
Detection and phylogenetic identification of labeled prokaryotic cells on mineral surfaces using...
11
Detection and phylogenetic identification of labeledprokaryotic cells on mineral surfaces using Scanning X-ray Microscopy Bénédicte Ménez a , , Céline Rommevaux-Jestin a , Murielle Salomé b , Y uheng Wang a , Pascal Philippot a , Alain Bonneville a , Emmanuelle Gérard a , c a IPGP , CNRS-UMR 7154/ Centre de Recherchessur le Stockage Géologique du CO 2 (IPGP/TOTAL/SCHLUMBERGER), Paris, France b European Synchrotron Radiation Facility, Grenoble, France c ESE, CNRS UMR 8079, Université Paris-Sud, Orsay, France Received6 July 2006; received in revised form 1 February 2007; accepted 10 February 2007 Editor: D. Rickard Abst ract The involvement of intraterrestrial microbes ingeochemical cycles is nowwell recognized. However,owing to the small number of appropriate methods for probing these ecosystems, theexploration of their metabolic diversity, energy sources, and biogeochemicaltransformations remains limited. Here we demonstrate theabilityof scanning X-ray microscopy using synchrotron radiation to localize and characterize the phylogeneticaffiliation of individual prokaryotic cells on variousmineral surfaces (e.g. carbonates,basaltic glass) when combined with a newlydevelopedprotocol based on fluorescence in situ hybridization coupled to ultra-small immunogold. The possibility to associate simultaneously the phylogenetic identification of microorganisms with the chemical and structural characteristics of associated mineral phases (i.e. inorganic substrateand biomineralizations),offers great interest for assessing the geochemicalimpact of subsurface microbial communities and unraveling microbe-mineralinteractions in the deepbiosphere. © 2007 Elsevier B.V . All rights reserved. Keywords: Prokaryotic cells; Mineralized environment; X-ray microscopy; Synchrotron radiation; Fluorescent in situ hybridization; Immunodetection 1. In t roduc ti on Itis nowwell recognized thatthe Earth subsurface is a major habitat for prokaryotes, and the number of microbes thatthrive deepbelow the ground probably exceeds the numberfound inother components of the biosphere (Whitman et al., 1998). Indeed, microbial lifecolonized all lithosphericenvironments wherever carbon and energy sources, subsurface porosity, and temperature permit. As highlighted these last years in several reviews (Kieft and Phelps, 1997; Krumholz, 2000; Pedersen, 2000; Fre- drickson and Onstott, 2001; Parkes and Wellsbury, 2004; Amend and T eske, 2005), the deep subsurface biosphere contributessignificantly to the overall biomass and biodiversity on Earth. The study of the deepbiosphere therefore has importantimplications for our understand- ing of global biogeochemical cycles and interactions between the biosphereand the geosphere. Our knowledge is however limited by the inaccessibilityof subsurface Chemical Geology 240 (2007) 182 – 192 www.elsevier .com/locate/chemgeo Corresponding author . Current address: IPGP , Equipe Géobio- sphère Actuelleet Primitive, case 89, 4 place Jussieu, 75252 Paris cedex 05, France. Tel.: +33 1 44 27 77 23; fax: +33 1 44 27 99 69. E-mail address: menez@ipgp.jussieu.fr (B. Ménez). 0009-2541/$ - see front matter © 2007 Elsevier B.V . All rights reserved. doi:10.1016/j.chemgeo.2007.02.009
Transcript of Detection and phylogenetic identification of labeled prokaryotic cells on mineral surfaces using...

Detection and phylogenetic identification of labeled prokaryotic cellson mineral surfaces using Scanning X-ray Microscopy
Bénédicte Ménez a, , Céline Rommevaux-Jestin a, Murielle Salomé b, Yuheng Wang a,Pascal Philippot a, Alain Bonneville a, Emmanuelle Gérard a,c
a IPGP, CNRS-UMR 7154/ Centre de Recherches sur le Stockage Géologique du CO2 (IPGP/TOTAL/SCHLUMBERGER), Paris, Franceb European Synchrotron Radiation Facility, Grenoble, Francec ESE, CNRS UMR 8079, Université Paris-Sud, Orsay, France
Received 6 July 2006; received in revised form 1 February 2007; accepted 10 February 2007
Editor: D. Rickard
A bstract
The involvement of intraterrestrial microbes in geochemical cycles is now well recognized. However, owing to the smallnumber of appropriate methods for probing these ecosystems, the exploration of their metabolic diversity, energy sources, andbiogeochemical transformations remains limited. Here we demonstrate the ability of scanning X-ray microscopy using synchrotronradiation to localize and characterize the phylogenetic affiliation of individual prokaryotic cells on various mineral surfaces (e.g.carbonates, basaltic glass) when combined with a newly developed protocol based on fluorescence in situ hybridization coupled toultra-small immunogold. The possibility to associate simultaneously the phylogenetic identification of microorganisms with thechemical and structural characteristics of associated mineral phases (i.e. inorganic substrate and biomineralizations), offers greatinterest for assessing the geochemical impact of subsurface microbial communities and unraveling microbe-mineral interactions inthe deep biosphere.© 2007 Elsevier B.V. All rights reserved.
