
Developmental Responses to Temperature and Photoperiod in Ecotypes of Medicago polymorpha L....
6
Developmental Responses to Temperature and Photoperiod in Ecotypes of Medicago polymorpha L. Collected Along an Environmental Gradient in Central Chile ALEJANDRO DEL POZO* {, CARLOS OVALLE {, JAMES ARONSON } and JULIA AVENDAN ˜ Ok {Departamento de Produccio ´n Vegetal, Facultad de Agronomı´a, Universidad de Concepcio ´n, Casilla 537, Chilla ´n, Chile, {Centro Regional de Investigacio ´n Quilamapu, Instituto de Investigaciones Agropecuarias (INIA), Casilla 426, Chilla ´n, Chile, }CNRS UPR 9056, CEFE, 1919, Route de Mende, 34293 Montpellier cedex 5, France and kCentro Experimental Cauquenes, Instituto de Investigaciones Agropecuarias (INIA), Casilla 165, Cauquenes, Chile Received: 27 September 1999 Returned for revision: 10 November 1999 Accepted: 10 February 2000 The phenological development of nine Chilean accessions of Medicago polymorpha, collected along a north–south aridity gradient, and of two commercial cultivars of the same species, were compared in 12 sequential outdoor sowings at Cauquenes (35858 0 S, 72817 0 W, elev. 177 m), in the sub-humid Mediterranean climate zone of Chile. A glasshouse experiment was also conducted to evaluate the eect of photoperiod on phenophase timing. There was a clear gradient in precocity among the Chilean accessions in both experiments: accessions MPO-9-88 and MPO-7-88, from the arid zone, were the earliest-flowering accessions, whereas MPO-36-88 from the humid Mediterranean zone was the latest. Both experiments revealed significant variation among the Chilean accessions in the response of flowering time to variation in photoperiod regime. Dierences in days to flowering between the least- (8 h) and the most- (16 h) inductive photoperiods were lower in precocious accessions from arid and semi-arid zones, than in late- flowering accessions from more humid zones. Rate of progress to flowering, defined as the inverse of time from emergence to first flower appearance (1/f ), was related to mean diurnal temperature, or to both mean diurnal temperature and mean photoperiod. In two early-flowering accessions from the arid zone, and in the Australian cultivar ‘Circle Valley’, 1/f was aected significantly (P 5 0 . 05) by both temperature and photoperiod. In the remaining accessions, no significant responses to temperatures were detected; 1/f was influenced significantly by photoperiod only. # 2000 Annals of Botany Company Key words: Annual medic, aridity gradient, Medicago polymorpha, flowering time, rate of development. INTRODUCTION The circum-Mediterranean burr medic (Medicago polymor- pha L.) is widely naturalized throughout the Mediterra- nean-climate region of central Chile (Del Pozo et al., 1989) and is locally considered to be the most valuable nitrogen- fixing legume for pasture improvement (Avendan˜o and Ovalle, 1992), and ley-farming style rotations with cereal or other annual field crops (Ovalle et al., 1990; Del Pozo et al., 1999). This species is currently sown on about 1 million ha year 1 in Western Australia (Ewing and Howieson, 1989; Ewing, 1999) and is increasingly grown in other countries with a Mediterranean-type climate. Yet little is known of its ecophysiology, adaptive strategies under varying environ- mental conditions, or range of genetic expression. Ehrman and Cocks (1990, 1996) did include burr medic in their ecogeographic study of 71 annual legumes in Syria, and an in-depth study of this species has been carried out in Sardinia (Loi et al., 1993, 1995). Some relevant ecological and agronomic literature exists on several other annual medics and clovers (e.g. Clarkson and Russell, 1975, 1979; Junttila et al., 1990a, b; Piano et al., 1996), but much remains to be discovered about naturalized annual legume species in Chile, and M. polymorpha in particular. It is in Chile that some of the most promising commercial accessions have originated (Ewing and Howieson, 1989), and where M. polymorpha, and eight other annual medic species, occur along a pronounced, 1000-km-long, aridity gradient, which is highly conducive to rapid ecotypic dierentiation. Advancing knowledge on diversity and recurrent patterns in populations of burr medic along this gradient will be of great importance not only to annual legume breeders, but also to theoretical ecologists. Medicago spp. have only been present in Chile since the late 16th or early 17th century, when they were inadver- tently introduced from southern Spain or Portugal, in the coats of livestock or mixed with seeds or hay carried by ships. Despite its relatively short history in Chile (approx. 400–450 years), the great range of bioclimatic and edaphic conditions in which this species now occurs has generated clear ecotypic dierentiation of many adaptive traits, including reproductive phenology and winter vigour, as well as spiny vs. non-spiny pods (Del Pozo et al., 1989). There is a clear gradient in flowering precocity among burr medic accessions along the natural aridity gradient in the Mediterranean-climate region of Chile, with early- flowering (precocious) populations occurring at the drier, northern end, and late-flowering ecotypes at the southern, sub-humid end (Ovalle et al., 1993, 1997). This paper Annals of Botany 85: 809–814, 2000 doi:10.1006/anbo.2000.1141, available online at http://www.idealibrary.com on 0305-7364/00/060809+06 $35.00/00 # 2000 Annals of Botany Company * For correspondence. Fax 56-42-227517, [email protected]
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Developmental Responses to Temperature and Photoperiod in Ecotypes of Medicago polymorpha L....

