
Description of four new species of Neolepton Monterosato, 1875 (Mollusca: Bivalvia: Neoleptonidae),...
37
This article was downloaded by: [UMA University of Malaga] On: 22 February 2012, At: 06:50 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Ophelia Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/smar19 Description of four new species of Neolepton Monterosato, 1875 (Mollusca: Bivalvia: Neoleptonidae), with comments on the genus and on its affinity with the Veneracea Carmen Salas a & Serge Gofas b a Departamento de Biologia Animal, Universidad de Málaga, E-29071, Málaga, Spain b Laboratoire de Biologie des Invertébrés marins et Malacologie, Muséum National d'Histoire Naturelle, 55 rue Buffon, F-75005, Paris, France Available online: 20 Jan 2012 To cite this article: Carmen Salas & Serge Gofas (1998): Description of four new species of Neolepton Monterosato, 1875 (Mollusca: Bivalvia: Neoleptonidae), with comments on the genus and on its affinity with the Veneracea, Ophelia, 48:1, 35-70 To link to this article: http://dx.doi.org/10.1080/00785236.1998.10428676 PLEASE SCROLL DOWN FOR ARTICLE Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. The publisher does not give any warranty express or implied or make any representation that the contents will be complete or accurate or up to date. The accuracy of any instructions, formulae, and drug doses should be independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings, demand, or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with or arising out of the use of this material.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Description of four new species of Neolepton Monterosato, 1875 (Mollusca: Bivalvia: Neoleptonidae),...

This article was downloaded by: [UMA University of Malaga]On: 22 February 2012, At: 06:50Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
OpheliaPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/smar19
Description of four new species of NeoleptonMonterosato, 1875 (Mollusca: Bivalvia: Neoleptonidae),with comments on the genus and on its affinity withthe VeneraceaCarmen Salas a & Serge Gofas ba Departamento de Biologia Animal, Universidad de Málaga, E-29071, Málaga, Spainb Laboratoire de Biologie des Invertébrés marins et Malacologie, Muséum National d'HistoireNaturelle, 55 rue Buffon, F-75005, Paris, France
Available online: 20 Jan 2012
To cite this article: Carmen Salas & Serge Gofas (1998): Description of four new species of Neolepton Monterosato, 1875(Mollusca: Bivalvia: Neoleptonidae), with comments on the genus and on its affinity with the Veneracea, Ophelia, 48:1, 35-70
To link to this article: http://dx.doi.org/10.1080/00785236.1998.10428676
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form toanyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae, and drug doses shouldbe independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims,proceedings, demand, or costs or damages whatsoever or howsoever caused arising directly or indirectly inconnection with or arising out of the use of this material.

OPHELIA 48 (1): 35-70 (March 1998)
DESCRIPTION OF FOUR NEW SPECIES OFNEOLEPTONMONTEROSATO, 1875 (MOLLUSCA:BIVALVIA: NEOLEPTONIDAE), WITH COMMENTS
ON THE GENUS AND ON ITS AFFINITYWITHTHE VENERACEA
Carmen Salas} & Serge Gofas2
I Departamento de Biologia Animal, Universidad de Malaga, £-29071 Malaga, Spain2Museum National d'Histoire Naturelle, Laboratoire de Biologie des Invertebres marins et
Malacologie, 55 rue Buffon, F-75005 Paris, France
ABSTRACT
The type species of Neolepton Monterosato, 1875, Lepton sulcatulumJeffreys, 1859, is redescribed using SEM of shells, critical-point dried soft parts, and semi-thin sections. Four new species (N.gl.lanchefrom the Canary Is., N. cancellatum from the Azores, N. benguelensisfrom Angola, N. caledonicum. from New Caledonia) are described; for the latter species is presented data on living animalsand anatomy. NotoleptonFinlay, 1927 and Neodavisia Chavan in Moore, 1969 are confirmed as synonyms of Neolepton, and the shells of their type species are figured. Kellya atlantica Smith, 1890 (withRochefortia milda Bartsch, 1915 as a new synonym) from St Helena and South Africa, and Halodakrasubtrigona (Carpenter, 1857) from California, are newly assigned to Neolepton. Notolepton atlanticumSoot-Ryen, 1960 from Tristan da Cunha is renamed Neoleptonsootryeni n. nov.
The hinges of postlarval Veneridae, Gouldia minima (Montagu, 1803) and Chamelea stsiatula (daCosta, 1778) were found to share essential traits with the hinge of adult Neolepum. The morphologyof the mantle edge in Neolepton. sulwtulum was found to fit the descriptions given for the Verieridae. The two short siphonal apertures are also similar to those found in shallow infaunal venerids(e.g. Gouldia) and similarly have a delicate siphonal membrane inside the exhalant siphon. Theseobservations are given as arguments to consider the Neoleptonidae as paedomorphic Veneracea,instead of Leptonacea or Cyamiacea as stated in the literature. It is further suggested that all Cyamiacea should be reevaluated for their systematic position with respect to Veneracea.
Key words: Neoleptonidae, Veneracea, Cyamiacea, Taxonomy, Paedomorphy.
INTRODUCTION
Neolepton is a genus of minute bivalves found in the algal mat of intertidal orshallow subtidal rocky shores, in various parts of the world. The name was introduced, as a section of Lepton, by Monterosato (1875) for Lepton sulcatulumJeffreys, 1859, L. clarkiaeClark, 1852, L. glabrum Fischer, 1873, and a proposed newspecies L. obliquatum, nomen nudum, validated by Chaster (1897). Crosse(1885) subsequently designated L. sulcatulum as the type species.
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2

36 CARMEN SALAS & SERGE GOFAS
The systematic position of Neolepum has been debated until quite recently.The type species was initially included byJeffreys (1863) in the family KelliidaeForbes & Hanley, 1849. Thiele (1934) introduced the family Neoleptonidae inthe superfamily Cyamiacea Philippi, 1845, to include the genera Neolepton Monterosato, 1875, Pachykellya Bernard, 1898 and Puysegl.tria Powell, 1927. Nevertheless, Tebble (1966) still placed the genus Neolepton in the family LeptonidaeGray, 1847, and this family in the superfamily Erycinacea Deshayes, 1850. Nordsieck (1969) introduced a new subfamily Neoleptoninae in the Leptonidae (ignoring Thiele's family name) and this one in the superfamily Leptonacea Gray,1847. Deroux (1961) described the anatomy of Epilepton clarkiae, consideredEpilepton as a subgenus of Neolepton, and on these bases questioned Thiele'splacement of the Neoleptonidae. Chavan (in Moore, 1969) nevertheless maintained the family Neoleptonidae Thiele, 1934 in the Cyamiacea. The latter classification has gained general acceptance and was used by Bowden & Heppell(1968), Powell (1979) and Sabclli et at. (1990).
The affinity of Neoleptori with the Veneracea has been already suspected byOckelmann (in Bowden & Heppell, 1968). We have addressed this questionthrough the examination of specimens of several species of the genus Neolepton,and comparison with juveniles of Veneridae with particular atten tion to the ontogeny of the hinge, and morphology of the mantle edge. A full taxonomictreatmen t will be given for the type species of Neolepton and of the genera whichwe consider synonyms, for Kellya atlantica Smith, 1890 newly transferred to Neolepton, and for the new species described herein.
This study has been partially supported by grants DGICYf PB 89-0081 (Fauna I) and DGICYf PB92-0415 (Barbate faunal survey) of the Spanish government. The first author acknowledges facilities given by the Laboratoire de Biologie des Invertebres marins et Malacologie (MNHN, Paris)during the preparation of this paper.
We are grateful to the curators of marine collections in the Auckland Museum; Natal Museum;United States National Museum, Washington; and Zoologisk Museum, Oslo, for loan of relevantmaterial. We thank Gustav Paulay, Winston Ponder and Anders Waren for useful comments - although this does not imply agreement with all our views - on an earlier draft of this paper.
We are particularly indebted to our colleague David Macias (University of Malaga) for thr- preparation of semithin sections of Neolepton. The SEM micrographs were taken byJuan:Jose Cuencaand Gregorio Caballero, University of Malaga.
MATERIAL AND METHODS
Most of the material of Neolepton used for this study is housed in Museum National d'Histoire Naturelle (MNHN), Paris, and was collected during the following expeditions:
- Intertidal and shallow collecting in the Canary Islands, by P. Bouchet and S.Cofas (1980, 1981)
- Workshops in the Azores organized by Pro A. Frias Martins (Universidade dos
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2

NEW SPECIES OF NE.DL£PTON 37
Acores): Vila Franca do Campo, Sao Miguel (1988), Flores (1989), Santa Maria (1990)
- The second author's collecting in Angola (1981-1987)- "Montrouzier" workshop in New Caledonia organized by P. Bouchet (1993)- Collecting at the type locality of Neolepton sulcatulum in Guernsey, in Septem-
ber 1994 following an indication by Phorson (1988).
Early growth stages of several species of Veneridae (Gouldia minima (Montagu,1803), Chamelea striatula (Da Costa, 1778), Clausinella fasciata (Da Costa, 1778)and Pitar rudis (Poli, 1795» were collected during a monthly survey during oneyear ofa soft bottom community from Barbate (36°08.3'N 05°53.6', 29 m, province of Cadiz, Spain). Growth series of the same species were obtained alsooff Redondela, S. Spain (Fauna I expedition, 1990, st. 70A, 37°08, 91'N 07°13.50'W. and st. 71A, 37°09.38'N - 07°16.00'W). This material is depositedin Museo Nacional de Ciencias naturales, Madrid (MNCN).
Specimens from Guernsey and from New Caledonia were picked out frombottom material sieved in sea water, relaxed with isotonic magnesium chloridesolution (71 g of MgCI2, 6H 20 /litre), and drawn.
Specimens from the same localities were fixed in 6% formaldehyde solutionin sea water for several days and then preserved in 70% alcohol neutralizedwith sodium tetraborate. Several such specimens were critical-point dried andexamined under SEM.
For histological study, specimens fixed and preserved as above were postfixedin Os04 (1%) for one hour at 4°C, washed, dehydrated and embedded in Araldite 502. Semithin sections of 1 urn were obtained in a Reichert UMO-2 ultramicrotome and stained with toluidine blue.
The terminology of Bernard (1895, 1898) is used in hinge descriptions (seeCox in Moore, 1969: N55-N57), and the teeth numbers are indicated on figures1-2. Our generic description is based on all the reviewed species for the shell;for the soft parts only N. sulcatulum and N. caledonicum sp. n. were available.
TAXONOMY
Genus Neolepton Monterosato, 1875
Lepton (Neoleplon) , new section: Monterosato, 1875: 12. Type species: Lepton sulcatulumJeffreys,1859, subsequent designation by Crosse (l885).
NeodavisiaChavan in Moore, 1969 (new name for Daoisin Cooper & Preston, 1910, non DaoisiaDelGuercio, 1909. Type species: Dauisia cobtn Cooper and Preston, 1910, by monotypy. Synonymizedby Dell (1964) but maintained as valid by Chavan in Moore (1969).
Notolepton Finlay, 1927. Type species: Kellya antipode Filhol, 1880, by original designation. Synonymized by Thiele (1934).
Halodakra Olsson, 1961. Type species: ? Circe subtrigona Carpenter, 1857, by original designation.New synonym.
Diagnosis. - Shell minute, equivalve, slightly inflated, subcircular to suboval,
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2
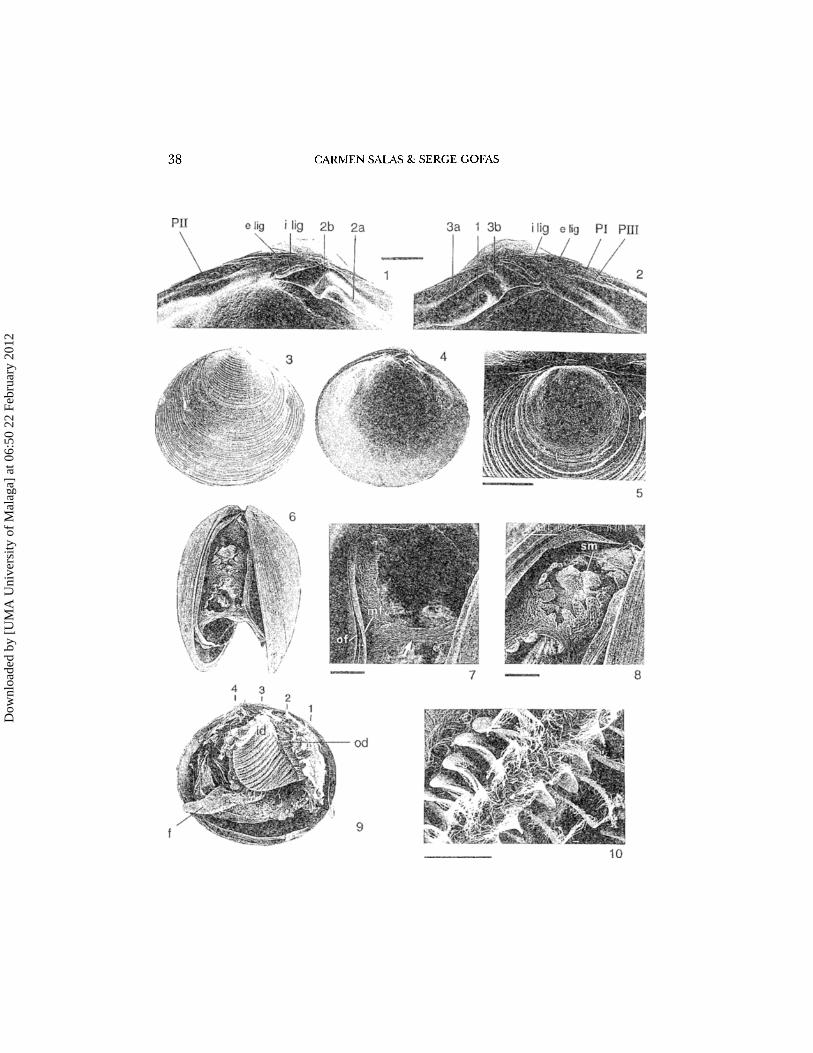
38 CARMEN SALAS & SERGE GOFAS
PH 2b 2a 3a 3b i lig eli!) PI
4 3I I
7
9
8
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2