Keywords: Prokaryotic cells; Mineralized environment; X-ray microscopy; Synchrotron radiation; Fluorescent in situ hybridization;Immunodetection
1. Introduction
It is now well recognized that the Earth subsurface is amajor habitat for prokaryotes, and the number ofmicrobesthat thrive deep below the ground probably exceeds thenumber found in other components of the biosphere(Whitman et al., 1998). Indeed, microbial life colonized
all lithospheric environmentswherever carbon and energysources, subsurface porosity, and temperature permit. Ashighlighted these last years in several reviews (Kieft andPhelps, 1997; Krumholz, 2000; Pedersen, 2000; Fre-drickson and Onstott, 2001; Parkes and Wellsbury, 2004;Amend and Teske, 2005), the deep subsurface biospherecontributes significantly to the overall biomass andbiodiversity on Earth. The study of the deep biospheretherefore has important implications for our understand-ing of global biogeochemical cycles and interactionsbetween the biosphere and the geosphere. Our knowledgeis however limited by the inaccessibility of subsurface
Chemical Geology 240 (2007) 182 – 192www.elsevier.com/locate/chemgeo
Corresponding author. Current address: IPGP, Equipe Géobio-sphère Actuelle et Primitive, case 89, 4 place Jussieu, 75252 Pariscedex 05, France. Tel.: +33 1 44 27 77 23; fax: +33 1 44 27 99 69.
E-mail address: [email protected] (B. Ménez).
0009-2541/$ - see front matter © 2007 Elsevier B.V. All rights reserved.doi:10.1016/j.chemgeo.2007.02.009

microbial niches, the omnipresent risk of contaminationand the low number of appropriate methods for the in situprobing of these ecosystems.
One of the key challenges in the understanding of thecomplexities of the subsurface and its microbialinhabitants is the exploration of the metabolic diversityof the prokaryotic populations, their energy sources, andbiogeochemical transformations. Molecular methods areparticularly useful for localizing and identifying thephylogenetic affiliation of microorganisms in theirnatural environments and have become an essentialtool in microbial ecology. In particular, fluorescentlylabeled rRNA-targeted nucleic acid probes allow torapidly determine specifically the abundance, location,and activity of individual microbial cells in situ (Amannet al., 1997). However, fluorescence in situ hybridiza-tion (FISH) has several limitations with regards to thestudy of microbes–mineral interaction and particularlythe evaluation of the impact of microorganisms on theformation or dissolution of minerals. These limitationsinclude the autofluorescence of the surrounding miner-alized environment and the inability to simultaneouslyobtain chemical, crystallographic or spectroscopicinformation on associated mineral phases.
Over the past decade, advances in spectroscopictechniques have provided a large variety of opportunitiesto identify new microbiologically-mediated processesinvolving minerals, as recently reviewed in Geesey et al.(2002). In particular, owing to its high sensitivity, non-destructive character, high spatial resolution, and in situmulti-element analytical capability, third generationsynchrotron light sources have been recognized as apowerful and promising new tool. Several methods im-plementing these small and brilliant X-ray beams weresuccessfully applied to image individual cells. The mostimportant ones include (cryo-) scanning transmission X-ray microscopy (STXM) using soft X-rays which allowsto investigate single cells in 2D (Benzerara et al., 2005b)or 3D (Larabell and Le Gros, 2004), synchrotron infraredmicrospectrometry (Jamin et al., 1998) and single-particleX-ray diffraction (Miao et al., 2003). X-ray imaging basedon X-ray fluorescence measurements has also been per-formed on whole cells with low energy X-rays (i.e. a fewkeV; Ueki et al., 2002; Foriel et al., 2004; Lemelle et al.,2004). In the hardX-ray range, single human (Bohic et al.,2001), yeast (Ortega et al., 2004), protist (Twining et al.,2003), and prokaryotic (Kemner et al., 2004) cells weremapped at the micrometric level, thus allowing the celllocalization and identification by its light-elementcontent. Notably, Kemner et al. (2004) succeeded inlow-Z element localization of individual hydratedbacterial cells at a spatial resolution of 150 nm. However,
the imaging approaches developed in these studies are notspecific and consequently, do not permit cell phylogeneticaffiliation. Moreover, the inframicrometric sample thick-ness required by some of the experimental set-up (e.g.STXM) does not allow to work on mineral surfaceswithout particular sample preparation (e.g. focused ionbeam system, Benzerara et al., 2005a).
Coupling cell hybridization with specific ribosomalRNA targeted probes to spectroscopic techniquesshould open new venues of research in microbialphysiology and ecology, particularly in the domain ofgeomicrobiology. It will allow the coupling of thephylogenetic identification of individual cells with thechemical and structural characterization of the mineralenvironment surrounding the cells. This direct mineralenvironment could act as a metabolic reactant and/orcould have formed as a product of metabolic activity(biomineralization). We present here the first successfulattempt at localizing and phylogenetically identifyingprokaryotic cells in a mineralized environment bycombining Scanning X-ray Microscopy (SXM) usingsynchrotron radiation with a newly developed protocolutilizing FISH technique coupled to ultra-small immu-nogold detection (Gérard et al., 2005). Sensitivity andresolution of this method were evaluated on the X-rayMicroscopy beamline ID21 of the European Synchro-tron Radiation Facility (ESRF, Grenoble, France) wherewe demonstrated the efficient specific identification ofsilver–gold-labeled cells on a variety of commonmineral substrates such as Ca- and Fe-carbonates, quartzand basaltic glasses.
2. M ater ials and methods
2.1. Sample preparation
To evaluate the ability of SXM to localize andinvestigate the phylogenetic affiliation of prokaryoticcells on mineral surfaces, we used a simple modelcomposed of a mixture of cultivable bacteria from dif-ferent taxonomic divisions, readily distinguishable bytheir morphologies, as described in detail by Gérard et al.(2005). Escherichia coli (small rod) and Deinococcusradiodurans (large cocci), belonging respectively to theGammaproteobacteria subdivision and the Thermus/Deinococcus group, were selected for this purpose. Thecells were deposited on different types of substratesincluding polymer films (i.e. Ultralene® films), glassslides, basaltic glasses, calcite (CaCO3), magnesiansiderite ((Fe, Mg)CO3), and quartz (SiO2). Followingthe protocol described in Gérard et al. (2005), cells weregrown in appropriate media and chemically fixed with
183B. Ménez et al. / Chemical Geology 240 (2007) 182–192

formaldehyde (methanol free, Ultra Pure; Polysciences).A 1:1 mix of E. coli:D. radiodurans cells suspended inphosphate-buffered saline pH 7.2 (PBS; 130 mM NaCl,7 mM Na2HPO4, 30 mM NaH2PO4) with 0.5% lowmelting agarose (Seaplaque GTG® agarose) weredeposited on the substrates of interest. Cells were sub-sequently prepared for hybridization. This impliessequential dehydration with ethanol and permeabiliza-tion by mean of lysozyme solution and proteinase K.
