
Developmental exposure to decabrominated diphenyl ether (BDE-209): Effects on sperm oxidative stress...
10
Developmental Exposure to Decabrominated Diphenyl Ether (BDE-209): Effects on Sperm Oxidative Stress and Chromatin DNA Damage in Mouse Offspring Li-Ho Tseng, 1 Ping-Chi Hsu, 2 Chia-Wei Lee, 2 Shinn-Shyong Tsai, 3 Min-Hsiung Pan, 4 Mei-Hui Li 5 1 Department of Occupational Safety and Hygiene, Tajen University, Pingtung, Taiwan 2 Department of Safety, Health, and Environmental Engineering, National Kaohsiung First University of Science and Technology,Kaohsiung, Taiwan 3 Department of Veterinary Medicine, National Pingtung University of Science and Technology, Pingtung, Taiwan 4 Department of Seafood Science, National Kaohsiung Marine University, Kaohsiung, Taiwan 5 Environmental Toxicology Lab, Department of Geography,National Taiwan University, Taipei, Taiwan Received 15 December 2010; revised 27 March 2011; accepted 30 March 2011 ABSTRACT: Polybrominated diphenyl ethers (PBDEs) are used as brominated flame retardants and have been found in human milk in recent years. This study investigates whether prenatal exposure to decabro- minated diphenyl ether (BDE-209) induces sperm dysfunction in male offspring. Pregnant CD-1 mice were gavaged once daily with corn oil (control), 10, 500, and 1500 mg kg 21 body weight of BDE-209 from day 0 of gestation to day 17. The outcomes of male reproductive parameters were assessed on postnatal day 71. Anogenital distance, sperm-head abnormalities, and testicular histopathology were significantly affected in male offspring prenatally exposed to 1500 mg kg 21 . Significant increases in the tendency for sperm DNA denaturation (aT) induction and the DNA fragmentation index (DFI) were found in those exposed to 10, 500, and 1500 mg kg 21 (P \ 0.05). We observed a significant increase of sperm hydrogen peroxide (H 2 O 2 ) generation in the 10 and 1500 mg/kg/day groups compared to the control group (P \ 0.05). Although our findings suggested that the mechanisms underlying BDE-209-induced sperm DNA damage and H 2 O 2 generation might not be represented as a dose-response relationship, we found that the greater the excess production of sperm H 2 O 2 , the greater the sperm aT(r 5 0.65, P 5 0.0155) and DFI (r 5 0.53, P 5 0.002). In conclusion, developmental exposure to BDE-209 induced sperm-head abnormal- ity, oxidative stress, chromatin DNA damage, and testicular histopathological changes. These findings suggest that BDE-209-induced male reproductive effects might involve the formation of sperm H 2 O 2 which attacks nucleic acids via H 2 O 2 generation. # 2011 Wiley Periodicals, Inc. Environ Toxicol 28: 380–389, 2013. Correspondence to: Dr. Ping-Chi Hsu; e-mail: [email protected] Published online 27 May 2011 in Wiley Online Library (wileyonlinelibrary.com). DOI 10.1002/tox.20729 Contract grant sponsor: National Science Council of Taiwan Contract grant number: NSC 96-2314-B-327-001 MY3, NSC 97-2221- E-327-009-MY3 C 2011 Wiley Periodicals, Inc. 380
Transcript of Developmental exposure to decabrominated diphenyl ether (BDE-209): Effects on sperm oxidative stress...

Developmental Exposure to DecabrominatedDiphenyl Ether (BDE-209): Effects on SpermOxidative Stress and Chromatin DNA Damage inMouse Offspring
Li-Ho Tseng,1 Ping-Chi Hsu,2 Chia-Wei Lee,2 Shinn-Shyong Tsai,3 Min-Hsiung Pan,4
Mei-Hui Li5
1Department of Occupational Safety and Hygiene, Tajen University, Pingtung, Taiwan
2Department of Safety, Health, and Environmental Engineering, National Kaohsiung FirstUniversity of Science and Technology, Kaohsiung, Taiwan
3Department of Veterinary Medicine, National Pingtung University of Science and Technology,Pingtung, Taiwan
4Department of Seafood Science, National Kaohsiung Marine University, Kaohsiung, Taiwan
5Environmental Toxicology Lab, Department of Geography, National Taiwan University, Taipei,Taiwan
Received 15 December 2010; revised 27 March 2011; accepted 30 March 2011
ABSTRACT: Polybrominated diphenyl ethers (PBDEs) are used as brominated flame retardants and havebeen found in human milk in recent years. This study investigates whether prenatal exposure to decabro-minated diphenyl ether (BDE-209) induces sperm dysfunction in male offspring. Pregnant CD-1 mice weregavaged once daily with corn oil (control), 10, 500, and 1500 mg kg21 body weight of BDE-209 from day 0of gestation to day 17. The outcomes of male reproductive parameters were assessed on postnatal day71. Anogenital distance, sperm-head abnormalities, and testicular histopathology were significantlyaffected in male offspring prenatally exposed to 1500 mg kg21. Significant increases in the tendency forsperm DNA denaturation (aT) induction and the DNA fragmentation index (DFI) were found in thoseexposed to 10, 500, and 1500 mg kg21 (P\ 0.05). We observed a significant increase of sperm hydrogenperoxide (H2O2) generation in the 10 and 1500 mg/kg/day groups compared to the control group (P \0.05). Although our findings suggested that the mechanisms underlying BDE-209-induced sperm DNAdamage and H2O2 generation might not be represented as a dose-response relationship, we found thatthe greater the excess production of sperm H2O2, the greater the sperm aT (r 5 0.65, P 5 0.0155) and DFI(r5 0.53, P 5 0.002). In conclusion, developmental exposure to BDE-209 induced sperm-head abnormal-ity, oxidative stress, chromatin DNA damage, and testicular histopathological changes. These findingssuggest that BDE-209-induced male reproductive effects might involve the formation of sperm H2O2
which attacks nucleic acids via H2O2 generation. # 2011 Wiley Periodicals, Inc. Environ Toxicol 28: 380–389, 2013.
Correspondence to: Dr. Ping-Chi Hsu; e-mail: [email protected]
Published online 27 May 2011 in Wiley Online Library
(wileyonlinelibrary.com). DOI 10.1002/tox.20729
Contract grant sponsor: National Science Council of Taiwan
Contract grant number: NSC 96-2314-B-327-001 MY3, NSC 97-2221-
E-327-009-MY3
�C 2011 Wiley Periodicals, Inc.
