
Different Binding of NF-Y Transcriptional Factor to DQA1 Promoter Variants
10
Different Binding of NF-Y Transcriptional Factor to DQA1 Promoter Variants Paola Indovina, Francesca Megiorni, Paola Ferrante, Ilaria Apollonio, Fiorella Petronzelli, and Maria Cristina Mazzilli ABSTRACT: Polymorphism in the HLA-DQA1 pro- moter (QAP) sequences could influence the gene expres- sion through a differential binding of transcriptional fac- tors. Considering the main role played by the Y-box in the transcription, we focused on the QAP4 variants dif- fering for a G vs A transition from the QAP Y-box consensus sequence. Electrophoretic Mobility Shift Assay using the two Y-box sequences was performed to deter- mine whether this mutation could be reflected in an allele-specific binding of transcriptional factors. Indeed, the NF-Y specific band, recognised by supershift experi- ments, was clearly observed using the Y-box consensus probe but it was barely detectable with the QAP4 one. On the contrary, two other complexes were found to more strongly interact with QAP4 Y-box in comparison to the consensus sequence. The analysis of a selected panel of HLA homozygous lymphoblastoid cell lines by competitive RT-PCR and by Northern blotting revealed that the DQA1 *0401, *0501,*0601 alleles regulated by the QAP4 promoters were less expressed at the mRNA level than the DQA1* 0201 allele regulated by the QAP2.1 variant. In conclu- sion, these results show an evident reduction of NF-Y binding to the mutated QAP4 Y-box and a decreased mRNA accumulation of the DQA1 alleles regulated by these variants. Human Immunology 59, 758 –767 (1998). © American Society for Histocompatibility and Immu- nogenetics, 1998. Published by Elsevier Science Inc. ABBREVIATIONS QAP DQA1 promoter variant RT-PCR reverse transcriptase-polymerase chain reaction MHC major histocompatibility complex IFN-g interferon-gamma TRE TPA response element CRE cAMP response element EMSA electrophoretic mobility shift assay IHW international histocompatibility workshop SSOP sequence-specific oligo-probing APC antigen presenting cells INTRODUCTION Polymorphism of the Major Histocompatibility Complex (MHC) class II genes leads to the formation of different molecules able to bind a wide range of antigenic peptides and to present them to CD41 T lymphocytes. Consti- tutive expression of these genes is limited to the antigen presenting cells which include macrophages, B lympho- cytes, dendritic cells and thymic epithelial cells [1]. A multiplicity of external signals, of which IFN-g is the most potent, can induce class II gene expression render- ing most cell types immunocompetent [2]. Aberrant expression of these antigens is associated with immune dysfunction in many neoplastic diseases or in Bare Lym- phocytes Syndrome where the patients are unable to mount a specific immune response to soluble antigens [3–5]. All class II gene promoters share cis-acting elements, called W, X1, X2 (similar to TRE/CRE sequences) and Y boxes, which are responsible for the control of their expression [1]. Furthermore, W and X boxes confer B-cell specificity and are involved in IFN-g-induction. Nuclear factors binding to these sequences have been identified and some of them have been cloned. In par- ticular RF-X complexes binds to the X1 box while NF-S From the Department of Experimental Medicine and Pathology, “La Sapienza” University of Rome, Italy Address reprint requests to: Prof. M.C. Mazzilli, Department of Exper- imental Medicine and Pathology, “La Sapienza” University, Viale Regina Elena, 324 00161 Roma, Italy; Tel: 139-064451286 Fax: 139- 064454820, E-Mail: [email protected]. Received June 18, 1998; accepted August 4, 1998. Human Immunology 59, 758 –767 (1998) 0198-8859/98/$–see front matter © American Society for Histocompatibility and Immunogenetics, 1998 Published by Elsevier Science Inc. PII S0198-8859(98)00082-2
Transcript of Different Binding of NF-Y Transcriptional Factor to DQA1 Promoter Variants

Different Binding of NF-Y TranscriptionalFactor to DQA1 Promoter Variants
Paola Indovina, Francesca Megiorni, Paola Ferrante,Ilaria Apollonio, Fiorella Petronzelli, andMaria Cristina Mazzilli
ABSTRACT: Polymorphism in the HLA-DQA1 pro-moter (QAP) sequences could influence the gene expres-sion through a differential binding of transcriptional fac-tors. Considering the main role played by the Y-box inthe transcription, we focused on the QAP4 variants dif-fering for a G vs A transition from the QAP Y-boxconsensus sequence. Electrophoretic Mobility Shift Assayusing the two Y-box sequences was performed to deter-mine whether this mutation could be reflected in anallele-specific binding of transcriptional factors. Indeed,the NF-Y specific band, recognised by supershift experi-ments, was clearly observed using the Y-box consensusprobe but it was barely detectable with the QAP4 one. Onthe contrary, two other complexes were found to more
strongly interact with QAP4 Y-box in comparison to theconsensus sequence.
The analysis of a selected panel of HLA homozygouslymphoblastoid cell lines by competitive RT-PCR and byNorthern blotting revealed that the DQA1 *0401,*0501,*0601 alleles regulated by the QAP4 promoterswere less expressed at the mRNA level than the DQA1*0201 allele regulated by the QAP2.1 variant. In conclu-sion, these results show an evident reduction of NF-Ybinding to the mutated QAP4 Y-box and a decreasedmRNA accumulation of the DQA1 alleles regulated bythese variants. Human Immunology 59, 758–767 (1998).© American Society for Histocompatibility and Immu-nogenetics, 1998. Published by Elsevier Science Inc.