Annals of Botany 85: 809±814, 2000doi:10.1006/anbo.2000.1141, available online at http://www.idealibrary.com on
Developmental Responses to Temperature and Photoperiod in Ecotypes of Medicago
pokCentro E
species in
0305-7364/
* For corr
lymorpha L. Collected Along an Environmental Gradient in Central Chile
ALEJANDRO DEL P , JAMES ARONSON}
xperimental Cauquenes, Institu
Received: 27 September 1999 R
Key words: Annual medic, aridity gr
Chile, and M. polymorpha in pa
00/060809+06 $35.00/00
espondence. Fax �56-42-227517, e-ma
OZO*{, CARLOS OVALLE{
and JULIA AVENDANÄ Ok{Departamento de ProduccioÂn Vegetal, Facultad de AgronomõÂa, Universidad de ConcepcioÂn, Casilla 537, ChillaÂn,Chile, {Centro Regional de InvestigacioÂn Quilamapu, Instituto de Investigaciones Agropecuarias (INIA), Casilla 426,
ChillaÂn, Chile, }CNRS UPR 9056, CEFE, 1919, Route de Mende, 34293 Montpellier cedex 5, France and
to de Investigaciones Agropecuarias (INIA), Casilla 165, Cauquenes, Chileeturned for revision: 10 November 1999 Accepted: 10 February 2000
The phenological development of nine Chilean accessions of Medicago polymorpha, collected along a north±southaridity gradient, and of two commercial cultivars of the same species, were compared in 12 sequential outdoorsowings at Cauquenes (358580S, 728170W, elev. 177 m), in the sub-humid Mediterranean climate zone of Chile. Aglasshouse experiment was also conducted to evaluate the e�ect of photoperiod on phenophase timing. There was aclear gradient in precocity among the Chilean accessions in both experiments: accessions MPO-9-88 and MPO-7-88,from the arid zone, were the earliest-¯owering accessions, whereas MPO-36-88 from the humid Mediterranean zonewas the latest. Both experiments revealed signi®cant variation among the Chilean accessions in the response of¯owering time to variation in photoperiod regime. Di�erences in days to ¯owering between the least- (8 h) and themost- (16 h) inductive photoperiods were lower in precocious accessions from arid and semi-arid zones, than in late-¯owering accessions from more humid zones. Rate of progress to ¯owering, de®ned as the inverse of time fromemergence to ®rst ¯ower appearance (1/f), was related to mean diurnal temperature, or to both mean diurnaltemperature and mean photoperiod. In two early-¯owering accessions from the arid zone, and in the Australiancultivar `Circle Valley', 1/f was a�ected signi®cantly (P5 0.05) by both temperature and photoperiod. In theremaining accessions, no signi®cant responses to temperatures were detected; 1/f was in¯uenced signi®cantly byphotoperiod only. # 2000 Annals of Botany Company
adient, Medicago polymorpha, ¯owering time, rate of development.
INTRODUCTION
The circum-Mediterranean burr medic (Medicago polymor-pha L.) is widely naturalized throughout the Mediterra-nean-climate region of central Chile (Del Pozo et al., 1989)and is locally considered to be the most valuable nitrogen-®xing legume for pasture improvement (AvendanÄ o andOvalle, 1992), and ley-farming style rotations with cereal orother annual ®eld crops (Ovalle et al., 1990; Del Pozo et al.,1999). This species is currently sown on about 1 million hayearÿ1 in Western Australia (Ewing and Howieson, 1989;Ewing, 1999) and is increasingly grown in other countrieswith a Mediterranean-type climate. Yet little is known of itsecophysiology, adaptive strategies under varying environ-mental conditions, or range of genetic expression. Ehrmanand Cocks (1990, 1996) did include burr medic in theirecogeographic study of 71 annual legumes in Syria, and anin-depth study of this species has been carried out inSardinia (Loi et al., 1993, 1995). Some relevant ecologicaland agronomic literature exists on several other annualmedics and clovers (e.g. Clarkson and Russell, 1975, 1979;Junttila et al., 1990a, b; Piano et al., 1996), but muchremains to be discovered about naturalized annual legume
rticular. It is in
Chile that some of the most promising commercialaccessions have originated (Ewing and Howieson, 1989),and where M. polymorpha, and eight other annual medicspecies, occur along a pronounced, 1000-km-long, ariditygradient, which is highly conducive to rapid ecotypicdi�erentiation. Advancing knowledge on diversity andrecurrent patterns in populations of burr medic along thisgradient will be of great importance not only to annuallegume breeders, but also to theoretical ecologists.
