
Development of a heat treatment to enhance the antimicrobial ...
169
Development of a heat treatment to enhance the antimicrobial properties of wood based mulches and animal bedding materials By Mina Kalantarzadeh UNIVERSITY OF SURREY SUBMITTED FOR THE DEGREE OF DOCTOR OF PHILOSOPHY NATURAL PRODUCTS RESEARCH GROUP DEPARTMENT OF CHEMISTRY FACULTY OF ENGINEERING AND PHYSICAL SCIENCES UNIVERSITY OF SURERRY December 2013 © M. K
-
Upload
khangminh22 -
Category
Documents
-
view
4 -
download
0
Transcript of Development of a heat treatment to enhance the antimicrobial ...

Development of a heat treatment to enhance the antimicrobial properties of wood based mulches and
animal bedding materials
By
Mina Kalantarzadeh
UNIVERSITY OF
SURREY
SUBMITTED FOR THE DEGREEOF
DOCTOR OF PHILOSOPHY
N A T U R A L PR O D U C T S R ESE A R C H G R O U P
D E PA R T M E N T O F C H E M IST R Y
FA C U L T Y O F E N G IN E E R IN G A N D PH Y SIC A L SC IE N C E S
U N IV E R SIT Y O F SU R E R R Y
D ecem ber 2013
© M. K

ProQuest Number: 27598828
All rights reserved
INFORMATION TO ALL USERS The quality of this reproduction is dependent upon the quality of the copy submitted.
In the unlikely event that the author did not send a com p le te manuscript and there are missing pages, these will be noted. Also, if material had to be removed,
a note will indicate the deletion.
uestProQuest 27598828
Published by ProQuest LLO (2019). Copyright of the Dissertation is held by the Author.
All rights reserved.This work is protected against unauthorized copying under Title 17, United States C ode
Microform Edition © ProQuest LLO.
ProQuest LLO.789 East Eisenhower Parkway
P.Q. Box 1346 Ann Arbor, Ml 48106- 1346

Acknowledgements
I would like to express my sincere gratitude to my supervisor. Professor Dulcie Mulholland,
for their support, time and guidance through my PhD. She provided me the opportunity o f
working in Natural Product Group which I will forever appreciate. I would not be able to
continue with my PhD project if she would not stand by me all along the changes and
difficulties during my study at University o f Surrey. I thank you Professor M ulholland for
your generous and loving heart.
I also would like to particularly thank Dr Joan Webber, the project manager for
Phythophthora control programme in Forest Research for their continuous support during the
work on P. ramorum. I appreciate the time and effort she put in supervising me with drafting
my thesis. I thank you Dr W ebber for your understanding and consideration. I should also
thank my former and co-supervisors Dr Frans de Leij and Professor La Ragione for the time
that they lent to me.
I also wish to thank Dr Moses Langat and Dr Dan Driscoll at Chemistry Department,
University o f Surrey, for their help and supervision with NM R spectroscopy and GC-MS
analyses o f the wood materials that are the core to this study. Appreciation also goes to the
members o f Pathology Department at Forest Research, who were all kind supportive and
welcoming particularly M r Andy Jeeves, for their laboratory technical support and M r Tony
Reeves for their kind and priceless support with the field trials in Cornwall. I would like to
thank the head o f Land Regeneration and Urban Greenspace at Forest Research, M r Tony
Hutchings and Ms Vicki Lawrence for their help and support.
The journey through this PhD life has enabled me to meet with people from all parts o f the
world. Many o f whom I have had a chance to make friends with. I would like to thank the
following people for their support and encouragement during the past three years, W atcharee
W aratchareeyakul, Katerina Ridge and Dorota Nawrot. I always will remember you.
I would like to thank my family members, specially my daughter and my sisters for their
love, patience and consideration during this journey. I would like to thank Robert and his
family for their support.
Finally, I would like to acknowledge Forest Research Centre (Alice Holt Lodge) for funding
my research.
IV

Table of content
Declaration............................................................................................................................... ii
Dedication .........................................................................................................................iii
Acknowledgements........................... iv
Table of content................................................ v
List of figures.................................................................................................................. xi
List of tables............................................ xiii
List of abbreviations..............................................................................................................xv
Abstract...................................................................................................................................xvi
CHAPTER 1 : INTRODUCTION...........................................................................................1
1.1 Heat-treated woods and their applications......................................................................1
1.1.1 Characteristics of heated wood m aterials................................................................... 2
1.1.2 Antimicrobial chemicals................................................................................................2
1.1.3 Chemical characteristic of wood before and after heating........................................ 3
1.1.3.1 Wood extractives and their chemistry......................................................................3
1.1.3.2 Aliphatic compounds..................................................................................................4
1.1.3.3 Terpenes and terpenoids........................ 4
1.1.3.4 Phenolic extractives.................................................................................................... 5
1.1.3.5 Wood structural components...................... 6
1.1.4 Changes in thermally modified w ood................................................................... 8
1.1.4.1 Hydrolysis of wood structural components as a result o f heating...................... 8
1.1.4.2 Colour change............................................................................... 9
1.1.4.3 Formation of acid........................... 9
1.1.5 Novel applications for heat treated w ood................................................................. 10
1.2 Background to poultry production and its economic significance............................10
1.2 Heath risks associated with poultry .........................................................................11
1.2.1 Salmonellosis and public health..................................................................................11
1.2.1.1 Broiler production and associated risk factors of Salmonellosis .......... 12
(1) Broiler production .......................................................................................................... 12
(2) Major sources of Salmonella in broiler houses............................................................13
1.2.1.2 Significance of litter moisture on Salmonella survival......................................... 14
1.2.1.3 Control of Salmonella in the gastrointestinal tract of growing broilers............ 15

(1) Competitive exclusion......................................................................................................15
(2) Probiotics ......... 16
(3) V accines.............................................................................................................................16
(4) Biosecurity measures................................................................. 16
1.2.1.4 Litter control m easures.......................................................................................... 17
Moisture control .......................................................... 17
Litter pH control.......................................................................................... 17
Bedding m aterials............................................................................. .:...................................18
1.2.2 Ammonia emissions..................... 18
1.2.2.1 Animal welfare and human health....................................................... 18
1.2.2.2 Environmental impacts of ammonia emission from broiler houses...................19
1.2.2.3 Production of ammonia in broiler un its............... 20
1.2.2.4 Ammonia production control and litter management..........................................21
(1) Moisture control............................................................................................................... 21
(2) Litter amendments............................................................................................................22
(3) Characteristics of wood materials as animal bedding m aterials ......................... 23
1.2.2.5 Limitation in using bedding materials and alternative approaches....................24
1.3 Background to the impact and biology of Phytophthora ramorum.................... ..25
1.3.1 The causal agent of Sudden Oak Death: Phytophthora ramorum ........................26
1.3.2 Symptoms caused by Phytophthora ramorum................................ 27
1.3.3 Risk factors of Phytophthora ramorum....................................................................28
1.3.3.1 Distribution................................................................................................................ 28
1.3.3.2 Host factors.............................. 29
(1) Range of Plant hosts........................................................................................................ 29
(2) Role of foliar hosts on disease cycle and development.............................................. 30
1.3.3.3 Environmental factors............................................................................................ 31
1.3.4 D ispersal........................................................................................................................32
1.3.5 Survival.................................................................... 32
1.3.6 Disease current management......................................................................................33
1.3.6.1 Exclusion..................................................................... 33
1.3.6.2 Best management practice.......................................................................................34
1.3.6.3 Eradication.......................................................................................................... 34
VI

1.3.6.4 Heat treatment.................................................... 35
1.3.6.5 Chemical treatments ................... 35
1.3.6.6 Natural products.........................................................................................................36
1.4 Summary and research objectives...............................................................................37
CHAPTER 2: MATERIALS & METHODS..................................................... 38
2.1 Bacterial isolates and media ............................................... 38
2.1.1 Solid agar m edia.............................. 38
2.1.2 Liquid growth media.................................. ...38
2.1.3 Bacterial enumeration (cfu ml’^)................................................................................ 38
2.2 Thermal treatment technique.......................................................................................38
2.2.1 Wood Materials...................................................... 38
2.2.2 Wood treatment temperature.......................................................................................39
2.2.3 Wood treatment time.................................................................................................... 39
2.2.4 Wood species................ 39
2.3 CO2 production............................ 39
2.4 Production of ammonium ions.......................................................................................40
2.5 pH ....................................................................................................................................... 40
2.6 Chemical analysis techniques........................................................................................40
2.6.1.1 Wood extraction.........................................................................................................40
2.6.1.2 Water extraction.........................................................................................................41
2.6.3 Methanol extraction...................................................................................................41
2.7 GC-MS analysis o f heated and non-heated wood shavings ............................. 41
2.8 Isolation of active compounds derived from the methanol crude extract............... 41
2.8.1 Thin layer chromatography .............................................................................. 41
2.8.2 Column chromatography............................................................................................ 41
2.9 In vitro assays.................................................................................................................. 42
2.9.1 Direct contact assay .................................................... 42
2.9.2 Synergistic effect assay........................... ...42
2.9.3 pH activity assay...........................................................................................................43
2.9.4 Bioautographic T L C ............................................................................................. 44
2.10 Determination of the effect of heating of rhododendron and larch wood shaving on P. ramorum............................................... 44
Vll

2.10.1 Wood species of tested materials.............................................................................44
2.10.2 Preparation of crude extracts................................................................... 44
2.10.3 Test isolates................................................................................................................. 44
2.10.3.1 Inoculum preparation............... 45
2.10.4 vitro assays.............................................................................................................46
CHAPTER 3: EVALUATION OF HEAT-TREATED WOOD AS LITTER .............47
3.1 Introduction........................................................ 47
3.2 Experimental.....................................................................................................................49
3.2.1 Wood inoculation..........................................................................................................49
3.2.2 Litter model preparation............................................................................................. 49
3.3 Results........................................................... 50
3.3.1 Effect of heating regime on the anti-microbial properties of pine bark............... 50
3.3.2. Effect o f heated pine shavings on microbial breakdown in modelled litte r 51
3.3.3 Effect of heated pine wood on CO2 production from wood shavings based litter (as indicator of microbial activity)............................................................................................ 52
3.3.4 Effect of heating duration at 120°C on CO2 production, pH and ammonia concentration ................. 53
3.3.5 Spectrum o f enhanced antimicrobial activity of heated pine shavings................ 56
3.3.6 Evaluation of the antimicrobial activity o f different wood types after heating ..57
3.3.7 Effect o f time of exposure to heat treatment of 120°C on pH of pine wood shavings 57
3.4 Discussion................................................................................................................... ...58
CHAPTER 4; CHEMICAL ANALYSIS AND EVALUATION OF ENHANCED ANTIMICROBIAL ACTIVITY OF HEAT-TREATED WOOD.................................................................... 62
4.1 Introduction.......................................................................................................................62
4.2 Experimental.....................................................................................................................64
4.2.1 Extract assays......................... 64
4.2.2. Chemical analysis........................................................................................................65
4.3 Results................................................................................................................................65
4.3 .1 Extract assays of the methanol crude extract........................ 65
4.3.2 GCMS analysis of crude extracts of p in e .................................................................66
4.3.3 Active fractions from column chromatography ................... 67
4.3.4 Yield of different molecules o f interest from heated Scots pine wood shavings 68
4.3.5 Synergistic effect o f dehydroabietic acid on the antimicrobial activity of vanillin69
v i i i

4.3.6 Effect of low pH on antimicrobial activity o f vanillin using citric acid ..............70
4.3.7 Antimicrobial activity of commercial vanillin and D H A A ...................................70
4.3.8 Effect of pH and storage temperature on antimicrobial activity o f commercial vanillin/DH AA..................................... 71
4.3.9 Spectrum of antimicrobial activity o f the methanol crude extract and isolated fractions of heated wood of pine............................................................... 72
4.4 Discussion.............................................................................. 72
CHAPTER 5: EFFECT OF HEATED WOOD ON P. RAMORUM ............................. 76
5.1 Reasons for concern.........................................................................................................76
5.2 Alternative approaches,to control............... 76
5.2 Experimental.....................................................................................................................79
5.2.1 Micro-cosm trails..........................................................................................................79
5.2.1.1 Effect of larch, pine and rhododendron heated woodchips on the survival of Phytophthora ramorum zoospores.......................................................................................79
5.2.1.2 Effect o f incorporating heated woodchips made from larch into soil infected with P. ramorum................... ...79
5.2.1.3 Effect of incorporated heated woodchips made from larch on the survival o f P. ramorum chlamydospores.......................... 80
5.2.2 In vitro assays........................................... 81
5.2.2.1 Assessment o f Minimum Inhibitory Concentration (MIC) of crude extracts against P. ramorum zoospores................................................................................................................ 81
5.2.2.2 Effect of crude extracts derived from heated and air-dried wood on chlamydospore germination of P. ramorum................................................................................................... 81
5.2.2.3 MIC of chemicals in crude extracts derived from heated pine wood against hyphae, zoospores and chlamydospores o f P. ramorum................................................. 82
5.2.3 Field studies............................................................... 82
5.2.3.1 Largin Wood field evaluation........................ 82
5.2.3.2 Druid’s Hill field evaluation....................................................................................83
5.2.4 Chemical analyses.........................................................................................................86
5.2.4.1 Chemical analysis of heated larch, pine and rhododendron wood.....................86
5.2.4.2 Chemical analysis of discoloured bark of a Japanese larch................................86
5.3 Results............................................................................................................................. 87
5.3.1 Microcosm studies........................................................................................................87
5.3.1.1 Evaluation of woodchips derived from larch, pine and rhododendron............. 87
IX

5.3.1.2 Effect o f incorporating heated woodchips into soil infested with zoospores ..88
5.3.1.3 Effect of incorporated heated larch woodchips on the survival of chlamydospores 88
5.3.2 In vitro assays............................ ...............................................................................89
5.3.2.1 Assessment o f minimum inhibitory concentration (MIC) of crude extracts against zoospores..................................................................................................................................89
5.3.2.2 Effect of crude extracts derived from heated and air-dried wood on chlamydospore germination............................................................................................................................. 89
5.3.2.3 Minimum Inhibitory Concentration (MIC) of different chemicals found in crude extracts derived from heated pine w ood...................... ...................................... ...............90
5.3.3 Field studies..................................................................................................................91
5.3.3.1 Largin Wood evaluation..........................................................................................91
5.3.3.2 Druid’s Hill field evaluation................................................................................... 92
5.3.4 Chemical analyses........................................................................................................ 93
5.3.4.1 Chemical analysis of heated larch, pine and rhododendron wood used in the field 93
5.3.4.2 Chemical analysis of discoloured bark of a Japanese larch............................... 93
5.4 Discussion...................... ....96
CHAPTER 6: DISCUSSION..............................................................................................103
Future w ork....................... 110
CHAPTER 7: REFERENCES.............................. ............................................................ 112
Appendices................................................................................................................... 128

List of figures
Figure 1.1 Examples of monoterepenes and their derivatives in softwood ................. 4
Figure 1.2 Examples o f resin acids ...............................................................................................5
Figure 1.3 Examples of phenolic extractives........................................................... 6
Figure 1.4 Arrangements of fibrils, microfibrils and cellulose in cell w a lls ...............................6
Figure 1.5 The most important hemicelluloses in softwoods; xylan and glucomannan............7
Figure 1.6 Structure of softwood lignin............................................................................................ 7
Figure 1.7 Examples of chromophores in wood according to Sundqvist, 2004.......................... 9
Figure 1.8 A broiler growing house................................................................................................. 13
Figure 1.9 Illustration of processes related to ammonia emissions from litter based manure 21
Figure 1.10 Symptoms of sudden oak disease in woodland and forest...................................... 26
Figure 1.11 Sporulation of Phytophthora ramorum...................................................... 27
Figure 1.12 The symptoms of infection by Phytophthora ramorum ..........................................28
Figure 1.13 The disease cycle of P. ramorum ............................................................................... 31
Figure 2.1 The methanol extracts of heat-treated (left) and air-dried (right) wood shavings of pine......................................................................................................................................................... 45
Figure 3.1 Litter models made in 2.5 liter plastic containers...................... 50
Figure 3.2 Change of pH in modelled litter over time.............................. 52
Figure 3.3 Comparison o f pH between 8 different model litters over time (n=4).................... 55
Figure 3.4 Changing pH values of pine shavings after different times of heat treatment........58
Figure 4.1 A comparison between two GC-MS chromatograms of heated and air dried pine wood.......................................................................................................................................................67
Figure 4.2 Compounds isolated from Pinus sylvestris after heat treatment at 120°C..............69
Figure 5.1 Dead larch trees infected by Phytophthora ramorum ............................................... 76
Figure 5.2 Larch trees infected by Phytophthora ramorum, Crymant, near N eath ................. 77
Figure 5.3 Field site in Largin Wood....................................................................... 84
Figure 5.4 Example of trays with wood chips placed underneath a rhododendron bush at Druid’s Hill, Cornwall............... 85
Figure 5.5 Bark (phloem) of a Japanese larch tree showing different stages of discolouration. ; 86
Figure 5.6 Examples of an assay plate (96-well plate) after the chemically treated zoospores are plated on SMA (a), and (b) an enlarged picture of normal growth of P. ramorum in sterile water (control)..............................................'....................................................................... 90
Figure 5.7 Comparison of mass spectra of methanol crude extracts of (a) heated pine (b) larch and (c) rhododendron..........................................................................................................;..... 94
Figure 5.8 Comparison o f mass spectra of crude extract o f discoloured bark from Japanese larch........................................................................................................................................................ 95
XI

Figure 6.1 Interactions of secondary metabolites with biomembrane and membrane proteins. 108
XU

List of tables
Table 1.1 Examples of aliphatic extractives in xylem and bark.....................................................4
Table 3.1 Effect of different heating temperatures on the survival o f Salmonella Enteritidis in ground pine bark ......................................................................................................................50
Table 3.2 Effect of source materials on the survival of Salmonella Enteritidis in pine shavings before and after heat treatment..................................................... ;................................... 51
Table 3.3 Effect of different heating temperature on the survival of Salmonella Enteritidis o n , filter paper........................................................................................................................ 51
Table 3.4 Effect of heating at 150°C and 200°C on the pH of pine shavings made into litter. ........................................................................................................................ 52
Table 3.5 Effect of heating at 150°C and 200°C for one hour on the microbial activity expressed as CO2 production in a pine shavings base litter...........................................................53
Table 3.6 Effect of heating pine shavings at 120°C for increasing periods of time (1 to 72 hours) on antimicrobial activity expressed as CO2 production..................................................... 54
Table 3.7 Effect o f heating pine shavings at 120°C for increasing periods of time (1 to 72 hours) on antimicrobial activity of the treated shavings expressed as the pH .............. 54
Table 3.8 Effect o f heating pine shavings at 120°C for increasing periods of time (1 to 72 hours) on antimicrobial activity of the treated shavings expressed as ammonium ion concentration .................................................................................... 56
Table 3.9 Survivals of S. aureus, and E. coli and on heat treated and air-dried wood o f pine and air dried and heated filter paper. ..............................................................................................56
Table 3.10 Recovery o f Salmonella Enteritidis from inoculated heated and air-dried wood shavings derived from 8 different wood species............................................................................. 57
Table 4.1 Recovery of SE from inoculated filter paper treated with extracts from heated and air-dried pine shavings.........................................................................................................................65
Table 4.2 Recovery of Salmonella (log cfu g' ̂± SE) from inoculated, heated and air-dried pine shavings..................................................... 66
Table 4.3 Yield of different compounds isolated from heated wood o f P. sylvestris...............68
Table 4.4 A comparison of antimicrobial activity of vanillin and combinations of vanillin and dehydroabietic acid on the survival O f SE.............................................................. 70
Table 4.5 Effect of citric acid on the antimicrobial activity of vanillin against Salmonella Enteritidis...............................................................................................................................................71
Table 4.6 Antimicrobial activity of commercial vanillin alone and in combination with dehydroabietic acid on the survival of Salmonella Enteritidis......................................................71
Table 4.7 Effect of pH and temperature on antimicrobial activity of on vanillin/ DHAA solution...................................................................................................................................................72
Table 5.1 Survival of P. ramorum zoospores inoculated on air-dried and heated woodchips. ............................................................................................................................................. 87
Table 5.2 Effect of incorporation rate of heated larch woodchips into soil on the survival oftwo isolates of P. ramorum, 3 days and 8 weeks after inoculation (n=3).....................................87
X lll

Table 5.3 Effect of heated woodchips of larch on the germination o f leaf disks of rhododendron....................................... 88
Table 5.4 Minimum inhibitory concentration o f the methanol crude extract (mg ml'^) of air- dried and heated wood of pine, larch and rhododendron (n=3).............................. ....89
Table 5.5 Effect of methanol crude extracts of air-dried and heated wood o f pine, larch and rhododendron on germination o f pre-colonized leaf disks o f rhododendron with chlamydospores of two isolates of P. ramorum.................................................................. 90
Table 5.6 Minimum inhibitory concentration (mg mf^) of the compounds derived from heated wood of pine on mycelia, zoospores and chlamydospores (in colonized leaf disks) of two isolates o f P. ramorum................................................................................................................. 90
Table 5.7 Minimum inhibitory concentrations of acetovanillone, coniferaldehyde and vanillin, with and without 20 mM dehydroabietic acid (DHAA) tested against zoospores of P. ramorum (n=3).............................................................................. 91
Table 5.8 Minimum inhibitory concentration o f acetovanillone, coniferaldeyde and vanillin with or without either 20mM of abietic, dehydroabietic or pimaric acid tested against zoospores of P. ramorum isolate B RCO l........................................................................................ 91
Table 5.9 Effect of heat treatments on survivals of P. ramorum on woodchips of larch, rhododendron and pine .......................................................................................................... 92
XIV
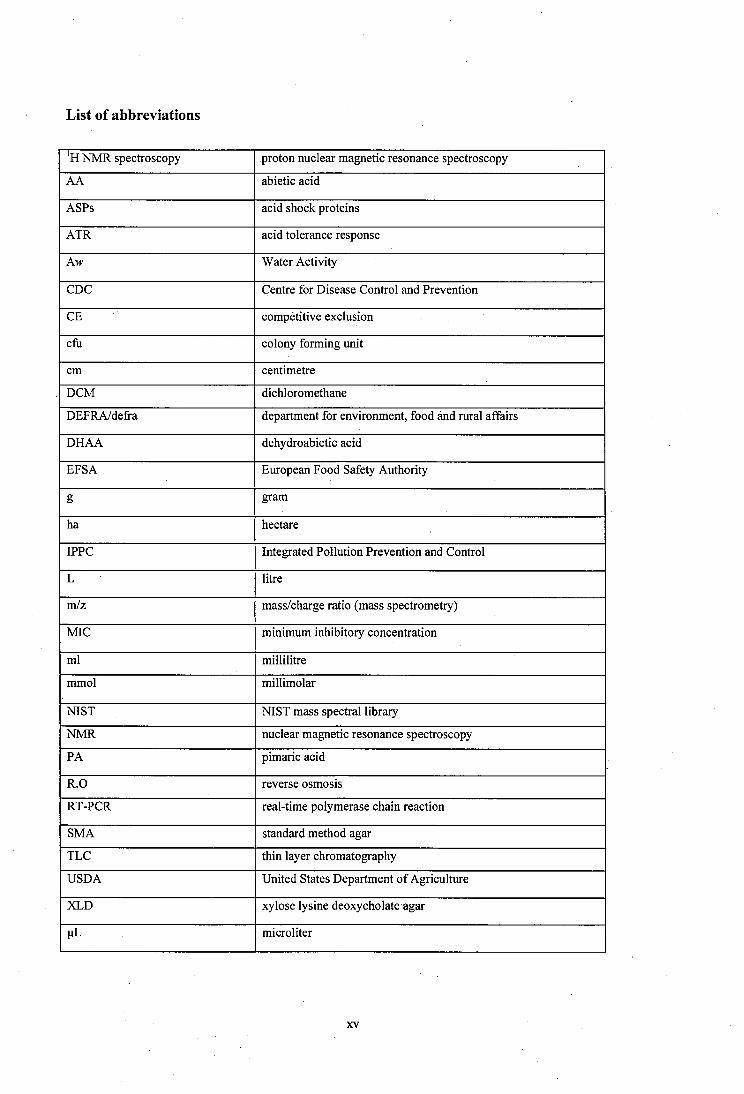
List of abbreviations
N M R spectroscopy proton nuclear magnetic resonance spectroscopy
AA abietic acid
ASPs acid shock proteins
ATR acid tolerance response
Aw Water Activity
CDC Centre for Disease Control and Prevention
CE compétitive exclusion
cfli colony forming unit
cm centimetre
DCM dichloromethane
DEFRA/defra department for environment, food and rural affairs
DHAA dehydroabietic acid
EFSA European Food Safety Authority
g gram
ha hectare
IPPC Integrated Pollution Prevention and Control
L litre
m/z mass/charge ratio (mass spectrometry)
MIC minimum inhibitory concentration
ml millilitre
mmol millimolar
NIST NIST mass spectral library
NM R nuclear magnetic resonance spectroscopy
PA pimaric acid
R .0 reverse osm osis
RT-PCR real-time polymerase chain reaction
SMA standard method agar
TLC thin layer chromatography
U SDA United States Department o f Agriculture
XLD xylose lysine deoxycholate agar
pL microliter
XV

Abstract
Heat treatment is the most commonly practiced technology to add value to products derived
from wood materials. Traditionally, heat treatment enhances the resistance of wood materials
to weathering and decay fungi, but despite its wide use in sectors such as construction, wood
preservation and food packaging, there has been little attempt to apply heat treatment in other
areas to manage food-home and plant diseases. A preliminary study indicated that pre
heating increased the inhibitory effect o f pine wood shavings against the important food-
home pathogen o f broilers, Salmonella Enteritidis (SE). Thus the first objective of this study
was to optimise a heating regime to evaluate the effect o f the enhanced inhibitory activity on
SE in direct contact and also reducing ammonia emission of a constructed litter using heated
wood. Heating to 120°C for 72 hours was the most effective treatment. In addition, heated
pine wood had antimicrobial activity against other known microorganisms in poultry bedding
including Staphylococcus aureus, Escherichia coli and Saccharomyces cerevisiae as well as
SE. Hardwood shavings of ash, beech, red oak also showed antimicrobial activity against SE
as did similarly treated softwood species such as pine and cypress. In vitro assays of
dominant chemicals derived from fractionation o f the extract demonstrated that bio-active
fractions containing vanillin and dehydroabietic acid exhibited antimicrobial activity, but
weak or no activity after purification. Overall, the in vitro assays suggested that a synergistic
interaction occurred between vanillin and dehydroabietic acid decreasing the minimum
inhibitor levels of vanillin. A second objective of the study was to investigate the potential of
heat-treated woodchips of Japanese larch and rhododendron to control Phytophthora
ramorum (PR) an invasive pathogen of larch in UK forests. Results demonstrated that heat-
treated pine, larch and rhododendron woodchips inhibited the recovery o f zoospores o f PR
compared with air-dried woodchips. This inhibition was maintained even if the larch
woodchips were diluted with soil. The in vitro assays revealed that their methanol cmde
extracts had an inhibitory effect on PR zoospores and also reduced germination of
chlamydospores compared with extracts from air-dried wood. Chemical analysis o f crude
extracts of all three showed some of the induced compounds were present in all the extracts
but differed in concentration. Coniferaldehyde was the most active compound against all
three propagules. The dominant resin acids, DHAA and abietic acid (AB) decreased the
minimum inhibitory level of the tested phenolic compounds against PR but had no effect
when tested alone. Results of a field trial using heat-treated and air-dried woodchips were
consistent with the bioassay results of crude extracts and indicated that heat treated materials
have potential to reduce the survival of PR under natural conditions. Finally, based on
XVI

overall results of bioassays in both parts of this study a mechanism of action for the
synergism between aromatic compounds such as vanillin and coniferaldehyde and resin acids
dehydroabietic and abietic acid was hypothesized.
XVll

CH APTER 1; INTRO DUCTIO N
1.1 Heat-treated woods and their applications
Wood and wood products are the most important source of renewable bio-mass and they have
a wide range of applications. The versatile nature of wood makes it possible for processing
industries to produce a vast number o f products with different applications. However, wood
materials are likely to become the habitat o f different microorganisms such as pathogenic
bacteria and moulds because of their porous structure. They can also be susceptible to decay
fungi. Wood materials are sanitised using different techniques such as washing, radiation
steam and dry heating. Dry heating techniques are widely used in industry as simple and
economic techniques which give heat-treated materials an advantage over raw material in
use. These heated wood materials have a wide range of different applications, in construction
and furniture, musical instruments, children playgrounds and food packaging.
In construction and furniture manufacture, heating is mainly used to improve wood
dimensional stability and its resistance to weathering conditions and decay fungi. Heating is
commonly used to improve hygiene of wood materials in contact with food (Beyer and
Gudbjomsdottir, 2002; Sjostrom, 1981). Wood packaging materials are required to be heat-
treated to prevent the spread of harmful microorganisms.
These improvements have been shown to be due to physical and chemical changes taking
place during heating and depend on two critical factors, temperature and time o f exposure
(Esteves et a l, 2008; Sundqvist, 2004). Lower temperatures improve hygiene conditions of
wood simply by making the wood a less favourable substrate for colonization by microbes.
According to the FAO, a core heat treatment of 56°C for a minimum of 30 minutes is
essential to pasteurize wood materials used in food packaging (FAO, 2002). Meat handling
pallets and vegetable boxes are heat-treated to clean the raw materials against mould before
use (Gudbjomsdottir et ah, 2000). Microwave techniques can also be used to sanitise
wooden pallets and egg trays and boxes (Gudbjomsdottir et a l, 2000). Examples include egg
trays heated at 90°C to pasteurise them and prevent transmission of poultry pathogens, such
as Salmonella, between sites (Gudbjomsdottir et a l, 2000). High temperatures of 100-115°C
are effective at reducing food pathogenic agents. Wooden kitchen utensils and cutting boards
are usually heat treated at the end of manufacturing (Beyer and Gudbjomsdottir, 2002). High
temperature treatments are also applied when a high degree of resistance toward target
microorganisms is required in products, such as outdoor fumiture and construction-aimed
materials. Temperatures above 200°C are usually known to be effective with these kinds of

applications (Esteves et a l, 2008; Kamdem et a l, 2002). When wood materials are heated at
very high temperatures they can be chemically modified and exhibit different characteristics
depending on the type of process applied (Esteves et a l, 2008; Esteves et a l, 2011).
1.1.1 Characteristics of heated wood materials
Heat treatment is one o f the wood industry’s main processes used to change the colour,
improve durability to biodégradation and weathering and to improve dimensional stability of
wood (Sundqvist, 2004). Among these, durability o f thermally modified wood against fungi
is of a great interest. Heating of wood seems to be an effective method to improve its
resistance against fungal attacks. It has been found that heating at 200°C, leads to wood
resistance against blue-stain, white-rot and brown-rot fungi (Kamdem et al, 2002; Savluchinske-
Feio et al, 2007). Some heat treatment techniques are also shown to have the potential of
replacing wood preservatives with high environmental impacts (Savluchinske-Feio et a l,
2007). Thermally modified woods show improved antimicrobial activities compared to
untreated sources. Previous researches indicate that the bioactivity is partly due to new
chemicals induced in wood materials after heating (Esteves et a l, 2008; Kamdem et a l,
2002; Sundqvist, 2004). In some cases, identified compounds are known for their potential in
a variety o f other antimicrobial applications and are produced from wood in large quantities
by heating techniques (MacKay et a l, 2009a; Murzin et a l, 2007).
1.1.2 Antimicrobial chemicals
There are literally hundreds o f chemical products derived from wood species which are used
in medicines, as food additives, in cosmetics, or act as preservatives, fungicides and
insecticides (Voda et a l, 2003; Wink, 2006). Chemical products o f wood are divided into
two general groups, wood extractives and chemicals derived from wood processing
techniques. Wood extractives are secondary metabolites which are extracted from raw
materials. Many of these show antimicrobial activity and are used widely as medicines, food
additives, spices, etc. Valuable chemicals derived from wood processing are also used as
food preservatives, many others are manufactured and are used as antimicrobial agents or
intermediate chemicals in the production of pharmaceuticals.
Wood processing chemicals are induced when wood undergoes different chemical reactions
upon different processing. Many o f these processes have been practiced for years and many
others have received substantial research and development efforts to introduce new products
or understand how to best obtain them. Depending on the type of heating process, wood

materials with different eharacteristics and chemicals are produced. These include high
energy processes such as pyrolysis and gasification that consume a lot o f energy in a range of
400°C or even higher. They are used to produce fuels or fuel intermediates such as Syngas
that is used to produce methanol, ammonia and bio-fuel. Some by-products of high energy
processing exhibit antimicrobial properties and are widely manufactured; such an example is
wood vinegar (MacKay et a l, 2009a; Williams, 2012). Wood vinegar is used in cosmetic
products such as shampoos and lotions and acts against bacteria and fungi that cause skin
damage (Williams, 2012).
Low energy processes like fermentation, extraction and torrefacation are applied to produce
different chemicals such as solvents and resins in large quantities. Some others are used to
produce bioactive chemicals with application in food flavourings, preservatives, pharmaceuticals
and their intermediate chemicals. Acetic acid is produced by either fermentation or dry
heating (different wood materials) and has application in the preparation o f pharmaceuticals
such as aspirin and some fungicides (MacKay et a l, 2009a). Phenolic aldehydes such as
syringaldéhyde and vanillin are pharmaceutical precursors (MacKay et a l, 2009a).
Syringaldéhyde is also used as hair and fibre dye. Vanillin is used as a preservative and
flavouring agent in food products such as cakes and ice creams. Others are used as smoke
flavours and meat preservatives, such as guaiacol and syringol. These chemicals are
produced as a result o f chemical modification in wood structural components (MacKay et a l,
2009a). Regardless o f chemical differences between wood species, desirable chemicals are
produced under the same heating regime.
1.1.3 Chemical characteristic of wood before and after heating
1.1.3.1 Wood extractives and their chemistry
Extractives are chemicals in wood that can be removed using solvents. They mainly consist
of fat, fatty acids, fatty alcohols, phenols, terpenes, steroids, resin acids, rosins, waxes and
other minor organic compounds. Tannins and inorganic salts and certain carbohydrates can
be extracted from wood with water. Most of the extractives exist in heartwood o f both
softwoods and hardwoods. However, softwoods have a higher content of extractives
(Sjostrom, 1981). Most of the extractives may not be essential for plant growth but they play
an important role in defending against wood decay fungi, insects and biological damage
(Céspedes et a l, 2006; Zwenger and Basu, 2008). Three major groups of wood chemicals
are aliphatic compounds, terpenes and terpenoids, and phenolic compounds.

1.1.3.2 Aliphatic compounds
Wood resin contains a large variety o f aliphatic compounds such as alkanes, fatty acids, fatty
alcohol, fats and waxes (Table 1.1). Fatty acids are the major components of parenchyma
resin in both hardwoods and softwoods. They are either saturated or unsaturated. Linoleic
acid and oleic acids are common representatives o f fatty acids. The most important esters are
fats (glycerol esters), usually occurring as triglycerides. Waxes are esters of other alcohols
which are usually aliphatic alcohols or o f terpenoid nature (Sjostrom, 1981).
Table 1.1 Examples o f aliphatic extractives in xylem and bark (Sjostrom, 1981)
Group Structure
n-AIkanes CHs—(CHa)»—CHî n - @-30Fatty alcohols C H ,-<C Fy,-C H ,O H n = 16-22Fatty adds CM r-fCH,),-CDOH n = 10-24Fats CH*-OR R, R', arid R"
(glycerol esters) 1 can be fattyCH— OR' acid residues
talkyl--C(>4CHf-OR* or hydrogen
(mono-, di-, and triglycerides)
Waxes RO—(CHjj),—GHj: R is fatty acid(estere of other alcohols) RO—-sterol
RO—terpene alcoholresidue
Subefln Characteristic; n = 18-28(polyestolides) 1— O — (CHj)n---CO — 1
1.1.3.3 Terpenes and terpenoidsThe constituents of both resin acids and volatile oils are o f a terpenoid nature. The volatile
oil o f conifers contains monoterpenes and their hydroxy derivatives (Figure 1.1). Pinene is
the most common monoterpene in softwood conifers. The resin acids present in the oleoresin
o f coniferous woods are derivatives o f tricyclic diterpenes (Figure 1.2).
OR
Figure 1.1 Examples o f monoterepenes and their derivatives in softwoodl,a-pinene; 2, P-pinene;3, 3-carene; 4, camphene; 5, bomeol; 6, limonene; a-terpineol; 8, dipentene (Sjostrom, 1981)

They are mainly found in heartwood and have a free carboxylic acid functional group.
Dehydroabietic acid is one example of a resin acid (Figure 1.2). Triterpenoids exist in
hardwood and closely related sterols occur in softwoods. Sterols also appear as the alcohol
component in waxes. Polyterpenes and their derivatives also exist in some trees and are
known as polyprenols. They contain various functional groups such as carbonyl, carboxyl,
hydroxyl and ester groups (Sjostrom, 1981).
COOH
C O OH
Figure 1.2 Examples o f resin acids.1, pimaric acid; 2, sandaracopimaric acid; 3, isopimaric acid; 4, abietic acid; 5, levopimaric acid; 6, palustric acid; 7, neoabietic acid; 8, dehydroabietic acid (Sjostrom, 1981)
1.1.3.4 Phenolic extractives
Heartwoods usually contains a large variety o f phenolic substances (Figure 1.3). They have
fungicidal properties and therefore protect the tree against microbiological attack. Phenolic
constituents of wood can be divided into five different groups; hydrolyzable tannins,
flavonoids, lignans, stilbene derivatives and tropolones. Although the phenolic substances
are concentrated in heartwood, only small amounts exist in xylem (Sjostrom, 1981).
OH
4
HO,.OH
OHOH
6OH
5
Figure 1.3 Exam ples o f phenolic extractives.1, gallic acid; 2, ellagic acid; 3, chrysin, 4, taxifolin; 5, catechin; 6, genistein (Sjostrom , 1981 ).
1.1.3.5 Wood structural components
Cellulose, hemicellulose and lignin are structural components o f wood and together they are
regarded as holocellulose. About 40-50% of the wood consists of cellulose which is a linear
polymer chain containing 7,000-15,000 monomeric units of y?-D-glucopyranose, linked by
glycosidic bonds. A group of cellulose chains held together by hydrogen bonding between
their hydroxyl groups is called a microfibril (Figure 1.4).
Cell wall
FibrilPlant cell
MicrofibrilH OH CHjOHH OH CHjOH H OH CH.OH
CHjOH H OH CH,OH H OH CH,OH
C ellulose
Figure 1.4 A irangem ents o f fibrils, micro fibrils and cellulose in cell walls
(Botany Resource Library, 1998)
Hemicellulose is a polysaccharide composed o f monomeric units that are highly branched
and held together by glycoside bonds (Figure 1.5). Each chain consists o f 100 to 400
monomeric units. Glucose and mannose or glucose and xylose are the sugar units o f the main

chain. High numbers of side branches make the hydroxyl groups easily accessible and
moisture in wood is commonly bonded to hemicellulose.
q A HO0 P
GlucomanttaftOH ■ OH
RO"
OH
PHIHO-
Figure 1.5 The most important hem icelluloses in softwoods; xylan and glucomannan (D u ttae /a /., 2012)
Lignin is a three-dimensional network polymer o f wood which consists o f three different
phenol units linked together (Figure 1.6). Lignin comprises 15-25% of the dry weight of
woody plants and is the most complex polymer o f wood because o f the random coupling of
different units. Lignin provides mechanical support to bind plant fibres together and is
known as wood glue.
Figure 1.6 Structure o f softwood lignin (Brunow et al., 1998)

1.1.4 Changes in therm ally modified wood
1.1.4.1 Hydrolysis of wood structu ra l components as a result of heating
The basic characteristics of thermally modified wood have been described by many
researchers. These changes are temperature related. According to Manninen et al., below
40°C, the changes that occur are mainly physical, such as reducing wood moisture and
emission of volatile components. Extractives tend to be more susceptible to heat as they
occur in resin canals and tracheid rays from which they can evaporate and they are less
bonded with structural wood components (Manninen et a l, 2002). As temperature increases,
the changes in chemical composition of wood take place. These changes are due to
degradation of both extractable and structural compounds. Results of many research studies
show that at temperatures above 100°C, all the original extractives have mostly disappeared
and as the temperature reaches above 130°C, structural components start to change and new
chemicals start to appear at certain temperatures. The new chemical properties may result
from changing the ratio between different structural components in the wood (Gonzalez-Pena
efa/.,2009).
Cellulose is the most embedded component in wood and therefore more resistant to
hydrolysis than hemicellulose and lignin (Sjostrom, 1981). However, crystalline cellulose
increases due to the degradation o f amorphous cellulose which is hardly affected by heat
below temperatures of 300°C. The chemical modification starts with the decomposition o f
hemicellulose which is bonded to the surfaces and between cellulose microfibrils when
temperatures reach 180-200°C. This is followed by deacetylation and finally
depolymerisation catalysed by the induced acetic acid. As a result, small organic compounds
with low molecular mass are induced. Lignin degradation starts at around 180-200°C and the
formation of aldehydes occurs because of the decrease in the content o f hydroxyl groups.
Homolytic cleavage o f y5-ether linkages in lignin is affected by heat treatment. Also
formation of condensed structures occurs as methoxyl content is reduced (Tjeerdsma and
Militz, 2005). Possible cross links between lignin and polysaccharides have been reported as
a result o f heating at relatively low temperatures (Gonzalez-Pena et a l, 2009; Niemz et a l,
2010). In addition, the condensation of induced low molecular compounds leads to the
formation of chromophores which are the cause of change in the colour of heated wood
(Beyer et a l, 2006; Sundqvist, 2004).

Pa/'fl-Quinones Stilbenes
■CHj
ort/io-Quinones phenols coniferaldehyde
Figure 1.7 Examples o f chromophores in w ood according to Sundqvist, 2004.(Ri to Re; H or continuing organic chains)
1.1.4.2 Colour change
Colour changes of the wood develop at around 70°C and not only depend on the temperature
but also on the wood species. Softwood becomes slightly darker while hardwood becomes
significantly darker when exposed to heat. The colour change is due to compounds produced
as a result of hydrolysis of carbohydrates (Fengel and Wegener, 1989) and their possible
reactions with other components in the wood. Research on colour change in wood shows that
resistance to biodégradation after heating is dependent on the degree o f change in colour,
which becomes darker with increasing temperature. It has been found that formation of
chromophores such as coniferaldehyde which is a lignin degradation product is a possible
cause of the change of colour in wood (Sundqvist, 2002, 2004).
1.1.4.3 Formation of acid
Wood is naturally acidic, with a pH o f 3 to 6. Extractives such as acidic phenols, low
molecular weight acids and fatty acids give wood its acidic properties. Upon heating of
wood, the formation of further acidic compounds occurs. Acetic acid and formic acid have
been found in emissions from the drying stage of radiata pine at 100°C (MacDonald et a i,
2002). Acetic acid is the predominant acid produced when the temperature is raised to
150°C. Acetic acid, itself, will further accelerate the hydrolysis rate o f wood polysaccharides
(Sundqvist, 2004). A change in quantity and type o f resin acid content of softwoods is
reported by several researchers. Unsaturated resin acids such as dehydroabietic acid are
shown to increase while more saturated forms such as pimaric, linoleic and abietic acids are
decreased (Micales et a i, 1994).