NEW SPECIES OF NEOUrrrON 39
slightly inequilateral with beaks anterior to the vertical midline, slightly prominent, prosogyrous. Protoconch almost circular, with a smooth or granulatedsurface, surrounded by a clearly demarcated smooth rim along its margin. Teleoconch with concentric ridges (Fig. 3), sometimes also with radial sculpture ormicrosculpture (Fig. 34). Periostracum thin, inconspicuous. Hinge line (Figs1, 2) occupying nearly all the dorsal margin. Left valve with two anterior divergent cardinals (2a, 2b) fused proximally tu form a hook-shaped structure, andan elongate PH, close to the dorsal margin. Right valve with three divergent cardinals (1, 3a, 3b): the 3b joined with the upper end of the elongate 3a to forma hook-shaped structure; the 1 short, stronger and close to the lower margin ofthe hinge plate, well separated from the fused 3a-b by a chevron-shaped 2a'-2b'socket; two posterior laterals (PI and PIlI), the upper one very narrow and closeto the dorsal margin. A well developed internal ligament (Figs 1, 2), below thebeaks and directed obliquely backwards, occupying a small part of the availablespace between cardinals and posterior laterals, A very small external ligament,between the beaks, extending slightly onto the dorsal margin. Interior marginof the shell smooth. Two suboval adductor muscle scars, impressed beneath theanterior and posterior ends of the hinge plate. Pallial line entire or weakly deflected posteriorly, generally well defined. Three openings to the pallial cavity(Figs 6, 11, 58); one anterior-ventral (pedal) and two posterior (siphonal), thelatter bordered by short tentacles. A very thin, transparent siphonal membranein the exhalant siphon, surrounded basally by the tentacles, not always visible asit may be retracted into the proximal part of the siphon. Mantle edge formedby four folds (Figs 7, 16, 17,57): two marginal folds (outer fold and middlefold) with a smaller secondary fold between them, a large inner fold, fused posteriorly to form the siphons, and an innermost fold directed inwards. Foot welldeveloped (Figs 11, 15), relatively long and finger-like, without byssal gland.Ctenidia composed of two reflected demibranchs (Figs 9, 10,56); inner dernibranch occupying nearly the whole length between the two adductors frompractically below the hinge down to near the pallial line; outer demibrachshorter....Figs 1-10. Neolepton stllcalulum]effreys, from Vazon Bay, Guernsey; 1&2, hinge of left and rightvalves (scale bar 100 urn: 2a, 2b: cardinal teeth ofleft valve; PH: posterior lateral tooth of left valve;I, 3a, 3b: cardinal teeth of right valve; PI, PIlI: posterior lateral teeth of right valve; i Iig: inner ligament; e Iig: outer ligament); 3, outside of right valve (length 1.2 mm); 4, inside of left valve(same specimen as Figs 1-2, length 1.25 mm); 5, protoconch of another specimen (scale bar 100prn): 6, postero-ventral view of a critical-point dried animal showing the two posterior siphonalopenings. Length of shell 1.3 mm; 7, detail of the inhalant siphon, to show the fragile fused portion (arrow) of the inner mantle lobe closing it anteriorly. To the left, the outer (of) and middle(mf) lobes of the mantle can be seen, detached from the shell (scale bar 100 pm); 8, detail of theexhalant siphon, to show the siphonal membrane (sm) inside the fringe of tentacles (scale bar 100urn): 9, lateral view of another specimen, with left valve and left mantle lobe removed to show thefoot (f), inner dernibranch (id), outer demibranch (od) and positions (1-4) of sections Figs 12-15
(length of shell 1.2 mm); 10, detail of the inner demibranch (scale bar 10 prn).
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2

40
sm
CARMEN SAL-\S & SERGE GOFAS
ex in poFig. II. Oblique ventral view of living Neolepton sulcatulum Jeffreys from Vazon Bay, Guernsey.Length of shell 1.3 mrn, ex : exhalant siphon, with its siphonal membrane (srn) : ill: in halant si
phon; po : pedal opening; f: fOOL
Neolep uni sulcat.u lu m (Jeffreys, 1859)
(Figsl-J?)
Lepton 5utW1ULulIlJdfn:ys, 1859: 34-35, Fig. 2 a·g.
Type material. - Syntypes, Guernsey. 1 sp ecimen USNM 199456 and many shellsUSNM 199475 (fide Waren, ]980; not seen) . Possible syntypes, received by Petitde la Saussaye from Jeffreys, Guernsey, 5 specimens and 4 valves, MNHN.
Type locality. - Guernsey.
Material examined. - English Channel: Cuernsey, Vazon bay (49°28 ' N, 02.37 'W)in coralline algae at extreme low water mark, 30 specimens collected alive, Salas& Gofas leg. 9.1994. Roscoff, 1 specimen, Rodriguez Babio leg. 1975 . - SWSpain: Tarifa, Torre de la Pefia, 3 specimens and 12 valves, Gofas Icg., 1981. Getares, 3 specimens and 25 valves, Gofas leg ., 1981. Barbate, from low water markand shallow subtidal, 20 specimens, Salas & Gofas leg., 4.1994. - MediterraneanFrance: Nice, 5 valves, Locard collection . Frejus 20-30 m, 3 specirnens.]. Pelorceleg. 1994. - Sicily: Palermo, 4 specimens, Monterosato leg. 1872. Palermo, 12specimens, Petit de la Saussaye collection. - Algeria: Algiers, 6 valves, Locardcollection. All specimens in MNHN.
Redescription. - Shell 1-1.5 mm long, almost circular (length/height ratio 1.10),slightly inequilateral. Posterior end broadly rounded . Protoconch almost circular (maximum diameter 226 to 240 prn) with apical area flattened , smooth withirregular surface; with a very narrow commarginal rim. Sculpture of te1eoconch
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2

NEW SPECIES OF NEOLEPTON 41
with numerous flat concentric ridges, broader than the interspaces, slightlystronger near the margin, sometimes also with an extremely inconspicuous radial microsculpture towards the anterior end. Colour white. Hinge plate narrowing slightly below the beaks. Left valve (Figs 1(3) with cardinal 2b pointingbackwards, joined near its proximal end with a longer 2a to form a hook; anelongate PII, close to the dorsal margin. Right valve (Fig. 2) with 3b narrow,pointing backwards, joined with the upper end of the elongate 3a to form ahook; 3a almost fused with the dorsal margin of the hinge; cardinal 1 ratherelongate. Two posterior laterals, the PI narrow, the PIn longer, very narrowand close to the dorsal margin. Siphons short and relatively broad, hardly extending beyond the edge of the shell (Fig. 11), colourless with a pair of orangespots and smaller orange dots around the exhalant. Mantle edge with the fourfolds characteristic of the genus (Figs 7, 16, 17), and the inside of the edge ofthe pedal aperture with a thick layer of glandular cells (Fig. 17). Outer dernibranch short, less than one-third of the length of the inner one. Gill filamentssloping towards the anterior side. Sexes separate.
Remarks. - The mantle fusion anteriorly demarcating the inhalant siphon (Fig.7, arrow) is fragile: in 26 out of 30 living specimens examined in Guernsey, thisbridge was broken and allowed movement of the foot to reach the posteriorend of the inhalant siphon. The split appears to be healed and the portion ofthe mantle edge with tentacles assumes in life a position to form the inhalant siphon.
Neolepton guanche sp. n.
(Figs 18-27)
Type material. - Holotype (MNHN) and 30 paratypes from the type locality, Bouchet & Cofas leg. 7.1980 (10 MNHN, 10 MNCN, 10 Museo Insular de CienciasNaturales, Tenerife).
Type locality. - Palm-Mar (28°02'N, 16°42'W, intertidal rocks with algal mat),Tenerife, Canary Islands.
Other material examined. - Tenerife: Punta del Hidalgo, 9 specimens, Bouchet &Cofas leg. 7.1980. Las Calletas, 5 specimens and 10 valves, Bouchet & Cofas leg.7.1980. Punta de Teno, intertidal, 3 specimens and 10 valves, Bouchet & Cofasleg. 7.1980. Los Cristianos, 0-1 m, 2 specimens, Bouchet & Gofas leg. 7.1980.Lanzarote: Arrecife, 3 specimens, Bouchet leg. 12.1981. - Selvagens Islands: Selvagen Grande, intertidal, 50 specimens and 100 valves, Segonzac leg. 6.1989.Selvagen Pequena, 16 specimens and 40 valves, Segonzac leg. 6.1989. All specimens in MNHN.
Description. - Shell 1-1.5 mm long, subcircular to suboval (length/height ratio1.10 to 1.16), slightly to moderately inequilateral with posterior part longer.Posterior end broadly rounded; dorsal posterior margin rising higher than the
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2
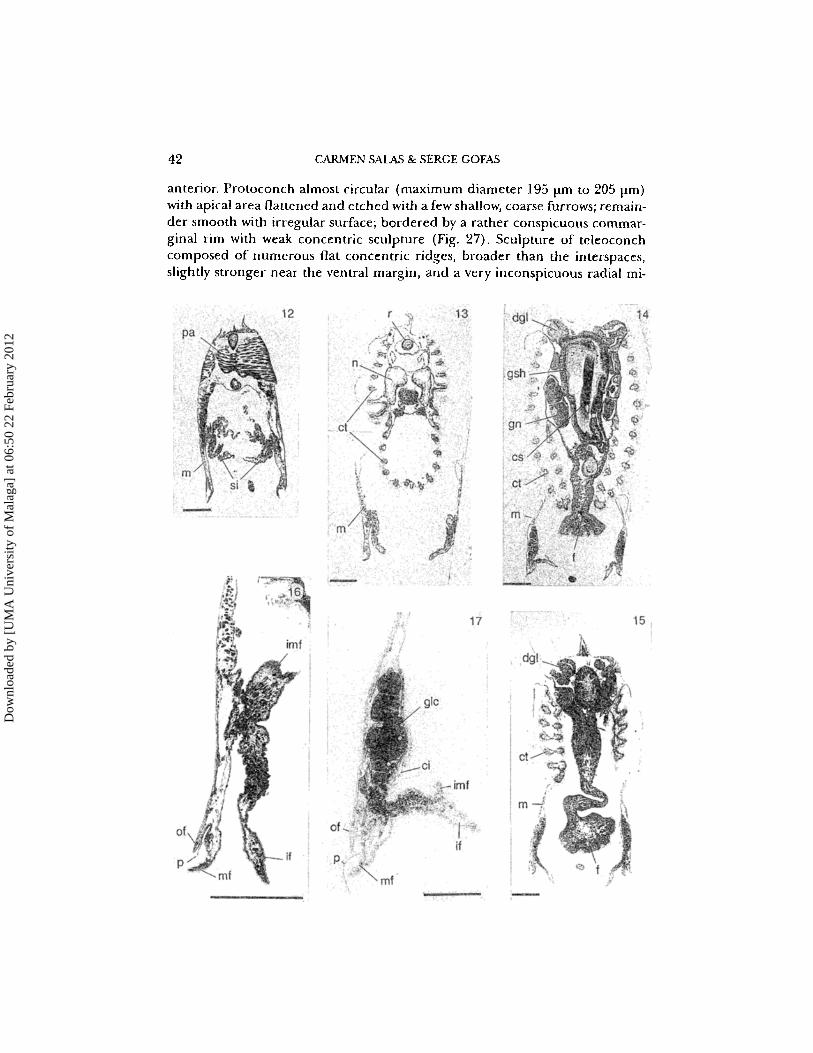
42 CARMEN SAlAS & SERGE GOFAS
anterior. Protoconch almost circular (maximum diameter 195 \lm to 205 prn)with apical area flattened and etched with a few shallow, coarse furrows; remainder smooth with irregular surface ; bordered by a rather conspicuous cornmarginal rim with weak concentric sculpture (Fig. 27). Sculpture of teleoconchcomposed of numerous flat concentric ridges, broader than the interspaces,slightly stronger near the ventral margin, and a very inconspicuous radial rni-
13
,}}
!
17
..' i•
mf
15
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2

