
Developmental prosopagnosia: A study of three patients
19
Developmental prosopagnosia: A study of three patients Jason J.S. Barton, a,b,c, * Mariya V. Cherkasova, a Daniel Z. Press, a James M. Intriligator, a and Margaret OÕConnor a a Department of Neurology, Beth Israel Deaconess Medical Center and Harvard Medical School, Boston, MA, USA b Department of Ophthalmology, Beth Israel Deaconess Medical Center and Harvard Medical School, Boston, MA, USA c Department of Biomedical Engineering, Boston University, Boston, MA, USA Accepted 1 July 2002 Abstract We studied perception in three patients with prosopagnosia of childhood onset. All had trouble with other Ôwithin-categoryÕ judgments. All were deficient on face matching tests and severely impaired on tests of perception of the spatial relations of facial features and abstract designs, indicating a deficit in the encoding of coordinate relationships, similar to adult-onset prosopagnosia with lesions of the fusiform face area. Two had difficulty perceiving feature colour, which correlated with reduced luminance sensitivity. In contrast to adult-onset pa- tients, saturation discrimination was spared in two and spatial resolution impaired in two. Curvature discrimination was relatively spared. Contrast sensitivity showed variable reduc- tions at different spatial frequencies. We conclude that developmental prosopagnosia is similar to the adult-onset form in encoding deficits for the spatial arrangement of facial el- ements. Deficits in luminance perception and spatial resolution are more associated with defective encoding for basic object-level recognition, as shown on tests of object and spatial perception. Ó 2003 Elsevier Science (USA). All rights reserved. Keywords: Perception; Prosopagnosia; Developmental 1. Introduction Prosopagnosia is the inability to recognize familiar faces in the absence of more generalized cognitive dysfunction. While some degree of visual (de Haan, Young, & Newcombe, 1987) or memory disturbance (Bauer & Verfaellie, 1988; Bruyer et al., 1983; de Haan et al., 1987; Malone, Morris, Kay, & Levin, 1982; Rizzo, Hurtig, & Damasio, 1987) for stimuli other than faces can be present, the recognition deficit is more severe and sometimes highly specific for faces. Most cases of this unusual problem are acquired through lesions of the right (de Renzi, 1986; Landis, Brain and Cognition 51 (2003) 12–30 www.elsevier.com/locate/b&c * Corresponding author. Fax: 1-617-975-5322. E-mail address: [email protected] (J.J.S. Barton). 0278-2626/03/$ - see front matter Ó 2003 Elsevier Science (USA). All rights reserved. doi:10.1016/S0278-2626(02)00516-X
Transcript of Developmental prosopagnosia: A study of three patients

Developmental prosopagnosia: A study ofthree patients
Jason J.S. Barton,a,b,c,* Mariya V. Cherkasova,a Daniel Z. Press,a
James M. Intriligator,a and Margaret O�Connora
a Department of Neurology, Beth Israel Deaconess Medical Center and Harvard Medical School,
Boston, MA, USAb Department of Ophthalmology, Beth Israel Deaconess Medical Center and Harvard Medical School,
Boston, MA, USAc Department of Biomedical Engineering, Boston University, Boston, MA, USA
Accepted 1 July 2002
Abstract
We studied perception in three patients with prosopagnosia of childhood onset. All had
trouble with other �within-category� judgments. All were deficient on face matching tests andseverely impaired on tests of perception of the spatial relations of facial features and abstract
designs, indicating a deficit in the encoding of coordinate relationships, similar to adult-onset
prosopagnosia with lesions of the fusiform face area. Two had difficulty perceiving feature
colour, which correlated with reduced luminance sensitivity. In contrast to adult-onset pa-
tients, saturation discrimination was spared in two and spatial resolution impaired in two.
Curvature discrimination was relatively spared. Contrast sensitivity showed variable reduc-
tions at different spatial frequencies. We conclude that developmental prosopagnosia is
similar to the adult-onset form in encoding deficits for the spatial arrangement of facial el-
ements. Deficits in luminance perception and spatial resolution are more associated with
defective encoding for basic object-level recognition, as shown on tests of object and spatial
perception.
� 2003 Elsevier Science (USA). All rights reserved.
Keywords: Perception; Prosopagnosia; Developmental
1. Introduction
Prosopagnosia is the inability to recognize familiar faces in the absence of more
generalized cognitive dysfunction. While some degree of visual (de Haan, Young, &Newcombe, 1987) or memory disturbance (Bauer & Verfaellie, 1988; Bruyer et al.,
1983; de Haan et al., 1987; Malone, Morris, Kay, & Levin, 1982; Rizzo, Hurtig, &
Damasio, 1987) for stimuli other than faces can be present, the recognition deficit is
more severe and sometimes highly specific for faces. Most cases of this unusual
problem are acquired through lesions of the right (de Renzi, 1986; Landis,
Brain and Cognition 51 (2003) 12–30
www.elsevier.com/locate/b&c
*Corresponding author. Fax: 1-617-975-5322.
E-mail address: [email protected] (J.J.S. Barton).
0278-2626/03/$ - see front matter � 2003 Elsevier Science (USA). All rights reserved.
doi:10.1016/S0278-2626(02)00516-X

Cummings, Christen, Bogen, & Imbof, 1986; Takahashi, Kawamura, Hirayama,
Shiota, & Isono, 1995) or both(Damasio, Damasio, & van Hoessen, 1982; Meadows,
1974) occipitotemporal cortices, involving regions ranging from the anterior tem-
poral lobe to the fusiform and lingual gyri. Just as normal face recognition is thought
to involve a series of processing stages, acquired prosopagnosia is considered now to
have several functional subtypes (Damasio, 1985; Takahashi et al., 1995). The
broadest and most frequently used division is into two categories (Damasio, Tranel,
& Damasio, 1990; de Renzi, Faglioni, Grossi, & Nichelli, 1991): apperceptiveprosopagnosia, in which impaired formation of the facial percept prevents recog-
nition, and associative prosopagnosia, in which an adequately formed facial percept
cannot be compared with stores of facial memories.
Though rare, there are numerous well-studied cases of acquired adult-onset
prosopagnosia in the literature. Developmental or childhood-onset prosopagnosia,
however, has not been reported as often. The first case of this developmental form,
AB, was described in 1976 (McConachie, 1976) and in later reports (Campbell, 1992;
de Haan & Campbell, 1991). Since then, at least seven additional patients have beenreported in single case studies. These cases are summarized in Table 1.
Although the numbers are few, comparisons with adult-onset prosopagnosia re-
veal some interesting points. The majority of the developmental cases are thought to
have deficits at the level of structural encoding of the facial percept. Supportive
evidence for this stems mainly from impaired performance on the matching of un-
familiar faces (the Benton Face Recognition Test, or BFRT), poor judgments of
facial affect, gender or age (Ariel & Sadeh, 1996; de Haan & Campbell, 1991; Kracke,
1994; Young & Ellis, 1989), and deficient recognition of objects under more de-manding conditions, as in line drawings, unusual views, and overlapping or in-
complete figures (Ariel & Sadeh, 1996; de Haan & Campbell, 1991; Young & Ellis,
1989). Also, within-category judgments for non-face objects were impaired in at least
three patients (de Haan & Campbell, 1991; Temple, 1992; Young & Ellis, 1989),
suggesting that the defect was not as face-specific as reported for some adult-onset
prosopagnosic patients (Farah, Levinson, & Klein, 1995; McNeil & Warrington,
1993).
Despite these parallels with adult-onset prosopagnosia, imaging in the few de-velopmental cases in which this was obtained has not shown lesions of occipito-
temporal cortex similar to those in the adult-onset form. Also, other deficits
commonly associated with occipitotemporal lesions are unusual in childhood pros-
opagnosia. Only one patient had dyschromatopsia and superior altitudinal field
defects (Young & Ellis, 1989), and only two had topographagnosia (de Haan &
Campbell, 1991; Young & Ellis, 1989) while two others did not (Kracke, 1994;
Temple, 1992). Residual covert or unconscious knowledge of familiar faces has not
been found in any developmental prosopagnosic patients tested (Bentin et al., 1999;de Haan & Campbell, 1991; Young & Ellis, 1989), though it has been shown in
several adult-onset cases (Bruyer, 1991; Young, 1994).
The paucity of other functional markers or imaging evidence of occipitotemporal
damage raise the question of whether the type of perceptual encoding deficits in
developmental prosopagnosia are similar to those in adult-onset prosopagnosia. It is
not always the case that developmental disorders share the same pathophysiology as
their adult counterparts. For example, childhood-onset dyslexia and the acquired
pure alexia of left occipital lesions are considered different disorders. In adult-onsetprosopagnosia, the type of perceptual encoding dysfunction continues to be debated.