380

Keywords: polybrominated diphenyl ethers (PBDEs); decabrominated diphenyl ether (BDE-209);prenatal exposure; oxidative stress; sperm DNA damage
INTRODUCTION
Brominated flame retardants (BFRs) have been widely used
in many products for fire prevention purposes since the late
1970s (WHO, 1994). The most commonly used BFRs
are the polybrominated diphenyl ethers (PBDE), hexabro-
mocyclododecane (HBCD), and tetrabromobisphenol A
(TBBPA). Decabrominated diphenyl ether (BDE-209), the
second most used BFR after TBBPA, is used in the produc-
tion of electronic enclosures and upholstery textiles and
foams (Hardy, 2002). During the manufacturing process,
use, or disposal of such products, PBDEs may leach into
the environment (Sjodin et al., 2001).
These persistent and bioaccumulative compounds
(Fangstrom et al., 2005; Streets et al., 2006) have been
found not only in the environment (Ikonomou et al.,
2002; Kelly et al., 2008) but also in the blood of preg-
nant women (Bradman et al., 2007) and in breast milk
(Kalantzi et al., 2009). Elevated concentrations of BDE-
209 have also been found in waterbird egg samples (Lam
et al., 2007), wild harbor seals (Shaw et al., 2008), and
plasma from polar bears and glaucous gulls (Verreault
et al., 2005).
The effects on human health of increased of PBDE
levels in the environment on the health of humans is a
cause for concern (Zoeller, 2005; Main et al., 2007).
Although the EU banned the manufacture and use of
penta-BDE and octa-BDE (EU, 2003) and both products
were subsequently withdrawn from the US market (Betts,
2008), the environmental and occupational pollution of
PBDEs remain a major concern. There is, therefore, a
continuing need to study their effects. Workers disman-
tling electronics (Muenhor et al., 2010), computer techni-
cians (Jakobsson et al., 2002), and laborers involved in
the manufacturing or handling of rubber (Thuresson
et al., 2005) have been found to be exposed to BDE-209.
Recently, a survey in the general population indicated
that certain higher brominated PBDEs appear to affect
the menstruation characteristics of reproductive-age
females (Chao et al., 2010). However, the toxicity of
BDE-209 is not clearly understood.
A study from Indiana found a close association between
maternal and fetal cord blood PBDE concentrations
(Mazdai et al., 2003). Another study, from the EU, found a
similar relative distribution of PBDEs, especially highly
brominated PBDEs, between maternal and fetal serum
(Antignac et al., 2009). These studies showed that PBDEs
may be able to cross the placental barrier and enter fetal cir-
culation. PBDEs are potential endocrine disruptors and
may have an effect on the reproductive system. In a study
of the reproductive effects of PBDEs on mice, Kuriyama
et al. (2005) reported that prenatal exposure to a single low
dose of PBDE 99 on gestation day (GD) 6 resulted in sig-
nificantly decreased sperm counts in offspring. Lilienthal
et al. (2006) have also reported that prenatal exposure to
PBDE 99 resulted in reduced testosterone and anogenital
distance (AGD) in adult male rat offspring. To the best of
our knowledge, however, very few studies have investi-
gated the effects of BDE-209 exposure during pregnancy
on the relationship between sperm oxidative stress and
DNA damage in adult offspring.
In this study, we examined the effects of prenatal expo-
sure to BDE-209 on sperm and testis dysfunction in the off-
spring of mice. To do this, we:
1. evaluated infant development, including body and repro-
ductive organ weights, AGD, anogenital index (AGI,
weight-adjusted AGD), and testis index (TI),
2. characterized sperm function—including sperm count,
motility, motion, morphology, sperm chromatin struc-
ture analysis (SCSA), and sperm reactive oxygen species
(ROS) generation,
3. assessed testicular functions, including testosterone hor-
mone levels and histopathology.
The results of this study may provide additional useful
information on the extent of BDE-209-related adverse
effects on the environmental toxicology and help make pos-
sible the assessment of health risk.
MATERIALS AND METHODS
Overview of Study Design
Adult male (12-weeks old) and female (10-weeks old) CD-
1 mice were provided by the Animal Center of National
Cheng Kung University Medical Center (Tainan, Taiwan).
The animal chamber was supplied with UV sterilized air
and maintained at 248C 6 28C with a constant humidity of
60% 6 10%. The mice were maintained on a 12-h light/
dark cycle. Food (Laboratory Rodent Diet 5001, LabDiet,
Richmond, IN) and distilled water were provided ad libi-tum. After 2 weeks of acclimation, the breeding process
was conducted by placing one female mouse into a male
mouse’s cage. The day of vaginal plug was defined as GD
0. Twenty pregnant females were randomly divided into
four groups, each containing with five mice. Each mouse
was housed individually. Animals from each group were
gavaged once daily with corn oil (control), 10, 500, and
381DEVELOPMENTAL EXPOSURE TO DECABROMINATED DIPHENYL ETHER (BDE-209)
Environmental Toxicology DOI 10.1002/tox

1500 mg kg21 of body weight of BDE-209 from GD 0 to
GD 17. Dosages were adjusted daily on the basis of
changes in body-weight. On postnatal day (PND) 71, three
male offspring from each litter were randomly selected, to
make a total of 15 from each group. Their AGD and AGI
were then measured. AGD was defined as the distance from
the center of the anus to the anterior base of the penis. AGI
was estimated after adjusting for body weight. The animals
were euthanized by CO2. The testis, epididymis, and semi-
nal vesicles were removed and weighed. Blood samples
were taken by heart puncture for testosterone analysis. The
left cauda epididymis was used to perform SCSA. The right
cauda epididymis was prepared for sperm suspension in
order to measure sperm count, motility, morphology, and
ROS generation.
Chemicals
BDE-209 (98%, CAS no. 1163-19-5) was obtained from
Sigma-Aldrich (St. Louis, MO). The BDE-209 solution
was prepared by mixing the BDE-209 compound with
corn oil and sonicating the mixture at room temperature
for 30 min. The desired doses (10, 500, or 1500 mg
kg21) were prepared weekly. The suspensions were vor-
texed during dosing for at least 30 min. Hydroethidine
(HE) and 20,70-dichlorofluorescin diacetate (DCFH-DA)
were purchased from Molecular Probes (Eugene, OR).