1.1.5 Novel applications for heat treated wood
The market for animal bedding materials from the UK total wood machine waste usage is
estimated to be approximately 400,000 tons of recycled materials, around 56% of the total
(Fletcher et a l, 2010). In general, softwoods are preferred as they do not support bacterial
growth as well as hardwoods and, for example, pine shavings are known to exhibit
antimicrobial activity against food-borne pathogens such as Salmonella and E. coli when
used as animal bedding materials. However, even when natural inhibitors are present in
wood, controlling microbial activity can still be difficult and highlights the need to
investigate how heat treatment could enhance the antimicrobial characteristics in wood-based
bedding materials or mulches.
For this reason the potential beneficial effects o f heat treated wood to be used in two very
different woodehip based systems are considered:
• Poultry rearing systems where food-borne pathogens, primarily Salmonella, must be
controlled as part o f an integrated poultry litter management.
• In woodehip mulches as part of a control strategy directed against the recently arrived
tree pathogen Phytophthora ramorum.
1.2 Background to poultry production and its economic significance
Poultry production is of substantial importance to worldwide economy and agriculture. The
increasing demand for poultry products has resulted in the expansion of animal production
systems in order to meet the protein needs of the human population. The annual poultry meat
production in the U.S., the world's largest poultry producer, totals over 43 billion pounds
(USDA, 2012a). In this 20 billion-dollar industry, circa 65% of the production consists of
broiler meat whilst turkey meat accounts for most of the remainder (USDA, 2012b). Every
year the UK poultry meat industry rears over 850 million chickens resulting in around 1.5
million tonnes of chicken meat. The average poultry meat consumption in the UK is 31kg
per person per year, with a total consumption o f circa 1.9 million tonnes (FSA, 2011). In
2010, the UK poultry production sector comprised 29% breeding and laying fowls, 63% table
chickens and 8% of other poultry including ducks, geese, and turkeys. The broiler share of
poultry meat production increased to a high of 1.32 thousand tonnes or 84% of total
production in 2010 with a value exceeding £2 billion. The versatility o f poultry meat,
particularly chicken, together with its perception as a healthy meat option, has increased the
importance o f poultry in western diets (Crane et a l, 2010).
10

1.2 Heath risks associated with poultry
Despite its economic benefits, intensive poultry production is associated with several major
health issues for humans, food animals and the environment.
1.2.1 Salmonellosis and public health
Bacteria belonging to the species Salmonella are the leading cause of intestinal diseases in
humans and animals worldwide (Castiglioni Tessari et a l, 2012). Estimated costs o f medical
expenses, sick leave and loss of productivity related to the high incidence of salmonellosis in
the US range from 1.3 to 4.0 billion US dollars a year (Taitt et a l, 2004). The annual report
on zoonoses and food-home outbreaks in the European Union reported that Salmonella
accounted for 99,020 reported human food poisoning cases in 2010. Salmonella was found
most often in chicken and turkey meat (EPSA, 2010a, b). In 2009, around 26% of
Salmonella infections reported in the UK were associated with consumption of poultry meat.
In the UK it is estimated that each year approximately one million people suffer from a food-
home disease, accounting for 20,000 hospitalization cases and 500 deaths. The health costs
are likely to be in the region of £1.5 billion (FSA, 2011). In developing countries.
Salmonella is the main cause o f food home disease outbreaks (Majowicz et a l, 2010). It
causes severe intestinal infection in humans accompanied by diarrhoea, fever and abdominal
cramps, symptoms may last for a week or more. In some instances, affected persons later
develop joint pains and arthritis. People of all ages are susceptible but a higher incidence is
reported for infants. Infection may spread to the blood stream, the bone marrow or the
meningeal linings of the brain in people with deficient immune systems causing severe or
fatal illnesses (Tauxe, 1996).
The causal agent
Salmonella was first discovered by Theobald Smith in 1885 and named in honour o f his
director, Daniel Elmer Salmon. Species of the genus Salmonella are characterised as rod
shaped, non-spore-forming, flagellated. Gram negative bacteria belong to the
Enterobacteriaceae family. There are more than 2,400 different serovars of the bacteria that
all are believed to cause illnesses in humans. According to the current system of
nomenclature adopted by Centres for Disease Control and Prevention (CDC), the genus
Salmonella comprises two species. Salmonella enterica and Salmonella bangori, which each
include several serotypes (Brenner et a l, 2000). Salmonella enterica is divided into six
11

different subspecies. The CDC system uses names for the serotypes of subspecies I, for
example serotypes Enteritidis and Typhimurium.
Salmonella Enteritidis (SE) is the most common serovar (43.5%) of Salmonella isolated from
human worldwide (Hendriksen et a l, 2011). Their ability to survive extreme conditions
combined with a wide distribution makes their control very challenging (Santos et a l, 2008;
White et a l, 1997). In the 1980s, the presence of SE in foodstuffs of poultry origin raised
concern about public health when several outbreaks of foodbome diseases occurred in
England. In the US, there were 189 outbreaks caused by SE with 6,604 patients and 43 deaths
between 1985 and 1989 (Castiglioni Tessari et a l, 2012). According to the European Food
Safety Authority, SE is one o f the most important serovars for public health in the European
Union (EFSA, 2010a). Salmonella Enteritidis is also an important pathogen for the layer
industry primarily because of its ability to infect hens and ultimately contaminate egg
contents (Holt, 1995).
1.2.1.1 Broiler production and associated risk factors of Salmonellosis
(1) Broiler production
The broilers sector o f the poultry industry rears chickens for meat. A typical broiler shed is a
large house housing 10,000-30,000 birds at one time. Chicks are transferred from a hatchery
seetion to the shed at one day old and are kept indoors until they reach the desired slaughter
weight which is around 6 weeks (in some cases birds may be reared outside i.e. free range).
Sheds are heated until ehicks are three weeks old and their body heat can keep them warm.
Food and water are provided. The house condition is monitored and temperature is
maintained by ventilation. The house floor is usually covered with a type o f bedding material
for the insulation from the cold and damp which becomes litter after the chick’s placement
(Figure 1.8). Wood shavings are the most common type o f bedding material but peanut
hulls, rice hulls, straw, shredded papers, and sand are also used. Different poultry production
systems use bedding materials with different depth. In intensive broiler production, 5-10 cm
o f bedding material is layered on the floor prior to placement of chicks. Under the EU
regulations, the entire litter is removed from a house after each flock and before new litter is
applied, the poultry shed is disinfeeted and cleaned thoroughly. In the USA litter from a
previous broiler floek is reused for multiple flocks (for up to one year). Broiler production is
the largest sector of agriculture in the UK. Although free range, bam and organic sectors
have seen growth since 1994, they still remain relatively small (Crane et a l, 2010).
12

Figure 1.8 A broiler grow ing house(http://w w w .bigdutchm an.de/en/poultry-production/hom e/pr-section/photos/poultry-grow ing.htm l)
(2) M ajor sources of Salmonella in broiler houses
Consumption o f meat o f poultry origin is known to be responsible for the major outbreaks of
enteric pathogens (EFSA, 2010a). Contamination o f poultry products may occur at any point
along the food chain from production (broiler houses, layers) to food processing
(slaughterhouses) and preparation (kitchen). Fresh broiler chicken meat is known to be a
major risk factor for SE infections (CCFH-Working-Group, 2007). Like other enteric
pathogens, SE can be disseminated to chickens through a variety o f sources in broiler houses.
Chicken’s gastrointestinal tract
O f the many potential sources reported for Salmonella contamination which include feed,
water, dust, insects, rodents, equipment, faeces and litter, the chicken gut is known to be a
main reservoir in poultry facilities. In conventional production systems, newly hatched
chicks in the clean and controlled environment o f a hatchery will be transferred to broiler
sheds without any contact with adult chickens. Therefore, development of gastrointestinal
microflora (Gl) is largely dependent on environmental sources. A balanced G 1-microflora is
essential to out-compete pathogens and protects the host against enteric pathogens. In the
case o f pathogen appearance in the environment, the sterile intestinal tract o f the newly
hatched chick represents an empty ecological niche for the pathogen to occupy. Salmonella
species can survive the acidic environment of chicken gut (with a pH as low as 3.3 or less) by
an adaption process called the acid tolerance response (ATR) (Foster and Hall, 1990). The
process requires the synthesis o f several acid shock proteins (ASPs) which induces pH
homeostasis and protein repair systems (Foster, 2001). Salmonellae multiply and colonize

the intestinal tract of chicks. Salmonella Enteritidis, like other serovars, causes only a
localized gastro enteric infection in birds (Cogan and Humphrey, 2003). Poultry may carry
different serovars of the pathogen and show slight or no symptoms. Salmonellae can survive
in the gastrointestinal tract of carcasses after slaughter, spreading the contamination into the
processing plant. This can lead to contamination of the whole slaughter line and has the
potential for contamination o f other foods and food preparation utensils. Furthermore, by
law, the detection o f the important serotypes of Salmonella including S. Enteritidis and S.
Typhimurium in early points during production and processing may result in destruction of
the whole flock or slaughter line in some countries. This leads to loss of financial returns for
the poultry producers. Therefore, primary control measures and strategies are established to
control or reduce the pathogen growth in the chickens gut. These include interventions that
aim to assist the development o f a balanced Gl-microflora in chickens growing under
conventional production systems.
Chicken manure
During the growth period, litter is a potential source of Salmonella contamination (Davies,
2005). Enteric pathogens o f the poultry gut are excreted into the litter with manure and
faeces. As chickens continue to live in contact with the litter, it will be seeded with higher
levels o f microbes in the chicken manure as they grow. Outside o f the chicken gut.
Salmonella survival and proliferation is dependent on the temperature and moisture o f the
surrounding environment. In intensive production with a large numbers of birds, litter has an
enormous potential o f becoming reservoirs o f Salmonella.
1.2.1.2 Significance of litter moisture on Salmonella survival
The effect o f high moisture levels in litter can have a dramatic effect on the growth and
spread of Salmonella within the house. Poorly managed litter also has the potential to become
the main source and route o f Salmonella transmission. It can spread the contamination by
sticking to the chicken bodies and carrying the contamination to the processing plants. The
feathers and skin, crop and cloaca o f birds are often highly contaminated (Kotula and Pandya,
1995). The results of extensive research by the United States Department of Agriculture
(USDA) shows that up to 75% of birds in Salmonella positive sites carry the pathogen on
their feathers and feet to processing plants while live birds were only infected at a rate of 1 0 -
15% (Marsh, 1999b). Transmission of Salmonella between (vertical) and among (horizontal)
the flocks is also facilitated by contaminated litter, particularly when litter is reused for
14

raising several flocks. It is reported that 4% of litter is consumed by boilers during grow-out
which is associated with a higher incidence o f Salmonella (Payne and Watkins, 2005). As
chickens contaminated with Salmonella (with the exception o f host restricted serovars) do not
show any symptoms, they can act as vectors, spreading the contamination between flocks
undetected. They further contaminate feed and water with Salmonella by direct contact.
Wet litter has also a direct effect on the microbial composition of litter (Dumas et a l, 2011;
Lovanh et al, 2007). Higher microbial diversity is linked with moisture content o f litter over
the length of the broiler house (Lovanh et a l, 2007). High diversity is believed to increase
the risk of emergence of antibiotic resistant strains of food-borne pathogens o f poultry
(Dumas et a l, 2011). Resistant non-pathogenic bacteria on farms can also act as a pool of
transferrable genes (DeFrancesco et a l, 2004) increasing the emergence o f resistant strains of
pathogens causing food-home diseases such as Salmonella (Angulo et a l, 2004).
The numerous potential sources o f Salmonella contamination in poultry systems increase the
risk of the pathogen carriage in livestock and contamination of food products. As a result, the
impacts of Salmonella incidence are added to public health. Furthermore, strict poultry
legislation of zero tolerance for Salmonella leads to flnancial loss for poultry producers and
growers (the destruction of entire Salmonella positive sites after inspection followed by costs
of substantial sanitation of the broilers house and removal and disposal o f contaminated
litter). As a result, reducing pathogen contamination on the farm seems to be receiving more
emphasis and is in need of interventions that effectively deerease further consequences of
primary contamination carried over to the food processing stage.
1.2.1.3 Control of Salmonella in the gastrointestinal tract of growing broilers
(1) Competitive exclusion
Control of Salmonella infection by competitive exclusion (CE) was first achieved by oral
administration of a suspension of gut content o f Salmonella-frQQ adult birds to 2-days-old
chicks (Mead and Barrow, 1990). Exclusion is considered to result from a range o f direct
effects such as the production volatile fatty acids, competition for colonization sites or
indireet effects such as the stimulation of the host immune system (Doyle and Erickson,
2012). CE treatment is prophylactic and is effective under controlled laboratory experiments
when used against 10 ̂cells/chick (Mead and Barrow, 1990). Under commercial conditions,
however, CE success has been variable due to the absence o f efficacy requirements such as
15

Salmonella-fvQQ chicks, good bio security and low-levels o f stress during the first few days of
treatment (Cox and Pavic, 2010; Goren et a i, 1988).
(2) Probiotics
Probiotics are live cultures of non-pathogenic bacteria that are mixed with bird feed or water.
Microorganisms that are used as animal feed probiotics are mainly bacterial strains o f several
genera such a.s Lactobacillus, Enterococcus, Pediococcus and Bacillus spp.(Gaggia et a l,
2010; Vandeplas et a l, 2010). These treatments attempt to manipulate the microbiota within
the gut, whieh can provide protection against colonization by Salmonella in several ways,
including production of antimicrobial substances such as volatile fatty acids, bacteriocins, or
hydrogen peroxide, reduction of the availability of sites for colonization, competition for
limited nutrients, and stimulation of the immune system (Foley et a l, 2011). Probiotics
provide protection against Salmonella, E. coli. Yersinia enterocolitica, and Campylobacter
jejuni (Schneitz, 2005). However, they have limited suecess in the control of Salmonella for
various reasons. As such, their applications are restricted to minimize the risk of emerging
virulent species and invasive serovars and their potential for antibiotic resistance (EFSA,
2 0 1 0 a).
(3) Vaccines
Another control method is use of a vaccine to reduce Salmonella in the gut as well as in the
reproductive organs. The use o f vaccines against poultry pathogens is increasing (Doyle and
Erickson, 2012). Due to public health problems associated with Salmonella Enteritidis and
Salmonella Typhimurium, these serovars are the target of most vaccinations (Foley et a l,
2011). Both live and killed vaccines are commercially available (Russell, 2012). However,
regardless of the type, they do not guarantee complete protection against the pathogens
(Chambers and Lu, 2002; Gast, 2007) but may decrease the infection burden.
(4) Biosecurity measures
Effective control o f Salmonella outside of chicken gut in the primary production level relies
on biosecurity measures to a great extent. Apart from complementary strategies such as
competitive exclusion and probiotics, restricting movements of birds, people and equipment
combined with substantial sanitation, reduces the risk of zoonotic pathogens. There are
different codes of practiee and standard procedures used in different countries highlight the
significanee of biosecurity (White et a l, 1997). Measures generally include pest control,'
preventative measurements for feed and water contamination, transport sanitation, equipment
16

maintenance and staff training to reduce the risk o f Salmonella contamination (FSIS, 2010).
Management of litter condition during the grow-out period is a major part of biosecurity of
broiler systems (The-Environment-Agency, 2013).
1.2.1.4 Litter control measures
Moisture control
Water activity (Aw) of the litter, which is the level of unbound water, affects Salmonella
growth and survival (Russell, 2012). Since Salmonella is only able to grow at water activities
above 0.94, keeping litter dry is necessary to prevent establishment and proliferation. Results
of a research study by Carr and colleagues showed low water activity values (<0.84) were
associated with Salmonella-nQgaA.i\Q flocks (Carr et a l, 1995). Similarly, it was found that
higher Salmonella and Escherichia coli counts were detected in litter samples with Aw
greater than 0.90 and moisture content greater than 35% (De Rezende et a l, 2001).
However, in intensive production, moisture control is a big challenge and is only possible by
implementing a few strategies such as a low salt and fat diet, effeetive management of
watering systems and using bedding materials with high moisture absorbent properties.
Litter pH control
Salmonella spp. grow naturally between pH 6.5 and 7.5 (Chung and Goepfert, 1970). So the
pH of chicken manure is favourable for Salmonella growth. Mineralization o f organic
nitrogen due to microbial activity in the poultry litter leads to ammonia production in the
litter. Ammonia dissolves in litter moisture and converts to ammonium ions in the litter.
This shifts litter pH toward alkaline which provides the optimum growth conditions for litter
mieroorganisms including Salmonella. Lowering the litter pH has been found to be effective
at inhibiting bacterial growth (Choi et a l, 2008; Moore et a l, 1996; Payne et a l, 2002).
Different types of litter amendments are used to reduce ammonia by reducing pH or, numbers
o f ammonia producing bacteria and direct chernical interactions (Terzich et a l, 1998).
Although, litter amendments or treatments have been shown to be generally effective on
bacteria levels, their use against Salmonella has not been as effective. Line found that
aluminium sulphate and sodium bisulphate reduced Campylobacter colonization frequency
and population in the ceca but not Salmonella colonization (Line, 2002). The greatest
reduction in Salmonella was found when the pH was 4 or less, and showed no significant
reduction at pH of 7 and 9 (Payne et a l, 2007). In a previous study it was found that
aluminium sulphate and sodium bisulphate significantly decreased Campylobacter
colonization in broiler chickens but had no effect on Salmonella numbers (Line and Bailey,
17

2006). Pope and Cherry found no significant effect on Salmonella populations in broiler
litter when using commereially available sodium bisulphate, even though a pH of 1.2 was
achieved (Pope and Cherry, 2000). In another study high levels of both granulated sulphuric
acid and sodium bisulphate (45.4 kg 92.9 m^), which were used in various application levels
on used litter, were needed to obtain low pH for significantly reducing Salmonella
concentrations (Payne et a l, 2007).
Bedding materials
Effect of bedding materials on composition and development o f gut microbiota illustrates
their important role in broiler health and performanee (Garrido et a l, 2004; Ivanov, 2001;
Line, 2002; Torok et a l, 2009). Results of a study by Torok et al. (2009) showed the choice
of litter materials significantly influence caecal microbiota composition and as a consequence
live weight and feed intake. They investigated the effect of bedding type on gut microbiota
composition and broiler performance. Seven types of bedding, softwood and hardwood
sawdust, softwood shavings, shredded paper, chopped straw, rice hulls and refused tea were
tested. Aktan and Sagdic also found that litter type affeets the composition and development
of the microbiota during bird placement (Aktan and Sagdic, 2004). However, others found
microbiota in the gut o f broiler chickens raised on wood shavings and straw developed
regardless of litter type with a slight difference in the first 4 weeks (Fries et a l, 2005),
although, total aerobic bacterial counts were found to be lower for wood shavings than for
straw. Additionally, studies in respect of the effeet o f litter treatment also shows that litter
acidifieation can alter gut and the intestinal flora of broiler chickens and lowers the frequency
of Campylobacter and C. perfrigens, Enterococcus spp. and Lactobacillus spp., (Garrido et
a l, 2004; Line, 2002). Results of these studies illustrate the significance o f chemical
characteristics of bedding materials and their inhibitory effect on microbial activity o f broiler
litter. This potential can be used to effectively control Salmonella during grow-out. The
inhibitory characteristics of bedding materials that could be used to advantage include their
ability to absorb moisture, to effectively control microbial activity o f litter and reduce the
assoeiated health impacts.
1.2.2 Ammonia emissions
1.2.2.1 Animal welfare and human health
A further health risk to human and bird welfare in broiler systems is ammonia emission from
manure. Health and welfare problems associated with high ammonia concentrations in
poultry houses include damage to the respiratory tract o f chickens, increased susceptibility to
18

Newcastle disease, incidence o f airsacculitis,, increased Mycoplasma gallisepticum, and
incidence o f keratoconjunctivitis (Moore et a l, 1995). Adequate ventilation is usually
allowed to exhaust ammonia into the atmosphere. Although ventilation will improve the air
quality of in-house environment, it does not reduce the amount of ammonia in the litter,
leaving birds vulnerable to the negative effect o f high pH on their bodies in contact with the
litter. Skin conditions o f poultry such as breast blisters, ulcers, hulk and foot pad bums are
caused by the high ammonia content of poultry litter. Ulcers can act as gateway for other
infections caused by E. coli and Clostridium perfringens, such as necrotic dermatitis and
scabby hip syndrome, which make broilers suffer from painful inflammations in joints and
under the skin. High ammonia concentrations (20 ppm and above) in poultry houses also
reduees animal growth, feed efficiency, and egg production (Nahm, 2005).
The effects of high ammonia concentrations in poultry facilities on human health are o f great
concern (Moore et a l, 1995). Upper respiratory ailments and eye irritations o f farm workers
are reported to be caused by exposure to concentrations above 25 ppm depending on the time
of exposure and serious health risks are involved at high concentrations of 50-110 ppm
ammonia (Donham, 1996; Ritz et a l, 2009). In addition, ammonia acts as a precursor in the
formation o f ammonium nitrate and ammonium sulphate in the atmosphere which are major
causes of premature mortality, asthma attack and chronic bronchitis (Ritz et a l, 2004;
Tasistro et a l, 2007). By law, the ammonia concentration in broiler houses should not
exeeed 50 ppm as it poses health risks to human operators of poultry (Miles et a l, 2004).
1.2.2.2 Environmental impacts of ammonia emission from broiler houses
Common strategies for reducing the ammonia concentration in poultry units generally aim to
improve the air quality o f the in-house environment rather than inhibiting the formation of
ammonia. Extracting the excessive level of ammonia into the atmosphere has become an
important environmental issue. It has been shown to play an important role in acid rain and
the dominant source of ammonia in Europe is livestock waste (Apsimon et a l, 1987; Moore
et a l, 1996). Ammonia is also a major cause of soil and surface-water acidification, nitrogen
enrichment and aerosol formation when litter is used as a fertilizer. With inereasing growth
of the poultry industry, environmental impacts of ammonia emissions and their effect on air
quality is likely to increase. Strict emission regulations are enforced by environmental
authorities known as Integrated Pollution Prevention and Control Directive (IPPC). In
practical terms, the implementation o f IPPC means that farms that have more than 40,000
birds must ( 1 ) minimise emissions of ammonia, (2 ) reduce associated odour from poultry

houses and (3) have a manure management plan in place. Also, IPPC rules are constantly
being tightened, forcing existing farmers to adjust in the future by implementing new
technologies enabling them to comply with tougher IPPC guidelines (The-Environment-
Agency, 2006). In July 2010, the European Committee proposal to amend the IPPC required
a reduction in the size threshold of poultry units from 40,000 to 30,000 laying hens, to 24,000
for ducks and 11,500 for turkeys. The directive became law in November 2010 (NFU-online,
2013). With profit margins already under pressure, to comply with environmental
regulations, reducing primary production o f ammonia in the litter is only possible by reducing
microbial activity within the litter which results in production of ammonia in broiler houses.
1.2.2.3 Production of ammonia in broiler units
A large and diverse population of bacteria, fungi, viruses and protozoa exists in the poultry
litter. Bacillus spp. and Arthrobactev spp. are dominant mineralizing bacteria (Kim and
Patterson, 2003; Ritz et a l, 2004) and some fungal species such as Aspergillus spp. also
mineralize organic forms of nitrogen into ammonia (Cook et a l, 2008). Mineralization of
organic nitrogen naturally takes place due to microbial activity in the poultry litter and leads
to ammonia production (Figure 1.9). Undigested proteins, uric acid and urea are the main
source o f organic nitrogen excreted as manure. Microbial degradation of uric acid in the litter
occurs in two steps; first urea is formed as an intermediate compound. The decomposition
process requires uric acid, water, and oxygen to react, releasing NH 3 and carbon dioxide
(CO2). The water-soluble characteristic of NH 3 allows it to be dissolved in the moisture,
protonating ammonia NH 3 into the non-volatile ammonium ion (NJU^.
NH 3+ H 2 O ± = ^ NH4"^+0 H‘
In broiler units, high stocking density decreases the rate of moisture release from the litter.
However, the respiratory output o f large numbers o f birds especially toward the end o f the
grow-out period increases the relative humidity and temperature of the in-house environment.
Litter absorbs excess water in the atmosphere. High moisture content in conjunction with
high temperatures in the broiler houses facilitates microbial breakdown o f the manure leading
to a higher content of ammonium ions in the litter. Ammonium ions will be converted into
ammonia (NH3), so the pH of the litter shifts toward alkaline and ammonia volatilization
increases. The inactive life style o f chickens (as a reflection of high stocking density) means
longer exposure o f their body to direct contact with litter. It is therefore necessary to control
2 0

the condition of the litter during the grow-out period. Moisture and pH of the litter are the
main important factors in controlling ammonia production in broiler units.
1.2.2.4 Ammonia production control and litter management
(1) Moisture control
Best management practice guidelines are issued to help farmers and producers to manage
litter efficiently by the Environmental Agency (The-Environment-Agency, 2013). According
to BMP (Environmental Management Plans) guideline for intensive farming, maintaining
moisture content o f 20-25% is recommended to avoid odour problems of wet litter. A
substantial depth o f litter o f at least 10-15 cm together with managing water leaks or spillage
due to faulty drinkers or leaks from walls and ceilings will control the moisture of the litter to
optimal level.
Free air stream
g.»
IN H g (aq) + H
Biomass
N H 4 ̂(adsorbed)
L it te r solidM oistu re
layer
Convection mass transfer
Partitioning betw een aqueous and gaseous phase ammonia
Chemistry o f ammonia in aqueous solution
Ammonia generation
Partitioning betw een solid and aqueous phase ammonia
Figure 1.9 Illustration o f processes related to am m onia em issions from litter based manure (Liu et a i , 2007)
Absorbent amendments are also sometimes recommended to reduce extra water in the litter,
although their use is limited by cost and their variable seasonal effects. Ventilation is
traditionally a common practice to lower ammonia concentration o f poultry units. Air
scrubbers and bio filters are other technologies that aim to clean air by exhaustion. However,
their practical applications are limited by cost and size restrains o f conventional poultry
houses (Ritz et a i, 2004). Reducing nitrogen content o f manure based on diet manipulation
such as reducing protein uptake and amino acid requirements also results in lower ammonia
production. However, as litter ages, moisture control becomes very challenging for the
21

poultry producers, particularly in re-used production systems. Furthermore, to maintain the
temperatures of the in-house environment during cold seasons, ventilation is reduced.
Therefore, litter amendments are commonly used to reduce ammonia volatilization by
lowering the pH of broiler litter.
(2) Litter amendments
Litter amendments are chemical treatments that are used to reduce ammonia emissions in
broiler houses. Different types of litter amendments are used to reduce ammonia by
reduction in litter pH, numbers o f ammonia producing bacteria and direct chemical
interactions (Terzich et a l, 1998). Currently, the most effective products have been found to
be chemicals that lower the pH of the litter. When litter pH is at levels o f less than 7.0,
ammonia production is insignificant. But at pH levels of greater than 7.0, ammonia
formation increases and when greater than 8 , it provides optimum growth condition for
Bacillus pasteurii, one of the primary uricolytic bacteria that facilitates NHg production
(Bacharach and Evans, 1957; Carr et a l, 1990; Reece et a l, 1979; Scheffer, 1965).
Typically, the pH of poultry manure as excreted and the pH of litter are between 7.5 and 8.5.
Lowering the litter pH has been found to be effective at inhibiting bacterial growth (Choi et al,
2008; Moore et a l, 1996; Payne et a l, 2002).
Choi et al, (2008) found using aluminium sulphate, aluminium chloride, and ferro sulphate
significantly reduced total aerobes and Gram negative bacteria in all three treatments. They
studied the effect of chemical treatments on pH and bacteria level o f litter treated with alum,
AICI3 and FeS0 4 at the same concentration rate ( 8 g 100 g litter) for 3 weeks. All treatments
reduced numbers of aerobic bacteria by 22 to 87% and Gram negatives by 63-99% of
untreated litter. Aluminium sulphate has been also shown to be able to either eliminate or
reduce aerobic bacteria levels in the litter (Line and Bailey, 2006). Others have reported that
acidic treatments reduce the survival and reproduction of the total populations o f different
microorganisms in the litter.(Choi et a l, 2008; Moore, 2003). However, litter amendments
are more affective at controlling pH when they are used above the manufacture’s
recommended rate (Wheeler et a l, 2008). This has increased concern about emerging
virulent strains of bacteria as a result of being exposed to high levels of chemicals (Payne and
Watkins, 2005). Furthermore, the effect of litter amendments lasts only 10-14 days while the
broilers are susceptible to ammonia until they become 21 days old (Wheeler et a l, 2008).
2 2

(3) Characteristics of wood materials as animal bedding materials
Physical parameters
Bedding materials are traditionally used to provide a dry and warm environment between
birds and to protect them from the cold floor. Different types of bedding materials such as
saw dust, rice hulls, straw, shredded papers and wood shavings are commonly available as a
by-products of other industries (Ritz et a l, 2009). The effect o f bedding materials on litter
condition and broiler performance has been extensively studied (Atencio et a l, 2010; Bilgili
et a l, 1999; Carlile, 1984; Hess et a l, 2010; Macklin et a l, 2005; Malone et a l, 1983;
Maitland, 1985; Oliveira et a l, 1974; Willis et a l, 1987). From these studies it can be
concluded that the ability of bedding materials to absorb moisture effectively reduces health
risks caused by high microbial activity of the litter such as ammonia emissions. Microbial
composition of poultry litter has been shown to vary due to the physical parameters of
bedding materials such as moisture content, pH and temperature (Aktan and Sagdic, 2004;
Lovanh et a l, 2007). However, a study by Miles (2011) comparing organic and inorganic
materials shows water absorbance is not a reliable indicator for good bedding materials. In
this study, NH 3 content o f pine, rice husk, sand and vermiculate in two different level of
moisture were compared. Results showed that at the original moisture content, sand and
vermiculate produced the most NH 3 (sand: 5.3 mg, vermiculate: 9.1 mg) and commercial
wood shavings and rice husk produced the least (wood shavings: 0.9 mg, rice husk: 0.26 mg)
while vermiculate was the more absorbent compared to wood materials (Miles et a l, 2011a).
This has drawn the attention of the research to chemical characteristics o f bedding materials
and its effect on microbial population of chicken litter.
Chemical characteristics
Several studies have found that the reduction of ammonia in the litter is linked with lower
microbial activity due to inhibiting effects of bedding materials rather than physical
parameters (Atapattu et a l, 2008; Duqueza, 1996; Lien et a l, 1998; Miles et a l, 2011b).
Duqueza (1996) suggests that lower ammonia emissions from wood shavings are due to
higher lignin content. Lien (1998) also found that using wood shavings reduced ammonia
compared with peanut hulls. In another study by Atapattu (2008), the NH 3 emission o f three
bedding types, refused tea, sawdust and paddy husk were studied. The lowest emission was
recorded from refused tea (13 mg kg'^ per hour) compared with sawdust (54 mg kg" ̂ per
hour) and paddy husk (44 mg kg’* per hour) after 42 days. The authors suggest that the
polyphenol content of refused tea lowers microbial activity and ammonia concentration.
23

These results are in disagreement with those of Duqueza because paddy husk, with higher
lignin content, had higher ammonia emission compared to refused tea (44 vs. 13 mg kg'^ per
hour). Nevertheless, wood materials are known as the most effective and commonly used
bedding materials because o f their good moisture absorption properties (Embury, 2004; Miles
a/., 2 0 1 1 b).
1.2.2.5 Limitation in using bedding materials and alternative approaches
The efficiency o f wood materials is supported by results o f many research studies on their
effects on reducing ammonia emissions and their major impacts on broiler health and
reduction in mortality and skin conditions of broilers (Ferrante et a l, 2006; Miles et a l,
2011b). However, wood materials are likely to become the habitat of different
microorganisms such as pathogenic bacteria and mould because of their porous structure. In
other industries, where hygiene of wood is critical, such as in food packaging, wood materials
are sanitised using different techniques such as washing, radiation, steam and dry heating
before use. In contrast, wood materials used to raise poultry are usually the unprocessed end
products of other industries such as the timber industry (Fletcher et a l, 2010). New bedding
materials are shown to be a potential source o f Salmonella at the time o f placement o f new
flocks and untreated materials also increase the risk of contamination with Salmonella during
storage (Fries et a l, 2005; Volkova et a l, 2009).
Although hygiene of bedding materials and their storage condition is suggested to influence
litter condition during grow-out, the effect o f potential treatments such as heating which can
be done to improve the quality of wood materials as animal bedding has not been
investigated. Research on efficacy or potential applications o f wood materials has been
focused on improving the life duration and resistance toward wood decay fungi. Therefore,
most of the existing developments are based on the need in the market for commercially
profitable products and their applications although, the market for animal bedding materials
in the UK, is the second biggest market for recycling the wood materials o f other industries.
Little has been done to develop heating techniques that improve the hygienic condition of
bedding materials before use (Fletcher et a l, 2010). Heating also has the potential of
introducing valuable levels of chemicals with antimicrobial properties, thereby reducing
zoonoses diseases of poultry as well as improving welfare of animals and humans and air
quality by reducing ammonia emission.
24

1.3 Background to the im pact and biology oi Phytophthora ramorum
Sudden Oak Death is a relatively new disease caused by the invasive plant pathogen
Phytophthora ramorum. In the mid 1990’s the disease was noticed killing tanoak trees
{Lithocarpus densijlorus) over a matter of weeks in some forests of California in the USA
(Rizzo et a l, 2002). Since then, a similar phenomenon has been observed to occur in
fourteen counties from Monterey County in the south to Humboldt County in the north
(Kliejunas, 2010). The mysterious causal agent was unknown until it was isolated and
identified as a species of Phytophthora. It then transpired that it was the same species, named
P. ramorum in 2001 (Werres et a l, 2001), that had been isolated from nursery plants of
rhododendron and viburnum in Germany and Netherland in the mid 1990s.
P. ramorum has since been discovered mainly on nursery stock in a number of European
countries including Germany, Netherlands, Belgium, the Czech Republic, France, Ireland,
Italy, Spain, Slovenia, Sweden, Denmark, Switzerland, Norway and Serbia. The first case of
P. ramorum in the UK was confirmed in a nursery in April 2002 on Viburnum tinus (Lane et
al, 2003). At the time of this discovery, the disease was causing damage to the forests and
wildland in the USA but it was only confirmed in a number o f nurseries in Europe. In 2001,
however, the disease was found on rhododendron in a Californian nursery, followed by the
first report of an infected tree outside the United States in November 2003 when P. ramorum
was isolated from a mature southern red oak tree {Quercus falcata) in the UK (Brasier et a l ,
2004; Goheen et a l, 2002). Since 2003, and despite quarantine efforts and emergency
regulations put in place in both Europe and the USA, the disease has spread fast and infected
a broad range of plant species (Frankel, 2008). In 2004, 176 nurseries were reported positive
after inspection in 21 states of the United States, apparently as a result o f interstate shipments
out of infested nurseries in California (Garbelotto and Rizzo, 2005; Jones, 2006). In Oregon,
approximately, 971 ha of the United State forest has been subject to eradication o f all infected
and nearby host plants between 2001 and 2008 (Kanaskie, 2010).
In 2007, several tree species with symptoms of bleeding bark cankers were identified in the
United Kingdom in Cornwall, mainly beech, but with some other species showing symptoms
of foliar infection and dieback, for example ash and holm oak (Sansford, 2007). In 2009,
Japanese larch was found with both stem and foliar symptoms in southwest England (Webber
et a l, 2010 ). In 2010, infection of larch became widespread with the disease spreading to
south Wales, where it has killed large numbers o f mature larch trees. (Figure 1.10)
25

a - ê i ^%» ■- -«Ÿ* ' ,
i ^ f K k mr f
3r} .;:>,'V4!SS Î ■s
Figure 1.10 Sym ptom s o f sudden oak disease in woodland and forest.Left, dead and dying tanoak in California. July 2010. Right, similarly afflicted larch forests south w est England, May 2010 (Source: w w w .nature.com /.../n7308/fig ).
1.3.1 The causal agent of Sudden O ak Death: Phytophthora ramorum
The pathogen is a member of the genus Phytophthora, and many o f its species pose a serious
threat to a range o f agricultural crops. Phythophthora belongs to the Oomyctes or water
moulds which are diploid eukaryotic microorganisms. Oomycetes were traditionally
classified as fungi but are now assigned to the Kingdom Chromista. Most known
Phytophthora species are soil-borne, root-infecting plant pathogens (Erwin and Ribeiro,
1966) but P. ramorum is an exception in that it is an air-borne species. It produces two types
of spores: chlamydospores and zoospores and the latter are contained in sporangia. Both are
vital for the survival and successful, spread o f the disease (Figure 5.2). Chlamydospores are
resistant to adverse environmental conditions and are known as resting spores, allowing the
pathogen to overwinter or persist during adverse environmental conditions. Sporangia are
asexual reproductive structures that contain zoospores and both are able to germinate and
infect plants directly. Zoospores have flagella enabling them to swim in water films
(Judelson and Blanco, 2005). Phytophthora ramorum is heterothallic. needing two mating
types, Al and A2, to produce oospores. Oospores are sexual spores which, in case o f P.
ramorum. have not yet been found in nature (Grunwald et al.. 2012)
26

Figure 1.11 Sporulation o f Phytophthora ramorum.Hyphae grow through lea f tissue and produce sporangia and chlam ydospores (Source: w w w .suddenoakdeath.org/w p-content/gallery/p-r)
Until 2012, three lineages o f P. irtmontm were known, termed EUl (European lineage 1),
NAl and NA2 (North American lineages 1&2) (Grunwald et al., 2012). The EUl lineage has
been found in European nurseries and forests and it has caused the current epidemic on
Japanese larch in the UK. There are also reports o f its recovery from US west coast nurseries
and waterways. The NAl is commonly isolated from infected forests in California and on
occasions from US nurseries. The NA2 is recovered mostly from US nurseries and is the
dominant lineage in western Canada (Grunwald et a l, 2012). Recently, a fourth genetically
distinct lineage has been identified, and designated as EU2 and is currently limited to
southwest Scotland and Northern Ireland (Poucke el a l, 2012).
1.3.2 Symptoms caused by Phytophthora ramorum
Infection by Phytophthora ramorum causes two different types o f disease: bleeding cankers
on the trunks and branches o f tree species and dieback and leaf blight of mainly shrubs and
ornamentals. Symptoms of the disease on tree species appear on the lower trunk in the form
of bleeding cankers (Brasier et a l, 2002) (Figure 1.12). When P. ramorum grows into the
bark it destroys the cambium and can eventually girdle the tree. At this stage, older leaves
start to become pale green and in a matter of two to three weeks they turn brown while
remaining attached to the tree. Symptoms on foliar hosts include necrotic spots appearing
along the leaf margins and leaf tips throughout the crown. Spores that are formed on the
infected leaf and shoot tissue (Figure 1.11) of foliar hosts but these can then also infect
leaves and bark of other plant species (Figure 1.12).
27

Figure 1.12 The sym ptom s o f infection by Phytophthora ramorumRhododendron leaves (left) and on the right, bark cracks with black exudates in coast live oak caused by P. ramorum. (Source: http://w w w .forestresearch.gov.uk/fr/IN FD -7B 5K U 2 )
1.3.3 Risk factors of Phytophthora ramorum
1.3.3.1 D istribution
The native origin o f P. ramoimm is unknown, but there is evidence that the pathogen has been
introduced separately into North America and Europe from another undiscovered area. The
distribution o f P. ramorum now includes the United States, where the disease was first seen
in 1995, Canada and several European countries (Brasier, 2003; Grunwald et a l, 2012; Ivors
et a i, 2004; Rizzo et al., 2005).
In the United States, the disease was first reported from forests in California and Oregon.
Since the first disease signs in 1995. and despite control efforts, from 2001 through 2008
approximately 971 ha o f forest has been burnt or cut to prevent further spread. The first
report o f P. ramorum from nurseries was made in 2003, when it was isolated from plant
containers o f rhododendron in Santa Cruse County. In 2004, the number o f positive nurseries
increased markedly due to interstate shipments o f affected plants. In 2009. the US
Department o f Agriculture, Animal and Plant Health Inspection Service reported 11 states
where P. ramorum had been detected.
In Canada. P. ramorum has been limited to nurseries since the first report from potted
rhododendrons in British Columbia in 2003. Eradication treatments have been successful to
control the spread o f f . ramorum and it is not present in the wider environment.
Since the first reports in Germany and Netherlands (Werres et al.. 2001). P. ramorum has
been discovered mainly on nursery stocks in a number o f European countries including
2 8

Germany, the Netherlands, Belgium, the Czech Republic, France, Ireland, Italy, Spain,
Slovenia, Sweden, Denmark, Switzerland, Norway and Serbia. However, in Belgium,
Denmark, France, Germany, Ireland, Luxembourg, the Netherlands, Slovenia, Spain and the
United Kingdom infected plants have been reported outside nurseries, in managed gardens
and woodlands (Sansford et a l , 2009).
In the UK, P. ramorum was first found on a mature oak tree {Quercus falcata) in Sussex in
2003 (Brasier et a l, 2004). It was the first confirmation of a diseased tree outside the United
States. About the same time, infection was also found on several Quercus rubra trees in the
Netherlands (Werres et a l, 2001). Since then, it has been found on a wide range o f plant
species in the UK. By 2007, limited numbers of tree species were found infected with
symptoms of stem canker but foliage and dieback symptoms were mainly reported on
rhododendron, viburnum and camellia. In 2009, Japanese larch trees were found with both
stem and foliage symptoms in Devon, Cornwall, and Summerset in southwest England (FC,
2012; Webber et a l, 2010 ). Until these findings, the majority of woodland infections in the
UK were associated with rhododendron which acts as the sporulating host, producing many
spores which then lead to infection of other species. Interestingly, in the absence of
rhododendron in these newly infected sites, needles of infected Japanese larch were found to
be producing large numbers o f spores, even more than had been previously found to be
produced on rhododendron leaves (FC, 2012). Therefore, felling infected larch trees in
forests was commenced alongside removal o f rhododendron to prevent the production of
spores and thus reduce the risk o f further infections and spread. By mid 2010, P. ramorum
had been confirmed on larch in both England and Wales (Brasier and Webber, 2010). Despite
taking emergency action in cutting diseased trees and putting biosecurity measures in place,
infection has been also found affecting larch in southwest Scotland and Northern Ireland.
1.3.3.2 Host factors
(1) Range of Plant hosts
Phytophthora ramorum infects a wide range o f plant species, including shrubs and
ornamentals, heathland plants and many tree species. To date, more than 150 plant species
are known to be susceptible to P. ramorum worldwide. In the United States, where the
disease was first discovered, the main host is known to be oak {Quercus) but other tree
species are also reported as hosts such as pacific yew, Douglas fir, grand fir and coastal
redwood. However, as a result of the significant damage to oak trees, the disease is well-
29

known as sudden oak death in the United States, where P. ramorum has killed large numbers
of North American native and tanoak trees (Frankel, 2008).
In the UK, more than forty plant species have been found to be susceptible to attack by P.
ramorum including red oak, beech, ash, yew, rhododendron, viburnum, magnolia and
heathers and bilberry (Brasier et a l, 2002; Denman et a l, 2005a; Denman et a l, 2005b;
Denman et a l, 2005c). However, the two native English oak species {Q. robur and Q.
petraea) have been shown to be more resistant and are less affected by P. ramorum than
American native oak species so the disease is known as “ramorum disease” rather than
sudden oak death in the UK. Until 2009, woodland infection was not widespread and most
infections occurred in nurseries. In 2010, infection of Japanese larch became widespread in
some part of the UK. Hybrid and European larch are also infected occasionally. Sitka spruce
is also reported to be infected under high levels of inoculum pressure (Chastagner et a l,
2012).
(2) Role of foliar hosts on disease cycle and development
Foliage hosts play an important role in the dispersal and epidemiology o f ramorum disease as
they are the main source of spores (Rizzo et a l, 2002; Turner, 2007). Tree hosts that are only
susceptible to trunk bleeding canker, such as oak, do not produce spores from infected tissue
(Davidson et a l, 2005a; Garbelotto and Rizzo, 2005). Studies on the disease cycle show that
P. ramorum is reliant on foliar hosts for reproduction and survival (Figure 1.13). California
bay laurel (Umbellularia california) was found to be the significant sporulating host for large
numbers of oak trees infected by P. ramorum in California forests (Kelly and Meentemeyer,
2002; Swiecki and Bernhardt, 2001). The mortality rate in oak trees in California forests was
shown to increase with increase in the number o f bay laurel associated with high levels o f
inoculum (Condeso and Meentemeyer, 2007). Cushman and Meentemeyer suggest there is a
lag effect between infections on bays and appearance of symptoms on oak trees (Cushman
and Meentemeyer, 2006). In Europe, Rhododendron ponticum has been found to be the key
sporulating host for the infection on susceptible tree species. In the UK, Rhododendron
ponticum and Larix kaempferi provide primary inoculum for most stem infections o f tree
hosts (Brasier and Jung, 2006; Brown et a l, 2006; Turner, 2007). Other foliar hosts such as
bilberry, holm oak and sweet chestnut are also suggested as significant sporuator hosts for
ramorum infection in northern Europe (Inman et a l, 2005; Sansford et a l, 2009).
30

A. Sporulation on infected lear es of foliar host
C. Dying crown o f a bole canker host
B. Twigs and leaf infection
D. Necrotic lesion in inner bark under bleeding canker in outer bark
£. Sporulation on fallen leas es
(D Primary- inoculum (sporangia) produce on (D Local intensification o f disease caused by ® The pathogen can persist in soil forinfected leaves can be carried \tia rain splash and air currents, infecting new hosts.
(D Secondary inoculum (sporangia or zoospores) is carried down stems by rainwater to infect lower portions o f the tree, infecting the inner bark and sapwood. resulting in a bleeding canker.
secondary inoculum which is produced in the canopy
® Infected leaves can act as a source o f inoculum even after they fall to the ground
® Sporangia produced on fallen leaves are carried to leaves and lower stem o f shrubs by rain splash
several months presumably as ÇjUainydpsEprçs.
F igu re 1.13 The disease cycle o f P. ramorum
(Parke and Lucas, 2008)
1.3.3.3 Environm ental factors
Temperature and moisture are the two main environmental factors that play important roles in
P. ramorum epidemiology. Phytophthora ramorum is regarded as a eool-temperature
organism which has an optimum growth temperature o f between 18-22°C (Werres et al.,
2001 ). The asexual spores, zoospores and chlamydospores, are produced under a broad range
of temperatures. Laboratory studies show that NA and EU lineage isolates produce sporangia
at temperatures between 10 to 30°C and 6 to 26°C respectively (Englander et a l. 2006).
Similarly, chlamydospore production o f U.S and European isolates is favoured by a
temperature range o f 10 to 28°C and 10 to 26°C respectively depending on the isolate.
However, abundant production o f sporangia and successful germination o f zoospores is
dependent on humidity. The maximum level o f zoospore germination is observed al 100%
humidity at a temperature range o f between 1()°C to 30°C. The presence o f free water in
susceptible plant parts, such as at leaf tips, enhances the infection by zoospores (Garbelotto et
6//., 2003).

1.3.4 Dispersal
Inoculum of Phytophthora ramorum spreads by different means after production on host
tissue. The spread can be by rain splash, in wind-driven rain, and in turbulent air locally
(Davidson et a l, 2002; Davidson et a l, 2005a; Fichtner et a l, 2009; Hansen et a l, 2008).
Long distance dispersal is mainly due to the movement of infected materials by local,
national and international trade of ornamental plants (Davidson and Shaw, 2003) and via
water courses (Davidson et a l, 2005a; Turner, 2007). Human activities also contribute to the
introduction of the pathogen into uninfected areas (Cushman and Meentemeyer, 2006).
Infected plant matter and soil can stick to vehicle tyres, shoes and tools and thus can spread
the disease over great distances (Cushman et a l, 2008; Tjosvold et a l, 2002a; Webber and
Rose, 2008). During 2006, dispersal of the pathogen up to a distance of 50 meters away from
an infected host was reported in the UK, by using water traps to track spores of P. ramorum
(Turner, 2007).
1.3.5 Survival
The potential o f disease caused by P. ramorum is dependent on its survival during
unfavourable conditions. The pathogen is able to survive dry and hot conditions in host
tissue materials. Like some other species of Phytophthora, chlamydospores are the resting
structures in the disease cycle and these are produced abundantly in the leaf tissue o f infected
foliar hosts (see Figure 1.11). Laboratory studies show chlamydospores and zoospores are
also able to survive at least for a month in moist conditions outside host tissue (Davidson et
a l, 2002). Bay laurel leaves are known to be an important niche for the pathogen during hot
summers in California (Rizzo, 2006). Survival in abscised leaves is also significant in the
disease cycle, with the survival in buried leaf litter shown to be much higher than in leaves on
the ground surface (8 8 % vs. 26-66%) (McLaughlin et a l, 2006). Other studies have reported
survival o f P. ramorum in rhododendron leaf tissue buried in potted nursery stock after 150
days (Shishkoff and Tooley, 2004). It was also recovered from potting media and sand over
approximately a year with no significant difference among treatments over time (Shishkoff,
2007). The pathogen can also survive in artificially infected host leaves buried or in the
ground. Turner et al, (2005) reported recovery o f P. ramorum for two winters from an
experiment conducted in the UK when it could be isolated from at least 50% of infected
leaves in all treatments. Survival of P. ramorum in woody materials is also reported. Shelly
et al. (2006) recovered P. ramorum from 1 out of 30 specimens o f tanoak and coast live oak
logs that had been air-dried for 6 months. P. ramorum has also been shown to survive in the
32

exposed wood of trees for up to two years (Brown and Brasier, 2007). Similarly, P. ramorum
survives the summer season in Rhododendron ponticum leaves in the UK.
Phytophthora ramorum \s occasionally isolated from soil during hot summers (Davidson et
a l, 2002; Davidson et a l, 2005a). Failure or decrease in P. ramorum survival in soil is
known to be associated with gradual soil drying during hot summer months (Davidson et a l,
2002; Fichtner et a l, 2007b). Phytophthora ramorum has a high potential to survive in
potting media. In an experiment where sporangia and chlamydospores where artificially
inoculated with the media, up to 6 and 1 2 months survival was observed respectively
(Linderman and Davis, 2006; Linderman et a l, 2006). Survival in potting media and sand is
reported to be longer at 4°C compared with survival at room temperature (Jeffers, 2005). In
the UK, laboratory studies on the effect o f temperature on chlamydospore germination
showed complete inhibition at extreme temperatures (-25°C and 40°C). In a study by Swain
et al. (2006) on the effect o f temperature during composting, no chalamydospores were
recovered after exposure to 55°C for an hour or 40°C for 24 hours with similar results
obtained in another study (Swain et a l, 2006; Turner and Jennings, 2005).
1.3.6 Disease current management
Current disease management involves two main principles: prevention and suppression.
Prevention includes actions taken before the pathogen is introduced such as import control or
inspection. Exclusion and eradication are the primary strategies that are aimed at preventing
the spread of inoculum or to control disease outbreaks.
1.3.6.1 Exclusion
Regulatory actions are implemented worldwide to reduce or eliminate the spread o f P.
ramorum. Different countries have restricted the movement of plants or plant products from
the United States. Sixty-eight countries are listed in the European Union Pest Risk
Assessment for which P. ramorum is part o f their legislation or considered in their Plant
Health Regulation (Sansford et a l, 2009).
The United States has enforced interstate and international plant quarantine measures. In
2001, the first quarantine measures were issued by Oregon State to prohibit the introduction
of P. ramorum from California where the disease was first noticed. Also, in 2002, the
European Union took emergency action by restricting the import of susceptible hosts into
member countries from the United States (2002/757/EC). For the movement of the main
known hosts in the EU (rhododendron, viburnum and camellia), plants must be certified by a
33

system called a Plant Passport which aims to verify that plant materials are inspected and free
o f infection (Frankel, 2008; Rizzo et a l, 2005). Under EU regulations, all member states
must undertake surveys to keep legislation continuously updated for P. ramorum. In the UK,
monitoring of P. ramorum was started after the first reported outbreaks in large managed
gardens in 2003. DEFRA commissioned a research programme for monitoring
contamination in infected areas following eradication actions. Seasonal pathogen survival,
pathogen persistence in water, litter, and soil and inoculum dispersal were all investigated to
inform the pathogen risk analyses in the UK. A range of methods for monitoring P. ramorum
are used which includes leaf baiting followed by traditional culturing methods and quatitative
polymerase chain reaction method (PCR) to detect low levels of inoculum. If infected plants
are found they have to be destroyed, and other phytosanitary measures are applied to prevent
further spread. For primary detection of Phytophthora species in the field, an ELISA kit
known as Lateral Flow Device (LFD) is developed by Forsite Diagnostics Ltd., York, UK.
The kit is easy to use and gives results in a few minutes. LFD kits are widely used for field
diagnosis of Phytophthora species and their results are usually comparable with PCR and
other laboratory techniques.
1.3.6.2 Best management practice
To reduce the adverse effect o f the disease, different measures, methods and practices have
been introduced through experience, extensive research and background knowledge o f the
disease cycle. This information has been distilled into guidelines for preventing or managing
of P. ramorum outbreaks in both woodlands and nurseries including measures to prevent the
spread during pruning, planting and soil handling in regulated areas. The guidelines may
vary between woodlands and nurseries, although applying a combination o f the suggested
measures is usually most effective. A best practice guide for woodland, parks and gardens is
developed and published by DEFRA (Defra, 2008).
1.3.6.3 Eradication
The current strategy to control and reduce the spread o f P. ramorum where it is already
present is aimed at lowering the inoculum levels in the environment by removing infected
sporulating hosts. Although, these efforts are likely to be successful if the disease is notified
shortly after it has been introduced, they can be expensive and costly. In the United States,
2331 hectares of Oregon State lands around the disease centre were subjected to Oregon and
APHIS quarantines (Kliejunas, 2010). The results showed a reduction in the number o f new
infested areas over a few years, reducing the damage to forests. Since 2002, FERA (Food
34

and Environment Research Agency) has launched a programme to clear rhododendron from
woodland and other sites to control ramorum in the UK. The process has been successful at
some sites (Turner, 2007), but not completely at others. In response to the outbreak o f P.
ramorum on Japanese larch, the Forestry Commission felled 600,000 infected larch trees in
2010 which were in private and public sites in south west o f England, compared with more
than 60 ha of rhododendrons cleared from woodlands in 2007 (Brasier and Webber, 2010).
After removal of rhododendron, herbicides are applied on the remaining stumps to prevent
resprouting. Similar actions are taken as emergency requirement against P. ramorum
throughout European Union countries (Aveskamp et a l, 2006; Defra, 2008).
1.3.6.4 Heat treatment
Heat treatment has been examined to assess its potential to reduce the pathogen levels in
infected plant materials. Composting has been shown to be effective at reducing P. ramorum
in green waste (Swain et a l, 2002; Swain et a l, 2006). A temperature of 55°C for 2 weeks
has been demonstrated to be sufficient for complete elimination of P. ramorum from compost
piles. However, the risk o f contamination remains if the composting process is not done
correctly. In addition, the prolonged heat treatment is not practical for some hosts. Steam
heat has been shown to be more effective than dry heat to decontaminate the plant containers
or infested media (Linderman and Davis, 2006). Heat treatment is also used to
decontaminate conifer bark before it can be moved from non-EU countries. An hour at 55°C
is also considered to be sufficient heat treatment to stop viable P. ramorum remaining in
cankered stems and woodchips of affected trees (Shelly et a l, 2006).
1.3.6.5 Chemical treatments
A variety of different fungicides used against other species o f Phytophthora have been tested
on P. ramorum. Many o f these are found to be effective in vitro, and include metalaxyl, Al-
fosetyl, copper sulphate, copper hydroxide and phosphonate (Garbelotto and Rizzo, 2001;
Hardy et a l, 2001). In the USA, the effect of fungicides has been evaluated using different
application methods such as trunk injections, soil drenches, foliar spray and topical
applications (Garbelotto et a l, 2003). Trunk injection of the active ingredient (phosphonate
compounds) was found to be the most effective method for controlling P. ramorum.
Although chemical treatments are found to be effective, the practicality o f their application is
limited to individual high-value trees rather than widescale application in woodlands and
forests. Foliar treatment o f oaks and tanoaks was tested using phosphonate but it was found
to only provide short-term protection accompanied by some phytotoxicity (Garbelotto, 2004;
35