ABBREVIATIONSQAP DQA1 promoter variantRT-PCR reverse transcriptase-polymerase chain
reactionMHC major histocompatibility complexIFN-g interferon-gammaTRE TPA response element
CRE cAMP response elementEMSA electrophoretic mobility shift assayIHW international histocompatibility
workshopSSOP sequence-specific oligo-probingAPC antigen presenting cells
INTRODUCTIONPolymorphism of the Major Histocompatibility Complex(MHC) class II genes leads to the formation of differentmolecules able to bind a wide range of antigenic peptidesand to present them to CD41 T lymphocytes. Consti-tutive expression of these genes is limited to the antigenpresenting cells which include macrophages, B lympho-cytes, dendritic cells and thymic epithelial cells [1]. Amultiplicity of external signals, of which IFN-g is the
most potent, can induce class II gene expression render-ing most cell types immunocompetent [2]. Aberrantexpression of these antigens is associated with immunedysfunction in many neoplastic diseases or in Bare Lym-phocytes Syndrome where the patients are unable tomount a specific immune response to soluble antigens[3–5].
All class II gene promoters share cis-acting elements,called W, X1, X2 (similar to TRE/CRE sequences) andY boxes, which are responsible for the control of theirexpression [1]. Furthermore, W and X boxes conferB-cell specificity and are involved in IFN-g-induction.Nuclear factors binding to these sequences have beenidentified and some of them have been cloned. In par-ticular RF-X complexes binds to the X1 box while NF-S
From the Department of Experimental Medicine and Pathology, “LaSapienza” University of Rome, Italy
Address reprint requests to: Prof. M.C. Mazzilli, Department of Exper-imental Medicine and Pathology, “La Sapienza” University, Viale ReginaElena, 324 00161 Roma, Italy; Tel: 139-064451286 Fax: 139-064454820, E-Mail: [email protected].
Received June 18, 1998; accepted August 4, 1998.
Human Immunology 59, 758–767 (1998)0198-8859/98/$–see front matter© American Society for Histocompatibility and Immunogenetics, 1998
Published by Elsevier Science Inc. PII S0198-8859(98)00082-2

and X2BP occupy the X2 motif [5–7]. The Y boxelement, an inverted CCAAT sequence, is the target ofthe ubiquitously expressed heterotrimeric protein NF-Ythat is involved in the formation of the transcriptionalinitiation complex and is essential to stabilize the inter-actions of the nuclear factors with X and W boxes[8–11]. Another Y box binding factor is YB-1 thatseems to be a negative regulatory protein of class II genes[12, 13]. Transcriptional regulation also requires theclass II transactivator (CIITA) factor, which probablydoes not bind DNA directly but functions by interactingwith the other regulatory proteins [14, 15].
Despite the important role of these consensus regula-tory motifs, a certain degree of variability exists amongthe promoters of the class II genes and it has beenhypothesized that a differential binding of trans-actingfactors to these sequences could be at the basis of theexpression gradient reported with DR . DP . DQ[16–18]. The variability of the X boxes has already beenrelated to a different binding affinity of RF-X and NF-Stranscriptional factors [19, 20]. Furthermore, a transitionG vs A at position 2122 in the Y box of the DQA1promoter has been suggested to cause a minor interactionof the NF-Y nuclear factor and consequently a lowerexpression of DQA1 respect to DRA gene [21].
An intra-locus polymorphism in the control region ofmost of class II genes has been also described [22–27].These findings have raised the possibility that a differ-ential expression could exist among structural allelesregulated by different promoters. A different transcrip-tional efficiency of some promoter sequences has beendemonstrated by transient expression assays in the DR,DQB1 and DQA1 genes [28–30]. In particular, muta-tions in the Y box have been shown to have a great effecton the promoter strength of the DQA1 gene [30].
We focused on the Y-box of the DQA1 promotervariants (QAP) in order to analyze the influence of the Gvs A transition present in the CCAAT sequence of theQAP4 alleles (QAP4.1 and QAP4.2) on DNA-proteininteractions (Fig. 1). We evaluated by ElectrophoreticMobility Shifth Assay (EMSA) the effect of this single-site mutation on NF-Y binding efficiency. Furthermore,we compared by competitive RT-PCR and Northern blotassays the mRNA levels of the DQA1 alleles regulatedby the QAP4 variants to those of QAP2.1 positive celllines, representative of the X and Y boxes consensussequence.
MATERIALS AND METHODSCell Lines and Culture ConditionsA panel of human lymphoblastoid cell lines of 10thInternational Histocompatibility Workshop (IHW) wasmaintained in culture in RPMI 1640 medium supple-
mented with 10% fetal calf serum. The QAP polymor-phism was determined during the 12th IHW by se-quence-specific oligo-probing (SSOP). The panelincluded 11 HLA homozygous cell lines: five QAP 2.1(PITOUT, LBF, IBW9, MANN and MOU typed asDRB1*07-DQA1*0201-DQB1*02); four QAP 4.1(JBUSH and JVM, typed as DRB1*11-DQA1*0501-DQB1*0301, HERLUF, typed as DRB1*1201-DQA1*0501-DQB1*0301 and WT49 typed as DRB1*0301-DQA1*0501-DQB1*0201); two QAP 4.2 (OLL typedas DRB1*08022-DQA1*0401- DQB1*0402 and LUYtyped as DRB1*08032-DQA1*0601-DQB1*0301).
Electrophoretic Mobility Shift Assay
Nuclear extract was prepared according to Dignam et al.[31], except for the following modifications: proteaseinhibitors (0.5 mM PMSF and 5 mg/ml leupeptin) wereadded to all solutions; the pellet crude nuclei resultingfrom the centrifugation at 25000 g for 20 min wassuspended in 1 M NaCl, 0.5 mM MgCl2, 0.2 mMEDTA, 20 mM HEPES (pH 7.9), 25% glycerol, 0.5 mMDTT [32]. Final protein concentration was determinedaccording to manufacturer’s instructions (Bio-Rad). Nu-clear extract was divided into aliquots and stored at280°C.
Double-stranded DNA probes (Table 1) were pre-pared by annealing complementary oligonucleotides ex-cept for a 59 five-base single stranded tail. Labeling wasperformed by filling in these tails with Klenow DNApolymerase, [a-32P]dATP and the other deoxynucleosidetriphosphates unlabelled.