We have recently found that patients with lesions of the fusiform face area are se-
verely impaired in their ability to discern the spatial relations of facial features
(Barton, Press, Keenan, & O�Connor, 2002). These spatial relations are important indistinguishing the subtle variations in the basic configuration of features that all
faces share. An orientation-specific expertise with perceiving these spatial relations
J.J.S. Barton et al. / Brain and Cognition 51 (2003) 12–30 13
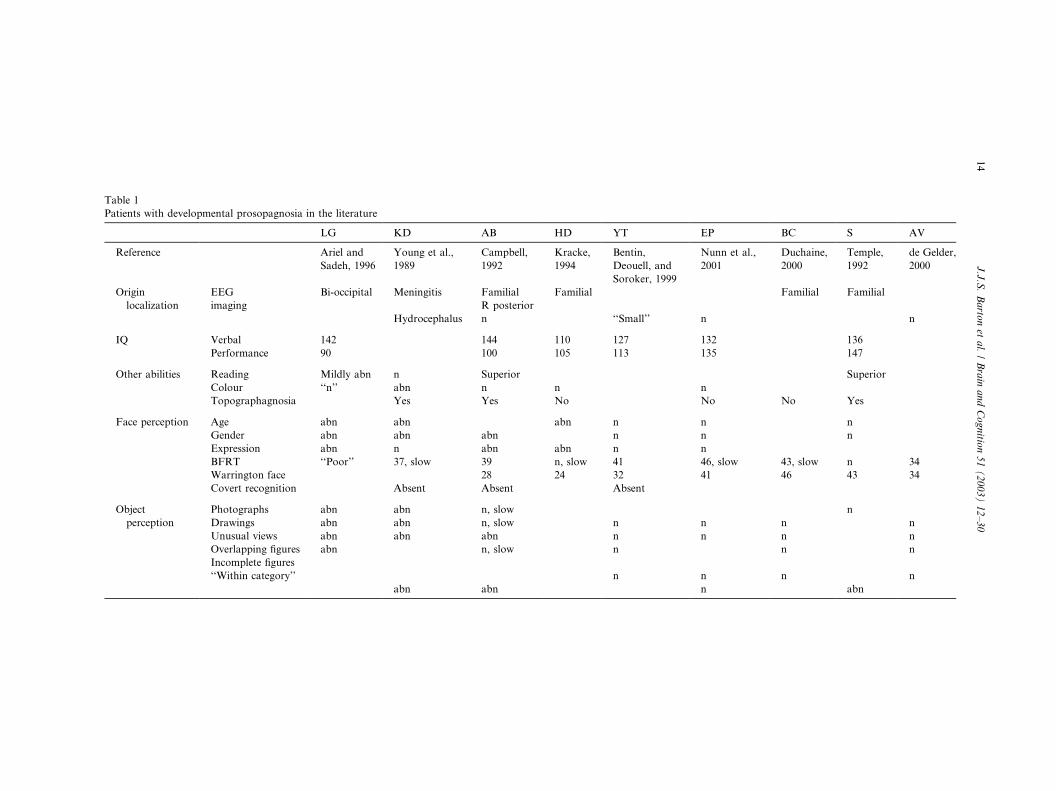
Table 1
Patients with developmental prosopagnosia in the literature
LG KD AB HD YT EP BC S AV
Reference Ariel and
Sadeh, 1996
Young et al.,
1989
Campbell,
1992
Kracke,
1994
Bentin,
Deouell, and
Soroker, 1999
Nunn et al.,
2001
Duchaine,
2000
Temple,
1992
de Gelder,
2000
Origin
localization
EEG
imaging
Bi-occipital Meningitis Familial
R posterior
Familial Familial Familial
Hydrocephalus n ‘‘Small’’ n n
IQ Verbal 142 144 110 127 132 136
Performance 90 100 105 113 135 147
Other abilities Reading Mildly abn n Superior Superior
Colour ‘‘n’’ abn n n n
Topographagnosia Yes Yes No No No Yes
Face perception Age abn abn abn n n n
Gender abn abn abn n n n
Expression abn n abn abn n n
BFRT ‘‘Poor’’ 37, slow 39 n, slow 41 46, slow 43, slow n 34
Warrington face 28 24 32 41 46 43 34
Covert recognition Absent Absent Absent
Object
perception
Photographs
Drawings
Unusual views
Overlapping figures
Incomplete figures
‘‘Within category’’
abn
abn
abn
abn
abn
abn
abn
abn
n, slow
n, slow
abn
n, slow
abn
n
n
n
n
n
n
n
n
n
n
n
n
n
abn
n
n
n
n
14
J.J.S.Bartonetal./BrainandCognitio
n51(2003)12–30

likely accounts for the difficulty normal subjects have in recognizing upside-down
faces—the �inversion effect� (Barton, Keenan, & Bass, 2001b; Cooper & Wojan, 2000;Freire, Lee, & Symons, 2000; Leder & Bruce, 2000).
Apart from the indirect evidence noted above, the nature of perceptual encoding
has rarely been studied in detail in developmental prosopagnosia. One patient, GA,
was found to have a deficit in curvature perception (Kosslyn, Hamilton, &
Bernstein, 1995). It was speculated that this reflected selective anoxic damage to
end-stopped cells in primary visual cortex. Our study of seven adult-onset pros-opagnosic patients (Barton, Cherkasova, Press, Intriligator, & O�Connor, 2003)found normal curvature perception in all but one, suggesting that curvature mis-
perception seldom contributes to defective perceptual encoding in adult cases with
defined extra-striate damage.
In this report we describe studies on GA and an additional two patients with
developmental prosopagnosia. The evidence suggests that all three have problems
with perceptual encoding of faces. We conducted a detailed review of many per-
ceptual functions, using the same tests employed in our sample of seven adult-onsetprosopagnosic patients with localizable lesions on imaging. These tests probed
perception of not only spatial relations but also a range of other basic functions of
potential importance to encoding complex three-dimensional structures, such as
luminance, contrast, saturation, spatial resolution, and curvature. Our goal was to
determine whether the pattern of deficits in these developmental patients was similar
to that in adult-onset prosopagnosia, or if it indicated a very different type of ap-
perception.
2. Methods
All patients and normal subjects gave informed consent in a protocol approved by
the hospital�s institutional review board. Normal subjects serving as controls had
corrected Snellen near acuities of better than 20/40 for participation, normal visual
fields by confrontation, and handedness was assessed with the Edinburgh battery.
Patients had further detailed neurologic and neuro-ophthalmologic assessment, in-cluding Goldmann perimetry and magnetic resonance imaging.
Detailed neuropsychological testing of these patients included indices of verbal
and performance IQ using the WAIS-R (Wechsler, 1981), Benton face recognition
test—BFRT (Benton & van Allen, 1972), Warrington recognition test for words and
faces—WFMT (Warrington, 1984), Warrington visual object and space perception
battery (VOSP) (Warrington & James, 1991), Benton line orientation test, Ghent
overlapping figures, and the logical memory and visual reproduction tests of the
Wechlser Memory Scale.Prosopagnosia was confirmed by the Famous Faces test (Albert, Butters, & Levin,
1979), as well as by a test of overt familiarity that used a series of famous and un-
known faces, reported elsewhere (Barton, Cherkasova, & O�Connor, 2001a). Thisasked patients to indicate which of a series of 20 famous and 20 anonymous faces
were familiar. Because this method yields both false positives and false negatives,
discriminative power can be assessed independent of criterion bias with signal de-
tection methodology. Because KBN is from a foreign culture, we confirmed her
prosopagnosia using faces of family members and actors on her favorite televisionshows. She showed no ability to identify any individual.
2.1. Case reports
GA, a 21-year-old right handed man, was the subject of a previous re-
port (Kosslyn et al., 1995). He had suffered a fall at one year of age, with
J.J.S. Barton et al. / Brain and Cognition 51 (2003) 12–30 15

cardiopulmonary arrest and coma. He has had lifelong difficulty recognizing people
and finding his way even in familiar surroundings, but no problems with colour
perception. He began a computer programming major at college, but did not
complete it. GA has a socially isolated life. His main friends are the individuals
who participate in his internet support group for developmental prosopagnosia. He
has never had a girlfriend. He continues to live at home with his mother and is
employed in computer work for her company. His speech tends towards the pe-
dantic side, with an insistence upon strict definitions, though he has a lively ifquirky sense of humour. Visual acuity was 20/20 in each eye with correction, and
visual fields were normal. He had a slightly microphthalmic appearance. Neuro-
logic examination was normal. Neuropsychologic evaluation showed superior
verbal intelligence. There were no stereotyped movement patterns. MRI of his
brain was normal.
KBN is a 31-year-old woman who had frequent seizures since two years of age.
These were initially generalized tonic-clonic seizures, but most since early childhood
are spells of unresponsiveness without motor manifestations. She was the product ofa normal pregnancy and delivery and achieved early developmental milestones at a
normal rate. However, she has always had difficulty recognizing faces, using cues
such as voice or clothing for identity instead. She also has trouble recognizing her
brother�s car in parking lots. She gets lost in familiar surroundings and her reading isslow, but she denied problems with colour perception. Socially she became pro-
gressively withdrawn and tended to engage in solitary activities, despite growing up
in a large family. She achieved the equivalent of grade 12 but has not worked or
studied at school since. She has no friends outside her family and has never had aromantic relationship.