NEW SPECIES OF NEOLEPTON 43
crosculpture, not overrunning the concentric ridges. Colour variable, white,pale yellowish, pinkish or brownish, with the central area usually opaque withnumerous and irregular translucent spots. Interior of shell white, with externalcolouration showing through. Hinge plate narrowing below the beaks. Leftvalve (Figs 18, 23) with cardinal 2b short, very strong, vertical or pointing slightly forwards, joined near its proximal end with a longer 2a to form a hook; anelongate PII, close to the dorsal margin. Right valve (Figs 19, 24) with 3b smalland narrow, vertical, joined with the upper end of the elongate 3a to form ahook; cardinal 1 rather elongate. Two posterior laterals, the PI narrow andweakly developed, the PIlI longer, very narrow and close to the dorsal margin.
Distribution. - Canary and Selvagen Islands.
Biotope. - In intertidal and shallow subtidal mats of coralline algae with accumulated particles of sediment.
Etymology. - The species is named after the indigenous people of the CanaryIslands.
Remarks. - This species is very similar to Neoleptoii sulcatulum (Jeffreys), but itsvalves have more prominent beaks and the posterior dorsal margin is moreraised. The shell colour is variable, white, golden or pink and its surface has amottled appearence due to the translucent spots, in contrast with the plainwhite colour of N. sulcatulum. Cardinal tooth 2b of N. guanche is short, nearlyvertical or pointing forwards, while that of N. sulcatulum is longer and pointingbackwards. The posterior lateral PIlI of the right valve is more developed in N.guanche than in N. sulcatulum. The protoconch is smaller (195 to 205 urn instead of 228 to 240 prn). Superficially, this species also resembles N. caledonicumsp. n. with which it shares the polychromatism, but the latter is readily distinguished by the much more developed radial microsculpture.
Jeffreys (1859) mentioned that he examined some minute shells "in Mr.McAndrew's cabinet from Orotava and Lancerote in the Canaries", and identified them as N. sulcatulum, but we suppose that they were N. guanche.
Figs 12-15. Semi-thin sections along vertical transverse planes of Neolepuni suLratulurnJeffreys, arranged from posterior to anterior. See Fig. 9 for the position of planes. cs: crystalline style. ct: ctenidia, dgl: digestive gland. f: foot. gn: female gonad. gsh: gastric shield. m: mantle. n: nephridium.pa: posterior adductor muscle. r: rectum. si: siphons. Scale bars 100 pm; 12, section at the level ofsiphons (plane 1); 13, section showing both inner (below) and outer (above) demibranchs (plane2); 14, section at the level of stomach (plane 3); 15, section at the level of foot (plane 4).Figs 16,17. Semi-thin sections through the mantle edge of Neolepton sulcatulum (nomenclature offolds following Ockelmann, 1964). ci: ciliary tract of the mantle. glc: glandular cells. of: outer fold.if: inner fold. imf: innermost fold. mf: middle fold. p: periostracum. Scale bars 100 urn; 16, sectionof a posterior part of the mantle edge, showing the four mantle folds characteristic of the Veneracea; 17, section of a median part of the mantle edge, with conspicuous glandular cells possibly re-
lated to brooding.
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2
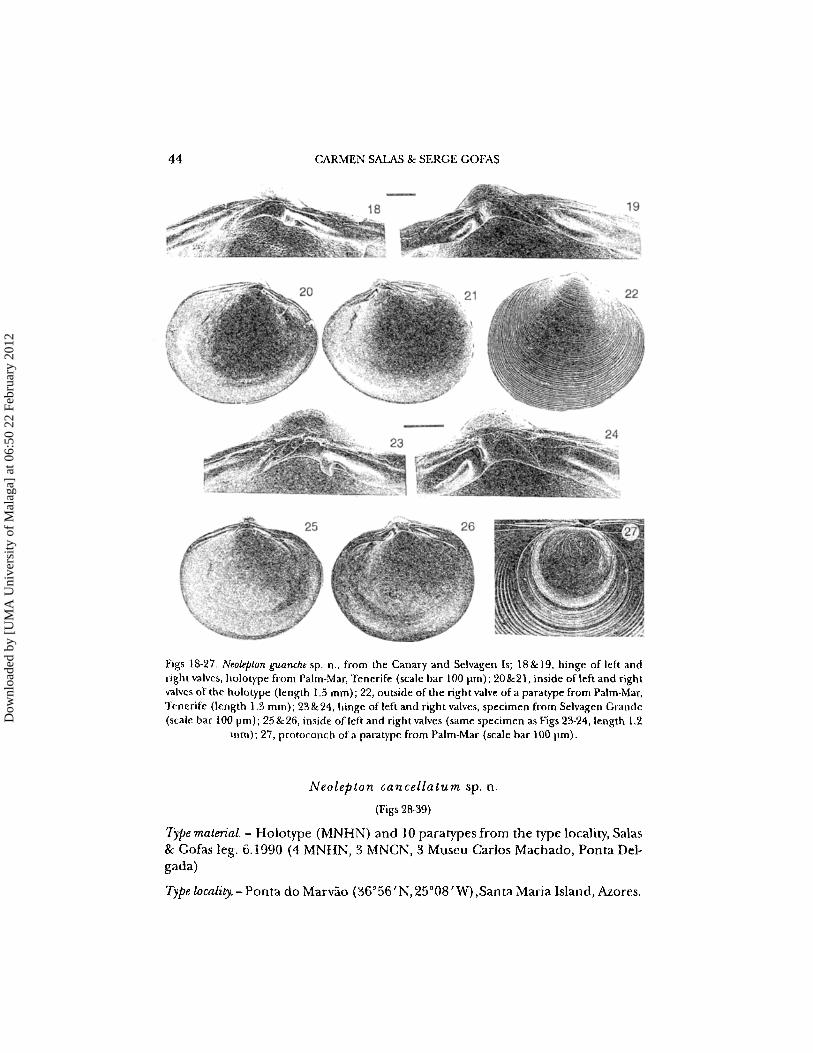
44 CARMEN SAlAS & SERGE GOFAS
Figs 18-27. Neolepton guanche sp. n., from the Canary and Selvagen Is; 18&19, hinge of left andright valves. holotype from Palm-Mar. Tenerife (scale bar 100 urn): 20&21, inside of left and rightvalves of the holotype (length 1.5 rnrn): 22, outside of the right valve of a para type from Palm-Mar.Tenerife (length 1.3 mm); 23&24, hinge ofleft and right valves. specimen from Selvagen Grande(scale bar 100 prn): 25 & 26, inside of/eft and right valves (same specimen asFigs 23·24. length 1.2
mm); 27, protoconch ofa paratype from Palm-Mar (scale bar 100 prn).
Neolepton cancellatum sp. n.
(Figs 28-39)
Type material. - Holotype (MNHN) and 10 paratypes from the type locality, Salas& Cofas leg. 6.1990 (4 MNHN, 3 MNCN, 3 Museu Carlos Machado, Ponta Delgada)
Type locality.- Ponta do Marvao (36°56' N, 25°08 'W),Santa Maria Island, Azores.
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2

NEW SPECIES OF NEOLEPTON 45
Other material examined. - Santa Maria: Vila do Porto, 6-9 rn, 5 specimens and 30valves, Azevedo leg. 6.1990. - Sao Miguel: Ponta da Piramide, 13 m, Azevedoleg. 7.1988, 20 specimens. Ilheu de Vila Franca do Campo, 0-1 m, 3 specimens,Cofas leg. 7.1988. - Flores: Santa Cruz, 20 m, 18 specimens and 6 valves, Azevedo leg. 7.1989.
Description. - Shell 1-1.2 mm long, subcircular (length/height ratio 1.12 to1.14), almost equilateral. Posterior end broadly rounded, anterior end with themaximum curvature closer to the dorsal margin. Protoconch almost circular(maximum diameter 176 to 186 prn) with granulated surface, and apical areaflattened with irregular depressions; bordered by a conspicuous rim with weakconcentric sculpture. Sculpture of teleoconch reticulate, with numerous concentric ridges, overrun by numerous, radial ribs, unequal in size and irregularly spaced, forming small knobs on the intersection points (Fig. 33). Colourwhite. Hinge plate narrowing very much below the beaks. Left valve (Figs 28,35) with cardinal 2b short, very strong, vertical, joined at the proximal third ofits length by a longer 2a to form a hook; a strong, elongate PII close to the dorsal margin. Right valve (Figs 29, 36) with 3b very small and narrow, pointingslightly backwards, joined with the upper end of the 3a to form a hook; the 3aelongate, very narrow and almost fused with the dorsal margin of the hinge;cardinal 1 moderately elongate. Two posterior laterals, the PIlI very narrow,hardly distinct from the dorsal margin.
Distribution. - Azores.
Biotope. - In shallow subtidal rocky bottoms (0-20 m) with algal mat and accumulated particles of sediment.
Etymology. - The species is named after the reticulated sculpture of the shell.
Remarks. - The hinges of N. cancellatum sp. n. and N. benguelensis sp. n. are peculiar by the cardinals of the left valve, fused at some distance along the posteriorcardinal to form a hammer-shaped structure. N. benguelensis is the most closelyrelated species, distinguished only by the much more irregular radial microsculpture, not dearly forming a reticulate pattern with the concentric one. Neolepton cancellatum differs from both N. sulcatulum and N. guanche by its reticulated te1eoconch sculpture and granulated surface of the central area of the protoconch; from N. guanche also by the white colour and the hinge plate muchmore narrowing below the beaks.
Neolepton benguelensis sp. n.
(Figs 40-47)
Type material. - Holotype (MNHN) and 2 paratypes from the type locality, Cofasleg. 9.1984 (MNHN). 4 paratypes from Caotinha, province of Benguela, Cofas
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2

46 CARtVlEN SALAS & SERGE GOFAS
Figs 28-39. Neolepton cancel/alum sp. n., from the Azores; 28&29, hinge of left and right valves, holotype from Ponta do Marvao, Santa Maria (scale bar 100 fun); 30&31, inside of left and rightvalves of the holotype (length 1.2 mm); 32, outside of the left valve of a paratype from Ponta doMarvao (length 1.1 mm); 33, protoconch and early part of teleoconch (same specimen as fig. 32;scale bar 100 11m);34, outside of the left valve of a specimen from Ilheu de Vilafranca, Sao Miguel(length 1.1 mm); 35 & 36, hinge of left and right valves, specimen from Santa Cruz de Flores (scalebar 100 pm): 37 &38, inside ofleft and right valves (same specimen as Figs 35-36, length 1.0 mm);
39, outside of a right valve from Santa Cruz de Flores (length 1.0 mm).
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2

NEW SPECIES OF NEOLEPTON 47
leg. 12.1985 (2 MNCN, 2 Universidade Agostinho Neto, Luanda). 2 paratypesfrom Praia Amelia, Gofas leg. (MNHN).
Type locality. - Chapeu Armado (14"27'S, 12"21 'E), province of Namibe, Angola, in algal mat on rocks at low tide.
Description. -Shell 1-1.3 mm long, subcircular (length/height ratio 1.13), slightly in equilateral. Posterior end broadly rounded; dorsal posterior margin risinga little higher than the anterior. Protoconch almost circular (maximum diameter 190 to 208 urn) with granulated surface, apical area flattened with irregulardepressions; bordered by a conspicuous rim with weak concentric sculpture(Fig. 47). Sculpture of teleoconch (Fig. 42) reticulated, with numerous concentric ridges, crossed by very numerous, discontinuous, unequal radial ribs, overrunning the concentric ribs towards the anterior and posterior ends of theshell. Colour white. Hinge plate narrowing below the beaks. Left valve (Fig. 43)with cardinal 2b short, very strong and thick, vertical or pointing slightly forward, joined at almost mid-length with a longer, curved 2a to form a hook; astrong, elongate PIl, close to the dorsal margin. Right valve (Fig. 44) with 3bvery small and narrow, pointing backwards, joined with the upper end of the 3ato form a hook; the 3a elongate, very narrow and almost fused with the dorsalmargin of the hinge; cardinal I moderately elongate. Two posterior laterals, thePI strong, the PIlI very narrow, hardly distinct from the dorsal margin.
Distribution. - Southern Angola.
Biotope. - In shallow subtidal algae (0-5 rn) with accumulated particles of sediment, on rocky shores.
Etymology. - The species is named after the province of Benguela.
Remarks. - This species most resembles N. cancellatum, with which it shares thewhite shell, granulated protoconch and reticulated teleoconch. It is slightlymore inequilateral, has the radial threads finer, more irregular and discontinuous, and not overrunning the concentric lines on the central surface of theshell. The protoconch is approximately 10% larger. N. benguelensis also resembles N. caledonicum by the microsculpture, but is much smaller, colourless andits hinge is different: the central anterior cardinal of the right valve is elongateinstead of triangular, and the fused cardinals of the left valve are hammershaped like in N. cancellatum.
Neolepton caledonicum sp. n.
(Figs 48-58)
Type material. - Holotype (MNHN) and 40 paratypes from the type locality,Montrouzier expedition, 9.1993 (10 MNHN, 10 Auckland Museum, 10 National Museum of New Zealand, 10 Australian Museum).
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2
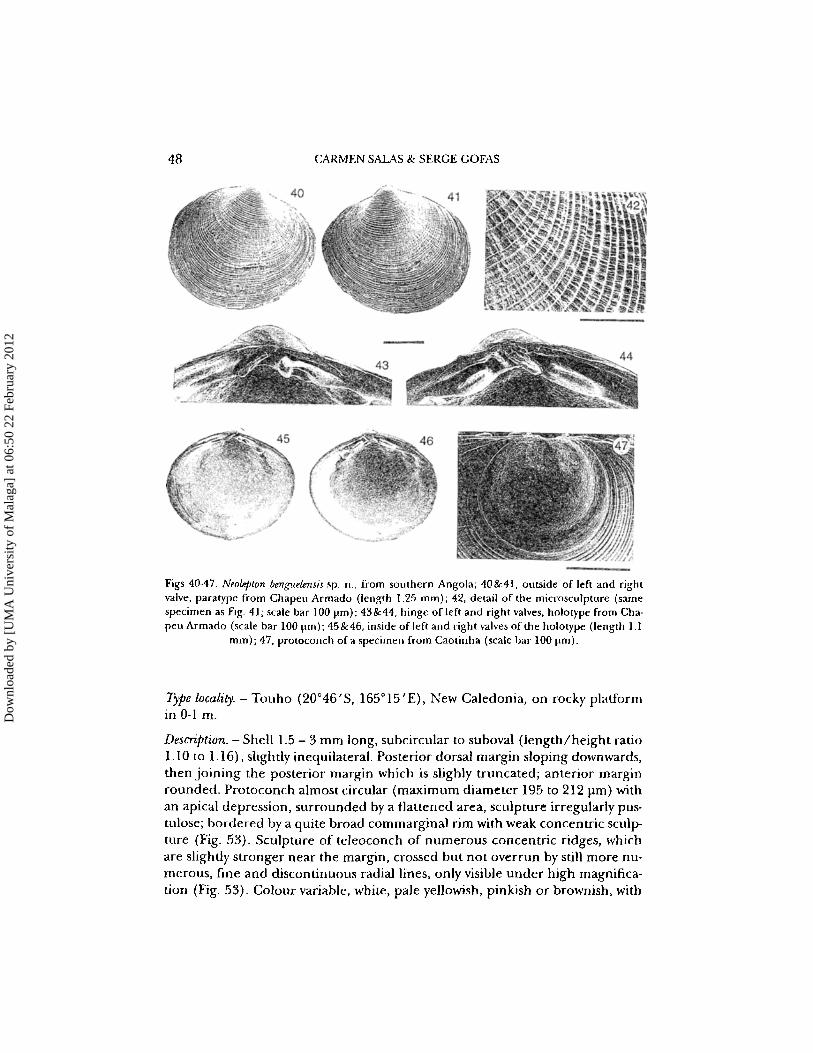
48 CARMEN SAlAS & SERGE GOFAS
~ .- -Figs 40-47. Neolepton benguelensis sp. n., from southern Angola; 40&41, outside of left and rightvalve, paratype from Chapeu Armado (length 1.2:> mm); 42, detail of the microsculpture (samespecimen as Fig. 41; scale bar) 00 prn): 43 &44, hinge of left and right valves, holotype from Chapeu Armado (scale bar 100 urn}; 45&46, inside of left and right valves of the holotype (length l.l
mm): 47, protoconch of a specimen from Caotinha (scale bar 100 prn).
Type locality. - Touho (20°46'5, 165°15'E), New Caledonia, on rocky platformin 0-1 m.
Description. - Shell 1.5 - 3 mm long, subcireular to suboval (length/height ratio1.10 to 1.16), slightly inequilateral, Posterior dorsal margin sloping downwards,then joining the posterior margin which is slighly truncated; anterior marginrounded. Protoconch almost circular (maximum diameter 195 to 212 prn) withan apical depression, surrounded by a flattened area, sculpture irregularly pustulose; bordered by a quite broad com marginal rim with weak concentric sculpture (Fig. 53). Sculpture of teleoconch of numerous concentric ridges, whichare slightly stronger near the margin, crossed but not overrun by still more numerous, fine and discontinuous radial lines, only visible under high magnification (Fig. 53). Colour variable, white, pale yellowish, pinkish or brownish, with
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2

