
Delivery of a cocktail DNA vaccine encoding cysteine proteinases type I, II and III with solid lipid...
9
Delivery of a cocktail DNA vaccine encoding cysteine proteinases type I, II and III with solid lipid nanoparticles potentiate protective immunity against Leishmania major infection ☆ Delaram Doroud a,b,1 , Farnaz Zahedifard a,1 , Alireza Vatanara b, ⁎ ,2 , Abdolhossein Rouholamini Najafabadi b , Yasaman Taslimi a , Rouholah Vahabpour c , Fatemeh Torkashvand d , Behrooz Vaziri d , Sima Rafati a, ⁎⁎ ,2 a Molecular Immunology and Vaccine Research Laboratory, Pasteur Institute of Iran, Tehran, Iran b Department of Pharmaceutics, School of Pharmacy, Tehran University of Medical Sciences, Tehran, Iran c Hepatitis and AIDS Department, Pasteur Institute of Iran, Tehran, Iran d Biotechnology Research Center, Pasteur Institute of Iran, Tehran, Iran abstract article info Article history: Received 3 February 2011 Accepted 11 April 2011 Available online 19 April 2011 Keywords: Cationic solid lipid nanoparticles Delivery system Leishmania Cocktail DNA vaccine Cysteine proteinases Earlier generations of Leishmania vaccines have reached the third-phase of clinical trials, however none of them have shown adequate efficacy due to lack of an appropriate adjuvant. In this study, cationic solid lipid nanoparticles (cSLNs) were used to formulate three pDNAs encoding L. major cysteine proteinase type I (cpa), II (cpb) and III (cpc). BALB/c mice were immunized twice with a 3-week interval, with SLN-pcDNA-cpa/b/c, pcDNA- cpa/b/c, SLN, SLN-pcDNA and PBS. Footpad assessments, parasite burden, cytokine and antibody responses were evaluated. Mice vaccinated with SLN-pcDNA-cpa/b/c significantly (p b 0.05) showed higher protection levels with specific Th1 immune response development compared to other groups. This is the first report demonstrating cSLNs as a nanoscale vehicle boosting immune response quality and quantity; in a designable trend. The nanomedical feature of this novel formulation can be applied for wide-spread use in genetic vaccination against leishmaniasis, which is currently managed only through relatively ineffectual therapeutic regimens. © 2011 Elsevier B.V. All rights reserved. 1. Introduction Sandfly-transmitted protozoan parasites of the genus Leishmania are associated with broad spectrum of diseases ranging from simple cutaneous to invasive visceral leishmaniasis. Considerable efforts have been devoted to vaccine development against this infection. Definitely, whole-parasite approaches have not shown to be helpful for establishment of a consistently effective, stable and reliable vaccine against leishmaniasis [1]. Crude or purified antigens have been shown to elicit certain degrees of protection in susceptible mouse strains such as BALB/c. In this regard, DNA immunizations have also been investigated in the recent years for this purpose [1,2]. These kinds of vaccines are potent activators of innate and adaptive immunity and prove to be promising immunization tools in a variety of experimental models. Furthermore, it is possible to formulate several antigens from different parasite life cycle stages or subspecies as one vaccine shot. It is also a cost-effective technology for developing countries [2]. Leishmania express large quantities of cysteine proteinases (CPs) which are members of the papain-superfamily [3]. In Leishmania major (L. major), two most important CPs have been described. CPB (type I) presents maximally at the amastigote developmental stage [4]. CPA (type II) is expressed at higher level in amastigote stage and stationary phase promastigote [5]. CPC (Type III) presents in all life cycle stages of Leishmania and its activity is necessary for the parasite's survival as amastigote forms within the host macrophages [6]. We have reported high immunogenicity and protective potential of L. infantum CPC by using a prime-boost strategy in BALB/c mice [7]. Furthermore, we demonstrat- ed that the combination of CPA/CPB is more protective against visceral and cutaneous leishmanial infection than the separate forms [8–11]. Nevertheless and despite all of these valuable data in experimental leishmaniasis, genetic vaccines are poorly immunogenic on their own and still faced with a number