammalian retinas. There was no staining in other
retinal layers.
Interaction of retinal glycans with Galb1–3GalNAca1-(Ta)
specific lectin—peanut agglutinin (PNA) (Figs. 22–24)
The PNA lectin preferentially bound to the region of cone
IS and OS. The binding pattern was basically the same in
the retina of rat, dog, and pig. There was no binding of
PNA in the other layers of the retina.
Interaction of retinal glycans with Galb1–3GalNAca1-
(Ta) and Galb1–4GlcNAcb1-(II) specific lectins
Ricinus communis toxin (Ricin) (Figs. 25–27)
Ricin stained weakly photoreceptor IS and OS in the rat
retina. In canine retina, there was moderate staining both in
the region of the photoreceptor OS and IS. In pig retina,
there was strong binding in photoreceptor OS, and mod-
erate staining at the IS region. Ricin labeled weakly blood
vessel walls in the retinas of dog and pig. However, no
Ricin binding to other layers of retina was found.
Abrus precatorius agglutinin (APA) (Figs. 28–30)
APA reacted weakly with RPE, photoreceptor OS and IS in
rat retina. APA also weakly labeled ganglion cell bodies,
and blood vessels in rat retina. There was stronger labeling
in the photoreceptor IS and OS in dog and pig retinas. RPE,
Figs. 16–30 (16–18) Agaricus bisporus agglutinin (ABA) binds to
retinal pigment epithelium, the outer segments and inner segments of
the photoreceptor layer, outer nuclear layer, outer plexiform layer,
inner nuclear layer, inner plexiform layer, ganglion cells layer, blood
vessel walls, nerve fiber layer, and internal limiting membrane. (19–
21) Amaranthus caudatus lectin (ACL) binds preferentially to
photoreceptor outer and inner segments. ACL also faintly stains
blood vessel wall (arrows). (22–24) Peanut agglutinin (PNA)
preferentially binds to the region of cone inner and outer segment
in the retinas of rat, dog, and pig (arrows). (25–27) Ricinus communis
toxin (Ricin) stains photoreceptor inner and outer segments. Ricin
also stains blood vessel walls weakly in the retinas of dog and pig
(arrows). (28–30) Abrus precatorius agglutinin (APA) reacts with
retinal pigment epithelium, photoreceptor inner and outer segments,
ganglion cells (arrows), and blood vessels (asterisk) in the three
mammalian retinas. Scale bar in figure 30 applies to all figures. Scale
bar=50 lm
624 Neurochem Res (2006) 31:619–628
123

ganglion cell bodies, and vascular endothelium cells were
also weakly labeled by APA in dog and pig retinas.
Interaction of retinal glycans with Galb1–3/
4GlcNAcb1-(I/II) specific lectins
Wheat germ agglutinin (WGA) (Figs. 31–33)
WGA bound strongly to the OS region of photoreceptor
cells, and moderately to IPL, ganglion cell bodies, and
blood vessels. WGA also bound weakly to RPE, ONL,
OPL, INL, NFL, and ILM. Except for weaker WGA binding
of photoreceptor OS and IS, the binding pattern of WGA in
rat retina was similar to dog and pig retina.
Erythrina cristagalli lectin (ECL) (Figs. 34–36)
ECL reacted weakly with blood vessel walls in the three
mammalian retinas. There was no ECL binding in other
layers of the retina.
Ricinus communis agglutinin (RCA1) (Figs. 37–39)
Retinal vessels were moderately stained by RCA1 in rat,
and intensely stained in dog and pig. There was no staining
in other structures of the retinas.
Interaction of retinal glycans with Gala1–4Gal (E)
and II specific lectin—Abrus precatorius toxin-a
(Abrin-a) (Figs. 40–42)
Abrin-a weakly labeled RPE and photoreceptor OS and IS,
and moderately labeled blood vessels walls of rat retina. In
the retinas of dog and pig, Abrin-a weakly labeled RPE, but
strongly labeled photoreceptor IS and OS and blood vessels
walls. The binding pattern of Abrin-a among rat, dog, and
pig was quite similar.