NEW SPECIES OF NEOLEPTON 49
numerous and irregular translucent spots, and brown mottles, in the centralarea. Hinge plate relatively broad and thick. Left valve (Fig. 48) with cardinal2bshorter, pointing slightly backwards, touching at its proximal end with a longer2a; an elongate PH, situated posteriorly and close to the dorsal margin. Rightvalve (Fig. 49) with 3b very small and narrow, vertical or pointing slightly backwards, not reaching the lower margin of the hinge plate, joined with the upperend of the elongate 3a to form a hook; the 3a very narrow and hardly distinctfrom the dorsal margin; cardinalI triangular, very strong. Two posterior laterals, the PIlI very narrow, close to the dorsal margin and situated posteriorly tothe more prominent PI. Siphons short, occupying nearly half of the anteriormargin, bordered each with ca. 20 small tentacles, sprinkled with yellow andwith purple patches inside. Siphonal membrane extending far outside the exhalantsiphon, reaching more than 1/3 of the shell length (Fig. 58). Remainderof the mantle edge smooth, colourless. Foot rather small and narrow. Gills (Fig.56) with outer demibranch shorter, less than half of the inner one, and filaments nearly horizontal.
Distribution. - New Caledonia.
Biotope. - In intertidal and shallow subtidal algal mat with accumulated particlesof sediment.
Etymology. - The species is named after the island of New Caledonia.
Remarks. - Neolepton caledonicum differs from the New Zealand species N. antipodum (Filhol, 1880) and N. subobliquum (Powell, 1937) in having a radial microsculpture, and by the hinge which has a strong, more triangular cardinal in theright valve. A peculiar feature is that the hinge plate does not narrow below thebeaks.
The microsculpture of N. caledonicum recalls that of N. benguelensis by the irregular, discontinuous radial lines, but the latter is smaller, colourless and has aquite different hinge. The colour pattern is strikingly similar to that of N.guanche but the latter has a smooth protoconch and lacks a conspicuous radialmicrosculpture.
Neolepton atlanticum (Smith, 1890). New combination
(Figs 59-68)
!<ellya atlantimSmilh, 1890: 313, PI. 24 Fig. 24.Rochefortia milda Bartsch, 1915: 200-201, PI. 47 Fig. 6, PI. 52 Fig. 9-10. New synonym.
Type localities. - Kellya atlantica, St. Helena island, on drifted kelp; Rochefortia milda, Port Alfred, South Africa.
Material examined. - St. Helena Is.: 1 specimen,Jousseaume collection (MNHN).- South Africa (mostly Natal Museum (NM), Pietermaritzburg): Cape Town,
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2
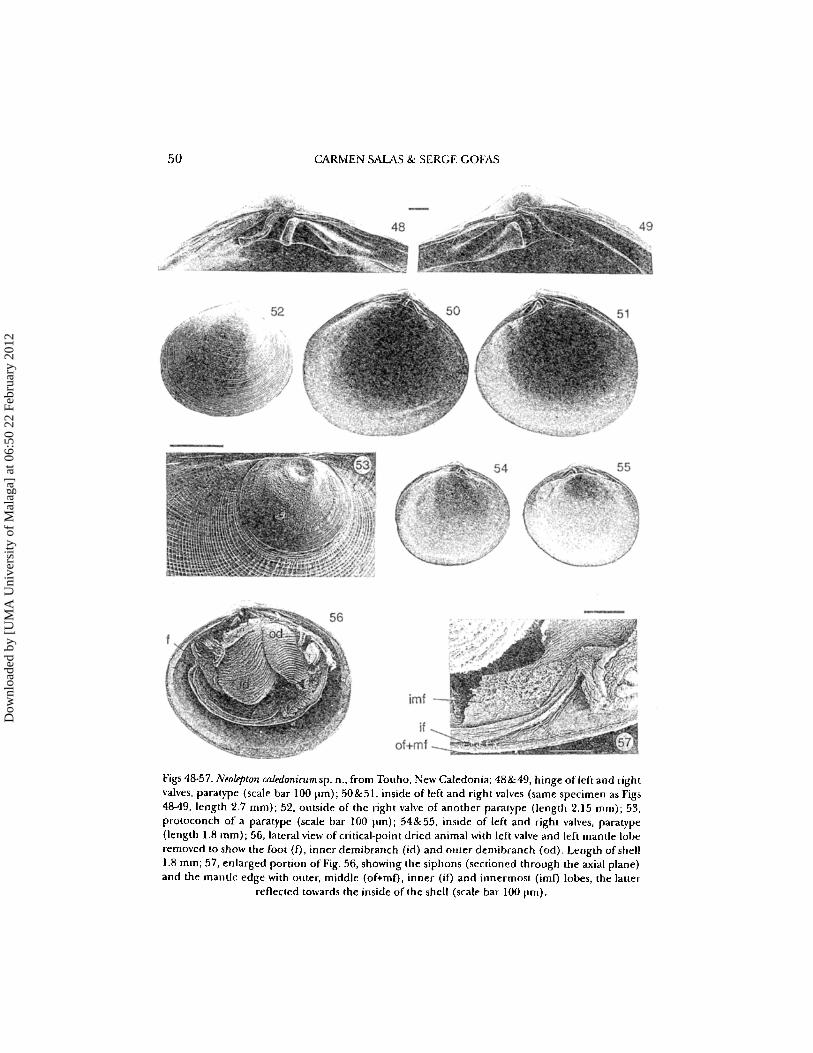
50 CARNlEN SALAS & SERG E GOFAS
Figs 18-57. Neolepton caledonicu m sn . n., from Touho, New Caledonia; 48&19, h inge often and rightvalves, paratype (scale bar 100 prn): 50&51. inside of left and right valves (same specimen as Figs48..49, length 2.7 mm); 52, outside of the right valve of another paratype (length 2.15 mm); 53,protoconch of a paratype (scale bar 100 11m); 54&55, inside of left and right valves, paratype(length 1.8 mrn) ; 56, lateral view of critical-point dried animal with left valve and left mantle loberemoved to show the foot ( f) , inner demibranch (id) and outer dernibranch (od) . Length of shell1.8 mm ; 57 , enlarged portion of Fig. 56, showing the siphons (sectioned through the axial plane)and the mantle edge with outer, middle (of+mf) , inner (if) and innermost (imf) lobes, the lauer
reflected towards the inside of the shell (scale bar 100 11m).
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2

NEW SPECIES OF NEOLEPTON 51
Sea Point, 1 valve, R. Kilburn leg. 1966 (NM, cat. A 6487). False Bay, off Macassar Beach, dredged ca. 18 m: 1 valve, C.M. Connolly leg. 1.1974 (NM, cat. A3115). False Bay, Gordons Bay, beach drift, 2 shells, C.M. Connolly leg. (NM,cat. B 6874). St. Sebastian bay, Breede river mouth, beach drift, 5 valves, C.D.Quickelberge leg. 9.1982 (NM, cat. E 2383). Port Alfred, 1 shell and 1 valve, figured syntypes of RocheJortia milda Bartsch, W.H. Turton leg., (USNM, cat. 249894). Port Alfred, 1 specimen and 5 valves, W.H. Turton leg. 1921 (MNHN).Port Alfred, 7 valves, W.H. Turton leg. (NM, cat. B 7620). East London,Hickman's river mouth, 1 shell, R. Kilburn leg. 1969 (NM, cat. A 1494). PortShepstone, 1 shell, Burnup leg. (NM, cat. A 6413). Natal South Coast, Aliwalshoal, Cracker reef, 23 m, 12 specs. + 1 shell, D. Herbert leg. 4.1989 (NM, cat. E7200, E 7204). Aliwal shoal, 9-15 m, 3 specs., D. Herbert & R. Emanuel leg.11.1988 (NM cat. E 6123). Durban, dredged sand 18-20 m, 1 valve, R. Kilburn &D. Herbert leg. 8.1974 (NM, cat. B 8990). Zululand, Salwana Bay, Two MileReef, stone wash 10-13 m, 8 specs + 1 v., D. Herbert leg. 10.1986 (NM, cat. D5392). Salwana Bay, Two Mile Reef, stone wash 15-17 m, 6 specs., D. Herbert &R. Emanuel leg. 10.1986 (NM, cat. D 5327).
Redescription. - Shell 1.8-3.0 mm long, suboval (length/height ratio 1.20 to1.21), slightly to definitely inequilateral, rather flat for the genus. Anterior endwith maximum curvature at mid-height; posterior dorsal margin rising higherthan the anterior, then joining the posterior margin which may be slightly truncated. Protoconch almost circular (maximum diameter 198 to 216 prn) with apical area flattened and smooth with irregular surface; with a flattened, smoothcommarginal rim (Fig. 64). Sculpture of teleoconch with numerous flat concentric ridges, broader than the interspaces, slightly stronger near the margin,and a radial microsculpture, sometimes very inconspicuous, on the posteriorpart of the shell, not overrunning the concentric ridges. Colour white. Hingeplate narrowing below the beaks. Left valve (Figs 59, 67) with cardinal 2b short,small, pointing backwards, joined at its proximal end with a very elongate 2a toform a hook; a narrow, elongate PII, close to the dorsal margin. Right valve(Figs 60, 68) with 3b small and narrow, pointing backwards, joined with theupper end of the very elongate 3a to form a hook; cardinal I rather elongate.Two posterior laterals, the PI narrow and quite strong, the PIlI indistinct and almost fused to the dorsal margin.
Remarks. - This species was described from drifting algae around St Helena island. The chevron-shaped tooth on the left valve and other features clearly allow identification as a Neolepton species. Specimens from St Helena (Figs 59-64)and from South Africa (Figs 65-68) are hardly separable from shell characters.Smith (1890) pointed out that his specimens were collected on drifting algae ofSouth African origin; the transport of small invertebrates on this kind of substrate has been reported on by Arnaud et al. (1976).
We could ascertain from the type material that the South African RocheJortia
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2

52 CARMEN SALAS & SERGE GOFAS
Fig. 58. Oblique ventral view ofliving Neoleptoa calfdonicum sp. n., holotype, from Touho, New Caledonia. Length of shell 1.9 mm. ex: exhalant siphon, with its long siphonal membrane (sm); in: in
halant siphon; po: pedal opening; f: fool.
milda Bartsch, 1915 is a Neolepton. Bartsch's (1915) figures 9 and 10 of the hingeare confusing because they do not show the characteristic hook of a Neolepton,but look very Mysella-like. This can be explained because cardinals 2b and 3bare very inconspicuous, but these cardinals do exist in the type specimens. Thetransfer of RocheJortia milda to Neolepton has no bearing on the systematic position of RocheJortia Velain, 1877 (type species R. australis Velain, 1877 by rnonotypy) in the Montacutidae. N. milda is the oldest name for a Neolepton with a SouthAfrican type locality, but we cannot see convincing differences which could leadto reject the stated synonymy with N. atlanticum.
There is much variation in the contour of the shell in South African populations, most specimens (e.g. Bartsch's figured syntype) being more rounded inoutline, others being more elongate than those figured herein. There is noc1earcut separation of two morphs in sympatric populations, and thus no indication of presence of more than one species in South Africa.
Neolepton sootryeni nom. n.
Notolepton atlanticum Soot-Ryen, 1960: 22-27, PI. 3 Figs 26-29.
Type locality. - Julia Point, Tristan da Cunha Island, southern Atlantic Ocean;intertidal.
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2