of significant hurdles mainly finding a safe and effective delivery system. Cationic solid-lipid nanoparticles (cSLN) represent one of the advanced delivery technologies with potential for formulating biologics to overcome shelf-life stability, cost and toxicity issues. These nanoparticles consist of physiologically Journal of Controlled Release 153 (2011) 154–162 ☆ Financial support information: This work was financially supported by grants from Tehran University of Medical Sciences and Pasteur Institute of Iran for D. Doroud's PhD studentship. The mentioned funding sources, had no involvement in study design; collection, analysis and interpretation of data; writing of the report; and decision to submit the paper for publication. ⁎ Corresponding author. Tel.: + 98 21 66953311x2112; fax: + 98 21 66496560. ⁎⁎ Corresponding author. Tel./fax: + 98 21 66959057. E-mail addresses: [email protected] (A. Vatanara), [email protected], [email protected] (S. Rafati). 1 Equally participated as first author. 2 S.R. and A.V. equally designed research. 0168-3659/$ – see front matter © 2011 Elsevier B.V. All rights reserved. doi:10.1016/j.jconrel.2011.04.011 Contents lists available at ScienceDirect Journal of Controlled Release journal homepage: www.elsevier.com/locate/jconrel GENE DELIVERY
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Delivery of a cocktail DNA vaccine encoding cysteine proteinases type I, II and III with solid lipid...

Journal of Controlled Release 153 (2011) 154–162
Contents lists available at ScienceDirect
Journal of Controlled Release
j ourna l homepage: www.e lsev ie r.com/ locate / jconre l
GENEDELIVERY
Delivery of a cocktail DNA vaccine encoding cysteine proteinases type I, II and IIIwith solid lipid nanoparticles potentiate protective immunity againstLeishmania major infection☆
Delaram Doroud a,b,1, Farnaz Zahedifard a,1, Alireza Vatanara b,⁎,2, Abdolhossein Rouholamini Najafabadi b,Yasaman Taslimi a, Rouholah Vahabpour c, Fatemeh Torkashvand d, Behrooz Vaziri d, Sima Rafati a,⁎⁎,2
a Molecular Immunology and Vaccine Research Laboratory, Pasteur Institute of Iran, Tehran, Iranb Department of Pharmaceutics, School of Pharmacy, Tehran University of Medical Sciences, Tehran, Iranc Hepatitis and AIDS Department, Pasteur Institute of Iran, Tehran, Irand Biotechnology Research Center, Pasteur Institute of Iran, Tehran, Iran
☆ Financial support information: This work was financTehran University of Medical Sciences and Pasteur Institstudentship. The mentioned funding sources, had nocollection, analysis and interpretation of data; writingsubmit the paper for publication.⁎ Corresponding author. Tel.: +98 21 66953311x211⁎⁎ Corresponding author. Tel./fax: +98 21 66959057
E-mail addresses: [email protected] (A. [email protected] (S. Rafati).
1 Equally participated as first author.2 S.R. and A.V. equally designed research.
0168-3659/$ – see front matter © 2011 Elsevier B.V. Adoi:10.1016/j.jconrel.2011.04.011
a b s t r a c t
a r t i c l e i n f oArticle history:Received 3 February 2011Accepted 11 April 2011Available online 19 April 2011
Keywords:Cationic solid lipid nanoparticlesDelivery systemLeishmaniaCocktail DNA vaccineCysteine proteinases
Earlier generations of Leishmania vaccines have reached the third-phase of clinical trials, however none of themhave shown adequate efficacy due to lack of an appropriate adjuvant. In this study, cationic solid lipidnanoparticles (cSLNs) were used to formulate three pDNAs encoding L. major cysteine proteinase type I (cpa), II(cpb) and III (cpc). BALB/cmicewere immunized twicewith a 3-week interval, with SLN-pcDNA-cpa/b/c, pcDNA-cpa/b/c, SLN, SLN-pcDNA and PBS. Footpad assessments, parasite burden, cytokine and antibody responses wereevaluated.Mice vaccinatedwith SLN-pcDNA-cpa/b/c significantly (pb0.05) showedhigher protection levelswithspecific Th1 immune response development compared to other groups. This is the first report demonstratingcSLNs as a nanoscale vehicle boosting immune response quality and quantity; in a designable trend. Thenanomedical feature of this novel formulation can be applied for wide-spread use in genetic vaccination againstleishmaniasis, which is currently managed only through relatively ineffectual therapeutic regimens.