Medicago spp. have only been present in Chile since thelate 16th or early 17th century, when they were inadver-tently introduced from southern Spain or Portugal, in thecoats of livestock or mixed with seeds or hay carried byships. Despite its relatively short history in Chile (approx.400±450 years), the great range of bioclimatic and edaphicconditions in which this species now occurs has generatedclear ecotypic di�erentiation of many adaptive traits,including reproductive phenology and winter vigour, aswell as spiny vs. non-spiny pods (Del Pozo et al., 1989).
There is a clear gradient in ¯owering precocity amongburr medic accessions along the natural aridity gradient inthe Mediterranean-climate region of Chile, with early-¯owering (precocious) populations occurring at the drier,northern end, and late-¯owering ecotypes at the southern,
sub-humid end (Ovalle et al., 1993, 1997). This paper# 2000 Annals of Botany Company

for theoretical and applied research.
emergence to ®rst ¯ower was taken to be 111 d.
en
addresses the speci®c environmental factors, especiallytemperature and photoperiod, which presumably a�ectecotypic and, to the extent it exists, genotypic di�er-entiation of reproductive phenology.
In Mediterranean-type climates, photoperiod (daylength)is considered to be an important environmental signal for¯ower initiation (Evenari and Gutterman, 1985). In someMedicago (Clarkson and Russell, 1975, 1979) and alsoTrifolium (Evans et al., 1992) species, photoperiod andtemperature strongly a�ect time to ¯owering, but not in thesame way for all species. Both temperature and photoperioddi�er greatly along the N±S gradient in central Chile, andtherefore accessions of a single, polymorphic Medicagospecies, collected along this gradient, may well showmarkedly-di�erent developmental responses to temperatureand photoperiod. In an experimental study of three wide-spread annuals occurring along a desert-Mediterraneantransect in Israel, photoperiod requirement for ¯oweringinduction decreased with increasing aridity and rainfallunpredictability (Aronson et al., 1992). Conversely, popu-lations of the same species, occurring in more stable andpredictable environments on the aridity gradient, respondedto both temperature and photoperiod. This study investi-gates how naturalized annual legumes respond to temp-erature and photoperiod along a longer, but also highlypronounced, aridity gradient i.e. the Mediterranean-climateregion of central Chile.
For long-day plants, the rate of progress to ¯owering (1/f,where f is days from emergence to ®rst ¯ower) increaseswhen the actual photoperiod (P) is greater than a `ceiling'photoperiod (Pce) and shorter than a `critical' photoperiod(Pc) (Summer®eld et al., 1991). Thus, photoperiod require-ments for ¯owering in populations from di�erent environ-ments may di�er due to di�erences in Pce or Pc , ordi�erences in responsiveness to daylength, i.e. the rate ofprogress to ¯owering.
This work tests the hypothesis that, despite its relativelyshort history in the country (probably 5450 years), theimpressive range of bioclimatic and edaphic conditionsunder which burr medic now occurs in Chile have promotedecotypic and, perhaps, genotypic di�erentiation in pheno-
810 Del Pozo et al.ÐPhenological Developm
logical response to varying temperature regime and
TABLE 1. Collection sites of nine Chilean accessions of Min centra
LatiAccession Collection site (8
MPO-9-88 Ovalle 308MPO-7-88 Combarbala 318MPO-1-88 Batuco 338MPO-21-88 Calleuque 348MPO-29-88 Chanco 358MPO-30-88 Santa Dolores 358MPO-43-88 Cauquenes 358MPO-33-88 CanÄ ete 378MPO-36-88 Traigue n 388
* According to di Castri and Hajek (1976); where the value in bracket
photoperiod, as well as other adaptive traits of interest
t in Ecotypes of Medicago polymorpha
MATERIALS AND METHODS
Glasshouse experiment
Nine Chilean accessions of the naturalized annual burrmedic Medicago polymorpha (Table 1) were grown in aheated glasshouse at ®ve photoperiods: 8, 10, 12, 14 and16 h of daylight. On 13 Aug. 1996, ten inoculated seeds ofeach accession were sown in 13 cm diameter pots ®lled with1 kg of a mixture of 1 :1 sand and a volcanic soil (12%organic matter, oven dry soil), previously fertilized with 90,77 and 69 mg kgÿ1 of N, P and K, respectively, and thinnedto ®ve seedlings after emergence. A completely-randomizeddesign was used, with a factorial arrangement and fourreplicate pots per accession and per photoperiod. Plantsreceived 8 h of natural light between 0900 and 1700 h, andwere then arti®cially illuminated with 0, 2, 4, 6 or 8 h oflight, provided by 60 W Plantilux light bulbs in darkchambers, from 21 August onwards. Mean minimum andmaximum temperatures during the experiment were 18 and288C, respectively. Phenological observations were carriedout on individual plants every 3 d. Flowering date wasrecorded when 60% of the plants in a pot had at least oneopen ¯ower. The mean time between emergence and theappearance of the ®rst ¯ower was then calculated for eachaccession and photoperiod. A two-way analysis of variancewas performed on days from emergence to ®rst ¯ower, andmeans were compared by LSD (log transformation was notrequired). In treatments where plants did not ¯ower beforethe end of the experiment (11 December), the period from
Sowing date experiment
The same nine accessions of burr medic, plus two widely-utilized Australian cultivars (`Circle Valley' and `Santiago')of the same species were sown outdoors in raised beds, ®lledwith 1 m of granitic soil, at Cauquenes, Chile (358580S,728170W, elev. 177 m), on 12 sequential sowing datesfrom May 1993 to April 1994. One row, 1.1 m long, was
sown to obtain 26 plants for each accession. There was oneedicago polymorpha along a north±south aridity gradientl Chile
tude LongitudeS) (8W) Climatic zone*
330 718070 Arid (134)090 718000 Arid (134)110 708460 Semi-arid (370)260 718270 Sub-humid (439)410 728290 Sub-humid (898)520 728110 Sub-humid (676)580 728190 Sub-humid (676)480 738230 Perhumid (1373)090 728410 Perhumid (1241)
s indicates mean annual rainfall in mm.

