
Diaphorobacter nitroreducens gen. nov., sp. nov., a poly(3-hydroxybutyrate)-degrading denitrifying...
10
Introduction Biological denitrification is the most widely used mi- crobial process for nitrogen removal in water and wastewater treatment. Conventional biological nitrogen removal systems use residual organic matter in waste- water or intentionally added liquid substrates, e.g., methanol and acetate, as electron donors for denitrifi- cation. On the other hand, several reports have in re- cent years introduced new biotechnology of nitrogen removal using solid biopolymers, such as poly(3-hy- droxybutyrate) (PHB) and poly(3-hydroxybutyrate-co- hydroxyvalerate) (PHBV), as an alternative substrate for microbial growth and denitrification (Biedermann et al.,1997; Boley et al., 2000; Khan and Hiraishi 2001; Mergaert et al., 2001; Müller et al., 1992; Schloe et al., 2000). In this connection, what kinds of PHBV-degrad- Diaphorobacter nitroreducens gen. nov., sp. nov., a poly(3-hydroxybutyrate)-degrading denitrifying bacterium isolated from activated sludge Shams Tabrez Khan and Akira Hiraishi* Department of Ecological Engineering, Toyohashi University of Technology, Toyohashi 441–8580, Japan (Received August 12, 2002; Accepted October 23, 2002) Three denitrifying strains of bacteria capable of degrading poly(3-hydroxybutyrate) (PHB) and poly(3-hydroxybutyrate-co-3-hydroxyvalerate) (PHBV) were isolated from activated sludge and characterized. All of the isolates had almost identical phenotypic characteristics. They were motile gram-negative rods with single polar flagella and grew well with simple organic com- pounds, as well as with PHB and PHBV, as carbon and energy sources under both aerobic and anaerobic denitrifying conditions. However, none of the sugars tested supported their growth. The cellular fatty acid profiles showed the presence of C16:1w7cis and C16:0 as the major com- ponents and of 3-OH-C10:0 as the sole component of hydroxy fatty acids. Ubiquinone-8 was de- tected as the major respiratory quinone. A 16S rDNA sequence-based phylogenetic analysis showed that all the isolates belonged to the family Comamonadaceae, a major group of b -Pro- teobacteria, but formed no monophyletic cluster with any previously known species of this fam- ily. The closest relative to our strains was an unidentified bacterium strain LW1 (DSM 13225) (99.9% similarity), reported previously as a 1-chloro-4-nitrobenzene degrading bacterium. DNA- DNA hybridization levels among the new isolates were more than 60%, whereas those between our isolates and strain DSM 13225 were less than 50%. The GC content of genomic DNA of the new strains was 64 to 65 mol%. Based on these results, we concluded that the PHBV-degrading denitrifying isolates should be classified as a new genus and a new species, for which we pro- pose the name Diaphorobacter nitroreducens. The type strain is strain NA10B (JCM 11421CIP 107294). We also propose to classify strain DSM 13225 as a genospecies of Diaphorobacter. Key Words——activated sludge; denitrifying bacteria; Diaphorobacter nitroreducens; PHB degradation J. Gen. Appl. Microbiol., 48, 299–308 (2002) Full Paper * Address reprints requests to: Dr. Akira Hiraishi, Department of Ecological Engineering, Toyohashi University of Technology, Toyohashi 441–8580, Japan. E-mail: [email protected]
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Diaphorobacter nitroreducens gen. nov., sp. nov., a poly(3-hydroxybutyrate)-degrading denitrifying...

Introduction
Biological denitrification is the most widely used mi-crobial process for nitrogen removal in water andwastewater treatment. Conventional biological nitrogenremoval systems use residual organic matter in waste-
water or intentionally added liquid substrates, e.g.,methanol and acetate, as electron donors for denitrifi-cation. On the other hand, several reports have in re-cent years introduced new biotechnology of nitrogenremoval using solid biopolymers, such as poly(3-hy-droxybutyrate) (PHB) and poly(3-hydroxybutyrate-co-hydroxyvalerate) (PHBV), as an alternative substratefor microbial growth and denitrification (Biedermann etal.,1997; Boley et al., 2000; Khan and Hiraishi 2001;Mergaert et al., 2001; Müller et al., 1992; Schloe et al.,2000). In this connection, what kinds of PHBV-degrad-
Diaphorobacter nitroreducens gen. nov., sp. nov., a poly(3-hydroxybutyrate)-degrading denitrifying bacterium isolated
from activated sludge
Shams Tabrez Khan and Akira Hiraishi*
Department of Ecological Engineering, Toyohashi University of Technology, Toyohashi 441–8580, Japan
(Received August 12, 2002; Accepted October 23, 2002)
Three denitrifying strains of bacteria capable of degrading poly(3-hydroxybutyrate) (PHB) andpoly(3-hydroxybutyrate-co-3-hydroxyvalerate) (PHBV) were isolated from activated sludge andcharacterized. All of the isolates had almost identical phenotypic characteristics. They weremotile gram-negative rods with single polar flagella and grew well with simple organic com-pounds, as well as with PHB and PHBV, as carbon and energy sources under both aerobic andanaerobic denitrifying conditions. However, none of the sugars tested supported their growth.The cellular fatty acid profiles showed the presence of C16:1ww7cis and C16:0 as the major com-ponents and of 3-OH-C10:0 as the sole component of hydroxy fatty acids. Ubiquinone-8 was de-tected as the major respiratory quinone. A 16S rDNA sequence-based phylogenetic analysisshowed that all the isolates belonged to the family Comamonadaceae, a major group of bb-Pro-teobacteria, but formed no monophyletic cluster with any previously known species of this fam-ily. The closest relative to our strains was an unidentified bacterium strain LW1 (�DSM 13225)(99.9% similarity), reported previously as a 1-chloro-4-nitrobenzene degrading bacterium. DNA-DNA hybridization levels among the new isolates were more than 60%, whereas those betweenour isolates and strain DSM 13225 were less than 50%. The G�C content of genomic DNA of thenew strains was 64 to 65 mol%. Based on these results, we concluded that the PHBV-degradingdenitrifying isolates should be classified as a new genus and a new species, for which we pro-pose the name Diaphorobacter nitroreducens. The type strain is strain NA10B (�JCM 11421�CIP107294). We also propose to classify strain DSM 13225 as a genospecies of Diaphorobacter.