Corn oil, phosphate-buffered saline (PBS), Ham’s F12
medium, eosin Y, propidium iodide (PI) stain, and all
other reagents were obtained from Sigma-Aldrich (St.
Louis, MO).
Sperm Count, Motility, Motion Analysis
The right cauda epididymis was removed and placed in a
medium composed of 1 mL Ham’s F12 and maintained
at 348C in an environment saturated with 5% CO2. After
5 min, cauda epididymis was minced with curved scissors
and the sperm was dispersed. After 5 min, the sperm was
collected and transferred to a fresh tube. Sperm suspen-
sion (10 lL) was placed on a prewarmed Makler cham-
ber (10 lm depth; Sefi-Medical Instruments, Haifa,
Israel). Spermatozoa were counted and the ratio between
the number of motile spermatozoa and total number was
used to express sperm motility. Sperm motion parameters
were analyzed from videotaped images using computer-
assisted sperm analysis (CASA) obtained for velocity
indices with a Hamilton Thorn Research motility analyzer
(version HTMIVOS Specification, Beverly, MA). Motion
parameters, including curvilinear velocity (VCL, lm
s21), angular progressive velocity (VAP, lm s21),
straight-line velocity (VSL, lm s21), lateral head ampli-
tude (ALH, lm), and beat-cross frequency (BCF, Hz)
were also measured.
Sperm Morphology Analysis
Morphological abnormalities were classified based on a
modification of a method reported in Wyrobek and Bruce
(1975). Briefly, the sperm suspension was diluted with
phosphate buffered saline (PBS) and then mixed with 1%
aqueous eosin Y (10:1), smeared onto glass slides, and air-
dried. Slides were briefly rinsed in methanol to remove
excess stain. They were air dried again and cover-slipped
with mounting medium. Two samples were made for each
mouse. At least 600 cells from each mouse were examined
for morphological abnormalities under a light microscope
(4003) (Zeiss, Axioskop2, Germany). Head was defined as
abnormal when it lacked the usual hook, or had a banana-
shaped, triangular, or giant head.
Flow Cytometry (FCM) Analysis for SpermChromatin DNA Damage
The SCSA described by Evenson et al. (2002) is used to
detect the susceptibility of sperm to in situ acid denatura-
tion of DNA. The sample was analyzed using FCM (BD
Immunocytometry Systems, San Jose, CA). A total of
10 000 spermatozoa were analyzed. AO is a metachromatic
fluorochrome used to distinguish stain double-stranded
from single-stranded nucleic acids. AO intercalates into
double-stranded DNA and has green fluorescence, and it
binds to single-stranded nucleic acids and had has red fluo-
rescence. In SCSA analysis, the metachromatic shift from
green to red fluorescence was expressed as aT, a ratio
between red and total fluorescence [red/(red 1 green)].
Each sample was calculated and results were expressed as
the mean of the aT distribution (X aT) reflecting the level
of sperm with DNA damage. Spermatozoa with abnormal
chromatin structure or DNA damage were represented by
DNA fragmentation index (DFI), formally called COMP aT(the percentage of cells outside the main population of aT).
Sperm ROS Assay
Modifying a method reported by Fisher et al. (2005), we
measured sperm hydrogen peroxide (H2O2) levels. Briefly,
sperm cells (1 3 106 cells mL21) were added into 2.5 mMDCFH-DA to a final concentration of 12.5 lM. DCFH-DA
is a stable compound that passively diffuses into cells and
is hydrolyzed by intracellular esterase to yield DCHF,
which is trapped inside cells. H2O2 produced by cells oxi-
dizes DCHF to the highly fluorescent compound, 20,70-dichlorofluorescein (DCF), which is fluorescent at 530 nm.
Sperm superoxide anion (O22l) levels were measured using
a modification of a previously described method (Marchetti
et al., 2002). Sperm cells at a concentration of 1 3 106 cells
mL21 were added into 0.33 mM HE to a final concentration
of 2 lM. The mixture was set aside and maintained at 348Cfor 30 min. Sperm H2O2 and O22l levels were measured
382 TSENG ET AL.
Environmental Toxicology DOI 10.1002/tox

using a FCM with excitation and emission set at 488 and
530 nm, respectively.
Assessment of TI and Serum TestosteroneAssay
TI was calculated for each mouse (testicular length 3 tes-
ticular width/body weight). Testosterone was measured
using Coat-a-Count radioimmunoassay kits obtained from
Diagnostic Products Corporation (Los Angeles, CA) and
performed following manufacturer’s directions. The assays
were performed in the Department of Nuclear Medicine at
National Cheng Kung University Medical Center (Tainan,
Taiwan).
Histopathology of Testis
The left testis was fixed in Bouin’s solution for at least
48 h. To prepare them for histological studies, the testes
were embedded in paraffin and sections (3 to 4 lm) were
stained with hematoxylin and eosin. The damage to semi-
niferous tubules was evaluated in cross sections. Sections
were observed under 4003 magnification with an Olympus
microscope (Milton Keynes, Buckinghamshire, UK) con-
nected to a computer on which the digital images were
saved. Each germ cell stage in the seminiferous epithelium
was categorized according to a classification system used
by Oakberg (1956).
Statistics
Results were expressed as mean 6 standard error of the
mean (SEM). For the analysis of male offspring, the litter
was the statistical unit. The body weights, reproductive
organ weights, AGD, TI, sperm count, motility, velocity,
morphology, SCSA, generation of H2O2 and O22l of the
groups exposed to BDE-209 and the control group were
compared using one-way analysis of variance (ANOVA),
followed by the Tukey-Kramer honestly significant differ-
ence (HSD). The serum testosterone data were not normally
distributed, so log transformed testosterone levels were per-
formed. A P-value of\0.05 was considered significant. All
statistical operations were performed using the JMP 5.0 sta-
tistical package (SAS Institute, Gary, NC).
RESULTS
Body and Reproductive Organ Weights ofMale Offspring
We found no significant differences between any of the
exposed groups or between those groups and the controls
with regard to body weight, absolute or relative weights of
testis, epididymis, cauda epididymis, and seminal vesicles
at PND 71 (data not shown).