Her visual acuity was 20/50. She saw 11 of 14 Ishihara pseudoisochromatic plates,
in the normal range. Confrontation visual fields were mildly constricted. She had left
superior oblique overaction but otherwise normal eye movements. Neurologic ex-
amination was normal. Neuropsychologic testing showed average verbal intelligence
and superior attention and working memory. Memory for verbal material was
normal, but that for non-verbal material was severely impaired. Object recognition
was impaired, particularly for unusual views and line drawings. Reading speed andcomprehension were slow. Note was made of her poor eye contact, monotonous
speech, lack of facial expression, and difficulty recognizing non-verbal aspects of
conversation. Interictal EEG showed generalized bursts of slowing. MRI showed
abnormal gyral patterns of both occipital lobes, interpreted as a mild focal devel-
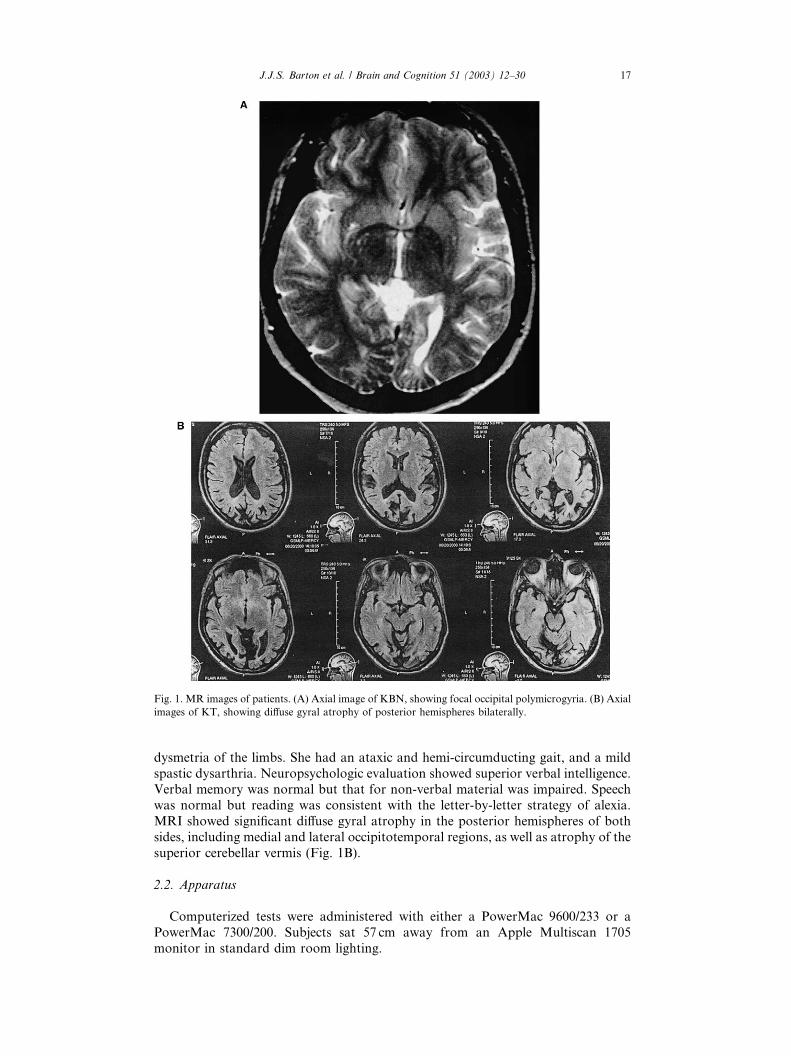
opmental polymicrogyria (Fig. 1A).
KT, a 36-year-old right handed woman, had a respiratory arrest at age 6 and was
in a vegetative state for 2months. On recovery she was nearly blind, had impaired
balance and limb coordination, and developed an exostrabismus. Her vision grad-ually improved to a visual acuity of about 20/50. Despite this, she continued to have
persistent problems with recognizing faces and finding her way around familiar
routes. She could not describe the route to a school she had been attending for four
years, for example. She denied problems with colour vision. She has difficulty
reading but graduated from college with an English major and learned seven lan-
guages, relying on audiotapes and volunteer readers. Her social circle is limited to
immediate family and academic mentors. She continues to live with her mother and
has not had a romantic relationship.Her visual acuity was 20/50 in the right eye and 20/200 in the amblyopic left eye.
She was impaired on Ishihara pseudoisochromatic plates, identifying only 4 of 14
plates. Goldmann perimetry shows a homonymous partial left inferior quadran-
tanopia. She had a large-angle exotropia, frequent square wave jerks, and impaired
smooth pursuit and cancellation of the vestibulo-ocular reflex. Neurologic exam
showed diffuse spasticity, mild left hemiparesis, startle myoclonus, and cerebellar
16 J.J.S. Barton et al. / Brain and Cognition 51 (2003) 12–30
dysmetria of the limbs. She had an ataxic and hemi-circumducting gait, and a mild
spastic dysarthria. Neuropsychologic evaluation showed superior verbal intelligence.Verbal memory was normal but that for non-verbal material was impaired. Speech
was normal but reading was consistent with the letter-by-letter strategy of alexia.
MRI showed significant diffuse gyral atrophy in the posterior hemispheres of both
sides, including medial and lateral occipitotemporal regions, as well as atrophy of the
superior cerebellar vermis (Fig. 1B).
2.2. Apparatus
Computerized tests were administered with either a PowerMac 9600/233 or a
PowerMac 7300/200. Subjects sat 57 cm away from an Apple Multiscan 1705
monitor in standard dim room lighting.
Fig. 1. MR images of patients. (A) Axial image of KBN, showing focal occipital polymicrogyria. (B) Axial
images of KT, showing diffuse gyral atrophy of posterior hemispheres bilaterally.
J.J.S. Barton et al. / Brain and Cognition 51 (2003) 12–30 17
2.3. Stimuli and tests
2.3.1. Characterization of prosopagnosia
In addition to assessment with the BFRT and WFMT, we administered tests to
determine the status of facial configuration perception and the face-specificity of the
disorder, by testing for �within category� identifications within another object do-main. Tests of covert face perception in these three subjects have been reported
elsewhere (Barton et al., 2001a, 2001b): none of these patients showed evidence ofcovert familiarity or covert semantic facial information.
1. Discrimination of facial features and spatial configuration. This tested whether
subjects were impaired in their perception of changes in facial structure or feature
colour. We created face stimuli that differed in a step-wise quantitative fashion along
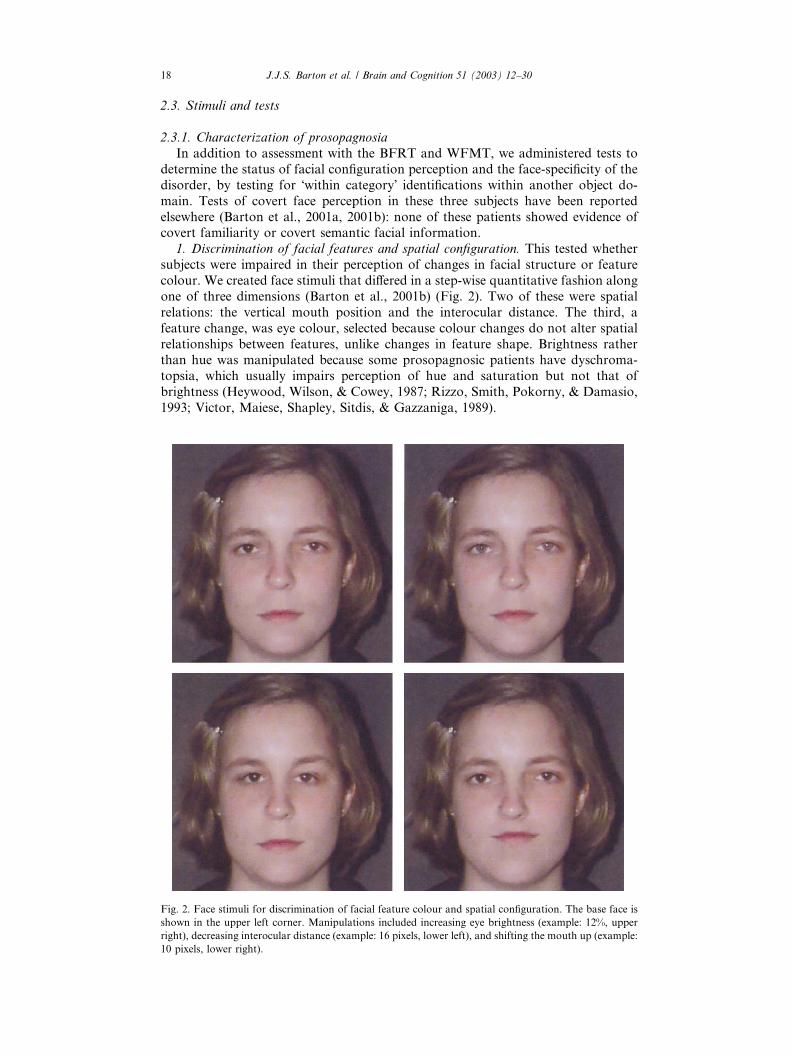
one of three dimensions (Barton et al., 2001b) (Fig. 2). Two of these were spatial
relations: the vertical mouth position and the interocular distance. The third, a
feature change, was eye colour, selected because colour changes do not alter spatial
relationships between features, unlike changes in feature shape. Brightness ratherthan hue was manipulated because some prosopagnosic patients have dyschroma-
topsia, which usually impairs perception of hue and saturation but not that of
brightness (Heywood, Wilson, & Cowey, 1987; Rizzo, Smith, Pokorny, & Damasio,
1993; Victor, Maiese, Shapley, Sitdis, & Gazzaniga, 1989).