ially supported by grants fromute of Iran for D. Doroud's PhDinvolvement in study design;of the report; and decision to
2; fax: +98 21 66496560..ara), [email protected],
ll rights reserved.
© 2011 Elsevier B.V. All rights reserved.
1. Introduction
Sandfly-transmitted protozoan parasites of the genus Leishmaniaare associated with broad spectrum of diseases ranging from simplecutaneous to invasive visceral leishmaniasis. Considerable effortshave been devoted to vaccine development against this infection.Definitely, whole-parasite approaches have not shown to be helpfulfor establishment of a consistently effective, stable and reliablevaccine against leishmaniasis [1].
Crude or purified antigens have been shown to elicit certaindegrees of protection in susceptible mouse strains such as BALB/c. Inthis regard, DNA immunizations have also been investigated in therecent years for this purpose [1,2]. These kinds of vaccines are potent
activators of innate and adaptive immunity and prove to be promisingimmunization tools in a variety of experimental models. Furthermore,it is possible to formulate several antigens from different parasite lifecycle stages or subspecies as one vaccine shot. It is also a cost-effectivetechnology for developing countries [2].
Leishmania express large quantities of cysteine proteinases (CPs)which are members of the papain-superfamily [3]. In Leishmania major(L. major), two most important CPs have been described. CPB (type I)presentsmaximally at the amastigotedevelopmental stage [4]. CPA (typeII) is expressed at higher level in amastigote stage and stationary phasepromastigote [5]. CPC (Type III) presents in all life cycle stages ofLeishmania and its activity is necessary for the parasite's survival asamastigote forms within the host macrophages [6]. We have reportedhigh immunogenicity andprotective potential of L. infantumCPCbyusinga prime-boost strategy in BALB/cmice [7]. Furthermore, we demonstrat-ed that the combination of CPA/CPB is more protective against visceraland cutaneous leishmanial infection than the separate forms [8–11].
Nevertheless and despite all of these valuable data in experimentalleishmaniasis, genetic vaccines are poorly immunogenic on their ownand still faced with a number of significant hurdles mainly finding asafe and effective delivery system. Cationic solid-lipid nanoparticles(cSLN) represent one of the advanced delivery technologies withpotential for formulating biologics to overcome shelf-life stability,cost and toxicity issues. These nanoparticles consist of physiologically

155D. Doroud et al. / Journal of Controlled Release 153 (2011) 154–162
GENEDELIVERY
well-tolerated ingredients mostly approved for pharmaceuticalapplication in humans [12]. In this regard, we have reported theformulation of immunogenic cpa and cpb genes with cSLNs consistingof cetyl palmitate, cholesterol, DOTAP and Tween 80, as a novel non-viral DNA vaccine formulation against leishmaniasis [13]. Our resultsrevealed that stable formulation of cSLN was able to protect pDNA inDNase I challenge assay. In vitro COS-7 cells transfection revealedthese formulations have low cytotoxicity and acceptable efficiency/cytotoxicity ratio comparable to linear PEI-25 kDa [13].
Herein, we evaluated the potency of CPs types I, II and III combinationformulated with cSLNs as DNA vaccination strategy against L. majorinfection in BALB/c as susceptible mice model.