precocious accessions from arid and semi-arid regions.
TABLE 2. Number of days from emergence to ®rst ¯ower innine accessions of M. polymorpha grown in a glasshouse
under ®ve photoperiods
Photoperiod (h)
Accession 8 10 12 14 16Mean foraccession
MPO-9-88 64 49 47 36 24 46a
MPO-7-88 66 58 56 41 39 49a
MPO-1-88 85 72 59 43 35 59b
MPO-21-88 111* 98 55 58 47 74cd
MPO-29-88 108 92 77 53 49 76d
MPO-30-88 105 89 69 53 41 71c
MPO-43-88 106* 91 73 50 40 72cd
MPO-33-88 111* 95 84 53 38 76d
MPO-36-88 111* 91 79 50 47 76d
Mean forphotoperiod
96a 82b 66c 48d 40e
* Plants had not ¯owered by the end of the experiment (111 d afteremergence).
Means with di�erent superscripts are signi®cantly di�erent atP50.01.
ent in Ecotypes of Medicago polymorpha 811
raised bed (4 � 1.1 m) per sowing date and the distancebetween rows (accessions) was 40 cm. Inoculum diluted inwater (1 g lÿ1) was applied immediately after sowing, andthe same strain of Rhizobium meliloti was used for allaccessions. Fertilizer was mixed into the soil before sowingat a rate of 4.4 g P mÿ2, as superphosphate. Plants werewatered once or twice a week from August to April.
Phenological observations were carried out every 3 d on22±25 individual plants for each accession. Emergence datewas recorded when half of the plants in the row hademerged. Days from emergence to ®fth leaf, appearance ofthe ®rst ¯ower, appearance of ®rst pod, and plantsenescence were recorded for each plant, and the meantime between phenophases was calculated for each acces-sion and sowing date. Maximum and minimum tempera-tures were recorded daily at a meteorological station located1 km from the experimental site.
The rate of plant development (1/t), de®ned as the inverseof time (t) from emergence to succeeding phenophases, wasrelated to mean diurnal temperature (T, 8C) or to bothmean diurnal temperature and mean photoperiod (P,h dÿ1), using two linear models (Hadley et al., 1983; DelPozo et al., 1987; Summer®eld et al., 1991). For pheno-phases insensitive to photoperiod, the (model 1) relation-ship is:
1=t � a � bT �1�where intercept a and slope b are constant for eachphenophase and accession.
In photoperiod-sensitive phenophases (andPce5 P5 Pc),such as the rate of progress to ¯owering, the rate ofdevelopment can be described (model 2) as:
1=t � a0 � b0T � c0P �2�where Pce and Pc are the ceiling and critical photoperiods,respectively, and a0, b0 and c0 are constants speci®c to eachaccession (Summer®eld et al., 1991). For a `long-day' plantsuch as burr medic, the rate of progress to ¯owering (1/f)will be reduced if P 5 Pc , and c0 is positive (Summer®eldet al., 1985).
The constants of both models were estimated throughlinear regression, using Statgraphics Plus (1994), for eachaccession. For the rate of progress to ¯owering, both
Del Pozo et al.ÐPhenological Developm
models were compared for all accessions.
(Table 3).