Fig. 2. Face stimuli for discrimination of facial feature colour and spatial configuration. The base face is
shown in the upper left corner. Manipulations included increasing eye brightness (example: 12%, upper
right), decreasing interocular distance (example: 16 pixels, lower left), and shifting the mouth up (example:
10 pixels, lower right).
18 J.J.S. Barton et al. / Brain and Cognition 51 (2003) 12–30

We used full-colour digitized frontal images of the faces of one male and one
female, not known to the subjects. Each face spanned 8.8�� 8.8� of visual angle.Spatial manipulations of the images were done with Paintshop Pro 3.0 (Shareware).
Target faces with altered eye displacement had both eyes moved nasally by an
amount ranging from 4 to 33min of angle. Those with altered mouth position had
the mouth moved up by 4–21min of angle. Those with altered eye colour had eye
brightness increased by 3–15%, using Adobe Photoshop 5.0 (Adobe, San Jose).
A trial stimulus consisted of three faces presented simultaneously in a triangulararrangement. Two were the base (unaltered) face and one a target face. The subject�stask was to indicate which face was the different one.
We constructed test blocks with the Superlab 1.71 (Cedrus, Phoenix, AZ) pro-
gram. Each trial stimulus was presented nine times for each of the two (male and
female) faces. Thus each data point represents the mean of 18 trials. Normal subjects
were tested at five gradations of change in each dimension, but some of the patients
were tested only at the easiest levels. Blocks could be given with different viewing
durations of 2, 4 s, or unlimited time. Reaction time was also recorded for the latter.The standard test contained equal numbers of trials with the three different types of
stimulus change.
An additional block tested subjects with trials that contained only changes in
mouth position. Subjects were informed that this was the case, so that the effects of
limiting attention to this single spatial change could be determined.
Control subjects were 10 men and 4 women ranging in age from 16 to 43 years: 12
were right handed. The control data was used to generate 95% prediction intervals
with which each patient�s data could be compared.2. Within-category recognition of fruits and vegetables. The aim of this test was to
determine whether the recognition deficit extended to another within-category
perceptual domain. Perceptual accuracy can reflect expertise gained from prior
experience (Diamond & Carey, 1986; Gauthier, Skudlarski, & Gore, 2000). One
difficulty in ascertaining the �face-specificity� of impaired recognition lies in ensuringthat a given subject has equal expertise with other object categories tested (Farah
et al., 1995). We reasoned that almost all people have experience with and interest
in food.We used segments of colour images of fruits and vegetables, printed on paper.
Many segments did not conform to the silhouette of the item portrayed, requiring
the subject to analyze the internal contours, texture and colour to identify the item.
This test included 14 red items and 10 green items to be identified by name and 12
items to be matched to a panel of 6 exemplars, which were fruits or vegetables of a
different variety or seen from a different viewpoint. There was no time limit. Accu-
racy but not reaction time was recorded. Control subjects were 4 men and 4 women
of mean age 30 years (range 19–43 years).
2.3.2. Characterization of basic perceptual functions
1. Contrast sensitivity. Subjects viewed vertical sinusoidal gratings within a two-
dimensional Gaussian envelope spanning 7� in diameter (Morphonome 2.7.9, ImagePsychophysics, San Francisco). The base contrast was zero. A method-of-adjustment
protocol was used. Approximate threshold was determined, and contrast was first set
below this level at a randomly determined point. Subjects then increased contrast
with a keypress until the display was just visible. Contrast was then reset at a randompoint above this level, and subjects decreased contrast with the keypress until the
display just disappeared. The procedure was repeated three times, to yield six values.
The inverse of these contrast threshold values were averaged to give the mean
contrast sensitivity.
We tested four different spatial frequencies: 0.1, 0.5, 4.1, and 8.2 cycles/�. Thestimulus was always viewed centrally.
J.J.S. Barton et al. / Brain and Cognition 51 (2003) 12–30 19

Control subjects for this test were 6 women and 3 men, of mean age 34 years
(range 22–44 years). The same controls were also used for the following tests of lu-
minance, saturation and spatial resolution.
2. Luminance discrimination. Luminance, saturation, and spatial resolution tests
were created by author JI using the Vision Shell programming platform
(www.kagi.com/visionshell). A square of 8� width was divided into upper and
lower halves, and seen against a black background. One half always had a lumi-
nance value of 20 cd/M2, and the luminance of the other half varied in the rangegreater than 20 cd/M2. To improve the sensitivity of the test, luminant noise was
added to the entire display. Which half was brighter varied randomly between
trials. The square was centered at fixation or 8� left or right of fixation, with lo-cation on a trial also randomly determined. The display lasted 75ms. Subjects
fixated on a central point and were asked to indicate whether the top or the
bottom half was the brighter, by a keypress. Normal subjects were tested on a
standard set of brightness values for the target half ranging from 21 to 35 cd/M2.
The values used to test patients were modified when necessary, extending to highervalues for patients impaired on the standard set. Twelve trials were used for each
luminance difference measured at each position. For saturation, luminance, and
spatial resolution, we estimated psychophysical thresholds corresponding to 75%
correct in a two-alternative forced-choice test, using least-squares linear regression
to z-scored data (Simpson, 1995). We then calculated the mean and 95% prediction
limits for thresholds in normal subjects, and compared patient thresholds to these
intervals.
3. Saturation discrimination. This test resembled the test of luminance discrimi-nation. The same square was used, only this time it was a slight pink hue, whose
percent saturation was set at zero for the standard half, while saturation increased in
the target half. A variable amount of luminance was added to each half randomly
among trials, to eliminate the use of luminant cues in the test. Subjects were asked to
indicate which half was the redder. Eight normal subjects were tested on a standard
set of saturation values ranging from 1 to 10%. Again, the values used to test the
patients were modified when they performed poorly with the standard set. As above,
12 trials were used for each saturation difference measured at each position, andthresholds estimated at 75% correct performance.
4. Spatial resolution (Landolt C). A white ring of 2� diameter and luminance of60 cd/M2 was seen at center or 8� left or right of center. This appeared for only150ms. The ring contained a gap whose size varied from 1 to 10 pixels (0.037�/pixel)at either its top or bottom (12 or 6 o�clock). The subject was asked to indicate with akeypress whether they perceived the gap at the top or bottom position. 16 trials were
used for each gap size at each position.
5. Dot displacement discrimination. This tested perception of spatial relations innon-facial stimuli.
To mimick the relevant dimension of our face tests, we created a design of four
grey dots, which if overlayed on one of our face stimuli would have covered the eyes,
tip of nose, and center of mouth. The procedure also mimicked the presentation in
our face trials. Three sets of this design were presented on a trial, in the same po-
sition as our face stimuli. Two designs were identical. The third differed in one of the
following ways: the horizontal distance between the upper two dots was reduced by
0.2, 0.4, 0.6 or 0.8�, the vertical distance between the lower two dots was reduced by0.1, 0.2, 0.3, or 0.4�, or all four dots were increased in brightness by one or two levelsin the drawing program. The subjects task was to indicate which of the three designs
was different, with viewing duration set at 2 s. Twelve trials were given for each
stimulus change.
For this test and that of curvature discrimination, we had five control subjects, 4
women and 1 man, of mean age 36 (range 22–44 years).
20 J.J.S. Barton et al. / Brain and Cognition 51 (2003) 12–30
6. Curvature discrimination. This test was included because of a prior demon-
stration of impaired curvature detection in a prosopagnosic patient (Kosslyn et al.,
1995).
Two curved line segments were created by showing a portion of the left side of
two black rings, placed side by side, through a rectangular aperture, whose height
and placement varied slightly between trials, being 2.5� on average. One curvealways belonged to a circle with a diameter of 6.6�. The other curve belonged tocircles whose diameter varied from trial to trial, from 6.3� to 3.5�. Eight differentcircle diameters were used, with 18 trials for each. The subject�s task was to in-dicate whether the left or right line segment was more curved. Viewing duration
was unlimited, and both the percent correct scores and reaction times were
measured.
3. Results (Table 2)
3.1. Characterization of prosopagnosia
GA and KBN were in the low normal range (11th percentile) on the BFRT,
whereas KT was severely impaired (<98% limit). All scored below the 95% limit on
the Warrington Face recognition test. On tests of covert recognition reported else-
where (Barton et al., 2001a), none of these subjects showed any residual unconscious
access to facial identity.
3.1.1. Discrimination of facial features and spatial configuration
All three patients were significantly impaired in judgments of the spatial position
of features, in both time-limited and unlimited duration trials (Fig. 3). Neither KBN
nor KT were able to reach threshold (67% correct) with even the largest shifts of
feature position. GA was able to achieve a borderline normal score for mouth
position with unlimited duration, but at the cost of very prolonged reaction times
(Fig. 4).
Fig. 3. Accuracy for discrimination of facial feature colour and spatial configuration. Data for normal
controls are shown in the top graphs, patients in the bottom graphs. Left graphs are for eye brightness
changes, middle graphs are for eye position shifts, and right graphs for mouth position shifts. Viewing
durations are: 2s, 2 seconds; 4s, 4 seconds; unlim, unlimited time.