We then hybridized universal and specific fluorescein-labeled oligonucleotide probes to the ribosomal RNA ofour cell mixture. The different probes used in separateexperiments were: fluorescein-labeled S-D-Bact-0338-a-A-18 (EUB338, 5 -GCTGCCTCCCGTAGGAGT-3 ) forBacteria, fluorescein-labeled L-C-gProt-1027-a-A-17(GAM42a, 5 -GCCTTCCCACATCGTTT-3 ) for Gam-maproteobacteria and the fluorescein-labeled non-Bact338 (NONEUB, 5 -ACTCCTACGGGAGGCAGC-3 ) as negative control (Loy et al., 2003). In practice, theEUB338 probe hybridises the ribosomal RNA ofboth species present in the mixture (i.e. E. coli andD. radiodurans) whereas the GAM42a probe only bindsthose of E. coli. When the NONEUB probe is used, noneof these species are hybridized. To check the attainment ofthis stage, control samples were DAPI stained andhybridized cells were observed in epifluorescence undera Zeiss Axioplan 2 Imaging microscope using specificfilters for different wavelength emission.
We finally used antibodies against fluoresceincoupled to subnanometer gold particles to label thehybridized probes in the ribosome (Ultra-Small Immu-nogold Mouse-anti-FITC, Aurion). To favor the anti-bodies penetration within the cells, the used goldparticles displayed an average diameter of 0.8 nm. Inorder to increase the size of the metal particles and inaccordance, optimize their detection, a subsequent stepof silver enhancement was carried out using the kit R-GENT SE-EM (Aurion). Silver enhancement reaction isa gold particle-catalyzed reduction in which silver ionsare reduced to metallic silver with a photographic de-veloping compound as the electron source.
Control samples were dehydrated with ethanol andcarbon coated for visualization of the specific gold–silversignal by Scanning Electron Microscopy (SEM), thusensuring reliable samples for X-ray microscopy prior toexperiment. Observationswere carried out on a JEOL JSM840A microscope operating at 20 kVaccelerating voltage.
2.2. X-ray microscopy
Experiments were carried out with the Scanning X-rayMicroscope operated at the ID21 beamline of the ESRF
(Grenoble, France). The ID21 beamline is dedicated toX-ray imaging andmicrospectroscopy in the range from2 to 7 keV, in both absorption and fluorescence modes(Susini et al., 2002). This energy range covers theK absorption edges of medium-light elements (e.g. Al, Si,P, S, Cl, K, Ca, Sc, Ti, V, Cr, Mn, and Fe) and the L or Mabsorption edges of heavier elements. This includes theL-absorption edges of Ag andM-absorption edges of Au,which are important for our work. Moreover, X-raymicroscopy in this energy range offers great advantages interms of penetration depth for the observation of bio-logical specimens up to 10 m-thick. In addition, phasecontrast X-ray imaging that exploits the differences in therefractive index of soft tissues can be used as a sensitivetechnique for visualizing phase information from lowabsorbing samples, which could not be imaged in simpletransmission mode, and as a result, allows the localizationof the cells (Wilkins et al., 1996; Wilhein et al., 2001;Kaulich et al., 2002; Di Fabrizio et al., 2005).
The energy of the X-ray beam emerging from thelinear undulator at the storage ring is selected by a fixedexit double crystal monochromator (Si b111N). Toensure good fluorescence yield and optimize thedetection of the elements of interest, the beam energywas set to 3.95 keV, above the L edges of Ag. A Fresnelzone plate, in association with a small pinhole aperture,was used to focus the X-ray beam on the sample down toa beamsize of 0.8 0.3 m2 (horizontal by vertical,full width at half maximum, FWHM). The transmittedX- rays are detected by a Si photodiode mounted down-stream of the sample holder, Samples under investigationwere positioned in the image plane at 60° with respect tothe incident photon beam. X-Ray Fluorescence (XRF)emerging from the samples as a result of the photonicexcitation is analyzed using a high resolution germaniumsolid state detector of 135 eV resolution (measured onMn K line with a shaping time of 6 s at an input countrate of 1000 counts/s), mounted in the horizontal planeperpendicular to the beam and calibrated with silverstandards. The energies of the emitted X-rays lines arecharacteristic of a given atom and their intensity isproportional to its concentration in the specimen. Toobtain phase contrast X-ray images, a 50 m aperturewas placed in front of the transmission photodiode,160 mm away from the zone plate focal plane andpartially covering the diverging beam cone generated bythe zone plate. This pinhole was used to convert themicrobeam deviations caused by the change of refractionindex when passing through the cells into variations ofintensity. To detect light-element fluorescence andminimize the absorption of the incident and emitted X-rays in air, measurements were performed in vacuum.
184 B. Ménez et al. / Chemical Geology 240 (2007) 182–192
Each sample was scanned horizontally and verticallyusing a piezoelectric stage and both the transmissionsignal and the fluorescence spectra were recorded toproduce a set of images. For each pixel of the maps, a fullfluorescence spectrum was available. The overallcollected XRF spectra were fitted using the spectralnon-linear least-squares deconvolution algorithm of theAXIL program package (Vekemans et al., 1994).Elemental images, produced by using the K lines,were then normalized by the intensity of the incomingbeam. Flat field correction was applied on phase contrastX-ray images.