Sperm Count, Motility, Motion, andMorphology Analysis
As can be seen in Table I, a summary of average sperm
count, motility, motion, and morphology for each of the
treatment groups, there was a significantly greater increase
in mean percentages of sperm morphological abnormalities
in the 1500 mg kg21-treated offspring than in the controls
(P\ 0.05), though there were no significant differences in
sperm count and motility among the treatment groups.
Based on our CASA motion analysis of cauda epididymal
TABLE I. Sperm count, motility, morphology, and velocity of CD-1 male mice prenatally exposed to decabrominateddiphenyl ether (BDE-209)
Parameters
Treatment of BDE-209 (mg kg21)
P valueControl 10 500 1500
Sperm count (106/mL) 15.5 6 1.9 13.1 6 1.1 13.9 6 0.7 16.2 6 1.2 0.3304
Sperm motility (%) 65.6 6 1.5 64.3 6 4.6 66.1 6 4.4 67.1 6 1.3 0.8826
Abnormal sperms head (%)a 10.3 6 2.5 14.9 6 1.4 15.5 6 0.4 18.2 6 1.3* 0.0207
Velocity of motionVCL (lm s21)b 145.8 6 7.6 147.9 6 7.3 149.3 6 7.7 148.5 6 9.0 0.9908
VAP (lm s21)c 80.3 6 4.9 82.6 6 3.0 79.6 6 3.3 80.9 6 4.8 0.9597
VSL (lm s21)d 59.8 6 5.1 63.7 6 3.0 59.4 6 3.0 62.3 6 4.1 0.8453
ALH (lm)e 6.46 6 0.2 6.7 6 0.4 6.8 6 0.5 7.0 6 0.9 0.8285
All data was expressed in means 6 SEM.
*P\0.05 as compared with control group.a Based on scoring 600 sperm per animal (mean 6 SEM).b VCL: curvilinear velocity.c VAP: Average path velocity.d VSL: straight line velocity.e ALH: amplitude of lateral head displacement.
383DEVELOPMENTAL EXPOSURE TO DECABROMINATED DIPHENYL ETHER (BDE-209)
Environmental Toxicology DOI 10.1002/tox

sperm from male offspring, there were no significant differ-
ences in sperm motion parameters, including VCL, VAP,
VSL, ALH, and BCF.
SCSA and ROS Generation
The sperm function of male offspring for each treatment
group is shown in Table II. Measurement of sperm chroma-
tin structure after in situ denaturation by flow cytometric
methods represents a relatively stable and sensitive indica-
tor of conventional sperm quality characteristics (concen-
tration, motility, and morphology). Epididymal sperm was
analyzed by FCM SCSA to investigate whether there was
any induced change in sperm chromatin DNA integrity af-
ter in utero exposure to BDE-209. We found a significant
increase in the two indicators of sperm chromatin DNA
damage, X aT and DFI, in the 10, 500, and 1500 mg kg21
day21 groups compared to the control group (P \ 0.05)
(Table II). To investigate sperm oxidation stress, we next
studied sperm H2O2 and O22l generation. We observed a
significant increase of sperm H2O2 generation in the 10 and
1500 mg kg21 day21 groups compared to the control group
(P \ 0.05) (Table II). However, there were no dose-
dependent effects in sperm DNA damage and H2O2
generation among BDE-209-treated groups and controls.
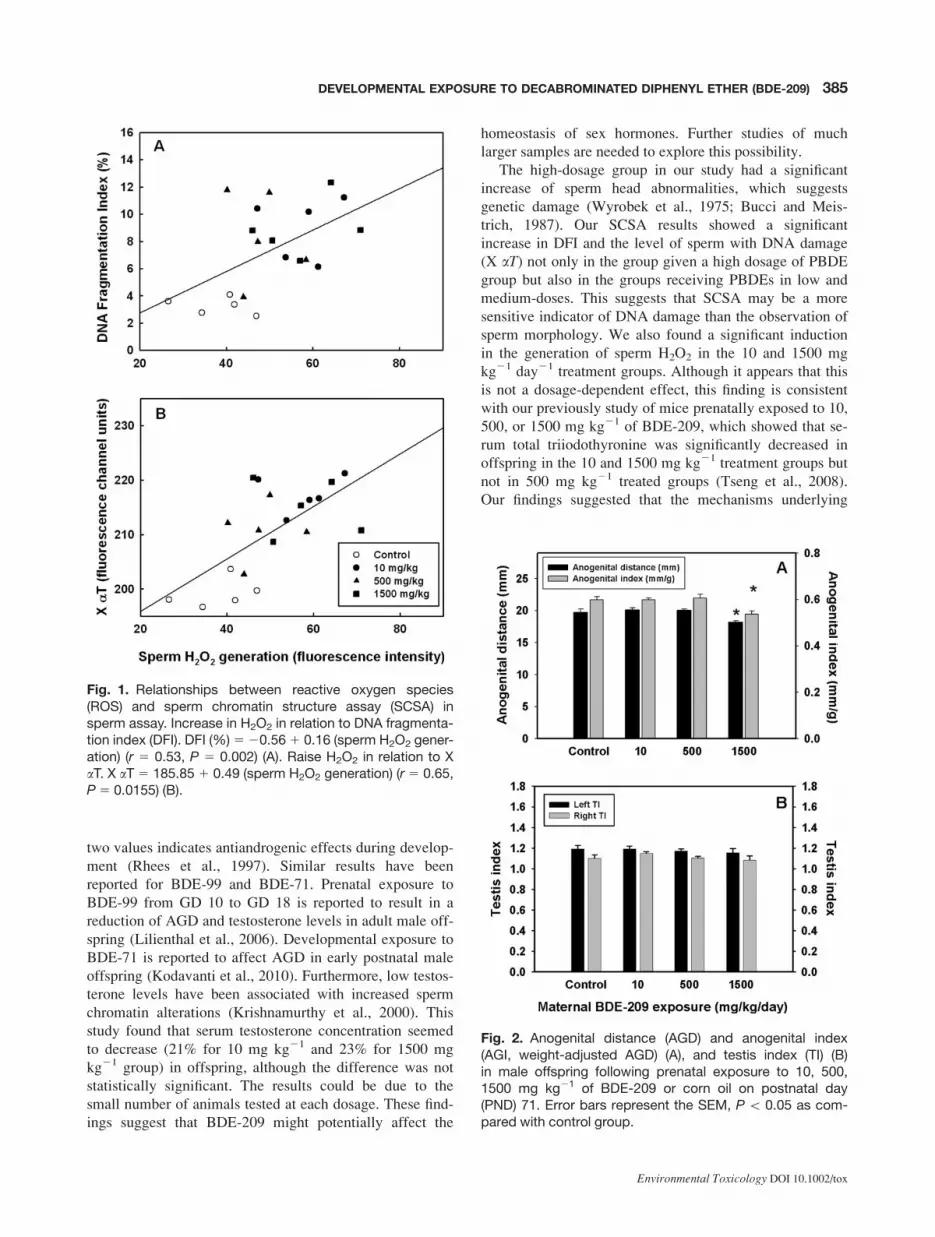
However, the percentage of sperm with excessive H2O2
generation was found to be significantly correlated with
DFI (r 5 0.53; P 5 0.002) [Fig. 1(A)] and X aT (r 5 0.65,
P 5 0.0155) [Fig. 1(B)].