Key Words——activated sludge; denitrifying bacteria; Diaphorobacter nitroreducens; PHB degradation
J. Gen. Appl. Microbiol., 48, 299–308 (2002)
Full Paper
* Address reprints requests to: Dr. Akira Hiraishi, Departmentof Ecological Engineering, Toyohashi University of Technology,Toyohashi 441–8580, Japan.
E-mail: [email protected]

ing denitrifiers exist and actually work in such a newdenitrification process is a subject of major concern.
PHBV-degrading denitrifying bacteria hitherto iso-lated have been identified as being Acidovorax facilis(Mergaert et al., 2001), Acidovorax sp. (Schloe et al.,2000), and Brevundimonas intermedia (Mergaert et al.,2001). In a previous study, we isolated a new PHB-and PHBV-degrading denitrifying bacterium that exhib-ited such a high denitrification rate as 19 mg NO3
�-N re-moved g dry cells�1 h�1 with PHBV as the electrondonor (Khan and Hiraishi, 2001). A phylogenetic analy-sis based on 16S rDNA sequences showed that thisbacterium, designated strain NA10BT, belongs to thefamily Comamonadaceae, a major group of b-Pro-teobacteria (Willems et al., 1991), but does not form aphylogenetic cluster with any species of the genera ofthis family. Our attempt to find further similar PHBV-de-grading denitrifiers in activated sludge was successful(Khan et al., 2002), and a total of three strains of thisnew group of the Comamonadaceae have been iso-lated and characterized. In this article, we report thetaxonomic characteristics of the new PHBV-degradingdenitrifying strains and propose to classify them into anew genus and species with the name Diaphorobacternitroreducens.
Materials and Methods
Bacterial strains. Strains NA10BT, KSP3 andKSP4, all of which were isolated from activated sludgetaken from sewage treatment plants in Japan (Khanand Hiraishi, 2001; Khan et al., 2002), were studied.The reference strains used were Acidovorax avenaesubsp. avenae DSM 7227T, Acidovorax facilis DSM649T, Acidovorax defulvi DSM 12644T, Acidovorax de-lafieldii DSM 64T, Acidovorax konjaci DSM 7481T, Aci-dovorax temperans DSM 7270T, [Aquaspirillum]sinosum DSM 11556T (bracketed name�misclassifiedgeneric name) (Wen et al., 1999), Brachymonas deni-trificans AS-P1T, Comamonas terrigena IAM 12421T,Comamonas testosteroni IAM 12419T and JCM 10170,Delftia acidovorans IAM 12409T, and Rhodoferx fer-mentans FR2T. In addition, an unidentified bacterium,strain DSM 13225 (�strain LW1) (Katsivela et al.,1999) were used. The strains with DSM, IAM, andJCM numbers were obtained from the DSMZ(Deutsche Summlung von Mikroorganismen und Zel-lkulturen GmbH), Braunschweig, Germany, IAM Cul-ture Collection, The University of Tokyo, Tokyo, Japan,
and the Japan Collection of Microorganisms, Wako,Japan, respectively. All other strains were from ourown collection.
Growth media and cultivation. A complex mediumdesignated PBY (0.5% Bacto-peptone (Difco Laborato-ries, Detroit, MI, USA), 0.3% beef extract (Difco) and0.1% yeast extract (Difco)) was used for maintenance,preculture, and main culture used for chemotaxonomicand molecular studies. For studying PHBV degrada-tion, mineral medium RM2 (Hiraishi and Kitamura,1984) supplemented with 0.2% PHBV and 0.01%yeast extract (Difco) was used. These media con-tained 1.8% agar when used as solidified media. Foranaerobic denitrifying growth, PBY and PHBV mediawere supplemented with 0.2% KNO3. Liquid cultureswere introduced into screw-capped test tubes (20-mlcapacity) or bottles (100–500 ml capacity) and incu-bated aerobically with vigorous shaking on a reciprocalshaker. For anaerobic growth, test tubes and bottleswere completely filled with a medium and incubatedwithout shaking. Anaerobic denitrifying growth on agarmedia was performed using the AnaeroPak system(Mitsubishi Gas Chemicals, Niigata, Japan). All cul-tures were incubated at 28 to 30°C unless otherwisespecified.
Microscopic studies. Morphological propertieswere studied using an Olympus BX-50 phase-contrastmicroscope. For more detailed morphological studies,a JEOL-1200EX transmission electron microscopewas used. Gram staining was carried out as describedby Magee et al. (1975).
Physiological and biochemical tests. Unless other-wise specified, all tests were carried out at 28 to 30°C.Degradation of PHB and PHBV was determined as de-scribed previously (Khan and Hiraishi, 2001). Carbonnutrition tests were performed in 20-ml screw-cappedtest tubes containing 8 ml of mineral medium RM2 (Hi-raishi and Kitamura, 1984) supplemented with an or-ganic compound as the carbon source. All carbonsources were filter-sterilized and added at a final con-centration of 0.2% as a neutralized form. For anaero-bic denitrifying growth, the test tubes were completelyfilled with the medium supplemented with 0.2% KNO3.The test tubes were incubated for 10 days with vigor-ous shaking for aerobic growth or without shaking foranaerobic growth. Growth was monitored by measur-ing the optical density at 660 nm. All other tests wereperformed as described by Hiraishi et al. (1995).
Fatty acid and quinone analyses. Whole-cell fatty
300 KHAN and HIRAISHI Vol. 48

acids were extracted with methanol-HCl, and gas chro-matographic analyses of their methyl esters were per-formed by NCIMB Japan Co., Shimizu, Japan. In somecases, 3-OH fatty acids in the samples extracted wereseparated by thin-layer chromatography and analyzedby gas chromatography as described previously (Hi-raishi et al., 1992). Quinones were extracted, fraction-ated, and analyzed by spectrochromatography andmass spectrometry as described previously (Hiraishi etal., 1996).