Interaction of retinal glycans with GalNAca1–3Gal/
GalNAc (A/F) specific lectin-Griffonia simplicifolia
I-A4 (GSI-A4), Gala1–3Gal (B) specific lectin-G.
simplicifolia I-B4 (GSI-B4) and LFuca1–2Gal (H)
specific lectin-Anguilla anguilla agglutinin (AAA)
GSI-A4 labeled blood vessel walls weakly to none in the
retinas of dog, pig and other layers of the three mammalian
retinas. Similarly, GSI-B4 and AAA did not react with any
structures of the three mammalian retinas (figure not
shown).
Inhibition of lectin–retinal glycan interactions
by glycoproteins and monosaccharides
To confirm that the observed binding of the lectins was not
due to non-specific adsorption or simply tissue entrapment
of a smaller molecular weight lectin, various glycoproteins
with known polyvalent carbohydrate structures and termi-
nal glycotopes were used to block lectin–carbohydrate
interactions. With all of the lectins, binding was totally
inhibited or markedly reduced (WGA) by polyvalent gly-
coproteins. For instance, in the case of Galb1–3GalNAca1
(T)-specific lectin ACL, the addition of T/Tn-containing
glycoprotein, such as asialo PSM, completely abolished its
binding. Nevertheless, inhibition with glycoproteins con-
taining other glycotopes, for example blood group A+H
[GalNAca1–3(LFuca1–2)Gal]-active glycoprotein from
hog gastric mucin #4 or cyst Beach phenol insoluble (blood
group B active gp) did not diminish the binding intensity of
Figs. 31–42 (31–33) Wheat
germ agglutinin (WGA) labels
all layers of retina and the blood
vessel walls. (34–36) Erythrina
cristagalli (ECL). ECL reacted
weakly with blood vessel walls
(arrows) in the retina. (37–39)
Ricinus communis (RCA1)
heavily stains retinal vessels in
rat, dot, and pig retina (arrows).
(40–42) Abrus precatorius
toxin-a (Abrin-a) labels retinal
pigment epithelium and
photoreceptor outer and inner
segments, and blood vessel
walls (arrows) obviously in the
three mammalian retinas
Neurochem Res (2006) 31:619–628 625
123

ACL. In addition, adding monosaccharides (Gal, GalNAc,
methyl a/b-Gal) or disaccharide (Galb1–4Glc) as inhibitors
did not abolish ACL staining. These results confirmed the
specific interaction of each lectin with the carbohydrates
within the retinal tissue.
Discussion
Lectins are carbohydrate-binding proteins of non-immune
origin [11, 12]. Those lectins that can be used as tools to
study glycobiological systems are defined as ‘applied’
lectins [9, 11–15]. Many have been used to study the dis-
tribution of glycoconjugates in normal [2, 12, 14, 16–23]
and diseased retinas [24–29]. However, most of these
studies used only one animal species and focused only on
part of the retina, such as the interphotoreceptor matrix and
photoreceptors. Information on the lectin binding patterns
of other retinal layers was not well organized and limited in
scope. Moreover, interpretation of these results was based
on monosaccharides, rather than on disaccharide structural
units or polyvalent glycotopes [30, 31], which may lead to
a false-conclusion [4]. Therefore, it would be much more
relevant to use both oligosaccharide and polyvalent gly-
cotopes present in macromolecules rather than monosac-
charides to confirm carbohydrate–lectin interactions,
especially for Agaricus bisporus agglutinin (ABA), which
is not easily inhibited by monosaccharides. In this study,
three representative mammalian retinal neurons were
studied.
The possible, distinct carbohydrate sequences and their
specified distribution in retinal layers were defined in this
study. The knowledge of carbohydrates has important
implications on the future retinal biology and pathology.
Retinal detachment (RD), for example, is an important
cause of severe visual loss and is a common cause of
blindness after cataract surgery [32, 33]. The formation of
RD is due to separation of photoreceptors from the
underlying RPE. Carbohydrate structures in the junction of
retina and RPE may play valuable role in the maintenance
of the retinal adhesion to its underlying RPE structure. In
previous study, glycoconjugates are identified in IPM, of
which the major structural component is chondroitin 6-
sulfate glycosaminoglycan, a form of proteoglycan [34].