3. Results
To check the sensitivity of the method and detectpotential interferences, we proceeded sequentially by
using various types of substrates. In particular, westarted with thin Ultralene® film that allows combiningphase contrast X-ray imaging with X-ray fluorescencemeasurements, owing to its thickness (i.e. 4 m) and itsX-ray low absorption. For the other substrates, espe-cially for minerals, the absorption of the incident X-raysis too strong to permit measurements in transmission.
3.1. Specimens on polymer films
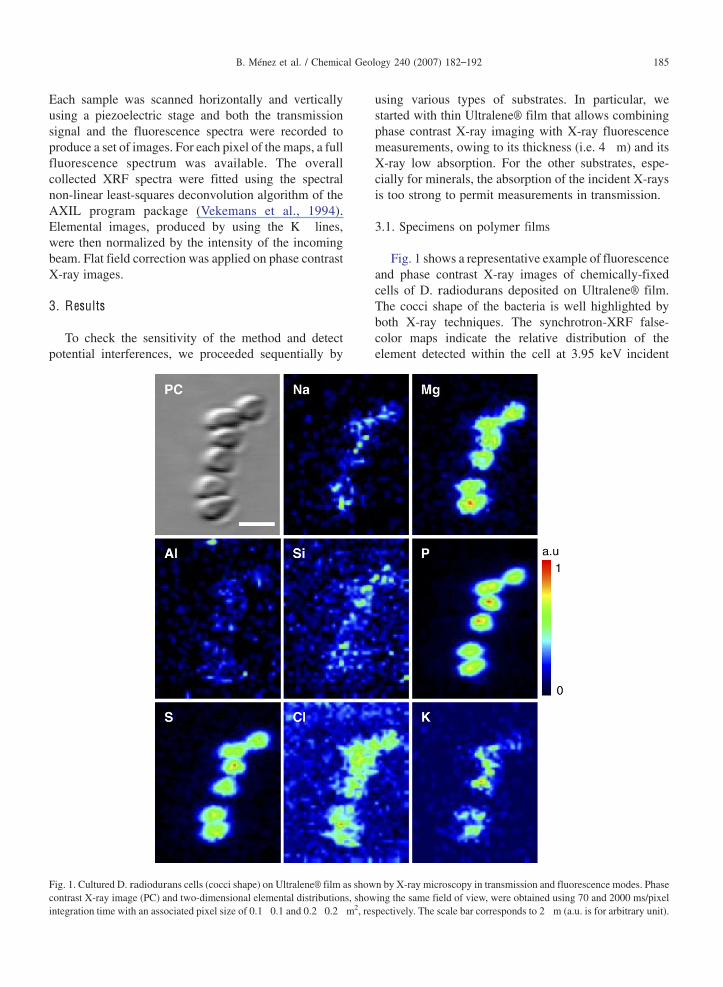
Fig. 1 shows a representative example of fluorescenceand phase contrast X-ray images of chemically-fixedcells of D. radiodurans deposited on Ultralene® film.The cocci shape of the bacteria is well highlighted byboth X-ray techniques. The synchrotron-XRF false-color maps indicate the relative distribution of theelement detected within the cell at 3.95 keV incident
Fig. 1. CulturedD. radiodurans cells (cocci shape) on Ultralene® film as shown by X-ray microscopy in transmission and fluorescence modes. Phasecontrast X-ray image (PC) and two-dimensional elemental distributions, showing the same field of view, were obtained using 70 and 2000 ms/pixelintegration time with an associated pixel size of 0.1 0.1 and 0.2 0.2 m2, respectively. The scale bar corresponds to 2 m (a.u. is for arbitrary unit).
185B. Ménez et al. / Chemical Geology 240 (2007) 182–192

photon energy, which include Na, Mg, Al, Si, P, S, Cl, K.Among these, Mg, P, S, Cl, K, as major and minor cellcomponents are particularly concentrated whereas Na,Al, Si are only slightly detected and their distributiondoes not mimic the cell morphology. For Na, this is likelydue to its weak fluorescence yield that does not allowaccurate XRF measurements in the present case. For Siand Al, the limiting parameter could be their very lowconcentrations (i.e. less than 0.01% of the cellular mass)close to detection limits. All these observations result inestablishing a correspondence between the spatiallocation of the bacteria and the collected elementalfluorescence that constitutes a chemical fingerprint of thebacteria. In order to evaluate the effects of cellpermeabilization on this signature, similar images wereacquired on E. coli cells on Ultralene® films. Results arepresented in Fig. 2 which highlights, as previously, avery high resolution of the technique together with agood agreement between phase contrast X-ray image andelemental maps. In particular, the rod shape of E. coli iswell represented. Only S and P display a high count ratewhereas Cl and K appear significantly less concentrated.Mg was not detected.