J.J.S. Barton et al. / Brain and Cognition 51 (2003) 12–30 21
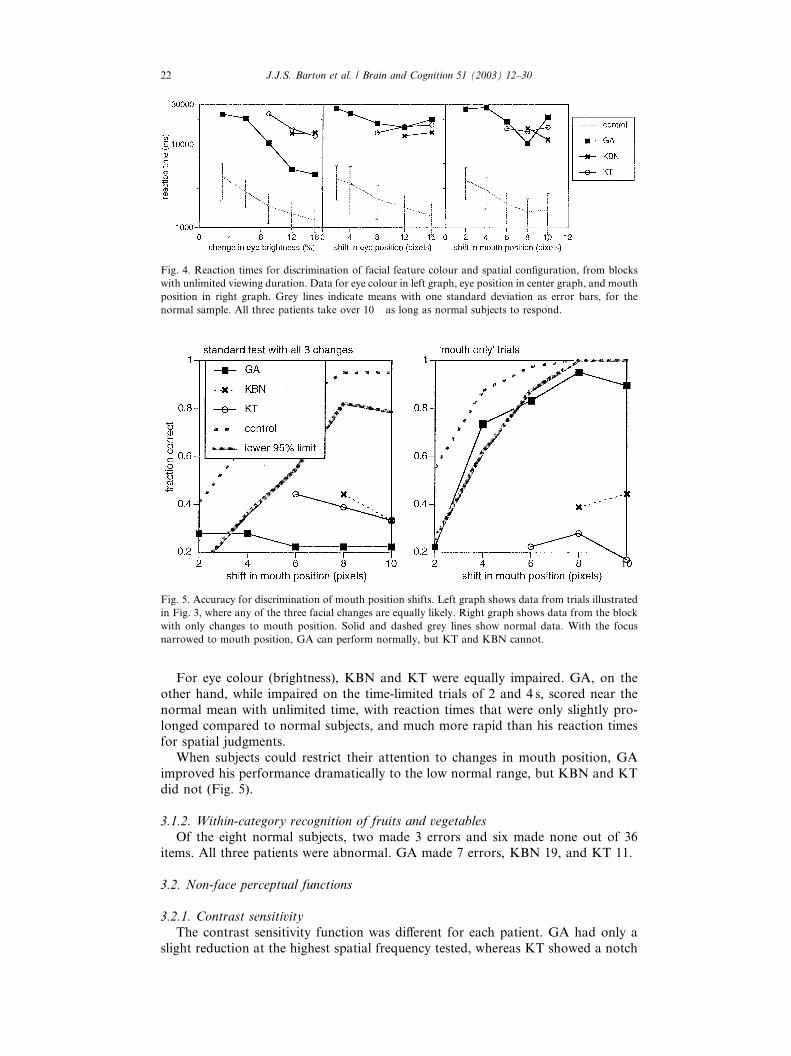
For eye colour (brightness), KBN and KT were equally impaired. GA, on the
other hand, while impaired on the time-limited trials of 2 and 4 s, scored near the
normal mean with unlimited time, with reaction times that were only slightly pro-
longed compared to normal subjects, and much more rapid than his reaction times
for spatial judgments.
When subjects could restrict their attention to changes in mouth position, GA
improved his performance dramatically to the low normal range, but KBN and KTdid not (Fig. 5).
3.1.2. Within-category recognition of fruits and vegetables
Of the eight normal subjects, two made 3 errors and six made none out of 36
items. All three patients were abnormal. GA made 7 errors, KBN 19, and KT 11.
3.2. Non-face perceptual functions
3.2.1. Contrast sensitivity
The contrast sensitivity function was different for each patient. GA had only a
slight reduction at the highest spatial frequency tested, whereas KT showed a notch
Fig. 5. Accuracy for discrimination of mouth position shifts. Left graph shows data from trials illustrated
in Fig. 3, where any of the three facial changes are equally likely. Right graph shows data from the block
with only changes to mouth position. Solid and dashed grey lines show normal data. With the focus
narrowed to mouth position, GA can perform normally, but KT and KBN cannot.
Fig. 4. Reaction times for discrimination of facial feature colour and spatial configuration, from blocks
with unlimited viewing duration. Data for eye colour in left graph, eye position in center graph, and mouth
position in right graph. Grey lines indicate means with one standard deviation as error bars, for the
normal sample. All three patients take over 10� as long as normal subjects to respond.
22 J.J.S. Barton et al. / Brain and Cognition 51 (2003) 12–30
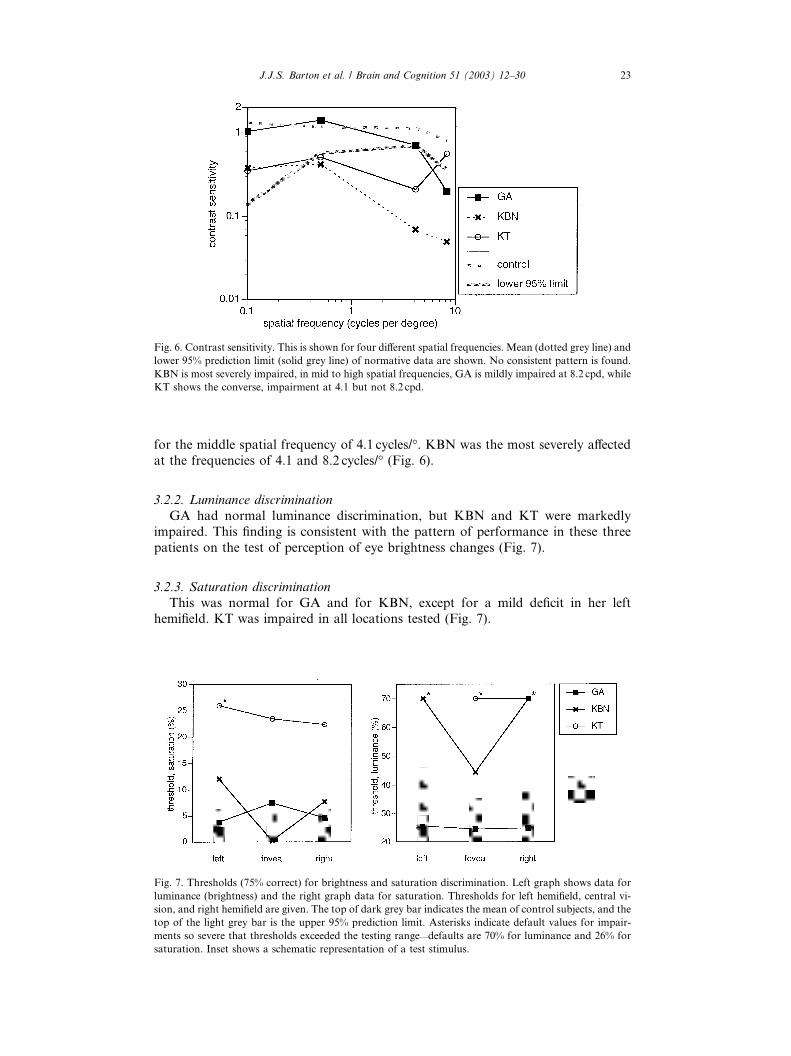
for the middle spatial frequency of 4.1 cycles/�. KBN was the most severely affected
at the frequencies of 4.1 and 8.2 cycles/� (Fig. 6).
3.2.2. Luminance discrimination
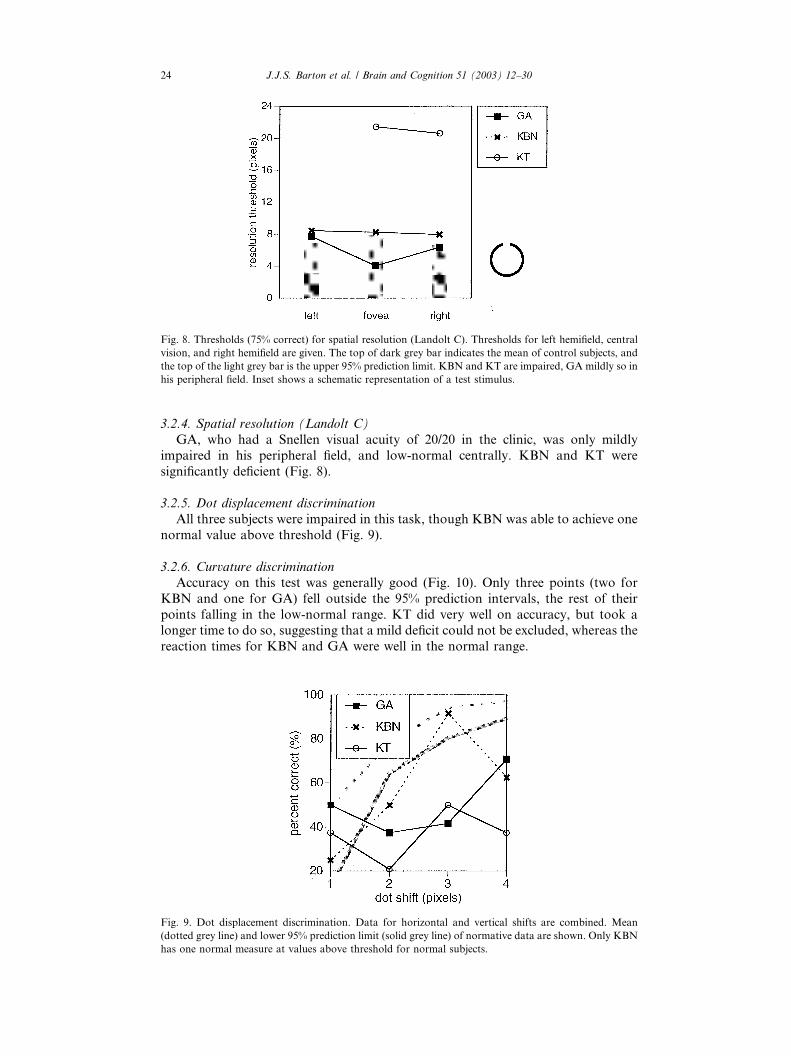
GA had normal luminance discrimination, but KBN and KT were markedly
impaired. This finding is consistent with the pattern of performance in these three
patients on the test of perception of eye brightness changes (Fig. 7).