Glycans on IPM closely adhere to cone photoreceptors and
to the RPE and sustain the retinal attachment [35, 36], and
inhibition of proteoglycan synthesis in IPM results in RD
[36]. Therefore, glycoconjugates on the IPM-related
structures and their exfoliate substances which undergoes
glycosylation change during RD could be a good marker
predicting the prognosis of the disease or the mediators
facilitating the progression of this disease. In addition,
carbohydrate also plays some role during retinal develop-
ment in the embryonic stage [37], and also involves path-
ologic conditions of the eye such as age-related macular
degeneration [38], and retinoblastoma [39]. The knowledge
of the carbohydrate in the retina may help elucidate the
pathogenesis of those diseases.
Our study finds retinal binding patterns of lectins with
similar specificities are not completely identical. For
instance, Jacalin and Peanut lectin have similar carbo-
hydrate specificities, but they show different binding
patterns in retina. One possible explanation is that Jac-
alin binds more strongly with T/Tn glycotopes present in
glycoproteins, while PNA shows high affinity for T and
II antigens. Moreover, for the proportional contributions
of individual residues of T disaccharides (Gal and Gal-
NAc of Galb1–3GalNAc), it has been reported that Gal
> GalNAc in Peanut, but GalNAc > Gal in Jacalin [5,
40]. This could be the key reason for the differences in
binding pattern between Jacalin and Peanut.
In most cases, the concentration of lectin can affect
the binding of lectins toward small amounts and low
affinity types of glycotopes present on macromolecules
[10]. The lectin binding profile in the retina at a very
high concentration of lectin can be different from those
at a low lectin concentration (unpublished data). When
the lectin concentration was increased, binding of lectin
to small quantities and poor affinity types of glycotopes
present on the macromolecules were shown. For differ-
ential purposes, optimal amounts of lectin were selected.
Only a major or dominant carbohydrate glycotope was
demonstrated in the retinal neurons. An area without
lectin staining reflects the fact that there is no or very
little carbohydrate present in that area in comparison to
an area with strong binding. Under this condition, the
binding of lectins in the retinal neurons could be com-
pletely inhibited by polyvalent inhibitory glycotopes
present on the macromolecules, which rules out non-
specific interactions.
In conclusion, our study demonstrates that: (i) the retina
of rat, dog, and pig are specifically stained by different
lectins, indicating that binding to the retinal structures
depends on the sugar-binding specificities of the different
lectins, showing the presence of different glycoconjugates
in different parts of the retina i.e. each layer had its own
binding specificity; (ii) all sections of the retinas reacted
well with Morniga M, indicating N-linked glycans are
present in all retinal layers; (iii) no detectable human blood
group ABH active glycotopes were found in the layers of
the retinas; (iv) outer and inner segments contained glyco-
conjugates rich in ligands reacting with Ta (Galb1–3Gal-
NAca1-Ser/Thr) and Tn (GalNAca1-Ser/Thr) specific
lectins; (v) the cone cells of the retina bound specifically to
peanut agglutinin (PNA) which mainly recognizes Ta
residues could be used as a specific marker for these
626 Neurochem Res (2006) 31:619–628
123

photoreceptor cells; and (vi) the binding intensity of retina
in the rat, dog and pig was similar, but with different
intensity. This information might be useful to understand
the structural and functional role of carbohydrates in the
retina of the eye and its related pathogenesis.
Acknowledgements This study was supported by grants from Na-
tional Science Council, Taiwan (NSC 94-2320-B-182-044, NSC 94-
2320-B-182-053). This work was also supported by grants from the
Chang-Gung Medical Research Project (CMRPD no. 33022, Kwei-
san, Tao-yuan, Taiwan) and grants from the Fund for Scientific
Research-Flanders (to E.J.M. Van Damme).