AGD, AGI, and TI Measurement and SerumTestosterone Hormone Levels
The mean AGD and AGI measured in male offspring on
PND 71 were significantly reduced in male mice in utero-exposed to the highest dose (1500 mg kg21) compared to
the controls [Fig. 2(A)]. We found no significant group dif-
ferences in TI [Fig. 2(B)]. Although there was a marked
reduction for all BDE-209 doses of testosterone levels,
according to the statistical analysis, they did not result as
significant changes as compared to the control group
(Table II).
Histopathology of Testis
On PND 72, control offspring mice were found to have nor-
mal testicular morphology and spermatogenesis [Fig. 3(A)].
In the BDE-209-treated offspring, however, pathological
lesions, mainly in the interstitial cells and/or seminiferous
tubules were found. The cytoyplasm of the interstitial cells
of male offspring treated with 10 mg kg21 BDE-209
showed moderate vacuolization [Fig. 3(B)]. Those treated
with 500 mg kg21 group had similar but less severe
changes of vacuolization [Fig. 3(C)]. Those treated with
1500 mg kg21 BDE-209 had many interstitial cells and
seminiferous tubules with severe vacuolization and had
almost lost all spermatozoa and spermatids. The vacuoliza-
tion in the tubules was probably Sertoli cells suggesting
direct toxicity and apoptosis of sperm, whereas vacuoliza-
tion in Leydig cells indicated an accumulation of hormone
precursors, though basement membrane remained intact
and a few spermatogonia remained [Fig. 3(D)]. Histopatho-
logic findings are summarized in Table III.
DISCUSSION
Studies have documented the transfer of PBDEs through
the placenta to the fetus (Mazdai et al., 2003; Schecter
et al., 2007). In this study, we hypothesized that maternal
exposure to BDE-209 might have adverse effects on the
reproductive system of male offspring. The adverse effects
are summarized in Table IV.
In this study, we observed that the highest BDE-209
dose (1500 mg kg21 day21) significantly decreased the
AGD and AGI of male offspring. The reduction of these
TABLE II. Sperm DNA damage, reactive oxygen species (ROS) generation, and serum testosterone levels frommalemice in prenatal exposed to decabrominated diphenyl ether (BDE-209)
Parameters
Treatment of BDE-209 (mg kg21)
P valueControl 10 500 1500
X aTa 199.2 6 1.2 217.4 6 1.5* 208.2 6 3.3* 215.0 6 2.3* 0.0001
DFI (%)b 3.3 6 0.3 8.9 6 1.0* 7.5 6 1.8* 8.9 6 0.9* 0.0081
Sperm H2O2 Generationc 38.1 6 3.5 57.7 6 3.4* 43.9 6 2.2 57.8 6 4.5* 0.0017
Sperm O22� Generationc 248.4 610.6 296.2 6 14.9 305.9 6 13.9 290.3 6 21.6 0.0899
Log serum testosterone levels (ng mL21) 0.95 6 0.19 0.35 6 0.10 0.46 6 0.24 0.32 6 0.18 0.1007
All data was expressed in means 6 SEM.
*P\0.05 as compared with control group.a X aT is the level of sperm with DNA damage.b DNA fragmentation index (DFI) is the % of sperm with chromatin DNA damage.c Fluorescence intensity.
384 TSENG ET AL.
Environmental Toxicology DOI 10.1002/tox
two values indicates antiandrogenic effects during develop-
ment (Rhees et al., 1997). Similar results have been
reported for BDE-99 and BDE-71. Prenatal exposure to
BDE-99 from GD 10 to GD 18 is reported to result in a
reduction of AGD and testosterone levels in adult male off-
spring (Lilienthal et al., 2006). Developmental exposure to
BDE-71 is reported to affect AGD in early postnatal male
offspring (Kodavanti et al., 2010). Furthermore, low testos-
terone levels have been associated with increased sperm
chromatin alterations (Krishnamurthy et al., 2000). This
study found that serum testosterone concentration seemed
to decrease (21% for 10 mg kg21 and 23% for 1500 mg
kg21 group) in offspring, although the difference was not
statistically significant. The results could be due to the
small number of animals tested at each dosage. These find-
ings suggest that BDE-209 might potentially affect the
homeostasis of sex hormones. Further studies of much
larger samples are needed to explore this possibility.
The high-dosage group in our study had a significant
increase of sperm head abnormalities, which suggests
genetic damage (Wyrobek et al., 1975; Bucci and Meis-
trich, 1987). Our SCSA results showed a significant
increase in DFI and the level of sperm with DNA damage
(X aT) not only in the group given a high dosage of PBDE
group but also in the groups receiving PBDEs in low and
medium-doses. This suggests that SCSA may be a more
sensitive indicator of DNA damage than the observation of
sperm morphology. We also found a significant induction
in the generation of sperm H2O2 in the 10 and 1500 mg
kg21 day21 treatment groups. Although it appears that this
is not a dosage-dependent effect, this finding is consistent
with our previously study of mice prenatally exposed to 10,
500, or 1500 mg kg21 of BDE-209, which showed that se-
rum total triiodothyronine was significantly decreased in
offspring in the 10 and 1500 mg kg21 treatment groups but
not in 500 mg kg21 treated groups (Tseng et al., 2008).