RESULTS
E�ect of photoperiod on ¯owering time
In the glasshouse experiment, time to ¯owering showedgreat plasticity in all accessions: the number of days fromemergence to ®rst ¯ower decreased with increasing day-length, as expected in long-day plants. Under 8 h ofdaylength, four accessions did not ¯ower before the end ofthe experiment, suggesting that these accessions wereobligatory long-day plants; the other ®ve accessions, fromarid, semi-arid and sub-humid zones, behaved as facultativelong-day plants. Accessions from the arid and semi-arid
end of the transect were more precocious than accessionsfrom the humid end of the transect in the south, regardlessof photoperiod (Table 2). Moreover, di�erences in thenumber of days to ®rst ¯ower between the least- (8 h) andthe most- (16 h) inductive photoperiods were lower in
E�ect of sowing date on plant development
As sowing date was delayed from late autumn (May) toearly summer (December), the number of days to theappearance of the ®fth leaf and the ®rst ¯ower decreased forall accessions, but then increased for summer and autumnsowing dates in January to April (Fig. 1). A clear gradient inprecocity among the Chilean accessions was observedfor all sowing dates, including those where the photo-thermal conditions were the most- and the least-inductivefor ¯owering (Table 3). MPO-9-88 (Ovalle) and MPO-7-88(Combarbala ), from the arid region, were the earliest-¯owering accessions, whereas MPO-36-88 (Traigue n), fromthe humid end of the transect, was the latest. The number ofdays to pod maturation followed the same pattern amongpopulations along the desert±Mediterranean transect (datanot shown). Of the two Australian cultivars, `Santiago' wasthe earliest-¯owering cultivar, showing a similar response tosowing date to MPO-7-88 and MPO-1-88; `Circle Valley'was phenologically similar to MPO-30-88 (Santa Dolores)
Rate of development as a function of temperature andphotoperiod
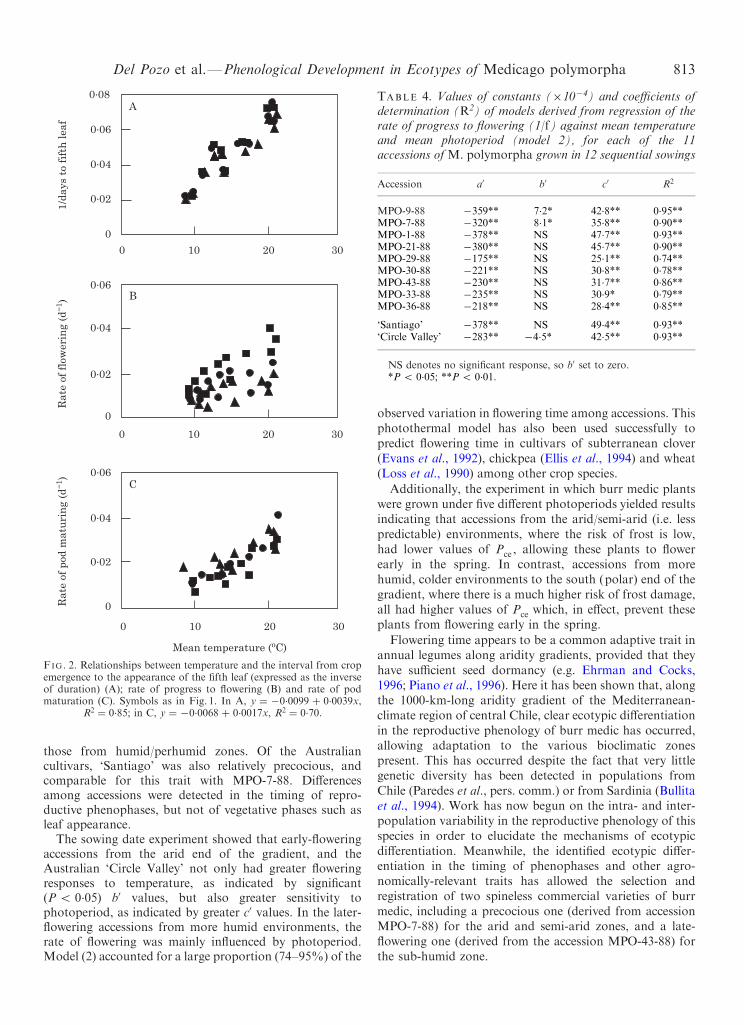
For all accessions, the interval from crop emergence tothe appearance of the ®fth leaf (expressed as the inverse ofduration) increased linearly with mean temperature
(Fig. 2A), as did time from ®rst ¯ower to plant senescence
environments.
60
50
40
30
20
10
0
200
180
160
140
120100
80
60
40
200
6/5 7/6 5/7 6/8 6/9 5/10 4/11 6/12 5/1 4/2 4/3 4/4
6/5 7/6 5/7 6/8 6/9 5/10 4/11 6/12 5/1 4/2 4/3 4/4
Sowing date
Day
s to
flo
wer
ing
Day
s to
fif
th le
af
A
B
FIG. 1. Variation in number of days to the appearance of the ®fth leaf(A) and number of days to ¯owering (B) of three accessions ofMedicago polymorpha sown outdoors on 12 sequential dates, atCauquenes, Chile (358580S, 728170W). (j), MPO-7-88; (d), MPO-
812 Del Pozo et al.ÐPhenological Developmen
(Fig. 2C). Depending on sowing date, the mean temperatureand mean photoperiod for the period from emergence to®rst ¯ower ranged from 9.1 to 218C, and from 9.8 to 14.4 h,
43-88; (m), MPO-36-88.
respectively (Table 2). The shortest mean photoperiod
TABLE 3. Phenological plasticity of nine Chilean acces
Most-inductive conditions*
Accession Sowing date f T (8C) P
MPO-9-88 06/12 24 20.3 14MPO-7-88 06/12 25 20.6 14MPO-1-88 06/12 29 20.7 14MPO-21-88 04/11 32 16.6 14MPO-30-88 06/12 40 21.0 14MPO-43-88 06/12 40 20.9 14MPO-29-88 06/12 49 21.1 14MPO-33-88 05/01 44 20.5 13MPO-36-88 06/12 49 21.0 14
`Santiago' 06/12 28 20.7 14`Circle Valley' 06/12 37 21.0 14
* The most- and least-inductive conditions are for the sowing date for wand highest, respectively.