2. Methods
2.1. Materials and chemicals
All solutions were prepared using MilliQ™ ultrapure (Milli-Q-System, Millipore, Molsheim, France) and apyrogenic water to avoidsurface-active impurities. Cetyl palmitate, Tween 80 and cholesterolwere purchased from Merck (Darmstadt, Germany). N-[1-(2,3-Dioleoyloxy) propyl]-N,N,N trimethylammonium chloride (DOTAP),Sodium dodecyl sulfate (SDS) were purchased from Sigma-Aldrich(Sigma, Deisenhofen, Germany). The materials applied for PCR,enzymatic digestion and agarose gel electrophoresis were acquiredfrom Roche Applied Sciences (Mannheim, Germany). Cell culturereagents including M199 medium, HEPES, L-glutamine, adenosine,hemin, gentamicin, and RPMI were sourced from Sigma (Darmstadt,Germany) and Gibco (Gibco, Life Technologies GmbH, Karlsruhe,Germany) respectively. Fetal Calf Sera (FCS) was purchased from(Gibco, Life Technologies GmbH, Karlsruhe, Germany).
2.2. cSLN preparation
The cSLN suspension was produced by a technique previouslydescribed byDoroud et al. [13]. Briefly, desired amount of DOTAP (0.4%w/v)was dissolved in hot aqueous phasewhichwas then added to themelted cetyl palmitate and cholesterol (5.1% w/v) phase containingTween 80 at 3.2:1 molar ratio. Emulsification was carried out bystirring themixture at 2000 g by amechanical stirrer (IKA®,Germany)for 10 min at 90 °C. Samples were then homogenized using a highshear homogenizer (IKA®, Germany) at 18,000 g for 15 min. cSLNdispersion was obtained by direct cooling of hot O/W microemulsionon an ice-bath while stirring at 1000 rpm. cSLNs were washed bycentrifugation (6000 g, 10 min, three times) using 100 kDa Amicon®Ultra centrifugal filters (Millipore GmbH, Schwalbach/Ts, Germany) topurify the suspension from the excess amounts of surfactant.
2.3. Plasmid construction and purification
Both pGEM-cpa and pGEM-cpb were available from our previousstudies [10] and each fragmentwas subcloned into pcDNA3.1(−) vector(Invitrogen). pDNAs were transformed into the DH5α E. coli strain andpurified by alkaline lysis method (QIAGEN Endofree Plasmid Giga Kit,Hilden, Germany) and then confirmed by PCR and digestion (data notshown). pcDNA-cpc was also available from our former study [7].
2.4. Vaccine formulation and in vitro characterization
cSLN–pDNA complexes were prepared by adding volumes corre-sponding to 650 μg of each purified pDNA (pcDNA-cpa, pcDNA-cpb,pcDNA-cpc) to cSLN suspension at DOTAP:pDNA ratio of 6:1 (w/w)and 60 min incubation at room temperature separately [13]. Completecondensation and complexation of pDNAs with cSLN were analyzedby agarose gel electrophoresis, as previously demonstrated [13]. Sizeand zeta potential measurements, gel retardation analysis and DNase I
protection study were all performed according to the conditionsdemonstrated in our previous study [13].
2.5. Ethics statement
All mouse experiments including maintenance, animals' handlingprogram and blood sample collection were approved by InstitutionalAnimal Care and Research Advisory Committee of Pasteur Institute ofIran (Education Office dated January, 2008), based on the SpecificNational Ethical Guidelines for Biomedical Research issued by theResearch and Technology Deputy of Ministry of Health and MedicinalEducation (MOHME) of Iran that was issued in 2005.
2.6. Mice and parasites
Female BALB/c mice, (6–8 weeks old, weighing 20±5 g) wereobtained from the Pasteur Institute of Iran, housed in plastic cageswith free access to tap water and standard rodent pellets in an air-conditioned room under a constant 12:12 hr light–dark cycle at roomtemperature with relative humidity (50–60%).