Our findings suggested that the mechanisms underlying
Fig. 1. Relationships between reactive oxygen species(ROS) and sperm chromatin structure assay (SCSA) insperm assay. Increase in H2O2 in relation to DNA fragmenta-tion index (DFI). DFI (%)520.561 0.16 (sperm H2O2 gener-ation) (r 5 0.53, P 5 0.002) (A). Raise H2O2 in relation to XaT. X aT 5 185.85 1 0.49 (sperm H2O2 generation) (r 5 0.65,P5 0.0155) (B).
Fig. 2. Anogenital distance (AGD) and anogenital index(AGI, weight-adjusted AGD) (A), and testis index (TI) (B)in male offspring following prenatal exposure to 10, 500,1500 mg kg21 of BDE-209 or corn oil on postnatal day(PND) 71. Error bars represent the SEM, P\ 0.05 as com-pared with control group.
385DEVELOPMENTAL EXPOSURE TO DECABROMINATED DIPHENYL ETHER (BDE-209)
Environmental Toxicology DOI 10.1002/tox

BDE-209-induced oxidative stress may not be represented
as a linear response. Shiner et al. (2006) found that cellular
oxidative stress had a biphasic U-shaped effect on the mac-
rophage antioxidant emzymatic activity, possibly because
toxins might lead to opposite effects when used at low and
high dosages, as has been found in the effects of other envi-
ronmental fluctuations (Arumugam et al., 2006; Ji et al.,
2006). Another study has found tissue and whole-body
elimination of PBDE 47 to be biphasic (Staskal et al.,
2005). Because it is not clear whether BDE-209 has bipha-
sic effects, however, further investigation is required.
An epidemiological study by Akutsu et al. (2008)
reported strong inverse correlations between the serum
2,20,4,40,5,50-hexabromodiphenyl ether levels and sperm
concentrations and testes sizes in Japanese. That study did
not provide clear the pathological evidence of the relation-
ship, however. We found the pathological lesions in the in-
terstitial cells and/or seminiferous tubules in BDE-209-
treated offspring. We also found vacuolization levels had a
tendency to be U-shaped in testicular interstitial cells in our
histopathological assays. Only the 1500 mg kg21 BDE-
209-treated group showed severe vacuolization of seminif-
erous tubules associated with almost complete loss of sper-
matozoa and spermatids. We did not, however, find signifi-
cant changes in epididymal sperm count in the 1500 mg
kg21 BDE-209-treated group. Additional large numbers of
animals tested at each dosage are needed to confirm the
results of the present study.
Fig. 3. Tissue sections of offspring testis stained with H&E were obtained from controland BDE-209-treated mice whose dams were exposed to BDE-209 from gestation day(GD) 0 to GD17. Control group shows normal testicular morphology (A). The interstitialcells of the testis treated with 10 mg kg21 BDE-209 show moderate vacuolar degeneration(B). Similar, but less severe, changes were found in the 500 mg kg21 BDE-209-treatedmice (C). In 1500 mg kg21 BDE-209 treated mice; the seminiferous tubules show severevacoualization associated with complete losses of spermatozoa and spermatids, but stillretained both basement membrane and a few spermatogonia. *, vacuolization of seminif-erous tubules; arrowhead, vacuolization of interstitial cells; Scale bar 5 100 lm. [Colorfigure can be viewed in the online issue, which is available at wileyonlinelibrary.com.]
386 TSENG ET AL.
Environmental Toxicology DOI 10.1002/tox

The low dose of BDE-209 administered to the dams was
�1.38 times higher than the human cord serum BDE-209
concentrations reported for women in France, who were
found to have a maximal level of about 363 ng g21 of body
fat weight for BDE-209 in serum lipid (Antignac et al.,
2009). Assuming that 20% of the body weight of an adult
mouse is composed of adipose tissue, such a concentration
could be achieved by administering a single dose of 0.073
mg BDE-209/kg BW. BDE-209 is poorly absorbed, rapidly
eliminated and marginally distributed to adipose tissue,
thus oral absorption has been reported to only be about 1%
of the BDE-209 dose in rats (Morck et al., 2003). Accord-
ing to our calculations, a dose of 10 mg kg21 was 1.38
times higher than maternal exposure from cord serum. We
administered 10 mg kg21 at the lowest dose to the pregnant
female mice in our study. The hypothesis of the high dos-
age of BDE-209 was used to predict the potential effects on
occupational exposure workers. Occupational exposure to
higher levels of BDE-209 can occur in workers manufactur-
ing or handling flame-retarded products. The highest BDE-
209 concentration was 3100 ng g21 lipid in electronics dis-
mantling workers (Bi et al., 2007). The high-dosage group
(1500 mg kg21) had a level of BDE-209 that was at least a
207 times higher level of BDE-209 than current maternal
cord serum levels and a 24 times higher level of BDE-209
than workers’ serum levels. From the reproduction toxicity
point of view, the chance of humans being exposed to such
a high dose of BDE-209 is very small. Findings in mice
prenatally exposed to high levels of BDE-209, however,
provide valuable information on male reproductive effects
of PBDEs and related chemicals in humans. The evidence
found concerning the adverse effects of PBDE congeners
both in our animal studies and those of others, suggests that
further studies using environmentally relevant doses are im-
portant for hazard identification.
Oxidative stress is one of the major contributors to DNA
damage in the male germ line. Moreover, DNA damage is a
major contributor to infertility, miscarriage and birth
defects in the offspring (reviewed by Aitken and De Iuliis,
2010). It has been proposed that the source of the oxidative
stress responsible for sperm DNA damage has been
involves a loss of antioxidant protection, infection, chemi-
cal toxicants exposure, and intrinsic radical generation by
spermatozoa. BDE-209-induced sperm oxidative damage
could therefore be associated with ROS generation. At pres-
ent, there is very little information available on the impact
of BDE-209 on ROS generation and DNA damage in sper-
matozoa. The mechanisms responsible for the association
between BDE-209 exposure and ROS generation are
unknown and validation of this concept is required.
This is the first study to document the lowest-observed-
adverse-effect level (LOAEL) for sperm DNA damage and
TABLE III. Pathological changes in testis tissue of male mice prenatally exposed to decabrominated diphenyl ether(BDE-209)
Pathological changes
Control (a/b) Treatment of BDE-209 (a/b)
0 10 mg kg21 500 mg kg21 1500 mg kg21
Vacuolization in interstitial cells 0/5 2/5 11 3/5 1 3/5 11
2/5 1 2/5 1
Vacuolization in seminiferous tubules 0/5 0/5 0/5 2/5 111
2/5 1
a/b: number of mice with positive pathological change/total number of mice in each group.