Number of days from emergence to ¯owering ( f) under the most- and leconditions.
(9.8 h) in this experiment was greater than the ceilingphotoperiod (8 h) established for some accessions in theglasshouse experiment.
Temperature alone (model 1) explained only a smallproportion of the observed variation in rate of progress to¯owering (1/f ), except for the earliest-¯owering accessions(MPO-1-88 and MPO-7-88) and `Santiago', for which R2
was greater than 0.7 (P 5 0.01 in the 3 accessions)(Fig. 2B).
When temperature and photoperiod were both includedin the analysis (model 2), signi®cant responses of 1/f tomean temperature and photoperiod were detected in twoearly-¯owering accessions (MPO-9-88 and MPO-7-88) fromthe arid zone, and in the Australian `Circle Valley'(Table 4). In contrast, in accessions from the semi-arid,sub-humid and perhumid zones, and in `Santiago', theresponses of 1/f to temperature (expressed by b0 in model 2)were not signi®cant (P 4 0.05), whereas the e�ect ofphotoperiod (expressed by c0 in model 2) was signi®cant(Table 4). The parameter c0 was positive and highlysigni®cant for all accessions, indicating that rate of progressto ¯owering was accelerated by longer photoperiods. Early-¯owering accessions from the northern (equatorial), drierenvironments tended to have greater values of c0 than later-¯owering accessions from the more humid, southern (polar)
t in Ecotypes of Medicago polymorpha
DISCUSSION
Ecotypic di�erentiation of phenology has occurred in burrmedic (M. polymorpha) along the 1000 km-long Mediterra-nean gradient of aridity in central Chile. In the presentpaper, it has been shown that di�erences in ¯owering timeamong accessions of burr medic collected along this ariditygradient can be explained by di�erential sensitivity of eco-types to photoperiod and temperature. In particular, therewas a clear gradient in precocity among accessions in boththe glasshouse and the outdoor experiments: accessions
from the arid/semi-arid zones were more precocious thansions and two Australian cultivars of M. polymorpha
Least-inductive conditions*
(h) Sowing date f T (8C) P (h)
.4 06/05 84 9.1 9.8
.4 06/05 92 9.2 9.9
.4 06/05 100 9.3 10.0
.2 04/04 149 11.0 10.3
.2 04/04 141 10.6 10.2
.3 04/04 118 10.9 10.0
.2 04/04 147 10.5 10.2
.4 04/04 148 10.5 10.2
.2 04/04 152 10.5 10.2
.4 04/04 100 11.3 10.0
.4 04/04 108 11.2 10.0
hich the number of days from crop emergence to ¯owering was lowest
ast-inductive photothermal (mean temperature, T, and photoperiod, P)
0 10 20 30
0 10 20 30
0 10 20 30
Mean temperature (oC)
1/da
ys t
o fi
fth
leaf
0.08
0.06
0.04
0.02
0
0.06
0.04
0.02
0
0.06
0.04
0.02
0
Rat
e of
flo
wer
ing
(d−1
)R
ate
of p
od m
atu
rin
g (d
−1)
A
B
C
FIG. 2. Relationships between temperature and the interval from cropemergence to the appearance of the ®fth leaf (expressed as the inverseof duration) (A); rate of progress to ¯owering (B) and rate of podmaturation (C). Symbols as in Fig. 1. In A, y � ÿ0.0099 � 0.0039x,
R2 � 0.85; in C, y � ÿ0.0068 � 0.0017x, R2 � 0.70.
TABLE 4. Values of constants (�10ÿ4) and coe�cients ofdetermination (R2) of models derived from regression of therate of progress to ¯owering (1/f) against mean temperatureand mean photoperiod (model 2), for each of the 11accessions of M. polymorpha grown in 12 sequential sowings
Accession a0 b0 c0 R2
MPO-9-88 ÿ359** 7.2* 42.8** 0.95**MPO-7-88 ÿ320** 8.1* 35.8** 0.90**MPO-1-88 ÿ378** NS 47.7** 0.93**MPO-21-88 ÿ380** NS 45.7** 0.90**MPO-29-88 ÿ175** NS 25.1** 0.74**MPO-30-88 ÿ221** NS 30.8** 0.78**MPO-43-88 ÿ230** NS 31.7** 0.86**MPO-33-88 ÿ235** NS 30.9* 0.79**MPO-36-88 ÿ218** NS 28.4** 0.85**
`Santiago' ÿ378** NS 49.4** 0.93**`Circle Valley' ÿ283** ÿ4.5* 42.5** 0.93**
NS denotes no signi®cant response, so b0 set to zero.*P 5 0.05; **P 5 0.01.