Garbelotto et a l, 2007). Chemical treatment o f irrigation water and soil with compounds
such as hydrogen peroxide and sodium hypocholorite can be effective at decontaminating
water containing P. ramorum propagules (Sansford and Woodhall, 2007). Chemicals
containing copper are also toxic to propagules of P. ramorum (Colburn and Jeffers, 2010). A
variety of different chemicals are used as drenches to control P. ramorum in soil such as
metalaxyl-M, chloropicrin and dazomet (Yakabe and Macdonald, 2008) and can either reduce
the number of propagules or eliminate the pathogen (Turner et a l, 2006; Yakabe and
Macdonald, 2008). However, there is concern over widespread application of metalaxyl-M
as development of emerging resistant isolates of P. ramorum has been detected (Heungens et
a l, 2006; Turner et a l, 2008). There are also concerns that the use of metalaxyl-M and other
systematic fungicides may mask the symptoms of infection and make inspections invalid for
the detection of P. ramorum (Chastagner et a l, 2010; Shishkôff, 2005).
1.3.6.6 Natural products
Some natural products have been found to have potential inhibitory effects on the
germination and growth of zoospores of P. ramorum. Rice bam extract and grapefruit extract
have both been shown to inhibit the sporulation and growth of P. ramorum (Orlikowski,
2004; Widmer, 2008). A previous research demonstrated that the active compounds of
yellow cedar, hinokitol and thymoquinone have an inhibitory effect on P. ramorum zoospores
and potential to be used as spray on foliar hosts (Manter et a l, 2007). Another study carried
out by Nagel and colleagues (2011) showed resistant coast live oaks contained higher levels
of ellagic acid and tyrosol and total soluble phenolic compounds, all o f which have an
inhibitory effect in vitro on four species o f Phytophthora (Nagle et a l, 2011). Another study
by which investigated the chemical defence response of infected oaks showed there was an
association between higher concentrations of phenolic compounds and resistant species of
oak toward P. ramorum (Ockels et al., 2007). Manter et al, (2007) investigated antimicrobial
activity of heartwood extracts of seven western conifer trees against P. ramorum. The results
of this study showed incense cedar, western redcedar, yellow cedar, western juniper and Port-
Orford-cedar exhibited antimicrobial activity. From 13 compounds isolated from the
bioactive extracts, 6 compounds showed antimicrobial activity in bioassay tests. In a field
trial, redcedar woodchips which were placed under infected California bay laurel on the
forest floor for 4 months reduced the accumulation of P. ramorum DNA in the litter
compared with redwood woodchips that its extract showed no activity against P. ramorum.
The authors had previously suggested that antimicrobial woodchips o f yellow cedar can be
36

spread over trails or footpath to reduce the movement and redistribution of the spores from
these areas (Manter et al., 2006).
1.4 Summary and research objectives
A preliminary study on the effect of heat-treatment of pine wood shavings on the survival of
Salmonella showed promising results. The overarching aim of this project therefore was to
examine the extent to which primary heating techniques can enhance the antimicrobial
activity of wood making it more relevant for a particular reason.
The first part of the project focused on pine wood shavings used as animal bedding materials
and the specific objectives were to:
• Develop and optimise the conditions needed to enhance the antimicrobial activity in
pine wood shavings (Chapter 3)
• Evaluate the inhibitory effect on the microbial activity of chicken litter using
ammonia production as a measure o f microbial activity (Chapter 3)
• Undertake chemical analysis and identification o f the bioactive compounds in heated
wood materials (Chapter 4)
• Screen the bioactive fractions for inhibitory effects on Salmonella Enteritidis (Chapter 4)
The second part of the project extended the concept o f inhibitory activity o f heat treated
wood by testing the activity of larch and rhododendron woodchips against the sudden oak
death pathogen Phytophthora ramorum. Specific objectives were to:
• Evaluate the potential of heated woodchips of P. ramorum hosts (larch and
rhododendron) to inhibit P. ramorum growth in the laboratory (Chapter 5)
• Define the bioactive compounds within heat treated pine, larch and rhododendron
• Examine the potential for the application o f the heated materials in the field aS part of
a management strategy to control P. ramorum
37

CH APTER 2: M ATERIALS & M ETH O DS
2.1 Bacterial isolates and media
Salmonella Enteritidis ATCC 4931 was the model organism used in all experiments when
assessing antimicrobial activity o f heated and non-heated wood materials. Initially, SE was
subcultured on Nutrient Agar (25°C, 48 hours) from the original culture supplied by the
microbiology laboratory. Faculty o f Health and Medical Sciences, University o f Surrey.
After, developing the thermal regime, other microorganisms were also investigated including:
Staphylococcus aureus ATCC 25923, Esherichia coli ATCC 11775, Saccharomyces
cerevisiaeV^CC \9Q%.
2.1.1 Solid agar and liquid media
Fresh cultures of all bacteria strains were routinely cultured on Nutrient Agar for 48-36 hours
at their optimum growth temperature. An overnight liquid culture of the test strains was
routinely prepared in Nutrient Broth for bacteria strains. Malt Extract Agar and Malt extract
were used to subculture Saccharomyces. Retrieved Colony Forming Units (CFU) recovered
from wood were cultured on XLD agar (Xylose Lysine Deoxycholate agar; Oxoid) for
Salmonella, Nutrient Agar for E. coli and Staphylococcus and Malt Extract Agar for
Saccharomyces under aerobic conditions.
2.1.2 Inoculum preparation
Bacteria strains were cultured in Nutrient Broth and incubated for 16 hours at their optimum
growth temperature routinely while shaking at 2 0 0 rpms and were used as starting inoculum
for inoculating wood or filter paper samples (approximately 10’̂ ). For bioassay of the
chemicals, the inoculum size was routinely adjusted at 1 0 ̂ cfu mf^ by measuring the optical
density at 600 nm (ODeoo) using a spectrophotometer.
2.1.3 Bacterial enumeration (cfu ml' )̂
The retrieved numbers of Colony Forming Unites (CFU) from wood samples were
determined by culturing a fixed volume of the suspension (0.1 ml). % strength Ringer’s
solutions was routinely added to one gram of inoculated materials and a 1 0 -fold dilution
series of viable inoculum tested on XLD agar for viability.
2.2 Thermal treatment technique
2.2.1 Wood Materials
The wood materials were provided in the form of 80 cm logs by Forest Research, Famham,
England, and were made into wood shavings by the workshop in Faculty of Health and
38

Medical Sciences, University of Surrey. The materials of different plant species were
individually bagged and kept at room temperature of 25°C for 2 days before they were either
air-dried or heat-treated.
2.2.2 Wood treatment temperature
Pine bark and pine wood were initially ground into a course powder and wrapped in several
layers o f aluminium foil before heating. Wood materials were placed in a muffle furnace
exposed to a range of temperatures (20, 50, 100, 150°C) and heated to the required
temperature for at least one hour. The heated material was allowed to cool before use in
experiments.
2.2.3 Wood treatment time
After the determination o f the efficient temperature, the effect of different heating times o f 1,
24, 48, and 36 and 72 hours at this temperature on the induced inhibitory effect o f heat
treated wood in contact with Salmonella was then assessed.
2.2.4 Wood species
Other wood materials derived from eight different species were also assessed for their
antimicrobial activities after heating. Species tested included: eucalyptus (Eucalyptus
gunnii), beech {Fagus sylvaticd), birch (Betula pendula), ash {Fraxinus excelsior), sweet
chestnut {Castanea sativd), red oak (Quercus rubra), cypress (Cupressus sempervirens) and
spruce (Picea abies).
2.3 CO2 production
For measuring microbial respiration, CO2 evolution within the prepared litter treatments was
determined using an ADC.225.MK3 infra-red gas analyser (Analytical Development Co. Ltd,
Hoddesdon, England). To accurately measure microbial activity, before measuring, all the jar
lids were removed and the head space was air flushed using the air pump on the analyser.
Then to allow sufficient accumulation of CO2 in the jars, the lids were replaced and
tightened. They were left for 60 minutes before the head space of the jars was extracted
using a 60 ml syringe (via a sampling port fitted above the head space). The extracted gas
from each treatment was injected into the gas analyser. A sample of ambient air was used as
a control. Dilutions were made where necessary with N 2 to bring the concentration o f the
samples into line with the measurement range o f the analyser. After the measurement, the
litter treatments were air flushed before incubation. The following equation was used to
39

convert CO2 concentration in the samples to CO2 production per hour per gram of the
materials (h'^ g'^).
CO2 production h'^s'LSamnle concentration (mg I'h - Air concentration (mg I'h
Incubation time (minutes) x Sample weight (g)
2.4 Production of ammonium ions
The concentration of ammonium ions in the litter treatments was measured using an
ammonium probe. One gram of each litter treatment was dissolved into 10 ml o f R.O. water
of a McCartney bottle. Samples were gently swirled around for one minute before the probe
was used to measure the conductivity of the sample. Since, the probe was not selective for
ammonium ions, other ions can interfere with the measurements, notably K^. However, as
the experiment was conducted in the pine wood shavings, the concentration in the
material were too low to cause any significant interference. To measure the actual
ammonium ion concentration in the bedding, a standard curve was made. For this purpose
ammonium chloride was dissolved in R.O. water and a concentration range of 1, 5, 10, 50,
100, 500, 1000, 5000 and 10,000 mg 1"̂ was made. The conductivity of each concentration
was measured and a log linear relationship between conductivity and ammonium ion
concentration was seen. Based on the constructed curve, the actual concentration of
ammonium ions in the litter could be calculated.
2.5 pH
Measurement of pH was carried out by suspending Ig o f each litter treatment in 10 ml of
sterile water. The pH meter (Hanna 210 Instruments, Leighton Buzzard) was calibrated using
pH 7.01 and pH 4.01 standard solutions provided by the manufacture. Between
measurements, the electrode was washed with distilled water. The pH readings were taken
after the mixtures of wood shavings and water had been gently swirled around for one
minute.
2.6 Chemical analysis techniques
2.6.1.1 Wood extraction
Three extracts of heated wood were prepared to determine if the compounds responsible for
inhibitory activity were soluble in hot water (90°C), cold water or methanol.
40

2.6.1.2 Water extraction
Material from each set o f heat-treated and air-dried pine shavings (6 g each) were soaked in
water by adding sterile water to shavings, 2 g in 1 0 ml set up in triplicate, using the following
soaking regimes: 24 hours soaking at room temperature for a cold water treatment and 24
hours soaking at 90°C in a water bath for a hot water treatment. The extracts were then
decanted into sterile vials and kept at 4°C before they were tested.
2.6.3 Methanol extraction
A methanol extract used the same procedure as was followed for water extraction, except
using laboratory grade methanol. Samples were soaked for 24 hours on a roller shaker
(SPERAMAX 5, DENLEY Instrument Ltd.) at room temperature.
2.7 GC-MS analysis of heated and non-heated wood shavings
Gas chromatography mass spectrometry analysis of the methanol crude extracts o f heat-
treated and air-dried pine shavings was carried out using an Agilent Technologies, 7890A,
GC system with an Agilent Technologies 5975 inert XL EI/CIM SD with triple axis detector.
The samples were dissolved in chloroform or methanol (depending on solubility), 2pi o f each
sample was injected and volatised at 250°C for 25 minutes. The column was a HP-5-MS, 30
m X 250pm x 0.25 pm, which was heated initially to 50°C for 3 minutes and then to 250°C at
10°C /minute for 2 minutes, and then the temperature was held for 30 minutes.
2.8 Isolation of active compounds derived from the methanol crude extract
2.8.1 Thin layer chromatography
Thin layer chromatography (TLC) was used to choose a solvent system for column
chromatography which induced a good separation o f the components present in the methanol
crude extract of heated wood shavings of Scots pine. 100% Dichloromethane was chosen to
be the first solvent system, to be followed by a gradient elution using a mixture o f methanol
and dichloromethane. For this purpose methanol was added to dichloromethane step wise in
1 % incremental steps.
2.8.2 Column chromatography
The methanol crude extract of Scots pine (30 g) was dissolved in 100% methanol and mixed
with 10 g silica gel (Merck 9385) followed by a drying step in the fume hood for 2 days. A
50 cm cylindrical gravity column with a 4 cm internal diameter was packed using a slurry o f
silica gel and 100% hexane. The dried crude extract/silica gel was added to the packed
column and eluted using 100% dichloromethane. The eluent was collected in 75ml portions
41

giving around 260 fractions. Thin Layer Chromatography (TLC) was used to analyse the
purity of each fraction. Subsequently, fractions that had similar TLC bands were combined.
A solution of />-anisaldehyde, sulphuric acid and cold methanol (1.5:2.5:96) was used to
visualise the TLC spots. Combined fractions were further purified using small gravity
columns (1cm internal diameter) or preparative TLC plates and each purified product was
identified using NMR spectroscopic techniques. NMR spectra were recorded at room
temperature on either 300 or 500 MHz Bruker AVANCE NMR spectrometer.
2.9 In vitro assays
2.9.1 D irect contact assay
This method was developed to assess the antimicrobial activity of the fractions and
compounds derived from column chromatography. Chemicals were added onto 10 mg of
ground perlite placed inside Eppendorf tubes (1.5 ml). For each fraction, 10 pi was dissolved
in 80:20 ethanol/water and was used to coat the perlite. Four replicates of each fraction were
made. The tubes were left in a laminar flow chamber overnight to allow the solvent to
evaporate. Each tube was inoculated with 1 pi (10^ cfu ml'^) of a log phase culture of
Salmonella Enteritidis. To keep the perlite moist during incubation, 10 pi of sterile distilled
water was dispensed into each tube. As controls, four tubes received distilled water only.
The tubes were incubated overnight and from each treatment tube, a two-fold dilution series
was prepared in Ringer’s solution. For each dilution, 20 pi was plated onto XLD agar to
quantify bacterial survival. This method was used also to evaluate the minimum inhibitory
concentration of purified and identified compounds derived from extracts as well as their
commercial forms.
2.9.2 Synergistic effect assay
To assess the effect o f dehydroabietic acid on the antimicrobial effect o f vanillin, a two-fold
dilution series of 20 mmol stock solution of vanillin purified from fraction Cr 94-100 was
prepared in ethanol and water (80/20: v/v) giving concentrations from 20 mmol to 0.009
mmol. A 20 mmol solution of dehydroabietic acid, which was also purified from the same
fraction, was made in ethanol and water (80/20 v/v). Ten pi of each vanillin dilution was
pipetted onto three, 1 0 mg replicates of ground perlite, thus giving the same concentration per
gram perlite as was in the solution together with 10 pi of 20 mmol dehydroabietic acid. One
set of control treatments did not receive dehydroabietic acid, another only 2 0 mmol
dehydroabieticacid, and a third control treatment received only 1 0 pi ethanol and water at a
ratio of 80/20. Once the alcohol had evaporated, all replicates received 1 pi o f a suspension
42

of Salmonella Enteritidis (10'^ cfu mf^). Tubes were incubated at 25°C for 24 hours before
each particle was dispersed in 1 ml of Ringer’s Solution. From each suspension, six 20 pi
droplets were placed onto XLD agar. The plates were incubated at 25°C for bacterial
enumeration.
2.9.3 pH activity assay
To evaluate the effect o f pH, 20 mmol solution of citric acid (analytical grade, Fisher
Scientific) was prepared in water and the above procedure was followed. The concentration
range for vanillin was kept the same.
2.9.4 Micro Broth Dilution Method
To test the effect of storage pH and temperature on antimicrobial activity of vanillin/DHAA
mixture. Vanillin and DHAA and the mixture stock solutions were made 48 hours before the
start of the experiment. To ensure that DHAA was dissolved two methods of dissolving the
compound were used; in one the compound was dissolved in a phosphate buffer with a pH of
4, in the other treatment DSMO was used with a pH of 7. Two batches of each stock solution
at pH 4 and 7 were made, one was kept in the fridge and the other was kept at 25 °C. The
bioassay test was carried out using a micro-broth dilution technique. Vanillin and vanillin/
DHAA stock solutions were diluted further down in a two-fold dilution series to make a
concentration range between lOmM - 0.625mM. lOOpL o f each concentration was added to
lOOpL of nutrient broth in each well o f a 96 well plate. Three replicates were made for each
concentration. Plates were kept in laminar chamber to allow evaporation o f the solvent..
Subsequently, each well was inoculated with 5 pi of a Salmonella enteritidis suspension
containing 10 ̂ cfu/ml that had grown overnight at 25°C. After inoculation, the plates were
incubated at 25°C for 24 hours. To determine if treatments were bacteriocidal, 0.01 ml from
each well was spotted onto nutrient agar. Plates were incubated for 24-48h and spots with no
bacterial growth were marked as ‘bacteriocidal’. To determine a bacteriostatic effect 50 pi
of lodonitrotetrazolium chloride solution (INT, Fluka, 58030) was added to each well to
determine metabolic activity. After overnight incubation at 25°C no change in colour was
defined as a bacteriostatic effect while metabolism (no effect) was indicated by a red to
purple colour.
43

2.9.5 Bioautographic TLC
The biological activity of the methanol crude extract of pine and active fractions were
evaluated using the direct bioautographic bioassay by Homans and Fuchs (1970) with a slight
modification. TLC plates were cut into square pieces (5cm x5 cm) and aliquots of the test
compounds (20 pi) in ethanol (5 mmol) were spotted in the centre of individual plates
separately using a micropipette. For treatments containing dehydroabietic acid, a mixture of
the two compounds was made (compound 5 mmol, 20 mmol DHAA) and spotted instead.
TLC plates were left overnight inside a fume hood to dry. A 10 ml cell suspension (10'^) of
all test microorganisms were prepared and sprayed evenly over TLC plates inside a fume
hood. Plates were incubated inside a Pyrex tank containing a moist cloth at room temperature
(25°C) for 3 days. Plates were sprayed over with 5ml water solution of iodonitrotetrazolium
chloride dye (INT, Fluka, 58030) each and incubated for 8 hours. Clear zones of inhibition
were measured from the centre o f TLC plates and mean area of inhibition was calculated.
2.10 Determination of the effect of heating of rhododendron and larch wood shaving on
P. ramorum
2.10.1 Wood species of tested materials
P. ramorum infects both larch and rhododendron, therefore, wood chips from rhododendron
{Rhododendron ponticum) and Japanese larch (Larix kaempferi) were heated and their
antimicrobial properties were assessed for development of an inhibitory mulch layer to
control the spread of P. ramorum in areas that were targeted for clearing.
2.10.2 Preparation of crude extracts
To extract the chemical in the heated woods using chromatography techniques, first wood
from Scots pine {Pinus sylvestris), rhododendron {Rhododendron ponticum) and Japanese
larch {Larix kaemopferi) were processed into fine shavings {ca 500 g for each wood type)
wrapped in aluminium foil and placed in a dry oven. Subsequently, the wood shavings were
extracted with methanol by following the procedure in 2.6.3 (Figure 2.1).
2.10.3 Test isolates
Two isolates of P. ramorum originating from two different hosts were selected from the P.
ramorum collection held at Forest Research, Famham, Surrey. Isolates P I578 and BRCOl
had been isolated from rhododendron and Japanese larch respectively. Both isolates belong
to the EUl lineage o f P. ramorum and later, one isolate from the EU2 lineage, P2460, was
also included in some of the in vitro assays.
44

Figure 2.1 The m ethanol extracts o f heat-treated (left) and air-dried (right) wood shavings o f pine.The wood materials w ere soaked in laboratory grade methanol (500 gram in 2.5 litres) on a platform shaker for 48 hours. The extracts were sieved through filter paper tw ice and were left inside a fum e hood for a week to dry.
2.10.3.1 Inoculum preparation
Zoospores
To produce a zoospore suspension, P. ramorum was sub-cultured on fifty carrot agar plates.
(50 g peeled, chopped carrots; 15 g technical agar; 1 litre of distilled water). Plates were kept
under continuous fluorescent light at room temperature for two weeks. Subsequently, 5ml of
distilled water was added to each plate and with a plastic spreader gently swirled around the
plates to dislodge the sporangia from the mycelia. The water containing the sporangia was
pipetted into a sterile beaker and kept in the fridge for 2 0 minutes, then moved to room
temperature for 45 minutes to release the zoospores. Spores from each isolate were counted
four times, using a haemocytometer according to Forest Research Standard Operating
Procedure (Forest-Research, 1997)
Chlamvdospores
Chlamydospores were prepared by the method o f (Tooley et a i. 2008). A spore suspension
of lO' ̂ zoospores m f' in sterile water (300 ml) were prepared and transferred to a zip-lock
plastic bag. Rhododendron leaf disks ( 6 mm diameter) were cut using a cork borer. Two
hundred leaf disks were added to the spore suspension and bags were sealed and kept at room
temperature for 5 days. The water content of each bag was removed using a sieve and the
leaf disks were rinsed with sterile water three times. Leaf disks were transferred to a
moistened tissue towel and placed inside a plastic container. The two containers were
incubated in the dark at 20°C for 4 weeks. Chlamydospores were observed on the leaf disk
using a light microscope.
45

Mycelia as the source of inoculum
A zoospore suspension (concentration of 10"̂ zoospores mf^) was made for each test isolate
and the wells of a 96-well plate were pre-loaded with 100 |il of sterile water per well, 10 pil of
spore suspension then added to each well, and the plates incubated in the dark for 3 days.
The mycelia forming inside the wells were examined after three days under a binocular
microscope and transferred to pre-loaded 96-well plates with known concentrations of the
crude extracts as well as the chemicals derived from the heated wood of the selected species.
2.10.4 In vitro assays
Micro broth dilution method
The inhibitory activity of crude extracts obtained from heated pine, rhododendron and larch
was tested on two isolates of P. ramorum using a micro broth dilution method. For this test,
each well of a 96 well micro titre plate was filled with 0.96 ml carrot broth. A stock solution
of 50 mg ml" ̂ o f each crude extract was made in acetone. A two fold dilution series (25,
12.5, 6.25 mg extract ml'^) was prepared and pipetted into the broth. Three replicates were
prepared for each concentration. Control wells received only acetone and carrot broth. Plates
were left in a laminar flow cabinet for 5 hours to allow the solvent to evaporate. A spore
suspension o f P. ramorum was prepared (as described in 2.10.3.1). Each well was inoculated
with 5 pi of a 10"̂ zoospores ml'^ spore suspension. After inoculation, the plates were
incubated at 20°C for 24 hours. To determine if treatments were fungicidal, 20 pi from each
well was spotted onto carrot agar plates which were incubated for 24-48 hours. Inoculum
spots with no growth were marked as positive fungicidal test results.
2.11 Data analysis
The data were analysed by a one-way Analysis o f Variance ANOVA, GraphPadPrism,
version lnStat3 (GraphPad-Software, 1998). The Tukey Post Analysis Test was performed to
assess the least significance difference between means at a P<0.05 level.
46

CH APTER 3: EVALUATIO N OF H EAT-TREATED W O O D AS LITTER
3.1 Introduction
Poultry litter is known to harbour high numbers of bacteria (Dumas et a l, 2011; Li et a l,
2003; Scheffer, 1965; Terzich et a l, 2000) and the total bacteria concentrations can vary
fromlO^^ and 10̂ ̂ colony forming units per gram of litter (Scheffer, 1965; Terzich et a l,
2000). Among these, pathogenic bacteria causing food-borne diseases are o f great concern to
the public and the poultry industry. Bacteria belonging to the species Salmonella are the
leading cause of intestinal diseases in humans and animals worldwide (Castiglioni Tessari et
a l, 2012). The annual report on zoonoses and food-home outbreaks in the European Union
shows Salmonella accounted for 99,020 reported human cases in 2010 (EPSA, 2010a).
Nearly 26% of Salmonella infections in the UK were associated with consumption of poultry
meat in 2009 (Defra, 2011).
Poultry litter is shown to play an important role in Salmonella survival and transmission in
broiler houses (Doyle and Erickson, 2006; Tucker, 1967). Tucker (1967) found that cycling
of Salmonella between the intestinal tract and litter seems to be significant in maintaining
intestinal infection of broilers. Salmonella can survive for up to 26 months in poultry litter
and spread the contamination among the flock in built-up litter. Higher carcass
contamination is also reported to be correlated with higher contamination o f Salmonella in
the litter during grow-out periods (Volkova et a l , 2009).
Extended survival of Salmonella and the high risk of its introduction into broiler units makes
control difficult. There is no single control strategy which is able to guarantee Salmonella-
free chickens. In the UK, DEFRA welfare codes of practice for prevention and control of
Salmonella are continuously updated to inform farmers and producers about the latest
changes and implementation. In 2011, the disease mitigation and control evidence plan
provided by DEFRA highlighted the need for on-farm cost-effective innovation and the
development of control strategies for zoonotic diseases based on current knowledge in Great
Britain (Defra, 2011). Although, national control programmes o f Salmonella have been
successful in reducing Salmonella incidences, further research is needed to avoid the risk of
new Salmonella strains becoming prevalent and possible changes of pathogen serovors
(Defra, 2011; Doyle and Erickson, 2012).
Ammonia production and volatilization in poultry units is another major health issue for
animals, humans and the environment. The litter condition has a direct influence on the
47

ammonia production level in broiler units. Natural break down of poultry manure by
mineralizing bacteria takes place within the litter environment which leads to ammonia
production and subsequently increasing pH and ammonia volatilization. Application of
acidifying amendments is the most effective current strategy to control pH of the litter
(Marsh, 1999a; Wheeler et a l, 2008). However, use of amendment above manufacturer’s
recommended rate has increased concern about emerging virulent strains of bacteria as a
result o f being exposed to high levels of chemicals (Wheeler et a l, 2008).
Effects of the type of bedding materials on survival of poultry pathogens and ammonia
formation have been studied extensively (Atapattu et a l, 2008; Atencio et a l, 2010; Bilgili et
a l, 1999; Duqueza, 1996; Macklin et a l, 2005; Miles et a l, 2004; Miles et a l, 2011b;
Tasistro et a l, 2007; Torok et o/., 2009). In many cases, the physical parameters o f bedding
materials such as the water holding capacity are found to make a significant difference
between different bedding materials (Atencio et a l, 2010; Bilgili et a l, 1999; Macklin et a l,
2005; Miles et a l, 2011a). Others consider the chemical characteristics o f bedding materials
to be responsible for inhibiting microbial activity (Atapattu et a l, 2008; Duqueza, 1996).
Chopped straw, peanut hulls, rice husks, sand, shredded paper, sawdust, and wood shavings
are commonly used in different countries. Wood materials have been shown to be
particularly efficient in reducing ammonia emissions and improving broiler health and
performance (Miles et a l, 2011b; Torok et a l, 2009). Other research on the survival of
pathogenic bacteria on wood materials have shown that wood exhibits antimicrobial action
against food-borne pathogens such as Salmonella and E. coli (Schonwalder et a l, 2000,
2002). In general, softwoods do not support bacterial growth as well as hardwoods which
may be due to differences in their chemical characteristics (Milling et a l, 2005; Schonwalder
et a l, 2000, 2002). However, choice of bedding materials is limited by what is available on
the market and by season (Embury, 2004). This makes it difficult to implement the
application o f antimicrobial bedding materials and highlights the need for investigating
alternative bedding materials that enhance antimicrobial characteristics.
A review o f the literature has shown that thermal treatments o f wood are well developed to
produce wood with desirable physical and chemical properties. Thermal treatment is mainly
used to improve wood dimensional stability and its resistance to weathering conditions and
decay fungi (Kamdem et a l, 2002; Stamm et a l, 1946; Voda et a l, 2003). Literally several
hundreds of chemicals are produced from wood processing technologies with different
applications in the pharmaceutical and other industries (Mackay et a l, 2009b; Murzin et a l,
48

2007). Up to the present time, research on efficacy or potential applications of heat treated
wood materials and their derived chemicals has been limited to improving the life duration of
wood itself and its resistance toward wood decay fungi.
This chapter aims to describe the development o f a heating regime to enhance the
antimicrobial activity o f pine wood shavings for use as bedding materials in broiler houses.
Due to the significance of food-home diseases caused by Salmonella strains. Salmonella
Enteritidis was chosen as the test organism to evaluate the inhibitory effect o f bedding
materials on Salmonella.
3.2 Experimental
Initially, survival of Salmonella on heated wood materials was tested. In later work to
measure the effect of antimicrobial activity o f treatments on ammonia production, a model
litter was prepared by homogenzing heated wood materials with an aliquot o f litter from a
broiler house to introduce the complex microbial population of chicken manure. Commercial
uric acid was added to provide the microbes with a source of nitrogen.
Full details of the experimental methods and methods o f statistical analysis are given below
and in Chapter 2, sections 2.11.
3.2.1 Wood inoculation
From heat-treated and air-dried materials three batches of one gram wood shavings were
weighed and transferred into McCartney bottles, giving three replicates for each treatment.
Each received 2 ml sterile distilled water one hour prior to inoculation and was then
inoculated with 1 ml of a suspension (10^ cfu ml"^) of Salmonella Enteritidis. Care was taken
to ensure that the suspension was completely absorbed by the test material. Inoculated
shavings were incubated at 25°C for 48 hours. One gram of materials from each treatment
was weighed into a clean McCartney bottle and received 10 ml sterile distilled water. A 10-
fold dilution series (neat - 1 0 '^) was prepared from the shavings and 0 . 1 ml of each dilution
plated onto XLD agar. Plates were incubated for 36 hours at 25°C for bacterial enumeration.
3.2.2 Litter model preparation
Fifty grams of test materials including heat-treated, air-dried and commercial wood shavings,
perlite were placed in 2.5 litre dispo-jars (four replicates for each treatment) that were fitted
with a sampling port for gas analysis. The materials were soaked to 60% of their water
holding capacity, and amended with 1 % spent chicken litter to provide a large microbial
49

inoculum and 1% uric acid that could be converted to ammonia by microbes. All model
treatments were incubated at 25°C for 48 hours to make a homogenized chicken litter (Figure
3.1). Each treatment was replicated four times and samples of gas and material were taken
every three days to assess microbial respiration (CO? evolution), ammonium ion
concentration and the pH of the material. Litter and air samples were taken 4, 7 and 11 days
after incubation and tested for ammonia production and changes in pH.
Figure 3.1 Litter models made in 2.5 liter plastic containersThe containers were fitted with a sam pling port for gas analysis and were kept inside an incubator at 25°C throughout the experim ent.
3.3 Results
3.3.1 Effect of heating regime on the antim icrobial properties of pine bark
The effect o f temperature on the antimicrobial properties o f pine shavings was assessed by
heating ground pine bark and pine wood shavings for 24 hours at temperatures ranging from
50 to 200°C. Antimicrobial activity was assessed by estimating the survival o f Salmonella
Enteritidis in the treated material 48 hours after inoculation. The survival of bacteria in the
heated bark decreased by 2 log units when the temperature o f the treatment increased from
50°C to 100°C (P<0.05) and was completely inhibited on pine wood shavings which had been
heated to 150°C and 200°C in contrast to the outcome at 50°C (P<0.0()1) (Table 3.1).
Table 3.1 Effect o f different heating tem peratures on the recovery o f SE from ground pine bark
Tem perature "C (n=3) Recovery o f SE (log cfu g ‘ ± SD)50 6.30 ± 0.04"*100 4.29 -t 0 .04”150 0.00 1: 0.00"200 0.00 t 0 .00“Significance PO.OOl
^D ifferent letters signify significant differences (P<0.05) between means. (D etection limit= 2 log cfu g ‘)
50
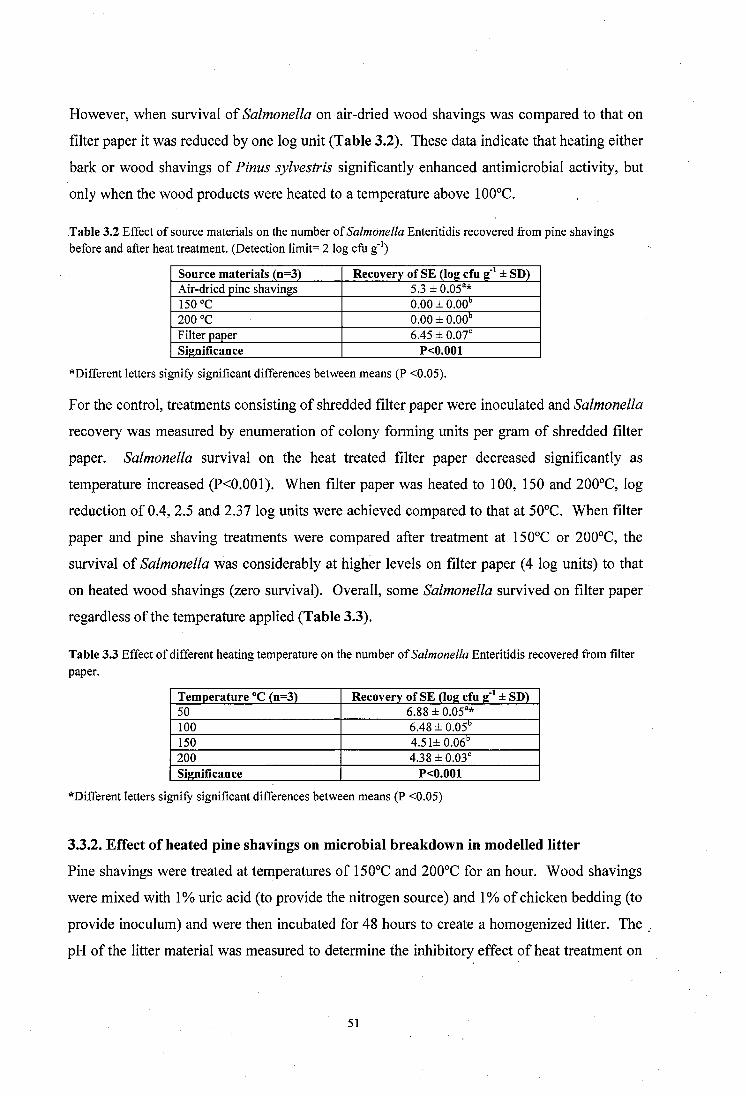
However, when survival of Salmonella on air-dried wood shavings was compared to that on
filter paper it was reduced by one log unit (Table 3.2). These data indicate that heating either
bark or wood shavings o f Pinus sylvestris significantly enhanced antimicrobial activity, but
only when the wood products were heated to a temperature above 100°C.
Table 3.2 Effect o f source materials on the number o f Salm onella Enteritidis recovered from pine shavings before and after heat treatment. (Detection limit= 2 log cfu g'^)
Source m aterials (n=3) R ecovery o f SE (log cfu g' ̂± SD)Air-dried pine shavings 5.3 ± 0.05"*150 °C 0.00 ± 0 .0 0 ”200 °C 0.00 ± 0.00”Filter paper 6.45 ± 0.07“Significance P <0.001
*Different letters signify significant differences between means (P <0.05).
For the control, treatments consisting of shredded filter paper were inoculated and Salmonella
recovery was measured by enumeration o f colony forming units per gram of shredded filter
paper. Salmonella survival on the heat treated filter paper decreased significantly as
temperature increased (P<0.001). When filter paper was heated to 100, 150 and 200°C, log
reduction of 0.4, 2.5 and 2.37 log units were achieved compared to that at 50°C. When filter
paper and pine shaving treatments were compared after treatment at 150°C or 200°C, the
survival of Salmonella was considerably at higher levels on filter paper (4 log units) to that
on heated wood shavings (zero survival). Overall, some Salmonella survived on filter paper
regardless of the temperature applied (Table 3.3).
T able 3.3 Effect o f different heating temperature on the number o f Salm onella Enteritidis recovered from filter paper.
Tem perature “C (n=3) R ecovery o f SE (log cfu g"̂ ± SD)50 6.88 ±0.05"*100 6.48 ± 0.05”150 4 .5 1± 0 .06”200 4.38 ± 0.03“Significance P <0.001
*Different letters signify significant differences between means (P <0.05)
3.3.2. Effect of heated pine shavings on microbial breakdown in modelled litter
Pine shavings were treated at temperatures of 150°C and 200°C for an hour. Wood shavings
were mixed with 1 % uric acid (to provide the nitrogen source) and 1 % of chicken bedding (to
provide inoculum) and were then incubated for 48 hours to create a homogenized litter. The
pH o f the litter material was measured to determine the inhibitory effect o f heat treatment on
51

the microbial breakdown of uric acid in the litter. Perlite and air-dried pine shavings were
modelled as control litters for eomparison.
Table 3.4 Effect o f heating at 150°C and 200°C on the pH o f pine shavings made into litter.
M odelled litter (n=4)
pH
4 days 7 days 11 daysPerlite 6.43 ± 0.05"* 8.33 ±0.02" 9.48 ±0.01"
A ir-dried pine shavings 6.68 ±0.03" 6.67 ±0.08” 8.75 ±0.10"”1 5 0 T 4.29 ± 0.03” 4.60 ± 0 .2 3 “ 8.27 ± 0 .4 1 ”2 0 0 T 5.84 ± 0 .1 f 7.27 ± 0.42"” 9.13 ±0 .02" ”
Significance P<0.001 P <0.001 P<0.001
^D ifferent letters signify significant differences between m eans (P <0.05)
As shown in Table 3.4, after 4 days the pH of the eontrol litters (perlite and air-dried
shavings) was signifieantly higher than the pH of both litters made with heat-treated shavings
heated to 150°C and 200°C (P<0.001 ). After 7 days, the pH value o f the heated pine shavings
at 200°C increased to 7.2 whieh was not significantly different from the pH of perlite and air-
dried shavings. In contrast, the litter with pine shavings heated to 150°C had the lowest pH
value of 4.6 (P<0.001 ). After 11 days, there were no significant differences between the pH
of air-dried or heat-treated litters at either 150°C and 200°C. Overall eomparison, wood
shavings heated to 150°C maintained the lowest pH values after 4. 7 and 11 days of
incubation (Figure 3.2).
pH of m odel litter
109.5
98.5
7.5
^6.5b
5.5 5
4.5 4
3.5 3
♦ perlite
■ air-dried
▲ 150°C
X 200°C
4 6 8 10
Incubation tim e (days)
12 14
Figure 3.2 Change o f pH in m odelled litter over time.
3.3.3 Effect of heated pine wood on CO 2 production from wood shavings based litter (as
indicator of microbial activity)
Heat treated shavings were made into a model chicken litter system (see 3.2.2) to assess the
effect o f heating pine shavings for an hour at either 15()°C or 200°C on microbial activity.
52

Controls consisted of perlite and air-dried wood shavings treated as described above.
Microbial activity was assessed by measuring CO2 production as an indicator of microbial
activity (mg CO iper kg of litter per hour) after 4 ,1 and 11 days.
As shown in Table 3.5, after 4 days o f incubation at 25°C, both 150°C and 200°C heated pine1 1supported significantly lower bacterial respiration rates (6.3 and 103 mg CO2 kg' h' )
compared with the two controls, air-dried pine wood (146 mg CO2 kg'^h'^) and perlite based
litter (272 CO2 mg kg" ̂ h'^) (P<0.001). After 7 and 11 days, the microbial respiration of
150°C heated pine was also significantly lower than perlite, air-dried and heat treated pine at
200°C (P<0.001). Although heated pine at 200°C had a higher microbial respiration rate
compared to 150°C, after 7 and 11 days it supported significantly lower microbial respiration
(174; 195 mg CO2 kg'^ h'^) compared to air-dried (241; 260 mg CO2 kg'^ h'^) and perlite ( 359
mg CO2 kg'^ h'^) treatments.
Table 3.5 Effect o f heating at 150°C and 200°C for one hour on the microbial activity expressed as CO2
production in a pine shavings base litter.
M odelled litter (n=4)
R espiration (m g CO 2 kg^ h'^)
4 days 7 days 11 daysPerlite 272 ±26"* 359.7± 20" 128.6 ± 8.2"Air-dried Pine shavings 146 ±10” 241 ± 13.4” 260.4 ± 1 1 .7 ”150°C 6.3 ± 3.2“ 32.1 ± 4 .5 “ 89.1 ± 5 .3 “200°C 1 0 3 ± 1 8 ” 174 ± 2 5 ” 195.8 ± 2 3 .2 ”Significance P<0.0 01 P<0.001 P<0.001
*Different letters signify significant differences between means (P <0.05)
3.3.4 Effect of heating duration at 120®C on CO2 production, pH and ammonia
concentration
The effect of heating pine shavings at 120°C for increasing periods o f time (1 to 72 hours) on
the antimicrobial activity of the treated shavings was measured, using the model chicken litter
system (see 3.2.2). Microbial activity was assessed by measuring microbial respiration (CO2
evolution), rise in pH (as a result of ammonium ion formation) and ammonium ion
concentrations in the litter after 4, 7 and 11 days incubation at 25°C. The results are
presented in Tables 3.6, 3.7 and 3.8.
53
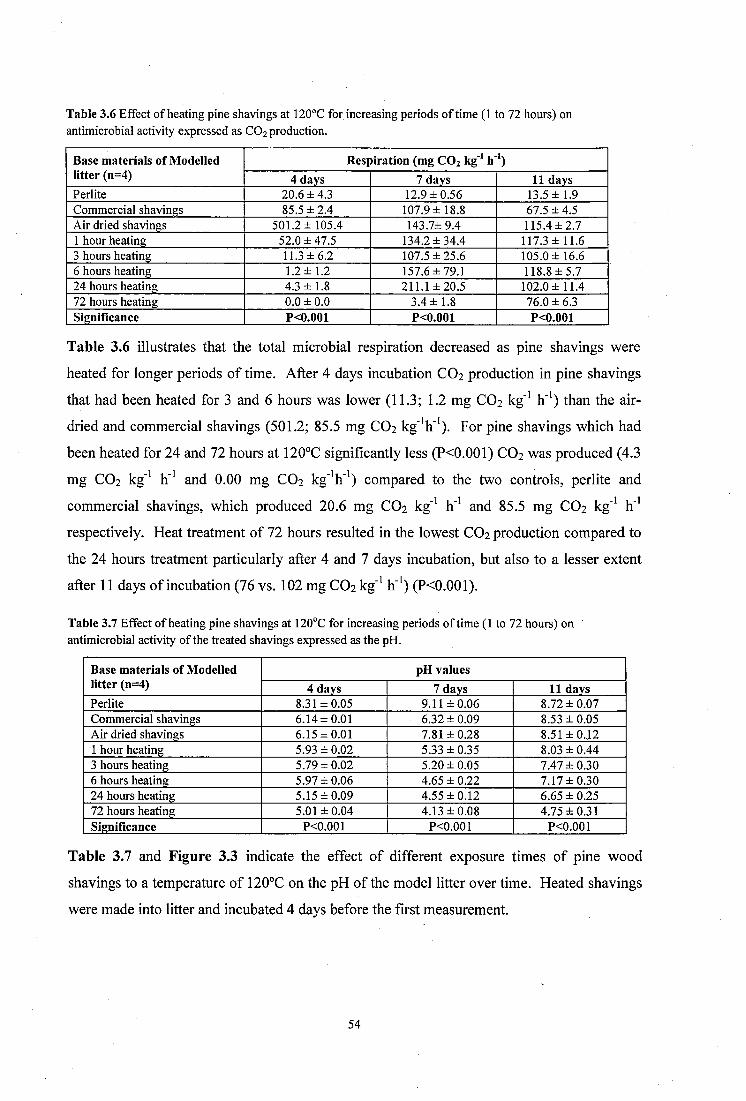
Table 3.6 Effect o f heating pine shavings at 120°C for increasing periods o f time (1 to 72 hours) onantimicrobial activity expressed as CO2 production.
Base m aterials o f M odelled litter (n=4)
R espiration (m g C O 2 kg'^ h'^)
4 days 7 days 11 daysPerlite 20.6 ± 4.3 12.9 ± 0 .5 6 13.5 ± 1.9Commercial shavings 85.5 ± 2.4 107.9 ± 18.8 67.5 ± 4.5Air dried shavings 501.2 ± 105.4 143.7± 9.4 115.4 ± 2 .71 hour heating 52.0 ± 47.5 134.2 ± 3 4 .4 117.3 ± 11.63 hours heating 11.3 ± 6 .2 107.5 ± 25.6 105.0 ± 16.66 hours heating 1.2 ± 1.2 157.6 ± 7 9 .1 118.8 ± 5 .724 hours heating 4.3 ± 1.8 211.1 ± 2 0 .5 102.0 ± 11.472 hours heating 0.0 ± 0.0 3.4 ± 1.8 76.0 ± 6.3Significance P<0.001 P<0.001 P<0.001
Table 3.6 illustrates that the total microbial respiration decreased as pine shavings were
heated for longer periods of time. After 4 days incubation CO2 production in pine shavings
that had been heated for 3 and 6 hours was lower (11.3; 1 . 2 mg CO2 kg'^ h'^) than the air-
dried and commercial shavings (501.2; 85.5 mg CO2 kg'^h'^). For pine shavings which had
been heated for 24 and 72 hours at 120°C significantly less (P<0.001) CO2 was produced (4.3
mg CO2 kg'^ h'^ and 0 . 0 0 mg CO2 kg'^h"^) compared to the two controls, perlite and
commercial shavings, which produced 2 0 . 6 mg CO2 kg" ̂ h'^ and 85.5 mg CO2 kg'^ h'^
respectively. Heat treatment of 72 hours resulted in the lowest CO2 production compared to
the 24 hours treatment particularly after 4 and 7 days incubation, but also to a lesser extent
after 1 1 days of incubation (76 vs. 1 0 2 mg CO2 kg'^ h" )̂ (P<0 .0 0 1 ).
Table 3.7 Effect o f heating pine shavings at 120°C for increasing periods o f time (1 to 72 hours) on antimicrobial activity o f the treated shavings expressed as the pH.
Base m aterials o f M odelled litter (n=4)
pH values
4 days 7 days 11 daysPerlite 8.31 ± 0 .0 5 9.11 ± 0 .0 6 8.72 ± 0 .0 7Commercial shavings 6.14 ± 0 .0 1 6.32 ± 0 .0 9 8.53 ± 0.05Air dried shavings 6.15 ± 0 .0 1 7.81 ± 0 .2 8 8.51 ± 0 ,1 21 hour heating 5.93 ± 0.02 5.33 ± 0.35 8.03 ± 0.443 hours heating 5.79 ± 0 .0 2 5.20 ± 0.05 7.47 ± 0.306 hours heating 5.97 ± 0.06 4.65 ± 0.22 7.17 ± 0 .3 024 hours heating 5.15 ± 0 .0 9 4.55 ± 0 .1 2 6.65 ± 0 .2 572 hours heating 5.01 ± 0 .0 4 4.13 ± 0 .0 8 4.75 ± 0 .3 1Significance P<0.001 P<0.001 P<0.001
Table 3.7 and Figure 3.3 indicate the effect of different exposure times of pine wood
shavings to a temperature of 120°C on the pH of the model litter over time. Heated shavings
were made into litter and incubated 4 days before the first measurement.
54
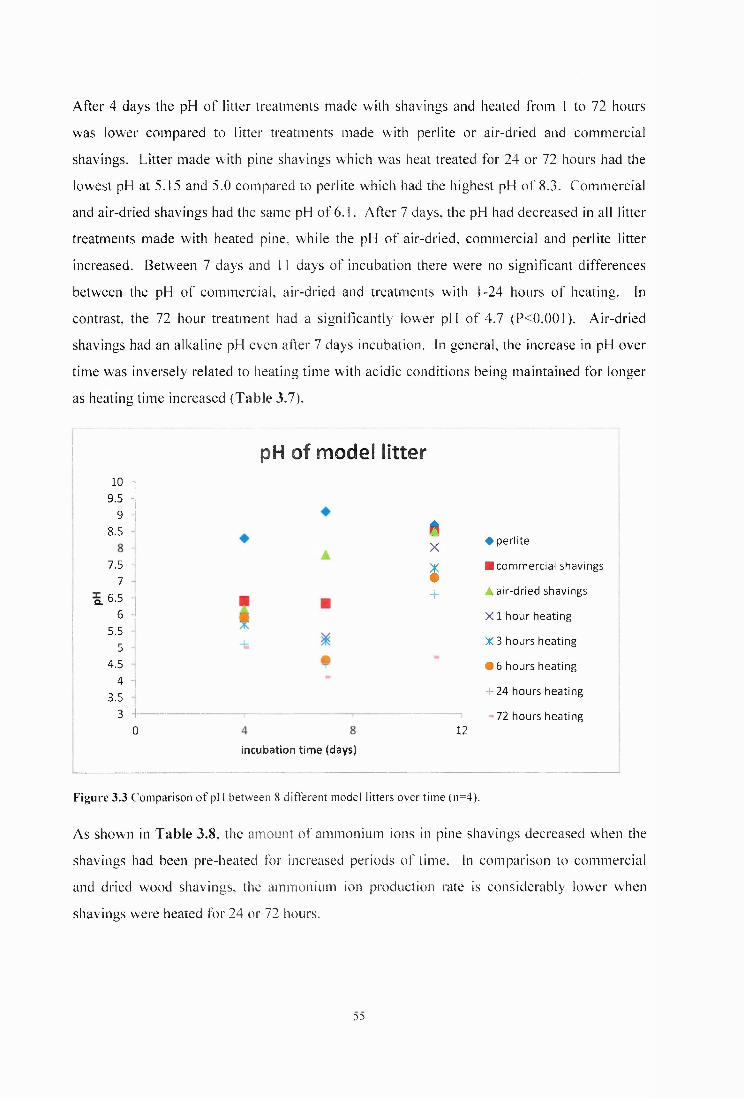
After 4 days the pH of litter treatments made with shavings and heated from I to 72 hours
was lower compared to litter treatments made with perlite or air-dried and commercial
shavings. Litter made with pine shavings which was heat treated for 24 or 72 hours had the
lowest pH at 5.15 and 5.0 compared to perlite which had the highest pH of 8.3. Commercial
and air-dried shavings had the same pH of 6 .1. After 7 days, the pH had decreased in all litter
treatments made with heated pine, while the pH of air-dried, commercial and perlite litter
increased. Between 7 days and 11 days of incubation there were no significant differences
between the pH o f commercial, air-dried and treatments with 1-24 hours o f heating. In
contrast, the 72 hour treatment had a significantly lower pH of 4.7 (P<0.001). Air-dried
shavings had an alkaline pH even after 7 days incubation. In general, the increase in pH over
time was inversely related to heating time with acidic conditions being maintained for longer
as heating time increased (Table 3.7).
I pH of m odel litter10 1
9.5 - 9
8.5
7.5 7
a 6.5 6
5.5 5
4.5
4 13.5
3 4
ftX
$+
♦ perlite
■ commercial shavings
A air-dried shavings
X 1 hour heating
X 3 hours heating
• 6 hours heating
+ 24 hours heating
- 7 2 hours heating0 12
incubation tim e (days)
Figure 3.3 Com parison o f pH between 8 different model litters over tim e (n=4).
As shown in Table 3.8. the amount o f ammonium ions in pine shavings decreased when the
shavings had been pre-heated for increased periods o f time. In comparison to commercial
and dried wood shavings, the ammonium ion production rate is considerably lower when
shavings were heated for 24 or 72 hours.
3 0
Table 3.8 Effect o f heating pine shavings at 120°C for increasing periods o f time (1 to 72 hours) onantimicrobial activity o f the treated shavings expressed as ammonium ion concentration.
Base m aterials o f M odelled litter (n=4)
A m m onium ion concentration (m g kg'^)
4 days 7 days 11 daysPerlite 1058 ± 7 5 455 ± 24 303 ± 17Commercial shavings 50 ± 0 508 ± 1 9 590 ± 42Dried shavings 8 0 ± 14 360 ± 27 3 1 0 ± 91 hour heating 8 0 ± 11 133 ± 96 315 ± 5 43 hours heating 70 ± 7 2 0 5 ± 152 378 ± 756 hours heating 1 1 7 ± 5 43 ± 5 323 ± 3324 hours heating 132 ± 4 0 75 ± 2 0 275 ± 1972 hours heating 90 ± 6 7 3 ± 10 108 ± 49Significance P<0.001 P<0.001 P<0.001
Wood that was heated for 72 hours at 120°C had the lowest concentration of ammonium ions
after 4, 7 and 11 days of incubation (90, 73, 108 mg ammonium ion kg’ ̂ respectively)
compared with perlite after 4, 7 and 11 days (1058, 455, 303 mg ammonium ion kg'^
respectively) and wood shavings that had been heated for 24 hours after 4, 7 and 11 days
(132, 75, 275 mg ammonium ion kg'^ respectively). These differences were highly
significant (P<0.001) (Table 3.8).
3.3.5 Spectrum of enhanced antimicrobial activity of heated pine shavings
The antimicrobial activity of heat treated pine shavings (120°C for 72 hours) was tested
against two other pathogens that are frequently isolated from poultry litter. Staphylococcus
aureus and Escherichia coli. The inhibitory effect o f heated shavings on each pathogen was
determined by inoculating one gram of heated shavings with 1 0 ̂ cells o f the test strains.
Numbers of the bacteria recovered from heated shavings was determined by enumeration of
colony forming units on agar plates. The results are shown in Table 3.9. Survival of both
pathogens was completely inhibited on heated shavings as compared with air-dried shavings
and filter paper (P< 0.001). Survival of both pathogens was higher on air-dried shavings
compared to that on filter paper.
T able 3.9 Survivals o f S. aureus, and E. coli and on heat treated and air-dried wood o f pine and air dried and heated filter paper. (Detection limit= 2 log cfii g'^)
Survival (log cfu g' ̂± SE)
Tested strains (n=3) H eated pine A ir-dried pine Filter paper SigniHcance
Staphylococcus aureus ATCC 25923 00 ± 0 0 7.78 ± 00 5.0 ± 00 P<0.001
Escherichia coli ATCC 11775 00 ± 0 0 7.34 ± 0 .0 2 6.72 ± 0 .0 1 P<0.001
56
3.3.6 Evaluation of the antimicrobial activity of different wood types after heating
A heating regime of 120°C for 72 hours was applied to wood shavings from eight different
tree species as follows: pine, spruee, cypress, ash, beech, red oak, bireh, eucalyptus. The
extent of antimicrobial activity was determined by inoculating 1 gram of each wood type with
10 ̂cells of Salmonella Enteritidis. Survival was measured after 48 hours incubation at 25°C
(Table 3.10). Heating at 120°C significantly inhibited the growth o f Salmonella on wood
shavings of pine, cypress, ash, beech, red oak and bireh (zero recovery) compared to survival
on air-dried wood shavings made from the same tree species (P<0.001). There were also
small, but significant differences in the antimicrobial properties of air-dried wood shavings.
Spruce, birch and oak appeared to have the least antimierobial activity, with large numbers of
bacteria (>10^ colonies g'^) recovered from the wood shavings of these tree species. Cyprus
and eucalyptus appeared to be the most antimicrobial before heat-treatment, with almost two
times less recovery of Salmonella compared with spruce. Heating resulted in a significant
increase (P<0.001) in the antimicrobial activity of all of the wood species that were tested,
resulting in a 4 fold increase in the antimierobial activity of eucalyptus wood shavings, a
1000 fold increase in antimicrobial activity in spruce, and complete inhibition of Salmonella
with pine, Cyprus, ash, beach, oak and birch (Table 3.10).
Table 3.10 Recovery o f Salm onella Enteritidis from inoculated heated and air-dried wood shavings derived from 8 different wood species.
W ood type (n=3) A ir-dried (log cfu g'b H eated (log cfu g'b SignificancePine (soft) 6.88 ± 0 .1 1 “* 0
Spruce (soft) 7.56 ± 0.06” 4.50 ± 0 .0 2 P<0.001Cypress (soft) 5.24 ± 0.07” 0
Ash (hard) 7.12 ± 0.06“ 0Beech (hard) 6.85 ± 0 .1 9 “ 0
Red Oak (hard) 7.22 ± 0 .1 3 “ 0Birch (hard) 7.32 ± 0.04“*̂ 0Eucalyptus 5.93 ± 0.07” 5.3 ± 0.06 P<0.001
Significance P<0.001 P<0.001
*Different letters indicate a significant difference (P<0.05) between means.
3.3.7 Effect of time of exposure to heat treatment of 120”C on pH of pine wood shavings
Wood shavings of pine were heated for 0, 0.5, 1, 2, 3, 4, 5, 6 , 7, 8 , 24, 30 and 48 hours and
the pH of each treatment was measured by suspending 2 grams of wood shavings in 20 ml
sterile water before pH was measured. The pH of wood shavings were found steadily to
decrease from an average of 4.6 before heating to 4.2 after 24 and 30 hours of exposure to
120°C. After 48 hours the pH value decreased to an average o f 3. Figure 3.4 shows the pH
57
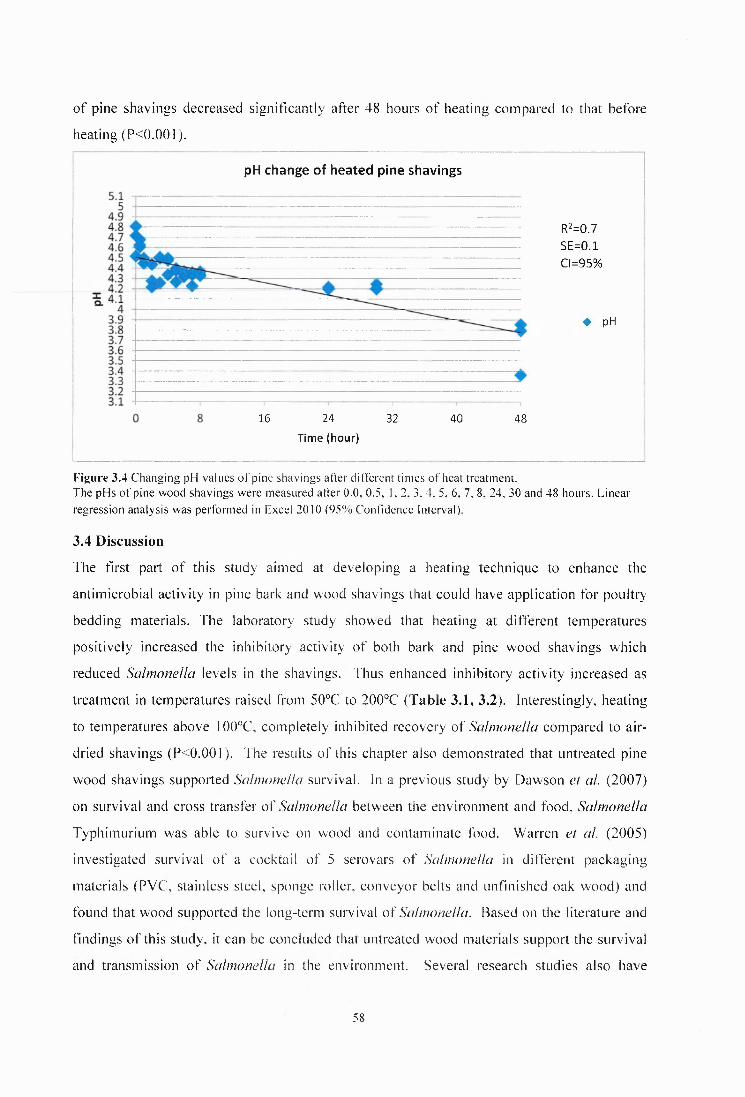
of pine shavings decreased significantly after 48 hours of heating compared to that before
heating (P<0.001 ).
pH change of heated pine shavings
R2=0.7SE=0.1Cl=95%
♦ pH
16 24
Tim e (hour)
32 40 48
Figure 3.4 Changing pH values o f pine shavings after different tim es o f heat treatm ent.The pHs o f pine wood shavings were measured after 0.0, 0.5, 1, 2. 3, 4. 5, 6, 7, 8. 24, 30 and 48 hours. Linear regression analysis was perform ed in Excel 2010 (95% Confidence Interval).
3.4 Discussion
The first part o f this study aimed at developing a heating technique to enhance the
antimicrobial activity in pine bark and wood shavings that could have application for poultry
bedding materials. The laboratory study showed that heating at different temperatures
positively increased the inhibitory activity o f both bark and pine wood shavings which
reduced Salmonella levels in the shavings. Thus enhanced inhibitory activity increased as
treatment in temperatures raised from 50°C to 200°C (Table 3.1, 3.2). Interestingly, heating
to temperatures above 100°C, completely inhibited recovery o f Salmonella compared to air-
dried shavings (P<0.001). The results of this chapter also demonstrated that untreated pine
wood shavings supported Salmonella survival. In a previous study by Dawson et al. (2007)
on survival and cross transfer o f Salmonella between the environment and food. Salmonella
Typhimurium was able to survive on wood and contaminate food. Warren el al. (2005)
investigated survival o f a cocktail of 5 serovars of Salmonella in different packaging
materials (PVC, stainless steel, sponge roller, conveyor belts and unfinished oak wood) and
found that wood supported the long-term survival o f Salmonella. Based on the literature and
findings of this study, it can be concluded that untreated wood materials support the survival
and transmission o f Salmonella in the environment. Several research studies also have
58