EMSA was carried out as described by Dorn et al. [32]with some modifications: a volume of 20 ml consisting in0.25 ng of labeled DNA probe (about 10000 cpm),0.2–0.4 mg of poly(dI-dC) to reduce non-specific proteinbinding, 5 mg of nuclear extract, 10 mM Tris-HCl pH7.1, 0.1 mM EDTA, 0.1% TritonX-100, 10 mM 2-mer-captothanol, 5% glycerol, 50 mM NaCl, 3 mM MgCl2,0.5 mM PMSF was incubated at 0°C for 1 h. Thespecificities of DNA-protein complexes were analyzed bycompetition experiments in which unlabelled specificand no specific competitors (50 and 400 fold excess toprobe) were added to the reaction mix. For supershiftexperiments 0.1 mg of the YB polyclonal antibody spe-cific for the B subunit of NF-Y [8] was added to reactionmix. An anti-b2-microglobulin monoclonal antibody(X 3.1) was used as control. A 5% polyacrylamide gel(acrylamide:bisacrylamide ratio of 29:1) was pre-runnedin 0.5 3 TBE (45 mM Tris-Borate, 1mM EDTA) at 200V for at least 90 min. The mix was loaded onto the geland electrophoresed for 75 min, dried and autoradio-graphed.
759NF-Y Binding to DQA1 Promoters

Northern BlottingCytoplasmic RNA from the panel of cell lines was pre-pared according to Chomczynski and Sacchi [33]. 20 mgof each RNA were electrophoresed in 1% agarose/form-aldehyde gel and transferred to nylon membranes (Boehr-inger-Mannhein) in 10x SSC.
cDNA probes specific for human HLA-DQA1 se-quence and for human b-actin gene (kindly provided byA. De Lerma) were radiolabeled with [a-32P]dCTP byrandom priming method (MegaprimeTM DNA labelingsystems RPN 1604, Amersham). Probes were added to
membrane at a concentration of 5 3 106 cpm/ml. Pre-hybridization and hybridization were performed accord-ing to recommended protocol of Rapid-hyb buffer(Amersham).
Quantitative Analysis by Competitive-PCR Method
cDNA was synthesized by 50U MuLV reverse transcrip-tase (Perkin-Elmer Cetus) using oligo dT 2.5 mM asprimer and 1 mg of total RNA in 10 mM Tris-HCl (pH8.3), 50 mM KCl, 5 mM MgCl2, 0.2 mM dNTPs, 20 URNase inhibitor (Perkin-Elmer Cetus). One cycle of am-plification consisting in 15 min at 42°C, 5 min at 99°C,5 min at 5°C was carried out using DNA Thermal Cycler480 (Perkin Elmer Cetus).
The DQA1 and b-actin competitive RT-PCRs wereperformed as previously described [34, 35]. Two coupleof primers for each gene were chosen, the external pairamplifying the target sequence and the internal primers,designed with 20 bp long tails complementary to each
FIGURE 1 Sequence alignement of three DQA1 promotervariants regulating the DQA1*0201 (QAP2.1), DQA1*0501(QAP4.1) and DQA1*0401 or *0601 (QAP4.2) structuralalleles. Nucleotide sequence identities are marked by dashes.Promoter sequences and designation of the major putativeboxes are as previously reported [24]. Regarding the Y-box,the locus-specific mutation is indicated by a dash and thepolymorphic position by bold typeface.
760 P. Indovina et al.

other, used for the construction of the competitive tem-plate (Table 2).
The coamplification of the cDNA to be tested and ofthe competitor DNA was carried out in several reactiontubes adding the same aliquot of target DNA and in-creasing known amounts of competitive sequence. ThePCR conditions for both genes consisted in 40 cycles runat 94°C for 40 s, 58°C for 30 s and 72°C for 30 s withreagents at standard concentration. The amplified prod-ucts of DQA1 (252–272 bp) and b-actin genes (227–247bp) were then electrophoresed onto 8% polyacrilamidegels.
The band intensities of target and competitive prod-ucts were measured by a densitometric analyzer and theratio of the two values was considered for each lane.These ratios were then plotted versus the amount ofcompetitor added in each tube and the correspondingpoints fitted by a straight line. In this way it was possibleto mathematically derive the amount of competitor cor-responding to a 1:1 ratio, equivalent to the amount ofinput cDNA in the sample [34].
Statistical AnalysisThe results obtained by competitive RT-PCR on the twogroups of QAP2.1 and QAP4 cell lines were analyzed byStudent’s t test for not appaired data. p 5 0.05 was takenas the upper limit of significance.
RESULTSNF-Y Binding Affinity Follows a QAP4 ,, QAPConsensus < DRA GradientIn order to establish if the mutation present in the Y-boxof QAP4 variants could influence the binding of tran-scriptional factors, double-stranded oligonucleotideprobes, Y-QAP4 and Y-QAPc representative of the Y-box consensus, were used in EMSA experiments. Thedifferent patterns observed using the two probes arereported in Fig. 2A. Among the three specific bands,those called A and B were observed with both probes,while the band corresponding to NF-Y (see below) wasonly visible with the Y-QAPc sequence. The NF-Ycomplex was barely detectable with the Y-QAP4 probeonly after prolonged exposure (Fig. 2B). Competitionexperiments, performed in the presence of unlabeleddouble-stranded oligonucleotides of the same sequences,showed that Y-QAP4 inhibited formation of both the Aand B complexes more efficiently than Y-QAPc (Fig. 2B:compare lanes 8 and 10, and lanes 9 and 11). NF-Ycomplex was not competed by Y-QAP4 (lane 5) while itwas weakly inhibited by Y-QAPc (lane 3). Neither of thecomplexes were competed away by an unrelated sequenceshowing the specificity of the interactions of A, B andNF-Y factors with Y-QAP4 and Y-QAPc probes (lanes 6and 12).