NEW SPECJES OF NEOLEPTON 53
Material examined. - Tristan da Cunha,]ulia Point (intertidal), Norwegian Scientific Expedition st. 74: 16 paratypes in alcohol (Zoologisk Museum, Oslo,D32308).
Remarks. - Soot-Ryen (1960) described Notolepton atlanticum from Tristan daCunha. The shells of the type specimens examined have been dissolved by acidic alcohol; consequently nothing could be added to the original description.We agree with Thiele (1934) that Notolepton is a synonym of Neolepton; thereforethe inclusion of both this species and Kellya atlantica Smith, 1890 in Neoleptonmakes a new name necessary. The species was tentatively placed by Soot-Ryen inthe family Neoleptonidae but he doubted whether the genus (and the wholefamily) may be better included in the Leptonacea.
Soot-Ryen (1960) gave a detailed anatomical account on his new species. Hestates that there is only one antero-ventral opening for the foot and one posterior opening. This situation seems similar to that we found in most of the livingN. sulcatulum examined; Soot-Ryen noted a row of 4-6 tentacle-like folds beforethe fused part of the free margin of the in ner folds, which would correspond tothe position of the inhalant siphon.
The large inner demibranch and the narrow posterior outer demibranch ofthe gills are in good agreement with our observation in N. sulcatulum. The inner fold of the mantle, reflected inwards as in N. sulcatulum, is clearly depictedon Soot-Ryen's figure 6. Eggs and larvae were found to be retained in the mantle cavity, glued to the inner mantle fold "apparently by a secrete from cells inthis fold". In one specimen, larvae were found brooded and measured about200 urn.
Neolepton antipodum (Filhol, 1880)
(Figs 69-73)
Kellia antipodum Filhol, 1880: 1095.Kellia citrina Hutton, 1883: 477.Kellia sanguinea Hutton, 1883: 478.
Type localities. - Kellia antipodum: Campbell island (52°30'S, 169°E), PacificOcean; Kellia citrina: Auckland, New Zealand; Kellia sanguineo: Foveaux Straits,New Zealand.
Type material. - Kellia antipodum: stated to be in the MNHN by Suter (1913) butlost. Kellia citnna and K. sanguinea: lectotypes (Ponder, 1969) in New ZealandNational Museum, Wellington.
Material examined. - New Zealand: Chatham Is., Waitangi, Powell leg. 1935,Staadt collection, 2 specimens and 12 valves (MNHN). Foveaux Straits, Powellleg. 1932, Staadt collection, 7 valves (MNHN). Auckland, Waiheke Is., A.G. Stevenson leg. 1935, Staadt collection, 50 specimens (MNHN). Auckland, Tiri TiriIs., Powell leg. 1939, Staadt collection, 7 specimens (MNHN). Otago, Dunedin
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2
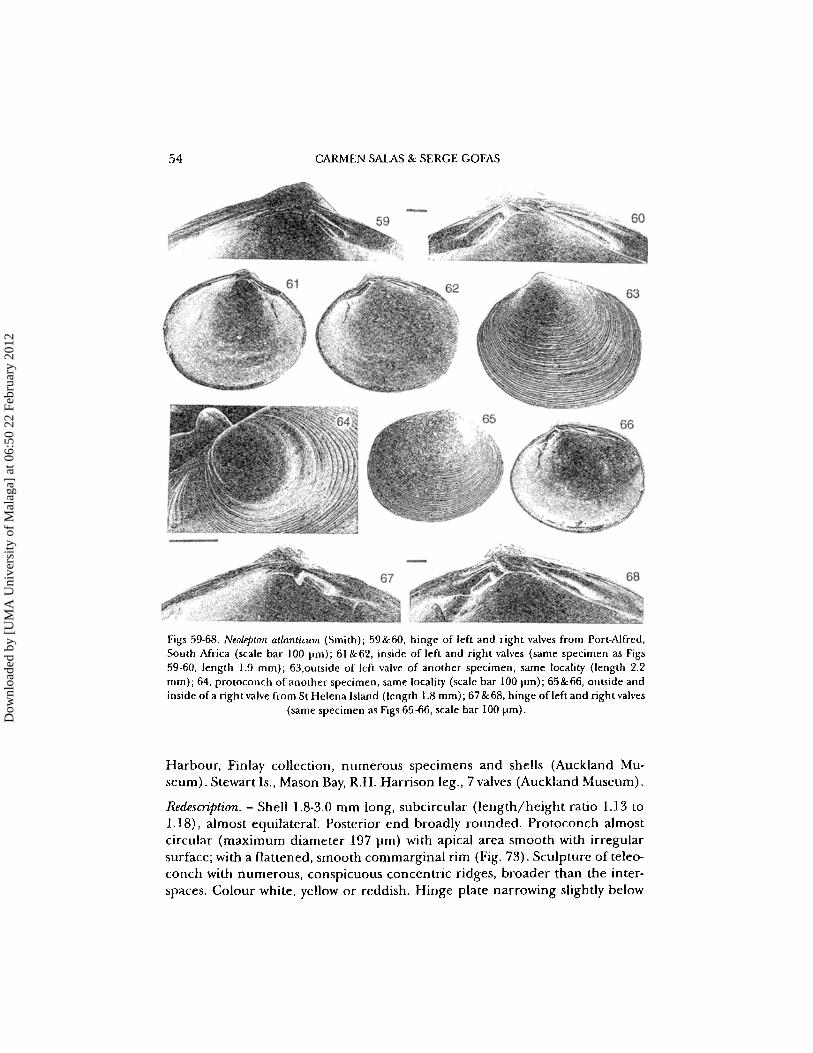
54 CARMEN SAlAS & SERGE GOFAS
Figs 59-68. Neolepton atlanticum (Smith); 59&60, hinge of left and right valves from Port-Alfred,South Africa (scale bar 100 prn): 61 &62, inside of left and right valves (same specimen as Figs59·60, length 1.9 mm); 63,outside of left valve of another specimen, same locality (length 2.2mm); 64, protoconch of another specimen, same locality (scale bar 100 pm); 65&66, outside andinside of a right valve from St Helena Island (length 1.8 mm); 67 &68, hinge of left and right valves
(same specimen as Figs 65-66, scale bar 100 pm).
Harbour, Finlay collection, numerous specimens and shells (Auckland Museum). Stewart Is., Mason Bay, R.H. Harrison leg., 7 valves (Auckland Museum).
Redescription. - Shell 1.8-3.0 mm long, subcircular (length/height ratio 1.13 to1.18), almost equilateral. Posterior end broadly rounded. Protoconch almostcircular (maximum diameter 197 11m) with apical area smooth with irregularsurface; with a flattened, smooth commarginal rim (Fig. 73). Sculpture of teleoconch with numerous, conspicuous concentric ridges, broader than the interspaces. Colour white, yellow or reddish. Hinge plate narrowing slightly below
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2

NEW SPECIES OF NEOLEPTON 55
the beaks. Left valve (Fig. 69) with cardinal 2b pointing backwards, joined at itsproximal end with a longer 2a to form a hook; an elongate PII, close to the dorsal margin. Right valve (Figs 70-72) with 3b very small and narrow, pointingbackwards, joined with the upper end of the very elongate 3a to form a hook;cardinal J rather elongate, strong. Two posterior laterals, the PI narrow andquite strong, the PIII very indistinct and almost fused to the dorsal margin.
Remarks. - This is the type species of Notolepton Finlay, 1927. It is a commonintertidal species in New Zealand (Powell 1979) , and Ponder (1969) gave an anatumical account of the species. N. antipodum has two short posterior siphons(the exhalant with a siphonal membrane), and gills with two demibranchs, theouter one very short. The mantle is lined with mucous glands inside the pedalaperture and, although this is not formally mentioned, the four folds of theedge can be seen on Ponder's (1969) figures 3E and 3F. The anatomy is similarto that of N. sulcatulum and suppo'rts further the synonymy of Notolepton withNeolepton.
We are reluctant to designate a neotype for N. antipodum without having access to material from the type locality. Neolepton antipodum, N. subobliquum (Powell, 1937), N. sublaevigatum (Powell, 1937), N. triangulate (Dell, 1956) and N.powelli Dell, 1964 are recognized as the valid species of Neoleptori for New Zealand waters by Powell (1979). We have examined paratypes of N. subobliquumand N. sublaeoigatusn from the Auckland Museum, and agree that there are atleast three different species. The two latter were described from valves dredgedin deeper water (Discovery II station 933, off Three King Islands, 260 m) butare likely to have been transported downslope. It is beyond the scope of thispaper to revise the New Zealand species of Neolepton, a purpose for which theexamination of living material would be most useful.
Bernard (1897) noted as anterior laterals (on Neolepton sanguineum fromStewart Island, southern New Zealand) the teeth that we have interpreted ascardinals. However, he states that the hinge "is not essentially different fromthat of Lutetina " and interpreted the same teeth as cardinals in the latter (Bernard 1898: 79).
Neolepton cobbi (Cooper & Preston, 1910). New combination
(Figs 74-77)
Daoisia cobbi Cooper & Preston, 1910: 113-114, PI. 4 Figs 9-10.Dauisia benneuiPreston, 1912: 639, PI. 21 Fig. 7.
Type locality. - Falkland Islands.
Material examined. - Falkland Is., 12 specs. coil. Staadt ex Cooper (MNHN). PortStanley, Falkland Is., syntypes of Davisia bennetti Preston, 1912 (MNHN).
Redescription. - Shell 2-3 mm long, equivalve, subcircular (length/height ratio
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2

56 CARMEN SALAS & SERGE GOFAS
Figs 69-73. Neolepton antipodum (Filhol) from Chatham Is., New Zealand; 69 & 70, hinge of left andright valves (scale bar 100 prn): 71, outside of right valve (length 1.8 mm) ; 72, inside of right valve(same specimen as Figs 69-70, length 1.8 mm) ; 73, protoconch and cady part of teleoconch of an-
other specimen (scale bar 100 urn) .
Figs 74-77. Neolepton cobbi (Cooper & Preston) from Falkland Is.; 74&75, hinge of left and rightvalves (scale bar 100 IJm); 76, outside of right valve of another specimen (length 2.1 mm); 77, in
side ofleft valve (same specimen as Figs 74-75, length 2.1 mm) .
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2

NEW SPECIES OF NEOLEPTON 57
1.06), slightly inequilateral with anterior part longer. Protoconch (worn on allexamined specimens). Sculpture of teleoconch of irregular growth lines, grading to irregular concentric striae towards the margins; periostracum rather conspicuous. Colour opaque white to yellowish. Hinge plate: left valve (Fig. 74)with cardinal 2b shorter, pointing slightly backwards, touching at its proximalend with a longer 2a; an elongate PH, situated posteriorly and close to the dorsal margin. Right valve (Fig. 75) with 3b very small and narrow, vertical or pointing slightly backwards, joined with the upper end of the elongate 3a to form ahook; the 3a very narrow and hardly distinct from the dorsal margin; cardinal Ielongate, very strong. Two posterior laterals, the PI quite strong.
Remarks. - This is the type species of Daoisia Cooper & Preston, 1910 (preoccupied, renamed Neodainsia by Chavan in Moore, 1969). It differs from all otherspecies discussed herein by having the beaks posterior rather than anterior tothe vertical midline, and by its more apparent yellowish periostracum. Theseare held as specific characters and we can see no reason, on the available evidence, to maintain Neodatnsia separate from Neolepton. Several of the larger examined specimens have a distorted margin, reflecting some obstacle to theirgrowth in the substrate in which they were trapped.
Preston (1912) later described two more species in Dauisia, collected in fishstomachs in the Falkland Islands. We can see no difference between the syntypes of D. bennetti and D. cobbi, except a slightly larger size.
The figured syntype of D. concentrica Preston, 1912 is also in the MNHN andseems to represent a distinct, valid species of Neolepton. It differs from D. cobbi bya marked concentric sculpture of the teleoconch and a flatter profile of thevalves.
Neolepton subtrigo n.u m (Carpenter, 1857). New combination
? Circe subtngona Carpenter, 1857: 82.
This is the type species of Halodakra Olsson, 1961, by original designation. Thedescription and excellent figures of the lectotype (BM(NH) 1857.6.4.413) andof another specimen given by Coan (1984) indicate that it belongs to Neolepton,and we thus consider Halodakra as a subjective synonym of Neolepton. This conclusion does not extend to the subgenus Stohleria Coan, 1984, which may beused with full generic status until the relationships of its type species (originaldesignation) Psephis salmonea Carpenter, 1864 are clarified.
ONTOGENESIS OF THE HINGES OF SOME VENERIDAE
We had noticed the resemblance of the hinge of Neolepton with early ontogenetic stages of the hinge ofVeneridae, and for this reason we have examined postlarval and juvenile shells of several venerid species, to be used for our discussion of the relationships of Neolepton.
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2
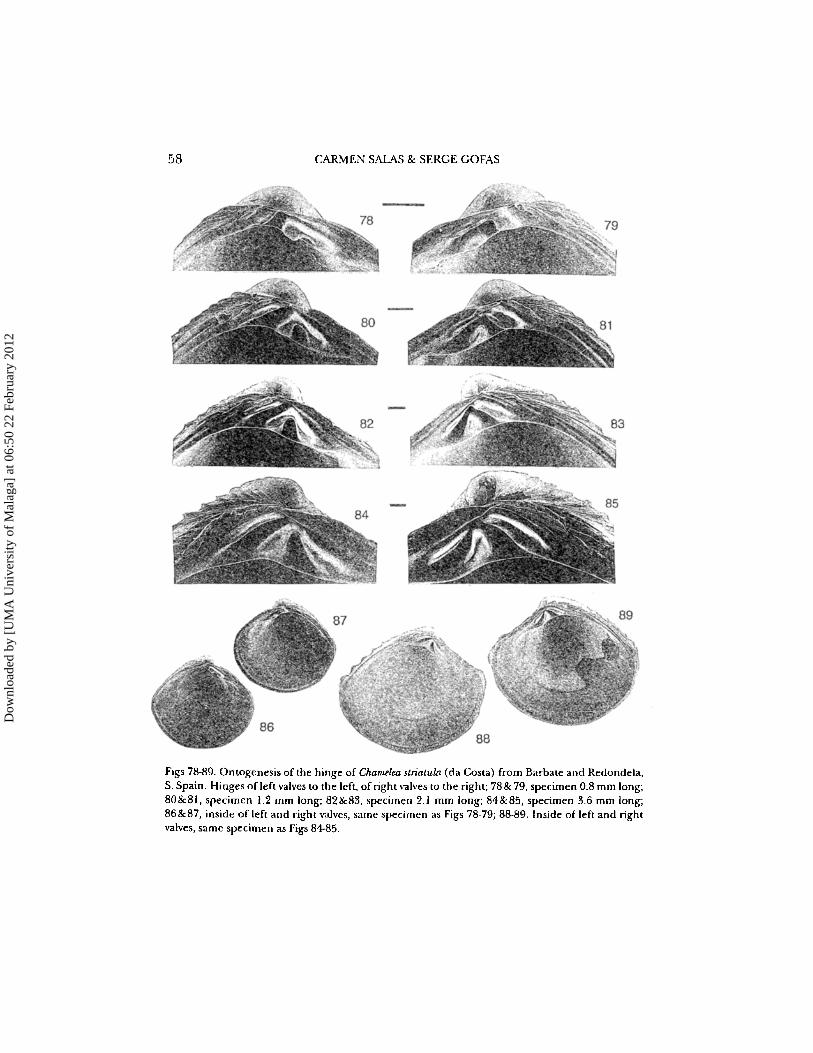
58 CARMEN SALA S & SERGE GOFAS
Figs 78-89. Ontogenesis of the hinge of Chamelea stn atula (da Costa) from Barbato and Red ondela,S. Spain. Hinges ofleft valves to the left, ofright valves to the right; 78&79, specimen 0.8 mm long;80 &81, specime n 1.2 mm long; 82&83, specimen 2.1 mm long; 84& 85, specimen 3.6 mm long;86&87, inside of left and right valves, same spec ime n as Figs 78·79; 88-89. Ins ide of left and rightvalves, same spe cim en as Figs 84-85.
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2