DNA base composition and DNA-DNA hybridization.Genomic DNA was extracted and purified by themethod of Marmur (1961). DNA base composition(guanine [G]�cytosine [C] ratios) was determined bythe HPLC method (Katayama et al., 1984). DNA-DNAhybridization studies were performed by the quantita-tive dot-blot hybridization method (Hiraishi et al., 1991)with alkaline phosphatase labeling and chemilumines-cence detection using an Amersham-PharmaciaAlkPhos kit. Hybridization signals were detected withan ECL Mini camera (Amersham-Pharmacia) andmeasured by an image analysis using the public do-main NIH Image program (http://rsb.info.nih.gov/nih-image/).
Phylogenetic analysis. The 16S rDNA sequencesof the new isolates were determined in our previousstudies (Khan and Hiraishi, 2001; Khan et al., 2002)and compared to those retrieved from theDDBJ/EMBL/GenBank databases. Multiple alignmentof sequence, calculation of the corrected evolutionarydistance (Kimura, 1980) and construction of a phyloge-netic tree by the neighbor-joining method (Saitou andNei, 1987) were performed using the CLUSTAL W pro-gram (Thompson et al., 1994). Branching patterns ofthe tree were evaluated by bootstrapping with 1,000resamplings (Felsenstein, 1985). Alignment positionswith gaps and unidentified bases were excluded forthe calculations. The tree was illustrated using theTreeView program (Page, 1996).
Results
Morphology and cultural characteristicsThe three new isolates had gram-negative, as-
porogenous, rod-shaped cells with rounded ends. Thecell size was 0.7 to 0.9 mm in width and 1.0 to 1.8 mmin length. Cells were actively motile with single polarflagella (Fig. 1). Pilus-like filamentous appendagesprojecting from the cell surface were also found in all
isolates.Cultivation on PBY agar media showed that strain
NA10BT formed slightly rough and raised colonies,whereas strains KSP3 and KSP4 had smooth colonieswith entire margins. The colonies and cell suspensionwere colorless at the early phase of growth but turnedcream to beige at older stages.
Phylogenetic and genetic analysesAlmost complete sequences (1,521–1,522 bases) of
the 16S rDNA of strains NA10BT, KSP3, and KSP4were determined in our previous studies. The se-quences of strains KSP3 and KSP4 were identical,and these were different in only two positions from thatof strain NA10BT. Homology search indicated that thethree new isolates had sequence similarities of 95.0–96.8% to Acidovorax species with Acidovorax avenaesubsp. citrulli as their closest relative, 96.3–96.4% toXylophilus ampelinus, 96.3% to [Aquaspirillum] anulus,95.9–96.0% to [Aquaspirillum] giesbergeri, 95.7–95.8%to [Aquaspirillum] sinosum, 94.1–95.7% to Coma-monas species, 95.2–95.4% to [Aquaspirillum] meta-morphum, and 95.1% to Delftia acidovorans. The newisolates also showed 99.9% similarity to an unidenti-fied bacterium strain LW1 (�DSM 13225), which isknown as a 1-chloro-4-nitrobenzene-degrading bac-terium (Katsivela et al., 1999).
2002 Diaphorobacter nitroreducens gen. nov., sp. nov. 301
Fig. 1. Transmission electron micrograph of a cell with asingle polar flagellum of strain NA10BT.
Scale bar�0.5 mm.

A neighbor-joining phylogenetic tree inferred fromthe 16S rDNA sequences is shown in Fig. 2. The treeclearly showed that all the isolates belong to the familyComamonadaceae and form a tight cluster togetherwith strain LW1 that is separable from any clusters ofthe previously known genera of this family. The groupof our isolates and strain LW1 clustered with an as-semblage of the misclassified Aquaspirillum species,[Aquaspirillum] anulus, [Aquaspirillum] giesbergeri,and [Aquaspirillum] sinosum (Ding and Yokota, 2002),as its sister group, but the bootstrap value supportingfor this cluster was low. Similar branching patterns onthe phylogenetic tree were obtained by the maximumlikelihood method as described previously (Khan andHiraishi, 2001).
Values of genomic DNA-DNA hybridization among
the new isolates and related organisms are shown inTable 1. DNA-DNA similarity levels among the newisolates were more than 60%, whereas those betweenthe former isolates and strain DSM 13225 (�strainLW1) were less than 50%. Much lower DNA-DNA re-latedness was found between our isolates and the au-thentic strains of Acidovorax avenae, Acidovorax fa-cilis, Acidovorax delafieldii, Comamonas testosteroni,and Delftia acidovorans. The G�C content of the newisolates ranged from 64 to 65 mol%. Strain DSM13225 also gave a G�C value within this range.
Fatty acid and quinone profilesThe three new isolates were homogeneous in cellu-
lar fatty acid composition (Table 2). In all the isolates,the primary component present was palmitoleic acid
302 KHAN and HIRAISHI Vol. 48
Fig. 2. Neighbor-joining distance matrix tree based on 16S rDNA sequences showing the phylogenetic relation-ships between the new PHBV-degrading denitrifying isolates and species of Comamonadaceae genera.
Accession numbers for the 16S rDNA sequences used are given in parentheses behind strain and species names.Bootstrap values are given at branching points of interest. Scale bar�1% nucleotide substitution rate by Kimura’s two-parameter model.