Among the complexes observed using Y-QAPc oligo-
TABLE 1 Double stranded oligonucleotides used in EMSA experiments
Probes Sequence Y-box
Y-QAPc 59-GATTTTTCTAATTGGCCAAAAAGATTAACCGGTTTTGGAC-59
QAP consensus
Y-QAP4 59-GATTTTTCTAATTAGCCAAAAAGATTAATCGGTTTTGGAC-59
QAP4 variants
Y-DRA 59-ATTTTTCTGATTGGCCAAAGAGACTAACCGGTTTCTCATT-59
DRA promoter
TABLE 2 Primers used for competitive RT-PCR assay
Gene Primers Sequence
DQA1 Ext A 59-GGTGTAAACTTGTACCAGT-39B 59-AAACACTGTGACCTCAGG-39
Int C-2T 5 9- (2T) -GTGCAAATTGCGGGTCAA-39D-1T 5 9- (1T) -TGACAAACATCGCTGTGC-39
b-actin Ext BA1 5 9-CATGTGCAAGGCCGGCTTCG-39BA4 59-GAAGGTGTGGTGCCAGATTT-39
Int BA-1T 5 9- (1T) -GGCGTGATGGTGGGCATGGG-39BA-2T 5 9- (2T) -CTGGTGCCTGGGGCGCCCCA-39
Tails 1T 5 9- ACCTGCAGGGATCCGTCGAC-392T 59- GTCGACGGATCCCTGCAGGT-39
761NF-Y Binding to DQA1 Promoters
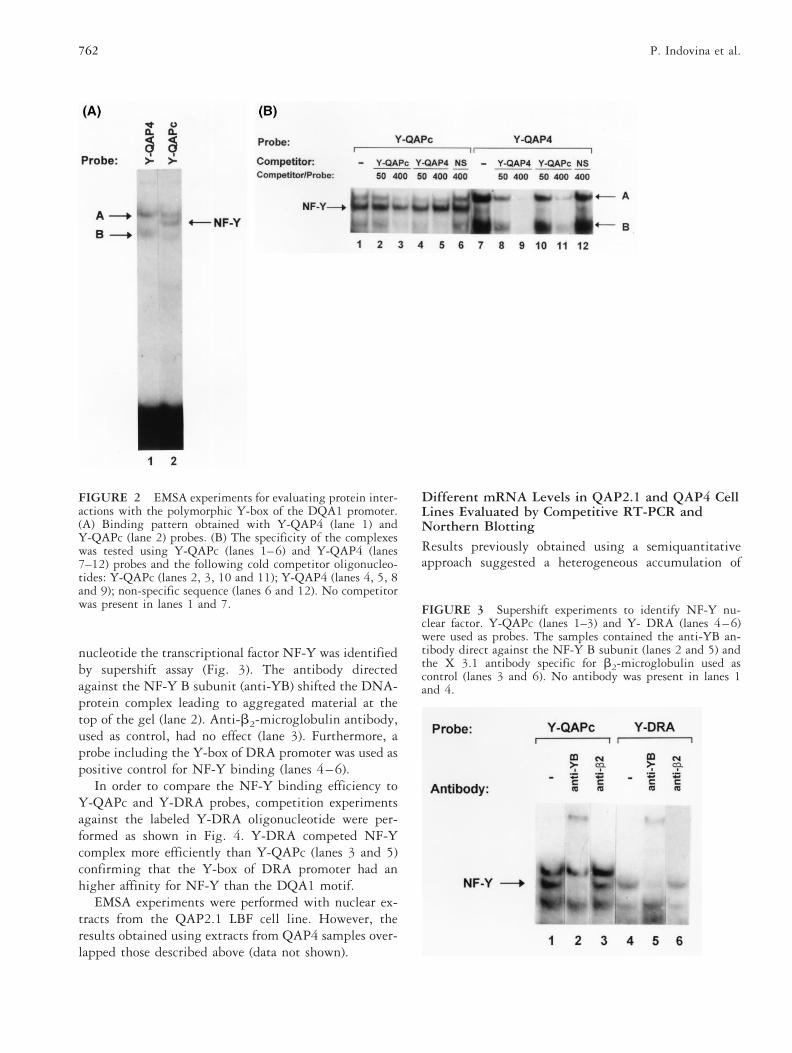
nucleotide the transcriptional factor NF-Y was identifiedby supershift assay (Fig. 3). The antibody directedagainst the NF-Y B subunit (anti-YB) shifted the DNA-protein complex leading to aggregated material at thetop of the gel (lane 2). Anti-b2-microglobulin antibody,used as control, had no effect (lane 3). Furthermore, aprobe including the Y-box of DRA promoter was used aspositive control for NF-Y binding (lanes 4–6).
In order to compare the NF-Y binding efficiency toY-QAPc and Y-DRA probes, competition experimentsagainst the labeled Y-DRA oligonucleotide were per-formed as shown in Fig. 4. Y-DRA competed NF-Ycomplex more efficiently than Y-QAPc (lanes 3 and 5)confirming that the Y-box of DRA promoter had anhigher affinity for NF-Y than the DQA1 motif.
EMSA experiments were performed with nuclear ex-tracts from the QAP2.1 LBF cell line. However, theresults obtained using extracts from QAP4 samples over-lapped those described above (data not shown).
Different mRNA Levels in QAP2.1 and QAP4 CellLines Evaluated by Competitive RT-PCR andNorthern BlottingResults previously obtained using a semiquantitativeapproach suggested a heterogeneous accumulation of
FIGURE 2 EMSA experiments for evaluating protein inter-actions with the polymorphic Y-box of the DQA1 promoter.(A) Binding pattern obtained with Y-QAP4 (lane 1) andY-QAPc (lane 2) probes. (B) The specificity of the complexeswas tested using Y-QAPc (lanes 1–6) and Y-QAP4 (lanes7–12) probes and the following cold competitor oligonucleo-tides: Y-QAPc (lanes 2, 3, 10 and 11); Y-QAP4 (lanes 4, 5, 8and 9); non-specific sequence (lanes 6 and 12). No competitorwas present in lanes 1 and 7. FIGURE 3 Supershift experiments to identify NF-Y nu-
clear factor. Y-QAPc (lanes 1–3) and Y- DRA (lanes 4–6)were used as probes. The samples contained the anti-YB an-tibody direct against the NF-Y B subunit (lanes 2 and 5) andthe X 3.1 antibody specific for b2-microglobulin used ascontrol (lanes 3 and 6). No antibody was present in lanes 1and 4.