Del Pozo et al.ÐPhenological Development in Ecotypes of Medicago polymorpha 813
those from humid/perhumid zones. Of the Australiancultivars, `Santiago' was also relatively precocious, andcomparable for this trait with MPO-7-88. Di�erencesamong accessions were detected in the timing of repro-ductive phenophases, but not of vegetative phases such asleaf appearance.
The sowing date experiment showed that early-¯oweringaccessions from the arid end of the gradient, and theAustralian `Circle Valley' not only had greater ¯oweringresponses to temperature, as indicated by signi®cant(P 5 0.05) b0 values, but also greater sensitivity tophotoperiod, as indicated by greater c0 values. In the later-¯owering accessions from more humid environments, therate of ¯owering was mainly in¯uenced by photoperiod.
Model (2) accounted for a large proportion (74±95%) of theobserved variation in ¯owering time among accessions. Thisphotothermal model has also been used successfully topredict ¯owering time in cultivars of subterranean clover(Evans et al., 1992), chickpea (Ellis et al., 1994) and wheat(Loss et al., 1990) among other crop species.
Additionally, the experiment in which burr medic plantswere grown under ®ve di�erent photoperiods yielded resultsindicating that accessions from the arid/semi-arid (i.e. lesspredictable) environments, where the risk of frost is low,had lower values of Pce , allowing these plants to ¯owerearly in the spring. In contrast, accessions from morehumid, colder environments to the south (polar) end of thegradient, where there is a much higher risk of frost damage,all had higher values of Pce which, in e�ect, prevent theseplants from ¯owering early in the spring.
Flowering time appears to be a common adaptive trait inannual legumes along aridity gradients, provided that theyhave su�cient seed dormancy (e.g. Ehrman and Cocks,1996; Piano et al., 1996). Here it has been shown that, alongthe 1000-km-long aridity gradient of the Mediterranean-climate region of central Chile, clear ecotypic di�erentiationin the reproductive phenology of burr medic has occurred,allowing adaptation to the various bioclimatic zonespresent. This has occurred despite the fact that very littlegenetic diversity has been detected in populations fromChile (Paredes et al., pers. comm.) or from Sardinia (Bullitaet al., 1994). Work has now begun on the intra- and inter-population variability in the reproductive phenology of thisspecies in order to elucidate the mechanisms of ecotypicdi�erentiation. Meanwhile, the identi®ed ecotypic di�er-entiation in the timing of phenophases and other agro-nomically-relevant traits has allowed the selection andregistration of two spineless commercial varieties of burrmedic, including a precocious one (derived from accessionMPO-7-88) for the arid and semi-arid zones, and a late-¯owering one (derived from the accession MPO-43-88) for
the sub-humid zone.
en
ACKNOWLEDGEMENTS
The authors gratefully acknowledge FONDECYT(Project No. 1961142), Comite ECOS-Sur-CONICYT(Action No. C95B05) and a CNRS-CONICYT Agreement(No. 7097) for ®nancial support. We also thank Iva nPedreros for assistance with the glasshouse experiment, andTeresa Aravena, Jose Cares and Marõ a Elena Dõ az fortechnical help in the outdoor experiment. Dr R. Ellis andDr B. Dear provided valuable comments on the manu-script. Some of the data presented in Table 4 werepreviously presented in the Proceedings of the Meeting ofthe Mediterranean Working Group of the FAO/CIHEAMInter-Regional Research and Development Network onPastures and Fodder Crops, Avignon (France),
814 Del Pozo et al.ÐPhenological Developm
29 May±2 June, 1995.
LITERATURE CITED
AvendanÄ o J, Ovalle J. 1992. Sistemas de regeneracio n de praderasanuales en espinales de la zona Mediterra nea subhu meda. I.Produccio n de pasto y cubrimiento de la vegetecio n. AgriculturaTeÂcnica (Chile) 52: 32±37.
Aronson J, Kigel J, Shmida A, Klein J. 1992. Adaptive phenology ofdesert and Mediterranean populations of annual plants grownwith and without water stress. Oecologia 89: 17±26.
Bullita S, Floris R, Hayward MD, Loi A, Poqueddu C, Veronesi F. 1994.Morphological variation in Sardinian populations of Medicagopolymorpha L. suitable for rainfed Mediterranean conditions.Euphytica 77: 262±268.
Clarkson NM, Russell JS. 1975. Flowering responses to vernalizationand photoperiod in annual medics (Medicago spp.). AustralianJournal of Agricultural Research 26: 831±838.
Clarkson NM, Russell JS. 1979. E�ect of temperature on thedevelopment of two annual medics. Australian Journal of Agricul-tural Research 30: 909±916.
Del Pozo A, AvendanÄ o J, Ovalle C. 1999. Long term productivity of aley farming system in the `secano interior' of central Chile. CahiersOptions MeÂditerraneÂennes 39: 235±238.
Del Pozo A, GarcõÂ a-Huidobro J, Novoa R, Villaseca S. 1987. Relation-ship of base temperature to development of spring wheat.Experimental Agriculture 23: 21±30.