1: slight; 11: moderate; 111: severe.
TABLE IV. A summary of adverse effects on various endpoints related to in male offspring prenatally exposed todecabrominated diphenyl ether (BDE-209)
Endpoints
Doses
10 mg kg21 500 mg kg21 1500 mg kg21
Anogenital distance (AGD) --- --- ;*
Abnormal sperms head --- --- :*
Sperm chromatin DNA damage levels (X aT) :* :* :*
DNA fragmentation index (DFI) :* :* :*
H2O2 generation :* --- :*
Histopathology of testis 11 1 111
---: no significant effects.* : P\0.05 as compared to control group.
1: slight; 11: moderate; 111: severe.
387DEVELOPMENTAL EXPOSURE TO DECABROMINATED DIPHENYL ETHER (BDE-209)
Environmental Toxicology DOI 10.1002/tox

excessive H2O2 production resulting from in utero exposure
to doses as low as 10 mg kg21 day21. In conclusion, our
study has found that lower concentrations of BDE-209
might cause sperm chromatin DNA damage via sperm oxi-
dative stress in mouse offspring, based on the evidence of
association between sperm H2O2 production and increased
fraction of chromatin defect. We believe that these results
make an important contribution to the issue of environmen-
tal pollution, and provide critical information for health-
risk assessments.
The authors thank Dr. Wei-Jen Yao from National Cheng
Keng University Medical Center for his technical assistance in the
serum hormone assay. They also thank Dr. Chee-Yin Chai and
Ms. Wan-Tzu Chen from Kaohsiung Medical University for shar-
ing their experience in histopathology.
REFERENCES
Aitken RJ, De Iuliis. 2010. On the possible origins of DNA dam-
age in human spermatozoa. Mol Hum Reprod 16:3–13.
Akutsu K, Takatori S, Nozawa S, Yoshiike M, Nakazawa H, Hay-
akawa K, Makino T, Iwamoto T. 2008. Polybrominated di-
phenyl ethers in human serum and sperm quality. Bull Environ
Contam Toxicol 80:345–350.
Antignac JP, Cariou R, Zalko D, Berrebi A, Cravedi JP, Maume
D, Marchand P, Monteau F, Riu A, Andre F, Le Bizec B. 2009.
Exposure assessment of French women and their newborn to
brominated flame retardants: Determination of tri- to deca- poly-
bromodiphenylethers (PBDE) in maternal adipose tissue, serum,
breast milk and cord serum. Environ Pollut 157:164–173.
Arumugam TV, Gleichmann M, Tang SC, Mattson MP. 2006.
Hormesis/preconditioning mechanisms, the nervous system
and aging. Ageing Res Rev 5:165–178.
Betts KS. 2008. New thinking on flame retardants. Environ Health
Perspect 116:A210–213.
Bi X, Thomas GO, Jones KC, Qu W, Sheng G, Martin FL, Fu J.
2007. Exposure of electronics dismantling workers to polybro-
minated diphenyl ethers, polychlorinated biphenyls, and orga-
nochlorine pesticides in south China. Environ Sci Technol
41:5647–5653.
Bradman A, Fenster L, Sjodin A, Jones RS, Patterson DG Jr,
Eskenazi B. 2007. Polybrominated diphenyl ether levels in the
blood of pregnant women living in an agricultural community
in California. Environ Health Perspect 115:71–74.
Bucci LR, Meistrich ML. 1987. Effects of busulfan on murine
spermatogenesis: Cytotoxicity, sterility, sperm abnormalities,
and dominant lethal mutations. Mutat Res 176:259–268.
Chao HR, Shy CG, Wang SL, Chen SC, Koh TW, Chen FA,
Chang-Chien GP, Tsou TC. 2010. Impact of non-occupational ex-
posure to polybrominated diphenyl ethers on menstruation char-
acteristics of reproductive-age females. Environ Int 36:728–735.
European Union. 2003. Directive 2003/11/EC of the European
Parliament and of the Council of 6 February 2003. Off J Eur
Union L42:45–46.
Evenson DP, Larson KL, Jost LK. 2002. Sperm chromatin struc-
ture assay: Its clinical use for detecting sperm DNA fragmenta-
tion in male infertility and comparisons with other techniques.
J Androl 23:25–43.
Fangstrom B, Athanasiadou M, Athanassiadis I, Bignert A, Grand-
jean P, Weihe P, Bergman A. 2005. Polybrominated diphenyl
ethers and traditional organochlorine pollutants in fulmars (Ful-marus glacialis) from the Faroe Islands. Chemosphere 60:836–
843.
Fisher MT, Nagarkatti M, Nagarkatti PS. 2005. Aryl hydro-
carbon receptor-dependent induction of loss of mitochon-
drial membrane potential in epididydimal spermatozoa by
2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD). Toxicol Lett 157:
99–107.
Hardy ML. 2002. A comparison of the properties of the major
commercial PBDPO/PBDE product to those of major PBB and
PCB products. Chemosphere 46:717–728.
Ikonomou MG, Rayne S, Fischer M, Fernandez MP, Cretney W.
2002. Occurrence and congener profiles of polybrominated di-
phenyl ethers (PBDEs) in environmental samples from coastal
British Columbia, Canada. Chemosphere 46:649–663.
Jakobsson K, Thuresson K, Rylander L, Sjodin A, Hagmar L,
Bergman A. 2002. Exposure to polybrominated diphenyl ethers
and tetrabromobisphenol A among computer technicians. Chemo-
sphere 46:709–716.
Ji LL, Gomez-Cabrera MC, Vina J. 2006. Exercise and hormesis:
Activation of cellular antioxidant signaling pathway. Ann N Y
Acad Sci 1067:425–435.
Kalantzi OI, Brown FR, Caleffi M, Goth-Goldstein R, Petreas M.
2009. Polybrominated diphenyl ethers and polychlorinated
biphenyls in human breast adipose samples from Brazil. Envi-
ron Int 35:113–117.
Kelly BC, Ikonomou MG, Blair JD, Gobas FA. 2008. Hydroxy-
lated and methoxylated polybrominated diphenyl ethers in a Ca-
nadian Arctic marine food web. Environ Sci Technol 42:7069–
7077.