The subsequent step consists of combining heavymetal fluorescence signals emerging specifically fromthe nanoparticles to the light-element fluorescence. Forthis purpose, fluorescent in situ hybridization was carriedout in E. coli cultures with a probe specific for bacteria(EUB338) on Ultralene® films. We first tried to visualizethe labeled cells without silver enhancement duringinvestigations at the M-absorption edges of Au but we
failed to detect any X-ray fluorescence signal from gold.The tiny size of these particles (0.8 nm in diameter whichcorresponds to a number of atoms per cluster b30)together with the poor fluorescence yields at the M edgescontributed to the difficulty in detecting the gold nano-particles by this method. Consequently, as for electronmicroscopy (Gérard et al., 2005), a subsequent step ofsignal amplification by silver was necessary. Results arepresented in Fig. 3, where cell positions are located bythe phase contrast X-ray image. Owing to the successivewashing procedure imposed by the protocol, the contrastof the phase contrast X-ray image is not as good as in theprevious experiment but cell positions are neverthelessdistinguishable. The elemental distributions obtained influorescence mode allow discrimination between thehybridized cells (displaying a good correlation betweenAg, S, and P) and the dispersed Ag individual particlesand aggregates (arrowheads on Fig. 3) that show highand localized count rates and form the experimentalnoise. This background signal is characterized by ran-domly distributed silver particles and is inherent to theemployed method. It corresponds to autonucleation andenhancement of non-specific gold immunodetection.The reduction of this background noise without de-creasing the specific cell signal, constituted one of themost critical steps of the optimisation process of theimmunogold approach and is described in detail byGérard et al. (2005). It was minimized by optimizingreagent and buffer concentrations together with washingconditions after incubation but is still present in allsample preparations.
Fig. 2. Chemically-fixed and permeabilized E. coli cells (rod shape) on Ultralene® film as shown by X-ray microscopy in transmission andfluorescence modes. Phase contrast X-ray image (PC) and two-dimensional elemental distributions showing the same field of view were obtainedusing 100 and 2000 ms/pixel integration time, with an associated pixel size of 0.1 0.1 and 0.25 0.25 m2, respectively. The scale bar corresponds to2 m (a.u. is for arbitrary unit).
186 B. Ménez et al. / Chemical Geology 240 (2007) 182–192

3.2. Specimens on mineral surfaces
In order to verify that prokaryotic cells present enoughchemical contrast to be mapped on mineral surface, onedrop of suspended cells ofD. radiodurans was depositedon a doubly-polished siderite crystal. X-ray fluorescenceimages of cells of D. radiodurans, presented in Fig. 4,revealed the cocci shape of individual cells underlined bythe distribution of major and minor cellular componentssuch as Na, P, S, Cl, and K as well as trace amounts of Al
and Si. Note the inverse distribution of O andMg, mainlyarising from the substrate and whose characteristic X-raysare mostly absorbed by the overlying cells. The horizontalshift between positive and negative images is due to thedetection angle.
SXM observations on siderite were then carried outwith silver-enhanced ultra-small immunogold particlesin D. radiodurans cells hybridized with fluorescein-
Fig. 3. X-ray microscopy observations of silver-enhanced ultra-smallimmunogold particles in E. coli hybridized with fluorescein-labeledbacterial probe EUB338 on Ultralene® film. Phase contrast X-rayimage (PC) and fluorescence images showing the same field of viewwere obtained by scanning cells at 0.2 0.2 m2 pixel size with 70and 2000 ms/pixel integration time, respectively. The scale barcorresponds to 5 m. Elemental distributions allows distinguishinghybridized cells displaying a good correlation between S, P, andAg from Ag grains belonging to the background signal and indicatedby arrowheads (a.u. is for arbitrary unit).
Fig. 4. X-ray micro-fluorescence elemental maps of culturedD. radiodurans cells deposited on siderite. The scanning time foreach pixel is 2000 ms, with 0.2 0.2 m2 pixel size. The scale barcorresponds to 5 m. Stars show an example of a D. radioduransindividual cell (a.u. is for arbitrary unit).
187B. Ménez et al. / Chemical Geology 240 (2007) 182–192

labeled bacterial probe EUB338. One of the mainadvantages of using D. radiodurans is that this cell typecan be identified by its morphological characteristics:they divide so that alternating planes of synchronizedcell division give rise to characteristic tetrads. Threeexamples are compiled in Fig. 5 where the single cells ofD. radiodurans were clearly identified by theirmorphology and their gold–silver particles content.Cell positions are located by the S and P distributions,which display a good correlation with that of Ag. In thiscase, the silver signal is distributed throughout the entirecell. Few Ag particles belonging to the surroundingnoise and presenting high count rate, are distinguishablefrom hybridized cells. Note however that a loss of the
fluorescence signal, induced by the numerous stages ofwashing during sample preparation, is observed in allsamples, in particular for P.
Control specimens were also investigated. For exam-ples, fluorescence in situ hybridization was performedin D. radiodurans cultures with (1) the fluorescein-labeled GAM42a probe, specific only for Gammapro-teobacteria, and (2) the fluorescein-labeled non-specificprobe NONEUB. Results are presented in Fig. 6. Spotsof few hundreds of nanometers, presenting high countrates of Ag and Cl and corresponding to nanogoldparticles surrounded by metallic silver, are present inboth samples as background noise. These particles, asheavy metals, induce a strong absorption of the X-rays
Fig. 5. Triplicates X-ray microscopy observations of silver-enhanced ultra-small immunogold particles in single tetrads of D. radiodurans hybridizedwith fluorescein-labeled bacterial probe EUB338 on siderite (Panels (A), (B), and (C)). Synchrotron X-Ray Fluorescence elemental maps wereobtained by scanning the surface at pixel size of 0.33 0.33 m2 for panels (A) and (B) and 0.25 0.25 m2 for panel (C) with integration time of2000 ms/pixel for panels (A) and (B) and 3000 ms/pixel for panel (C). The scale bars correspond to 5 m for each (a.u. is for arbitrary unit).