The enhanced GFP gene (egfp) transfected L. major strain, (MRHO/IR/75/ER) was kept in virulent state by continuous passage in BALB/cmice. A homogenized lymph node (LN) from an infected BALB/cmouse was cultured in RPMI-1640 media supplemented with 10%fetal calf serum and 100 μg of gentamycin/ml. Stationary phasepromastigotes were harvested, washed and resuspended at aconcentration of 3×106 cells/50 μl. 50 μl of this preparation wasinjected into the left hind footpad of each mouse.
2.7. Toxicity/safety profile evaluation of the SLN formulation in mice
Acute toxic effects (toxicity/safety profile) in BALB/c mice wereevaluated according to the international guidelines (WHO/OECD),after formulation administration. Briefly, animals weighed individu-ally on day one, after seven days and at weekly periods, subsequently.The animals were observed twice daily, for signs of morbidity anddeath. Clinical signs of toxicity noticed during these routine checkswere recorded. Feed consumption as well as formal clinical observa-tions (including mortality, monitoring of convulsions, lethargy, sleep,coma, salivation, diarrhea, cage side examination, skin color, fur, eyesand mucous membrane, spontaneous and voluntary motor activityand necropsy – in case the animal dies) were performed and recorded,weekly.
2.8. Vaccination schedules
Five groups (13 mice/group) were immunized in the right-hindfootpad. Group 1 was immunizedwith themixture of different Qiagenpurified pDNA vaccine candidates (50 μg of each plasmid) formulatedwith cSLN: (SLN-pcDNA-cpa/b/c). The second group (group 2) wasimmunized with a mixture of the same amount of pDNAs in PBS:(pcDNA-cpa/b/c). Control groups received cSLN (group 3), cSLN-pcDNA (group 4) and PBS (group 5). Three weeks later, the sameregimen was performed as booster immunization. Mice werechallenged 3 weeks after booster immunization and infection pro-gress was followed.
3. Estimation of disease progression
3.1. Footpad assessments
3.1.1. Footpad swellingFootpad swelling was monitored weekly by measuring the
increase in thickness of the inoculated footpad and the uninfectedcontra lateral footpad with a metric caliper.

Table 1cSLN formulations and their characteristics. Nanoparticles were formulated from cetylpalmitate, cholesterol and DOTAP hydrochloride. pDNA condensation was obtained at6:1, DOTAP:pDNA ratio. Results represent mean±SD of three independent SLNpreparations.
Formulation Size (nm) Zeta potential (mV) PDI
a) cSLN-pcDNA-cpa 244±12 +27±3 0.254b) cSLN-pcDNA-cpb 250±15 +22±3 0.264c) cSLN-pcDNA-cpc 240±12 +25±7 0.283
156 D. Doroud et al. / Journal of Controlled Release 153 (2011) 154–162
GENEDELIVERY
3.1.2. Footpad parasitic load by flow cytometryFlow cytometry was used to quantify cells by intracellular
fluorescence as an indicator of Leishmania infection, replication.Mice were euthanized at 9th week post challenge and infectedfootpads were homogenized under sterile conditions. Non-infectedfootpads were also removed and processed in the same manner ascontrol. The recovered viable cell suspensions were directly applied toa Partec PASIII flow cytometer (Partec GmbH, Germany) using abivariate scatter plot of fluorescence versus forward scatter, by gatesetting with non-infected footpad cells. A minimum of 10,000 eventsfrom the viable transfected cell population per sample was analyzed.Parasite inhibition was measured in terms of decrease in greenfluorescence intensity of footpads of vaccinated animals according tothe following formula:
Parasite inhibition %
= 1− Flourescence of the vaccinated footpadMean flourescence of the non� vaccinated footpads
� �× 100:
3.1.3. Footpad imagingTo demonstrate the in vivo level of infection, the infected footpads
were imaged at 9th week after challenge, with the KODAK Image-Station (4000) Digital Imaging System. Briefly, footpads were treatedwith a depilatory substance (Nair) to reduce background autofluorescence. Afterward mice were temporarily anesthetized with amixture of Xylazine 2% (7.5 μl), Ketamine 10% (30 μl) and salinesolution (260 μl)/mouse intraperitoneally and then imaged. Pixelcounting and measurement of the lesions were performed usingKODAK-molecular image software (version 5.3). Measurements werereported as "sum green intensity" defined as the number of greenpixels in a given area multiplied by the average intensity of each pixel.