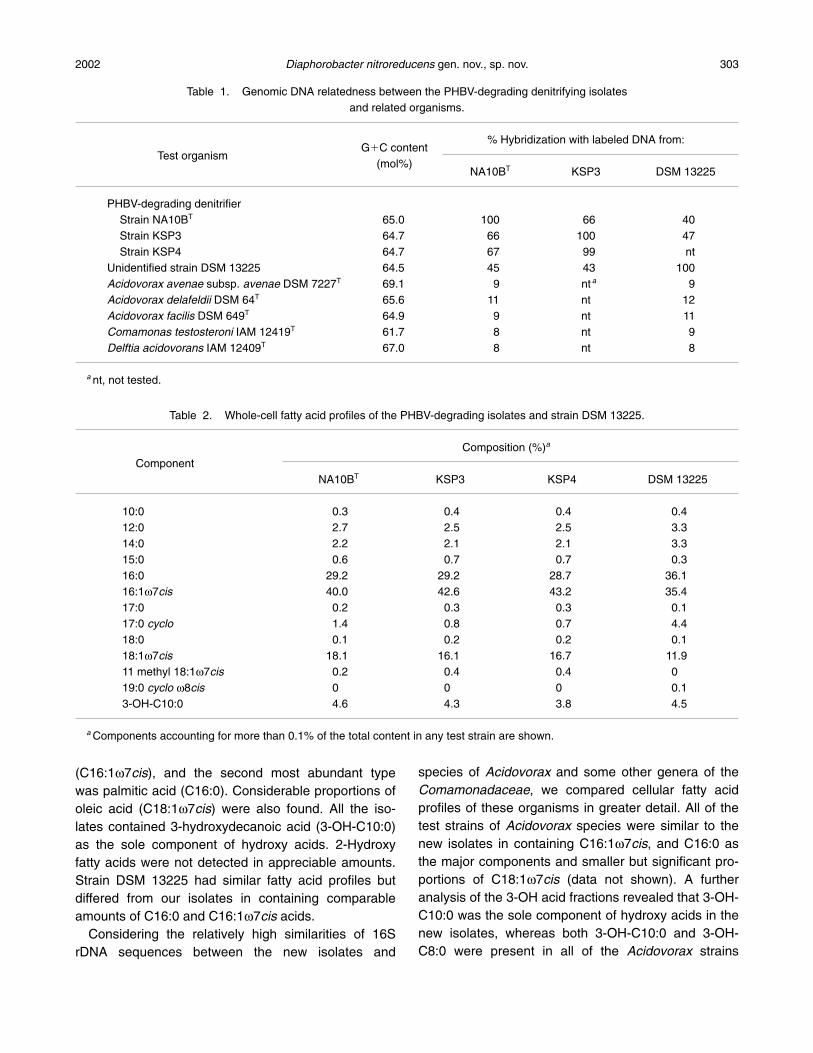
(C16:1w7cis), and the second most abundant typewas palmitic acid (C16:0). Considerable proportions ofoleic acid (C18:1w7cis) were also found. All the iso-lates contained 3-hydroxydecanoic acid (3-OH-C10:0)as the sole component of hydroxy acids. 2-Hydroxyfatty acids were not detected in appreciable amounts.Strain DSM 13225 had similar fatty acid profiles butdiffered from our isolates in containing comparableamounts of C16:0 and C16:1w7cis acids.
Considering the relatively high similarities of 16SrDNA sequences between the new isolates and
species of Acidovorax and some other genera of theComamonadaceae, we compared cellular fatty acidprofiles of these organisms in greater detail. All of thetest strains of Acidovorax species were similar to thenew isolates in containing C16:1w7cis, and C16:0 asthe major components and smaller but significant pro-portions of C18:1w7cis (data not shown). A furtheranalysis of the 3-OH acid fractions revealed that 3-OH-C10:0 was the sole component of hydroxy acids in thenew isolates, whereas both 3-OH-C10:0 and 3-OH-C8:0 were present in all of the Acidovorax strains
2002 Diaphorobacter nitroreducens gen. nov., sp. nov. 303
Table 1. Genomic DNA relatedness between the PHBV-degrading denitrifying isolates and related organisms.
G�C content% Hybridization with labeled DNA from:
Test organism(mol%)
NA10BT KSP3 DSM 13225
PHBV-degrading denitrifierStrain NA10BT 65.0 100 66 40Strain KSP3 64.7 66 100 47Strain KSP4 64.7 67 99 nt
Unidentified strain DSM 13225 64.5 45 43 100Acidovorax avenae subsp. avenae DSM 7227T 69.1 9 nt a 9Acidovorax delafeldii DSM 64T 65.6 11 nt 12Acidovorax facilis DSM 649T 64.9 9 nt 11Comamonas testosteroni IAM 12419T 61.7 8 nt 9Delftia acidovorans IAM 12409T 67.0 8 nt 8
a nt, not tested.
Table 2. Whole-cell fatty acid profiles of the PHBV-degrading isolates and strain DSM 13225.
Composition (%)a
ComponentNA10BT KSP3 KSP4 DSM 13225
10:0 0.3 0.4 0.4 0.412:0 2.7 2.5 2.5 3.314:0 2.2 2.1 2.1 3.315:0 0.6 0.7 0.7 0.316:0 29.2 29.2 28.7 36.116:1w7cis 40.0 42.6 43.2 35.417:0 0.2 0.3 0.3 0.117:0 cyclo 1.4 0.8 0.7 4.418:0 0.1 0.2 0.2 0.118:1w7cis 18.1 16.1 16.7 11.911 methyl 18:1w7cis 0.2 0.4 0.4 019:0 cyclo w8cis 0 0 0 0.13-OH-C10:0 4.6 4.3 3.8 4.5
a Components accounting for more than 0.1% of the total content in any test strain are shown.

tested (Table 3). These results for Acidovorax strainsagree well with those of previous studies (Schloe etal., 2000; Schulze et al., 1999; Willems et al., 1990).
HPLC experiments indicated that ubiquinone-8 (Q-8) accounted for 98 to 99% of the total quinone con-tents of the new isolates and strain DSM 13225 regardless of cultural conditions. The remainder de-tected was Q-7. No other quinone ring groups, such asmenaquinones and rhodoquinones, were detected.
PHBV degradation and denitrificationConsidering the phylogenetic and phenotypic simi-
larities between the new isolates and strain DSM13225, these organisms were compared as to the ca-pacity for hydrolysis of extracellular PHB and PHBVand denitrification. While all of the three new isolateswere capable of degrading PHB and PHBV, strainDSM 13225 was not. Among the reference strains ofComamonadaceae species tested, Acidovorax facilisDSM 649T, Acidovorax delafieldii DSM 64T, and Co-mamonas testosteroni JCM 10170 were able to de-grade PHB and PHBV. The capacity for denitrificationwas found in all the new isolates, strain DSM 13225,and B. denitrificans AS-P1T, whereas this characterwas absent in the type strains of the Acidovorax, Co-mamonas, Delftia, and Rhodoferax species tested.