188 B. Ménez et al. / Chemical Geology 240 (2007) 182–192

emerging from the substrate, thus leading to an inversedistribution for K. Nevertheless, the resolution of the X-ray microscope allows to visually determine that Ag and
Cl fluorescence signals are not co-localized with cellchemical fingerprint: as expected, the GAM42a andNONEUB probes do not bind the ribosomal RNA of D.radiodurans, recognizable by its tetrads and associatedP and S distributions. Note that Cl presents a strongaffinity for Ag in all samples. For panel B in Fig. 6, thebacteria appear larger than the cells on panel A. Thiscould be induced by a too long duration of the digestionstage during sample preparation causing damage of D.radiodurans cells and cytoplasm release. Despite theseuncertainties, Figs. 5 and 6 clearly prove that X-raymicroscopy allowed to discriminate the Ag-labeled fromthe unlabeled D. radiodurans cells against a backgroundof randomly distributed Ag-particles.
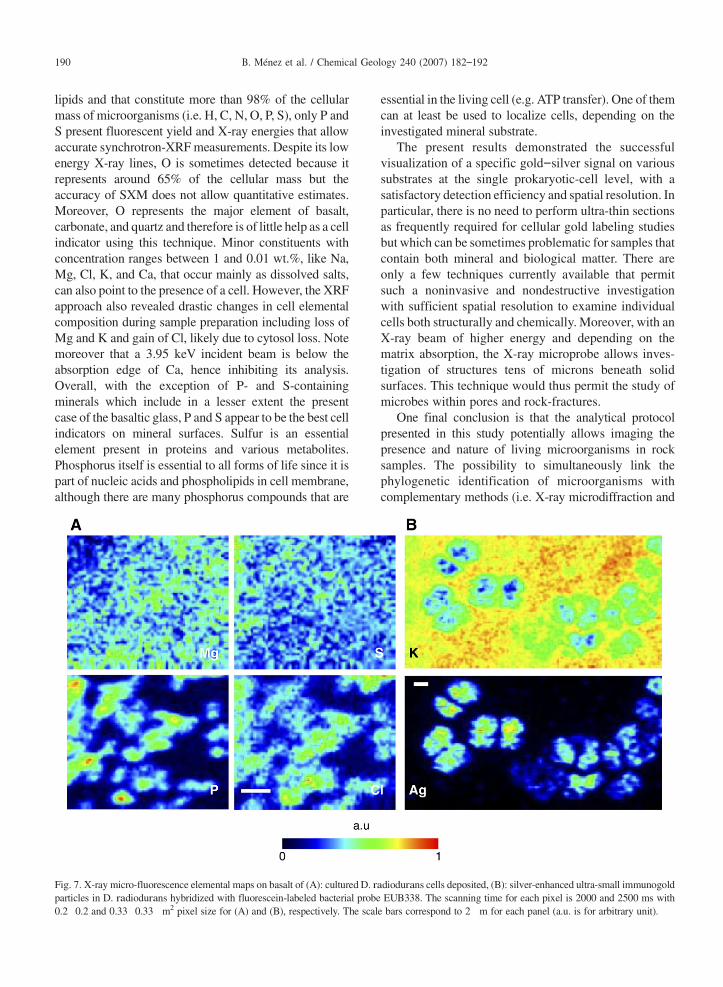
To enlarge the application field of this method, othersubstrates were investigated. As an example, Fig. 7 showsthe results obtained on basaltic glass following the sameprocedure described above for carbonates. Cells ofD. radiodurans were first deposited on a double-polishedchip of basaltic glass (Fig. 7A). Due to the complexbasaltic composition, the contrast is weaker than the oneobtained with the siderite. In particular, owing to the highS content of this basalt, resulting distribution maps do notconstitute in that case a strong indicator of cell presence.Mg associated with cells is also mapped with difficulty.Nevertheless, cells can be localized by means of P and Cldistributions. Fig. 7B corresponds to SXM observationobtained on silver-enhanced ultra-small immunogold par-ticles inD. radiodurans cells hybridized with fluorescein-labeled bacterial probe EUB338. As for Fig. 5, Ag dis-tribution covers the entire surface of single tetrads ofD. radiodurans thus clearly revealing the nature of thecells, whose positions are well indicated by the inverse Kdistributions.
4. D iscussion
In this study, we have shown that prokaryotic cellshybridized with fluorescein-labeled oligonucleotideprobes can be specifically detected on minerals byscanning X-ray microscopy after immunodetection byantibodies coupled to gold nanoparticles, enhanced bysilver. Practically, it implies associating heavy metalfluorescence signals emerging specifically from the gold–silver nanoparticles to the location of microbes, high-lighted by their light-element fluorescence (Mg, P, S, Cl,K). These spatial distributions constitute the cell chemicalfingerprint, firmly elaborated by taking advantage of themorphological and spatial information provided by thephase contrast X-ray imaging. Among the 6 majorelements that make up most biological moleculesincluding nucleic acids, proteins, polysaccharides, and
Fig. 6. X-ray microscopy observations of silver-enhanced ultra-smallimmunogold particles in D. radiodurans cells hybridized to fluorescentoligonucleotide probes on siderite. Panels correspond to cells hybridizedwith the fluorescein-labeled specific probe for GammaproteobacteriaGAM42a for (A) and with the fluorescein-labeled non-specific probeNONEUB for (B). Synchrotron X-Ray Fluorescence elemental mapswere obtained by scanning the surface at a pixel size of 0.2 0.2 m2 forpanels (A) and 0.25 0.25 m2 for panel (B) with integration time of2000 ms/pixel for panels (A) and 3000 ms/pixel for panel (B). The scalebars correspond to 2 m for each panel (a.u. is for arbitrary unit).