3.2. Determination of parasite burden
Three mice from each group were sacrificed 9 weeks afterchallenge and parasite burdens were determined as follows: LNswere excised, weighed and then homogenized with a tissue grinder in2 ml of Schneider's Drosophila medium supplemented with 20% heat-inactivated fetal calf serum and Gentamicin (0.1%). Under sterileconditions serial dilutions ranging from 1 to 10−20 were prepared in96-well micro titration plates. After 10 days of incubation at 26 °C,plates were examined with an inverted microscope at a magnificationof 40×. The presence or absence of mobile promastigotes wasrecorded in each well. The final titer was the last dilution for whichthe well contained at least one parasite (21, 22). The number ofparasites per gram was calculated through the following formula:
Parasite burden = − logParasite dilutionOrgan0s weight
� �:
In addition, 10 days after incubation of the LN cells, GFP expressingpromastigotes were visualized using a fluorescence microscope andthen evaluated for EGFP expression quantization by flow cytometryanalysis performed on a 10−3 dilution. Mean fluorescence intensities(MFIs)% were measured with respect to non-infected LNs cells ascontrol. PI% (parasite inhibition percent) was measured according tothe formula described in Section 1.2.
3.3. Cytokine assay
Prior to and 9 weeks post challenge, 3 mice from each group weresacrificed and lymph nodes were excised, homogenized and washed.Lymph node cells were plated in triplicate in 96-well microtiter platesin RPMI 5% (RPMI1640 supplementedwith 5% FCS, 1% L-glutamine, 1%
HEPES, 0.1% 2ME, 0.1% gentamicine) as final count of 106 per well andstimulated with 10 μg of the corresponding recombinant proteins(rCPA, rCPB, rCPC), SLA (soluble Leishmania antigen) or Concanvalin A(ConA), or medium alone. Cell culture plates were incubated for5 days at 37 °C in 5% CO2 humidified atmosphere. Production of IFN-γand IL-5 were measured in supernatant of stimulated lymph nodeculture by sandwich-based Elisa kits (R&D, Minneapolis, MN, USA)according to manufacturer's instructions. The lower detection limitsof IFN-γ and IL-5 were 2 and 7 pg/ml, respectively. All tests wereperformed in triplicate.
3.4. Determination of IgG1 and IgG2a antibodies
Pooled sera were prepared from each group of mice before, and9 weeks after challenge. Production of IgG1 and IgG2a antigen specificantibodies was measured against SLA, rCPA, rCPB and rCPC. Briefly96-well plates were coated with recombinant proteins or SLA antigen(10 μg/ml in PBS) and incubated at 4 °C overnight. After 3 washeswithwashing buffer (PBS plus 0.05% Tween 20), plates were blocked withblocking buffer (1% BSA in PBS) for 2 h at 37 °C then 100 μl of pooledsera were added and incubated for another 2 h at 37 °C (1:50dilution). Biotinylated anti mouse IgG1 (1:10,000, Zymed, Burlington,ON, Canada) and IgG2a (1:500, southern Biotech Birmingham, USA)were added and incubated for 2 h at 37 °C. Streptavidine-horseradishperoxidase (BRL, Gaithersburg, MD) was added (1:1000) andincubated for 1 h at 37 °C. Conjugates' binding were visualized withO-phenylenediamine (OPD) and the reaction was stopped by sulfuricacid (4 N) and the absorbance was measured at 492 nm.