NEW SPECIES OF NEOLEPTON 59
The larval and very early postlarval (less than 1 rnrn in shell diameter) hinges of \tenus uerrucosa, Clausinella [asciata and Chamelea striatula have been described by Le Pennec (1973, 1978, 1980); the primitive denticles were found tohave disappeared in shells larger than 400-450 pm, and the hinge to approximate its final form at sizes of2 - 2.5 mm. The hinges ofjuveniles offour speciesof venerids with sizes comparable to adult Neoleptoii are illustrated here (Figs 78109).
Ch amelea striatula (da Costa, 1778)
(Figs 78-89)
Adult specimens have three cardinal teeth on the left valve (2a, 2b and 4b) andthree on the right valve (1, 3a and 3b) placed below the beak; no laterals.
In early ontogenetic stages (Figs 78, 79: specimen 0.8 mm in diameter), theright valve has 4b inconspicuous; the 2b is short and fused proximally with theelongate 2a. The left valve has a short, rectangular 3b fused proximally with anelongate 3a, the latter very close to the dorsal margin of the hinge. Cardinal I issmall, close to the lower margin of the hinge plate and separated from the 3a3b by a broad, chevron-shaped 2a'-2b' socket. The ligament is external butsunken, bordering dorsally cardinal 4b.
In later stages (over 1 mm in diameter), the right valve has 4b more definitethough it remains narrow and very close to the dorsal edge of the hinge plate;the junction between 2b and 2a narrows when size increases and, in specimensover ca. 3.5 mm in diameter, the 2a and 2b are separated. The left valve shows3a and 3b junction narrowing, then both teeth separated; cardinalI reaches increasingly towards the upper part of the hinge plate, separating two distinct, triangular, 2a' and 2b' sockets. The ligament becomes more conspicuous fromoutside.
Gouldia m t n i m a (Montagu, 1803)
(Figs 90·101)
Adult specimens have three cardinal teeth on the left valve (2a, 2b and 4b) andthree on the right valve (1, 3a and 3b) placed below the beak; weak anterior laterals forming later than the cardinals.
In early ontogenetic stages (Figs 90,91: specimen 0.7 mm in diameter), theright valve has a small 4b; the 2b is strong and fused proximally with a weaker2a. The left valve has a rather elongate 3b fused proximally with an elongate 3a,the latter very close to the dorsal margin of the hinge. CardinalI is small, closeto the lower margin of the hinge plate and separated from the 3a-3b by a broad,chevron-shaped 2a'-2b' socket. The ligament is external but sunken, borderingdorsally cardinal 4b.
In later stages (1-2 mm in diameter), the right valve has anterior lateral AIlsplitting off 2a, but the hook-shaped 2a-2b remain fused proximally. On the left
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2
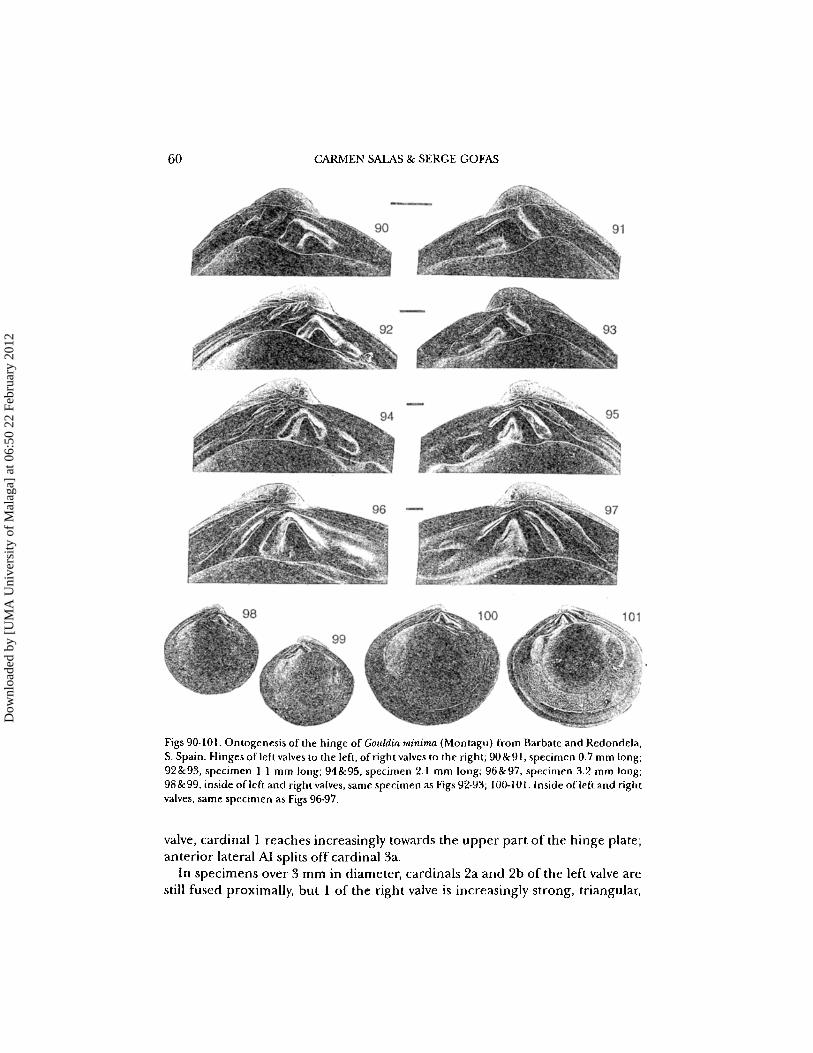
60 CARMEN SAlAS & SERGE GOFAS
Figs 90-101. Ontogenesis of the hinge of Gouldia minima (Montagu) from Barbate and Redondela,S. Spain. Hinges of left valves to the left, of right valves to the right; 90&91, specimen 0.7 mm long;92&93, specimen 1.1 mm long; 94&95, specimen 2.1 mm long; 96&97, specimen 3.2 mm long;98&99, inside oflefl and right valves, same specimen as Figs 92-93; 100-101. Inside of left and rightvalves, same specimen as Figs 96-97.
valve, cardinalI reaches increasingly towards the upper part of the hinge plate;anterior lateral AI splits off cardinal 3a.
In specimens over 3 mm in diameter, cardinals 2a and 2b of the left valve arestill fused proximally, but I of the right valve is increasingly strong, triangular,
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2

NEW SPECIES OF NEOLEPTON 61
and reaches high on the hinge plate. The ligament remains deeply sunken inthe nymph and bordering dorsally 4b.
Pitar ru di s (Poli, 1795)
(Figs 102, 103, 106, 107)
Adult specimens have three cardinal teeth on the left valve (2a, 2b and 4b), andthree on the right valve 0, 3a and 3b), placed below the beak. A weak, knoblike anterior lateral is formed later than the cardinals.
At a shell diameter of 1.4 mm (Figs 102, 103), the right valve has anterior lateral AIl splitting off 2a, but the hook-shaped 2a-2b remain fused proximally. Onthe left valve, cardinal 1 reaches moderately towards the upper part of thehinge plate and there still is a broad 2a'-2b' socket.
The ligament at these sizes is visible from outside when the shell is closed.
ClausineLLa fasciata (da Costa, 1778)
(Figs 104, 105, 108, 109)
Adult specimens have three cardinal teeth on the left valve (2a, 2b and 4b) andthree on the right valve 0, 3a and 3b) placed below the beak; no laterals.
At 0.86 rnrn length, (Figs 104, 105), the left valve has a well defined, thoughsmall 4b and quite strong, proximally fused 2a and 2b. Right valve with quite elongate 3a and 3b, still fused but narrowing very much at their proximal junction; cardinal l reaching moderately towards the upper part of the hinge plate.
DISCUSSION
Reproduction and larval development
The sexes are separate in the species where anatomical data are available (N.sulcatulum: this study; N. antipodum: Ponder, 1969; N. sootryeni: Soot-Ryen,1960).
The brooding of larvae has only been documented for N. sootryeni, and SootRyen (1960) observed that the eggs were glued to the glandular portion of theanterior inner surface of the mantle. We observed conspicuous glandular cellswith a similar position in N. sulcatulum (Figs 14, 17) and a similar glandular portion can bee seen in N. antipodum (Figs 3 E-F of Ponder, 1968). Considering thedata for N. sootryeni, we consider that this glandular area may have a function related with brooding; however Ponder (1968) interpreted them as mucousglands, which may be used to control the amount of silt particles entering themantle cavity. The morphology of the protoconch, quite unusual among the Bivalvia with its large, irregular central surface devoid of growth lines and narrowcom marginal rim, strongly suggests that there is no planktonic stage and thatthe larvae may be brooded in all the species studied.
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2

62 CARMEN SALAS & SERGE GOFAS
It is however surprising that no brooded larvae were found in the rather largenumber of live-taken specimens examined. This contrasts with species in othergenera known to brood (e.g. Dacrydium: Salas & Cofas, 1997) in which virtuallyevery other specimen opened contained larvae. A possible explanation is thatthe larvae have a short time of residence within the mantle cavity, possibly inspring. All our material of the Atlantic species was collected in summer, exceptfor the New Caledonian specimens collected at the end of the winter; conversely the material examined by Soot-Ryen (1960) was collected in spring. Amarked seasonality can be expected in species living in the littoral zone of temperate areas, and contrasts with the timing of reproduction in outer shelf ordeep-sea species such as Dacrydium spp., which may last all year round.
Systematic position
The currently accepted position of the Neoleptonidae is within the superfamilyCyamiacea Philippi, 1845. This superfamily was proposed by Thiele (1934) (nomen translatumfrom Cyamiidae) to accomodate the families Cyamiidae Philippi,1845, Sportellidae Dall, 1899, and Neoleptonidae Thiele, 1934. Coan (1984)has subsequently included the Bernardinidae Keen, 1963, initially placed in thesuperfamily Arcticacea Newton, 1891.
Ponder (1971) studied the hinges and morphology of several New Zealandand Subantarctic species belonging to the Cyamiacea and summarized the anatomical characters of the group:
a) There are two posterior pallial (branchial in Ponder's terminology) openings, usually bordered with short tentacles.
b) The outer demibranch is reflected over the gill axis and is shorter than theinner demibranch. The gills are attached to each other posteriorly and to theposterior mantle margin.
c) The foot is small or of moderate size and usually has a byssal gland and/orventral groove. There are no protractor pedal muscles but anterior and posterior retractor pedal muscles are present.
The superfamily Veneracea is currently accepted to include five families (Keenin Moore, 1969): Veneridae Rafinesque, 1815, Petricolidae Deshayes, 1839,Cooperellidae Dall, 1900, Clauconomidae Gary, 1853, and Rzehakiidae Korobkov, 1954. Most of the species of the superfamily belong to the Veneridae.
Ansell (1961, 1962) studied the functional morphology of the British speciesofVeneracea and noted as important characters:
a) The two posterior openings are siphons which vary in length, extent of fusion and development of tentacles. They are derived from the inner and middlefolds of the mantle edge. A constant feature of the siphons of the Veneridae isthe presence of siphonal membranes.
b) The outer demibranch is always well developed and about half as long asthe inner one. Both demibranchs are reflected and long cirri are present to-
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2