Other characteristicsThe new isolates and strain DSM 13225 shared a
number of phenotypic characteristics as follows. They
were mesophilic and neutrophilic. The temperaturerange for growth was 20 to 40°C (optimum 28–35°C)and the pH range was 5 to 9 (optimum pH 7.5–8.0).Catalase and cytochrome oxidase were present. Hy-drolytic activities against starch, casein, gelatin, andDNA were absent. Good carbon sources for aerobicgrowth were ethanol, acetate, propionate, butyrate,pyruvate, gluconate, glutamate, asparagine, and a-alanine. Moderate growth was found with caproate, fumarate, succinate, and b-alanine. However, little orno growth occurred with sugars including L-arabinose,D-ribose, D-xylose, D-fructose, D-glucose, D-galactose,D-mannose, cellobiose, lactose, sucrose, trehalose,mannitol, and sorbitol. Other organic compounds notutilized were methanol, caprylate, citrate, benzoate,serine, and histidine. These trends obtained for carbonnutrition were almost the same under anaerobic condi-tions when nitrate was added as a terminal oxidant. Asshown in Table 4, however, the new isolates differedfrom strain DSM 13225 in Tween hydrolysis and uti-lization of chlorobenzene and malonate in addition toPHB and PHBV degradation.
Discussion
As reported here and previously (Khan and Hiraishi,2001; Khan et al., 2002), the phylogenetic data clearlyindicate that the new isolates, NA10BT, KSP3 andKSP4, and strain DSM 13225, should be classified intoa single genus within the family Comamonadaceae.The chemotaxonomic profiles, i.e., the presence ofC16:1 and C16:0 as the major components of whole-cell fatty acids and Q-8 as the major respiratoryquinone, justify their placement in this family. At pres-ent, the family Comamonadaceae consists of 16 gen-
304 KHAN and HIRAISHI Vol. 48
Table 4. Differential phenotypic characteristics of the PHBV-degrading isolates and
strain DSM 13225.
NA10BT,DSM
Characteristic KSP3,KSP4
13225
PHB and PHBV degradation � �
Hydrolysis of Tween 20 and Tween 80 � �
Carbon source:Malonate � �
1-Chloro-4-nitrobenzene � �
Table 3. 3-Hydroxy fatty acid profiles of the PHBV-degradingdenitrifying strains, strains DSM 13225, and
related organisms.
Composition (%)Test organism
3-OH-C8:0 3-OH-C10:0
PHBV-degrading denitrifierStrain NA10BT 0 100Strain KSP3 0 100Strain KSP4 0 100
Unidentified strain DSM 13225 0 100Acidovorax defluvii DSM 12644T 40 60Acidovorax delafeldii DSM 64T 35 65Acidovorax facilis DSM 649T 34 66Acidovorax temperans DSM 7270T 31 69Comamonas testosteroni IAM 12419T 0 100Delftia acidovorans IAM 12409T 25 75

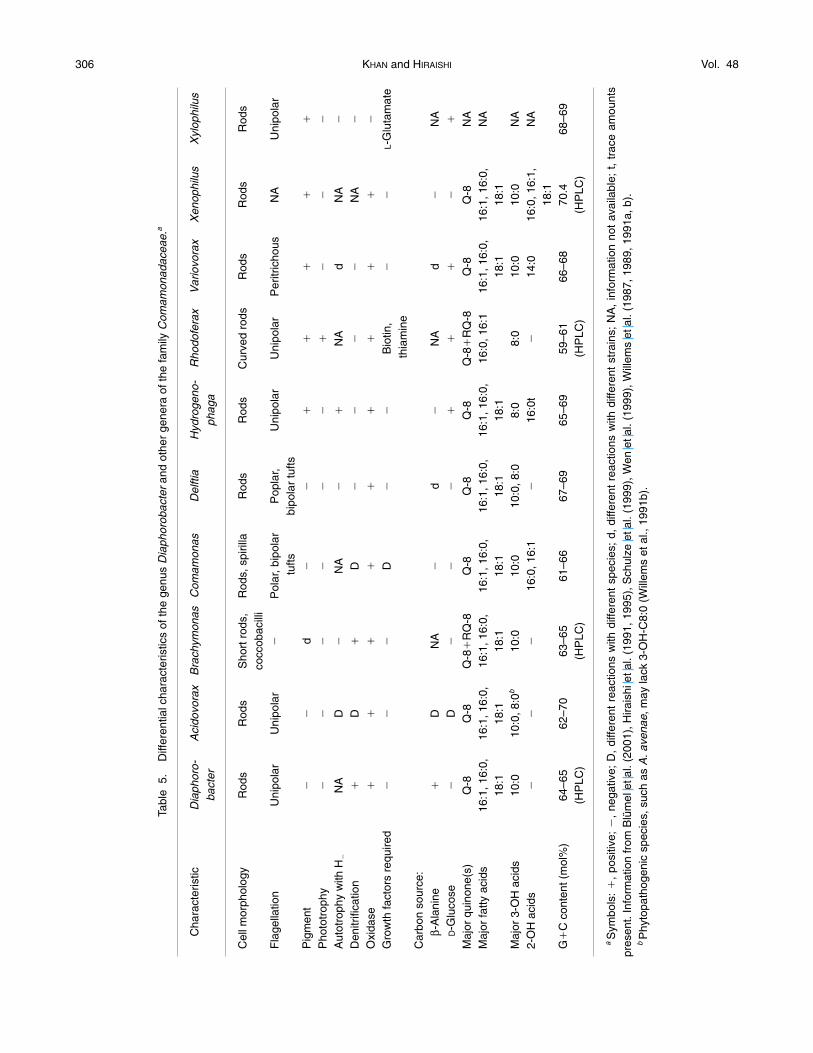
era, including the recently described genus Xenophilus(Blümel et al., 2001). Although the new isolates arephenotypically similar to members of some genera ofthe Comamonadaceae, in particular to denitrifyingspecies of the genera Acidovorax (Willems et al.,1990) and Comamonas (Etchebehere et al., 2001; Gu-maelius et al., 2001), the isolates form a phylogeneti-cally tight cluster distinguishable not only from thesetwo genera but also from any other established generaof this family. The average values of 16S rDNA se-quence similarity of the new isolates to each of theknown genera of the Comamonadaceae (e.g., Aci-dovorax, Comamonas, Delftia, and Xylophilus) are rel-atively equal, suggesting the difficulty of placing ournew isolates in a particular genus so far described.The phylogenetic neighbor of our isolates on the 16SrDNA distance matrix tree is the group of the threemisnamed Aquaspirillum species (Ding and Yokota,2002). However, the bootstrap value for the cluster ofour isolates with the Aquaspirillum species as its sistergroup is too low to justify its monophyly as a singlegenus. In view of these phylogenetic data, togetherwith the phenotypic diagnostic features discussedbelow, it is logical to classify the new isolates, togetherwith strain DSM 13225, into a new genus of the familyComamonadaceae. Here we propose the name Di-aphorobacter gen. nov. to accommodate these bacte-ria and D. nitroreducens sp. nov. for the PHBV-degrad-ing denitrifiers, strains NA10BT, KSP3, and KSP4.