189B. Ménez et al. / Chemical Geology 240 (2007) 182–192
lipids and that constitute more than 98% of the cellularmass of microorganisms (i.e. H, C, N, O, P, S), only P andS present fluorescent yield and X-ray energies that allowaccurate synchrotron-XRFmeasurements. Despite its lowenergy X-ray lines, O is sometimes detected because itrepresents around 65% of the cellular mass but theaccuracy of SXM does not allow quantitative estimates.Moreover, O represents the major element of basalt,carbonate, and quartz and therefore is of little help as a cellindicator using this technique. Minor constituents withconcentration ranges between 1 and 0.01 wt.%, like Na,Mg, Cl, K, and Ca, that occur mainly as dissolved salts,can also point to the presence of a cell. However, the XRFapproach also revealed drastic changes in cell elementalcomposition during sample preparation including loss ofMg and K and gain of Cl, likely due to cytosol loss. Notemoreover that a 3.95 keV incident beam is below theabsorption edge of Ca, hence inhibiting its analysis.Overall, with the exception of P- and S-containingminerals which include in a lesser extent the presentcase of the basaltic glass, P and S appear to be the best cellindicators on mineral surfaces. Sulfur is an essentialelement present in proteins and various metabolites.Phosphorus itself is essential to all forms of life since it ispart of nucleic acids and phospholipids in cell membrane,although there are many phosphorus compounds that are
essential in the living cell (e.g. ATP transfer). One of themcan at least be used to localize cells, depending on theinvestigated mineral substrate.
The present results demonstrated the successfulvisualization of a specific gold–silver signal on varioussubstrates at the single prokaryotic-cell level, with asatisfactory detection efficiency and spatial resolution. Inparticular, there is no need to perform ultra-thin sectionsas frequently required for cellular gold labeling studiesbut which can be sometimes problematic for samples thatcontain both mineral and biological matter. There areonly a few techniques currently available that permitsuch a noninvasive and nondestructive investigationwith sufficient spatial resolution to examine individualcells both structurally and chemically. Moreover, with anX-ray beam of higher energy and depending on thematrix absorption, the X-ray microprobe allows inves-tigation of structures tens of microns beneath solidsurfaces. This technique would thus permit the study ofmicrobes within pores and rock-fractures.
One final conclusion is that the analytical protocolpresented in this study potentially allows imaging thepresence and nature of living microorganisms in rocksamples. The possibility to simultaneously link thephylogenetic identification of microorganisms withcomplementary methods (i.e. X-ray microdiffraction and
Fig. 7. X-ray micro-fluorescence elemental maps on basalt of (A): culturedD. radiodurans cells deposited, (B): silver-enhanced ultra-small immunogoldparticles in D. radiodurans hybridized with fluorescein-labeled bacterial probe EUB338. The scanning time for each pixel is 2000 and 2500 ms with0.2 0.2 and 0.33 0.33 m2 pixel size for (A) and (B), respectively. The scale bars correspond to 2 m for each panel (a.u. is for arbitrary unit).
190 B. Ménez et al. / Chemical Geology 240 (2007) 182–192

X-ray absorption microspectroscopy) for the chemicaland structural characterization of the associated mineralphases (i.e. substrate and biomineralizations that consti-tute metabolic reactants and metabolic products), offers agreat innovation for studies of microbe-mineral interac-tions in the deep biosphere. A similar challenging study,combining molecular techniques with advanced imagingand spectroscopic methods, was performed byMagnusonet al. (2004) by using X-ray Photoelectron Spectroscopy(XPS) to investigate mineral surface-associated microbialactivities. No doubt that such interdisciplinary trendswould be a valuable outcome in the next future.
A cknowledgments
This work was funded by the Research Program onGeological Storage of CO2 (IPGP/TOTAL/SCHLUM-BERGER) and the “ACI Risques Naturels 2003 —stockage géologique du CO2”. We thank the ID21 stafffor their assistance during the experiment, and J. Cauzid,P. Lopez Garcia, D. Moreira, F. Guyot, I. Martinez andM. Van Zuilen for their help and insightful advices.(IPGP contribution No. 2193).
A ppendix A . Supplementary data
Supplementary data associated with this articlecan be found, in the online version, at doi:10.1016/j.chemgeo.2007.02.009.
References
Amann, R., Glöckner, F.O., Neef, A., 1997. Modern methods insubsurface microbiology: in situ identification of microorganismswith nucleic acid probes. FEMS Microbiol. Rev. 20, 191–200.
Amend, J.P., Teske, A., 2005. Expanding frontiers in deep subsurfacemicrobiology. Palaeogeogr. Palaeoclimatol. Palaeoecol. 219,131–155.
Benzerara, K., Yoon, T.H., Menguy, N., Tyliszczak, T., Brown, J.G.E.,2005a. Nanoscale environments associated with bioweathering of aMg–Fe-pyroxene. Proc. Natl. Acad. Sci. U. S. A. 102 (4), 979–982.
Benzerara, K., Yoon, T.H., Tyliszczak, T., Constantz, B., Spormann,A.M., Brown, J.G.E., 2005b. Scanning transmission X-ray micros-copy study of microbial calcification. Geobiology 2, 249–259.
Bohic, S., Simionovici, A., Snigirev, A., Ortega, R., Devès, G., Heymann,D., et al., 2001. Synchrotron hard x-ray microprobe: fluorescenceimaging of single cells. Appl. Phys. Lett. 78 (22), 3544–3546.
Di Fabrizio, E., Cojoc, D., Cabrini, S., Altissimo, M., Kaulich, B.,Wilhein, T., et al., 2005. Phase and intensity control throughdiffractive optical elements in X-ray microscopy. J. ElectronSpectrosc. Relat. Phenom. 144–147, 957–961.
Foriel, J., Philippot, P., Susini, J., Dumas, P., Somogyi, A., Salomé, M.,et al., 2004. High-resolution imaging of sulfur oxidation states,trace elements, and organic molecules distribution in individualmicrofossils and contemporary microbial filaments. Geochim.Cosmochim. Acta 68 (7), 1561–1569.