3.2.3. Saturation discrimination
This was normal for GA and for KBN, except for a mild deficit in her lefthemifield. KT was impaired in all locations tested (Fig. 7).
Fig. 7. Thresholds (75% correct) for brightness and saturation discrimination. Left graph shows data for
luminance (brightness) and the right graph data for saturation. Thresholds for left hemifield, central vi-
sion, and right hemifield are given. The top of dark grey bar indicates the mean of control subjects, and the
top of the light grey bar is the upper 95% prediction limit. Asterisks indicate default values for impair-
ments so severe that thresholds exceeded the testing range—defaults are 70% for luminance and 26% for
saturation. Inset shows a schematic representation of a test stimulus.
Fig. 6. Contrast sensitivity. This is shown for four different spatial frequencies. Mean (dotted grey line) and
lower 95% prediction limit (solid grey line) of normative data are shown. No consistent pattern is found.
KBN is most severely impaired, in mid to high spatial frequencies, GA is mildly impaired at 8.2 cpd, while
KT shows the converse, impairment at 4.1 but not 8.2 cpd.
J.J.S. Barton et al. / Brain and Cognition 51 (2003) 12–30 23
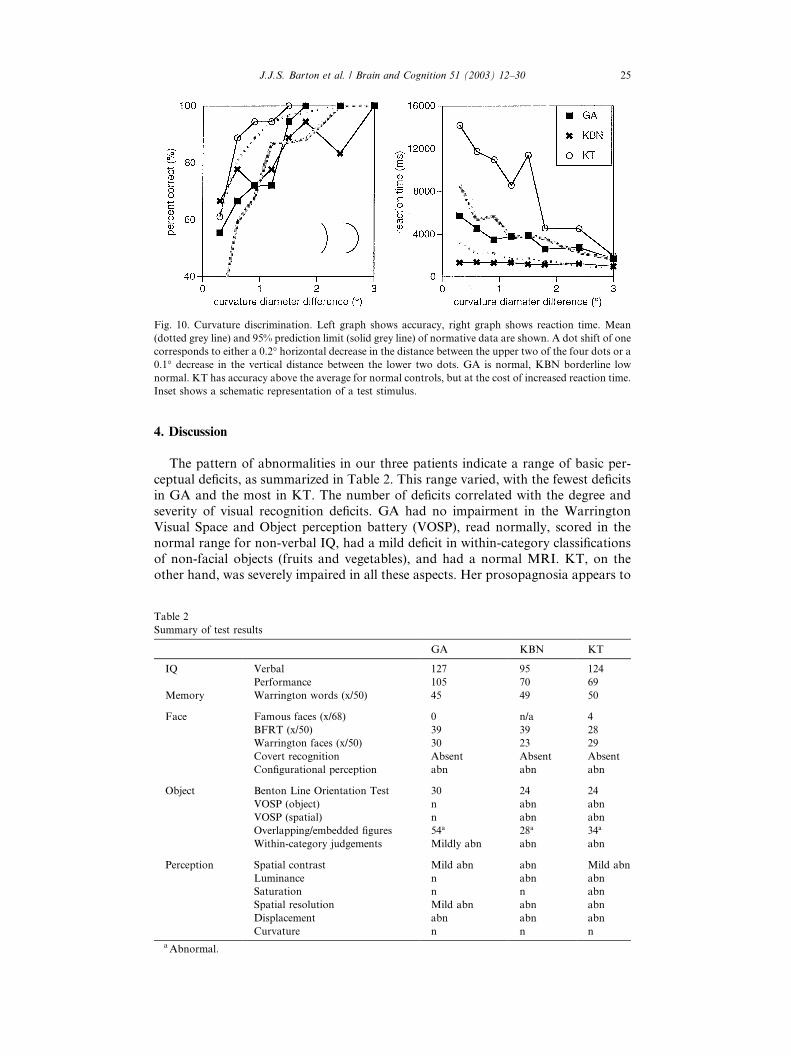
3.2.4. Spatial resolution (Landolt C)
GA, who had a Snellen visual acuity of 20/20 in the clinic, was only mildly
impaired in his peripheral field, and low-normal centrally. KBN and KT were
significantly deficient (Fig. 8).
3.2.5. Dot displacement discrimination
All three subjects were impaired in this task, though KBN was able to achieve one
normal value above threshold (Fig. 9).
3.2.6. Curvature discrimination
Accuracy on this test was generally good (Fig. 10). Only three points (two for
KBN and one for GA) fell outside the 95% prediction intervals, the rest of their
points falling in the low-normal range. KT did very well on accuracy, but took alonger time to do so, suggesting that a mild deficit could not be excluded, whereas the
reaction times for KBN and GA were well in the normal range.
Fig. 9. Dot displacement discrimination. Data for horizontal and vertical shifts are combined. Mean
(dotted grey line) and lower 95% prediction limit (solid grey line) of normative data are shown. Only KBN
has one normal measure at values above threshold for normal subjects.
Fig. 8. Thresholds (75% correct) for spatial resolution (Landolt C). Thresholds for left hemifield, central
vision, and right hemifield are given. The top of dark grey bar indicates the mean of control subjects, and
the top of the light grey bar is the upper 95% prediction limit. KBN and KT are impaired, GA mildly so in
his peripheral field. Inset shows a schematic representation of a test stimulus.
24 J.J.S. Barton et al. / Brain and Cognition 51 (2003) 12–30
4. Discussion
The pattern of abnormalities in our three patients indicate a range of basic per-
ceptual deficits, as summarized in Table 2. This range varied, with the fewest deficitsin GA and the most in KT. The number of deficits correlated with the degree and
severity of visual recognition deficits. GA had no impairment in the Warrington
Visual Space and Object perception battery (VOSP), read normally, scored in the
normal range for non-verbal IQ, had a mild deficit in within-category classifications
of non-facial objects (fruits and vegetables), and had a normal MRI. KT, on the
other hand, was severely impaired in all these aspects. Her prosopagnosia appears to
Table 2
Summary of test results
GA KBN KT
IQ Verbal 127 95 124
Performance 105 70 69
Memory Warrington words (x/50) 45 49 50
Face Famous faces (x/68) 0 n/a 4
BFRT (x/50) 39 39 28
Warrington faces (x/50) 30 23 29
Covert recognition Absent Absent Absent
Configurational perception abn abn abn
Object Benton Line Orientation Test 30 24 24
VOSP (object) n abn abn
VOSP (spatial) n abn abn
Overlapping/embedded figures 54a 28a 34a
Within-category judgements Mildly abn abn abn
Perception Spatial contrast Mild abn abn Mild abn
Luminance n abn abn
Saturation n n abn
Spatial resolution Mild abn abn abn
Displacement abn abn abn
Curvature n n n
aAbnormal.
Fig. 10. Curvature discrimination. Left graph shows accuracy, right graph shows reaction time. Mean
(dotted grey line) and 95% prediction limit (solid grey line) of normative data are shown. A dot shift of one
corresponds to either a 0.2� horizontal decrease in the distance between the upper two of the four dots or a0.1� decrease in the vertical distance between the lower two dots. GA is normal, KBN borderline low
normal. KT has accuracy above the average for normal controls, but at the cost of increased reaction time.
Inset shows a schematic representation of a test stimulus.
J.J.S. Barton et al. / Brain and Cognition 51 (2003) 12–30 25

be part of a triad of alexia, prosopagnosia and visual agnosia, a syndrome attributed
to bilateral occipital damage by others (Farah, 1990), and confirmed by her mark-
edly abnormal structural brain imaging.
Our prior studies of patients with adult-onset prosopagnosia revealed that lesions
involving the fusiform face area in the right occipitotemporal lobe were associated
with severe deficits in discriminating the spatial relationships of facial features
(Barton et al., 2002). These same deficits were present in all three patients with de-
velopmental prosopagnosia, even though no visible lesion of the fusiform gyrus waspresent, at least in KBN and GA. Evidence from functional imaging shows that the
fusiform face area is more specifically activated during face recognition than during
object recognition (Kanwisher, McDermott, & Chun, 1997) and other face pro-
cessing tasks like direction of gaze and expression (Haxby, Hoffman, & Gobbini,
2000). The fact that our developmental prosopagnosics share a perceptual deficit
similar to that caused by visible acquired lesions of the fusiform face area implies a
hypofunction of this region even in the absence of gross structural damage.