762 P. Indovina et al.

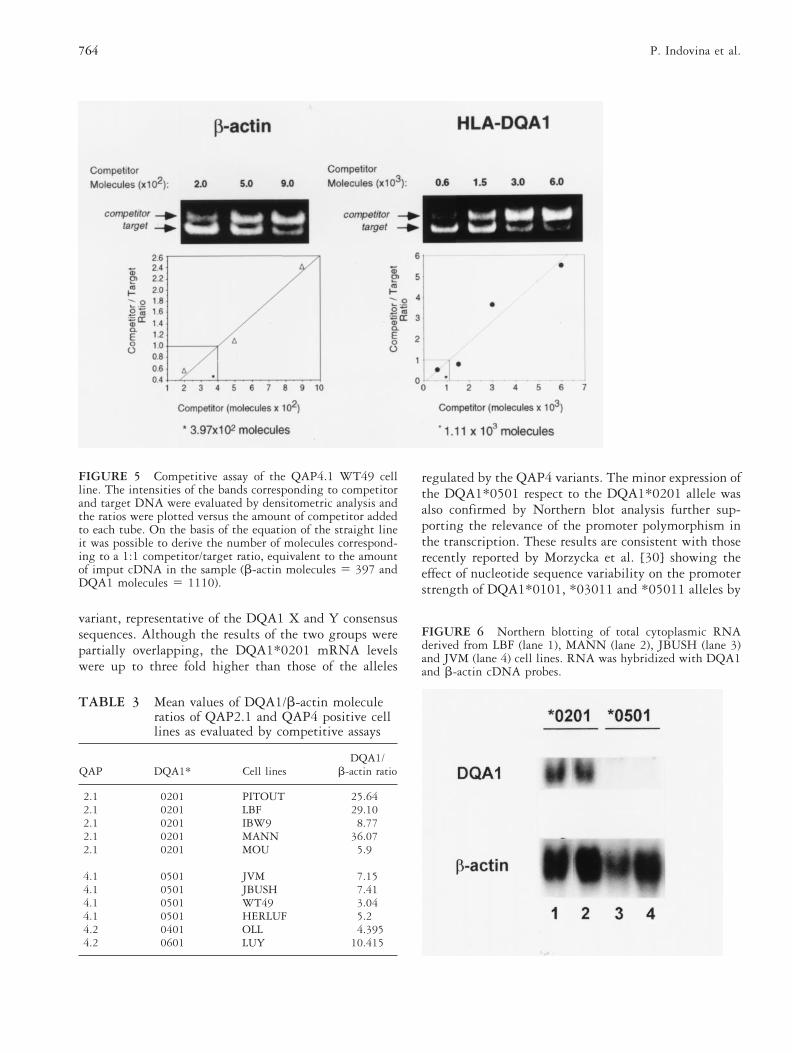
DQA1 mRNA molecules in samples carrying differentQAP variants [35]. In particular, the allelesDQA1*0401,*0501,*0601 regulated by the QAP4 pro-moters were the least expressed in comparison to theother DQA1 alleles. Based on these preliminary results amore precise evaluation of the number of DQA1 tran-script molecules has been performed using the compet-itive RT-PCR method [34]. The mRNA levels of allelesregulated by the QAP4 variants were compared to thoseof cell lines carrying the QAP2.1 promoter representa-tive of the X and Y consensus sequences. The constitu-tively expressed b-actin gene was chosen as control tonormalize the amount of cDNA used. A representativeresult relative to the WT49 cell line is reported in Fig.5. Since a certain degree of variability was observedtesting various cDNAs or RNA extractions performed atdifferent periods of time, the results relative to each cellline were expressed as mean values of DQA1/b-actinratios. As shown in Table 3, the QAP 2.1 cell linesreached three-fold higher levels of mRNA respect to theQAP 4 group (36.07 vs 10.415) although the observa-tions appeared very heterogeneous, ranging from 5.9 to36.07. On the contrary, a major degree of homogeneitywas observed in the QAP4 cell lines (3.04–10.415). Thestatistical analysis performed using Student’s t test fornot appaired data resulted significant with p 5 0.023.
The differences in the DQA1 mRNA levels of QAP4and QAP2.1 samples were also observed by Northernblot analysis using RNAs from LBF, MANN (QAP2.1)and JBUSH, JVM (QAP4.1) cell lines, as shown in Fig.6. The band relative to the DQA1 mRNA was clearlypresent in both QAP2.1 samples while seemed undetect-able in QAP4.1 cell lines. The b-actin mRNA used ascontrol appeared in all lanes without significant differ-ences.
DISCUSSIONA polymorphism in the DQA1 promoter has been de-scribed and the distribution of the ten variants in differ-ent populations and in relation to diseases has beenreported [22–27, 36, 37]. In order to investigate theeffect of the allelic differences in the regulatory boxes onthe binding of transcriptional factors, we analyzed theinteraction of the NF-Y protein with the Y box of DQA1promoter comparing the consensus sequence to theQAP4 variants which carry a G vs A transition in theATTGG/CCAAT motif. Interestingly, the NF-Y/DNAcomplex, present in the electrophoretic mobility shiftassay using the Y-QAP consensus sequence, was hardlydetectable using the Y-QAP4 probe. The DQA1 pro-moter also presents a locus-specific substitution imme-diately upstream to the inverted CCAAT motif and theNF-Y binding affinity seemed lower for Y-QAPc thanfor Y-DRA sequence as evident by competition experi-ments. It could be inferred that the NF-Y bindingefficiency followed a QAP4 ,, QAP consensus , DRAgradient since the mutation within the core motif almostprevented NF-Y interaction while that in the adjacentnucleotides only reduced its binding. These results aresupported by those obtained by in vitro mutagenesiswhere single-site transversions in the CCAAT sequencecompletely abolished the NF-Y binding, while flankingmutations affected the binding efficiency at a minorextent [32].