Strain DSM 13225 is phylogenetically closely relatedto D. nitroreducens, but DNA-DNA similarity levels be-tween the two are lower than those within the D. ni-troreducens strains. Also, strain DSM 13225 differsfrom D. nitroreducens in some phenotypic characteris-tics, e.g., PHBV degradation, Tween hydrolysis, and 1-chloro-4-nitrobenzene utilization. Therefore, it is appro-priate to classify strain DSM 13225 as another speciesof the genus Diaphorobacter. Nonetheless, we desig-nate strain DSM 13225 merely as a genospecies of Di-aphorobacter without a formal nomenclatural proposal,because this strain is the only strain available for thespecies at this time.
There is general agreement that coherent pheno-typic characteristics in addition to phylogenetic evi-dence are essential for both description and recogni-tion of a new genus of bacteria. We have found nophenotypic traits specific to the new genus Di-aphorobacter. However, this genus can be differenti-ated from all of the previously known genera of the
family Comamonadaceae by a combination of a num-ber of phenotypic characteristics (Table 5). One of thecharacteristic features of Diaphorobacter is the lack of3-OH-C8:0 and 2-OH acids as cellular fatty acid com-ponents, which is a useful chemotaxonomic criterionfor differentiating this genus from Acidovorax, Coma-monas, and Delftia (also see Tables 2 and 3). In addi-tion, Diaphorobacter can be differentiated from allspecies of Acidovorax, except A. defluvii (Schulze etal., 1999), by the inability to utilize sugars, including D-ribose and D-glucose; from Comamonas by the modeof flagellation; and from Delftia by denitrification andthe inability to utilize sugars. More clearly, the genusDiaphorobacter can be differentiated by major pheno-typic differences from all other genera of the Coma-monadaceae. Moreover, PHB and PHBV degradationunder both aerobic and anaerobic denitrifying condi-tions is a useful key character for the identification ofDiaphorobacter nitroreducens.
Descriptions
Description of Diaphorobacter gen. nov.Diaphorobacter (Di.a.pho.ro.bac’ter. Gr. adi. di-
aphoros, different, profitable; Gr. n. bacter, rod; M.L.masc. n. Diaphorobacter, distinguished and profitablerod, referring to usefulness in nitrogen removal).
Cells are straight rods with rounded ends. Motilewith single polar flagella. Gram negative. Aerobicchemoorganotroph having strictly respiratory type ofmetabolism with oxygen as the terminal electron ac-ceptor. Anaerobic growth with nitrate as the terminaloxidant is also possible. Denitrification positive. Nogrowth factor required. Catalase and cytochrome oxi-dase are produced. Amylolytic and proteolytic activitiesare absent. Simple organic compounds including b-alanine, but not sugars, are utilized as carbon sources.The major cellular fatty acids are C16:1w7cis andC16:0. 3-OH-C10:0 is present as the sole componentof hydroxy fatty acids. 2-OH acids are absent.Ubiquinone-8 is the major respiratory quinone. TheG�C content of genomic DNA ranges from 64 to 65mol% (HPLC). 16S rDNA sequence information placesthe genus in the family Comamonadaceae. Typespecies: Diaphorobacter nitroreducens.
Description of Diaphorobacter nitroreducens sp. nov.Diaphorobacter nitroreducens (nitro.re.du’cens. L. n.
nitrum nitrate; L. part. adj. reducens converting to a dif-
2002 Diaphorobacter nitroreducens gen. nov., sp. nov. 305
306 KHAN and HIRAISHI Vol. 48
Tabl
e5.
Diff
eren
tial c
hara
cter
istic
s of
the
genu
s D
iaph
orob
acte
ran
d ot
her
gene
ra o
f the
fam
ily C
omam
onad
acea
e.a
Cha
ract
eris
ticD
iaph
oro-
Aci
dovo
rax
Bra
chym
onas
Com
amon
asD
elfti
aH
ydro
geno
-R
hodo
fera
xV
ario
vora
xX
enop
hilu
sX
ylop
hilu
sba
cter
phag
a
Cel
l mor
phol
ogy
Rod
sR
ods
Sho
rt r
ods,
Rod
s, s
piril
laR
ods
Rod
sC
urve
d ro
dsR
ods
Rod
sR
ods
cocc
obac
illi
Fla
gella
tion
Uni
pola
rU
nipo
lar
�P
olar
, bip
olar
Pop
lar,
Uni
pola
rU
nipo
lar
Per
itric
hous
NA
Uni
pola
rtu
ftsbi
pola
r tu
ftsP
igm
ent
��
d�
��
��
��
Pho
totr
ophy
��
��
��
��
��
Aut
otro
phy
with
H�
NA
D�
NA
��
NA
dN
A�
Den
itrifi
catio
n�
D�
D�
��
�N
A�
Oxi
dase
��
��
��
��
��
Gro
wth
fact
ors
requ
ired
��
�D
��
Bio
tin,
��
L-G
luta
mat
eth
iam
ine
Car
bon
sour
ce:
b-A
lani
ne�
DN
A�
d�
NA
d�
NA
D-G
luco
se�
D�
��
��
��
�
Maj
or q
uino
ne(s
)Q
-8Q
-8Q
-8�
RQ
-8Q
-8Q
-8Q
-8Q
-8�
RQ
-8Q
-8Q
-8N
AM
ajor
fatty
aci
ds16
:1, 1
6:0,
16:1
, 16:
0,16
:1, 1
6:0,
16:1
, 16:
0,16
:1, 1
6:0,
16:1
, 16:
0,16
:0, 1
6:1
16:1
, 16:
0,16
:1, 1
6:0,
NA
18:1
18:1
18:1
18:1
18:1
18:1
18:1
18:1
Maj
or 3
-OH
aci
ds10
:010
:0, 8
:0b
10:0
10:0
10:0
, 8:0
8:0
8:0
10:0
10:0
NA
2-O
H a
cids
��
�16
:0, 1
6:1
�16
:0t
�14
:016
:0, 1
6:1,
NA
18:1
G�
C c
onte
nt (
mol
%)
64–6
562
–70
63–6
561
–66
67–6
965
–69
59–6
166
–68
70.4
68–6
9(H
PLC
)(H
PLC
)(H
PLC
)(H
PLC
)
aS
ymbo
ls:
�,
posi
tive;
�,
nega
tive;
D,
diffe
rent
rea
ctio
ns w
ith d
iffer
ent
spec
ies;
d,
diffe
rent
rea
ctio
ns w
ith d
iffer
ent
stra
ins;
NA
, in
form
atio
n no
t av
aila
ble;
t,
trac
e am
ount
spr
esen
t. In
form
atio
n fr
om B
lüm
el e
t al.