NEW SPECIES OF NEOLEPTON 63
wards the posterior end.c) The foot is well developed; the byssal gland is functional at settlement (An
sell, 1962) but only in Venerupis does it remain functional in the adult.Ansell (1961) pointed out that the mantle edge of the Veneridae is split into
four folds: an outer fold which secretes the outer calcareous layer of the shelland the periostracum from its outer and inner surface respectively, and threeothers, the innermost of which is a small flap-like fold directed dorsally. Anselldiscussed the homology of these folds with the usual three folds of the typicalbivalve mantle edge, and concluded that the middle fold is duplicated. Thisview was contended by Ockelmann (1964) on the basis that the siphons areformed by fusion of both innermost lobes. We agree and here call the innerfold the "m l " fold of Ansell (1961).
The Cyamiacea and the Veneracea are both members of the order Veneroidabut are not considered to be closely related, the former being in the Luciniacea-Leptonacea lineage, the latter in the Arcticacea-Veneracea lineage forwhich Newell (1965) erected a separate suborder Arcticina. Nevertheless, thecharacters discriminating Cyamiacea from Veneracea, according to the diagnoses given by Chavan in Moore (1969) and Keen in Moore (1969), respectively,are tenuous. Much is made of the configuration of the hinge, with interlockinghook-shaped elements in the Cyamiacea, three tubercular cardinal teeth ineach valve of the Veneracea, and the position of the ligament, which is internal,adjacent to the nymph in the Cyamiacea and external and opisthodetic in theadult Veneracea.
The differences between the hinge of the juvenile Veneridae here examined,together with those studied by Le Pennec (1973, 1978, 1980) and a Neoleptonare: a) The posterior laterals, well developed in Neolepum, are absent in the Veneridae. b) The posterior cardinal 4b of the left valve is very narrow and borders anteriorly the ligament in the Veneridae. In Neolepton, the cardinal 4b is absent and the ligament overhangs an empty space behind cardinal3b (see Fig. 2).
The ligament is opisthodetic in the Veneridae and develops from internal topartially external and then external. The internal ligament of Neolepton is hereinterpreted as homologous with the external ligament of Veneridae, the presence/absence of 4b and the internal vs. external positions of the ligament being viewed as the same character state transformation (compare Figs 1,2 withthe venerid hinges Figs 78-105). The absence of 4b and the internal ligamentare, in this hypothesis, larval characters of Veneridae retained in Neolepton.
Otherwise, the general "bauplan" for the hinges ofajuvenile Veneridae andNeolepton is similar. The peculiar hook-shaped structure formed by the cardinalsfused proximally is found in both; it is later lost in the ontogeny of a Veneridaeby progressive narrowing of the fused part, and ultimately separation of theteeth. On these grounds, we do not agree with Chavan (1969) to reject Tuttoniafrom the Veneracea on the grounds of having a hinge "not cyrenoid".
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2
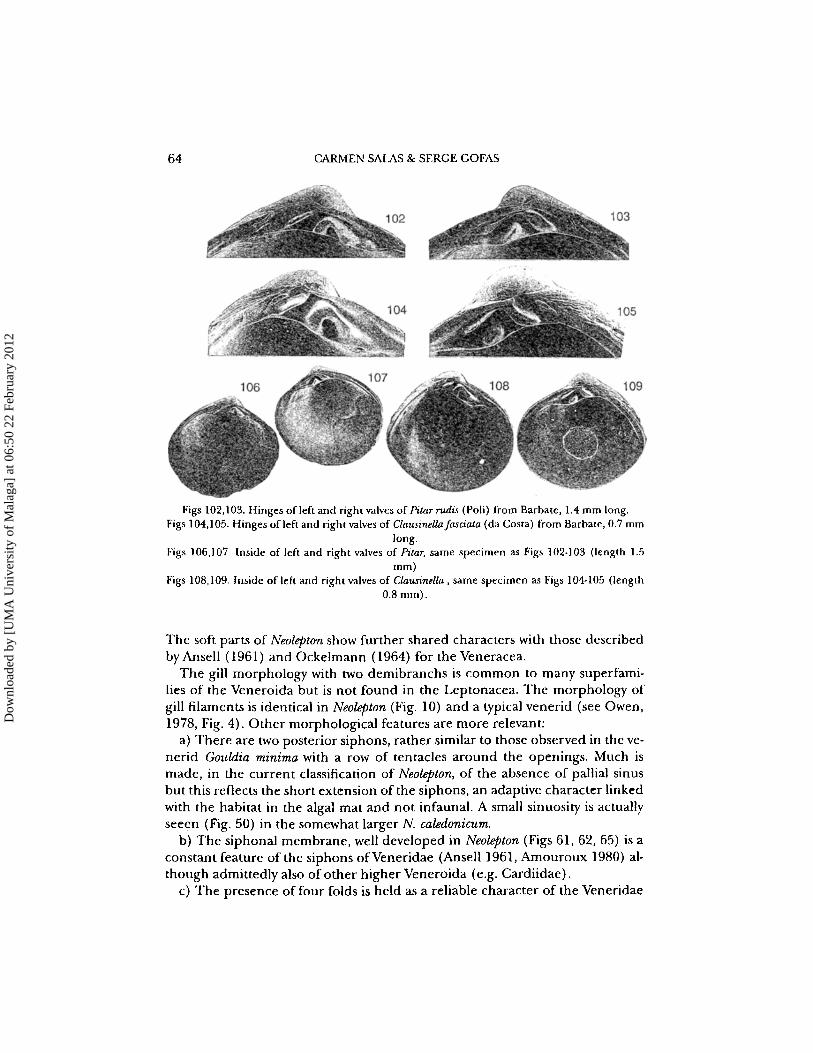
64 CARMEN SAlAS & SERGE GOFAS
Figs 102,103. Hinges of left and right valves of Pitar rudis (Poli) from Barbate, 1.4 mm long.Figs 104,105. Hinges of left and right valves of Clausinella[asciata (da Costa) from Barbate, 0.7 mm
long.Figs 106,107. Inside of left and right valves of Pitar, same specimen as Figs 102·103 (length 1.5
mm)Figs 108,109. Inside of left and right valves of Clausinella , same specimen as Figs 104-105 (length
0.8 mm).
The soft parts of Neolepton show further shared characters with those describedby Ansell (1961) and Ockelmann (1964) for the Veneracea.
The gill morphology with two demibranchs is common to many superfamilies of the Veneroida but is not found in the Leptonacea. The morphology ofgill filaments is identical in Neolepton (Fig. 10) and a typical venerid (see Owen,1978, Fig. 4). Other morphological features are more relevant:
a) There are two posterior siphons, rather similar to those observed in the venerid Gouldia minima with a row of tentacles around the openings. Much ismade, in the current classification of Neolepton, of the absence of pallial sinusbut this reflects the short extension of the siphons, an adaptive character linkedwith the habitat in the algal mat and not infaunal. A small sinuosity is actuallyseeen (Fig. 50) in the somewhat larger N. caledonicum.
b) The siphonal membrane, well developed in Neolepton (Figs 61,62,65) is aconstant feature of the siphons ofVeneridae (Ansell 1961, Amouroux 1980) although admittedly also of other higher Veneroida (e.g. Cardiidae).
c) The presence of four folds is held as a reliable character of the Veneridae
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2

NEW SPECIES OF NEOLEPTON 65
although a broader knowledge of all eulamellibranch superfamilies may showits occurrence elsewhere. Other bivalves studied (in the families Cardiidae, Carditidae, Vesicomyidae, Astartidae, Chamidae: unpublished data) for this character have only three mantle folds. The main elements of the venerid mantle margin can be recognized in Neolepton (Fig. 16).
The structure and mineralogy of the shell does not bring any evidence supporting nor objecting to the affinity of Neoleptonidae with the Veneracea. According to Taylor et al. (1973), the structure and mineralogy of the shell of Neoleptoncobbi (Cooper & Preston, 1910) is aragonitic with two homogeneous layers, similar to those found in the family Cyamiidae [Cyamium antarcticum Philippi,1845] of the Cyamiacea, as well as in the Cooperellidae (Cooperella subdiaphana(Carpenter, 1864)] of the Veneracea.
We conclude that Neoleptoii sulcatuLum - and thus the family Neoleptonidae may be more closely related to the Veneracea than its current classification suggests. This conclusion does not extend to all the other species originally or subsequently included in Neolepton or the Neoleptonidae (many are now includedin the Montacutidae or related families, see appendix). Using the terminologyof heterochrony of McNamara (1986), the genus Neolepton may have evolved bypaedomorphosis, retention of ancestral juvenile characters (here, of the hinge)in the descendant adult phase, through progenesis ("precocious sexual maturation"). The same - with more far-reaching transformations including suppression of the separate posterior openings - applies to Turtonia minuta (Fabricius),which would have evolved from the genus Venerupis as suggested by Ockelmann(1964) .
The observed differences separating Veneridae and Neoleptonidae involvetwo important transformations of the hinge, from that of a possible commonancestor: the loss of posterior laterals in Veneridae, and the loss of cardinal 4and correlated sinking of the ligament in Neoleptonidae. We consider this difference sufficient to maintain the two families separate. The inclusion of bothfamilies in the Veneracea will be ascertained if the shared characters discussedabove - reflected inner mantle lobe in particular - are not found in othersuperfamilies of the Veneroida, and can be retained as a synapomorphy.
The superfamily Cyamiacea currently includes three families in addition toNeoleptonidae: Cyamiidae, Sportellidae and Bernardinidae. The GaimardiidaeHedley, 1916 were included in the Cyamiidae by Ponder (1971) but maintainedas a separate, though closely related family, by Morton (1979). The anatomicalcharacters given by Ponder (1971) for the Cyamiidae, and those of the Veneracea from Ansell (1961), are not extremely different. The arguments developedabove for relating the hinge of Neolepum with that of the Veneridae may apply also to Cyamium and to many genera currently classified in the Cyamiacea. Coan(1984) drew attention to the possibility that the two bernardinid genera may be
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2

66 CARMEN SAlAS & SERGE GOFAS
reminiscent of some species of the Veneridae in which the ligament is somewhatsunken below the hinge margin and which brood their young, as Nutricola(=Transennella) tantilla (Gould, 1853); but he found that the lack of a pallial sinus, the less conspicuous beaks and the internal ligament, are characteristicswhich do not allow to consider the bernardinids as venerids. We have shownthat the lack of a pallial sinus is not an important character at higher level ifshort siphons remain present. Stohlena salmonea (Carpenter, 1864) presents asinuosity in the pallial line (Coan, 1984: Fig. 8b) quite similar to that found inNeolepton caledonicum or in juveniles of Gouldia minima. Thus, we consider it important to reevaluate the systematic position of the Bernardinidae, using livingmaterial. Sportellidae is based on SportellaDeshayes, 1858, with a fossil type species (Psammolea dubia Deshayes, 1824, from the Eocene of Paris Basin), and itsplacement is not very well documented. Other genera currently placed in Sportellidae (e.g. Basterotia Mayer in Homes, 1859) are strongly reminiscent of theLeptonaceajudging from hinge characters and pallial line.
We suspect that the superfamily Cyamiacea, as currently understood, is polyphyletic. It may be a catchall of unrelated paedomorphic species which, accordingly with their small sizes, show a simplification of the hinge characters andmorphology; some of them have the appearance of post-larval and juvenile veneraceans, while others may be related to the leptonacean lineage.
REFERENCES
Amouroux,j.-M., 1980. Etude monographique des siphons de quelques mollusques bivalves: adaptation et morphologie. - Oceanis 5 (1): 33-89.
Ansell, A.D., 1961. The functional morphology of the British species ofVeneracea (Eularnellibranchia). - Journal of the Marine Biological Association of the United Kingdom 41: 489-515.
Ansell, A.D., 1962. The functional morphology of the larva, and post-larval development of Venus
striatula (Da Costa). - Journal of the Marine Biological Association of the United Kingdom 42:419-443.
Arnaud, F., P. M. Arnaud, A. Intes & P. Le Loeuff, 1976. Transport dinvertebres benthiques entrel'Afrique du Sud et Sainte Helene par les larninaires (Phaeophyceae). - Bulletin du MuseumNational dHistoire Naturelle (3e serie) 384, Ecologic generate 30: 49-55.
Bartsch, P., 1915. Report on the Turton collection of South African marine mollusks, with additional notes on other South African shells contained in the United States National Museum. ~United States National Museum Bulletin 91: i-xii and 1-305, pI. I-54.
Bernard, F., 1895. Premiere note sur le developperneru et la morphologie de la coquille chez lesLarnellibranches. - Bulletin de la Societe Geologique de France (3) 23: 104-154.
Bernard, F., 1897_ Sur quelques coquilles de Larnellibranches de rile Stewart. - Bulletin duMuseum d'Histoire Naturelle 3(7}: 309-314.
Bernard, F., 1898. Sur quelques coquilles de Lamellibranches de rile Saint-Paul. - Bulletin duMuseum d'Histoire Naturelle 4(2}: 78-86.
Bogi, C., M. Coppini, & A. Margelli, 1992. Ritrovameruo di Litigiella glabra (P. Fischer, (873) nelleacque di Livorno (Bivalvia: Lasaeidae). - La Conchiglia 264: 43--44.
Bourne, G.c., 1906. Report on [ousseaumia. a new genus of Eularnellibranchs commensal with thecorals Heterocyathus and Heteropsammia. - Ceylon Pearl Oyster Fisheries Report 5: 243-266.
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2