Two more complexes (A and B) were shown to spe-cifically bind to the DQA1 Y-box and their affinitieswere greater for the Y-QAP4 sequence than for theconsensus one. The Y box binding protein YB-1 has beenpreviously described [12, 13], however, there were nodata to conclude if any of the observed complexes couldbe related to this factor.
Considering the key role played by the NF-Y nuclearfactor in the initiation of transcription and its coopera-tive interactions with the other DNA-binding proteins,we evaluated the mRNA levels of the DQA1*0401,*0501 and *0601 alleles regulated by QAP4 promotersin a panel of cell lines by competitive RT-PCR. Theresults were compared with those obtained inDQA1*0201 positive cell lines regulated by the QAP2.1
FIGURE 4 Comparison of NF-Y binding affinity to theY-box of the DQA1 and DRA promoters. EMSA experimentswere performed with Y-DRA probe. Cold competitors Y-DRA(lanes 2 and 3) and Y-QAPc (lanes 4 and 5) were used to testtheir relative ability to compete NF-Y/DNA complex. Nocompetitor was present in lane 1.
763NF-Y Binding to DQA1 Promoters
variant, representative of the DQA1 X and Y consensussequences. Although the results of the two groups werepartially overlapping, the DQA1*0201 mRNA levelswere up to three fold higher than those of the alleles
regulated by the QAP4 variants. The minor expression ofthe DQA1*0501 respect to the DQA1*0201 allele wasalso confirmed by Northern blot analysis further sup-porting the relevance of the promoter polymorphism inthe transcription. These results are consistent with thoserecently reported by Morzycka et al. [30] showing theeffect of nucleotide sequence variability on the promoterstrength of DQA1*0101, *03011 and *05011 alleles by
TABLE 3 Mean values of DQA1/b-actin moleculeratios of QAP2.1 and QAP4 positive celllines as evaluated by competitive assays
QAP DQA1* Cell linesDQA1/
b-actin ratio
2.1 0201 PITOUT 25.642.1 0201 LBF 29.102.1 0201 IBW9 8.772.1 0201 MANN 36.072.1 0201 MOU 5.9
4.1 0501 JVM 7.154.1 0501 JBUSH 7.414.1 0501 WT49 3.044.1 0501 HERLUF 5.24.2 0401 OLL 4.3954.2 0601 LUY 10.415
FIGURE 5 Competitive assay of the QAP4.1 WT49 cellline. The intensities of the bands corresponding to competitorand target DNA were evaluated by densitometric analysis andthe ratios were plotted versus the amount of competitor addedto each tube. On the basis of the equation of the straight lineit was possible to derive the number of molecules correspond-ing to a 1:1 competitor/target ratio, equivalent to the amountof imput cDNA in the sample (b-actin molecules 5 397 andDQA1 molecules 5 1110).
FIGURE 6 Northern blotting of total cytoplasmic RNAderived from LBF (lane 1), MANN (lane 2), JBUSH (lane 3)and JVM (lane 4) cell lines. RNA was hybridized with DQA1and b-actin cDNA probes.
764 P. Indovina et al.

transient transfection assays. Indeed, the plasmid con-struct carrying the QAP4.1 promoter presented the low-est level of transcription. Furthermore, the G vs A mu-tation of the QAP4 sequences introduced in the Y motifof the p03011 construct strongly reduced its level ofactivity, proving the crucial role of the Y-box polymor-phism on the promoter strength. Taken together, theNF-Y inability to bind the QAP4 Y- box and the minorexpression of constructs with the G vs A transition, let usassume that the low mRNA level of DQA1*0401,*0501 and *0601 alleles was, at least in part, due to thelack of the NF-Y binding. The presence of the two othercomplexes, A and B, specific for the Y box was notsufficient to raise the mRNA accumulation of thesealleles to the levels of DQA1*0201 one, besides theirstrong binding affinity for the QAP4 sequence.
Differences in the Y-box sequence could account notonly for allele variations but also for interloci diversity.As shown, the NF-Y protein had a lower binding affinityfor the Y-box consensus of the DQA1 promoter respectto the DRA sequence. Thus, the different binding of thistranscriptional factor to the regulatory Y-motif couldcontribute to the differential expression of DQ respect toDR antigens on APC cells [16]. However, intra andinterloci differences of transcription could also be corre-lated to the variability of other regulatory sequences.Previous studies relative to the X-box nucleotide varia-tions of DRA, DPA and DQA promoters showed thatthe DQA1 sequence had the lowest affinity for the RF-Xcomplex and the higher for the NF-S protein [19, 20].The X-box polymorphism of the DQA1 promoter couldhave a similar effect on the binding of nuclear factors,therefore, its functional relevance in the differences weobserved at mRNA level should be also considered.
Understanding the functional effect of the nucleotidepolymorphism of DQA1 promoter could be particularlyrelevant for the biological effects attributable to the DQmolecules. The conservation of linkage among allelicDQA1 sequences in the coding region and the upstreamregulatory region may reflect a selective pressure to keepspecific combinations together. Harboring a diversity ofpromoters could enrich the flexibility and variety of theHLA system in resisting infection [18]. On the otherhand, disease susceptibility ascribed to the structuralpolymorphism could be conferred by specific promoter-allele combinations. Among the DQA1 alleles, theDQA1*0501 one, regulated by the QAP4.1 promoter, isoften associated to autoimmune diseases such as theinsulin-dependent diabetes mellitus or celiac disease [38,39]. However, the strong association between this struc-tural allele and its regulatory region does not allow toestablish the real effect of the latter in disease suscepti-bility. A direct role of the promoter sequences has beensuggested in the early-onset pauciarticular juvenile
chronic arthritis found associated to the three DQA1alleles regulated by the QAP4 variants [36].