References
1. Blanks JC (2001) Morphology and topography of the retina. In:
Ryan SJ, Odgen TE, Hinton DR (eds) Basic science and inherited
retinal disease. Mosby, St. Louis, pp 32–53
2. Silver FH, Benedetto D (1996) Polysaccharides used in
ophthalmology. In: Dimitriu S (ed) Polysaccharides in medicine
and biotechnology Marcel Dekker, New York, pp 689–703
3. Lazarus HS, Hageman GS (1992) Xyloside-induced disruption of
interphotoreceptor matrix proteoglycans results in retinal
detachment. Invest Ophthalmol Vis Sci 33:364–376
4. Wu AM (2001) Expression of binding properties of Gal/GalNAc
reactive lectins by mammalian glycotopes. Adv Exp Med Biol
491:55–64
5. Wu AM, Song SC, Tsai MS, Herp A (2001) A guide to the
carbohydrate specificities of applied lectins-2. Adv Exp Med Biol
491:551–585
6. Wu AM, Wu JH, Singh T, Chu KC, Peumans WJ, Rouge P, Van
Damme EJM (2004) A novel lectin (Morniga M) from mulberry
(Morus nigra) bark recognizes oligomannosyl residues in
N-glycans. J Biomed Sci 11:874–885
7. Sarkar M, Wu AM, Kabat EA (1981) Immunochemical studies on
the carbohydrate specificity of Maclura pomifera lectin. Arch
Biochem Biophys 209:204–218
8. Duk M, Lisowska E, Wu JH, Wu AM (1994) The biotin/avidin-
mediated microtiter plate lectin assay with the use of chemically
modified glycoprotein ligand. Anal Biochem 221:266–272
9. Wu AM (2003) Carbohydrate structural units in glycoproteins
and polysaccharides as important ligands for Gal and GalNAc
reactive lectins. J Biomed Sci 10:676–688
10. Wu AM (2004) Polyvalency of Tn (GalNAca fi Ser/Thr) gly-
cotope as a critical factor for Vicia villosa B4 and glycoprotein
interactions. FEBS Lett 562:51–58
11. Gabius HJ, Gabius S (1993) Lectins as tools for the character-
ization of glycoconjugates and lectins and neoglycoconjugates in
histochemical and cytochemical analysis. In: Gabius HJ, Gabius
S (eds) Lectins and glycobiology. Springer, Berlin, pp 141–187
and 211–326
12. Sharon N, Lis H (eds) (2003) Lectins. Kluwer Academic Pub-
lishers, Dordrecht, Boston, London
13. Hakomori S, Kannagi R (1986) Handbook of experimental
immunology. In: Weir DM (ed) Immunochemistry. Blackwell
Scientific Oxford, London and Boston, pp 9.1–9.39
14. Sameshima M, Uehara F, Ohba N (1987) Specialization of the
interphotoreceptor matrices around cone and rod photoreceptor
cells in the monkey retina, as revealed by lectin cytochemistry.
Exp Eye Res 45:845–863
15. Wu AM, Sugii S (1988) Differential binding properties of Gal-
NAc and/or Gal specific lectins. Adv Exp Med Biol 228:205–263
16. Bishop PN, Boulton M, McLeod D, Stoddart RW (1993) Glycan
localization within the human interphotoreceptor matrix and
photoreceptor inner and outer segments. Glycobiology 3:403–412
17. Blanks JC, Johnson LV (1983) Selective lectin binding of the
developing mouse retina. J Comp Neurol 221:31–41
18. Bopp S, el-Hifnawi ES, Laqua H (1992) Lectin binding pattern in
human retinal pigment epithelium. Anat Anz 174:279–285
19. Kivela T, Tarkkanen A (1987) A lectin cytochemical study of
glycoconjugates in the human retina. Cell Tissue Res 249:277–
288
20. Koide H, Suganuma T, Murata F, Ohba N (1986) Ultrastructural
localization of lectin receptors in the monkey retinal photore-
ceptors and pigment epithelium: application of lectin-gold com-
plexes on thin sections. Exp Eye Res 43:343–354
21. McLaughlin BJ, Boykins LG (1984) Lectin cytochemistry and
freeze-fracture study of phagocytosis in the rat retina. J Comp
Neurol 223:77–87
22. Uehara F, Ohba N, Sameshima M, Unoki K, Okubo A, Yanagita
T, Sugata M, Iwakiri N, Nakagawa S (1994) Binding of ama-
ranthin in photoreceptors of monkey retina. Jpn J Ophthalmol
38:360–363
23. Yan Q, Bumsted K, Hendrickson A (1995) Differential peanut
agglutinin lectin labeling for S and L/M cone matrix sheaths in
adult primate retina. Exp Eye Res 61:763–766
24. Lazarus HS, Sly WS, Kyle JW, Hageman GS (1993) Photore-
ceptor degeneration and altered distribution of interphotoreceptor
matrix proteoglycans in the mucopolysaccharidosis VII mouse.