demonstrated a high incidence of Salmonella in new bedding material (Dougherty, 1976;
Long et a l, 1980). Therefore, a pre-heating treatment o f bedding materials may reduce the
risk o f Salmonella contamination in broiler at the time o f placement. This is very important
since younger chickens are more susceptible to Salmonella when placed on new bedding
material and use of contaminated wood shavings may increase the risk of Salmonella
outbreaks on-farm.
Further experiments on the effect of exposure time to heat treatment showed perlite and dried
shavings supported significantly higher levels of respiration compared with shavings given an
hour of heat treatment. However, dried shavings and 1 hour heat-treated shavings did not
differ in the amount of ammonia they produced (Table 3.8). The effect o f time exposure on
enhanced antimicrobial activity showed that longer time exposures to a temperature of 120°C
significantly decreased microbial respiration of the model litters when compared to perlite,
air-dried and commercial shavings litter over an 11 days test (Table 3.6). The differences
were more distinctive between the exposure times after 4 days, with 3, 6 and 24 hours
showing the most variation between 4 and 7 days o f incubation. However, the differences
between all heat treated shavings were not significant except with the 72 hour o f heat
treatment sample (Table 3.6). The 72 hours heat treatment sample also had the lowest
microbial respiration rate after 11 days o f incubation. Although, the inhibitory activity of
heated wood o f pine on mineralizing bacteria was not directly evaluated, the lower respiration
rate from heated wood material indicates that the activity o f microorganisms in the litter was
inhibited, including mineralizing bacteria. In addition, ammonia formation in heat treated
wood shavings samples was also significantly different from air-dried, commercial shavings
and perlite litters after 7 and 11 days, and particularly low for 24 and 72 hours exposure
(Table 3.8). Ammonia production of commercial shavings and air-dried shavings was
approximately 9-10 times and 5 times more than o f the 6 , 24 and 72 hours heat treated
shavings after 7 days (Table 3.8). Variation in microbial respiration in the 3, 6 and 24 hours
treatment was also reflected in ammonia production. However, after 11 days only the 72
hour heat treated shavings had a significantly lower value (108 mg ammonium ion kg'^)
compared to that of commercial shavings with a value of 590 mg ammonium ion kg (Table
3.8). In the tested litters, pH was directly correlated with higher ammonia production. These
findings are in agreement with a study by Pop and Cherry (2000) which found pH and
ammonia formation were positively correlated. Unexpectedly, pH values of all heat treated
shavings decreased at 7 days of incubation, while the pH of air-dried and commercial
59

shavings increased. In this experiment, 72 hour exposure had the lowest pH and maintained a
lower pH compare to air-dried and commercial shavings over time. In comparison, perlite
litter produced extremely significant levels of ammonia after just 4 days. This can be
explained by fast microbial breakdown over a short period o f time, but the microbial
respiration rates and pH of the perlite litter did not reflect the extreme predicted values.
Although heating pine wood shavings to 200°C was as effective as heating to 150°C in
reducing Salmonella survival, in further experiments when the 200°C treated shavings were
challenged with a complex inoculum of spent litter and uric acid, it resulted in higher
microbial respiration compared with the I50°C treatments (Table 3.5). Also, the model litter
system made with 150°C heated wood maintained an acidic pH after 7 days while the pH
value of that at 200°C reached pH>7 which was associated with a higher microbial respiration
(Table 3.4). It was interesting to note that the pH of perlite was not significantly different
from the pH of the air-dried shavings, although microbial respiration of perlite was
significantly higher. This may be related to a lower microbial activity on air-dried shavings
compared to perlite, but as the pH values are the same it may also indicate that the activity of
the mineralizing bacteria is unaffected by substrate.
Overall, a heating regime of I20°C for 72 hours was found to be optimum for enhancing the
antimicrobial effect in pine wood shavings to inhibit Salmonella, as well as reducing
ammonia formation and the pH of the model litter. The heating technique was also effective
at significantly inhibiting Staphylococcus aureus and Escherichia coli, two other pathogens
known to be present in poultry litter (Table 3.9). In addition. Salmonella hsid lower survival
on filter paper compared with air-dried shavings, which may be due to a lack o f nutrition in
the filter paper substrate (Table 3.2).
The heating technique also significantly increased antimicrobial activity o f hardwood
shavings o f ash, beech, red oak as well as softwood species such as pine and cypress
(P<0.001) (Table 3.10). This highlights the potential of heat treatment to enhance the
antimicrobial properties of hardwoods and use these as substitutes when there is a shortage o f
the most desirable wood materials as bedding for poultry. Air-dried eucalyptus and cypress
had also an inhibitory effect on Salmonella recovery compared to the other tested wood
species.
It is also noteworthy that the pH of the wood shavings after 24 hours o f heat treatment was
found to be lowered from an average of 4.8 to 4.2. Although, pH values o f less than 5 are
6 0

known to be unfavourable for Salmonella, its survival in treated litter with extremely acidic
pH as low as 2.1 has been recorded (Williams, 2012). Thus, it seems unlikely that only lower
pH of heat treated wood completely inhibited Salmonella growth. However, low pH in
combination with low water activity can be effective at reducing Salmonella survival in the
litter. In a previous study, Payne et al. (2007) achieved a 5 log unit reduction o f Salmonella
at pH 4, although this was combined with maintaining water activity at less than 0.84. To
maintain a water activity o f 0.84, a moisture content of 25-30 % is required (Ye et a l, 2006).
In the present study, moisture content o f the various wood treatments was set at 60% of their
water holding capacity, which is equivalent to a water activity of 1 (Ye et a l, 2006),
indicating the inhibitory activity of heated wood materials against Salmonella is in addition to
the physical parameters. However, the lower pH o f heated wood can be considered as an
advantage over air-dried materials, as it reduces the need for application o f litter amendments
in controlling pH.
In the next chapter chemical characteristics of heated wood of pine which were effective at
inhibiting Salmonella and microbial activity in the modelled litter, by reducing ammonia and
pH, will be investigated to determine the bioactive compounds responsible for enhanced
antimicrobial activity.
61

CHAPTER 4: CHEMICAL ANALYSIS AND EVALUATION OF ENHANCED ANTIMICROBIAL
ACTIVITY OF HEAT-TREATED WOOD
4.1 Introduction
Antimicrobial compounds are essential tools to control microorganisms in the environment so
that the quality o f human life can be maintained and animal health conserved. Industrial
processes, pharmaceuticals, plant health services, animal health services, hospitals, clinics,
water treatment facilities and public access facilities benefit enormously by using a variety of
antimicrobial agents (American-Chemistry-Council, 2010). Antimicrobials can have a
variety o f applications. They are used as medicines to cure or prevent human diseases, to
inhibit the spread of disease through the use of disinfectants and to protect manufactured
products and industrial equipment and systems from deterioration (American-Chemistry-
Council, 2010). They are also used in food and beverages in the form of food additives and
preservatives, to protect and treat natural resources, water supplies and forests through their
used as pesticides. Antimicrobials are of great importance for treatments o f infectious
diseases ever since they have been discovered. However, in the last decade antimicrobial
resistance has become a serious public health concern because o f the social and economic
implications. Increased mortality rates particularly in infants and the elderly, increase the
cost of treatments and prolong hospital stays and so are major impacts. (Levy, 2001; Martin
et a l, 2010; Shea, 2003)
Overuse of antimicrobials has resulted in the development of resistance in a number of
environments including clinical facilities, farms and hospitals (Martin et a l, 2010). Using
antimicrobials as growth promoters or for therapeutic reasons in poultry, animal husbandry
and agriculture is also known to contribute towards antimicrobial resistance in humans. The
residues o f antibiotics in meat and animal products may result in humans being exposed to
antimicrobials unnecessarily and this may result in resistance in the natural flora in humans
(Kapil, 2005). The use of antimicrobials shifts natural selection towards resistant bacteria in
the intestines of animals intended for consumption. Resistant non-pathogenic bacteria on-
farm can also act as a pool of transferrable genes (DeFrancesco et a l, 2004; Kelley et a l,
1998). Resistant strains of pathogens causing food-borne diseases such as Salmonella are
increasing (Angulo et a l, 2004). Through the increased incidence of resistant pathogens and
the rapid development of the processing industry, there is now an urgent need for new
sources of antimicrobial compounds.
6 2

Many of the well-known antimicrobials were originally derived from plants, bacteria and
fungi. A large numbers o f bioactive chemicals derived from different plant species have been
identified and used to control microorganisms (Davidson et a l, 2005b). Plant materials are
the most important source of natural antimicrobials due to their chemical diversity and mass
availability (Cowan, 1999). Many research studies focus on exploring the potential of new
species o f plants whilst, others review the general techniques and procedures used to discover
novel bioactive substances and their evaluations (MacKay et a l, 2009a). Fermentation,
extraction and torrefacation of wood materials are examples o f developed techniques applied
to produce different chemicals in large quantities, such as solvents and resins. Valuable
chemicals derived from wood by various processes including heating also provide great
advantages to human life, animal health and economic welfare (MacKay et a l, 2009a;
Murzin et a l, 2007). Many of these chemicals are manufactured and used as antimicrobial
agents, food preservatives or intermediate chemicals in pharmaceuticals (Murzin et a l,
2007). Because they are produced as a result o f chemical modification o f wood structural
components, desirable chemicals are produced under the same heating regime regardless of
the chemical differences between wood species. These chemicals are then extracted using
organic solvents and identified using analytical techniques including Nuclear Magnetic
Resonance spectroscopy (NMR) and gas chromatography-mass spectrometry (GC-MS) for
manufacturing.
Although, enormous numbers o f chemicals are derived from wood processing techniques,
research and development around the heating techniques has been focused on improving the
life duration of wood and its resistance toward wood decay fungi (Kamdem et a l, 2002;
Savluchinske-Feio et a l, 2006; Voda et a l, 2003). Therefore, most o f the existing
developments are based on need in the market for commercially profitable products and their
application in sectors such as furniture manufacture and construction. Little has been done in
examining or developing techniques that introduce valuable levels of antimicrobial
compounds which have the potential to inhibit food-borne pathogens. However, the heating
technique developed and applied to pine wood shavings described in Chapter 3, showed
promising results in inhibiting SE when in direct contact. The aim of this chapter was
therefore to investigate chemical properties of heated wood using chemical analysis
techniques.
63

4.2 Experimental
The data presented in Chapter 3 revealed that heat-treated wood shavings of pine exhibited
strong antimicrobial activity compared with air-dried shavings. In order to be able to isolate
and identify the chemicals responsible for the activity, pine wood shavings were heat treated
with the thermal regime developed in Chapter 3 and extracted with cold water, hot water and
methanol. The crude extracts were impregnated into shredded filter paper and inoculated
with SE as described below (4.2.1). After incubation, the survival rate of Salmonella was
measured by the enumeration of colony forming units recovered from one gram filter paper
after incubation at 25°C. The same procedure was applied to air-dried pine shavings for
comparison. To evaluate the change in antimicrobial activity o f heated wood after extraction,
the three types o f extracted wood shavings (cold water, hot water and methanol) were
allowed to dry for 3 days and then inoculated with Salmonella as described below (4.2.1).
After incubation, one gram of each type of extracted wood shavings was tested for the survival
of Salmonella (cfu g'^).
4.2.1 Extract assays
Water and methanol crude extracts of pine wood shaving were prepared as described in 2.5.2
and 2.5.3. For each extract, 1 ml was added onto shredded (autoclaved) filter paper in
McCartney bottles (12 samples, Igram each). Samples were left overnight to absorb the
extracts. Control treatments received 1 ml of Ringer’s solution strength). A suspension of
SB (10 ^cfti ml'^) in Ringer’s Solution was made to inoculate the bottles. Each sample bottle
was inoculated with 1 ml of the suspension and then incubated overnight at 25°C.
Subsequently, a dilution series (10'^ to 10'^) for each sample was plated onto XLD agar to
assess bacterial survival. The plates were incubated at 25 °C and the colony count was used
to compare inhibition against control treatments and extracts from air-dried shavings.
In a second experiment, the antimicrobial activity of heat treated wood shavings after
extraction was tested to prove that extraction removed the chemicals responsible for the
antimicrobial activity. In this experiment the extracted wood samples were left to dry at 50°C
for 3 days. Two grams of each extracted sample was then transferred to a McCartney bottle
and inoculated with a 2 ml suspension of Salmonella. Two grams shredded filter paper was
used as the control. All samples were incubated at 25°C for 24 hours after which time a
dilution series (10'^ to 10'^) was prepared and plated on XLD agar. Plates were incubated
over night at 25°C for bacterial enumeration.
64
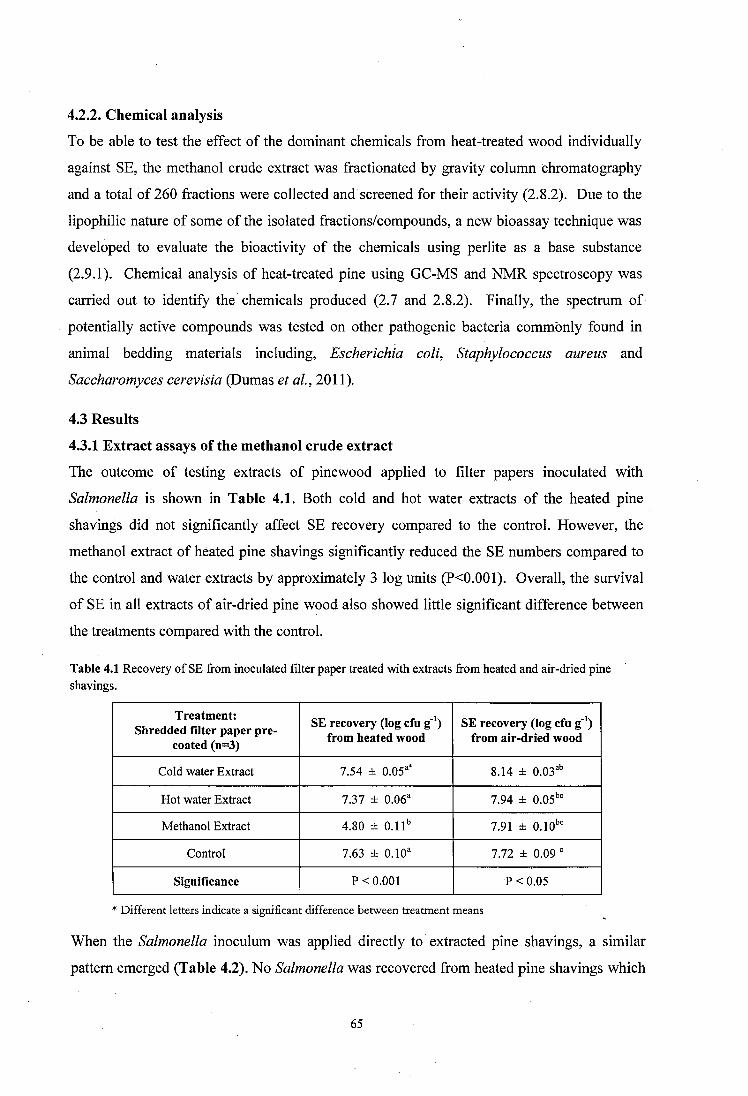
4.2.2. Chemical analysis
To be able to test the effect of the dominant chemicals from heat-treated wood individually
against SE, the methanol crude extract was fractionated by gravity column chromatography
and a total of 260 fractions were collected and screened for their activity (2.8.2). Due to the
lipophilic nature of some of the isolated fractions/compounds, a new bioassay technique was
developed to evaluate the bioactivity o f the chemicals using perlite as a base substance
(2.9.1). Chemical analysis of heat-treated pine using GC-MS and NMR spectroscopy was
carried out to identify the chemicals produced (2.7 and 2.8.2). Finally, the spectrum of
potentially active compounds was tested on other pathogenic bacteria commonly found in
animal bedding materials including, Escherichia coli. Staphylococcus aureus and
Saccharomyces cerevisia (Dumas et al., 2011).
4.3 Results
4.3.1 Extract assays of the methanol crude extract
The outcome of testing extracts o f pinewood applied to filter papers inoculated with
Salmonella is shown in Table 4.1. Both cold and hot water extracts of the heated pine
shavings did not significantly affect SE recovery compared to the control. However, the
methanol extract of heated pine shavings significantly reduced the SE numbers compared to
the control and water extracts by approximately 3 log units (P<0.001). Overall, the survival
of SE in all extracts o f air-dried pine wood also showed little significant difference between
the treatments compared with the control.
T able 4.1 Recovery o f SE from inoculated filter paper treated with extracts from heated and air-dried pine shavings.
Treatm ent: Shredded filter paper pre
coated (n=3)
SE recovery (log cfu g'^) from heated w ood
SE recovery (log cfu g’ )̂ from air-dried w ood
Cold water Extract 7.54 ± 0.05=* 8.14 ± 0.03=’’
Hot water Extract 7.37 ± 0.06= 7.94 ± 0.05*’°
Methanol Extract 4.80 ± O .lfr 7.91 ± 0.1 o'’"
Control 7.63 ± 0.10= 7.72 ± 0.09 "
Significance P < 0.001 P < 0.05
* D ifferent letters indicate a significant difference between treatm ent means
When the Salmonella inoculum was applied directly to extracted pine shavings, a similar
pattern emerged (Table 4.2). No Salmonella was recovered from heated pine shavings which
65
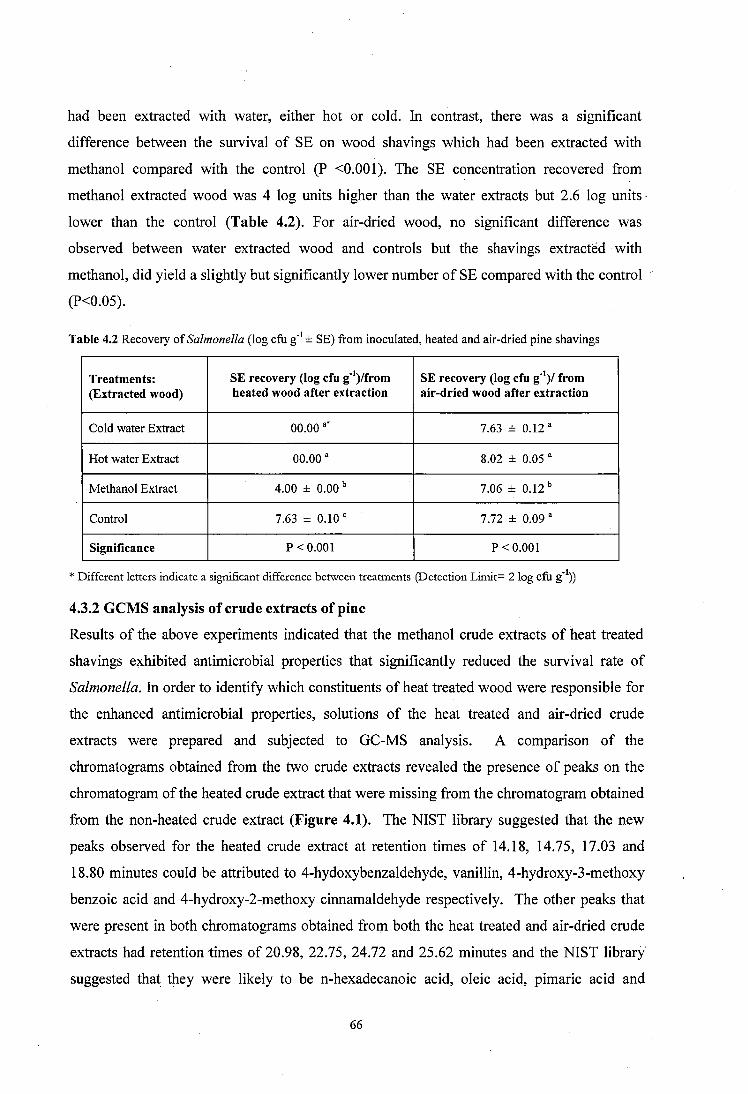
had been extracted with water, either hot or cold. In contrast, there was a significant
difference between the survival of SE on wood shavings which had been extracted with
methanol compared with the control (P <0.001). The SE concentration recovered from
methanol extracted wood was 4 log units higher than the water extracts but 2.6 log units
lower than the control (Table 4.2). For air-dried wood, no significant difference was
observed between water extracted wood and controls but the shavings extracted with
methanol, did yield a slightly but significantly lower number of SE compared with the control
(P<0.05).
Table 4.2 Recovery o f Salm onella (log cfu g'' ± SE) from inoculated, heated and air-dried pine shavings
T reatm ents: (Extracted w ood)
SE recovery (log cfu g’^)/from heated w ood after extraction
SE recovery (log cfu g^)t from air-dried w ood after extraction
Cold water Extract 00.00 =* 7.63 ± 0.12 =
Hot water Extract 00.00 = 8.02 ± 0.05 =
Methanol Extract 4.00 ± 0.00 ’’ 7.06 ± 0.12
Control 7.63 ± 0.10 " 7.72 ± 0.09 =
Significance P < 0.001 P < 0.001
* D ifferent letters indicate a significant difference between treatm ents (Detection Lim it= 2 log cfu g’ )̂)
4.3.2 GCMS analysis of crude extracts of pine
Results of the above experiments indicated that the methanol crude extracts o f heat treated
shavings exhibited antimicrobial properties that significantly reduced the survival rate of
Salmonella. In order to identify which constituents of heat treated wood were responsible for
the enhanced antimicrobial properties, solutions o f the heat treated and air-dried crude
extracts were prepared and subjected to GC-MS analysis. A comparison o f the
chromatograms obtained from the two crude extracts revealed the presence o f peaks on the
chromatogram of the heated crude extract that were missing from the chromatogram obtained
from the non-heated crude extract (Figure 4.1). The NIST library suggested that the new
peaks observed for the heated crude extract at retention times of 14.18, 14.75, 17.03 and
18.80 minutes could be attributed to 4-hydoxybenzaldehyde, vanillin, 4-hydroxy-3-methoxy
benzoic acid and 4-hydroxy-2-methoxy cinnamaldéhyde respectively. The other peaks that
were present in both chromatograms obtained from both the heat treated and air-dried crude
extracts had retention times of 20.98, 22.75, 24.72 and 25.62 minutes and the NIST library
suggested that they were likely to be n-hexadecanoic acid, oleic acid, pimaric acid and
66
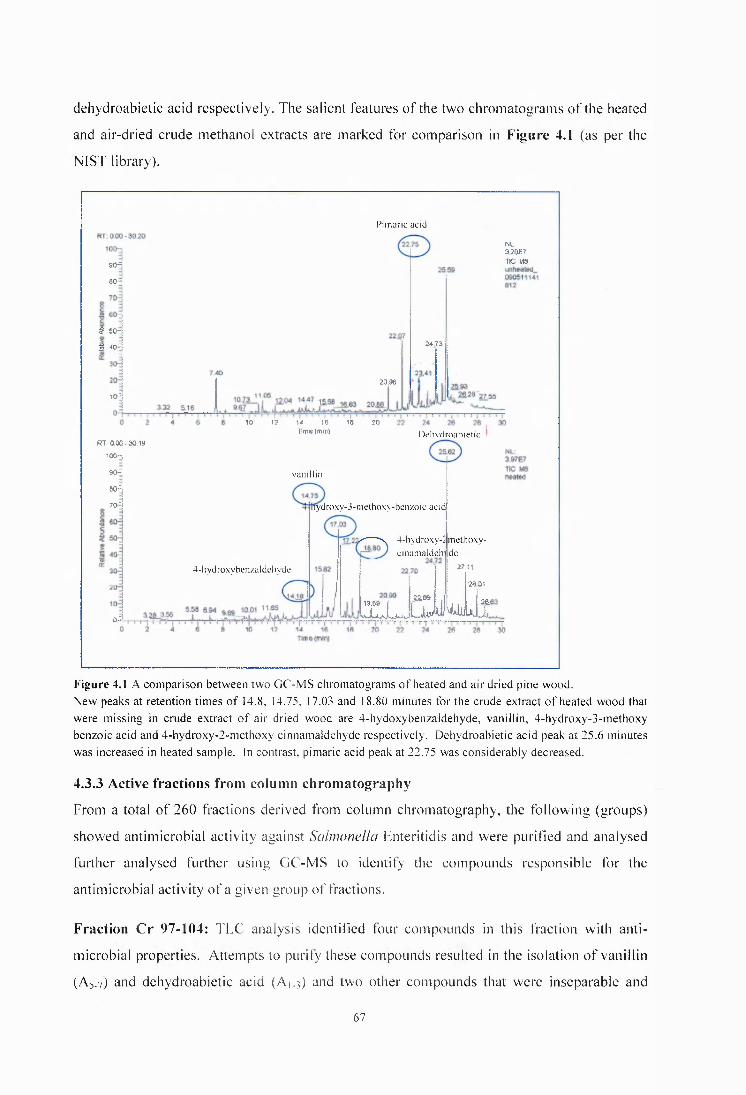
dehydroabietic acid respectively. The salient features o f the two chromatograms of the heated
and air-dried crude methanol extracts are marked for comparison in Figure 4.1 (as per the
NIST library).
Piinanc acid
NL:3 20E7 TC ke90-=
aoi
@ so-2473S 4CK
20.96
10 1? 1< 10 18 20 Time (fn in)
10=
nchvdroabieticRT: 0.00 . 30.19
lOOn
90-; vanillin
80=70= iydroxy-3-melhoxy-benzoic acid
z' \ 4-hydroxy-] methoxy-cinamaldeh de
y - 2801ziJdüLih
4-hydroxybenzaldeh\de
D=
Figure 4.1 A com parison between tw o G C-M S chrom atogram s o f heated and air dried pine wood.N ew peaks at retention tim es o f 14.8, 14.75, 17.03 and 18.80 m inutes for the crude extract o f heated w ood that were missing in crude extract o f air dried wood are 4-hydoxybenzaldehyde, vanillin, 4-hydroxy-3-m ethoxy benzoic acid and 4-hydroxy-2-m ethoxy cinnam aldéhyde respectively. Dehydroabietic acid peak at 25.6 m inutes was increased in heated sam ple. In contrast, pimaric acid peak at 22.75 was considerably decreased.
4.3.3 Active fractions from column chrom atography
From a total o f 260 fractions derived from column chromatography, the following (groups)
showed antimicrobial activity against Salmonella Enteritidis and were purified and analysed
further analysed further using GC-MS to identify the compounds responsible for the
antimicrobial activity o f a given group of fractions.
Fraction C r 97-104: TEC analysis identified four compounds in this fraction with anti
microbial properties. Attempts to purify these compounds resulted in the isolation o f vanillin
(A5 .7 ) and dehydroabietic acid (A 1.3 ) and two other compounds that were inseparable and
67

could therefore not be identified.
Fraction 2Cr 105-109; Purification of this fraction resulted in the isolation o f dehydroabietic
acid, picealactone-A (A4 ) and two other compounds that were inseparable.
Fraction 2Cr 110-115: This fraction yielded dehydroabietic acid, 3, 3’-dimethoxy-4, 4 ’-
dihydroxy stilbene (A 14.16) and also picealactone A (A4 ).
Fraction 2Cr 116-121: This fraction yielded dehydroabietic acid, 3, 3’-dimethoxy-4, 4 ’-
dihydroxy stilbene and acetovanillone (A n.12).
Fraction 2Cr 122-133: This fraction yielded dehydroabietic acid and acetovanillone.
Fraction 2Cr 138-139, Cr 140-160: This fraction yielded dehydroabietic acid and
coniferaldehyde (A 17.19). Dehydroabietic acid (DHAA) and vanillin were found to
prédominante in the methanol extract, but it was not possible to completely purify DHAA
and vanillin.
4.3.4 Yield of different molecules of interest from heated Scots pine wood shavings
As shown in Table 4.3, the most abundant compound isolated from heated wood shavings
was vanillin followed by dehydroabietic acid (0.26% and 0 .2 %) respectively.
Coniferaldehyde (30mg), stilbene (25 mg), acetovanillone (15 mg) and vanillic acid (As-io)
(15 mg) were the second most abundant compounds respectively. Guaiacol (A b) and
picealactone A were isolated in minor quantities (0.02% and 0.03% )). The chemical structure of
these compounds is shown in Figure 4.2.
Table 4.3 Y ield o f different compounds isolated from heated wood o f P. sylvestris.
C om pound Solvent system M ass obtained(30 g)
Acetovanillone 30% Hexane/ 70% DCM 15 mg (0.05% )
Coniferaldehyde 30% Hexane/ 70% DCM 30 mg (0.1%)
Dehydroabietic acid 30% Hexane/ 70% DCM 60 mg (0.2%)
Guaiacol 100% Hexane 6 mg (0.02% )
Picealactone A 30% Hexane/ 70% DCM 9 mg (0.03% )
Stilbene 30% Hexane/ 70% DCM 25 mg (0.08%)
Vanillic acid 30% Hexane/ 70% DCM 15 mg (0.05%)
Vanillin 30% Hexane/ 70% DCM 80 mg (0.26% )
68
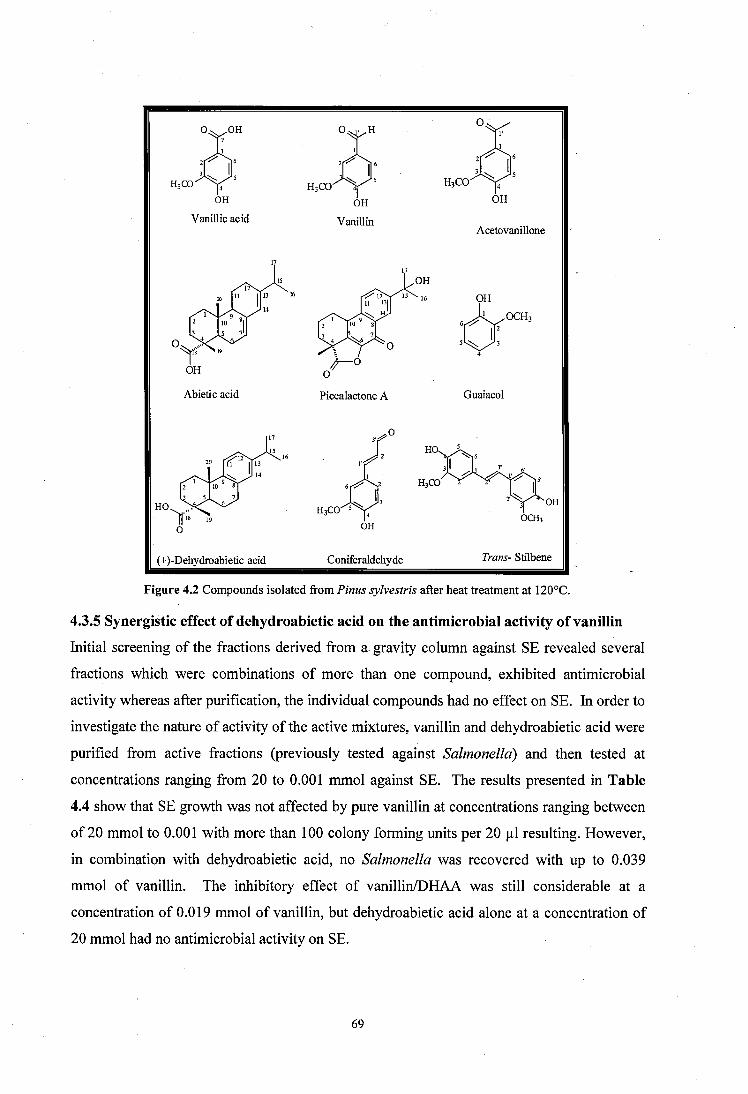
.OH
OHOH
Vanillic acidOH
VanillinAcetovanillone
OH
OH
OH
Abietic acid GuaiacolPicealactone A
HO,
OHHO.
OH
Trans- StilbeneConiferaldehyde(+)-Dehydroabietic acid
Figure 4.2 Compounds isolated from Pinus sylvestris after heat treatment at 120°C.
4.3.5 Synergistic effect of dehydroabietic acid on the antim icrobial activity of vanillin
Initial screening of the fractions derived from a gravity column against SE revealed several
fractions which were combinations of more than one compound, exhibited antimicrobial
activity whereas after purification, the individual compounds had no effect on SE. In order to
investigate the nature of activity of the active mixtures, vanillin and dehydroabietic acid were
purified from active fractions (previously tested against Salmonella) and then tested at
concentrations ranging from 20 to 0.001 mmol against SE. The results presented in Table
4.4 show that SE growth was not affected by pure vanillin at concentrations ranging between
of 20 mmol to 0.001 with more than 100 colony forming units per 20 pi resulting. However,
in combination with dehydroabietic acid, no Salmonella was recovered with up to 0.039
mmol of vanillin. The inhibitory effect o f vanillin/DHAA was still considerable at a
concentration of 0.019 mmol o f vanillin, but dehydroabietic acid alone at a concentration of
20 mmol had no antimicrobial activity on SE.
69
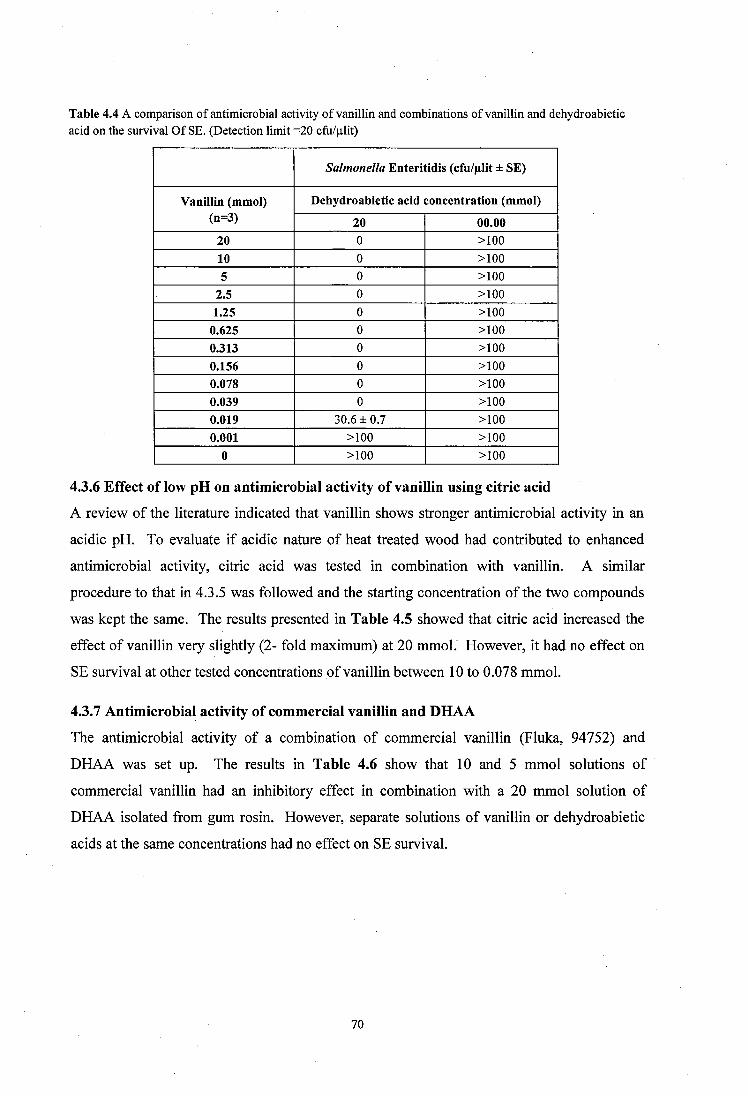
Table 4.4 A comparison o f antimicrobial activity o f vanillin and combinations o f vanillin and dehydroabieticacid on the survival O f SE. (Detection limit =20 cfli/plit)
Salm onella Enteritidis (cfn/plit ± SE)
V anillin (m m ol) (n=3)
D ehydroabietic acid concentration (m m ol)
20 00.0020 0 >10010 0 >1005 0 >100
2.5 0 >1001.25 0 >100
0.625 0 >1000.313 0 >1000.156 0 >1000.078 0 >1000.039 0 >1000.019 30.6 ± 0 .7 >1000.001 >100 >100
0 >100 >100
4.3.6 Effect of low pH on antimicrobial activity of vanillin using citric acid
A review of the literature indicated that vanillin shows stronger antimicrobial activity in an
acidic pH. To evaluate if acidic nature of heat treated wood had contributed to enhanced
antimicrobial activity, citric acid was tested in combination with vanillin. A similar
procedure to that in 4.3.5 was followed and the starting concentration o f the two compounds
was kept the same. The results presented in Table 4.5 showed that citric acid increased the
effect of vanillin very slightly (2- fold maximum) at 20 mmol. However, it had no effect on
SE survival at other tested concentrations of vanillin between 10 to 0.078 mmol.
4.3.7 Antimicrobial activity of commercial vanillin and DHAA
The antimicrobial activity of a combination of commercial vanillin (Fluka, 94752) and
DHAA was set up. The results in Table 4.6 show that 10 and 5 mmol solutions of
commercial vanillin had an inhibitory effect in combination with a 2 0 mmol solution of
DHAA isolated from gum rosin. However, separate solutions of vanillin or dehydroabietic
acids at the same concentrations had no effect on SE survival.
70
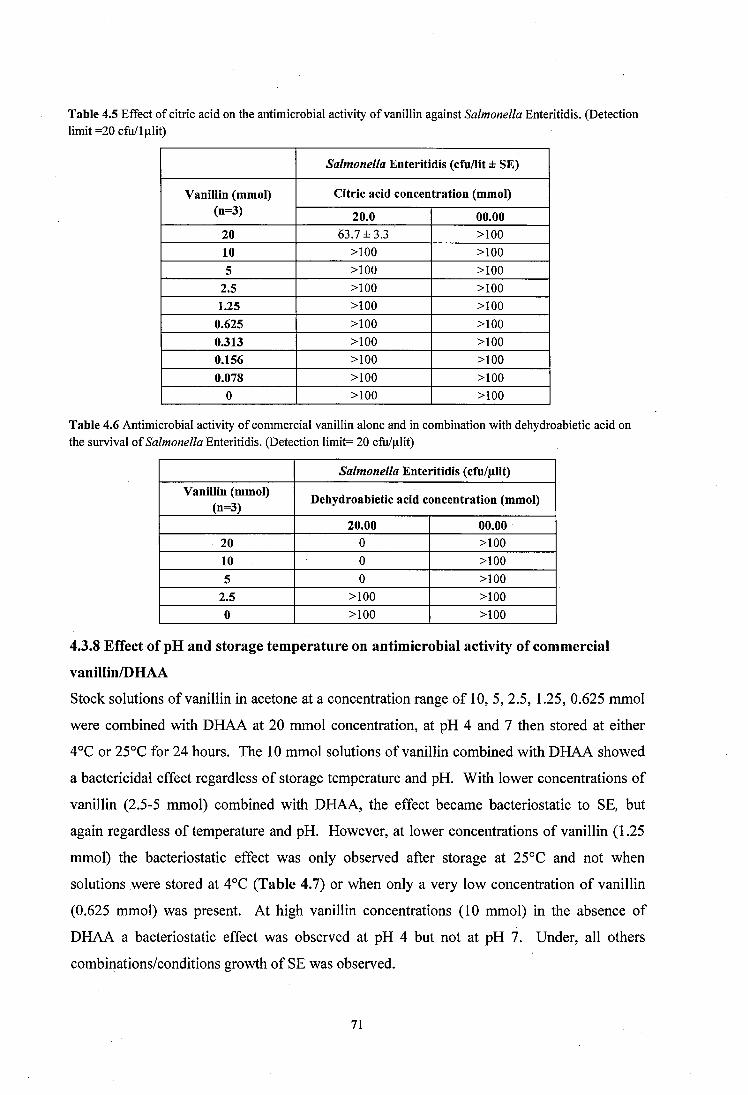
Table 4.5 Effect o f citric acid on the antimicrobial activity o f vanillin against Salmonella Enteritidis. (Detectionlimit =20 cfu /lp lit)
Salm onella Enteritidis (cfu/Iit ± SE)
V anillin (m m ol) (n=3)
C itric acid concentration (m m ol)
20.0 00.0020 63.7 ± 3 .3 >10010 >100 >1005 >100 >100
2.5 >100 >1001.25 >100 >100
0.625 >100 >1000.313 >100 >1000.156 >100 >1000.078 >100 >100
0 >100 >100
Table 4.6 Antimicrobial activity o f commercial vanillin alone and in combination with dehydroabietic acid on the survival o f Salm onella Enteritidis. (Detection limit= 20 cfli/plit)
Salm onella Enteritidis (cfu/plit)
V anillin (m m ol) (n=3)
D ehydroabietic acid concentration (m m ol)
20.00 00.0020 0 >10010 0 >1005 0 >100
2.5 >100 >1000 >100 >100
4.3.8 Effect of pH and storage tem perature on antim icrobial activity of commercial
vanillin/DHAA
Stock solutions of vanillin in acetone at a concentration range o f 10, 5, 2.5, 1.25, 0.625 mmol
were combined with DHAA at 20 mmol concentration, at pH 4 and 7 then stored at either
4°C or 25°C for 24 hours. The 10 mmol solutions of vanillin combined with DHAA showed
a bactericidal effect regardless of storage temperature and pH. With lower concentrations of
vanillin (2.5-5 mmol) combined with DHAA, the effect became bacteriostatic to SE, but
again regardless of temperature and pH. However, at lower concentrations of vanillin (1.25
mmol) the bacteriostatic effect was only observed after storage at 25°C and not when
solutions were stored at 4°C (Table 4.7) or when only a very low concentration o f vanillin
(0.625 mmol) was present. At high vanillin concentrations (10 mmol) in the absence o f
DHAA a bacteriostatic effect was observed at pH 4 but not at pH 7. Under, all others
combinations/conditions growth o f SE was observed.
71
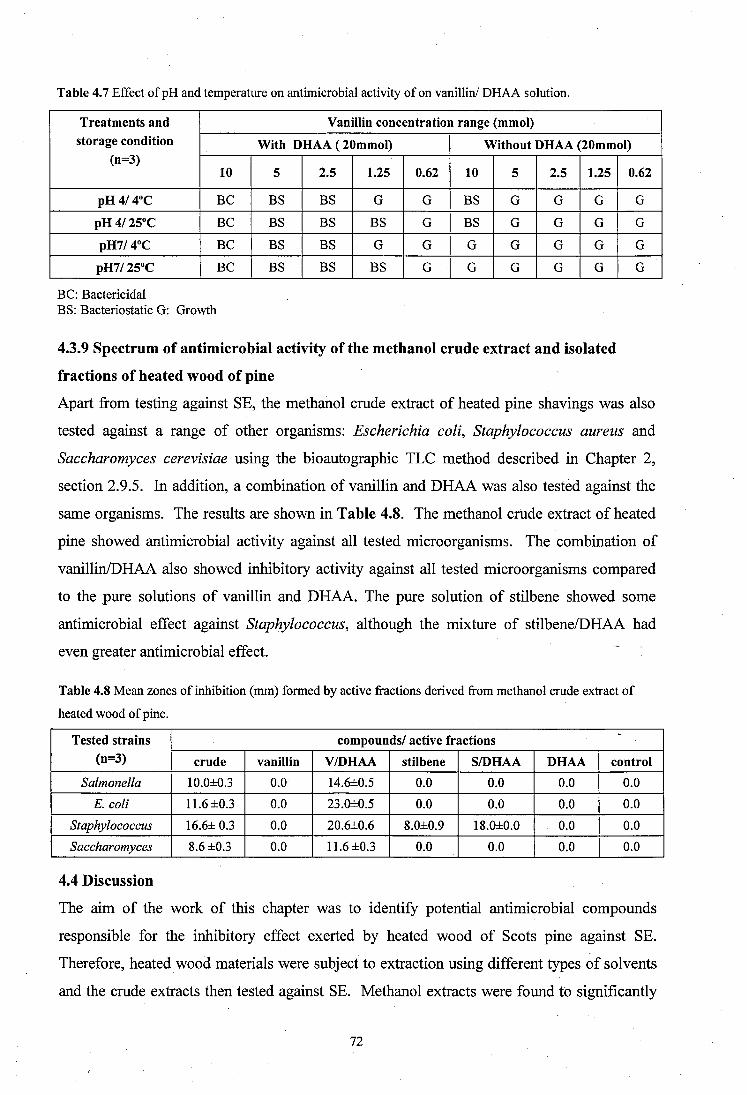
T ab le 4 .7 E ffect o f pH and temperature on antimicrobial activity o f on vanillin/ D H A A solution.
T reatm en ts and
storage cond ition
(n=3)
V an illin con cen tration ran ge (m m ol)
W ith D H A A ( 20m m ol) W ith ou t D H A A (20m m ol)
10 5 2 .5 1.25 0.62 10 5 2 .5 1.25 0.62
pH 4 / 4“C BC B S B S G G B S G G G G
p H 4 / 2 5 ”C BC B S B S B S G B S G G G G
p H 7/ 4“C BC B S B S G G G G G G G
p H 7/25" C BC B S B S B S G G G G G G
BC: BactericidalBS: Bacteriostatic G: Growth
4.3.9 Spectrum of antimicrobial activity of the methanol crude extract and isolated
fractions of heated wood of pine
Apart from testing against SE, the methanol crude extract o f heated pine shavings was also
tested against a range o f other organisms: Escherichia coli. Staphylococcus aureus and
Saccharomyces cerevisiae using the bioautographic TEC method described in Chapter 2,
section 2.9.5. In addition, a combination o f vanillin and DHAA was also tested against the
same organisms. The results are shown in Table 4.8. The methanol crude extract o f heated
pine showed antimicrobial activity against all tested microorganisms. The combination o f
vanillin/DHAA also showed inhibitory activity against all tested microorganisms compared
to the pure solutions o f vanillin and DHAA. The pure solution o f stilbene showed some
antimicrobial effect against Staphylococcus, although the mixture o f stilbene/DHAA had
even greater antimicrobial effect.
T able 4 .8 M ean zones o f inhibition (m m ) form ed by active fractions derived from m ethanol crude extract o f
heated w ood o f pine.
T ested stra ins com p ou n d s/ active fraction s(n=3) crud e van illin V /D H A A stilbene S /D H A A D H A A con tro l
Salm onella 10.0±0.3 0.0 14.6±0.5 0.0 0.0 0.0 0.0
E. co li 11.6 ±0.3 0.0 23 .0± 0 .5 0.0 0.0 0.0 0.0
Staphylococcus 16.6± 0.3 0.0 20 .6± 0 .6 8 .0±0.9 18.0±0.0 0.0 0.0
Saccharom yces 8.6 ±0.3 0.0 11.6 ±0.3 0.0 0.0 0.0 0.0
4.4 Discussion
The aim o f the work o f this chapter was to identify potential antimicrobial compounds
responsible for the inhibitory effect exerted by heated wood o f Scots pine against SE.
Therefore, heated wood materials were subject to extraction using different types o f solvents
and the crude extracts then tested against SE. Methanol extracts were found to significantly
72