4. Statistics
Statistics were performed using Graph-Pad Prism 5.0 for Windows(Graphpad Software Inc 2007, San Diego, Calif., USA). All the data wereanalyzed with one way ANOVA (Multiple-comparison Tukey posttest) when required, with the exception of size and zeta potentialmeasurements, which were analyzed with a Student's t-test. Ap-value of≤0.05 was considered significant.
5. Results
5.1. Physicochemical characteristics of the vaccine formulation
Formulations a, b and c were prepared by DNA adsorption on thesurface of cSLNs via direct complexation of each pDNA with cSLNs.Formulations were characterized according to their size and zetapotential and poly dispersity index (Table 1). The results indicatedthat formulations displayed an average size of 244±12, 250±15 and240±12 nm, respectively with no significant (pN0.05) differencebetween the sizes. The observed zeta potential revealed all theformulations are cationic (+22 to +27 mV).
Gel retardation assay for SLN-pDNAs confirmed complete com-plexation between pDNAS and cSLN at a DOTAP:pDNA:ratio of 6:1.Payloaded pDNAs in these formulations were completely protectedfrom DNase I digestion [13]. There was no sign of acute toxicityfollowing administration of these formulations to the mice.
0
20
40
60C
Different groups
GFP
exp
ress
ion
%
1 2 3 4 5 6 7 8 9 10 11 120
1
2
3
4
5
6
G1: SLN-pcDNA-cp/a/b/c
G2: pcDNA-cpa/b/c
G3: SLN
G4: SLN-pcDNA
G5: PBS
A
Weeks after challenge
foot
pad
thic
keni
ng (
mm
)
G1 G2 G3 G4 G5
G1 G2 G3 G4 G5
1.3×10 05
2.6×10 05
5.2×10 05
1.0×10 06
2.1×10 06
4.2×10 06
8.4×10 06
1.7×10 07
3.4×10 07B
Different groups
foot
pad
GFP
exp
ress
ion
net i
nten
sity
(pi
xel)
D
FL1
-
Gate: R1
FL1
-
%87.6:2Q%22.0:1Q
%71.67:4Q%48.61:3Q
Gate: R1%79.21:2Q%93.0:1Q
%58.67:4Q%08.9:3Q
Gate: R1%87.04:2Q%00.0:1Q
%22.95:4Q%00.0:3Q
Gate: R1%52.53:2Q%00.0:1Q
%57.46:4Q%00.0:3Q
Gate: R1%24.64:2Q%00.0:1Q
%85.35:4Q%00.0:3Q
G1 G2 G3 G4 G5
1
10
100
1000
0.10 100 200 300 400 500
FSC-
0 100 200 300 400 5000.1
1
10
100
1000
FSC -
FL
1 -
0.10 100 200 300 400 500
1
10
100
1000
FSC -
FL1
-
0 100 200 300 400 5000.1
1
10
100
1000
FSC -
FL1
-
0 100 200 300 400 5000.1
1
10
100
1000
FSC -
FL1
-
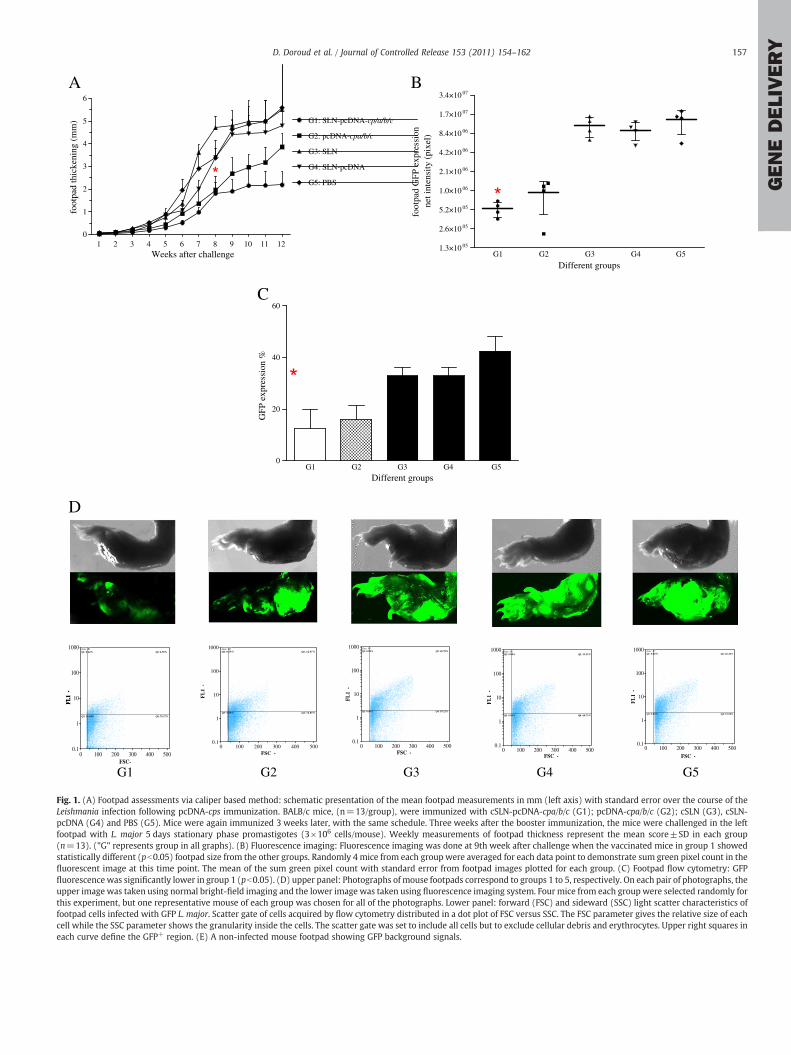