(200
1), H
irais
hi e
t al.
(199
1, 1
995)
, Sch
ulze
et a
l. (1
999)
, Wen
et a
l. (1
999)
, Will
ems
et a
l. (1
987,
198
9, 1
991a
, b).
bP
hyto
path
ogen
ic s
peci
es, s
uch
as A
. ave
nae,
may
lack
3-O
H-C
8:0
(Will
ems
et a
l., 1
991b
).

ferent state; N. L. adj. nitroreducens reducing nitrate).Cells are 0.7–0.9 mm wide and 1.0 to 1.8 mm long.
Colonies on agar media are rough with uneven mar-gins or smooth and entire margins. The color of thecolonies is transparent in earlier stages and turns paleto beige after longer incubation. Poly(3-hydroxybu-tyrate) and poly(3-hydroxybutyrate-co-hydroxyvaler-ate) are degraded under aerobic and anaerobic denitri-fying conditions. Hydrolysis of starch, gelatin, casein,DNA, Tween 20 and Tween 80 is negative. b-Galac-tosidase and b-xylosidase are absent. Mesophilic andneutrophilic. The optimum temperature for growth is 28to 35°C. The optimum pH ranges from 7.0 to 8.0. Carbon sources utilized are ethanol, acetate, propi-onate, butyrate, 3-hydroxybutyrate, caproate, gluconate,pyruvate, succinate, fumarate, a -alanine, asparagine,glutamate, casamino acids, peptone, and yeast extract.Not utilized are sugars, sugar alcohols, methanol,aminobutyrate, benzoate, caprylate, citrate, malonate,tartrate, histidine, leucine, serine, and 1-chloro-4-ben-zene. The G�C content of genomic DNA ranges from64 to 65 mol%. The habitat is activated sludge. Thetype strain is strain NA10B, which has been depositedwith the Japan Collection of Microorganisms (RIKEN),Wako, Japan, as JCM 11421 and with the Collection ofBacterial Strains of Institut Pasteur as CIP 107294.The accession number for the 16S rRNA gene se-quence of the type strain is AB064317.
Acknowledgments
We are grateful to Japan Monsanto Co., Tokyo, Japan, forproviding PHB and PHBV and M. Yamamoto and Y. Horiba forearly contributions to this study. We also thank Y. Matsuzawaand T. Kanbe for their guidance in electron microscopy. Thiswork was supported in part by a grand-in-aid from the Ministryof Education, Culture, Sports, Science and Technology, Japan(No. 14390028).
References
Biedermann, J., Owen, A. J., Schloe, K. T., Gassner, F., andSubmuth, R. (1997) Interaction between poly-3-hydroxybu-tyrate-co-3-hydroxyvalerate and a denitrifying Pseudomonasstrain. Can. J. Microbiol., 43, 561–568.
Blümel, S., Busse, H.-J., Stolz, A., and Kämpfer, P. (2001).Xenophilus azovorans gen. nov., sp. nov., a soil bacteriumthat is able to degrade azo dyes of the orange II type. Int.J. Syst. Evol. Microbiol., 51, 1831–1837.
Boley, A., Müller, W. R., and Haider, G. (2000) Biodegradablepolymers as solid substrate and biofilm carrier for denitrifi-cation in recirculated aquaculture systems. Aquacul. Eng.,
22, 75–86.Ding, L.-X. and Yokota, A. (2002) Phylogenetic analysis of the
genus Aquaspirillum based on 16S rRNA gene sequences.FEMS Microbiol. Lett., 212, 165–169.
Etchebehere, C., Errazquin, M. I., Dabert, P., Moletta, R., andMuxi, L. (2001) Comamonas nitrativorans sp. nov., a noveldenitrifier isolated from a denitrifying reactor treating landfillleachate. Int. J. Syst. Evol. Microbiol., 51, 977–983.
Felsenstein, J. (1985) Confidence limits on phylogenies: an ap-proach using the bootstrap. Evolution, 39, 783–791.
Gumaelius, L., Magnusson, G., Pettersson, B., and Dalhammar,G. (2001) Comamonas denitrificans sp. nov., an efficientdenitrifying bacterium isolated from activated sludge. Int. J.Syst. Evol. Microbiol., 51, 999–1006.
Hiraishi, A., Hoshino, Y., and Satoh, T. (1991) Rhodoferax fer-mentans gen. nov., sp. nov., a phototrophic purple nonsul-fur bacterium previously referred to as the “Rhodocyclusgelatinosus-like” group. Arch. Microbiol., 155, 330–336.
Hiraishi, A. and Kitamura, H. (1984) Distribution of phototrophicpurple nonsulfur bacteria in activated sludge systems andother aquatic environments. Bull. Jpn. Soc. Sci. Fish., 50,1929–1937.
Hiraishi, A., Shin, Y. K., and Sugiyama, J. (1995) Brachymonasdenitrificans gen. nov., sp. nov., a rhodoquinone-containingchemoorganotrophic bacterium, and evolutionary relation-ships of rhodoquinone producers to bacterial members withvarious quinone classes. J. Gen. Appl. Microbiol., 41, 99–117.
Hiraishi, A., Shin, Y. K., Sugiyama, J., and Komagata, K. (1992)Isoprenoid quinones and fatty acids of Zoogloea. Antonievan Leeuwenhoek, 61, 231–236.
Hiraishi, A., Ueda, Y., Ishihara, J., and Mori, T. (1996) Compara-tive lipoquinone analysis of influent sewage and activatedsludge by high-performance liquid chromatography andphotodiode array detection. J. Gen. Appl. Microbiol., 42,457–469.