Quantitative differences in the expression of DQA1alleles could be particularly relevant in the mechanism ofthymic selection and in the induction of T cell prolifer-ation at the basis of the immunological disturbances [40,41]. The analysis of cell surface expression in normal cellsas well as in pathological models will provide clear-cutevidence of the final significance of these promoter poly-morphism. Regarding celiac disease, the analysis of thedendritic cells, the main APC system of the intestinalmucosa, could represent an interesting model to inves-tigate the pathological relevance of the results describedin the present study.
In conclusion, we brought new evidence of differentialbinding of nuclear factors to the Y-box of DQA1 pro-moter variants differing for a single site mutation. Inparticular, the G vs A transition in the Y-QAP4 motifdramatically affected the NF-Y binding. Concurrently,the levels of mRNA transcription of the DQA1*0401,*0501 and *0601 alleles carrying the QAP4 reg-ulatory sequence appeared lower than those of DQA1*0201 positive cell lines. The evaluation of DQA1 expres-sion on peripheral blood lymphocytes as well as theanalysis of the other polymorphic boxes is currently inprogress.
ACKNOWLEDGMENTS
We thank Drs. M.P. Felli and P. Giacomini for the helpfuldiscussion. The cDNA probes and the anti-YB antibody werekindly given from Dr. A. De Lerma and Dr. R. Mantovani,respectively. P. Indovina and F. Megiorni contributed equallyto this work which was supported by the CNR Target Projecton “Biotechnology.”
REFERENCES
1. Benoist C, Mathis D: Regulation of major histocompati-bility complex class II genes: X, Y and other letters of thealphabet. Annu Rev Immunol 8:681, 1990.
2. Glimcher LH, Kara CJ: Sequences and factors: a guide toMHC class II transcription. Annu Rev Immunol 10:13,1992.
3. Garrido F, Cabrera T, Concha A, Glew S, Ruiz-Cabello F,Stern PL: Natural history of HLA expression during tu-mour development. Immunol Today 14:491, 1993.
4. Reith W, Steimle V, Mach B: Molecular defects in thebare lymphocyte syndrome and regulation of MHC class IIgenes. Immunol Today 16:539, 1995.
5. Mach B, Steimle V, Martinez-Soria E, Reith W: Regula-tion of MHC class II genes: lessons from a disease. AnnuRev Immunol 14:301, 1996.
6. Louis-Plence P, Moreno CS, Boss JM: Formation of aregulatory factor X/X2 box-binding protein/nuclear fac-
765NF-Y Binding to DQA1 Promoters

tor-Y multiprotein complex on the conserved regulatoryregions of HLA class II genes. J Immunol 159:3899,1997.
7. Fontes JD, Jabrane-Ferrat N, Peterlin BM: Assembly offunctional regulatory complexes on MHC class II promot-ers in vivo. J Mol Biol 270:336, 1997.
8. Mantovani R, Pessara U, Tronche F, Li XY, Knapp AM,Pasquali JL, Benoist C, Mathis D: Monoclonal antibodiesto NF-Y define its function in MHC class II and albumingene transcription. EMBO 11:3315, 1992.
9. Wright KL, Vilen BJ, Itoh-Lindstrom Y, Moore TL, Li G,Criscitiello M, Cogswell P, Badley Clarke J, Ting JPY:CCAAT box binding protein NF-Y facilitates in vivorecruitment of upstream DNA binding factors. EMBO13:4042, 1994.
10. Reith W, Siegrist CA, Durand B, Barras E, Mach B:Function of major histocompatibility complex class IIpromoters requires cooperative binding between factorsRFX and NF-Y. Proc Natl Acad Sci USA 91:554, 1994.
11. Bellorini M, Lee DK, Dantonel JC, Zemzoumi K, RoederRG, Tora L, Mantovani R: CCAAT binding NF-Y-TBPinteractions: NF-YB and NF-YC require short domainsadjacent to their histone fold motifs for association withTBP basic residues. Nucleic Acid Res. 25:2174, 1997.
12. Didier DK, Schiffenbauer J, Woulfe SL, Zacheis M,Schwartz BD: Characterization of the cDNA encoding aprotein binding to the major histocompatibility complexclass II Y box. Proc Natl Acad Sci USA 85:7322, 1988.
13. Ting JPY, Painter A, Zeleznik-Le NJ, MacDonald G,Moore TM, Brown A, Schwartz BD: YB-1 DNA bindingprotein represses interferon gamma activation of class IImajor histocompatibility complex genes. J Exp Med 179:1605, 1994.
14. Steimle V, Otten LA, Zufferey M, Mach B: Complemen-tation cloning of an MHC class II transactivator mutatedin hereditary MHC class II deficienti (or bare lymphocytesyndrome). Cell 75:135, 1993.
15. Riley JL, Westerheide SD, Price JA, Brown JA, Boss JM:Activation of class II MHC genes requires both the X boxregion and the class II transactivator (CIITA). Immunity2:533, 1995.
16. Collins T, Korman AJ, Wake CT, Boss JM, Kappes DJ,Fiers W, Ault KA, Gimbrone MA, Strominger JL, PoberJS: Immune interferon activates multiple class II majorhistocompatibility genes and the associated invariantchain in human endothelial cells and dermal fibroblasts.Proc Natl Acad Sci USA 81:4917, 1984.
17. De Preval C, Hadam MR, Mach B: Regulation of genesfor HLA class II antigens in cell lines from patients withsevere combined immunodeficiency. N Engl J Med 318:1295, 1988.
18. Guardiola J, Maffei A, Lauster R, Mitchinson NA, AccollaRS, Sartoris S: Functional significance of polymorphismamong MHC class II gene promoters. Tissue Antigens48:615, 1996.