reduce the number o f Salmonella cfu whereas cold and hot water extracts did not. However,
the inhibitory effect o f the methanol extract was only associated with the heat-treated pine
whilst the methanol extract o f air-dried wood had no inhibitory effect on SE numbers. These
results were consistent with the data obtained from a parallel experiment which measured the
recovery o f Salmonella on wood materials which were already water or methanol extracted.
They indicate that that methanol successfully extracts the active compound or compounds
which give heat-treated wood o f pine its antimicrobial activity when in direct contact with
SE.
GC-MS analysis revealed several differences between the chemical composition o f air-dried
and heat-treated pine wood. Heat-treated wood contained at least four new compounds:
vanillin, benzaldehyde, coniferaldehyde and benzoic acid and GC-MS analysis revealed that
vanillin was present in the highest concentration. Preliminary bioassay tests showed fractions
containing vanillin completely inhibited Salmonella recovery from m oist ground perlite.
Vanillin is known to exhibit antimicrobial activity against food-borne bacteria, moulds and
yeasts so it is used as a food preservative in fhiit juice and fruit puree. However, purification
o f the active fractions resulted in a reduction o f this activity. N M R analysis o f the active
fractions revealed that another compound was present in small quantity which was identified
as dehydroabietic acid (DHAA). Further, purification o f the compounds and their individual
bioassays showed the minimum inhibitory concentration o f purified vanillin was 2 0 mmol
when tested against SE and DHAA had no negative effect on SE survival (T able 4.4).
However, when DHAA was tested in combination with vanillin at different concentrations,
the MIC o f vanillin against SE decreased from 20 mmol to 0.019 mmol (Table 4.4).
Previous researches have also reported that DHAA has no antimicrobial activity on Gram
negative bacteria (Savluchinske-Feio et ah, 2006). Also, the minimum inhibitory
concentration o f vanillin against Gram negative bacteria including E. coli and Salmonella
Enteritidis is reported to be over 10 mmol and between 10 to 21 mmol against yeasts
(Fitzgerald et a l , 2004; Vasantha Rupasinghe et a l, 2006). However, the results o f this
study showed a minimum concentration o f 20 mmol o f DHAA in mixture with vanillin at a
concentration range o f between 20 to 0.019 mmol was essential but adequate to inhibit SE
completely. It is known that vanillin shows greater antimicrobial and antioxidant properties
when the pH is acidic (Mourtzinos et a l, 2009). Oxidization o f phenolic compounds during
lignin breakdown increases acidic content o f heated wood. To test the hypothesis that acidic
conditions might enhance the activity o f vanillin, DHAA was replaced with citric acid, but it
73

only increased the effect o f vanillin 2 fold (10 mmol) (Table 4.5). Therefore, the retained
activity of vanillin at low concentrations o f 0.019 mmol in mixture with DHAA may be
explained by the synergistic effect o f the mixture. The synergism between mixtures of
several phenols or abietane type resin acids has been reported previously (Cowan, 1999;
Mourtzinos et a l, 2009; Savluchinske-Feio et a l, 2007). However, the results of the present
study provide the first report o f synergism between an aromatic aldehyde and a diterpenoid
resin acid.
In testing the commercial forms o f aldehydes and resin acids, and finding that the
antimicrobial activity o f their mixture was weakened, it could be argued that other
requirements need to be met for a strong antimicrobial activity to be induced in the
commercial mixtures similar to that obtained in the bioassay o f original fractions. Two
factors are likely to have influenced the magnitude of activity in the original fractions based
on the environment of heat-treated wood and to a similar extent their methanol extracts.
These are (1) low pH caused by the presence o f a high content of organic acids; (2)
temperature effects on the miscibility or solubility o f the compounds. Therefore, mixtures of
the commercial forms were made prior to testing and the effect o f temperature and pH tested.
When vanillin was tested alone, at the same concentration the nature o f activity was only
bacteriostatic at pH 4 and showed no inhibitory effect at pH 7 (Table 4.7). The results also
showed the level o f inhibitory activity o f the mixture was dependent on the vanillin
concentration but it increased only 2 -fold which was less than half o f the antimicrobial
activity of the mixture derived from the methanol crude extract (Table 4.7). In addition, the
nature o f inhibitory activity of the mixture was also dependent on vanillin concentration.
When vanillin was at 10 mmol, the activity o f the mixture was bactericidal at both tested pH
and temperature. Fitzgerald et al (2004a) also reported that the bacteriostatic effect of
vanillin varied depending on the vanillin concentration and type of microorganisms. It is
well accepted that phenolic antimicrobials lose their activity with dilution (Russell, 2003).
However, the mixture derived from heated wood had a bactericidal effect even at the lowest
tested concentration of vanillin (0.019 mmol). This may indicate that the antimicrobial
activity of the mixture is in addition to pH or temperature effects and requires further
investigation. The results of a previous study on resin acid solubility in pulp mill suggest that
these chemicals form association with other organic molecules under acidic condition
(Werker and Hall, 1997). Therefore, the possibility o f such reactions and their significance
on the level of enhanced antimicrobial activity suggests further research in the future.
74

Mixtures of the two compounds originating from the crude extract showed an inhibitory
effect not only against SE but also against Escherichia, Staphylococcus and Saccharomyces.
Based on the mean zone of inhibition, the activity of vanillin/DHAA was greater than the
crude extract on each tested species. Maximum inhibition occurred with E. coli and the
minimum effect with Saccharomyces. Both vanillin and DHAA had no activity on any o f the
other tested species when tested alone. However, others have reported antimicrobial activity
of DHAA on Gram positive bacteria including S. aureus (Savluchinske-Feio et al., 1999).
However, when stilbene, another compound derived from crude extract, was tested in
combination with DHAA, it had an inhibitory effect but only on Staphylococcus. Also,
unlike vanillin, purified stilbene alone did have an inhibitory effect on Staphylococcus but to
a lesser extent than the stilbene/DHAA mixture. The antibacterial effect of stilbene on
Staphylococcus aureus has been reported elsewhere (Eloff et a l, 2005), and recently, the
synergistic effect o f stilbene in combination with antibiotics on pathogenic bacteria was
reported for the first time (Kumar et a l, 2012). This highlights the need for further
investigation on the mechanism by which the mixture o f DHAA with other compounds
results in a greater level of antimicrobial activity against different microorganism.
75

C H A P T E R 5: E F F E C T O F H E A T E D W O O D O N P. R A M O R U M
5.1 Reasons for concern
Identifying the outbreaks of the disease on Japanese larch in the UK increased concerns that
P. ramorum had the potential to severely affect biodiversity and the shape of the national
landscape throughout the UK. According to Brasier and Webber (2010) this was the first
time that the pathogen had established itself on a commercial plantation tree species,
infecting more than 600,000 mature larch trees (Figure 5.1). Currently, the Forestry
Commission and the private sector are felling all infected larches (Figure 5.2) to reduce the
risk of further damage to uninfected areas. The damage caused to larch has had a significant
economic effect on the value of timber in the UK. It also has raised the possibility that P.
ramorum could become a serious threat to European larch across other countries in Europe.
In 2011, a scientific opinion published by Plant Health Division of the European Food Safety
Authority (EFSA) stated “there are large regions across Europe that are climatically suitable
for spread of Phytopthora ramorum where susceptible hosts are also present” (EFSAP-PLH,
2011).
M '
iFigure 5.1 Dead larch trees infected by Phytophthora ramorum (Source:http://w w w .forestry.gov.uk/pdf/Phytophthoraram orum larchjune2011 .pdf)
5.2 Alternative approaches to control
Since the disease was discovered, researchers have developed control methods to limit the
impact of the disease to the nursery trade and to control its impact on woodland. Measures
include chemical treatment of plant stocks showing limited infection, irrigation water
disinfestation and soil disinfestation using chemical compounds. Such management
strategies have been successful in preventing significant damage to nursery plants. Despite
76

this, none of these treatments are guaranteed to eliminate the disease. Moreover, these
treatments are costly to the nursery trade due to the need to destroy infected stock (Frankel,
2008). Effective existing strategies are even more limited for infestations in the wider
environment. In heavily contaminated areas in the UK, trees are cut and logs transported to
licensed mills to be debarked and processed. Where clear cutting is not an option, removing
foliar hosts is carried out to reduce the spread of the disease to other trees. As the complete
clearance of foliar hosts in the wider environment is not economically viable, alternative
strategies are needed to limit inoculum levels and reduce the movement of inoculum from
contaminated litter to standing tree species.
Figure 5.2 Larch trees infected by Phytophthora ramorum, C rym ant, near N eath (Source: w w w .telegraph.co.uk/earth/countrvside/8298946)
One alternative strategy, covering the litter using a layer of woodchips has been practiced,
acting as a physical barrier to reduce the mobility of inoculum (Goheen et al., 2012).
However, using woodchips with antimicrobial activity potentially has more benefits than use
only as a physical barrier. Research on the chemical characteristics of woodchips used from
different tree species indicates the significance that different species may have on survival of
P. ramorum. For example, the survival of P. ramorum was investigated on woodchips of
rhododendron under natural conditions in a private garden in the Netherlands where the
disease was already presented (Boogert and Kessel, 2004). The study was undertaken in 16
plots over a period of 18 months. For each plot, 300 grams of woodchips were layered over
lm “ areas, 1 month prior to sampling in June 2004 when samples were taken to be tested for
77

presence of P. ramorum. Positive results were obtained throughout the study at each time of
sampling (variation between 1 to 10 samples). In contrast, woodchips from several western
conifer species and the identified compounds in the woodchips were found to have strong
antimicrobial activity toward P. ramorum (Manter et a l, 2006). The author suggested the
benefit of using antimicrobial woodchips as a part of an integrated programme to minimize
spore distribution in the area with higher human activities (Davidson et a l, 2005a; Tjosvold
et a l, 2 0 0 2 b).
Heating o f wood has been shown to be an effective method to improve its resistance against
fungal attacks. Heating to a temperature of 200°C, leads to wood becoming resistant against
blue stain, white rot and brown rot fungi (Kamdem et a l, 2002; Savluchinske-Feio et a l,
2007). Depending on the fungus and wood species, decaying o f wood can be described as the
depolymerisation or modification of the main wood components resulting in the release of
energy and metabolites for microbial growth. In the first instance, biological resistance of
heated wood can be explained by the changes caused by heating which ensure it is no longer
a favourable habitat for fungal growth. However, extensive research on the effect o f heat
treatments on biological resistance has shown the formation of new chemicals in thermally
modified wood contributes towards the toxicity o f heated wood for many fungi and bacteria.
In the earlier Chapters of the present study, chemical analysis of heated wood o f pine showed
the existence of potential antimicrobial compounds. Research reported in this chapter
therefore aims to evaluate the potential of antimicrobial activity of heat treated wood o f pine,
larch and rhododendron using the heating regime developed in Chapter 1 to control P.
ramorum. Initially, woodchips of pine were tested by direct contact with P. ramorum
progagules. In the later stages of the study, woodchips o f rhododendron and larch, the two
main hosts of P. ramorum in the UK, were then tested for their antimicrobial activity against
P. ramorum. Chemical analysis o f all three tested species was carried out and different
chemicals isolated and tested individually against P. ramorum to assess the active chemicals.
Finally, to evaluate the strength and magnitude of the enhanced activity under natural
conditions, field trials were set up in two different locations in south west England where P.
ramorum was present and the heated woodchips materials regularly monitored for the
presence of pathogen over a period of 6 month.
78

5.2 Experimental
5.2.1 Micro cosm trails
The antimicrobial activity of woodchips derived from pine, rhododendron and Japanese larch
was tested in a number of different trials. Heat-treated and air-dried woodchips were
prepared as described in Chapter 2 (2.10.1). Details o f test isolates of P. ramorum are also
given in Chapter 2 (2.10.3).
5.2.1.1 Effect of larch, pine and rhododendron heated woodchips on the survival of
Phytophthora ramorum zoospores
To assess the effect o f heat treated woodchips on growth and survival o f P. ramorum,
woodchips were obtained from the selected species using ten replicates for each treatment.
For each replicate, Ig of woodchips was placed in a Universal bottle, sealed and autoclaved
for 15 minutes at 120°C. Each was then inoculated with 1 ml of a spore suspension
(lO'^zoospores mf^) of the test P. ramorum isolates. Inoculated woodchips were then
incubated at 20°C for 3 weeks, then Ig of the woodchips of each treatment was placed on
Phytophthora selective medium (SMA) to monitor the survival of zoospores. Survival was
expressed as the number of treatments with positive growth of the pathogen from the
woodchips into the agar.
To detect the ability o f the pathogen (in the form of zoospores) to germinate and form
sporangia on the heated woodchips, one extra set of the same treatments was prepared and
received 1ml of the same spore suspension and an additional 1 ml o f sterile water. The
treatments were left under continuous light to induce sporangia for 2 weeks at 20°C. All the
bottles then received 1 0 ml of sterile water and were shaken well ( 2 1 bottles for each isolate)
before they were transferred to 4°C for an hour and moved back to ambient temperature for
45 minutes so zoospores were released from sporangia. The water content from each
universal bottle was transferred into a 25 ml sterile plastic bottles, 10 drops of 1 pi from each
bottle was placed on a glass slide. Using a microscope, the drops were then examined for
chlamydospores, zoospores and sporangia.
5.2.1.2 Effect of incorporating heated woodchips made from larch into soil infected with
P. ramorum
An experiment was designed to assess the potential of incorporating heated larch woodchips
to inhibit P. ramorum inoculum when combined. Two isolates BRCOl and P I578 were used
in the experiment. Heated and air-dried woodchips o f larch were ground to fine particles to
79

ensure a high level of interaction with soil. Soil free from P. ramorum was taken from a local
woodland (Alice Holt Forest, Famham, Surrey) and mixed with ground larch woodchips in
ratios of 25% and 75% (w:w) inside plastic bags. Each treatment received sterile water to its
water holding capacity, was sealed and left to settle at 25°C for 7 days. Plastic bags were
agitated and then the mixture was stirred every two days. A zoospore suspension which was
made as described in 2.10.3.1 (lO^zoospore mf^) and each bag received 10 ml o f the
suspension and was thoroughly mixed manually. Perlite was incorporated with soil in the
same ratios to act as controls. After the addition of the spore suspension, the bags were
sealed and stored at room temperature (20°C) for 4 weeks; thenl g o f the mixture was
removed from the bag, diluted with 1 0 ml sterile water and 1 ml o f the suspension plated onto
SMA. The same process was repeated after 8 weeks o f storage. The SMA plates were
incubated for 7 days at 20°C and any colonies o f P. ramorum were counted (cfu g" )̂. The
data were analysed by a one-way Analysis o f Variance ANOVA, GraphPadPrism, version
InStat3. The Tukey Post Analysis Test was performed to assess the least significance
difference between means (P<0.05). Survival of P. ramorum in the remaining mixtures in the
plastic bags was also tested.
5.2.1.3 Effect of incorporated heated woodchips made from larch on the survival of P.
ramorum chlamydospores
Apart from zoospores, P. ramorum produces chlamydospores that are considered to be long-
lived and resilient resting spores (Linderman et al., 2006). Chlamydospores of P. ramorum
are able to survive in soil for a long period o f time. To assess the inhibitory effect o f heated
woodchips when incorporated into the soil on the survival of chlamydospores, pre-colonized
leaf disks of rhododendron containing chlamydospores of P. ramorum were incorporated into
woodland soil (see above) which was then mixed with 75% heated larch woodchips.
Controls consisted of 100% soil and 100% heated woodchips. Treatments were compared
with an inert soil amendment consisting of 25 and 75% perlite. For each treatment, 12 leaf
disks previously colonized with isolates BRCOl and P I578 were used. The leaf disks were
recovered from the pots 4 weeks and 12 weeks after incorporation, rinsed with sterile water
and then plated onto SMA. Plates were incubated for at least 3 weeks at 20°C before each
disk was assessed for growth of P. ramorum. When the assays were completed, to assess if
P. ramorum had established in the soil itself, the soil from each pot was placed in another
container, flooded with sterile water and baited with 12 fresh rhododendron leaf disks. After
5 days incubation at 20°C, the bait disks were plated on SMA to check for any outgrowth o f
80

p. ramorum. The data were analysed by one-way ANOVA, GraphPadPrism, version InStatS.
The Tukey Post Analysis Test was performed to assess the least significance difference
between means (P<0.05).
5.2.2 In vitro assays
5.2.2.1 Assessment of Minimum Inhibitory Concentration (MIC) of crude extracts
against P, ramorum zoospores
For the assessment of the Minimum Inhibitory Concentration (MIC) o f the methanol crude
extracts derived from air-dried and heated wood were tested against two P. ramorum isolates
BRCOl and P I578. Concentrations o f 50, 25, 12.5, 6.25 and 3.12 mg o f extract per ml in
acetone were prepared. Half strength carrot broth was also prepared to provide a growth
medium for P. ramorum and was dispensed into each well of the assay plate and the acetone
evaporated in a fume hood. After evaporation, each well was inoculated with 5 pi of a fresh
zoospore suspension of P. ramorum containing 10"̂ zoospores ml"\ Each well contained a
final concentration range of 50, 25, 12.5, 6.25 or 3.12 mg of crude extract per ml o f carrot
broth. Plates were incubated for 24 hour before 20pi of the broth was removed and plated
onto SMA. The SMA plates were incubated at 20°C and scored as positive if growth o f P.
ramorum was detected. Each treatment was replicated three times.
5.2.2.2 Effect of crude extracts derived from heated and air-dried wood on
chlamydospore germination of P. ramorum
An experiment was set up to assess the effect o f the methanol crude extract derived from air-
dried and heated wood of pine, larch and rhododendron against P. ramorum chlamydospores
from isolates BRCOl and P I578. To test the effect of the different methanol extracts, each
crude extract was made in acetone. The test used leaf disks of rhododendron pre-colonized
with chlamydospores of P. ramorum. To expose the chlamydospores to the crude extract, a
six well plate was filled with 1 ml of sterile water in each well. Subsequently 1 ml o f extract
was pipetted into each well and the plates placed in a fume hood for 16 hours to allow the
acetone to evaporate giving a crude extract concentration in water of 10 mg m l '\ For each
treatment, two wells were used, each containing six pre-colonized leaf disks. To ensure good
contact between the chalamydospores and the extract, plates were incubated for 48 hours on a
platform shaker at ambient temperature. After 48 hours incubation, disks were removed from
the solution, rinsed in sterile water for Ih before being placed onto SMA. The SMA plates
were incubated for 3 weeks and the number of disks (average out of 6 disks from each o f two
wells) showing no outgrowth of P. ramorum, were calculated.
81

5.2.2.3 M IC of chemicals in crude extracts derived from heated pine wood against
hyphae, zoospores and chlamydospores of P. ramorum
The Minimum Inhibitory Concentration (MIC) of the compounds found in the crude
methanol extract derived from heated pine wood was assessed. The compounds included
coniferaldehyde, acetovanillone and vanillin which were tested against mycelia, zoospores
and chlamydospores of two isolates of P. ramorum, BRCOl and P I578. A two fold dilution
series of each compound (range from 20 to 0.009 mmol) in DMSO (dimethyl sulfoxide) was
prepared and dispensed in multi-well plates containing zoospores, chalamydospores and
mycelia which were prepared as described in 2.10.3.1. Each treatment was replicated three
times. Plates were incubated at 20°C for 24 hours for zoospores and 48 h for chlamydospores
and mycelia. After incubation the content of each well was plated onto SMA plates and
incubated for 5 days before assessing growth of P. ramorum. The minimum concentration at
which no P. ramorum colonies developed was recorded as the MIC.
To assess if resin acids lowered the MIC o f the various compounds, the assay was repeated
with combinations of the compounds plus dehydroabietic acid tested against zoospores of
three P. ramorum isolates (BRCOl, P1578 and P2460). The dehydroabietic acid
concentration was kept at 20 mmol. A further test was set up to assess if different resin acids
(dehydroabietic acid, abietic acid and pimaric acid) differed in their ability to increase the
activity of acetovanillin, coniferaldehyde and vanillin.
5.2.3 Field studies
5.2.3.1 Largin Wood field evaluation
Experimental assays described above indicated that heat treated woodchips particularly of
larch and pine, could be inhibitory to zoospores and chlamydospores of P. ramorum when
tested in the lab. Therefore, trials were designed to assess whether heat treated woodchips of
natural hosts of P. ramorum (rhododendron and larch) would inhibit the growth o f P.
ramorum under conditions in the field with variable inoculum loads. In November 2011, a
field study was conducted at Largin Wood, Forestry Commission, forest to assess the ability
of heated woodchips to inhibit the establishment of P. ramorum in soil and the litter layer.
The location was chosen as it was likely that a source o f natural P. ramorum inoculum
existed. Prior to placement of wood samples, several foliage and soil samples were collected
from three different spots at the site and were baited using rhododendron leaf disks. All three
spots proved P. ramorum positive. The selected points were labelled and wood samples were
prepared and placed on the litter in the site (Figure 5.3).
82

To prepare the sample, woodchips of rhododendron, larch and pine were wrapped in
aluminium foil and heated for 72 hours at 120°C in a dry oven. The heated samples of each
wood species were then split into 30 batches o f 150 g o f woodchips. Each batch was sealed
in a fine mesh plastic bag and labelled. The same number of air-dried woodchip samples of
pine, larch and rhododendron were also bagged in 150 g batches and sealed. All the bags
were transferred to the field and 1 0 bags o f each treatment (heated v& air-dried) of each wood"
species were placed on the litter in each spot (60 bags) (Figure 5.3). A set o f 18 sample bags
( 6 from each spot) were collected from the field on a monthly basis; for analysis each bag
was transferred to a plastic container and baited using rhododendron leaf disks which were
placed onto SMA after 5 days. The numbers o f leaf disks with positive outgrowth of P.
ramorum (n=30) were recorded for comparison. The data were analysed by a one-way
ANOVA, GraphPadPrism, version InStat3. The Tukey Post Analysis Test was performed to
assess the least significance difference between means (P<0.05).
5.2.3.2 Druid’s Hill field evaluation
A second experiment was set up between May and December 2012 to assess the inhibitory
effect of heat treated larch alone. The principle of this study was based on the fact that
woodchips or litter can become infected with P. ramorum when exposed to aerial inoculum
coming from infected foliar hosts. A5 size seed trays with drainage holes in the bottom were
filled with a 5cm layer o f wood chips made from heated larch woodchips. Air-dried
rhododendron wood chips were used as a control. Whole rhododendron leaves were used as
baits and placed on the top o f the woodchips. The trays were also covered with a fine mesh
net to protect them from animal damage. To minimise the risk of inoculum being washed off
by heavy rain, each tray was placed in a larger seed tray (without any drainage holes). In this
study site, the inoculum came from infected foliar o f R. ponticum and woodchips were placed
underneath infected rhododendron bushes (Figure 5.4).
83

r.
Figure 5.3 Field site in Largin W ood.
(a) landscape view o f the field site; (b) and (c) Nylon net bags containing w oodchips on site; (d) bagged sam ples o f w oodchips prior to analysis in the lab.
Samples were taken directly from the trays, and from the water that collected in the bottom
tray. To detect the presence of any viable P. ramorum, samples were baited with
rhododendron disks or subjected to RT-PCR for the detection of mRNA.
No P. ramorum could be detected in any of the treatments (air-dried or heat-treated wood
chips of larch and rhododendron) during May, June, July and August 2012. Test methods for
detection were both baiting and detection of live cells of P. ramorum using an mRNA based
RT-PCR assay (Chimento et al., 2011). Samples were negative (24 samples of larch and
rhododendron tested in July and all 36 tested samples in November).
84

Figure 5.4 Exam ple o f trays w ith w ood chips placed underneath a rhododendron bush at D ru id ’s H ill, Cornwall
(a) at the tim e o f set up in M ay 2012, (b) in D ecem ber 2012.
To determine the minimum detection limit of the RT-PCR assay (Chimento et al., 2011), a
dilution series of zoospores from the isolate of P. ramorum originating from foliage at the
survey site was prepared. After vacuum filtration, a concentration range of 1, 10, 100, 1000,
10,000, 100,000 and 1,000,000 zoospores m f’ in sterile water was made. The zoospore
concentrations of 1 , 1 0 and 1 0 0 zoospores m f’ were adjusted using a microliter pipette by
following the method described by Xu and Ko (1998). Five drops of the spore suspension
(each 1 pi) were placed on a glass slide and the number of spores was counted using a
microscope as a check. The drops were then recovered using a micropipette and transferred
to RNase free tubes. Concentrations above 100 zoospores m f’ were adjusted to the mean
value of four counts of spore suspensions made by a haemocytometer. RNA was extracted
from the samples using RNeasy Plant MiniKit (Qiagen Inc, Valencia, CA, USA) for total
RNA isolation from plants and filamentous fungi, following the manufacture’s protocol.
After elimination of genomic DNA, reverse transcription was performed by two-step RT-
PCR using QuantiTect® Reverse Transcription kit (Qiagen) following the manufacture’s
procedure and RT Primer Mix (Qiagen). Then, cDNA was amplified using
ACPramF/ACPramR primer set. PCR was performed in a total volume of 25 pi containing
6.26 pi cDNA, with 12.5 pi of SYBER® Green Mastermix containing 0.5 pmol of each
primer. Real-time amplification was carried out in a Real Time thermal cycler (LightCycler®
480 II) using the following program: one cycle at 50°C for 10 minutes, one cycle at 95°C for
3 minutes, 35 cycles at 95°C for 15 second and 63°C for 1 minutes. Ramp rate was 3.3°C/s
heating and 2°C. PCR amplification with Ct values (threshold cycle) lower than 31 were
considered positive.
85

5.2.4 Chemical analyses
5.2.4.1 Chemical analysis of heated larch, pine and rhododendron wood
GC-MS analysis of the methanol crude extracts of heat-treated pine, larch and rhododendron
(1 mg ml'* methanol) was carried out to compare the chemical constitutes (Figure 5.9). All
three crude extracts were dissolved in methanol as described in Chapter 2 (2.9.2) and were
subjected to Gas Chromatography Mass Spectrometry.
5.2.4.2 Chemical analysis of discoloured bark of a Japanese larch
When bark of Japanese larch is infected by P. ramorum, the phloem tissue undergoes a
number of colour changes that appear to be the response of the tree as it defends itself against
the pathogen (Figure 5.5). To assess what chemical changes in the bark could be associated
with the alternation in colour; a GC-MS analysis of the different bark samples was
performed. The analysis used DCM crude extracts of bark at different stages of
discolouration following invasion by P. ramorum. These consisted of: white tissue not yet
invaded; red tissue indicating a response to P. ramorum but usually LTD negative; dark
brown tissue, LTD positive and P. ramorum could usually be cultured from this tissue. Apart
from these samples, a DCM crude extract of the white phloem tissue from a healthy tree was
included for comparison.
f iFigure 5.5 Bark (phloem ) o f a Japanese larch tree show ing different stages o f discolouration, (a) white (b) red and dark brown tissue formed as a response to invasion by P. ramorum.
86
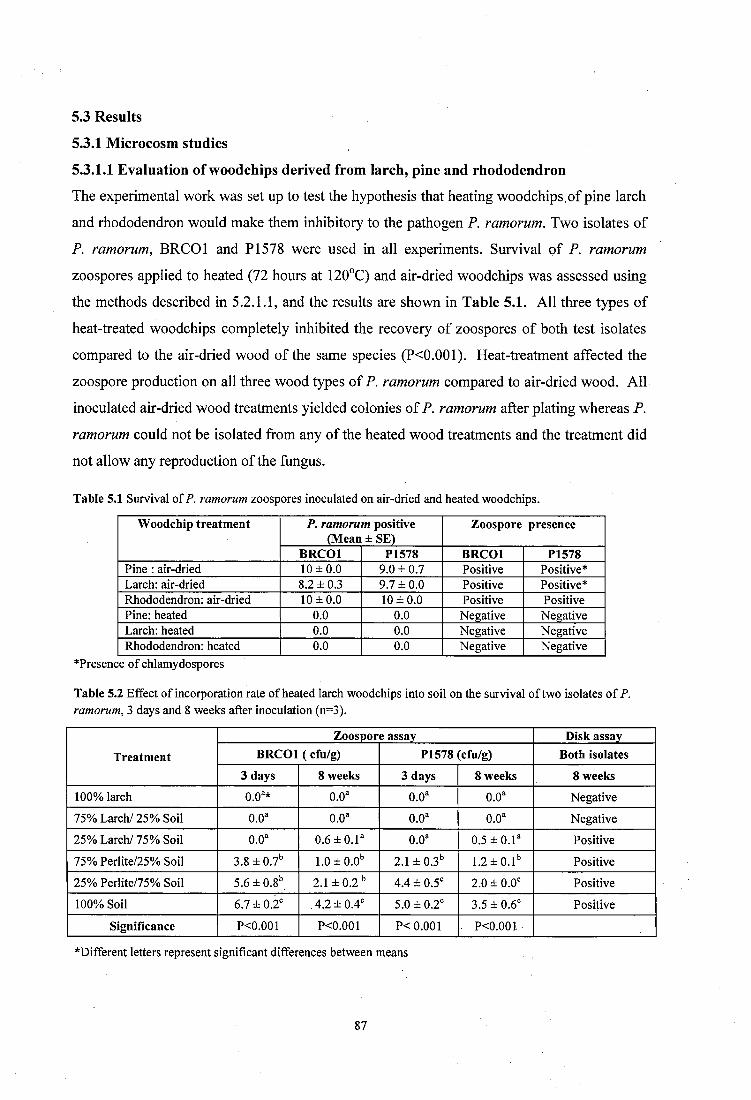
5.3 Results
5.3.1 Microcosm studies
5.3.1.1 Evaluation of woodchips derived from larch, pine and rhododendron
The experimental work was set up to test the hypothesis that heating woodchips , of pine larch
and rhododendron would make them inhibitory to the pathogen P. ramorum. Two isolates of
P. ramorum, BRCOl and P I578 were used in all experiments. Survival o f P. ramorum
zoospores applied to heated (72 hours at 120°C) and air-dried woodchips was assessed using
the methods described in 5.2.1.1, and the results are shown in Table 5.1. All three types of
heat-treated woodchips completely inhibited the recovery of zoospores of both test isolates
compared to the air-dried wood of the same species (P<0.001). Heat-treatment affected the
zoospore production on all three wood types of P. ramorum compared to air-dried wood. All
inoculated air-dried wood treatments yielded colonies o f P. ramorum after plating whereas P.
ramorum could not be isolated from any of the heated wood treatments and the treatment did
not allow any reproduction of the fungus.
Table 5.1 Survival o f P. ramorum zoospores inoculated on air-dried and heated woodchips.
W oodchip treatm ent P. ram orum positive (M ean ± SE)
Z oospore presence
B R C O l P1578 B R C O l P1578Pine : air-dried 10 ± 0 .0 9.0 ± 0.7 Positive Positive*Larch: air-dried 8.2 ± 0.3 9.7 ± 0.0 Positive Positive*Rhododendron: air-dried 10 ± 0 .0 10 ± 0 .0 Positive PositivePine: heated 0.0 0.0 Negative NegativeLarch: heated 0.0 0.0 Negative NegativeRhododendron: heated 0.0 0.0 Negative Negative
*Presence o f chlamydospores
Table 5.2 Effect o f incorporation rate o f heated larch woodchips into soil on the survival o f two isolates o f P. ramorum, 3 days and 8 w eeks after inoculation (n=3).
Treatm ent
Z oospore assay D isk assayB R C O l ( cfu/g) P1578 (cfu/g) B oth isolates
3 days 8 w eeks 3 days 8 w eeks 8 w eeks
100% larch 0.0“* 0.0“ 0.0“ 0.0“ N egative
75% Larch/ 25% Soil 0.0“ 0.0“ 0.0“ 0.0“ Negative
25% Larch/ 75% Soil 0.0“ 0.6 ± 0 .1 “ 0.0“ 0.5 ± 0 .1 “ Positive
75% Perlite/25% Soil 3.8 ± 0.7^ 1.0 ± 0 .0 '’ 2.1 ± 0 .3 '’ 1.2 ±0.1^ Positive
25% Perlite/75% Soil 5.6 ± 0.8'’. 2.1 ± 0 .2 ' ’ 4.4 ± 0.5° 2.0 ± 0.0° Positive
100% Soil 6.7 ± 0.2° . 4.2 ± 0.4° 5.0 ± 0.2° 3.5 ± 0.6° Positive
Significance P<0.001 P<0.001 P< 0.001 P<0.001
*Different letters represent significant differences between means
87
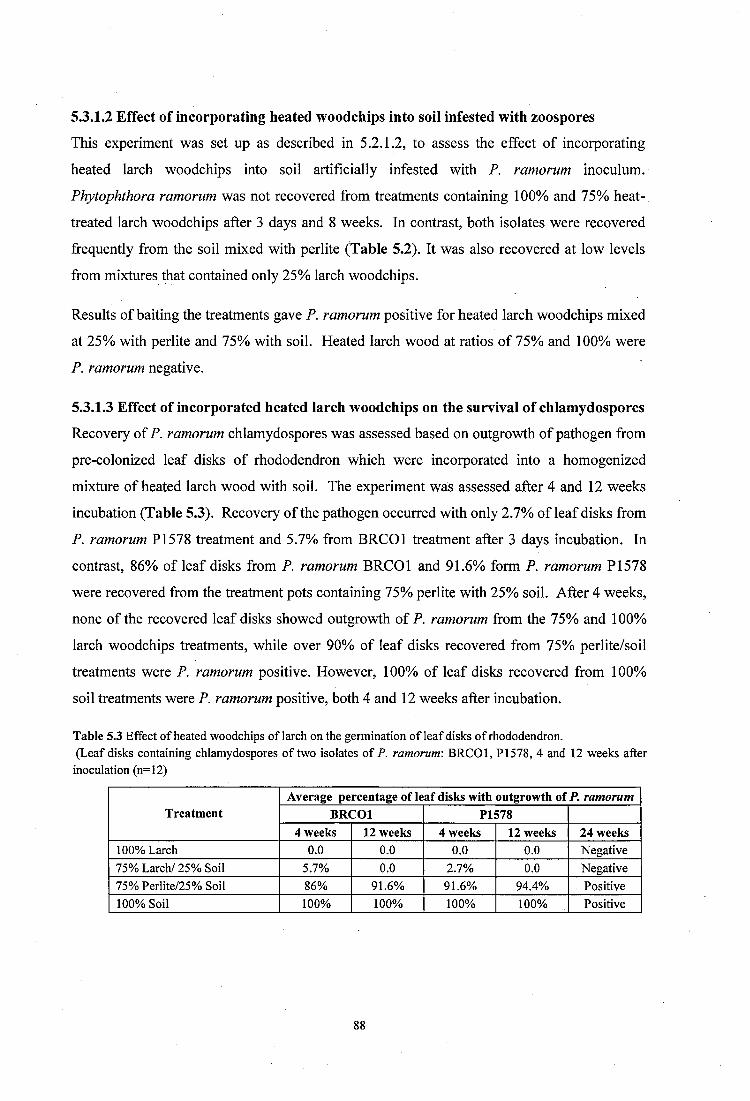
5.3.1.2 Effect of incorporating heated woodchips into soil infested with zoospores
This experiment was set up as described in 5.2.1.2, to assess the effect o f incorporating
heated larch woodchips into soil artificially infested with P. ramorum inoculum.
Phytophthora ramorum was not recovered from treatments containing 100% and 75% heat-
treated larch woodchips after 3 days and 8 weeks. In contrast, both isolates were recovered
frequently from the soil mixed with perlite (Table 5.2). It was also recovered at low levels
from mixtures that contained only 25% larch woodchips.
Results of baiting the treatments gave P. ramorum positive for heated larch woodchips mixed
at 25% with perlite and 75% with soil. Heated larch wood at ratios of 75% and 100% were
P. ramorum negative.
5.3.1.3 Effect of incorporated heated larch woodchips on the survival of chlamydospores
Recovery of P. ramorum chlamydospores was assessed based on outgrowth of pathogen from
pre-colonized leaf disks of rhododendron which were incorporated into a homogenized
mixture of heated larch wood with soil. The experiment was assessed after 4 and 12 weeks
incubation (Table 5.3). Recovery o f the pathogen occurred with only 2.7% of leaf disks from
P. ramorum P I578 treatment and 5.7% from BRCOl treatment after 3 days incubation. In
contrast, 8 6 % of leaf disks from P. ramorum BRCOl and 91.6% form P. ramorum P I578
were recovered from the treatment pots containing 75% perlite with 25% soil. After 4 weeks,
none of the recovered leaf disks showed outgrowth of P. ramorum from the 75% and 100%
larch woodchips treatments, while over 90% of leaf disks recovered from 75% perlite/soil
treatments were P. ramorum positive. However, 100% of leaf disks recovered from 100%
soil treatments were P. ramorum positive, both 4 and 12 weeks after incubation.
Table 5.3 Effect o f heated woodchips o f larch on the germination o f lea f disks o f rhododendron.(L eaf disks containing chlamydospores o f two isolates o f P. ramorum: B R C O l, P1578, 4 and 12 w eeks after
inoculation (n=12)
T reatm entA verage percentage o f lea f disks w ith outgrow th o f P. ram orum
B R C O l P15784 w eeks 12 w eeks 4 w eeks 12 w eeks 24 w eeks
100% Larch 0.0 0.0 0.0 0.0 N egative75% Larch/ 25% Soil 5.7% 0.0 2.7% 0.0 Negative75% Perlite/25% Soil 86% 91.6% 91.694 94.4% Positive100% Soil 100% 100% 100% 100% Positive
88
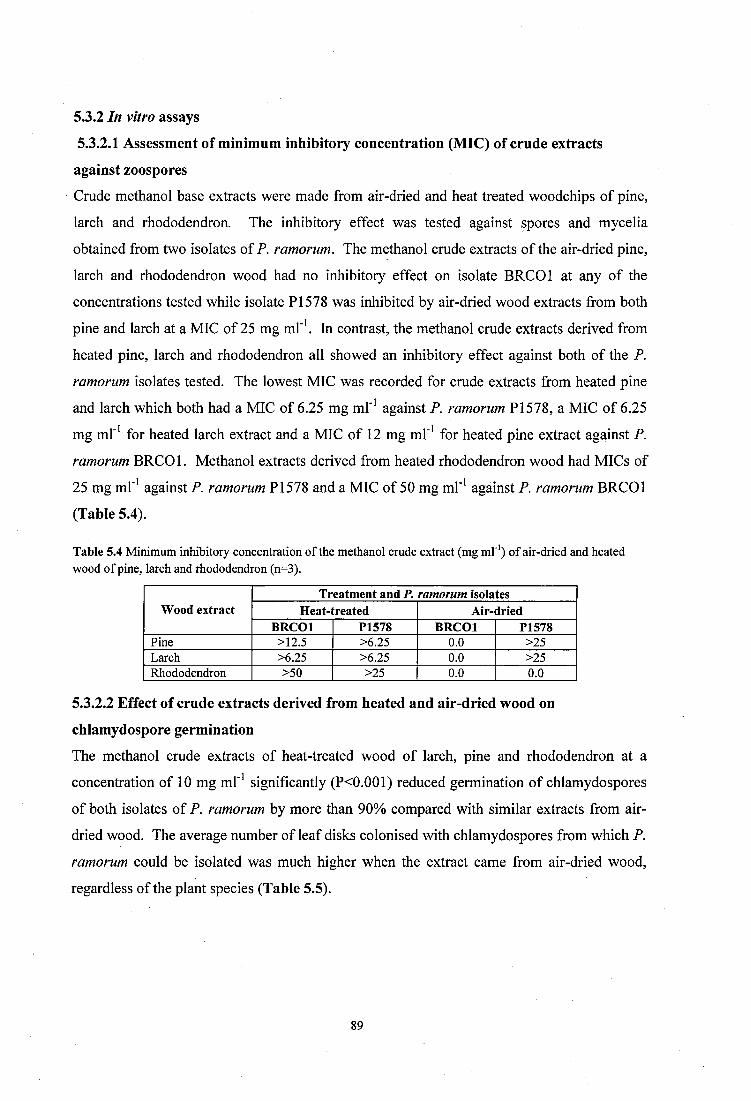
5.3.2 In vitro assays
5.3.2.1 Assessment of minimum inhibitory concentration (MIC) of crude extracts
against zoospores
Crude methanol base extracts were made from air-dried and heat treated woodehips of pine,
lareh and rhododendron. The inhibitory effeet was tested against spores and myeelia
obtained from two isolates of P. ramorum. The methanol crude extracts o f the air-dried pine,
larch and rhododendron wood had no inhibitory effect on isolate BRCOl at any o f the
concentrations tested while isolate P I578 was inhibited by air-dried wood extracts from both
pine and lareh at a MIC o f 25 mg ml"\ In contrast, the methanol erude extracts derived from
heated pine, lareh and rhododendron all showed an inhibitory effect against both of the P.
ramorum isolates tested. The lowest MIC was reeorded for crude extraets from heated pine
and larch which both had a MIC of 6.25 mg ml'^ against P. ramorum P1578, a MIC o f 6.25
mg ml'^ for heated larch extract and a MIC o f 12 mg ml'^ for heated pine extraet against P.
ramorum BRCOl. Methanol extracts derived from heated rhododendron wood had MICs of
25 mg ml'^ against P. ramorum P1578 and a MIC o f 50 mg ml'^ against P. ramorum BRCOl
(Table 5.4).
Table 5.4 Minimum inhibitory concentration o f the methanol crude extract (mg ml ') o f air-dried and heated wood o f pine, larch and rhododendron (n=3).
W ood extractTreatm ent and P. ram orum isolates
H eat-treated A ir-driedB R C O l P1578 B R C O l P1578
Pine >12.5 >&25 0.0 >25Larch >&25 >&25 0.0 >25Rhododendron >50 >25 0.0 0.0
5.3.2.2 Effect of crude extracts derived from heated and air-dried wood on
chlamydospore germination
The methanol crude extracts o f heat-treated wood of larch, pine and rhododendron at a
eoncentration of 10 mg ml'^ significantly (P<0.001) reduced germination o f ehlamydospores
of both isolates of P. ramorum by more than 90% eompared with similar extraets from air-
dried wood. The average number of leaf disks eolonised with ehlamydospores from which P.
ramorum could be isolated was much higher when the extract came from air-dried wood,
regardless of the plant species (Table 5.5).
89
Table 5.5 Effect o f m ethanol crude extracts o f air-dried and heated w ood o f pine, larch and rhododendron on germ ination o f pre-colonized lea f disks o f rhododendron with chlam ydospores o f tw o isolates o f P. ramorum. Data presented represent num ber o f lea f disks (out Of 6) that failed to yield any P. ramorum in two replicates o f each treatm ent.
IsolatesM ean num ber o f lea f disks w ithout outgrow th o f P. ram orum
H eat treated wood A ir-dried w oodPine Larch Rhododen Pine Larch Rhododen
P1578 5.75 ± 0 .2 6.00± 00 5.25± 0.2 1.0 ± 0 .7 0.5 ± 0.2 0.25 ± 0 .2
B RC O l 5.5 ± 0.5 5.5 ±0.2 5.0 ± 0 .4 0.0 0.0 0.0
Effect o f Isolate NS NS N S NS NS N SEffect o f W ood type NS NSEffect o f heat treatm ent P<0.001
5.3.2.3 Minimum Inhibitory Concentration (MIC) of different chemicals found in crude
extracts derived from heated pine wood
Pure compounds were 2-10 times more active against zoospores, chlamydosores and myeelia
of P. ramorum than the erude extract (Table 5.5). Coniferaldehyde was the most active
compound with a MIC of 1.7 mg m f' against myeelia and chlamydosores of P. ramorum and
a MIC of 0.15mg m f’ against zoospores (Table 5.6). Aeetovanillone and vanillin had similar
anti-microbial properties with MICs of 3.4-6 . 6 mg m f’ against myeelia and ehlamydospores
and MICs of between 1.5 and 1.6 mg m f’ against zoospores. All compounds (including the
crude extract) showed similar activities against both P. ramorum isolates tested (Table 5.6).
Table 5.6 M inim um inhibitory concentration (m g m l ' ) o f the com pounds derived from heated wood o f pine on m yeelia, zoospores and chlam ydospores (in colonized lea f disks) o f tw o isolates o f P. ramorum.
C om poundsM inim um inhibitory concentration (m g ml ')
M yeelia Zoospores C hlam ydosporesIsolates BR C O l PI 578 B R C O l PI 578 B R C O l P1578
Crude extract 25 25 6 2 5 6 2 5 10 10
Aeetovanillone 6.6 3.3 1.6 1.6 6.6 3.3
Coniferaldehyde 1.7 1.7 0.15 0.15 1.7 1.7
Vanillin 3.4 3.4 1.52 1.52 3.4 3.4
(a) (b)
Figure 5.6 Exam ples o f an assay plate (96-well plate) after the chem ically treated zoospores are plated on SMA (a), and (b) an enlarged picture o f normal grow th o f P. ramorum in sterile w ater (control).
90
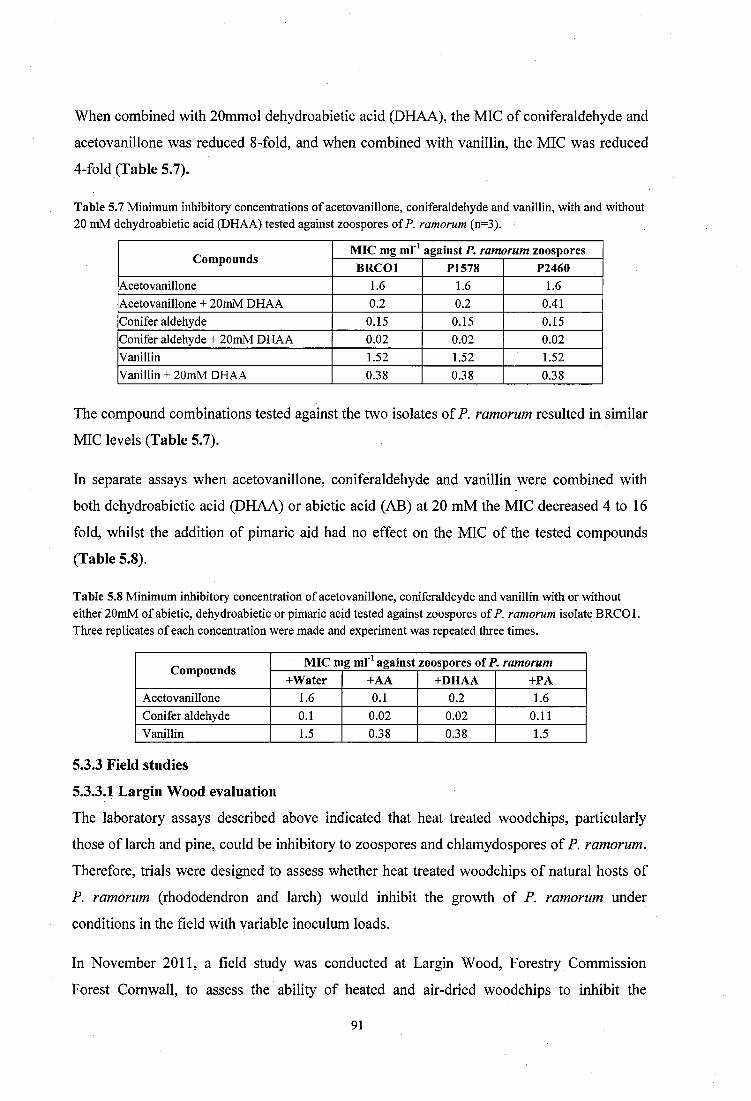
When combined with 20mmol dehydroabietic acid (DHAA), the MIC of coniferaldehyde and
aeetovanillone was reduced 8 -fold, and when combined with vanillin, the MIC was reduced
4-fold (Table 5.7).
Table 5.7 Minimum inhibitory concentrations o f aeetovanillone, coniferaldehyde and vanillin, with and without 20 mM dehydroabietic acid (DHAA) tested against zoospores o f P. ramorum (n=3). •
C om poundsM IC m g ml ' against P. ram orum zoospores
B R C O l P1578 P2460Aeetovanillone 1.6 1.6 1.6Aeetovanillone + 20mM DHAA 0.2 0.2 0.41Conifer aldehyde 0.15 0.15 0.15Conifer aldehyde + 20mM DHAA 0.02 0.02 0.02Vanillin 1.52 1.52 1.52Vanillin + 20mM DHAA 0.38 0 3 8 0.38
The compound combinations tested against the two isolates of P. ramorum resulted in similar
MIC levels (Table 5.7).
In separate assays when aeetovanillone, coniferaldehyde and vanillin were combined with
both dehydroabietic acid (DHAA) or abietic acid (AB) at 20 mM the MIC decreased 4 to 16
fold, whilst the addition of pimaric aid had no effect on the MIC of the tested compounds
(Table 5.8).
Table 5.8 Minimum inhibitory concentration o f aeetovanillone, coniferaldeyde and vanillin with or without either 20mM o f abietic, dehydroabietic or pimaric acid tested against zoospores o f P. ramorum isolate B R C O l. Three replicates o f each concentration were made and experiment was repeated three times.
C om poundsM IC m g m l ' against zoospores o f P. ram orum
+W ater +A A +D H A A +PAAeetovanillone 1.6 0.1 0.2 1.6Conifer aldehyde 0.1 0.02 0.02 0.11Vanillin 1.5 0.38 0.38 1.5
5.3.3 Field studies
5.3.3.1 Largin Wood evaluation
The laboratory assays described above indicated that heat treated woodchips, particularly
those of larch and pine, could be inhibitory to zoospores and chlamydospores of P. ramorum.
Therefore, trials were designed to assess whether heat treated woodchips of natural hosts of
P. ramorum (rhododendron and larch) would inhibit the growth o f P. ramorum under
conditions in the field with variable inoculum loads.
In November 2011, a field study was conducted at Largin Wood, Forestry Commission
Forest Cornwall, to assess the ability of heated and air-dried woodchips to inhibit the
91
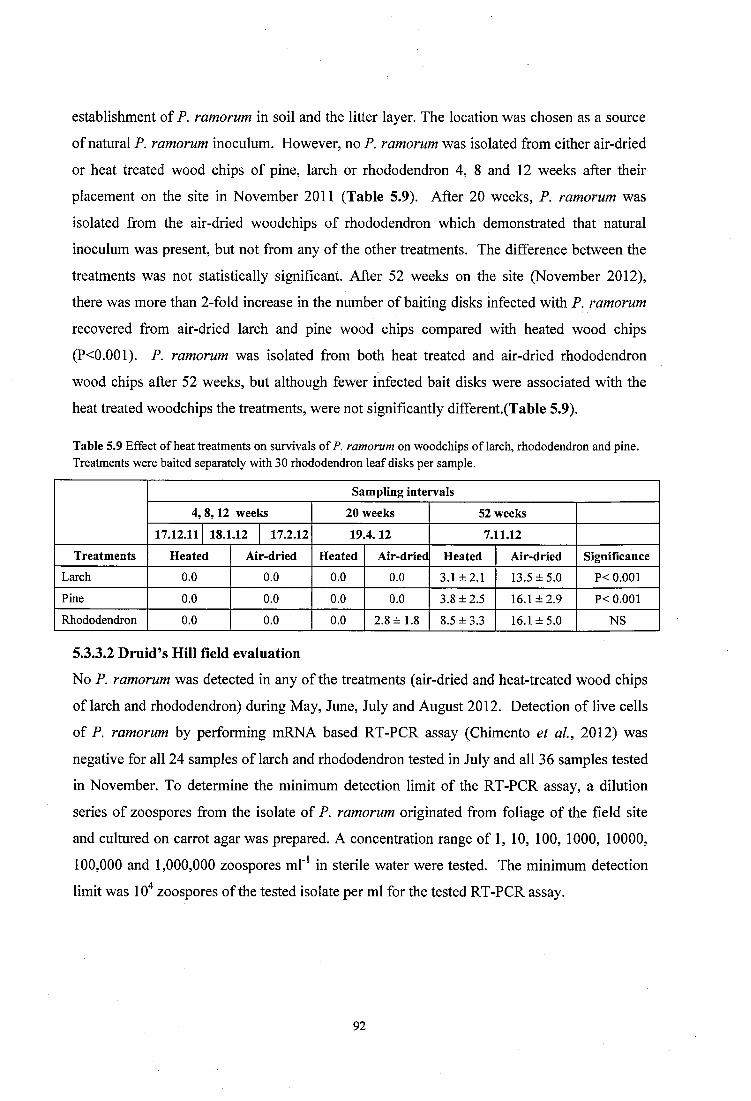
establishment of P. ramorum in soil and the litter layer. The location was chosen as a source
o f natural P. ramorum inoculum. However, no P. ramorum was isolated from either air-dried
or heat treated wood chips o f pine, larch or rhododendron 4, 8 and 12 weeks after their
placement on the site in November 2011 (Table 5.9). After 20 weeks, P. ramorum was
isolated from the air-dried woodchips of rhododendron which demonstrated that natural
inoculum was present, but not from any o f the other treatments. The difference between the
treatments was not statistically significant. After 52 weeks on the site (November 2012),
there was more than 2-fold increase in the number of baiting disks infected with P. ramorum
recovered from air-dried larch and pine wood chips compared with heated wood chips
(P<0.001). P. ramorum was isolated from both heat treated and air-dried rhododendron
wood chips after 52 weeks, but although fewer infected bait disks were associated with the
heat treated woodchips the treatments, were not significantly different.(Table 5.9).
Table 5.9 Effect o f heat treatments on survivals o f T. ramorum on woodchips o f larch, rhododendron and pine. Treatments were baited separately with 30 rhododendron lea f disks per sample.
Sam pling intervals
4, 8 ,1 2 w eeks 20 w eeks 52 w eeks
17.12.11 18.1.12 17.2.12 1 9 .4 .1 2 7.11.12
Treatm ents H eated A ir-dried H eated A ir-dried H eated A ir-dried Significance
Larch 0.0 0.0 0.0 0.0 3.1 ± 2 .1 13.5 ± 5 .0 P< 0.001
Pine 0.0 0.0 0.0 0.0 3.8 ± 2 .5 16.1 ± 2 .9 P< 0.001
Rhododendron 0.0 0.0 0.0 2.8 ± 1 .8 8.5 ± 3.3 16.1 ± 5 .0 N S
5.3.S.2 D ruid’s Hill field evaluation
No P. ramorum was detected in any of the treatments (air-dried and heat-treated wood chips
of larch and rhododendron) during May, June, July and August 2012. Detection o f live cells
of P. ramorum by performing mRNA based RT-PCR assay (Chimento et a l, 2012) was
negative for all 24 samples of larch and rhododendron tested in July and all 36 samples tested
in November. To determine the minimum detection limit of the RT-PCR assay, a dilution
series o f zoospores from the isolate of P. ramorum originated from foliage o f the field site
and cultured on carrot agar was prepared. A concentration range of 1, 10, 100, 1000, 10000,
100,000 and 1,000,000 zoospores mf^ in sterile water were tested. The minimum detection
limit was 10"̂ zoospores of the tested isolate per ml for the tested RT-PCR assay.
92