Kodavanti PR, Coburn CG, Moser VC, MacPhail RC, Fenton SE,
Stoker TE, Rayner JL, Kannan K, Linda S, Birnbaum LS. 2010.
Developmental exposure to a commercial PBDE mixture. DE-
71: Neurobehavioral, hormonal, and reproductive effects. Toxi-
col Sci 116:297–312.
Krishnamurthy H, Danilovich N, Morales CR, Sairam MR. 2000.
Qualitative and quantitative decline in spermatogenesis of the
follicle-stimulating hormone receptor nnockout (FORKO)
mouse. Biol Reprod 62:1146–1159.
Kuriyama SN, Talsness CE, Grote K, Chahoud I. 2005. Develop-
mental exposure to low dose PBDE 99: Effects on male fertility
and neurobehavior in rat offspring. Environ Health Perspect
113:149–154.
Lam JC, Kajiwara N, Ramu K, Tanabe S, Lam PK. 2007. Assess-
ment of polybrominated diphenyl ethers in eggs of waterbirds
from South China. Environ Pollut 148:258–267.
Lilienthal H, Hack A, Roth-Harer A, Grande SW, Talsness CE.
2006. Effects of developmental exposure to 2, 20,4,40,5-pentab-
romodiphenyl ether (PBDE-99) on sex steroids, sexual develop-
ment, and sexually dimorphic behavior in rats. Environ Health
Perspect 114:194–201.
388 TSENG ET AL.
Environmental Toxicology DOI 10.1002/tox

Main KM, Kiviranta H, Virtanen HE, Sundqvist E, Tuomisto JT,
Tuomisto J, Vartiainen T, Skakkebæk NE, Toppari J. 2007.
Flame retardants in placenta and breast milk and cryptorchidism
in newborn boys. Environ Health Perspect 115:1519–1526.
Marchetti C, Obert G, Deffosez A, Formstecher P, Marchetti P.
2002. Study of mitochondrial membrane potential, reactive
oxygen species. DNA fragmentation and cell viability by flow
cytometry in human sperm. Hum Reprod 17:1257–1265.
Mazdai A, Dodder NG, Abernathy MP, Hites RA, Bigsby RM.
2003. Polybrominated diphenyl ethers in maternal and fetal
blood samples. Environ Health Perspect 111:1249–1252.
Morck A, Hakk H, Orn U, Klasson WE. 2003. Decabromodi-
phenyl ether in the rat: Absorption, distribution, metabolism,
and excretion. Drug Metab Dispos 31:900–907.
Muenhor D, Harrad S, Ali N, Covaci A. 2010. Brominated flame
retardants (BFRs) in air and dust from electronic waste storage
facilities in Thailand. Environ Int 36:690–698.
Oakberg EF. 1956. A description of spermiogenesis in the mouse
and its use in analysis of the cycle of the seminiferous epithe-
lium and germ cell renewal. Am J Anat 99:391–413.
Rhees RW, Kirk BA, Sephton S, Lephart ED. 1997. Effects of pre-
natal testosterone on sexual behavior, reproductive morphology
and LH secretion in the female rat. Dev Neurosci 19:430–437.
Schecter A, Johnson-Welch S, Tung KC, Harris TR, Papke O,
Rosen R. 2007. Polybrominated diphenyl ether (PBDE) levels
in livers of US human fetuses and newborns. J Toxicol Environ
Health A 70:1–6.
Shaw SD, Brenner D, Berger ML, Fang F, Hong CS, Addink R,
Hilker D. 2008. Bioaccumulation of polybrominated diphenyl
ethers in harbor seals from the northwest Atlantic. Chemo-
sphere 73:1773–1180.
Shiner M, Fuhrman B, Aviram M. 2006. A biphasic U-shape
effect of cellular oxidative stress on the macrophage anti-oxi-
dant paraoxonase 2 (PON2) enzymatic activity. Biochem Bio-
phys Res Commun 349:1094–1099.
Sjodin A, Carlsson H, Thuresson K, Sjolin S, Bergman A, Ostman
C. 2001. Flame retardants in indoor air at an electronics recy-
cling plant and at other work environments. Environ Sci Tech-
nol 35:448–454.
Staskal DF, Diliberto JJ, DeVito MJ, Birnbaum LS. 2005. Toxico-
kinetics of BDE 47 in female mice: Effect of dose, route of ex-
posure, and time. Toxicol Sci 83:215–223.
Streets SS, Henderson SA, Stoner AD, Carlson DL, Simcik MF,
Swackhamer DL. 2006. Partitioning and bioaccumulation of
PBDEs and PCBs in Lake Michigan. Environ Sci Technol
40:7263–7269.
Thuresson K, Bergman A, Jakobsson K. 2005. Occupational expo-
sure to commercial decabromodiphenyl ether in workers manu-
facturing or handling flame-retarded rubber. Environ Sci Tech-
nol 39:1980–1986.
Tseng LH, Li MH, Tsai SS, Lee CW, Pan MH, Yao WJ, Hsu PC.
2008. Developmental exposure to decabromodiphenyl ether
(PBDE 209): Effects on thyroid hormone and hepatic enzyme
activity in male mouse offspring. Chemosphere 70:640–647.
Verreault J, Gabrielsen GW, Chu S, Muir DC, Andersen M, Ham-
aed A, Letcher RJ. 2005. Flame retardants and methoxylated
and hydroxylated polybrominated diphenyl ethers in two Nor-
wegian Arctic top predators: Glaucous gulls and polar bears.
Environ Sci Technol 39:6021–6028.
World Health Organization (WHO). 1994. Brominated Diphenyl
Ethers. Environmental Health Criteria 162. Geneva: WHO.
Wyrobek AJ, Bruce WR. 1975. Chemical induction of sperm
abnormalities in mice. Proc Natl Acad Sci USA 72:4425–4429.
Wyrobek AJ, Heddle JA, Bruce WR. 1975. Chromosomal abnor-
malities and the morphology of mouse sperm heads. Can J
Genet Cytol 17:675–681.
Zoeller RT. 2005. Environmental chemicals as thyroid hormone
analogues: New studies indicate that thyroid hormone receptors
are targets of industrial chemicals? Mol Cell Endocrinol
242:10–15.
389DEVELOPMENTAL EXPOSURE TO DECABROMINATED DIPHENYL ETHER (BDE-209)
Environmental Toxicology DOI 10.1002/tox




