Exp Eye Res 56:531–541
25. Long KO, Aguirre GD (1991) The cone matrix sheath in the
normal and diseased retina: cytochemical and biochemical stud-
ies of peanut agglutinin-binding proteins in cone and rod-cone
degeneration. Exp Eye Res 52:699–713
26. McLaughlin BJ, Wood JG (1980) The localization of lectin
binding sites on photoreceptor outer segments and pigment epi-
thelium of dystrophic retinas. Invest Ophthalmol Vis Sci 19:728–
742
27. Mieziewska K, Van Veen T, Aguirre GD (1993) Development
and fate of interphotoreceptor matrix components during dys-
plastic photoreceptor differentiation: a lectin cytochemical study
of rod-cone dysplasia 1. Exp Eye Res 56:429–441
28. Mieziewska K, Van Veen T, Aguirre GD (1993) Structural
changes of the interphotoreceptor matrix in an inherited retinal
degeneration: a lectin cytochemical study of progressive rod-cone
degeneration. Invest Ophthalmol Vis Sci 34:3056–3067
29. Kivela T 1987 Glycoconjugates in retinoblastoma. A lectin his-
tochemical study of ten formalin-fixed and paraffin-embedded
tumours. Virchows Arch A Pathol Anat Histopathol 410:471–479
30. Lee RT, Lee YC (2000) Affinity enhancement by multivalent
lectin-carbohydrate interaction. Glycoconj J 17:543–551
31. Wu AM, Wu JH, Herp A, Liu JH (2003) Effect of polyvalencies
of glycotopes on the binding of a lectin from the edible mush-
room, Agaricus bisporus. Biochem J 371:311–320
32. Lewis H (2003) Peripheral retinal degenerations and the risk of
retinal detachment. Am J Ophthalmol 136:155–160
33. The Eye Disease Case-Control Study Group (1993) Risk factors
for idiopathic rhegmatogenous retinal detachment. Am J Epi-
demiol 137:749–757
34. Hageman GS, Johnson LV (1987) Chondroitin 6-sulfate glycos-
aminoglycan is a major constituent of primate cone photoreceptor
matrix sheaths. Curr Eye Res 6:639–646
35. Hollyfield JG, Varner HH, Rayborn ME (1990) Regional varia-
tion within the interphotoreceptor matrix from fovea to the retinal
periphery. Eye 4:333–339
36. Lazarus HS, Hageman GS (1992) Xyloside-induced disruption
of interphotoreceptor matrix proteoglycans results in retinal
detachment. Invest Ophthalmol Vis Sci 33:364–736
Neurochem Res (2006) 31:619–628 627
123

37. Mintz G, Gottlieb DI, Reitman ML, et al. (1981) Developmental
changes in glycoproteins of the chick nervous system. Brain Res
206:51–70
38. Russell SR, Mullins RF, Schneider BL, Hageman GS (2000)
Location, substructure, and composition of basal laminar drusen
compared with drusen associated with aging and age-related
macular degeneration. Am J Ophthalmol 129:205–214
39. Yamashita T, Uehara F, Ozawa M, Ohba N (2002) Further
characterization of human mucin-like glycoprotein associated
with photoreceptor cells by its introduction into Y79 retinoblas-
toma cells. Ophthalmic Res 34:70–76
40. Wu AM, Wu JH, Lin LH, Lin SH, Liu JH (2003) Binding profile
of Artocarpus integrifolia agglutinin (Jacalin). Life Sci 72:2285–
2302
628 Neurochem Res (2006) 31:619–628
123
Copyright © 2022 FDOKUMEN




