5.3.4 Chemical analyses
5.3.4.1 Chemical analysis of heated larch, pine and rhododendron wood used in the field
GCMS analysis o f the methanol crude extracts o f heat-treated pine, larch and rhododendron
(Im g ml'^ methanol) was carried out to compare the chemical constitutents (Figure 5.7).
Spectra o f all three crude extracts had peaks for vanillin at 13.7 minutes, vanillic acid at 15.8
minutes, and syringaldéhyde at 16.9 minutes. The peak for vanillin was considerably higher
in the crude extract o f pine and larch compared to rhododendron. One peak at 17.7 minutes,
identified as coniferaldehyde (NIST library), was only present in larch and pine extracts. In
contrast, benzoic acid (18.67 minutes), syringic acid (18.73 minutes) and sinapaldehyde
(20.34 minutes) were only present in the crude extract o f rhododendron (Figure 5.7).
5.3.4.2 Chemical analysis of discoloured bark of a Japanese larch
The analysis o f the DCM crude extracts revealed that prior to detection o f the pathogen in
invaded bark, white phloem tissue showed an abundance o f resin acid 2 (2 4 .8 minutes) and
much lower amounts o f resin acid 1 (24 minutes) compared with the phloem from the
healthy control (Figure 5.8 a, b). However, in the crude extract o f the red-discoloured bark,
resin acids 1 and 2 were both present in considerable quantities, whereas in the brown tissue
resin acid 2 was at levels similar to the control (Figure 5.8 c, d). Due to trace quantity o f
resin acids, they were identified firom a chromatogram o f a spiked sample o f DCM crude
extract with 10 mg o f synthetic DHAA and AB. The resin acid number 2 was identified as
dehydroabietic acid (A2 1 ) and resin acid number 1 was identified as methyl abietate (A2 0 ), a
derivative o f abietic acid. It was also striking that particularly the red discoloured tissue had
raised levels o f many other compounds compared with the control and white coloured
phloem tissue. The monoterepenes, a-pinene (6.14 minutes) and 3-carene (7.58 minutes)
both were only present in the extract o f red and dark brown phloem which were formed
during and after pathogen invasion (A2 2 and A 2 3 ).
93
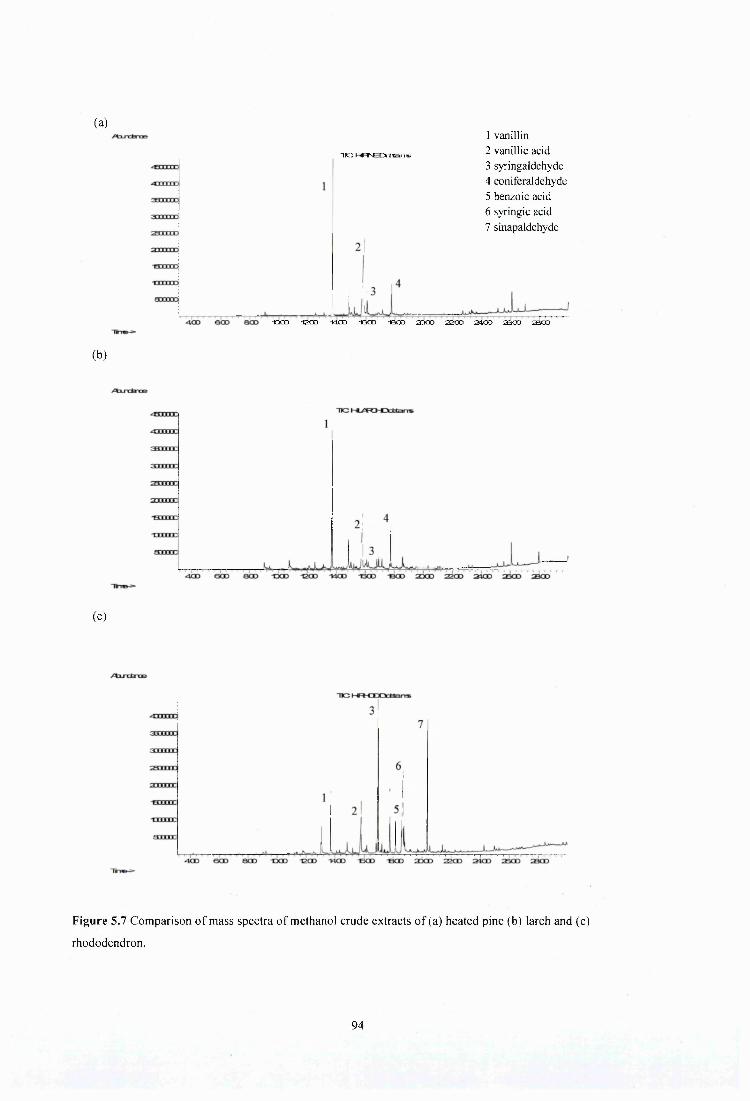
(a)
I K ; t 41 ffsl I \ i t: if T?.
1 vanillin2 vanillic acid3 syringaldéhyde4 coniferaldehyde5 benzoic acid6 syringic acid7 sinapaldehyde
-OCD 12CD 1400 -BOD 1BOD 3000 2200 3400 2SQO 3300
(b)
(c)
F igu re 5.7 C om parison o f mass spectra o f methanol crude extracts o f (a) heated pine (b) larch and (c)
rhododendron.
94

CLP
S"
O
IOÆ -Q CL c
IIcC>1
o
Di
E
-g-Q1301 -Ot+xo
g<u
I0
1CL
3E
L -oc
. iraCLEo
U00« /
i
I Ii l
îi<u
Iii!S 3IE
ooT|-(N3on ÛO
î i
! 1
î fI I
: gcS u
CL
CL

5.4 Discussion
The research described in this chapter was aimed at testing the hypothesis that heating
woodchips o f pine (Pinus sylvestris), larch {Larix kaempferi) and rhododendron
{Rhododendron ponticum) would make them inhibitory to the pathogen P. ramorum.
Starting with micro-cosm studies in the laboratory, all three types of heat-treated
woodchips significantly inhibited the recovery of zoospores o f P. ramorum compared
with air-dried woodchips of the same species. Results of leaf disk assays o f the
inoculated woodchips also showed that not only zoospores of P. ramorum were able
to survive on air-dried wood chips, they were also able to give rise to sporangia with
all three species tested. A previous study on survival of P. ramorum in air-dried
firewood of tanoak has suggested that spores o f P. ramorum are able to survive up to
6 months in firewood under wet conditions. Twenty pieces of firewood (exposed to
natural inoculum and rainfall) were tested for the presence of viable zoospores as well
as chlamydospores using a similar baiting method to that used in this study (Shelly et
a l, 2006). Although, the rate of positive samples was low (14 out of 80), the results
suggested a 12% chance of the pathogen spreading from a piece of firewood to a
susceptible host under favourable conditions. The results o f the present study also
indicate that air-dried wood materials (in this case pine, larch and rhododendron) may
be able to support P. ramorum sporulation on the forest floor under condition o f high
inoculum levels. In contrast, when the same materials were heat-treated, they no
longer supported growth and zoospore production. Thus heating processed wood
after harvesting infected trees could be an effective method of reducing the risk of
pathogen establishment and spread when prolonged storage is needed. Furthermore,
since woodchips can be used as physical barriers to reduce the spread o f pathogens
such as P. ramorum, the use o f woodchips should be limited to species that do not
support survival and production of P. ramorum when used against this pathogen.
In the next step of the study, the extent to which the inhibitory effect might be altered
by mixing with soil was assessed. As all three heat-treated types of woodchip showed
similar inhibitory effects, only larch was tested in a mixture with soil. Heated larch
woodchips completely inhibited the recovery o f zoospores mixed in a 75:25% ratio
with soil, whereas when perlite was substituted for the woodchips, P. ramorum was
frequently recovered. These results are important since P. ramorum is known to be
able to survive and even produce sporangia in the litter interface with soil and foliage
96

(Fichtner et a l, 2007a). Survival and sporulation of P. ramorum in foliage is an
important stage in the pathogen life cycle. It allows spore inoculum to build up in
sufficient quantities to cause infection o f mature trees when temperature and moisture
conditions are favourable. Therefore, it is crucial to control pathogen populations
through control measures which reduce inoculum levels. The results o f using heated
larch woodchips mixed with soil also showed their inhibitory activity is retained
under moist conditions. This suggests the heat-treated woodchips could have the
potential to control pathogen survival under wet conditions, and even when mixed or
in contact with soil interface. However, the inhibitory effect may vary under natural
soil moisture conditions and needs to be investigated further.
Similarly, mixtures of heated wood of larch with soil (75:25) were inhibitory to
chlamydospores o f P. ramorum. After 4 weeks in the larch/soil mix only 5% of
recovered leaf disks containing chlamydospres supported outgrowth o f the pathogen,
compared with 90% recovery with perlite/soil treatments and 100% recovery with soil
only. Results were consistent even after 12 weeks of incubation. Lack o f recovery of
P. ramorum from pre-colonized rhododendron baits placed in different potting media
has been also reported (Linderman and Davis, 2006). Insufficient moisture and
presence o f inhibitors in the substrate are considered to be the major factors causing
this. This study suggests that the inhibitory effect o f heated wood o f larch could give
long term suppression o f the germination o f chlamydospores at least in a moisture
controlled environment. However, the long term effect could reduce under natural
conditions, as a result of rainfall leaching the inhibitory compounds from the
woodchips.
In order to identify potential inhibitory components of heated wood, crude extracts
were initially tested on zoospores, and later tested on myeelia and chlamydospores.
The preliminary tests showed the responses o f two P. ramorum isolates to crude
extracts are dose dependent. Although, all three crude extracts of heat-treated wood
species had an inhibitory effect on P. ramorum zoospores, pine and larch had a
greater effect than rhododendron (6.25 mg ml" ̂ vs. 50 mg ml'^). Phytophthora
ramorum isolate BRCOl also showed more tolerance in comparison with isolate
P I578. Unexpectedly, air-dried pine and larch also had inhibitory effect on isolate
PI 578 while none of air-dried crude extracts had any effect on BRCOl.
97

The same extracts significantly reduced germination of chlamydospores by more than
90% compared with extracts from air-dried wood. This may be inhibition rather than
lethal effect of the extracts. Since all three tested wood species showed similar
inhibitory effects, the potential inhibitors are likely to be produced through chemical
modification in structural components of wood upon heating. Chemical analysis of
the three crude extracts showed some o f the induced compounds are present in the all
three crude extracts but in different concentrations. Spectra of the three crude extracts
had vanillin, vanillic acid and syringaldéhyde, but the peak for vanillin was
considerably higher in the extracts of pine and larch compared to rhododendron.
Interestingly, coniferaldehyde was only present in larch and pine extracts, whereas,
benzoic acid, syringic acid and sinapaldehyde were only present in the extract of
rhododendron. In general, pine and larch extracts contained the same compounds in
similar concentrations.
To determine precisely the active compounds of the extracts, purified compounds
dominant in the crude extract of heated pine including vanillin, aeetovanillone and
coniferaldehyde were tested on zoospores, chlamydospores and mycelium o f the two
isolates of P. ramorum. Pure compounds were 2-10 times more active against all three
propagules types compared with the crude extracts. Coniferaldehyde was the most
active compound against myeelia, chlamydospores and zoospores o f P. ramorum.
Since coniferaldehyde was not found in the crude extract o f heat-treated
rhododendron, this may explain why pine and larch extracts had much a greater effect
compared to rhododendron. Aeetovanillone and vanillin had similar antimicrobial
properties against myeelia, chlamydospores and zoospores. Both coniferaldehyde and
vanillin have previously been found to exhibit antifungal activity against the wood
decay fungi Trametes versicolor and Coniphoraputeana (Voda et a l, 2003). Garbber
et al., (1998) conducted a cell-wall model system to evaluate what effect aldehyde-
containing lignins have on the hydrolysis o f maize cell walls by fungal enzymes
(Grabber et a l, 1998) and their finding revealed severe inhibition of wall degradation
occured with coniferaldehyde. As phenolic aldehydes, both compounds are known to
be the most toxic heartwood components of conifers. Also, a previous study by Nagle
et al (2011) suggests Quercus spp trees with field-resistance to P. ramorum contain
high level of phenolic components compared with susceptible trees. Overall it can be
concluded that high levels of phenolic aldehydes that are induced as a result o f
98

heating are likely to be responsible for the enhanced inhibitory effect of heat treated
woods. However, results of chemical analysis and the bioassays o f identified
compounds already presented in Chapter 4 suggest that the magnitude of
antimicrobial activity may not only be due to aldehydes since their effects are known
to be dose dependent. As Chapter 4 revealed, resin acids contribute a greater effect
when working in conjunction with aldehydes.
To test the hypothesis that resin acids contribute toward enhanced antimicrobial
activity, individual compounds were tested in combination with the resin acids
already identified in Chapter 4. Dehydroabietic acid, decreased the minimum
inhibitory effect o f all three tested phenolic compounds while it showed no effect
when tested separately against P. ramorum. Dehydroabietic acid, the dominant resin
acid present in heated crude extracts o f both pine and larch has shown antifungal
activity against Trametes versicolor and Trichoderma species (Savluchinske-Feio et
a l, 2007). The oxidized derivatives of dehydroabietic acid are reported to have good
antimicrobial activity against Dothistroma pini, a pathogen of Pinus species and
Heterobasidion annosum, a root pathogen of conifers (Franich et a l, 1983; Henriks et
a l, 1979; Savluchinske-Feio et a l, 1999). When Savluchinske Feio et a l, (2007)
studied the potential o f synthetic derivatives of dehydroabietic acid as a conventional
fungicides, they found it inhibited normal growth of Botrytis cinerea and
Lophodermium seditiosum. Dehydroabietic acid derivatives exhibit activity against a
wide range of bacteria and fungi (Savluchinske-Feio, et a l, 1999). These derivatives
may be potential replacement for wood preservatives with less environmental impact
(Savluchinske-Feio et a l , 2007).
Interestingly, abietic acid showed similar results with just one isolate of P. ramorum
(BRCOl) while pimaric acid had no effect in combination with any of the tested
compounds. The tested resin acids are diterpenoids which are found in oleoresin
produced by conifers. They are known to have low toxicity toward fungal pathogens
but are of importance in sealing wounds and trapping insect invaders (Phillips and
Croteau, 1999). Abietic acid and its derivatives have also been found to show
antifungal activity against Fusarium moniliforme and Aphanomyces euteiches
(Spessard et a l, 1995). Moreover, there is evidence that resins are constitutively
produced and stored in conifers and are part o f their defence mechanisms towards
insect and fungi invaders (Lewinsohn et al., 1991). In some conifers, resin ducts are
99

produced as an inducible defence mechanism, including western red cedar in response
to Armillaria ostoyae or European larch in response to physical injury (Bollschweiler
et a l, 2007; Cleary, 2010).
Based on this theory, the red discoloured bark o f Japanese larch suspected o f being a
defence reaction toward P. ramorum, was chemically analysed to provide evidence o f
compounds induced during pathogen invasion. The analysis o f the DCM crude
extracts revealed that prior to detection o f the pathogen in invaded bark, white phloem
tissue has an abundance o f resin acid 2 (dehydroabietic acid) and much lower
amounts o f resin acid 1 (methyl abietate, an AB derivative) compared with the
phloem from the healthy tissue (A 2 o,2 i)- However, in the crude extract o f the red-
discoloured tissue where the pathogen could not be detected, methyl abietate and
DHAA were both present in considerable quantities, whereas in the brown tissue
where detection o f the pathogen was successful, DHAA had reduced to levels similar
to the control. It was also striking that particularly the red discoloured tissue had
raised levels o f many other compounds when compared with healthy and white
coloured phloem tissue. One other noticeable change in the chemical components o f
discoloured bark was the presence o f a-pinene and 3-carene in the extract o f red and
dark brown phloem (A2 2 ,2 3 ). A similar phenomenon was reported after artificial
inoculation o f pine seedlings ^vith six different strains o f fungi which resulted in
higher biosynthesis o f volatile compounds especially a-pinene and 3-carene
(Napierala Filipiak et a l, 2002). Both compounds are monoterpenes found in the
essential oils o f conifers that exhibit toxic activity against fimgi and insects (Phillips
and Croteau, 1999). These results suggest that resin acids may play an important role
in inhibiting the growth o f P. ramorum after invasion but the overall inhibitory effect
could potentially be due to the presence o f other antimicrobial compounds that are
induced in small quantities in conjunction with higher concentrations o f resin acids.
Results o f the first field trial using heat-treated and air-dried woodchips were
consistent with the bioassay results o f crude extracts. A t fifty-two weeks after the
placement o f the bags o f woodchips on site, there was a more than two-fold increase
in recovery o f P. ramorum from air-dried larch and pine compared with heat-treated
samples. In contrast, P. ramorum was isolated from both heat treated and air-dried
rhododendron wood chips after 52 weeks, but at lower levels from the heat treated
woodchips although this was not significantly different from the air-dried woodchips.
100

The outcome can be explained by the results of the chemical analysis that revealed the
lack of coniferaldehyde in rhododendron wood which had the greatest inhibitory
effect in bioassays of isolated compounds. The crude extract of heat treated wood of
rhododendron also contained a lower concentration of vanillin in eomparison with
larch and rhododendron. Another major factor could be the lack o f diterpenoid resin
acids that naturally exist in conifers and were shown to considerably decrease the
MIC of active compounds. An inhibitory effect of the heated crude extract of
rhododendron was achieved at a high concentration of 50 mg ml'^ in bioassay tests, so
it can be concluded this was due to additive effect of the complex mixture of
compounds found in the crude extract. These are probably less effective under high
inoculums level of P. ramorum which can occur at certain time of year under natural
conditions. The lack o f differences between air-dried and heated woodchips after 4
and 1 2 weeks after natural exposure could also be explained by low levels of
inoculum at that time of sampling during the field trail.
In the second field trial, traditional baiting combined with a PCR methods using m-
RNA to detect live cells of P. ramorum was used to improve detection. However, the
field trial still failed to provide information about the inhibitory effect o f heated
materials apparently because too little natural inoculum was generated during the trial.
To determine the minimum detection limit of the RT-PCR assay, a dilution series
between 1 to 1,000,000 zoospores ml'^ in sterile water was tested. The minimum
detection limit was 10"̂ zoospore in 1 ml of water for the RT-PCR assay, suggesting
that the amount o f inoculum sampled from the field exposed woodchips was often
below the detection limit of tested PCR assay. Although it seems unlikely that the
negative results of the baiting method were also associated with a lack o f spores, an
alternative method of detecting inoculums could improve the chance o f comparing
treatments more effectively.
Extensive research on the management of P. ramorum indicates that there is no single
strategy to eradicate the pathogen once it has already established in the wider
environment. The best management praetices are focused on slowing the spread o f the
disease particularly in woodland and forest, but bio-security and quarantine efforts are
more difficult to implement successfully in these environments. As the spores are the
key factor in pathogen survival and spread, methods which can reduce sporulation and
dispersal are potentially the most effective. One strategy that has, been suggested is to
101

cover infected leaf litter on the floor o f a contaminated area by a layer o f woodchips
to reduce the movement of inoculum and prevent spread to nearby susceptible plants
(Goheen et a l, 2012; Manter et a l, 2007). This has potential to be effective in places
where human activities are concentrated, such as roads, walk ways in woodlands and
paths in parks and gardens. Moreover, use of woodchips material with antimicrobial
properties could give added protection if used as a part of disease management
practices. A research study by Manter et a l, (2007) evaluating antimicrobial activity
of yellow cedar against P. ramorum showed consistency o f bioassay results of
chemical components compared with woodchips used in field treatments. Similarly,
results of the present study showed heat-treated wood of larch and pine exhibit
inhibitory effect against the pathogen under natural conditions as well as laboratory
experiments. These results suggest that using heat treated materials has the potential
to be an effective method to reduce survival of the pathogen under natural conditions.
Moreover, the active chemicals of heated material can be the potential candidates for
further studies in the development of chemicals treatment for sudden oak disease.
102

C H APTER 6: DISCUSSIO N
Despite the wide application of heating techniques to improve the way that wood
materials are used in sectors such as construction, wood preservation and food
paekaging, a review of the literature revealed that there has been little attention to the
use of heat treatment in other areas such as animal bedding or disease control and
management in agriculture or forestry. Therefore, the studies reported here aimed to
investigate the potential of a heating technique to enhance the antimicrobial activity of
wood materials with potential application in two different fields: animal husbandry
and forestry.
The first objective of this study was to optimise a heating regime for pine wood
shaving materials. A preliminary study indicated that pre-heated wood shavings can
inhibit an important food-borne pathogen of chickens. Salmonella Enteritidis. The
second objective was to investigate and evaluate the effect o f enhanced inhibitory
activity on ammonia emission of heated wood materials in poultry bedding. The
investigation of inhibitory activity was performed by extraction of heat-treated and
air-dried wood materials using organic solvents and water. Chemical analysis
techniques (GC-MS and NMR spectroscopy) were employed to study the chemical
modification o f heat-treated pine wood shavings and its association with any new
inhibitory effect in a bio-active methanol extract. The dominant chemicals derived
from fractionation o f methanol extract were tested for their antimicrobial activity on
Salmonella Enteritidis, but the wider spectrum of antimicrobial activity was also
tested against other poultry bedding microorganisms including: Staphylococcus
aureus, E. coli and Saccharomyces cerevisiae. In the final part o f the study, the
application of heat-treated wood materials was extended to Phytophthora ramorum as
a potential control for this invasive pathogen of larch in UK forests. The study
consisted of applying the heating technique to wood materials o f Japanese larch and
rhododendron, the two main hosts of P. ramorum. The potential inhibitory effect o f
the heat-treated materials on P. ramorum was studied and tested by chemical analysis
techniques and bioassay development using some of the approaches developed for
poultry bedding and Salmonella Enteritidis.
To expand on the findings of a preliminary study which demonstrated that heated pine
wood shavings exhibit antimicrobial activity against Salmonella, temperatures
ranging from 20°C to 200°C was tested to evaluate and optimize the effect to enhance
103

antimicrobial activity. Interestingly, raising the temperature (from 50°C to 200°C)
and the length of time of heat treatment increased the inhibitory effect o f pine wood
against Salmonella Enteritidis. Further evaluation studies revealed a heating regime
o f 120°C for 72 hours reduced the number o f Salmonella recovered from inoculated
heated wood shavings and also reduced levels o f ammonia produced in the chicken
litter over at least 11 days. In addition, hardwoods such as beech and ash exhibited
antimicrobial activity against Salmonella Enteritidis as well as softwoods such as pine
and cypress, indicating the potential o f heat treatment to produce a range of suitable
poultry bedding materials in response to shortages in the market by introducing
antimicrobial properties across a wide range of wood species. However, this study
did not investigate the efficacy o f heated wood materials in the natural environment of
a broilers house, but it is worth noting that the heating technique also reduced the pH
of pine wood. This process could be used to produce bedding which is a much less
favourable environment for Salmonella.
In investigating the potential of using similar heating techniques to produce
antimicrobial woodchips against P. ramorum, heated wood of two susceptible hosts,
larch and rhododendron was found to inhibit P. ramorum zoospores when in direct
contact. Therefore, heat-treated larch wood showed promise for further evaluation in
the laboratory experiments. Heat-treated larch wood in mixtures with soil was also
effective at inhibiting P. ramorum zoospores and chlamydospores, indicating the
potential of the inhibitory effect can be extended to an ecologically more complex
environment. The field evaluation (Largin Wood, Cornwall) using pine and larch
woodchips indicated that the inhibitory activity brought about by heating could be
effective in reducing the recovery of P. ramorum from retrieved wood samples left in
the field for several weeks. In comparison, rhododendron woodchips were not as
effective as larch or pine even after heat treatment. Overall, the results suggested that
pressure of high inoculum in the field may affect the effectiveness of inhibition for
different wood species, and this outcome was later found to be associated with
differences in the chemical content of each wood species.
In identifying the chemical components responsible for the antimicrobial activity in
pine, larch and rhododendron, heated wood materials were extracted and the methanol
crude extracts showed antimicrobial activity in in vitro assays. Antimicrobial activity
was associated with the presence of aromatic compounds and an increase in more
104

saturated types of resin acid in the heat-treated wood. Chemical analysis of the active
methanol crude extract of heat treated wood of pine showed that it contains three
different groups of chemicals; a) aromatic compounds including benzaldehyde,
coniferaldehyde, aeetovanillone, vanillin and stilbene; b) organic acids including
formic acid, acetic acid, and vanillic acid; c) diterpenoid resin acids including
pimaric, abietic and dehydroabietic acid. Vanillin and coniferaldehyde were the
dominant compounds of the methanol crude extracts of pine and larch and are known
to be formed as a result o f lignin breakdown. They also occur naturally in essential
oils, herbs and spices and have previously been reported to inhibit S. Enteritidis, E.
coli, and S. aureus (Seydim and Sarikus, 2006). Extracted from vanilla beans,
vanillin exhibits several bioactive properties including antimicrobial activity against
yeasts, moulds and bacteria including Salmonella Enteritidis (Fitzgerald, 2004;
Mourtzinos et ah, 2009). Coniferaldehyde gives whisky and wine their fruity flavour
and has antioxidant and anticancer properties (Connors et a l, 1970). Both
coniferaldehyde and vanillin have previously been found to exhibit antifungal activity
against the wood decay fungi Trametes versicolor and Coniphora puteana (Voda et
a l, 2003). The overall results of bioassay tests of the fractionated compounds and
mixtures derived from the methanol crude extract in the present study demonstrated
however, that the enhanced antimicrobial effect was not usually due to one individual
compound. Fractions containing aromatic compounds and resin acids including
dehydroabietic acid and abietic acid showed a stronger effect on the tested pathogens
than when used singly. For example vanillin and stillbene were less effective when
they were tested individually on Salmonella Enteritidis and Staphylococcus aureus,
respectively. Similarly when investigating the role o f chemical components of heat-
treated larch and rhododendron and their inhibitory properties on P. ramorum,
coniferaldehyde, vanillin, aeetovanillone and resin acids including abietic and
dehydroabietic acid, were found to work best in combination. Dehydroabietic acid
was identified as the dominant compound present in active fractions containing
vanillin and stillbene. It is interesting that the resin acids dehydroabietic acid or
abietic acid had no inhibitory effect on Salmonella or P. ramorum when tested alone,
suggesting that mixtures of the active compounds and resin acids synergize to produce
a greater level of activity against both pathogens. An extensive literature review did
not reveal any similar findings so this is the first report of synergistic effect between
aromatic compounds and dehydroabietic or abietic acid.
105

These findings appear to be further supported by the results of chemical analysis of
discoloured larch bark in the present study. This showed that the increase in resin acid
content of bark is followed by the production of new compounds, a-pinene and 3-
carene, in the extract o f red and dark brown phloem which are formed after invasion
by P. ramorum. Both compounds are monoterpenes found in essential oils of conifers
that exhibit toxic activity against fungi and insects (Phillips and Croteau, 1999).
Furthermore, in some conifers, resin ducts are produced as an inducible defence
meehanism, such as in western red cedar in response to Armillaria ostoyae infection
(Cleary, 2010). Also higher concentrations of resin acids have been reported in Scots
pine heartwood which is decay-resistant to brown-rot, with abietic acid being the
most abundant (Harju et a l, 2005). It is notable that the complex composition of
oleoresin, which is known to be an important part of their successful defence
mechanism of conifer species against wide range of pests and pathogens, contains
both resin acids and monoterpenes (Lewinsohn et a l, 1991). The composition of
oleoresin from conifers has also been shown to contain significant amounts o f resin
acids (60-70%).
The results of bioassay tests of the chemical constituents o f heat treated wood on P.
ramorum may also suggest that the aromatic content o f different wood species
(particularly phenols) work in conjunction with resin acids to induce resistance
against different pathogens. A previous study conducted by Nagle et al. (2011)
showed that oak trees with field-resistance to P. ramorum contained high levels of
phenolic components compared with susceptible trees. In contrast, the inner bark of
conifers with higher phenolic content can be more susceptible to pathogen
colonization and wood decay (Kelsey and Harmon, 1989). This may indicate that the
phenolic content of conifers is not the only chemical constituent o f their defence
mechanisms. A research study by Harris and Webber (unpublished work) on the
susceptibility of larch to P. ramorum examined colonization o f Japanese, hybrid and
European larch by the pathogen, and found that at certain times of year, Japanese and
hybrid larch were more resistant than European larch. Research on the chemieal
constituents of the three species of larch indicates that Japanese and hybrid larch both
contain more resin acids, mainly abietic acid, compared with European larch. There
is evidence that resins are constitutively produced and stored in conifers and are part
o f their defence mechanisms toward insect and fungi invaders (Lewinsohn et a l.
106

1991). Abietic acid is known to have low toxicity toward fungal pathogens but is of
great importance in sealing wounds and trapping insect invaders (Phillips and
Croteau, 1999). It has also been shown to have phytoalexin properties which appear
to have an important role in inducing resistance in plant hosts. In addition,
dehydroabietic acid, the dominant resin acid present in heated crude extract of pine
and larch, has shown antifungal activity against Trametes versicolor and Trichoderma
species. Therefore, presence of these resin acids may explain the difficulty of
isolating P. ramorum from bark of infected larch species. However, unexpectedly,
Japanese larch has proved more susceptible to P. ramorum in the natural environment
in the UK, despite higher levels o f resin acids. This may be because infected foliage
of Japanese larch produces large numbers of spores and counteracts the inhibitory
effectors o f the resins acids. Alternatively, it could be explained by insufficient
concentrations o f resin acids produced by Japanese larch growing under UK
environmental conditions. To produce resin acids, conifer species need adequate
sunlight and this or other environmental conditions may play a significant part in tree
susceptibility in ways that are not yet fully understood.
Although, the mechanism of action of bio-active resin acids is not well understood,
research studies hypothesize that their antibacterial and antifungal properties result
from their interaction with the cell membrane of the target microorganism (Urzua et
a l, 2008) (Figure 6.1). The lipophilicity of these chemicals is thought to be
associated with their relative activity in the disruption of cell membranes
(Savluchinske-Feio et a l, 2006). Higher lipophilicity means a higher concentration of
the chemicals are able to bind with the cell membrane layer, resulting in stronger
activity. Resin acids with a high lipophilicity are used as the starting material in the
development of surfactants (Piispanen et a l, 2002). These chemicals act as soluble
surfactants in alkaline conditions, but in acidic conditions they only dissociate partly
depending on their pK values (Werker and Hall, 1997). They also are less lipophilic
and more water soluble under acidic conditions. This suggests that the mechanism by
which dehydroabietic acid/aldehyde showed greater antimicrobial activity in this
study was not due to dehydroabietic acid interfering with cell membrane function.
Instead, because the effect of temperature on heat-treated wood showed the pH of
pine could reduce to a value of 3.5 after 72 hours, the effect could be due to the acidic
107

conditions. On the other hand, vanillin showed antimicrobial activity against
Salmonella, but it was dose dependent and the activity was lost by dilution.
W (Hin€llllg
POG
0 0
/ \
Figure 6.1 Interactions o f secondary m etabolites w ith biom em brane and m em brane proteins.1, Sarsaparilloside (a steroidal saponine), 2, ocim ene (linear m onoterpene), 3, a -p inene (bicyclic m onoterpene), 4, cadinene (sesquiterpene), 5, phyllocladene (diterpene); (A) ion channel, (B) transporter, (C) m em brane receptor (W ink, 2006).
Currently, the mode of action of vanillin is unclear, although other phenolic aldehydes
act by disrupting cell membrane function (Fitzgerald et a l, 2004). Therefore, it can
be hypothesized that vanillin disrupts the cell membrane and allows dehydroabietic
acid to pass through cell membrane which may affect metabolic processes inside the
cell. The weak solubility of resin acids, which causes variability in the results of their
bioassay outcome, supports the idea that if disrupting membrane function is facilitated
by cell membrane active compounds such as vanillin, they should be more effective.
This also can be extended to the strong antimicrobial activity of coniferaldehyde/
DHHA mixture against P. ramorum. Grabber et al. (1998) demonstrated that severe
inhibition of wall degradation occurred with coniferaldehyde, using a cell-wall model
system to evaluate what effect aldehyde-containing lignins have on the hydrolysis of
maize cell walls by fungal enzymes (Grabber et a l, 1998). These findings are
important as they bring insight to the evaluation and application of resin acids and
their unique potential in development of antimicrobial pesticides and drug design.
108

The results o f the first field trial using heat-treated and air-dried woodchips on P.
ramorum were consistent with the bioassay results of crude extracts, providing further
evidence in the role of dominant aldehyde/resin acids on the inherent inhibitory
activity of heat-treated wood. The lack of coniferaldehyde which had the greatest
inhibitory effect in bioassays o f isolated compounds in heat-treated rhododendron
together with the lack o f diterpenoid resin acids (both major differences revealed by
chemical analysis) could explain the insignificant effect o f heat-treatment on
rhododendron in both laboratory experiments and also under field conditions. These
findings may indicate that all types of wood treated materials (softwood or hardwood)
can exhibit antimicrobial effects after heat-treatment because of the high content of
active aromatic compounds, but the level of activity may change under environmental
circumstances. Therefore, softwoods which contain diterpenoid resins could provide
stronger activity over a longer period.
In the latest review of wood waste carried out by DEFRA, in 2012, approximately 4.1
million tonnes of wood material was transferred into the waste stream in the UK
(Defra, 2012). O f this, the market for animal bedding materials from the UK total
wood machine waste usage was 56% which is estimated to be approximately 400,000
tons of recycled materials (Fletcher et a l, 2010). Results of this study suggest that,
heating wood shavings o f pine (I20°C for 72 hours) is potentially an effective
technique to produce antimicrobial wood materials for use as bedding in poultry units
and it could return larger amounts of wood waste into new uses. However, it is not
easy to estimate the efficiency level because o f the potential running costs of various
methods of heating. In addition, the efficiency of these materials still needs to be
tested for their effect in a real broiler environment for final evaluation. The benefits
gained from heat treatment of wood materials aimed as broiler bedding with improved
control of Salmonella and ammonia production could improve the life standards of
chickens and quality of poultry products.
Heat-treated wood of larch and pine was shown to exhibit an inhibitory effect against
P. ramorum under natural as well as under laboratory conditions. Use o f such
material in P. ramorum affected areas has the potential to be very effective in places
where human activities are concentrated such as roads, walk ways in woodlands, as
well as paths in parks and gardens. To prevent transfer of spores by people, use of
woodchip material with enhanced antimicrobial properties could give added
109

protection as a part o f disease management practices. Finally, the active chemicals of
heated material can yield potential candidates for further studies in the development
of chemicals for treatment of sudden oak death. It is also worth noting that large
quantities of wood waste are unrecyclable because of the bonding resins they contain.
(Fletcher et a l, 2010), so finding uses for this material would be economically and
environmentally useful.
Future work
The results of this study demonstrated that the heating technique made different wood
species antimicrobial against a number of organisms. Pine wood shavings showed
antimicrobial activity against Salmonella in both laboratory experiments and also
microcosom systems. Further studies are required to investigate the true value of
antimicrobial wood shavings under the natural environment o f broilers units. To be
able to evaluate the potential of antimicrobial wood shaving, trials using recycled pine
that is available in the market as poultry bedding should be set and run in parallel for
comparison. Also the true potential o f heated wood materials in reducing ammonia
production in poultry litter should be tested in a real broiler system. In this study
there was insufficient time for the potential o f other heated wood species to reduce
ammonia production to be investigated. It would be interesting to examine the wood
species that showed antimicrobial activity against Salmonella for their effect on
ammonia production.
Based on the results of this study, the pH of wood decreased to a value of 3.2 after the
heat treatment. The presence o f organic acids such as formic, acetic and vanillic acid
in the heated materials gives heated wood its acidic pH and may also be used to
advantage for controlling Salmonella in broilers units. Acetic acid, formic acid, lactic
and propionic acids are already commonly used to suppress Salmonella in animal
feeds (Martin and Maris, 2005). In addition, the low pH o f heated wood materials
may reduce the need for litter amendments to control pH of the litter. It may also help
to reduce the risk of emerging resistant pathogens of poultry which is believed to be
driven by exposure to excessive amount of chemicals used to control litter conditions.
One approach therefore might be to undertake comparative studies to evaluate which
of the heated wood species or acidifying amendments are more effective in
controlling litter pH.
110

Based on results of in vitro assays in this study, the magnitude of antimicrobial
activity was due to the presence of bio active diterpenoid resin acids operating in
conjunction with phenolic aldehydes in softwoods such as pine and larch. It would be
necessary to investigate the mechanism of this antimicrobial activity further. This
study hypothesized that the synergism between vanillin and dehydroabietic acid may
be explained by the role of vanillin in disrupting cell membrane and therefore
facilitating dehydroabietic acid transition through the cell membrane. A better
understanding of interactions between mixtures of bioactive compounds and their
target sites would provide important information on extending the application o f both
aldehydes and the resin acids. Further studies could also be extended to the
evaluation of antimicrobial activity o f other cell membrane active compounds in
combination with the resin acids. Using cell membrane active compounds could help
to reveal the potential of other antimicrobials which cannot actively pass through the
cell membrane. As such, there is no report on the antimicrobial activity of
dehydroabietic acid on Gram negatives, which in mixture with vanillin, exhibited
strong antimicrobial effects in the present study.
Finally, apart from antibacterial and antifungal activity, abietane and dehydroabietane
derivatives have been found to have antiviral and antitumor properties, although only
oxidative forms are found to have cytotoxicity. More o f their oxidative derivatives
are synthesized and investigated as part o f screening new bioactive compounds with
antiviral and anticancer properties (G onzalez et a l, 2010). Therefore, another
approach in taking advantage of their unique potential could be to investigate their
true value in combination with cell membrane active compounds. This also may
encourage further studies in the formulation o f bioactive compounds in the search for
new antimicrobials and anticancer drugs.
I l l

CHAPTER 7; REFERENCES
Aktan, S., Sagdic, O., 2004, Dried rose {Rosa damascena Mill.) dreg: an alternative litter material in broiler production. S Afr J Anim Sci 34, 75-79.
American-Chemistry-Council, 2010. Benefits of antimicrobial pesticides in public health and industrial uses. Biocides Panel, American Chemistry Council, Arlington, U.S.A, p. 28.
Angulo, F. J., Nunnery, J. A., Bair, H. D., 2004. Antimicrobial resistance in zoonotic enteric pathogens. Rev Sci Tech Oie 23, 485-496.
Apsimon, H. M., Kruse, M., Bell, J. N. B., 1987. Ammonia Emissions and Their Role in Acid Deposition. Atmos Environ 21,1939-1946.
Atapattu, N. S. B. M., Senaratna, D., Belpagodagamage, U. D., 2008. Comparison of Ammonia Emission Rates from Three Types of Broiler Litters. Poultry Sci 87,2436-2440.
Atencio, J. L., Fernandez, J. A., Gemat, A. G., Murillo, J. G., 2010. Effect of pine wood shavings, rice hulls and river bed sand on broiler productivity when used as a litter sources. International Journal of Poultry Science 9, 240-243.
Aveskamp, M. M., van Baal, P. J. M., de Gruyter, J., 2006. Effect of sanitary measures on the survival of Phytophthora ramorwn in soil. In: Frankel, S. J. S., P.J.; Haverty, M.I (Ed.), The sudden oak death second science symposium: the state of our knowledge. Department of Agriculture, Forest Service, Pacific Southwest Research Station, Albany, CA: U.S., pp. 279- 2M.
Bacharach, T., Evans, J. R., 1957. Enlargement of the Colon Secondary to Hypothyroidism. Ann Intern Med 47,121-124.
Beyer, G., Gudbjomsdottir, B., 2002. Wood in the food industry - guidelines for handling wooden pallets and packaging. Project part report no. 8. Nordisk Industrifonc.
Beyer, M., Koch, H., Fischer, K., 2006. Role of hemicelluloses in the formation of chromophores during heat treatment of bleached chemical pulps. Macromol Symp 232, 98- 106.
Bilgili, S. P., Montenegro, G. I., Hess, J. B., Eckman, M. K., 1999. Sand as litter for rearing broiler chickens. J Appl Poultry Res 8, 345-351.
Bollschweiler, M., Stoffel, M., Schneuwly, D. M., Bourqui, K., 2007. Traumatic resin ducts in Larix decidua stems impacted by debris flows. Tree Physiology 28 255-263.
Boogert, P., Kessel, G., 2004. Assessment of foci in natural habitats: Survival of Phytophthora ramorum in wood chipped materials. Project report (2004-2007), pp. 123-124.
Brasier, C., 2003. Sudden oak death: Phytophthora ramorum exhibits transatlantic differences. Mycol Res 107,258-259.
Brasier, C. M., Denman, S., Rose, J., Kirk, S. A., Hughes, K. J. D., Griffin, R. L., Lane, C. R., Inman, A. J., Webber, J. F., 2004. First report of ramorum bleeding canker on Quercus falcata, caused by Phytophthora ramorum. Plant Pathol 53, 804-804.
Brasier, C. M., Jung, T., 2006. Recent developments in Phytophthora diseases of trees and natural ecosystems in Europe. In: Brasier, C. M. J., T.; Osswald, W., (Ed.), The third International Union of Forest Research Organization (lUFRO) Working Party 07.02.09., Famham, UK: Forest Research Agency pp. 5-16.
112