As with our adult-onset prosopagnosics (Barton et al., 2002), these developmentalpatients also varied in whether their deficit in perceiving spatial relationships in faces
could be reduced by focusing attention upon a single region (the mouth). GA per-
formed normally with a narrowing of focus, whereas KT and KBN did not benefit
from this. The enhanced performance with focusing has been interpreted as evidence
of processing failure under conditions of distributed attention, a higher level re-
duction in processing capacity. The two adult-onset patients who did not improve
with narrowing of focus had more peristriate damage, just as KBN and KT have
more imaging evidence of occipital damage than GA, and more extensive visualdysfunction on our battery. Thus their spatial relational perception may be impaired
at a lower hierarchical level than that of GA.
Tests of their non-facial perception yielded interesting similarities and differences
compared with the adult-onset patients (Barton et al., 2003). Like the adult onset
group, the problem with spatial relational judgements occurred even with geometric
dot arrangements: hence they are not part of a face-specific processing mechanism.
Luminance discrimination and contrast sensitivity were also variable in the group,
being least affected in GA. GA did have difficulty with discriminating eye luminancechanges in the face perception test when viewing duration was short. Thus he could
not apply his normal luminance mechanisms effectively in a situation requiring rapid
processing of multiple facial regions. This again implies a failure of processing ca-
pacity under conditions of distributed attention, extending not only to spatial rela-
tions but also to other feature information.
While all our adult-onset patients had normal spatial resolution, this was im-
paired in all our developmental prosopagnosic patients, again, least so in GA. The
contribution of reduction in such high spatial frequency information to face pro-cessing is debatable, particularly given the lack of deficit in the adult-onset group. It
does suggest more pervasive apperceptive deficits in our developmental patients,
though, as shown in the defects on VOSP (object) and embedded figures tests of
KBN and KT (Table 2).
One interesting finding was the good accuracy for curvature discrimination by all
three patients. Impaired curvature perception had been reported in GA previously
(Kosslyn et al., 1995). However, the test used in that study that was most similar to
our curvature test showed only prolonged reaction times, not impaired accuracy, byGA. This pattern is reproduced by KT to a degree, but not by GA at present. It may
be that his prior extensive testing served to practice his performance. In the prior
report (Kosslyn et al., 1995) GA was also impaired in accuracy on more complex
tests involving curved stimuli—but notably, many of these required him to make
some spatial relational judgement about a target superimposed upon curved designs.
It may be that the severe defect in spatial judgments we uncovered played a role in
26 J.J.S. Barton et al. / Brain and Cognition 51 (2003) 12–30

his poor performance on these tests. Certainly, at the basic level, his deficit with
simple curvature judgements are much less than his deficits with judging spatial
relations, and this is true also of KBN and KT.
What about defects classically associated with occipitotemporal lesions causing
adult-onset prosopagnosia? All our patients clinically described topographagnosia.
On the other hand, saturation discrimination was quite good in GA and KBN. KT
was impaired, consistent with an achromatopsia suggested by her impaired perfor-
mance with pseudo-isochromatic plates. The sparing of colour perception in GA andKBN contrasts with the universal impairment of saturation discrimination in our
adult-onset cases (Barton et al., 2003). While achromatopsia is a frequent accom-
paniment of prosopagnosia, the findings in GA and KBN confirm the independence
of the two deficits, and indicate that face and colour processing, while perhaps part
of the same ventral processing stream, are nevertheless separate functional modules.
The findings in KBN also confirm that the luminance axis of colour space can be
affected in isolation from hue and saturation—a pattern observed in one patient with
form agnosia (Milner & Heywood, 1989)—just as achromatopsic defects are said toaffect hue and saturation but not brightness perception (Heywood et al., 1987;
Heywood, Cowey, & Newcombe, 1991; Rizzo et al., 1993; Victor et al., 1989).
A review of the few other patients reported with developmental prosopagnosia
shows that, like adult-onset prosopagnosia, there may be significant variability in the
extent of associated perceptual impairment. In adult-onset cases, this variability is
part of the evidence that subtypes of prosopagnosia exist. Compared with other
patients with developmental prosopagnosia, our patients fall towards the apper-
ceptive end of the spectrum. KT in particular is likely among the most apperceptiveof cases, and has the agnosic defect that is the least specific for faces. This contrasts
with patients S (Temple, 1992) and BC (Duchaine, 2000), who have normal or near-
normal performance on the BFRT, Warrington Recognition test for faces, normal
object perception, and, in S at least, normal perception of facial age and sex. While
this constitutes only indirect evidence against an apperceptive basis to their proso-
pagnosia, S and BC are the two patients for whom an associative argument is most
plausible.
There is good direct evidence that our three patients all have perceptual encodingdeficits contributing to their prosopagnosia. All are impaired on unfamiliar face-
matching tests and the nature of this encoding deficit appears to be due to a uni-
formly severe failure in perceiving the spatial relationships of facial elements. Despite
the fact that they have other perceptual deficits on our battery, the spatial relational
defect is the most consistent finding they share with adult-onset prosopagnosics with
lesions of the fusiform face area. This argues for a common functional mechanism,
and suggests a hypofunctionality of the fusiform face area in these patients, a hy-
pothesis that can be tested with functional magnetic resonance imaging.A last point of comment is directed towards the observations made about their
social development. All three have evidence of social isolation and difficulty with
peer relationships leading to occupational and social problems. Minor abnormalities
in the social use of language were observed in GA and KBN. KBN exhibited ab-
normal non-verbal communication, with poor eye contact and paucity of expression
and gesture. KT exhibited a marked focus of interest in languages, which might
speak to an attempt to concentrate on residual abilities and strengths, but which
might also meet the DSM IV Asperger criterion of �restricted pattern of interest thatis abnormal either in intensity or focus�. These features are reminiscent of somepervasive developmental disorders, particularly Asperger syndrome. The potential
association between developmental prosopagnosia and Asperger syndrome has been
raised by others (Kracke, 1994). Furthermore, autistic traits have also been described
in congenital blindness (Brown, Hobson, & Lee, 1997; Cass, Sonksen, & McCona-
chie, 1994; Goodman & Ashby, 1990) and early onset general apperceptive visual
J.J.S. Barton et al. / Brain and Cognition 51 (2003) 12–30 27

agnosia (Jambaque, Mottron, Ponsot, & Chiron, 1998; Mottron et al., 1997). These
reports raise the possibility that impaired face recognition at a young age may im-
pede the development of social behaviour. While abnormalities of matching unfa-
miliar faces have been found in autism and Asperger syndrome (Davies, Bishop,
Manstead, & Tantam, 1994; Klin et al., 1999; Szatmari, Tuff, Finlayson, & Bart-
olucci, 1990; Tantam, Monaghan, Nicholson, & Stirling, 1989), there are fewer at-
tempts to show prosopagnosia, and the evidence is mixed (Boucher, Lewis, & Collis,
1998; Teunisse & de Gelder, 1994).If prosopagnosia can be demonstrated in Asperger patients, though, it can also be
questioned whether the face recognition deficit causes the social dysfunction, or if the
social dysfunction limits the experience and interest in faces necessary to develop
expert face processing (Carey, 1992). In our patients, the demonstration that the
perceptual impairment in spatial relation perception extends beyond faces to simple
dot patterns would argue for a primary perceptual encoding deficit rather than a
secondary one. Our data suggests that more studies of face perception and identi-
fication in Asperger syndrome are desirable, and that explorations of the general-izability of perceptual encoding defects would be valuable in any subgroup found to
have a prosopagnosic element to their disorder.
References
Albert, M., Butters, N., & Levin, J. (1979). Temporal gradients in retrograde amnesia of patients with
alcoholic Korsakoff�s disease. Archives of Neurology, 36, 211–216.Ariel, R., & Sadeh, M. (1996). Congenital visual agnosia and prosopagnosia in a child: A case report.
Cortex, 32, 221–240.
Barton, J., Cherkasova, M., & O�Connor, M. (2001a). Covert recognition in acquired and developmentalprosopagnosia. Neurology, 57, 1161–1167.
Barton, J., Cherkasova, M., Press, D., Intriligator, J., O�Connor, M. (2003). Perceptual function inprosopagnosia (in press).
Barton, J., Keenan, J., & Bass, T. (2001b). Discrimination of spatial relations and features in faces: Effects
of inversion and viewing duration. British Journal of Psychology, 92, 527–549.
Barton, J., Press, D., Keenan, J., & O�Connor, M. (2002). Lesions of the fusiform face area impair
perception of facial configuration in prosopagnosia. Neurology, 58, 71–78.
Bauer, R., & Verfaellie, M. (1988). Electrodermal discrimination of familiar but not unfamiliar faces in
prosopagnosia. Brain and Cognition, 8, 240–252.
Bentin, S., Deouell, L., & Soroker, N. (1999). Selective visual streaming in face recognition: Evidence from
developmental prosopagnosia. NeuroReport, 10, 823–827.
Benton, A., & van Allen, M. (1972). Prosopagnosia and facial discrimination. Journal of Neurological
Sciences, 15, 167–172.
Boucher, J., Lewis, V., & Collis, G. (1998). Familiar face and voice matching and recognition in children
with autism. Journal of Child Psychology and Psychiatry, 39, 171–181.
Brown, R., Hobson, R., & Lee, A. (1997). Are there ‘‘autistic-like’’ features in congenitally blind children?
Journal of Child Psychology and Psychiatry, 38, 693–703.