Katayama-Fujimura, Y., Komatsu, Y., Kuraishi, H., and Kaneko,T. (1984) Estimation of DNA base composition by high per-formance liquid chromatography of its nuclease P1 hy-drolysate. Agric. Biol. Chem., 48, 3169–3172.
Katsivela, E., Wray, V., Pieper, D. H., and Wittich, R.-M. (1999)Initial reactions in the biodegradation of 1-chloro-4-ni-trobenzene by a newly isolated bacterium, strain LW1.Appl. Environ. Microbiol., 65, 1405–1412.
Khan, S. T. and Hiraishi, A. (2001) Isolation and characteriza-tion of a new poly(3-hydroxybutyrate)-degrading, denitrify-ing bacterium from activated sludge. FEMS Microbiol. Lett.,205, 253–257.
Khan, S. T., Horiba, Y., Yamamoto, M., and Hiraishi, A. (2002)Members of the family Comamonadaceae as primarypoly(3-hydroxybutyrate-co-3-hydroxyvalerate)-degradingdenitrifiers in activated sludge as revealed by a polyphasicapproach. Appl. Environ. Microbiol., 68, 3206–3214.
Kimura, M. (1980) A simple method for estimating evolutionaryrates of base substitution through comparative studies of
2002 Diaphorobacter nitroreducens gen. nov., sp. nov. 307

nucleotide sequences. J. Mol. Evol., 16, 111–120.Magee, C. M., Rodeheaver, G., and Edgerton, R. F. (1975) A
more reliable Gram staining technique for diagnosis of sur-gical infections. Am. J. Surg., 130, 341–346.
Marmur, J. (1961) A procedure for the isolation of deoxyribonu-cleic acid from micro-organisms. J. Mol. Biol., 3, 208–218.
Mergaert, J., Boley, A., Cnockaert, M. C., Müller, W. R., andSwings, J. (2001) Identity and potential functions of het-erotrophic bacterial isolates from a continuous-upflowfixed-bed reactor for denitrification of drinking water withbacterial polyester as source of carbon and electron donor.Syst. Appl. Microbiol., 24, 303–310.
Müller, W. R., Heinemann, A., Schäfer, C., Wurmthaler, J., andReutter, T. (1992) Aspects of PHA (poly-b-hydroxy-butyric-acid) as an H-donor for denitrification in water treatmentprocesses. Water Supply, 10, 79–90.
Page, R. D. M. (1996) TREEVIEW: An application to displayphylogenetic trees on personal computers. Comp. Applic.Biosci., 12, 357–358.
Saitou, N. and Nei, M. (1987) The neighbor-joining method: anew method for reconstructing phylogenetic trees. Mol.Biol. Evol., 4, 406–425.
Schloe, K., Gillis, M., Hoste, B., Pot, B., Vancanneyt, M., Mer-gaert, J., Swings, J., Biedermann, J., and Sussmuth, R.(2000) Polyphasic characterization of poly-3-hydroxybu-tyrate-co-3-hydroxyvalerate (P(HB-co-HV)) metabolisingand denitrifying Acidovorax sp. strains. Syst. Appl. Micro-biol., 23, 364–372.
Schulze, R., Spring, S., Amann, R., Huber, I., Ludwig, W.,Schleifer, K.-H., and Kämpfer, P. (1999) Genotypic diversityof Acidovorax strains isolated from activated sludge anddescription of Acidovorax defluvii sp. nov. Syst. Appl. Mi-crobiol., 22, 205–214.
Thompson, J. D., Higgins, D. G., and Gibson, T. J. (1994)CLUSTAL W: Improving the sensitivity of progressive multi-ple sequence alignment through sequence weighting, posi-
tion-specific gap penalties and weight matrix choice. Nu-cleic Acids Res., 22, 4673–4680.
Wen, A., Fegan, M., Hayward, C., Chakraborty, S., and Sly, I. L.(1999) Phylogenetic relationships among members of theComamonadaceae, and description of Delftia acidovorans(den Dooren de Jong 1926 and Tamaoka et al., 1987) gen.nov., comb. nov. Int. J. Syst. Bacteriol., 49, 567–576.
Willems, A., Busse, J., Goor, M., Pot, B., Falsen, E., Jantzen,E., Hoste, B., Gillis, M., Kersters, K., Auling, G., and De Ley, J. (1989) Hydrogenophaga, a new genus of hydrogen-oxidizing bacteria that includes Hydrogenophagaflava comb. nov. (formerly Pseudomonas flava), Hy-drogenophaga palleronii (formerly Pseudomonas palleronii),Hydrogenophaga pseudoflava (formerly Pseudomonaspseudoflava and “Pseudomonas carboxydoflava”) and Hy-drogenophaga taeniospiralis (formerly Pseudomonas tae-niospiralis). Int. J. Syst. Bacteriol., 39, 319–333.
Willems, A., De Ley, J., Gillis, M., and Kersters, K. (1991) Co-mamonadaceae, a new family encompassing the acidovo-rans rRNA complex, including Variovorax paradoxus gen.nov. comb. nov., for Alcaligenes paradoxus (Davis 1969).Int. J. Syst. Bacteriol., 41, 427–444.
Willems, A., Falsen, E., Pot, B., Jantzen, E., Hoste, B., Van-damme, P., Gillis, M., Kersters, K., and De Ley, J. (1990)Acidovorax, a new genus for Pseudomonas facilis,Pseudomonas delafiedii, E. Falsen (EF) Group 13, EFGroup 16, and several clinical isolates, with the speciesAcidovorax facilis comb. nov., Acidovorax delafieldii comb.nov., and Acidovorax temperans sp. nov. Int. J. Syst. Bac-teriol., 40, 384–398.
Willems, A., Gillis, M., Kersters, K., Van Den Broecke, L., andDe Ley, J. (1987) Transfer of Xanthomonas ampelinaPanagopoulos 1969 to a new genus, Xylophilus gen. nov.,as Xylophilus ampelinus (Panagopoulos 1969) comb. nov.Int. J. Syst. Bacteriol., 37, 422–430.
308 KHAN and HIRAISHI Vol. 48




