Brasier, C. M., Rose, J., Kirk, S. A., Webber, J., 2002. Pathogenicity of Phytophthora ramorum isolates from North America and Europe to bark of European Fagaceae, American Quercus rubra, and other forest trees. In: Proceedings of the sudden oak death science symposium. Department of Agriculture, Forest Service and University of California, Berkeley, Berkeley, CA: U.S.
Brasier, C. M., Webber, J. F., 2010. Sudden larch death. Nature 466, 824-825.
Brenner, F. W., Villar, R. G., Angulo, F. J., Tauxe, R., Swaminathan, B., 2000. Salmonella nomenclature - Guest commentary. J Clin Microbiol 38, 2465-2467.
Brown, A., Brasier, C., Denman, S., Rose, J., Kirk, S., Webber, J., 2006. Distribution and etiology of aerial stem infections of Phytophthora ramorum and Phytophthora kernoviae at three woodland sites in the U.K. In: Frankel, S. J. S., P.J.; Haverty, M.I., (Ed.), The sudden oak death second science symposium: the state of our knowledge. Department of Agriculture, Forest Service. Pacific Southwest Research Station:, Albany, CA: U.S., pp. 105-108.
Brown, A. V., Brasier, C. M., 2007. Colonization of tree xylem by Phytophthora ramorum, P. kernoviae and other Phytophthora species. Plant Pathol 56, 227-241.
Brunow, G., Kilpelainen, I., Sipila, J., Syijanen, K., Karhunen, P., Setala, H., Rummakko, P., 1998. Oxidative coupling of phenols and the biosynthesis of lignin. Lignin and lignan biosynthesis. Amercian Chemical Society, pp. 131-147.
Carlile, F. S., 1984. Ammonia in Poultry Houses - a Literature-Review. World Poultry Sci J 40, 99-113.
Carr, L. E., Mallinson, E. T., Tate, C. R., Miller, R. G., Russekcohen, E., Stewart, L. E., Opara, O. O., Joseph, S. W., 1995. Prevalence of Salmonella in Broiler Flocks - Effect of Litter Water Activity, House Construction, and Watering Devices. Avian Dis 39, 39-44.
Carr, L. E., Wheaton, F. W., Douglass, L. W., 1990. Empirical-Models to Determine Ammonia Concentrations from Broiler Chicken Litter. T Asae 33,1337-1342.
Castiglioni Tessari, E. N., Kanashiro, A. M. I., Stoppa, G. F. Z., Luciano, R. L., Castro, A. G. M. D., Cardoso, A. L. S. P., 2012. Salmonella - A dangerous foodbome pathogen: Important aspects of Salmonella in the poultry industry and in public health. InTech, DOI: 10.5772/1308.
CCFH-Working-Group, 2007. Food safely risk profile for Salmonella species in broiler (young) chickens. Food and Agriculture Organization, p. 30.
Céspedes, C. L., Avila, J. G., Marin, J. C., Dominguez, M., Torres, P., Aranda, E., 2006. Natural compounds as antioxidant and molting inhibitors can play a role as a model for search of new botanical pesticides. Adv Phytomed 3,1-27.
Chambers, J. R., Lu, X., 2002. Probiotics and maternal vaccination for Salmonella control inbroiler chickens. J Appl Poultry Res 11, 320-327.
Chastagner, G., DeBauw, A., Riley, K., 2010. Effect of fungicides on the isolation ofPhytophthora ramorum from symptomatic and asymptomatic rhododendron leaf tissue. In: Frankel, S. J. K., J.T.; Palmieri, K.M. (Ed.), The sudden oak death fourth science symposium. Department of Agriculture, Forest Service, Pacific Southwest Research Station, Albany, CA: U.S., pp. 302-304.
Chastagner, G., Riley, K., Elliott, E., 2012. Susceptibility of Larch, Hemlock, Sitka Spruce, and Douglas-fir to Phytophthora ramorum. Proceedings of the Sudden Oak Death Fifth
113

Science Symposium. Pacific Southwest Research Station, Forest Service, Petaluma, California, p. 169.
Chimento, A., Cacciola, S. O., Garbelotto, M., 2012. Detection of mRNA by reverse- transcription PCR as an indicator of viability in Phytophthora ramorum. Forest Pathol 42, 14- 2 1 .
Choi, I. H., Kim, J. N., Kwon, Y. M., 2008. Effects of chemical treatments on pH and bacterial population in poultry litter: a laboratory experiment. Brit Poultry Sci 49, 497-501.
Chung, K. C., Goepfert, J. M., 1970. Growth of Salmonella at low pH. J Food Sci 35, 326.
Cleary, M. R., 2010. Traumatic resin duct formation in the phloem of western red cedar and other resistance mechanisms effective against Armillaria root disease The International Symposium - A Tale of Two Cedars: Western redcedar and yellow-cedar Forest Service Pacific Northwest Research Station, University of Victoria, Victoria British Columbia, pp. 53-56.
Cogan, T. A., Humphrey, T. J., 2003. The rise and fall of Salmonella Enteritidis in the UK. J Appl Microbiol 94 ,114s-119.
Colburn, G. C., Jeffers, S. N., 2010. Efficacy of commercial algaecides to manage species of Phytophthora in suburban waterways. In: Frankel, S. J. K., J.T.; Palmieri, K.M. (Ed.), The sudden oak death fourth science symposium. Department of Agriculture, Forest Service, Pacific Southwest Research Station, Albany, CA: U.S., pp. 223-224.
Condeso, T. E., Meentemeyer, R. K., 2007. Effects of landscape heterogeneity on the emerging forest disease sudden oak death. Journal of Ecology 95, 364-375.
Connors, W. J., Chen, C., Pew, J. C., 1970. Enzymic dehydrogenation of the lignin model coniferaldehyde. Journal of Organic Chemistry 35,1920-1924.
Cook, K. L., Rothrock, M. J., Warren, J. G., Sistani, K. R., Moore, P. A., 2008. Effect of alum treatment on the concentration of total and ureolytic microorganisms in poultry litter. J Environ Qual 37, 2360-2367.
Cowan, M. M., 1999. Plant products as antimicrobial agents. Clin Microbiol Rev 12, 564-+.
Cox, J. M., Pavic, A., 2010. Advances in enteropathogen control in poultry production. J Appl Microbiol 108, 745-755.
Crane, R., Davenport, R., Vaughan, R., 2010. Farm Business Survey 2009/2010 Poultry Production in England. Agriculture and Food Investigation Team (AFIT). Rural Business Research, University of Reading, p. 52.
Cushman, J. H., Cooper, M., Meentemeyer, R. K., Benson, S., 2008. Human activity and the spread of Phytophthora ramorum. In: Frankel, S. J. K., J.T.; Palmieri, K.M. (Ed.), The sudden oak death third science symposium. Department of Agriculture, Forest Service, Pacific Southwest Research Station, Albany, CA: U.S., pp. 179-180.
Cushman, J. H., Meentemeyer, R., 2006. The importance of humans in the dispersal and spread of Phytophthora ramorum at local, landscape, and regional scales. Sudden oak death second science symposium: the state of our knowledge. Department of Agriculture, Forest Service. Pacific Southwest Research Station, Albany, CA: U.S. Pacific Southwest Research Station, pp. 161-163.
Davidson, J. M., Maloney, P. E., Wickland, A. C., Morse, A. C., Jensen, C. E., Rizzo, D. M., 2002. Transmission of Phytophthora ramorum via Umbellularia californica (California Bay)
114

leaves in California oak woodlands. Proceeding of he sudden oak death science symposium: the state of our knowledge. Department of Agriculture, Forest Service and University of California, Berkeley, Berkeley, CA: U.S., p. 571.
Davidson, J. M., Shaw, C. G., 2003. Pathways of movement for Phytophthora ramorum, the causal agent of sudden oak death. Sudden oak death online symposium, http://www.forestphvtophthoras.org/references/pathwavs-movement-phvtophthora-ramorum- causal-agent-sudden-oak-death.
Davidson, J. M., Wickland, A. C., Patterson, H. A., Falk, K. R., Rizzo, D. M., 2005a. Transmission of Phytophthora ramorum in mixed-evergreen forest in California. Phytopathology 95, 587-596.
Davidson, P. M., Sotos, J. N., Branen, L. A., 2005b. Antimicrobials in food. CRC Press, http://www.crcnetbase.eom/isbn/9781420028737.
Davies, R. H., 2005. Pathogen populations on poultry farms. Food Safety Control in the Poultry Industry, 101-152.
Dawson, P., Han, I., Cox, M., Black, C., Simmons, L., 2007. Residence time and food contact time effects on transferof Salmonella Typhimurium from tile, wood and carpet: testing the five-second rule. J Appl Microbiol 102, 945-953.
De Rezende, C. L. E., Mallinson, E. T., Tablante, N. L., Morales, R., Park, A., Carr, L. E., Joseph, S. W., 2001. Effect of dry litter and airflow in reducing Salmonella and Escherichia coli populations in the broiler production environment. J Appl Poultry Res 10, 245-251.
Defra, 2008. Phytophthora ramorum. A Practical Guide for Established Parks &Gardens, Amenity Landscape and Woodland Areas. Department for Environment, Food and Rural Affairs, London.
Defra, 2011. Disease nitigation and control evidence plan 2011. Department for Environment Food and Rural Affairs, Richmond, Surrey.
Defra, 2012. Wood waste: A short review of recent research. Department for Environment, Food and Rural Affairs, London.
DeFrancesco, K. A., Cobbold, R. N., Rice, D. H., Besser, T. E., Hancock, D. D., 2004. Antimicrobial resistance of commensal Escherichia coli from dairy cattle associated with recent multi-resistant salmonellosis outbreaks. Vet Microbiol 98, 55-61.
Denman, S., Kirk, S. A., Brasier, C. M., Barton, V. C., Hughes, K. J. D., Webber, J. F., 2005a. Phytophthora ramorum on Quercus ilex in the United Kingdom. Plant Dis 89, 1241- 1241.
Denman, S., Kirk, S. A., Brasier, C. M., Hughes, K. J. D., Griffin, R., Hobdon, E., Webber, J. F., 2005b. Foliar infection of sweet chestnut (Castanea sativa) by Phytophthora ramorum in the UK. Plant Pathol 54, 581-581.
Denman, S., Kirk, S. A., Brasier, C. M., Webber, J. F., 2005c. In vitro leaf inoculation studies as an indication of tree foliage susceptibility to Phytophthora ramorum in the UK. Plant Pathol 54,512-521.
Donham, K. J., 1996. Air quality relationships to occupational health in the poultry industry. Proceedings of the 1996 national poultry waste management symposium. Auburn University, pp. 24-28.
115

Dougherty, T. J., 1976. Study of Salmonella Contamination in Broiler Flocks. Poultry Sci 55, 1811-1815.
Doyle, M. P., Erickson, M. C., 2006. Reducing the carriage of foodbome pathogens in livestock and poultry. Poultry Sci 85, 960-973.
Doyle, M. P., Erickson, M. C., 2012. Opportunities for mitigating pathogen contamination during on-farm food production. Int J Food Microbiol 152, 54-74.
Dumas, M. D., Poison, S. W., Ritter, D., Ravel, J., Gelb, J., Morgan, R., Wommack, K. E., 2011. Impacts of Poultry House Environment on Poultry Litter Bacterial Community Composition. Plos One 6, e24785.
Duqueza, M. C., 1996. Evaluation of nitrogen bioavailability indices for poultry wastes vol. PhD. Auburn University, p. 102.
Dutta, S., De, S., Saha, B., Alam, M., 2012. Advances in conversion of hemicellulosic biomass to furfural and upgrading to biofuels. Catal. Sci. Technol. 2, 2025-2036.
EFSA, 2010a. The European Union summary report on trends and source of zoonoses, zoonotic agent and food-bome outbreaks in 2010. European Food Safety Authority Journal 10,2597.
EFSA, 2010b. The European Union summary report on trends and source of zoonoses, zoonotic agent and food-borne outbreaks in 2010. European Food Safety Authority Journal 10, e2597.
EFSAP-PLH, 2011. Scientific Opinion on the Pest Risk Analysis on Phytophthora ramorum prepared by the FP6 project RAPRA. European Food Safety Authority Journal 9 .108.
Eloff, J. N., Famakin, J. O., Katerere, D. R. P., 2005. Isolation of an antibacterial stilbene from Combretum woodii (Combretaceae) leaves. Afr J Biotechnol 4,1167-1171.
Embury, I. S., 2004. Alternative litter materials for poultry. In: Georgia, U. o. (Ed.), Agfacts. Division of Agricultural Services, U.S.A.
Englander, L., Browning, M., Tooley, P. W., 2006. Growth and sporulation of Phytophthora ramorum in vitro in response to temperature and light. Mycologia 98, 365-373.
Erwin, D. C., Ribeiro, O. K., 1966. Phytophthora diseases worldwide. American Phytopathological Society Press, St. Paul, MN.
Esteves, B., Marques, A. V., Domingos, I., Pereira, H., 2008. Heat-induced colour changes of pine {Pinus pinaster) and eucalypt {Eucalyptus globulus) wood. Wood Sci Technol 42, 369- 384.
Esteves, B., Videira, R., Pereira, H., 2011. Chemistry and ecotoxicity of heat-treated pine wood extractives. Wood Sci Technol 45, 661-676.
F AO, 2002. Guidelines for regulating wood packaging materials in international trade. Food and Agriculture Organization, p. 11.
FC, 2012. Phytophthora ramorum in larch trees - update. National update. Forestry Commission, http://www.forestrv.gov.uk/forestrv/INFD-8EJKP4.
Fengel, D., Wegener, G., 1989. Wood: Chemistry, Ultrastructure, Reactions. Walter de Gruyter, New york.
116

Ferrante, V., Lolli, S., Marelli, S., Vezzoli, G., Sirri, F,, Cavalchini, L. G., 2006. Effect of light programmes, bird densities and litter types on broilers welfare. . Proc. XII European Poultry Conference. World’s Poult. Sci. Assoc., Beekbergen, the Netherlands, Verona, Italy.
Fichtner, E. J., Lynch, S. C., Rizzo, D. M., 2007a. Detection, distribution, sporulation, and survival of Phytophthora ramorum in a california redwood-tanoak forest soil. Phytopathology 97,1366-1375.
Fichtner, E. J., Lynch, S. C., Rizzo, D. M., 2009. Survival, Dispersal, and Potential Soil- Mediated Suppression of Phytophthora ramorum in a California Redwood-Tanoak Forest. Phytopathology 99, 608-619.
Fichtner, E. J., Rizzo, D. M., Lynch, S. C., Davidson, J. M., Buckles, G., Parke, J. L., 2007b. Summer survival of Phytophthora ramorum in California forests. Phytopathology 97, S36- S36.
Fitzgerald, D. J., Stratford, M., Gasson, M. J., Ueckert, J., Bos, A., Narbad, A., 2004. Mode of antimicrobial action of vanillin against Escherichia coli, Lactobacillus plantarum and Listeria innocua. J Appl Microbiol 97,104-113.
Fletcher, J., Thomback, J., Blofeld, S., 2010. Joinery: a resource efficiency action plan. Strategy for Sustainable Construction: A contribution to delivering the targets in the joint government and industry. British Woodworking Federation,www.bis.gov.uk/policies/business-sectors/construction/sustainable-construction. p. 57.
Foley, S. L., Nayak, R., Hanning, I. B., Johnson, T. J., Han, J., Ricke, S. C., 2011. Population dynamics of Salmonella enterica serotypes in commercial egg and poultry production. Appl Environ Microb 77,4273-4279.
Forest-Research, 1997. Quantification of fungal spores using a haemocytometer. Standard Operating Procedure (SOP0014v2). Forest Research, England.
Foster, J. W., 2001. Acid stress responses of Salmonella and E. coli: Survival mechanisms, regulation, and implications for pathogenesis. J Microbiol 39, 89-94.
Foster, J. W., Hall, H. K., 1990. Adaptive acidification tolerance response of Salmonella Typhimurium. J Bacteriol 172, 771-778.
Franich, R. A., Gadgil, P. D., Shain, L., 1983. Fungistatic effects of Pinus radiata needlQ epicuticular fatty and resin acids on Dothistroma pini. Physiol Plant Pathol 23,183-195.
Frankel, S. J., 2008. Sudden oak death and Phytophthora ramorum in the USA: a management challenge. Australas Plant Path 37,19-25.
Fries, R., Akcan, M., Bandick, N., Kobe, A., 2005. Microflora of two different types of poultry litter. Brit Poultry Sci 46, 668-672.
FSA, 2011. An FSA programme for the reduction of foodbome disease in the UK. Foodbome disease strategy 2010-15, www.food.gov.uk/multimedia/pdfs/fds2015.pdf. p. 24.
FSIS, 2010. Compliance guideline for controlling Salmonella and Campylobacter in poultry. Final bulletin for agency good guidance practices, www.fsis.usda.gov/Significant Guidance/index.asp. p. 52.
Gaggia, F., Mattarelli, P., Biavati, B., 2010. Probiotics and prebiotics in animal feeding for safe food production. Int J Food Microbiol 141, S15-S28.
Garbelotto, M., 2004. Sudden oak death: a tale of two continents. Outlooks on Pest Management 15, 85-89.
117

Garbelotto, M., Davidson, J. M., Ivors, K., Maloney, P. E., Hüberli, D., Rizzo, D. M., 2003. Non-oak native plants are the main hosts for the sudden oak death pathogen in California. California Agriculture 52,18-23.
Garbelotto, M., Rizzo, D., 2005. A California-based chronological review (1995-2004) of research on Phytophthora ramorum, the causal agent of sudden oak death. Phytopathologia Mediterranea 44, 127-143.
Garbelotto, M., Rizzo, D. M., 2001. Preliminary studies on chemical and cultural control of Phytophthora associated with sudden oak death Phytopathology 91, 1057-1065.
Garbelotto, M., Schmidt, D. J., Hamik, T. Y., 2007. Phosphite injections and bark application of phosphite + Pentrabark' '̂' ̂ control sudden oak death in coast live oak. Arboriculture and Urban Forestry 33, 309-317.
Garrido, M. N., Skjervheim, M., Oppegaard, H., Sorum, H., 2004. Acidified litter benefits the intestinal flora balance of broiler chickens. Appl Environ Microb 70, 5208-5213.
Gast, R. K., 2007. Serotype-specific and serotype-independent strategies for preharvest control of food-borne Salmonella in poultry. Avian Dis 51, 817-828.
Goheen, E. M., Hansen, E. M., Kanaskie, A., McWilliams, M. G., 2002. Sudden oak death caused by Phytophthora ramorum in Oregon. Plant Dis 86, 441.
Goheen, E. M., Reeser, P., Sutton, W., 2012. Reducing the spread of Phytophthora ramorum on the Redwood Nature Trail, Curry County Oregon: A case study. Phytophthora in Forests and Natural Ecosystems. 6th lUFRO Working Party 7.02.09, Cordoba-Spain.
Gonzalez, M. A., Pe'rez-Guaita, D., Correa-Royero, J., Zapata, B., Agudelo, L., Mesa- Arango, A., Betancur-Galvis, L., 2010. Synthesis and biological evaluation of dehydroabietic acid derivatives. Eur J Med Chem 45 811-816.
Gonzalez-Pena, M. M., Curling, S. F., Hale, M. D. C., 2009. On the effect of heat on the chemical composition and dimensions of thermally-modified wood. Polym Degrad Stabil 94, 2184-2193.
Goren, E., Dejong, W. A., Doornenbal, P., Bolder, N. M., Mulder, R. W. A. W., Jansen, A., 1988. Reduction of Salmonella infection of broilers by spray application of intestinal microflora : A longitudinal study. Vet Quart 10, 249-255.
Grabber, J. H., Ralph, J., Hatfield, R. D., 1998. Severe inhibition of maize wall degradation by synthetic lignins formed with coniferaldehyde. J Sci Food Agr 78, 81-87.
GraphPad-Software, 1998. InStat guide to choosing and interpreting statistical tests. GraphPad Software, Inc., San Diego, California, USA. www.graphpad.com. p. 126.
Grunwald, N. J., Garbelotto, M., Goss, E. M., Heungens, K., Prospero, S., 2012. Emergence of the sudden oak death pathogen Phytophthora ramorum. Trends Microbiol 20,131-138.
Gudbjomsdottir, B., Arason, S., Beyer, G., Weider, I., Lorentzen, G., Pia, A., Koch, A. P., 2000. Wood in the Food Industry. Nordic Wood 2. Nordisk Industrifond.
Hansen, E. M., Kanaskie, A., Prospero, S., McWilliams, M., Goheen, E. M., Osterbauer, N., Reeser, P., Sutton, W., 2008. Epidemiology of Phytophthora ramorum in Oregon tanoak forests. Can J Forest Res 38, 1133-1143.
Hardy, G. E., Barrett, S., Shearer, B. L., 2001. The future of phosphite as a fungicide to control the soilborne plant pathogen Phytophthora cinnamomi in natural ecosystems. Australas Plant Path 39, 133-139.
118

Garbelotto, M., Davidson, J. M,, Ivors, K., Maloney, P. E., Hüberli, D., Rizzo, D. M., 2003. Non-oak native plants are the main hosts for the sudden oak death pathogen in California, California Agriculture 52,18-23.
Garbelotto, M., Rizzo, D., 2005. A California-based chronological review (1995-2004) of research on Phytophthora ramorum, the causal agent of sudden oak death. Phytopathologia Mediterranea 44,127-143.
Garbelotto, M., Rizzo, D. M., 2001. Preliminary studies on chemical and cultural control of Phytophthora associated with sudden oak death Phytopathology 91, 1057-1065.
Garbelotto, M., Schmidt, D. J., Hamik, T. Y., 2007. Phosphite injections and bark application of phosphite + Pentrabark™ control sudden oak death in coast live oak. Arboriculture and Urban Forestry 33, 309-317.
Garrido, M. N., Skjervheim, M., Oppegaard, H., Somm, H., 2004. Acidified litter benefits the intestinal flora balance of broiler chickens. Appl Environ Microb 70, 5208-5213.
Gast, R. K., 2007. Serotype-specific and serotype-independent strategies for preharvest control of food-bome Salmonella in poultry. Avian Dis 51, 817-828.
Goheen, E. M., Hansen, E. M., Kanaskie, A., McWilliams, M. G., 2002. Sudden oak death caused by Phytophthora ramorum in Oregon. Plant Dis 8 6 , 441.
Goheen, E. M., Reeser, P., Sutton, W., 2012. Reducing the spread of Phytophthora ramorum on the Redwood Nature Trail, Curry County Oregon: A case study. Phytophthora in Forests and Natural Ecosystems. 6 th lUFRO Working Party 7.02.09, Cordoba-Spain.
Gonzalez, M. A., Pe'rez-Guaita, D., Correa-Royero, J., Zapata, B., Agudelo, L., Mesa- Arango, A., Betancur-Galvis, L., 2010. Synthesis and biological evaluation of dehydroabietic acid derivatives. Eur J Med Chem 45 811-816.
Gonzalez-Pena, M. M., Curling, S. F., Hale, M. D. C., 2009. On the effect of heat on the chemical composition and dimensions of thermally-modified wood. Polym Degrad Stabil 94, 2184-2193.
Goren, E., Dejong, W. A., Doomenbal, P., Bolder, N. M., Mulder, R. W. A. W., Jansen, A., 1988. Reduction of Salmonella infection of broilers by spray application of intestinal microflora : A longitudinal study. Vet Quart 10, 249-255.
Grabber, J. H., Ralph, J., Hatfield, R. D., 1998. Severe inhibition of maize wall degradation by synthetic lignins formed with coniferaldehyde. J Sci Food Agr 78, 81-87.
GraphPad-Software, 1998. InStat guide to choosing and interpreting statistical tests. GraphPad Software, Inc., San Diego, California, USA. www.granhnad.com. p. 126.
Gmnwald, N. J., Garbelotto, M., Goss, E. M., Heungens, K., Prospero, S., 2012. Emergence of the sudden oak death pathogen Phytophthora ramorum. Trends Microbiol 20,131-138.
Gudbjomsdottir, B., Arason, S., Beyer, G., Weider, I., Lorentzen, G., Pia, A., Koch, A. P., 2000. Wood in the Food Industry. Nordic Wood 2. Nordisk Industrifond.
Hansen, E. M., Kanaskie, A., Prospero, S., McWilliams, M., Goheen, E. M., Osterbauer, N., Reeser, P., Sutton, W., 2008. Epidemiology of Phytophthora ramorum in Oregon tanoak forests. Can J Forest Res 38,1133-1143.
Hardy, G. E., Barrett, S., Shearer, B. L., 2001. The future of phosphite as a fungicide to control the soilbome plant pathogen Phytophthora cinnamomi in natural ecosystems. Australas Plant Path 39,133-139.
118

Harju, A. M., Kainulainen, P., Venalainen, M., Tiitta, M., Viitanen, H., 2005. Differences in resin acid concentration between brown-rot resistant and susceptible Scots pine heartwood. Holzforschung 56, 479-486.
Hendriksen, R. S., Vieira, A. R., Karlsmose, S., Wong, D. M. A. L. P., Jensen, A. B., Wegener, H. C., Aarestrup, F. M., 2011. Global monitoring of Salmonella serovar distribution from the World Health Organization Global Foodbome Infections Network Country Data Bank: Results of Quality Assured Laboratories from 2001 to 2007. Foodbome Pathog Dis 8 , 887-900.
Henriks, M. L., Ekman, R., von Weissenberg, K., Aboensis, 1979. Bioassay does not seem to be strictly related to the carbonyl of some resin and fatty acids with Fomes annosus. Acta Academiae Aboensis 39B, 1-7.
Hess, J., Macklin, K., Bilgili, S., Blake, J., Norton, R., 2010. In-house litter composition between flocks. Poultry Science Department, Aubum University, Aubum, AL.
Heungens, K., De Debbelaere, I., Maes, M., 2006. Fungicide control of Phytophthora ramorum on Rhododendron. In: Frankel, S. J. S., P.J.; Haverty, M.I. (Ed.), The sudden oak death second science symposium: the state of our knowledge. Department of Agriculture, Forest Service, Pacific Southwest Research Station, Albany, CA: U.S., pp. 241-257.
Holt, P. S., 1995. Horizontal transmission of Salmonella Enteritidis in molted and nnmolted laying chickens. Avian Dis 39,239-249.
Homans, A. L., Fuchs, A., 1970. Direct bioautography on thin layer chromatography as a method for detecting fimgitoxic substances. Joumal of Chromatography 51, 327-329.
Inman, A. J., Hughes, K. J. D., Beales, P. A., Lane, C. R., Rizvi, R., Bames, A. V., Barton, V. C., R.J., B., Griffin, R , 2005. Non-tree host range and diagnosis of Phytophthora ramorum (sudden oak death) in support of pest risk assessment, management and policy. Project Report Department of Environment, Food and Rural Affairs, London
Ivanov, I. E., 2001. Treatment of broiler litter with organic acids. Res Vet Sci 70,169-173.
Ivors, K. L., Hayden, K. J., Bonants, P. J. M., Rizzo, D. M., Garbelotto, M., 2004. AFLP and phylogenetic analyses of North American and European populations of Phytophthora ramorum. Mycol Res 108, 378-392.
Jeffers, S., 2005. Recovery of Phytophthora ramorum from soilless mixes around container- grown omamental plants. Phytopathology 95, S48.
Jones, J. M., 2006. APHIS Phytophthora ramorum regulatory strategy for nurseries. In: Frankel, S. J. S., P.J.; Haverty, M.I. (Ed.), Sudden oak death second science symposium. Department of Agriculture, Forest Service, Albany, CA: U.S., pp. 45-46.
Judelson, H. S., Blanco, F. A., 2005. The spores of Phytophthora: Weapons of the .plant destroyer. Nat Rev Microbiol 3,47-58.
Kamdem, D. P., Pizzi, A., Jermannaud, A., 2002. Durability of heat-treated wood. Holz Roh Werkst 60,1-6.
Kanaskie, A. H., E.; Goheen, E. Michaels.; Osterbauer, N.; McWilliams, M.; Laine, J.; Thompson, M.; Savona, S.; Timeus, H.; Woosley, B.; Sutton, W.; Reeser, P.; Schultz, R.; Hilbum, D., 2010. Detection and eradication of Phytophthora ramorum from Oregon forests, 2001-2008. In: Frankel, S. J. K., J.T.; Palmieri, K.M., (Ed.), Proceedings of the sudden oak death fourth science symposium. Gen. Tech. Rep. PSW-GTR-229. United States Department of Agriculture, Forest Service, Albany, CA: U.S., pp. 259-261.
119

Kapil, A., 2005. The challenge of antibiotic resistance: Need to contemplate. Indian J Med Res 121, 83-91.
Kelley, T. R., Pancorbo, O. C., Merka, W. C., Barnhart, H. M., 1998. Antibiotic resistance of bacterial litter isolates. Poultry Sci 77, 243-247.
Kelly, N. M., Meentemeyer, R. K., 2002. Landscape dynamics of the spread of sudden oak death. Photogrammetric Engineering and Remote Sensing 6 8 , 1001-1009.
Kelsey, R. G., Harmon, M. E., 1989. Distribution and variation of extractable total phenols and tannins in the logs of four conifers after 1 year on the ground. Canadian Joumal of Forest Research, 19, 1030-1036.
Kim, W. K., Patterson, P. H., 2003. Effect of minerals on activity of microbial uricase to reduce ammonia volatilization in poultry manure. Poultry Sci 82, 223-231.
Kliejunas, J. T., 2010. Sudden Oak Death and Phytophthora ramorum: A summary of the Literature. Gen. Tech. Rep. PSW-GTR-234, vol. 1. United States Department of Agriculture, Forest service, Portland, OR: U.S., pp. 11-12.
Kotula, K. L., Pandya, Y., 1995. Bacterial contamination of broiler chickens before scalding. J Food Protect 58,1326-1329.
Kumar, R., Sharma, P. K., Mishra, P. S., 2012. A review on the vanillin derivatives showing various biological activities. PharmTech Research 4, 266-279.
Lane, C. R , Beales, P. A., Hughes, K. J. D., Griffin, R. L., Munro, D., Brasier, C. M., Webber, J. F., 2003. First outbreak of Phytophthora ramorum in England, on Viburnum tinus. Plant Pathol 52, 414-414.
Levy, S. B., 2001. Antibacterial household products: Cause for concern. Emerg Infect Dis 7, 512-515.
Lewinsohn, E., Gijzen, M., Savage, T. J., Croteau, R., 1991. Defense-Mechanisms of Conifers - Relationship of Monoterpene Cyclase Activity to Anatomical Specialization and Oleoresin Monoterpene Content. Plant Physiol 96, 38-43.
Li, J. R., Sanchez, S., Hofacre, C., Maurer, J. J., Harmon, B. G., Lee, M. D., 2003. Evaluation of broiler litter with reference to the microbial composition as assessed by using 16S rRNA and functional gene markers. Appl Environ Microb 69, 901-908.
Lien, R. J., Hess, J. B., Conner, D. E., Wood, C, W., Shelby, R. A., 1998. Peanut hulls as a litter source for broiler breeder replacement pullets. Poultry Sci 77,41-46.
Linderman, R. G., Davis, E. A., 2006. Survival of Phytophthora ramorum compared to other species of Phytophthora in potting media components, compost, and soil. Horttechnology 16, 502-507.
Linderman, R. G., Davis, E. A., Marlow, J. L., 2006. Response of selected nursery crop plants to inoculation with isolates of Phytophthora ramorum and other Phytophthora species. Horttechnology 16, 216-224.
Line, J. E., 2002. Campylobacter and Salmonella populations associated with chickens raised on acidified litter. Poultry Sci 81,1473-1477.
Line, J. E., Bailey, J. S., 2006. Effect of on-farm litter acidification treatments on Campylobacter and Salmonella populations in commercial broiler houses in northeast Georgia. Poultry Sci 85,1529-1534.
120

Liu, Z. Y., Wang, L., Beasley, D., Oviedo, E., 2007. Effect of moisture content on ammonia emissions from broiler litter: A laboratory study. Joumal of atmospheric chemistry 58, 41-53.
Long, J. R., Dewitt, W. F., Ruet, J. L., 1980. Studies on Salmonella from Floor Litter of 60 Broiler Chicken Houses in Nova-Scotia. Can Vet J 21, 91-94.
Lovanh, N., Cook, K. L., Rothrock, M. J., Miles, D. M., Sistani, K., 2007. Spatial shifts in microbial population structure within poultry litter associated with physicochemical properties. Poultry Sci 86,1840-1849.
MacDonald, A. G., Dare, P. H., Gifford, J. S., Steward, D., Riley, S., 2002. Assessment of air emissions from industrial kiln drying of Pinus radiata Wood. Holz Roh Werkst 60, 181-190.
MacKay, D. G., Cole, B. J. W., Fort, R. C., Mares, A., 2009a. Potential markets for chemicals and pharamceuticals from woody biomass in Maine. Forest research LLC, Maine, p. 26.
Mackay, D. G., Cole, B. J. W., Fort, R. C., Mares, A., 2009b. Potential markets for chemicals and pharmaceuticals from woody biomass in maine. In: Marine, U. o. (Ed.).
Macklin, K. S., Hess, J. B., Bilgili, S. F., Norton, R. A., 2005. Bacterial levels of pineshavings and sand used as poultry litter. J Appl Poultiy Res 14, 238-245.
Majowicz, S. E., Musto, J., Scallan, E., Angulo, F. J., Kirk, M., O'Brien, S. J., Jones, T. F., Fazil, A., Hoekstra, R. M., Burd, I. C. E. D., 2010. The Global Burden of Nontyphoidal Salmonella Gastroenteritis. Clin Infect Dis 50, 882-889.
Malone, G. W., Chaloupka, G. W., Saylor, W. W., 1983. Influence of Litter Type and Size onBroiler Performance .1. Factors Affecting Litter Consumption. Poultry Sci 62, 1741-1746.
Manninen, A. M., Pasanen, P., Holopainen, J. K., 2002. Comparing the VOC emissions between air-dried and heat-treated Scots pine wood. Atmos Environ 36, 1763-1768.
Manter, D. K., Karchesy, J. J., Kelsey, R. G., 2006. The sporicidal activity of yellow-cedar heartwood, essential oil and wood constituents towards Phytophthora ramorum in culture. Forest Pathol 36, 297-308.
Manter, D. K., Kelsey, R. G., Karchesy, J. J., 2007. Antimicrobial activity of extractable conifer heartwood compounds toward Phytophthora ramorum. J Chem Ecol 33, 2133-2147.
Marsh, E. N. G., 1999a. Coenzyme B-12 (cobalamin)-dependent enzymes. Essays Biochem 34,139-154.
Marsh, T. E., 1999b. The role of litter management in pre-harvest pathogen reduction programmes. Poultry International 38, 42.
Martin, H., Maris, P., 2005. An assessment of the bactericidal and fungicidal efficacy of seventeen mineral and organic acids on bacterial and fungal food industry contaminants. Sci Aliment 25,105-127.
Martin, S. J., Micek, S. T., Wood, G. C., 2010. Antimicrobial resistance: Consideration as an adverse drug event. Crit Care Med 38, S155-S161.
Martland, M. F., 1985. Ulcerative dermatitis in broiler-chickens : the effects of wet Litter. Avian Pathol 14, 353-364.
McLaughlin, I., Sutton, W., Hansen, E., 2006. Survival of Phytophthora ramorum in tanoak and rhododendron leaves. In: Frankel, S. J. S., P.J.; Haverty, M.I. (Ed.), The sudden oak death second science symposium: the state of our knowledge. Department of Agriculture, Forest Service, Pacific Southwest Research Station, Albany, CA: U.S., p. 529.
121

Mead, G, G,, Barrow, P. A., 1990. Salmonella control in poultry by 'Competitive Exclusion' or Immunization. Lett Appl Microbiol 10, 221-227.
Micales, J. A., Han, J. S., Davis, J. L., Young, R. A., 1994. Chemical-Composition and Fungitoxic Activities of Pine-Cone Extractives. Biodeterioration Research 4, 317-332.
Miles, D. M., Branton, S. L., Lott, B. D., 2004. Atmospheric ammonia is detrimental to the performance of modem commercial broilers. Poultry Sci 83,1650-1654.
Miles, D. M., Rowe, D. E., Cathcart, T. C., 2011a. High litter moisture content suppresses litter ammonia volatilization. Poultry Sci 90,1397-1405.
Miles, D. M., Rowe, D. E., Cathcart, T. C., 2011b. Litter ammonia generation: Moisture content and organic versus inorganic bedding materials. Poultry Sci 90, 1162-1169.
Milling, A., Kehr, R., Wulf, A., Smalla, K., 2005. Survival of bacteria on wood and plastic particles: Dependence on wood species and environmental conditions. Holzforschung 59, 72- 81.
Moore, P. A., 2003. Reducing ammonia emissions and phosphorus runoff from animal manure with aluminum compounds. Proce. Taegu University Life Science Institute, pp. 15- 27.
Moore, P. A., Daniel, T. C., Edwards, D. R., Miller, D. M., 1995. Effect of chemical amendments on ammonia volatilization from poultry litter. J Environ Qual 24, 293-300.
Moore, P. A., Daniel, T. C., Edwards, D. R., Miller, D. M., 1996. Evaluation of chemical amendments to reduce ammonia volatilization from poultry litter. Poultry Sci 75, 315-320.
Mourtzinos, I., Konteles, S., Kalogeropoulos, N., Karathanos, V. T., 2009. Thermal oxidation of vanillin affects its antioxidant and antimicrobial properties. Food Chem 114, 791-797.
Murzin, D. Y., Maki-Arvela, P., Salmi, T., Holmbom, B., 2007. Catalytic trans-formations for production of fine chemicals and pharmaceuticals from wood-derived raw materials. Chem Eng Technol 30, 569-576.
Nagle, A. M., Mcpherson, B. A., Wood, D. L., Garbelotto, M., Bonello, P., 2011. Relationship between field resistance to Phytophthora ramorum and constitutive phenolic chemistry of coast live oak. Forest Pathol 41, 464-469.
Nahm, K. H., 2005. Environmental effects of chemical additives used in poultry litter and swine manure. Crit Rev Env Sci Tec 35,487-513.
Napierala Filipiak, A., Werner, A., Mardarowicz, M., Gawdzik, J., 2002. Concentrations of terpenes in mycorrhizal roots of Scots pine {Pinus sylvestris) seedlings grown in vitro. J Plant Physiol 24, 137-143.
NFU-online, 2013. NFU lobbying success for livestock sector. Poultry Highlights. National Farmers' Union of England and Wales, http://www.nfuonline.com/news/press-centre/nfu- lobbving-success-for-livestock-sector/.
Niemz, P., Hofmann, T., Retfalvi, T., 2010. Investigation of chemical changes in the structure of thermally modified wood. Maderas-Cienc Tecnol 12, 69-78.
Ockels, F. S., Eyles, A., McPherson, B. A., Wood, D. L., Bonello, P., 2007. Phenolic chemistry of Coast Live oak response to Phytophthora ramorum infection. Chemical Ecology 33,1721-1732.
122

Oliveira, S. C., Cavlheiro, A, C. L., Trindade, D. S., Lopez, L, Cdrrea-Oliveira, S., 1974. Comparison between types of litter in broiler production in Rio Grande do Sul Brasil. 15th World’s Poultry Congress. World’s Poultry Science Association, Washington, USA, pp. 342- 344.
Orlikowski, L. B., 2004. Biological control of Phytophthora ramorum on rhododendron. Communications in Agricultural and Applied Biological Sciences 69, 687-692.
Parke, J. L., Lucas, S., 2008. Sudden oak death and ramorum blight. The Plant Health Instructor.
Payne, J. B., Kroger, E. C., Watkins, S. E., 2002. Evaluation of litter treatments on Salmonella recovery from poultry litter. J Appl Poultry Res 11, 239-243.
Payne, J. B., Osborne, J. A., Jenkins, P. K., Sheldon, B. W., 2007. Modeling the growth and death kinetics of Salmonella in poultry litter as a function of pH and water activity. Poultry Sci 86,191-201.
Payne, J. B., Watkins, S. E., 2005. Evaluation of litter treatments on Salmonella recovery in poultry litter. Spring 7,10-11.
Phillips, M. A., Croteau, R. B., 1999. Resin-based defences in conifers. Trends in Plant Science 4,184-190.
Piispanen, P. S., Hedman, B., Norin, T., 2002. Synthesis and characterization of dehydroabietic acid derivatives suitable for surfactant synthesis. J Surfactants Deterg 5, 165- 168.
Pope, M. J., Cherry, T. E., 2000. An evaluation of the presence of pathogens on broilers raised on Poultry Litter Treatment (R)-treated litter. Poultry Sci 79,1351-1355.
Poucke, K. V., Franceschini, S., Webber, J., Heungens, K., Brasier, C., 2012. Discovery of a fourth evolutionary lineage of Phytophthora ramorum: EU2. Fungal Biology 116, 1178-1191.
Reece, F. N., Bates, B. J., Lott, B. D., 1979. Ammonia Control in Broiler Houses. Poultry Sci 58, 754-755.
Ritz, C. W., Fairchild, B. D., Lacy, M. P., 2004. Implications of ammonia production and emissions from commercial poultry facilities: A review. J Appl Poultry Res 13, 684-692.
Ritz, C, W., Fairchild, B. D., Lacy, M. P., 2009. Litter quality and broiler performance. In: Scientists, E. P. (Ed.), Cooperative Extension Service/ The University of Georgia College of Agricultural and Environmental Sciences, pp. 1-7.
Rizzo, D. M., 2006. Research on the epidemiology, ecology and management of Phytophthora ramorum in California forests hi: Frankel, S. J. S., P.J.; Haverty, M.I. (Ed.), The sudden oak death second science symposium: the state of our knowledge. Department of Agriculture, Forest Service, Pacific Southwest Research Station, Albany, CA: US, pp. 21-25.
Rizzo, D. M., Garbelotto, M., Davidson, J. M., Slaughter, G. W., Koike, S. T., 2002. Phytophthora ramorum as the cause of extensive mortality of Quercus spp. and Lithocarpus densiflorus in California. Plant Dis 8 6 , 205-214.
Rizzo, D. M., Garbelotto, M., Hansen, E. A., 2005. Phytophthora ramorum: Integrative research and management of an emerging pathogen in California and Oregon forests. Annu Rev Phytopathol 43, 309-335.
Russell, A. D., 2003. Biocide use and antibiotic resistance: the relevance of laboratory findings to clinical and environmental situations. Lancet Infect Dis 3, 794-803.
123

Russell, S. M., 2012. Salmonella intervention during grow out. CRC Press, U.S.A. http://www.crcpress.com.
Sansford, C. E., Inman, A. J., Baker, R , Brasier, C., Frankel, S., de Gruyter, J., Husson, C., Kehlenbeck, H., Kessel, G., Moralejo, E., Steeghs, M., Webber, J., Werres, S., 2009. Report on the risk of entry, establishment, spread and socio-economic loss and environmental impact and the appropriate level of management for Phytophthora ramorum for the EU. Deliverable Report Forest Research, Central Science Laboratory, Sand Hutton, York, UK.
Sansford, C. E., Woodhall, J. W., 2007. Datasheet for Phytophthora ramorum. Central Science Laboratory, Department of Environment, Forestry, and Rural Affairs, Sand Hutton, York p. 43.
Sansford, C. E. W., J.W. , 2007. Datasheet for Phytophthora ramorum. Central Science Laboratory, Department of Environment, Forestry, and Rural Affairs., Sand Hutton, York.
Santos, F. B. O., Sheldon, B. W., Santos, A. A., Ferket, P. R , 2008. Influence of housing system, grain type, and particle size on Salmonella colonization and shedding of broilers fed triticale or corn-soybean meal diets. Poultry Sci 87,405-420.
Savluchinske-Feio, S., Marcelo Curto, M. J., Gigante, B., Roseiro, J. C., 2006. Antimicrobial activity of resin acid derivatives. Appl Microbiol Biot 72, 430-436.
Savluchinske-Feio, S., Nunes, L., Pereira, P. T., Silva, A. M., Roseiro, J. C., Gigante, B., Curto, M. J. M., 2007. Activity of dehydroabietic acid derivatives against wood contaminant fungi. J Microbiol Meth 70, 465-470.
Savluchinske-Feio, S. S., Gigante, B., Roseiro, J. C., Marcelo-Curto, M. J., 1999. Antimicrobial activity of diterpene resin acid derivatives. J Microbiol Meth 35, 201-206.
Scheffer, E., 1965. Microbiology of Built up Poultry Litter. J Appl Bacteriol 28, 403.
Schneitz, C., 2005. Competitive exclusion in poultry - 30 years of research. Food Control 16, 657-667.
Schdnwalder, A., Kehr, R., Wulf, A., Smalla, K., 2000. Remarkable antibacterial properties of wood. Holz Zentral-blatt 147, 2037-2038.
Schdnwalder, A., Kehr, R., Wulf, A., Smalla, K., 2002. Wooden boards affecting the survival of bacteria? Holz Roh-Werkst 60, 249-257.
Seydim, A. C., Sarikus, G., 2006. Antimicrobial activity of whey protein based edible films incorporated with oregano, rosemary and garlic essential oils. Food Research International 39, 639-644.
Shea, K. M., 2003. Antibiotic resistance: What is the impact of agricultural uses of antibiotics on children's health? Pediatrics 112, 253-258.
Shelly, J., Singh, R., Langford, C., Mason, T., 2006. Evaluating the survival of Phytophthora ramorum in firewood. In: Frankel, S. J. S., P.J.; Haverty, M.I. (Ed.), sudden oak death second science symposium: the state of our knowledge. Department of Agriculture, Forest Service, Pacific Southwest Research Station, Albany, CA: U.S, p. 540.
Shishkoff, N., 2005. The effect of systemic fungicides on detection by culturing of Phytophthora ramorum. Phytopathology 95, S96-S96.
Shishkoff, N., 2007. Persistence of Phytophthora ramorum in soil mix and roots of nursery ornamentals. Plant Dis 91,1245-1249.
124

Shishkoff, N., Tooley, P., 2004. Persistence of Phytophthora ramorum in nursery plants and soil. Phytopathology 94, S95-S95.
Sjôstrôm, E., 1981. Wood chemistry: Fundamentals and Applications. Academic Press, London.
Spessard, G. O., Matthews, D. R., Nelson, M. D., Rajtora, T. C., Possum, M. J., Giannini, J. L., 1995. Phytoalexin-like activity of abietic acid and Its derivatives. J Agr Food Chem 43, 1690-1694.
Stamm, A. J., Burr, H. K., Kline, A. A., 1946. Staybwood...Heat-Stabilized Wood. Ind Eng Chem 38, 630-634.
Sundqvist, B., 2002. Wood color control during kiln-drying. Forest Prod J 52, 30-37.
Sundqvist, B., 2004. Colour changes and Acid formation in wood during heating. Division of Wood Material Science. Lulea University of Technology, Sweden, p. 154.
Swain, S., Hamik, T., Mejia-Chang, M., Creque, J., Garbelotto, M., 2002. Survivability of Phytophthora ramorum in the composting process. Sudden oak death science symposium, Berkeley, CA: University of California.
Swain, S., Harnik, T., Mejia-Chang, M., Hayden, K., Bakx, W., Creque, J., Garbelotto, M., 2006. Composting is an effective treatment option for sanitization of Phytophthora ramorum- infected plant material. J Appl Microbiol 101, 815-827.
Swiecki, T. J., Bernhardt, E. A., 2001. Evaluation of stem water potential and other tree and stand variables as risk factors for Phytophthora ramorum canker development in coast live oak and tanoak. Phytosphere Research. Department of Agriculture, Berkeley, CA., Vacaville, CA: U.S.
Taitt, C. R., Shubin, Y. S., Angel, R., Ligler, F. S., 2004. Detection of Salmonella enterica serovar typhimurium by using a rapid, array-based immunosensor. Appl Environ Microb 70, 152-158.
Tasistro, A. S., Cabrera, M. L., Zhao, Y. B., Kissel, D. E., Xia, K., Franklin, D. H., 2007. Soluble phosphorus released by poultry wastes in acidified aqueous extracts. Commun Soil Sci Plan 38, 1395-1410.
Tauxe, R., 1996. An update on Salmonella. Health and Environmental Digestion 10, 1-4.
Terzich, M., Pope, M. J., Cherry, T. E., Hollinger, J., 2000. Survey of pathogens in poultry litter in the United States. J Appl Poultry Res 9, 287-291.
Terzich, M., Quarles, C., Goodwin, M. A., Brown, J., 1998. Effect of poultry litter treatment (R) (PET (R)) on the development of respiratory tract lesions in broilers. Avian Pathol 27, 566-569.
The-Environment-Agency, 2006. Integrated Pollution Prevention and Control (IPPC): Intensive Farming. In: Unit, D. a. I. E. (Ed.), vol. 1. Environment Agency, Bristol, p. 106.
The-Environment-Agency, 2013. Best Farming Practices, vol. 2013. The Environment Agency, http://www.environment-agencv.gov.uk/business/sectors/bestfarmingpractices.aspx.
Tjeerdsma, B. F., Militz, H., 2005. Chemical changes in hydrothermal treated wood: FTIR analysis of combined hydrothermal and dry heat-treated wood. Holz Roh Werkst 63, 102-111.
Tjosvold, S. A., Chambers, D. L., Davidson, J. M., Rizzo, D. M., 2002a. Incidence of Phytophthora ramorum inoculum found in soil collected from a hiking trail and hiker’s shoes
125

in a California park. The sudden oak death science symposium: the state of our knowledge. Department of Agriculture, Forest Service and University of California, Berkeley, Berkeley, CA: U.S.
Tjosvold, S. A., Chambers, D. L., Davidson, J. M., Rizzo, D. M., 2002b. Incidence of Phytophthora ramorum inoculum found in soil collected from a hiking trail and hiker’s shoes in a California park. The sudden oak death science symposium: the state of our knowledge. Department of Agriculture, Forest Service and University of California, Berkeley, Berkeley, CA: U.S.
Tooley, P. W., Browning, M., Berner, D., 2008. Recovery of Phytophthora ramorum following exposure to temperature extremes. Plant Dis 92,431-437.
Torok, V. A., Hughes, R. J., Ophel-Keller, K., Ali, M., MacAlpine, R., 2009. Influence of different litter materials on cecal microbiota colonization in broiler chickens. Poultry Sci 8 8 , 2474-2481.
Tucker, J. F., 1967. Survival of Salmonellae in built-up litter for housing of rearing and laying fowls. Brit Vet J 123, 92-&.
Turner, J., 2007. Phytophthora ramorum and Phytophthora kernoviae: development of post eradication strategies for management/treatment of contaminated substrates and inoculum at outbreak sites. Annual Project Report. Central Science Laboratory, Forest Research, Sand Hutton, York, UK.
Turner, J., Jennings, P., 2005. Epidemiology of Phytophthora ramorum in relation to risk and policy.. Project Work Package 3 Report. Forest Research, Central Science Laboratory, Sand Hutton, York, UK.
Turner, J., Jennings, P., McDonough, S., Liddell, D., Stonehouse, J., 2006. Chemical control of Phytophthora ramorum causing foliar disease in hardy nursery stock in the United Kingdom. In: Frankel, S. J. S., P.J.; Haverty, M.I. (Ed.), The sudden oak death second science symposium: the state of our knowledge. Department of Agriculture, Forest Service, Pacific Southwest Research Station, Albany, CA: U.S., pp. 273-274.
Turner, J., Jennings, P., Werres, S., Wagner, S., 2008. Report on response of isolates of P. ramorum exposed to fungicides commonly used to control Phytophthora diseases in nurseries. Deliverable Report Forest Research, Central Sciences Laboratory, Sand Hutton, York, UK.
Urzua, A., Rezende, M. C., Mascayano, C., Vasquez, L., 2008. A structure-activity study of antibacterial diterpenoids. Molecules 13, 882-891.
USDA, 2012a. Economic Data. United States Department of Agriculture, http://www.uspoultrv.org/economic data/.
USDA, 2012b. Poultry & Eggs. United States Department of Agriculture, http://www.ers.usda.gov/topics/animal-products/poultrv-eggs/background.aspx.
Vandeplas, S., Dauphin, R. D., Beckers, Y., Thonart, P., Thewis, A., 2010. Salmonella in chicken: current and developing strategies to reduce contamination at farm level. J Food Protect 73, 774-785.
Vasantha Rupasinghe, H. P., Boulter-Bitzer, J., Ahn , T., Odumeru, J. A., 2006. Vanillin inhibits pathogenic and spoilage microorganisms in vitro and aerobic microbial growth in fresh-cut apples. Food Research International 39, 575-580.
126