Fredrickson, J.K., Onstott, T.C., 2001. Biogeochemical and geologicalsignificance of subsurface microbiology. In: Fredrickson, J.K.,Fletcher, M. (Eds.), Subsurface Microbiology and Biogeochemis-try, New York, pp. 3–37.
Geesey, G.G., Neal, A.L., Suci, P.A., Peyton, B.M., 2002. A review ofspectroscopic methods for characterizing microbial transforma-tions of minerals. J. Microbiol. Methods 51, 125–139.
Gérard, E., Guyot, F., Philippot, P., Lopez-Garcia, P., 2005.Fluorescence in situ hybridisation coupled to ultra small immuno-gold detection to identify prokaryotic cells using transmission andscanning electron microscopy. J. Microbiol. Methods 63, 20–28.
Jamin, N., Dumas, P., Moncuits, J., Fridman, W.H., Teillaud, J.L.,Lawrence Carr, G., et al., 1998. Highly resolved chemical imagingof living cells by using synchrotron infrared microspectrometry.Proc. Natl. Acad. Sci. U. S. A. 95, 4837–4840.
Kaulich, B., Polack, F., Neuhäusler, U., Susini, J., Di Fabrizio, E.,Wilhein, T., 2002. Diffracting aperture based differential phasecontrast for scanning X-ray microscopy. Opt. Express 10 (20),1111–1117.
Kemner, K.M., Kelly, S.D., Lai, B., Maser, J., O'Loughlin, E.J.,Sholto-Douglas, D., et al., 2004. Elemental and redox analysis ofsingle bacterial cells by X-ray microbeam analysis. Science 306,686–687.
Kieft, T.L., Phelps, T.J., 1997. Life in the slow lane. In: Amy, P.S.,Haldeman, D.L. (Eds.), The Microbiology of the TerrestrialSubsurface. CRC Press, Boca Raton, FL, pp. 137–164.
Krumholz, L., 2000. Microbial communities in the deep subsurface.Hydrogeol. J. 8, 4–10.
Larabell, C.A., Le Gros, M.A., 2004. X-ray tomography generates 3-Dreconstructions of the yeast, Saccharomyces cerevisiae, at 60-nmresolution. Mol. Biol. Cell 15 (3), 957–962.
Lemelle, L., Salomé, M., Fialin, M., Simionovici, A., Gillet, P., 2004.In situ identification and X-ray imaging of microorganismsdistribution on the Tatahouine meteorite. Spectrochim. Acta B59, 1703–1710.
Loy, A., Horn, M., Wagner, M., 2003. ProbeBase: an online resourcefor rRNA-targeted oligonucleotide probes. Nucleic Acids Res. 31,514–516.
Magnuson, T.S., Neal, A.L., Geesey, G.G., 2004. Combining in situreverse transcriptase polymerase chain reaction, optical microscopy,and X-ray photoelectron spectroscopy to investigate mineral surface-associated microbial activities. Microb. Ecol. 48 (4), 578–588.
Miao, J., Hodgson, K.O., Ishikawa, T., Larabell, C.A., LeGros, M.A.,Nishino, Y., 2003. Imaging whole Escherichia coli bacteria by usingsingle-particle X-ray diffraction. Proc. Natl. Acad. Sci. U. S. A. 100(1), 110–112.
Ortega, R., Bohic, S., Tucoulou, R., Somogyi, A., Devès, G., 2004.Microchemical element imaging of yeast and human cells usingsynchrotron X-ray microprobe with Kirkpatrick-Baez optics. Anal.Chem. 76 (2), 309–314.
Parkes, R.J., Wellsbury, P., 2004. Deep biospheres. In: Bull, A.T. (Ed.),Microbial Diversity and Bioprospecting. ASM Press, Washington,D. C., pp. 120–129.
Pedersen, K., 2000. Exploration of deep intraterrestrial microbial life:current perspectives. FEMS Microbiol. Lett. 185, 9–16.
Susini, J., Salomé, M., Fayard, B., Ortega, R., Kaulich, B., 2002. Thescanning X-ray microprobe at the ESRF X-ray microscopybeamline. Surf. Rev. Lett. 9 (1), 203–211.
Twining, B., Baines, S.B., Fisher, N.S., Maser, J., Vogt, S., Jacobsen,C., et al., 2003. Quantifying trace elements in individual aquaticprotist cells with a Synchrotron X-ray Fluorescence microprobe.Anal. Chem. 75, 3806–3816.
191B. Ménez et al. / Chemical Geology 240 (2007) 182–192

Ueki, T., Takemoto, K., Fayard, B., Salomé, M., Yamamoto, A.,Kihara, H., et al., 2002. Scanning X-ray microscopy of living andfreeze-dried blood cells in two vanadium-rich Ascidian species,Phallusia mammillata and Ascidia sydneiensis samea. Zool. Sci.19, 27–35.
Vekemans, B., Janssens, K., Vincze, L., Adams, F., Van Espen, P.,1994. Analysis of X-ray spectra by iterative least squares (AXIL):new developments. X-Ray Spectrom. 23 (6), 278–285.
Whitman, W.B., Coleman, D.C., Wiebe, W.J., 1998. Prokaryotes: theunseen majority. Proc. Natl. Acad. Sci. U. S. A. 95 (12), 6578–6583.
Wilhein, T., Kaulich, B., Di Fabrizio, E., Romanato, F., Cabrini, S.,Susini, J., 2001. Differential interference contrast X-ray microscopywith submicron resolution. Appl. Phys. Lett. 78 (14), 2082–2084.
Wilkins, S.W., Gureyev, T.E., Gao, D., Pogany, A., Stevenson, A.W.,1996. Phase-contrast imaging using polychromatic hard X-rays.Nature 384, 335–338.
192 B. Ménez et al. / Chemical Geology 240 (2007) 182–192




