19. Reith W, Barras E, Satola S, Kobr M, Reinhart D, SanchezCH, Mach B: Cloning of the major histocompatibilitycomplex class II promoter binding protein affected in ahereditary defect in class II gene regulation. Proc NatlAcad Sci USA 86:4200, 1989.
20. Kobr M, Reith W, Herrero-Sanchez C, Mach B: TwoDNA-binding proteins discriminate between the promot-ers of different members of the major histocompatibilitycomplex class II multigene family. Mol Cell Biol 10:965,1990.
21. Kimura A, Sasazuki T: HLA-DQA gene is differentlyregulated from other HLA class II genes. In Yooshida TO,Wilson JM (eds): Molecular approaches to the study andtreatment of human disease. Amsterdam, Elsevier Press,1992.
22. Kimura A, Sasazuki T: Polymorphism in the 5’ flankingregion of the HLA-DQA1 gene and its relation to theDR-DQ haplotype. In Tsuji K, Aizawa M (eds): HLA1991: Proceedings of the Eleventh International Histo-compatibility Workshop and Conference. Oxford Univer-sity Press, 1992.
23. Morzycka-Wroblewska E, Harwood JI, Smith JR,Kagnoff M: Structure and evolution of the promoter re-gions of the DQA genes. Immunogenetics 37:364, 1993.
24. Brunnler G, Haas JP, Fan LA, Petzl-Erler ML, Volgger A,Yao Z, Wassmuth R, Albert ED, Middleton D, BarboniF, Contu L, Alvarez LR, Ferrara GB, Adorno D, Goro-dezky C, Alaez C, Cerna M, Mazzilli MC: DQA1 pro-moter polymorphism: 12th International Histocompati-bility Workshop study. In Charron D (ed): Geneticdiversity of HLA, functional and medical implication,Vol. 1. Paris, EDK publisher, 1997.
25. Haas JP, Kimura A, Andreas A, Hochberger M, Keller E,Brunnler G, de la Paz Bettinotti M, Nevinny-Stickel C,Hildebrandt B, Sierp G, Sasazuki T, Albert E: Polymor-phism in the upstream regulatory region of DQA1 genesand DRB1, QAP, DQA1, and DQB1 haplotypes in theGerman population. Hum Immunol 39:31, 1994.
26. Reichstetter S, Krellner PH, Meenzen CM, Kalden JR,Wassmuth R: Comparative analysis of sequence variabil-ity in the upstream regulatory region of the HLA-DQB1gene. Immunogenetics 39:207, 1994.
27. Petronzelli F, Kimura A, Ferrante P, Mazzilli MC: Poly-morphism in the upstream regulatory region of DQA1gene in the Italian population. Tissue Antigens 45:258,1995.
28. Louis P, Vincent R, Cavadore P, Clot J, Eliaou JF: Dif-ferential transcriptional activities of HLA-DR genes inthe various haplotypes. J Immunol 153:5059, 1994.
29. Beaty JS, West KA, Nepom GT: Functional effects of anatural polymorphism in the transcriptional regulatorysequence of HLA-DQB1. Mol Cell Biol 15:4771, 1995.
30. Morzycka-Wroblewska E, Munshi A, Ostermayer M, Har-wood JI, Kagnoff M: Differential expression of HLA-
766 P. Indovina et al.

DQA1 alleles associated with promoter polymorphism.Immunogenetics 45:163, 1997.
31. Dignam JD, Lebovitz RM, Roeder RG: Accurate tran-scription inititation by RNA polymerase II in a solubleextract from isolated mammalian nuclei. Nucleic AcidsRes 11:1475, 1983.
32. Dorn A, Bollekens J, Staub A, Benoist C, Mathis D: Amultiplicity of CCAAT box- binding proteins. Cell 50:863, 1987.
33. Chomcwynski P, Sacchi N: Single-step method of RNAisolation by acid guanidinium thiocyanate-phenol-chloro-form extraction. Anal Biochem 162:156, 1987.
34. Diviacco S, Norio P, Zentilin L, Menzo S, Clementi M,Biamonti G, Riva S, Norio P, Zentilin L, Giacca M: Anovel procedure for quantitative polymerase chain reac-tion by coamplification of competitive templates. Gene122:3013, 1992.
35. Ferrante P, Apollonio I, Del Nero A, Petronzelli F,Mazzilli MC: Regulation of HLA gene expression. Min-erva Biotec 8:32, 1996.
36. Haas JP, Kimura A, Truckenbrodt H, Suschke J, SasazukiT, Volgger A, Albert ED: Early-onset pauciarticular ju-venile chronic arthritis is associated with a mutation in
the Y-box of the HLA-DQA1 promoter. Tissue Antigens45:317, 1995.
37. Yao Z, Kimura A, Hartun K, Haas PJ, Volgger A, Brunn-ler G, Bonisch J, Albert ED, SLE study group members:Polymorphism of the DQA1 promoter region (QAP) andDRB1, QAP, DQA1, DQB1 haplotypes in systemic lupuserythematosus. Immunogenetics 38:421, 1993.
38. She JX: Susceptibility to type 1 diabetes: HLA-DQ andDR revisited. Immunol Today 17:323, 1996.
39. Sollid ML, Markussen G, Ek J, Gjerde H, Vardtal F,Thorsby E: Evidence for a primary association of celiacdisease to a particular HLA-DQ a/b heterodimer. J ExpMed 169:345, 1989.
40. Matis LA, Glimcher LH, Paul WE, Schwartz RH: Mag-nitude of response of histocompatibility-restricted T cellclones is a function of the product of the concentrations ofantigen and Ia molecules. Proc Natl Acad Sci USA 80:6019, 1983.
41. Janeway CA, Bottomly K, Babich J, Conrad P, Conzen S,Jones B, Kaye J, Katz M, McVay L, Murphy DB, Tite J:Quantitative variation in Ia antigen expression plays acentral role in immune regulation. Immunol Today 5:99,1984.
767NF-Y Binding to DQA1 Promoters




