Del Pozo A, Ovalle C, AvendanÄ o J, Del Canto P. 1989. Adaptation ofMedicago polymorpha to the subhumid Mediterranean zone ofChile. Proceedings of the XVI International Grassland Congress,Nice, France, 1139±1540.
di Castri F, Hajek E. 1976. BioclimatologõÂa de Chile. Santiago: Vice-rectoria Acade mica de la Universidad Cato lica de Chile.
Ehrman T, Cocks PS. 1990. Ecogeography of annual legumes in Syria:distribution patterns. Journal of Applied Ecology 27: 578±591.
Ehrman T, Cocks PS. 1996. Reproductive patterns in annual legumespecies on an aridity gradient. Vegetatio 122: 47±59.
Ellis RH, Lawn RJ, Summer®eld RJ, Qi A, Roberts EH, Chay PM,Brouwer JB, Rose JL, Yeates SJ, Sandover S. 1994. Toward
the reliable production of time to ¯owering in six annual crops.V. Chickpea (Cicer arietinum). Experimental Agriculture 30:271±282.
Evans PM, Lawn RJ, Watkinson AR. 1992. Use of linear models topredict the date of ¯owering in cultivars of subterranean clover.Australian Journal of Agricultural Research 43: 1547±1558.
Evenari M, Gutterman Y. 1985.Desert plants. In: Halevy AH, ed. CRChandbook of ¯owering plants, Vol. 1. Boca Raton, Florida: CRCPress, 41±59.
Ewing MA. 1999. Annual pastures legumes. A vital componentstabilising and rehabilitating low rainfall Mediterranean ecosys-tems. Arid Soil Research and Rehabilitation 13: 327±342.
Ewing MA, Howieson JG. 1989. The development of Medicago poly-morpha L. as an important pasture species for southern Australia.Proceedings of the XVI International Grasslands Congress, Nice,France, 197±198.
Hadley P, Roberts EH, Summer®eld RJ, Minchin FR. 1983. Aquantitative model of reproductive development in cowpea(Vigna unguiculata (L.) Walp.) in relation to photoperiod andtemperature and implications for screening germoplasm. Annals ofBotany 51: 531±543.
Junttila O, Svenning MM, Solheim B. 1990a. E�ects of temperatureand photoperiod on vegetative growth of white clover (Trifoliumrepens) ecotypes. Physiologia Plantarum 79: 427±434.
Junttila O, Svenning MM, Solheim B. 1990b. E�ects of temperatureand photoperiod on frost resistance of white clover (Trifoliumrepens) ecotypes. Physiologia Plantarum 79: 435±438.
Loi A, Howieson JG, Cocks PS, Caredda S. 1993. The adaptation ofMedicago polymorpha to a range of edaphic and environmentalconditions: e�ect of temperature on growth, and acidity stress onnodulation and nod gene induction. Australian Journal of Agricul-tural Research 33: 25±30.
Loi A, Porqueddu C, Veronesi F, Cocks PS. 1995. Distribution,diversity and potential agronomic value of Medicago polymorphain Sardinia. Journal of Agricultural Science 124: 419±426.
Loss SP, Perry MW, Anderson WK. 1990. Flowering times of wheats insouth-western Australia: a modelling approach. Australian Journalof Agricultural Research 41: 213±224.
Ovalle C, Aronson J, Del Pozo A, AvendanÄ o J. 1990. The espinal:agroforestry systems of the Mediterranean-type climate region ofChile. Agroforestry Systems 10: 213±239.
Ovalle C, AvendanÄ o J, Del Pozo A, Crespo D. 1993. Germoplasmcollection, evaluation and selection of naturalized Medicago poly-morpha in the Mediterranean zone of Chile. XVII InternationalGrassland Congress, New Zealand, 222±223.
Ovalle C, Del Pozo A, AvendanÄ o J, Aronson J. 1997. Caracterõ sticasfenolo gicas y productivas de 34 accesiones de hualputra (Medi-cago polymorpha) colectadas en la zona mediterra nea de Chile.Agricultura TeÂcnica (Chile) 57: 261±271.
Piano E, Pecetti L, Carroni A. 1996. Climatic adaptation in subter-ranean clover populations. Euphytica 92: 39±44.
Statgraphics Plus. 1994. User manual. Rockville, Maryland, USA:Manugistics Inc.
Summer®eld RJ, Roberts EH, Ellis RH, Lawn RJ. 1991. Toward thereliable prediction of time to ¯owering in six annual crops. I. Thedevelopment of simple models for ¯uctuating ®eld environments.Experimental Agriculture 27: 11±31.
Summer®eld RJ, Roberts EH, Erskine W, Ellis RH. 1985. E�ects oftemperature and photoperiod on ¯owering in lentils (Lens
t in Ecotypes of Medicago polymorpha
culinaris). Annals of Botany 56: 659±671.












![Genotoxicity assessment and detoxification induction in Dreissena polymorpha exposed to benzo[a]pyrene](https://static.fdokumen.com/doc/165x107/6344d92703a48733920b14f7/genotoxicity-assessment-and-detoxification-induction-in-dreissena-polymorpha-exposed.jpg)