Bruyer, R. (1991). Covert facial recognition in prosopagnosia: A review. Brain and Cognition, 15, 223–235.
Bruyer, R., Laterre, C., Seron, X., Feyereisen, P., Strypstein, E., Pierrard, E., & Rectem, D. (1983). A case
of prosopagnosia with some preserved covert remembrance of familiar faces. Brain and Cognition, 2,
257–284.
Campbell, R. (1992). Face to face: Interpreting a case of developmental prosopagnosia. Case studies in
cognition. In R. Campbell (Ed.), Mental lives (pp. 216–236). Oxford, UK: Blackwell.
Carey, S. (1992). Becoming a face expert. Philosophical Transactions of the Royal Society of London. Series
B: Biological Sciences, 335, 95–103.
Cass, H., Sonksen, P., & McConachie, H. (1994). Developmental setback in severe visual impairment.
Archives of Diseases in Childhood, 70, 192–196.
Cooper, E., & Wojan, T. (2000). Differences in the coding of spatial relations in face identification and
basic-level object recognition. Journal of Experimental Psychology: Learning, Memory, and Cognition,
26, 470–488.
Damasio, A. (1985). Prosopagnosia. Trends in Neuroscience, 132–135.
Damasio, A., Damasio, H., & van Hoessen, G. (1982). Prosopagnosia: Anatomic basis and behavioral
mechanisms. Neurology, 32, 331–341.
28 J.J.S. Barton et al. / Brain and Cognition 51 (2003) 12–30

Damasio, A., Tranel, D., & Damasio, H. (1990). Face agnosia and the neural substrates of memory.
Annual Review of Neuroscience, 13, 89–109.
Davies, S., Bishop, D., Manstead, A., & Tantam, D. (1994). Face perception in children with autism and
Asperger�s syndrome. Journal of Child Psychology and Psychiatry, 35, 1033–1057.de Haan, E., & Campbell, R. (1991). A fifteen year follow-up of a case of developmental prosopagnosia.
Cortex, 27, 489–509.
de Haan, E., Young, A., & Newcombe, F. (1987). Faces interfere with name classification in a
prosopagnosic patient. Cortex, 23, 309–316.
de Renzi, E. (1986). Prosopagnosia in two patients with CT scan evidence of damage confined to the right
hemisphere. Neuropsychologia, 24, 385–389.
de Renzi, E., Faglioni, P., Grossi, D., & Nichelli, P. (1991). Apperceptive and associative forms of
prosopagnosia. Cortex, 27, 213–221.
Diamond, R., & Carey, S. (1986). Why faces are and are not special: An effect of expertise. Journal of
Experimental Psychology: General, 115, 107–117.
Duchaine, B. C. (2000). Developmental prosopagnosia with normal configural processing. NeuRoreport,
11, 79–83.
Farah, M. (1990). Visual agnosia: Disorders of visual recognition and what they tell us about normal vision.
Cambridge, MA: MIT Press.
Farah, M., Levinson, K., & Klein, K. (1995). Face perception and within-category discrimination in
prosopagnosia. Neuropsychologia, 33, 661–674.
Freire, A., Lee, K., & Symons, L. A. (2000). The face-inversion effect as a deficit in the encoding of
configural information: Direct evidence. Perception, 29, 159–170.
Gauthier, I., Skudlarski, P., & Gore, J. (2000). Expertise for cars and birds recruits areas involved in face
recognition. Nature Neuroscience, 3, 191–197.
Goodman, R., & Ashby, L. (1990). Delayed visual maturation and autism. Developmental Medicine and
Child Neurology, 32, 808–819.
Haxby, J., Hoffman, E., & Gobbini, M. (2000). The distributed human neural system for face perception.
Trends in Cognition Science, 4, 223–233.
Heywood, C., Wilson, B., & Cowey, A. (1987). A case study of cortical colour �blindness� withrelatively intact achromatic discrimination. Journal of Neurology, Neurosurgery and Psychiatry, 50,
22–29.
Heywood, C. A., Cowey, A., & Newcombe, F. (1991). Chromatic discrimination in a cortically colour
blind observer. European Journal of Neuroscience, 3, 802–812.
Jambaque, I., Mottron, L., Ponsot, G., & Chiron, C. (1998). Autism and visual agnosia in a child with
right occipital lobectomy. Journal of Neurology, Neurosurgery and Psychiatry, 65, 555–560.
Kanwisher, N., McDermott, J., & Chun, M. (1997). The fusiform face area: A module in human
extrastriate cortex specialized for face perception. Journal of Neuroscience, 17, 4302–4311.
Klin, A., Sparrow, S. S., de Bildt, A., Cicchetti, D. V., Cohen, D. J., & Volkmar, F. R. (1999). A normed
study of face recognition in autism and related disorders. Journal of Autism and Developmental
Disorders, 29, 499–508.
Kosslyn, S., Hamilton, S., & Bernstein, J. (1995). The perception of curvature can be selectively disrupted
in prosopagnosia. Brain and Cognition, 27, 36–58.
Kracke, I. (1994). Developmental prosopagnosia in Asperger syndrome: Presentation and discussion of an
individual case. Developmental Medicine and Child Neurology, 36, 873–886.
Landis, T., Cummings, J., Christen, L., Bogen, J., & Imbof, H.-G. (1986). Are unilateral right posterior
cerebral lesions sufficient to cause prosopagnosia? Clinical and radiological findings in six additional
patients. Cortex, 22, 243–252.
Leder, H., & Bruce, V. (2000). When inverted faces are recognized: The role of configural information in
face recognition. Quarterly Journal of Experimental Psychology, 53A, 513–536.
Malone, D., Morris, H., Kay, M., & Levin, H. (1982). Prosopagnosia: A double dissociation between
recognition of familiar and unfamiliar faces. Journal of Neurology, Neurosurgery and Psychiatry, 45,
802–812.
McConachie, H. (1976). Developmental prosopagnosia, a single case report. Cortex, 12, 76–82.
McNeil, J., & Warrington, E. (1993). Prosopagnosia: A face-specific disorder. Quarterly Journal of
Experimental Psychology, 46A, 1–10.
Meadows, J. (1974). The anatomical basis of prosopagnosia. Journal of Neurology, Neurosurgery and
Psychiatry, 37, 489–501.
Milner, A., & Heywood, C. (1989). A disorder of lightness discrimination in a case of visual form agnosia.
Cortex, 25, 489–494.
Mottron, L., Mineau, S., D�eecarie, J.-C., Jambaqu�ee, I., Labrecque, R., P�eepin, J.-P., & Aroichane, M.
(1997). Visual agnosia with bilateral temporo-occipital brain lesions in a child with autistic disorder: A
case study. Developmental Medicine and Child Neurology, 39, 699–705.
Rizzo, M., Hurtig, R., & Damasio, A. (1987). The role of scanpaths in facial recognition and learning.
Annals of Neurology, 22, 41–45.
J.J.S. Barton et al. / Brain and Cognition 51 (2003) 12–30 29

Rizzo, M., Smith, V., Pokorny, J., & Damasio, A. (1993). Color perception profiles in central
achromatopsia. Neurology, 43, 995–1001.
Simpson, T. (1995). Vision thresholds from psychometric analyses: Alternatives to probit analysis.
Optometry and Vision Science, 72, 371–377.
Szatmari, P., Tuff, L., Finlayson, M., & Bartolucci, G. (1990). Asperger�s syndrome and autism:
Neurocognitive aspects. Journal of the American Academy of Child and Adolescent Psychiatry, 29, 130–
136.
Takahashi, N., Kawamura, M., Hirayama, K., Shiota, J., & Isono, O. (1995). Prosopagnosia: A clinical
and anatomic study of four patients. Cortex, 31, 317–329.
Tantam, D., Monaghan, L., Nicholson, H., & Stirling, J. (1989). Autistic children�s ability to interpretfaces: A research note. Journal of Child Psychology and Psychiatry, 30, 623–630.
Temple, C. (1992). Developmental memory impairment: Faces and patterns. In R. Campbell (Ed.),Mental
lives. Case studies in cognition (pp. 99–215). Oxford, UK: Blackwell.
Teunisse, J.-P., & de Gelder, B. (1994). Do autistics have a generalized face processing deficit?
International Journal of Neuroscience, 77, 1–10.
Victor, J., Maiese, K., Shapley, R., Sitdis, J., & Gazzaniga, M. (1989). Acquired central dyschromatopsia:
Analysis of a case with preservation of color discrimination. Clinical Vision Science, 4, 183–196.
Warrington, E. (1984).Warrington recognition memory test. Los Angeles: Western Psychological Services.
Warrington, E., & James, M. (1991). The visual object and space perception battery. Bury St. Edmunds:
Thames Valley Test Company.
Wechsler, D. (1981). The Wechsler adult intelligence scale-revised. New York: Psychological Corporation.
Young, A. (1994). Covert recognition. In M. Farah, & G. Ratcliff (Eds.), The neuropsychology of high-level
vision. Hillsdale, NJ: Lawrence Erlbaum Associates.
Young, A., & Ellis, H. (1989). Childhood prosopagnosia. Brain and Cognition, 9, 16–47.
30 J.J.S. Barton et al. / Brain and Cognition 51 (2003) 12–30




