Voda, K., Boh, B., Vrtacnik, M., Pohleven, F., 2003. Effect of the antifungal activity of oxygenated aromatic essential oil compounds on the white-rot Trametes versicolor and the brown-rot Coniophoraputeana. Int Biodeter Biodegr 51, 51-59.
Volkova, V. V., Bailey, R. H., Wills, R. W., 2009. Salmonella in broiler litter and properties of soil at farm location. Plos One, vol. 4, p. e6403.
Webber, J. F., Mullett, M., Brasier, C. M., 2010 Dieback and mortality of plantation Japanese larch {Larix kaempferi) associated with infection by Phytophthora ramorum. New Disease Reports 22.
Webber, J. F., Rose, J., 2008. Dissemination of aerial and root infecting Phytophthoras by human vectors.. The sudden oak death third science symposium. Department of Agriculture, Forest Service, Pacific Southwest Research Station, Albany, CA: U.S., pp. 195-198.
Werker, A. G., Hall, E. R., 1997. The influence of pH on resin acid solubility related to biodégradation kinetics of resin acid in pulp mill effluent. P Tech as P, 19-26.
Werres, S., Marwitz, R., Veld, W. A. M. I., De Cock, A. W. A. M., Bonants, P. J. M., De Weerdt, M., Themann, K., Ilieva, E., Baayen, R. P., 2001. Phytophthora ramorum sp nov., a new pathogen on Rhododendron and Viburnum. Mycol Res 105,1155-1165.
Wheeler, E., Casey, K., Gates, R., Xin, H., Liang, Y., Topper, P., 2008. Litter management strategies in relation to ammonia emissions from floor-raised birds. Proc.Mitigating Air Emissions from Animal Feeding Operations conference, Iowa State University.
White, P. L., Baker, A. R., James, W. O., 1997. Strategies to control Salmonella and Campylobacter in raw poultry products. Rev Sci Tech Oie 16, 525-541.
Widmer, T., 2008. Investigating the potential of biological control against Phytophthora ramorum. In: Frankel, S. J. K., J.T.; Palmieri, K.M. (Ed.), The sudden oak death third science symposium. Department of Agriculture, Forest Service, Pacific Southwest Research Station, Albany, CA: U.S., p. 491.
Williams, Z. T., 2012. Effects of Management on the Bacterial community present in poultry litter. Aubum University, Alabama, p. 128.
Willis, W. L., Ouart, M. D., Quarles, C. L., 1987. Effect of an Evaporative Cooling and Dust Control-System on Rearing Environment and Performance of Male Broiler-Chickens. Poultry Sci 66,1590-1593.
Wink, M., 2006. Importance of plant secondary metabolites for protection against insects and microbial infections. Adv Phytomed 3,251 -268,
Xu, X. H., Ko, W., 1998. A quantitative confined inoculation method for studies of pathogenicity of fungi on plants. Bot. Bull. Acad. Sin 39, 187-190,
Yakabe, L., E., Macdonald, J. D., 2008. Soil treatments for the elimination of Phytophthora ramorum from nursery beds: current knowledge from the laboratory and the field. In: Frankel, S. J. K., J.T.; Palmieri, K.M. (Ed.), The sudden oak death third science symposium. Department of Agriculture, Forest Service, Pacific Southwest Research Station, Albany, CA: U.S, pp. 113-114.
Ye, X., Wang, S., Ruan, R., Qi, J., Womac, A. R., Doona, C. J., 2006. Water mobility and mold susceptibility of engineered wood products. T Asabe 49, 1159-1165.
Zwenger, S., Basu, B., 2008. Plant terpenoids: applications and future potentials. Biotechnology and Molecular Biology Reviews 3, 001-007.
127

Appendices
Spectrum 1.1: Mass spectrum of (+)-dehydroabietic acid (CDCI3) ...............................1
Spectrum 1.2: NMR spectrum of (+)-dehydroabietic acid (CDCI3) ......................... 2
Spectrum 1.3: NMR spectrum of (+)-dehydroabietic acid (CDCI3) ........................ 3
Spectrum 2: NMR spectrum of picealactone A (CDCI3) ............................................4
Spectrum 3.1: Mass spectrum of vanillin (CDGI3) ............................................................ 5
Spectrum 3.2: NMR spectrum of vanillin (CDCI3) ....................................................... 6
Spectrum 3.3: NMR spectrum of vanillin (CDCI3) .......................................................7
Spectrum 4.1: Mass spectrum of vanillic acid (CDCI3) .....................................................8
Spectrum 4.2 :^H NMR spectrum of vanillic acid (CDCI3) ............................................... 9
Spectrum 4.3: NMR spectrum of vanillic acid (CDCI3) .............................................10
Spectrum 5.1: Mass spectrum of acetovanillone (CDC13)............................................. 11
Spectrum 5.2: NMR spectrum of acetovanillone (CDCI3) ........................................ 12
Spectrum 6 : NMR spectrum of guaicol (CDCI3) ...................... 13
Spectrum 7.1: Mass spectrum of stilbene (CDCI3) ......................... 14
Spectrum 7.3: NMR spectrum o f 3, 3 ’-dimethoxy-4, 4’-dihydroxy-stilbene(CD3O D ).................................................................................................................................. 15
Spectrum 7.2: NMR spectrum of 3, 3 ’-dimethoxy-4, 4’-dihydroxy-stilbene(CD3O D )........................................................................................................ 16
Spectrum 8.1: Mass spectrum of coniferaldehyde (CDCI3) ........................................... 17
Spectrum 8.2: NMR spectrum of coniferaldehyde (CDCI3) ......................... 18
Spectra 9.1: Mass spectrum of methyl abietate (DCM)................................................20
Spectra 9.2: Mass spectrum of dehydroabietic acid (DCM)........................................ 21
Spectra 9.3: Mass spectrum of a-pinene (D CM )............................... 22
Spectra 9.4: Mass spectrum of 3-carene (DCM)............................................................. 23
128
Scan 3738 (24.476 min): PINE.D\data.ms 23$.1
Abundance
285.185000
80000!
75000
70000
65000
60000
55000!
50000'
45000
40000
35000
197.130000
300.125000!
141.0
20000
15000 155.0117.091.0
10000 55.0 183.1169.0
69.15000
257.1
50 60 70 80 90 1 0 0 1 1 0 1 2 0 1 3 0 1 4 0 1 5 0 1 6 0 1 7 0 1 8 0 1 9 0 2 0 0 2 1 0 2 2 0 230240 250 260 270280 290 300311m /z->
Spectrum 1.1: Mass spectrum o f (+)-dehydroabietic acid (CDCI3)
1
Ilfij] c
u8es
a«£%
0
1 aiSZX
ClC/3
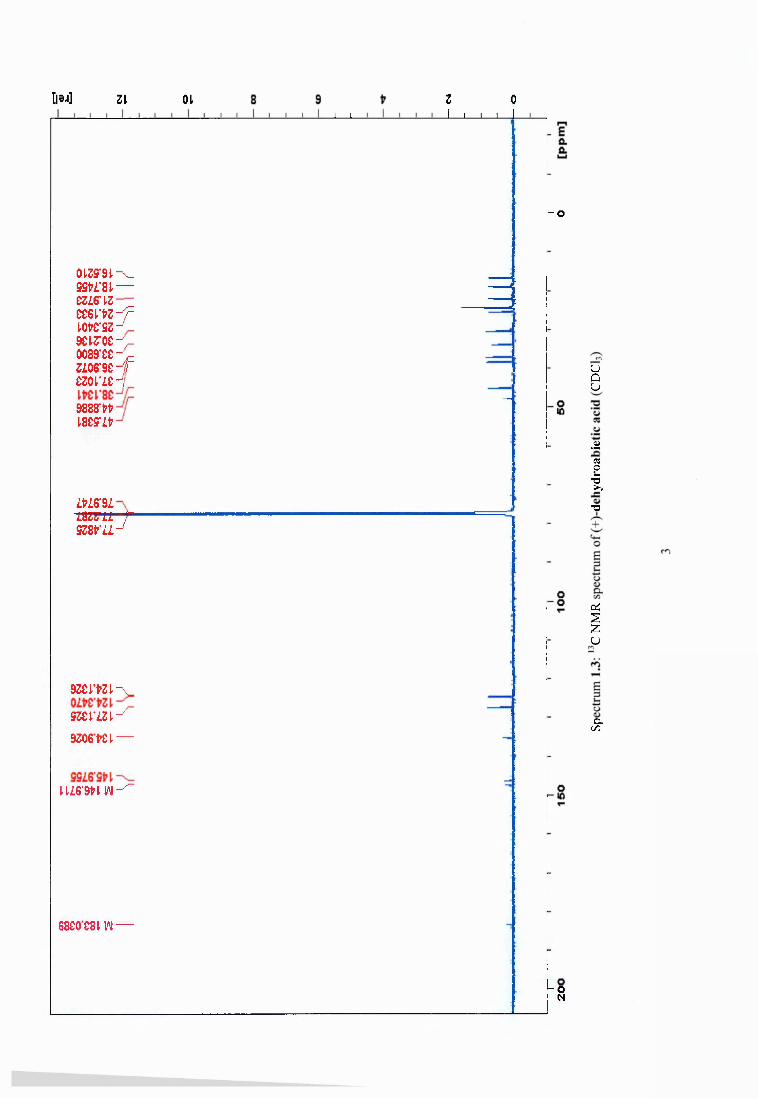
[|9J] 31 OL
OiZQ'Si-^ 99W81 — tZLÎ'iZ —t z % v n ^IOK'93-'9ei3'oe-v0089'££ 3Z06‘I 8301'.. ,
9888'f^ 188911'
)'8 8 - _Î-9 8 #L18i
it'i6-9i 183T 938M1É T
938l’W l- \
9381131^ 9306fr8l —
Ui6'9fHA|-'"
6880 881 W —
J L3 0
J I I L
m
-
- o
^8
uÛu
. 2
«2%■o
aiSzu
CLc/2
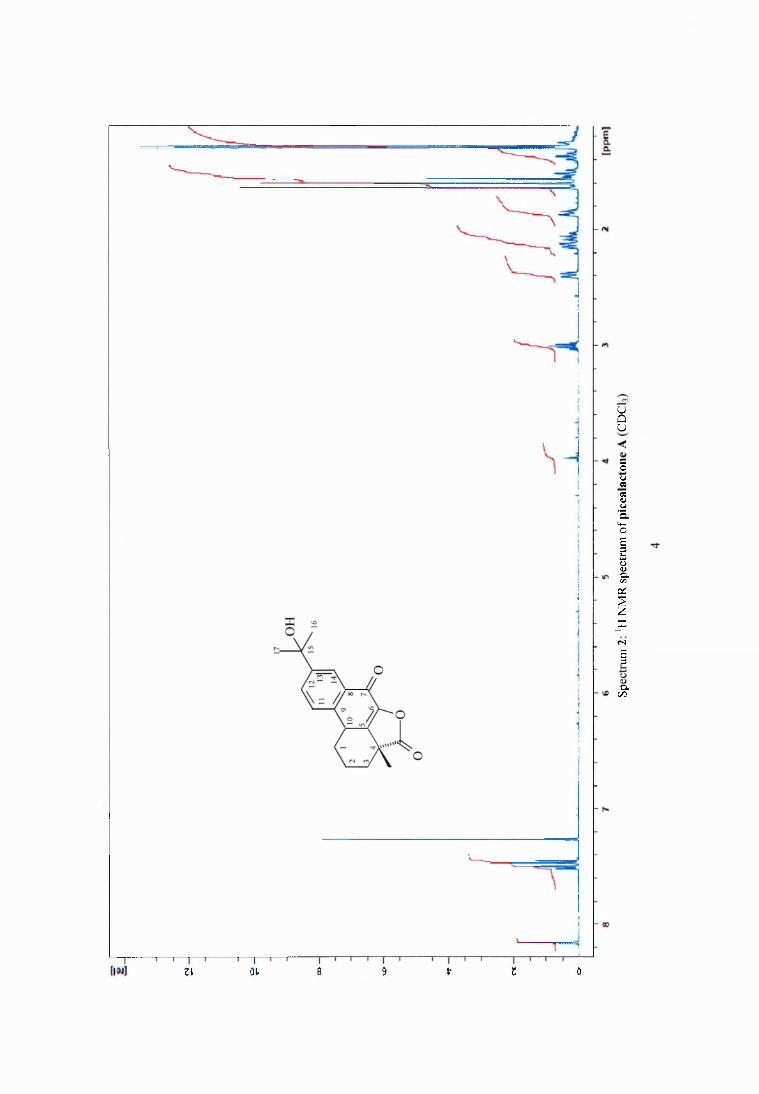
901. 8 tr Z 0z\
uûu
cou«S•I<4-0
1s-aiszX< s
I
CuC /3
Abundance Average of 13.701 to 13.730 min.: MS Vanilin.D\data.msis;.o
900000
800000
700000
600000
500000
400000
300000
200000
100000
000000
900000
800000
700000
600000
500000
81.0400000
109.0300000
20000053.0
100000
252.9168.1 190.9207.0222. 280.9100 120 140 160 180 200 220 240 260 280 300
Spectrum 3.1: Mass spectrum o f vanillin (CDCI3)
5
uÛu
&R>0
1&
sX
<s
:
[|9J] 02 St 01.
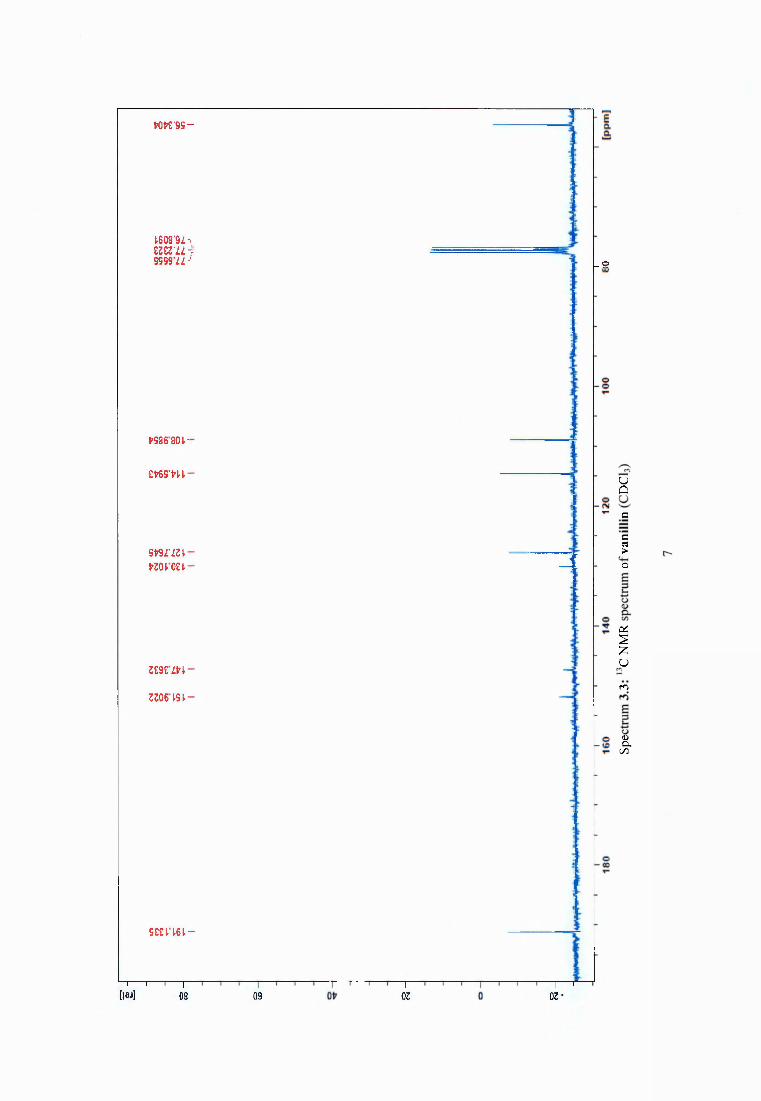
W3K-9S-
t608'9Z-., iZZZ'LL-r SSS9 Z2 J
PS86 801-
Ewsm
swrzzitzoioci
ZESEZtl
ZZ06191
a
_ g
UQ
’■ ccR>O
Qi2ZU
n
<u(%
SEEIWGI-
T r[|0J]
T08 09
TOZ
1 r OZ-
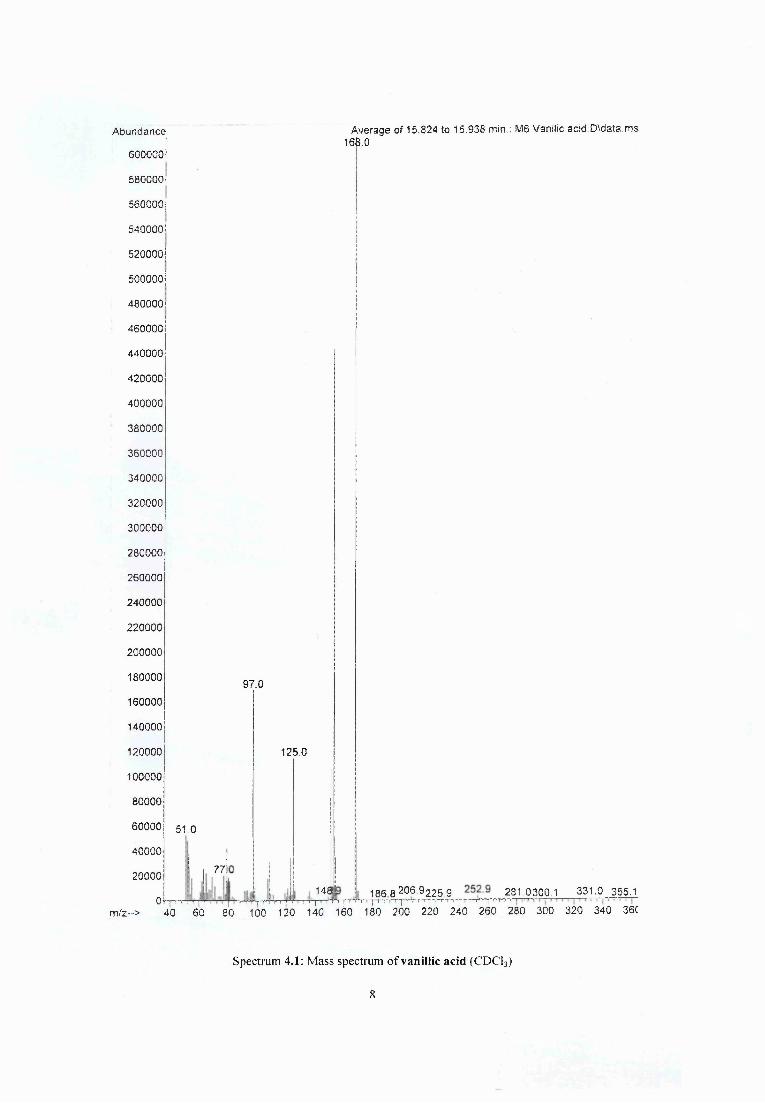
AbundanceI
600000j
580000!
560000j
540000Î
520000|
500000{
4800001
460000
440000
420000
400000
380000
360000
340000
320000
300000
280000
260000
240000
220000
200000
180000
160000
140000
120000
100000
80000
60000
40000
20000
0
Average of 15.824 to 15.938 min.: M6 Vanilic acid D\data.ms168.0
5 1 0
9 7 0
77
1250
148 8 206.9225.9 281.0300.1 3 8 1 0 355.11WSU ' T ' ! V T ' T T I— I T T - r r - ^ 1 ■ r-T -r -T - i- j - r T - n - j - i — r r - | - r f - ! ~ r j r - i ' f r - f , , , ' | .
m /z-> 40 60 80 100 120 140 160 180 200 220 240 260 280 300 320 340 36C
Spectrum 4.1: M ass spectrum o f vanillic acid (CDCI3)
R
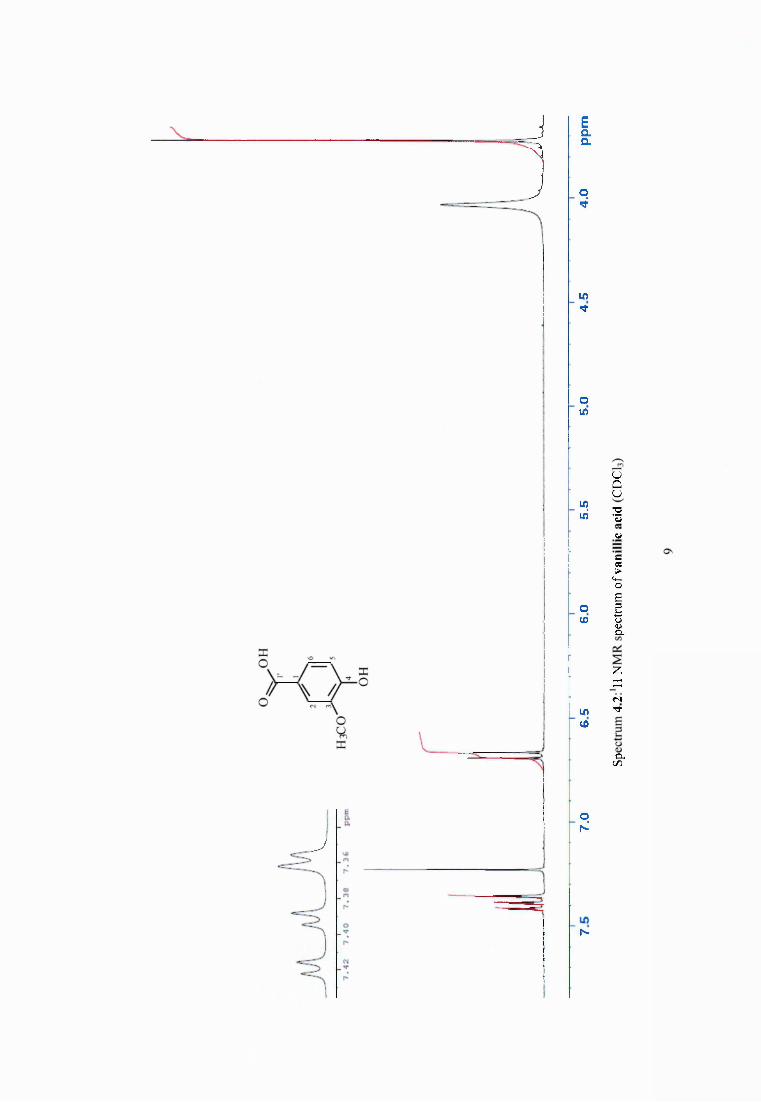
J]
EQ.a
q4"
in4"
oin
qin
uQU
•1
u
OCO
sCQ><+-0
1<L>Cl
. J
inCO
qN
ZX«STf
d.on
N
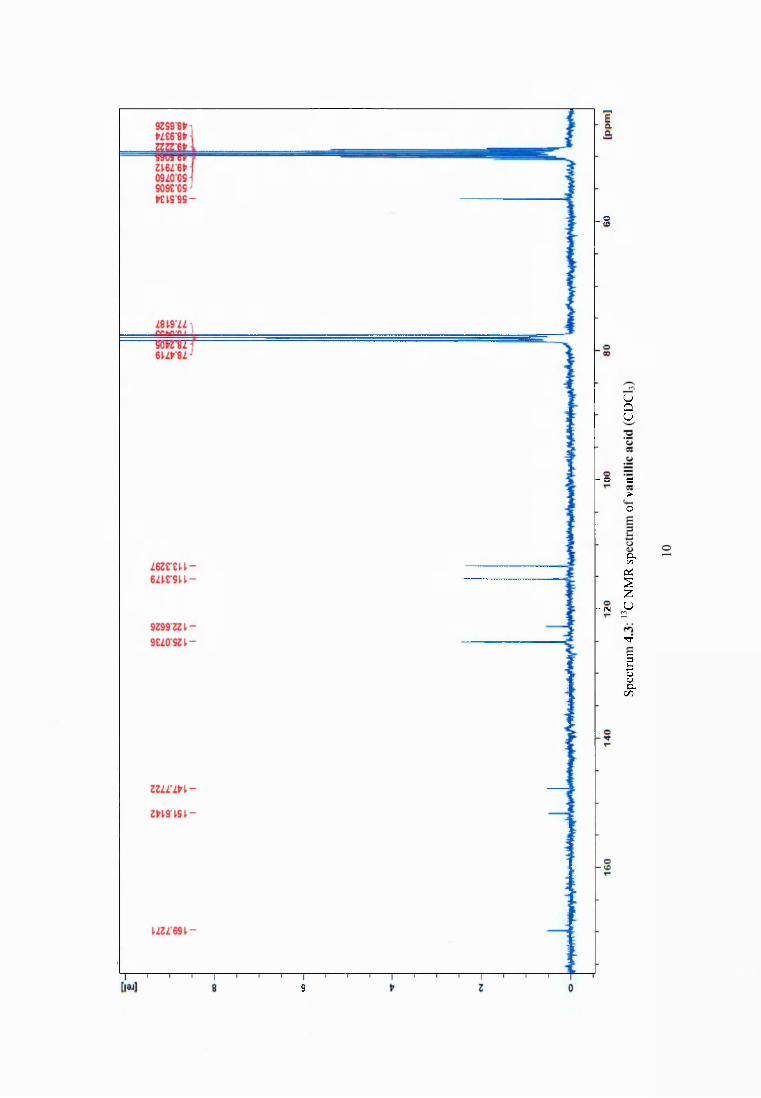
- 1
- o
- fN
9
UQU
c>o
&aiSZU
TT
iQ.ùO
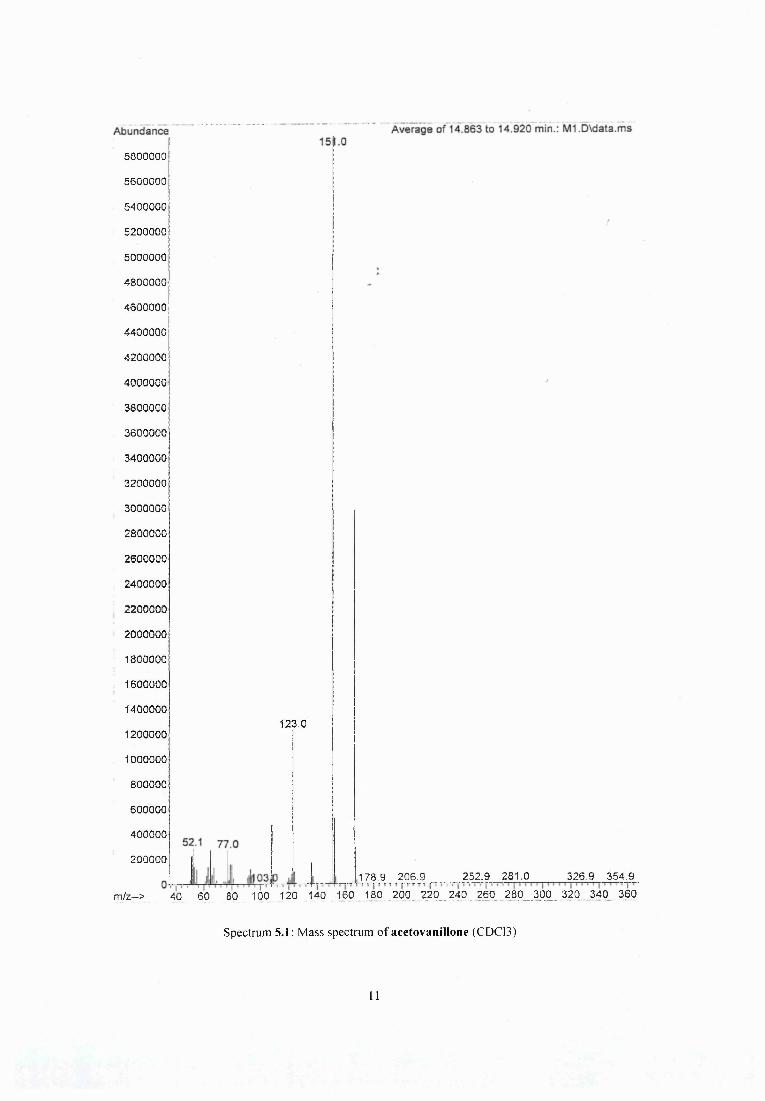
5800000
5600000
5400000
5200000
5000000
4800000
4600000
4400000
4200000
4000000
3800000
3600000
3400000
3200000
3000000
2800000
2600000
2400000
: 2200000 :
2000000
1800000
1600000
1400000123.0
1200000
1000000
800000
600000
400000
200000326.9 354.92 5 2 9 2 8 1 0178.9 206.9
40 60 80 100 120 140 160 180 ,2qq_ 2 2 p _ 2 4 q 2 6 q _ 2 8 p _ 3 p g _ 320 340_ 360m/z-->
Spectrum 5.1: Mass spectrum o f acetovanillone (CDC13)
11
uQ
O)cs'a
Ica0 E
1CL
ZXfSiX£3
CLüC
[|aj] ZL 01
__
uÛu
1«3b£
I0 m &SZX
1
CLüO
[IM 01.
Abundance
8000000
7500000
7000000
6500000
6000000
5500000
5000000
4500000
4000000
3500000
3000000
2 500000
2000000
1500000
1000000
500000
0
51.1
89.0
180
152.1
115.0
Scan 2528 (17 .552 min): M4 stilbene.DXdata.ms
207.0 253 .0 281.0 327 .0
m /z -> 40 60 80 100 120 140 160 180 200 220 240 260 280 300 320 340
Spectrum 7.1 : Mass spectrum o f stilbene (CDCI3)
14
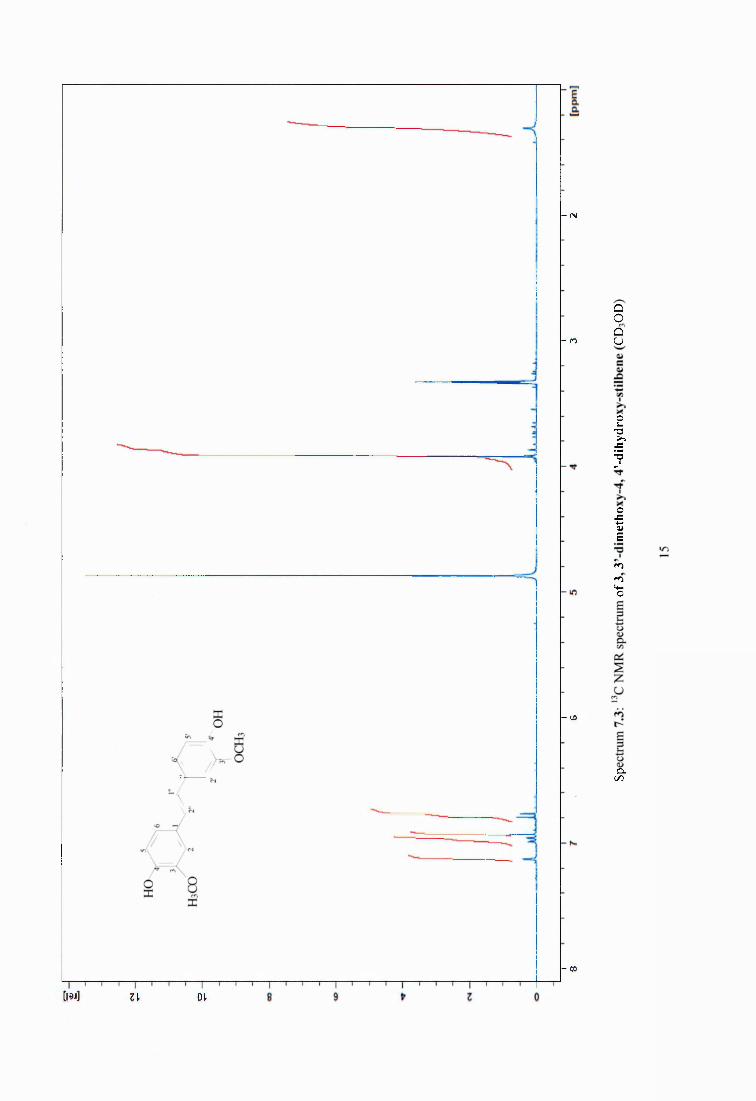
- (\
- m
- to
- »
aoQu
213. ty
01E
rorn«4-o
D-oo
[|9J] Zl 01
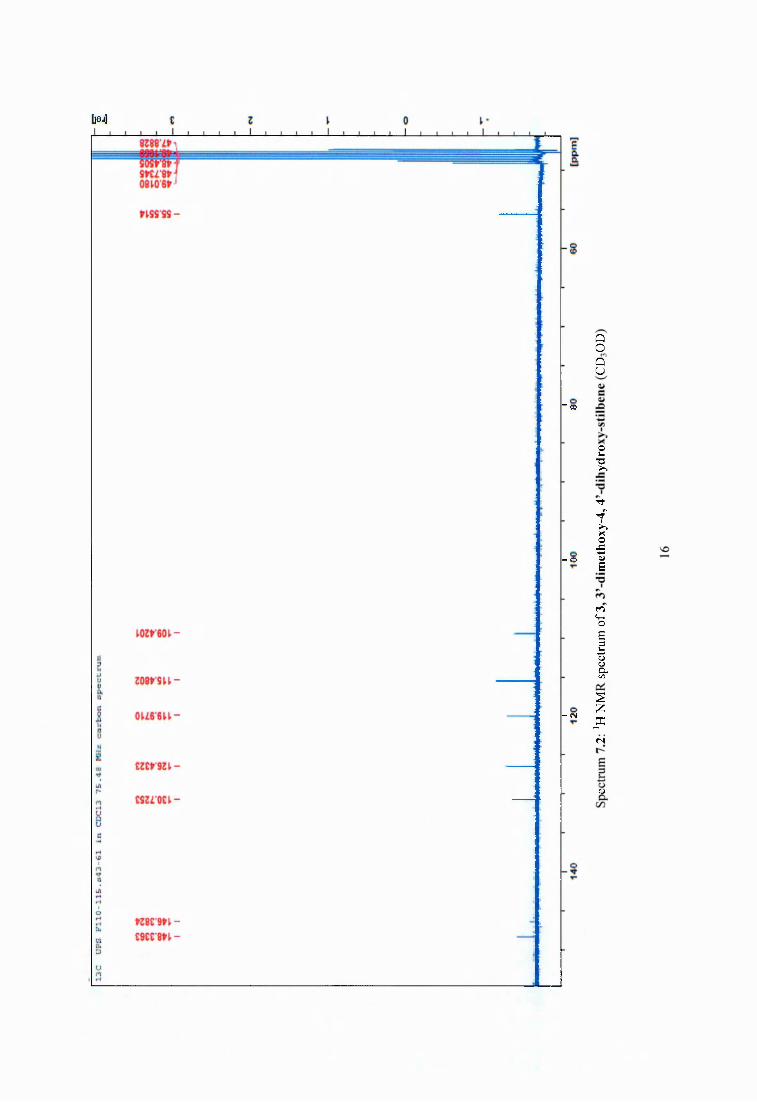
l|OJ]
- to
- o
- <M
QOOU
£%
TT-rfk01S
m(4-,0 E
1(U&
SzJE<s
<u
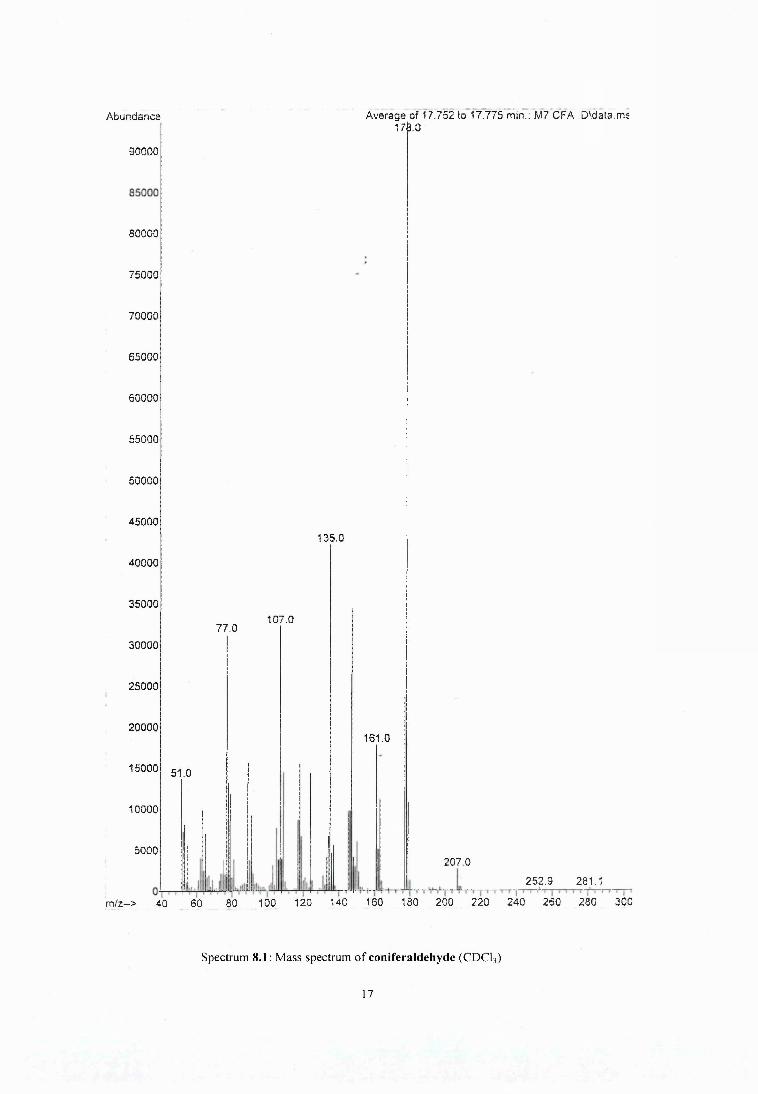
Abundance
90000
80000
75000
70000
65000
60000
55000
50000
45000
40000
35000
30000
25000
20000
15000
10000
5000
51.0
Average of 17.752 to 17.775 min.: M7 CFA .DXdata.m;178.0
135.0
77.0107.0
161.0
207.0
252.9 28 1 .1
m /z-> 40 60 80 100 120 140 160 180 200 220 240 260 280 300
Spectrum 8.1: Mass spectrum o f coniferaldehyde (CDCI3)
17
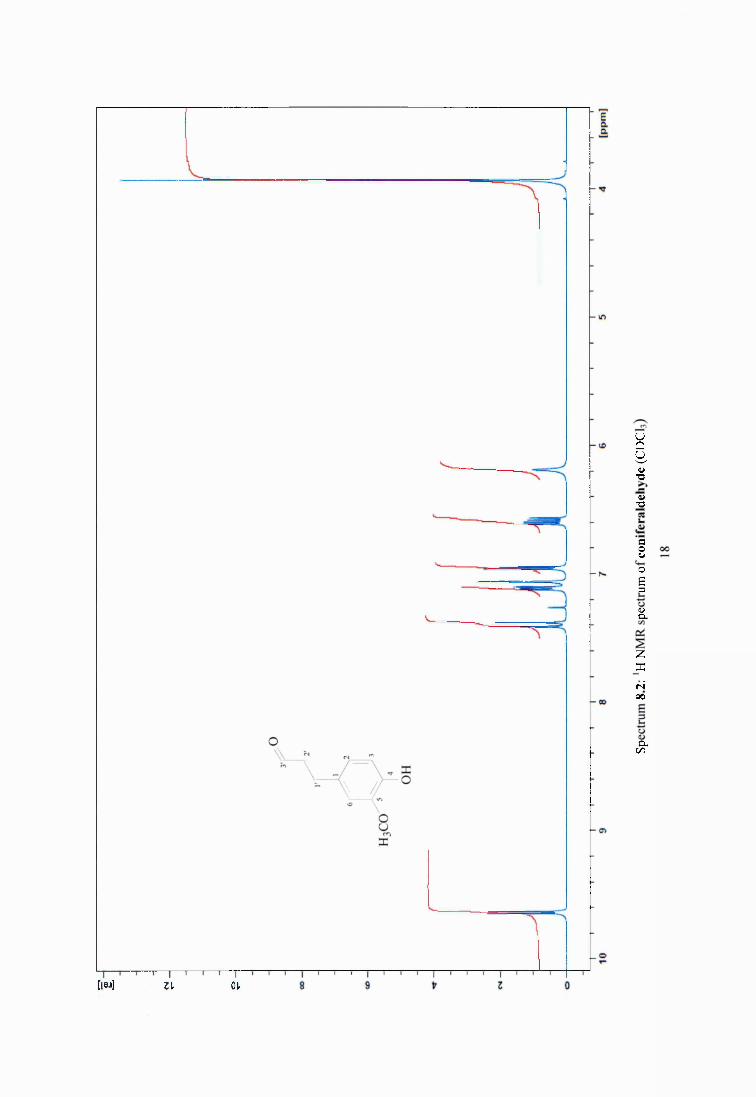
oÛs«■o
«■a
sou010)a .
ZX
rioc
CLOO
[|3J] 21. 01.
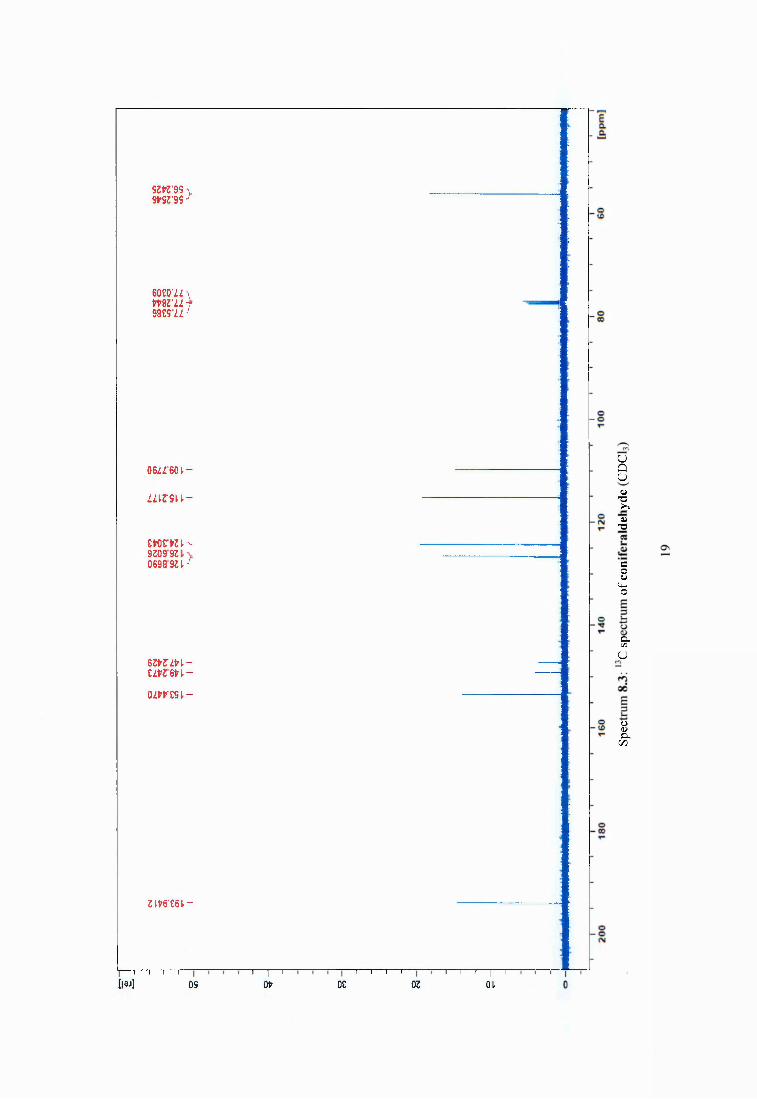
S2W9S V 9t'S2’9S "
6090"221, mZ'ZiT 9899/2̂
0622 601-
2 2 kZ'S^-
9W9'Wt - 9209 92 0698 921^
UQU4̂%«■o
cou(4-o
BZVZ'IV̂ -iLn%n-OlfVÎSi-
&u
2 tWE6 l -
Koo
“I— I— I— r[|SJ] 09
1 I r Ofr
1—I—I—r09 02
1 I I I r
01
> \ f c > u r » c l a r i œ
Unknown: Scan 3655 (24,001 min): brW"G1’15-fib-da.D\data.«ns Compound In Library Factor = -235
143100-
121
207
15550-
171193
0ISO 210120 150
256
241
275
316
301
394
210 240 270 300 330 360 390
Hit 1 : Methyl abietateC21H3202; MF; 7S2; RMF; 8Û7; Pmb 59.5%; CAS: 127-25-3; Lib: repiib; ID: 28885.
1 0 0
121105
21350- 131 185143
157171 I 195
04-180 210150120
256
241
273
4' I 270
31.
O301-)l', îjl̂ I ; I I’.-30Ô 330 360 390
Spectra 9.1 : Mass spectrum o f m ethyl abietate (DCM)
20
Unknown: Scan 3803 (24.847 min): h-0l-15-ati-cta,0\data,iins Compound In Library Factor = 640
100 -
50-
197
141
55 81 , j l I I, J i l l f 1 211225
285
I 300
Jii. ̂Ji.̂ 39660 90 120 150 ISO 210 240 270 300 330 360 390
Hit 1 : Detrydroabtedc addC20H2SO2; MF: 924; RMF: 968; Prob 63.6%; CAS; 1740-19-8; Lib: matnlib; 10:194581.
100-
50-
197
55 69 B'iVJ . .,1. 1,1,.. ?...
^^^55 Î IT169183M l 'lib -i
239I
257271211225
60 90 120 150 180 210 240 270 300 330 360 390
285
V V, . ^ x o r
300
z n I 7-^VV V"l ' < I I
OH
Spectra 9.2: Mass spectrum o f dehydroabietic acid (DCM)
2 1
/ k b u n d a n c
Unknown: Sean 527 {§.102 min): brw_<tam2.D\data.ms Compound In Library Factor = 101
100-
50-
83
77
53 .,1,1
60 60
sr
70 80
91
89■ • ’T90
> I ' 100
105 121
u s iig l^
Hitl : (iR)-2,6,§-Triïïî0thyiblcydoJ3.1.1ltifipt-2-eneC10H16; MF: 957; RMR 059: Pmb 25.2%; CAS: 7785-70-8; Lib: repiib; ID: 13150.
136
110 120 130 140 150
100-j
504
93
53, I
50
67
60 70
79
105 121
115119|.136
80 100 110 120 130 140 150
Spectra 9.3: Mass spectrum o f a-p inene (DCM )
22
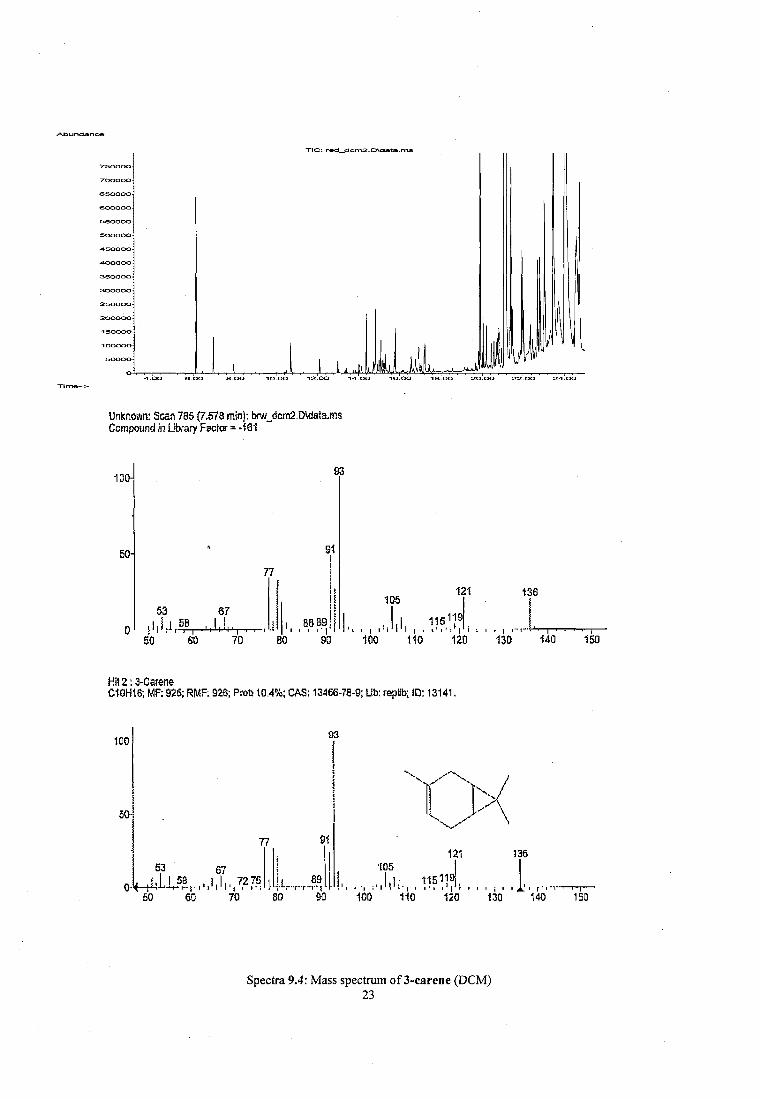
/ A b u n d a n c e
Unknown: Scan 795 {7,578 min); brw_ {̂Jcm2.D\data,ms Compound in Library Facfor = -101
100-
50-
53
50 60
67u
77
93
60
01
| i89 89 i. .- |'i
90
105
. , .
100 110
115
121
119 = . 12 0
136
, I l-T-130 lîô" 150
Hit 2 : 3-CareneC1ÛH16; MF: 925; RMF; 926; Pmb 10.4%; CAS; 13466-76-0; Lib: æpiib; ID: 13141.
100
60-
77
5358
5772 75
I-- 5 • I ' I ' I M ' i I I - ;60 70
03
91
89r ' r " I "' I
80 90
121105
, i ' IMI -1 1 0 0
11511
136
110 120 1301 r - 1 - -I—<—T-i—140 150
Spectra 9.4: Mass spectrum o f 3-carene (DCM) 23

24




