NEW SPECIES OF NEOLEPTON 67
Bowderi.]. & D. Heppell, 1968. Revised list of British Mollusca. 2. Unionacea - Cardiacea. - Journal of Conchology 26: 237-272.
Carrozza, E, 1983. Microdoride di malacologia mediterranea. - Bollettino Malacologico 19: 65-70.Chaster, G.W., 1897. III. Notes on the marine Mollusca of Rathlin Island. - The Irish Naturalist:
184-187.Chavan, A, 1969. Superfamily Cyamiacea Philippi, 1845. Pp. N537-N543. - In R C. Moore (Ed.):
Treatise on Invertebrate Paleontology. Part N, Mollusca 6, Bivalvia, vol.2. Geological Society ofAmerica and University of Kansas Press, Lawrence, Kansas.
Cianfanelli, S. & E. Talenti, 1987. Ulteriori conoscenze sulla malacofauna del Golfo di Baratti (Piornbino-Li). - Bollettino Malacologico 23: 263-268.
Coan, E., 1984. The Bernardinidae of the Eastern Pacific (Molllusca: Bivalvia). - The Veliger 27(2): 227-237.
Cooper,j.E. & H.B. Preston, 1910. Diagnoses of new species of marine and freshwater shells fromthe Falkland Islands, including descriptions of two new genera of marine Pelecypoda. - Annalsand Magazine of Natural History (ser. 8) 5: 110-114, pl. 4.
Cox, L.R, 1969. General features of the Bivalvia. Pp. N2-N129. - In R C. Moore (Ed.): Treatise onInvertebrate Paleontology. Part N, Mollusca 6, Bivalvia, vol. 1. Geological Society of America andUniversity of Kansas Press, Lawrence, Kansas.
Crosse, H., 1885. Nomenclatura generica e specifica di a\cune conchiglie mediterranee, pel Marchese di Monterosato [book review]. - Journal de Conchyliologie 33: 139-142.
Dell, RK., 1964. Antarctic and subantarctic Mollusca: Amphineura, Scaphopoda and Bivalvia. Discovery Reports 33: 93-250, plates II-VII.
Dell, RK., 1990. Antarctic Mollusca with special reference to the fauna of the Ross Sea. - Bulletinof the Royal Society of New Zealand 27: 1-3] 1.
Deroux, G., ]961. Rapports taxonomiques d'un leptonace non decrit "Lepton JUbtrigOnLLm" Jeffreys(nomen nudum-1873). - Cahiers de Biologie Marine 2: 99-]53.
Filhol, H., 1880. Mollusques marins vivant sur les cotes de l'i1e Campbell. - Comptes Rendus deI'Acadernie des Sciences 91: 1094-1095.
Finlay, HJ., 1927. A further commentary on New Zealand molluscan systematics. - Transactionsand Proceedings of the New Zealand Institute 57: 320-485, pl. 18-23.
Hutton, EW., 1883. Notes on some marine mollusca. - New ZealandJournal of Science 1: 477-478.Jeffreys, j.G., 1859. Further gleanings in British conchology. - Annals and Magazine of Natural
History (ser. 3) 3: 30-43.Jeffreys,j.e., ]863. British conchology. - j. van Voorst, London. Vol. II, 479 pp.Keen, A.M., ]963. Marine molluscan genera of western North America; an illustrated key. - Stan
ford University Press, Stanford, California. ]26 pp.Keen, AM., ]969. Superfamily Veneracea Rafinesque, 18]5. Pp. N670-N690. - In R C. Moore
(Ed.): Treatise on Invertebrate Paleontology. Part N, Mollusca 6, Bivalvia, vol.2. Geological Society of America and University of Kansas Press, Lawrence, Kansas.
Laseron, C.F., ]953. Minute bivalves from New South Wales. - Records of the Australian Museum23 (2): 33-53.
Le Pennec, M., ]973. Morphogenese de la charniere chez 5 especes de Veneridae. - Malacologia]2 (2): 225-245.
Le Pennec, M., 1978. Genese de la coquille larvaire et postlarvaire chez divers bivalves marins. Doctoral thesis, Universite de Bretagne Occidentale, Brest. 229 pp., 108 plates.
Le Pennec, M., 1980. The larval and post-larval hinge of some families of bivalve molluscs. - Journal of the Marine Biological Association of the United Kingdom 60: 601-6] 7.
McNamara, K, 1986. A guide to the nomenclature of heterochrony. - Journal of Paleontology 60(1): 4-13; 4 figs.
Monterosato, T. A di, 1875. Nuova rivista delle conchiglie mediterranee. - Aui della AccaderniaPalermitana di Scienze, Lettere ed Arti, Palermo (sez. II) 5:1-50.
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2

68 CARMEN SALAS & SERGE GOFAS
Monterosato, T.A di, 1908. Note sur l'Etycilla cuenoti. -Journal de Conchyliologie 56: 253-255.Morton, 8., 1979. The biology, functional morphology and taxonomic status of Gai11lal"dia (Neogai
mardia} finlayi (Bivalvia: Gaimardiidae). - Journal of Zoology 188: 123-142.Newell, N.D., 1965. Classification of the Bivalvia. - American Museum Novitates 2206: 1-25.Nicol, D., 1966. Description, ecology, and geographic distribution of some Antarctic Pelecypods.
Bulletins of American Paleontology 51 (231): 1-102.Nordsieck, E, 1969_ Die europaischen Meeresmuscheln. Vom Eismeer bis Kapverden, Miuelmeer
und Schwarzes Meer, - G. Fischer, Stuttgart. XIII + 256 pp.Ockelmann, K.W., 1964. Turtonia min uta (Fabricius), a neotenous veneracean bivalve. - Ophelia 1
(1): 121-146.Owen, G., 1978. Classification and the bivalve gill. - Philosophical Transactions of the Royal Soci
ety of London, ser. B, 284: 377-385.Pelseneer P., 1909. Phylogenie des Lamellibranches cornrnensaux. - Bulletin de I'Acadernie Roy
ale de Belgique (classe des Sciences) 1909 (12): I 14~1150.Pelseneer, P., 1911. Les Lamellibranches de I'expedition du Siboga. Partie anatomique. - Siboga
Expeditie 53a: 125 p., 25 pl. Leiden, EJ.Brill.Pelseneer, P., 1925. Un larnellibranche commensal de lamellibranche et quelques autres larnelli
branches commensaux. - Travaux de la Station Zoologique de Wimereux 9: 166-182.Phorson,j. E., 1988. Neolepton sykesiand Neolepton sulcatulum in Guernsey. - Conchological Newslet
ter 106: 117-120.Ponder, W.E, 1969. Notes on two neoleptonid bivalve molluscs. - New Zealand Journal of Marine
and Freshwater Research 3(2): 262-272.Ponder, W. F., 1971. Some New Zealand and subantarctic bivalves of the Cyamiacea and Leptona
cea with descriptions of new taxa. - Records of the Dominion Museum 7 (3): 119-141.Powell, A.W.B., 1957. Mollusca of Kerguelen and Macquarie Islands. - Reports of the B.A.N.Z. Ant
arctic Research Expedition 1929-1931, ser, B, 6 (7): 107-150,2 pl.Powell, AW.B., 1979. New Zealand Mollusca, marine, land and freshwater shells. - William Col
lins, Auckland. 500 pp.Preston, H.B., 1912. Characters of six new pelecypods and two new gastropods from the Falkland
Islands. - Annals and Magazine of Natural History (ser. 8) 9: 636-640, pl. 21.Sabelli, B., R. Ciannuzzi-Savelli & D. Bedulli, 1990. Catalogo annotate dei Molluschi marini del
Mediterraneo. - Libreria Naturalistica Bolognese, Bologna. Vol. I, 348 p.Salas, C. & S. Cofas, 1997. Brooding and non-brooding Dacrydium: a review of the Atlantic species.
- Journal of Molluscan Studies 63: 261-283.Smith, EA., 1890. Report on the marine molluscan fauna of the island of St. Helena. - Proceed
ings of the Zoological Society of London: 247-317, pI. 21-24.Soot-Ryen, T., 1960. Pelecypods from Tristan da Cunha. - Results of the Norwegian expedition to
Tristan da Cunha 1937-1938 no. 49. H. Aschehoug & Co., Oslo. 47 pp.Suter, H., 1913. Manual of the New Zealand Mollusca. - John Mackay, Wellington. H20 pp.Taylor,J.D., , w.j. Kennedy & A Hall, 1973. The shell structure and mineralogy of the Bivalvia. II.
Lucinacea-Clavagellacea. Conclusions. - Bulletin of the British Museum (Natural History),Zoology 22 (9): 253-294, pUS.
Tebble, N., 1966. British bivalve seashells. - Trustees of the British Museum (Natural History).London. 212 pp.
Thiele,]., 1934. Handbuch der systematischen Weichtierkunde. - Gustav Fischer,Jena. Bd. 2, Dritter Teil, pp. I-V, 779-1022.
Waren, A., 1980. Marine Mollusca described byJohn Gwyn Jeffreys, with the location of the typematerial. - Conchological Society of Great Britain and Ireland. Special publication 1,60 pp.
Waren, A. & F. Carrozza, 1993. Arculus sykesi (Chaster), a leptonacean bivalve living on a tan aidCrustacea in the Gulf of Genova. - Bollettino Malacologico 29: 303-306.
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2

NEW SPECIES OF NEOLEPTON
APPENDIX 1
69
A review of some species which have been assigned to the Neoleptonidae
These are the species originally included in Neoleptorc by Monterosato (1875), and type species ofthe genera currently accepted in the Neoleptonidae (Chavan in Moore, 1969).
A. Species which are excluded from the Neoleptonidae
Lepton tlarkiae Clark, 1852. Type species of Epilepton Dall, 1899 by original designation. Type locality: Exmouth, SW England.
Epilepton is included in the Neoleptonidae by Thiele (1934) and Chavan in Moore (1969). Deroux (1961: 147-150) described the animal as having a single dernibranch and a single posterioropening, and transferred it to a new genus Potidoma in the Montacutidae. We agree with this familial placement. Pelseneer (1925) and Deroux (1961) have shown this species to be an externalcommensal on sipunculids.
Lepton glalJrum Fischel', 1873 (= Erycina cuenoti Lamy, 1908. Type species of Litigiella Monterosato,1909, by original designation =Montacuta perezi Pelseneer, 1909). Type locality: Arcachon, France.
Pelseneer (1911: 46 and XTV) has shown that it has a single demibranch and a single posterioropening, and correctly classified it in the Montacutidae. It is a commensal on the sipunculid Sipunculus nudus (Linne, 1758) on sandy shores (Lamy 1908; Pelseneer 1911). The hinge is very similarto that of Epilepton clarkiae, so that the genera Litigiella and Epilepton are probably synonyms. Bogi,Coppini & Margelli (1992) give a review and illustration of the species.
Neolepum ohliquatuni Chaster, 1897. Type locality: Rathlin Island, Northern Ireland.No living specimens are known to us but the hinge is very similar to that of Mysella. It may be
provisionally retained as Mysella obliquata in the Montacutidae. Shells are illustrated by Carrozza(1983) and by Cianfanelli & Talenti (1987).
Lepton sykesi Chaster, 1895. Type species of Arculus Monterosato, 1909. Type locality: Guernsey,Channel Islands.
It is placed in Neoleptonidae by Thiele (1934) and Chavan in Moore (1969); in Neolepuniby Tebble (1966). Its family placement is unsettled, but its mode of life attached to the ventral part of atanaid (Waren & Carrozza 1993) strongly suggests that it belongs to the Leptonacea.
Lepton parasiticum Dall, 1876. Type locality: Kerguelen Islands, southern Indian Ocean.Nicol (1966) studied the type series and queried its placement in Neoleptoti (=Notolepton) by Pow
ell (1957) and Dell (1964). He proposed for this species a new genus Waldo, still tentatively retained in the Neoleptonidae. Dell (1990) turned back Lepton parasiticum to its original binomenawaiting further studies. We examined specimens from Kerguelen, which have a mantle edge protruding outside the valves and provided with few, large tentacles; a small siphon-like posterioraperture, and the remainder of the posterior margin fused; the pedal gape extending posteriorlyuntil the vertical midline, and only one demibranch. It is an external commensal on echinids, andmost likely belongs to the Montacutidae or Galeommatidae.
[ousseaumiella keterocyathi (Bourne, 1906). Type species ofjousseaumiella Bourne, 1907 = jousseoumiaBourne, 1906 non Sacco, 1894; subsequent designation by Chavan m Moore, 1969. Type locality:Ceylon.
The anatomical characters are discussed by Pelseneer (l909) and point to a placement in theMontacutidae. There is only one demibranch and one posterior aperture. It is, like Epilepton, commensal on the sipunculid Aspidosiphon embedded in corals.
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2

70 CARMEN SALAS & SERGE GOFAS
B. Species which are retained in the Neoleptonidae
Lutetina antarctica Velain, 1876. Type species of Lutetina Velain, 1876. Type locality: St Paul Island,southern Indian Ocean.
Thiele (1934) placed Luietina in the Neoleptonidae, and treated it as a possible synonym of Neolepton following Bernard (1898). Chavan in Moore (1969) placed it in the Cyamiidae and reproduced Bernard's (1898) excellent illustration of the hinge. We have examined the syntypes (3specimens and 3 valves) from St Paul Island in the MNHN, and agree with Bernard that there is noessential difference from the hinge of Neolepton, except for the relative size of teeth. The hinge ofLutetina has very strong, protruding 2a and 1 cardinals, and strong posterior laterals reaching highadapically, making the ligament still more internal than in typical Neolepum. Based on shell characters, Lutetina should definitely be maintained in the Neoleptonidae and may even be a synonym ofNeolepton, but other characters, including siphons and mantle edge, should be examined.
Calvitium glabrum Laseron, 1953. Type species of Caloitium Laseron, 1953, by original designation.Type locality: Port Hacking, New South Wales, Australia.
This genus was placed in the Neoleptonidae by Chavan in Moore, 1969, although Laseron(1953) suggested relationship with Cyamiomactra (currently in the Cyamiidae).
Pachykellya eduiardsi Bernard, 1898. Type species of Pachskellsa Bernard, 1898, by monotypy. Typelocality: Stewart Island, southern New Zealand.
This genus was placed in the Neoleptonidae by Thiele (1934) and Chavan in Moore (1969).
Puyseguria cuneata Powell, 1927. Type species of Puyseguria Powell, 1927 by original designation.Type locality: Puysegur point, SW Otago, southern New Zealand.
Also placed in the Neolcptonidae by Thiele (1934, misspelled Puysegeria) and Chavan in Moore(1969).
Dow
nloa
ded
by [
UM
A U
nive
rsity
of
Mal
aga]
at 0
6:50
22
Febr
uary
201
2




