Fig. 1. (A) Footpad assessments via caliper based method: schematic presentation of the mean footpad measurements in mm (left axis) with standard error over the course of theLeishmania infection following pcDNA-cps immunization. BALB/c mice, (n=13/group), were immunized with cSLN-pcDNA-cpa/b/c (G1); pcDNA-cpa/b/c (G2); cSLN (G3), cSLN-pcDNA (G4) and PBS (G5). Mice were again immunized 3 weeks later, with the same schedule. Three weeks after the booster immunization, the mice were challenged in the leftfootpad with L. major 5 days stationary phase promastigotes (3×106 cells/mouse). Weekly measurements of footpad thickness represent the mean score±SD in each group(n=13). ("G" represents group in all graphs). (B) Fluorescence imaging: Fluorescence imaging was done at 9th week after challenge when the vaccinated mice in group 1 showedstatistically different (pb0.05) footpad size from the other groups. Randomly 4 mice from each group were averaged for each data point to demonstrate sum green pixel count in thefluorescent image at this time point. The mean of the sum green pixel count with standard error from footpad images plotted for each group. (C) Footpad flow cytometry: GFPfluorescence was significantly lower in group 1 (pb0.05). (D) upper panel: Photographs ofmouse footpads correspond to groups 1 to 5, respectively. On each pair of photographs, theupper image was taken using normal bright-field imaging and the lower image was taken using fluorescence imaging system. Four mice from each group were selected randomly forthis experiment, but one representative mouse of each group was chosen for all of the photographs. Lower panel: forward (FSC) and sideward (SSC) light scatter characteristics offootpad cells infected with GFP L. major. Scatter gate of cells acquired by flow cytometry distributed in a dot plot of FSC versus SSC. The FSC parameter gives the relative size of eachcell while the SSC parameter shows the granularity inside the cells. The scatter gate was set to include all cells but to exclude cellular debris and erythrocytes. Upper right squares ineach curve define the GFP+ region. (E) A non-infected mouse footpad showing GFP background signals.
157D. Doroud et al. / Journal of Controlled Release 153 (2011) 154–162
GENEDELIVERY

G1 G2 G3 G4 G50
5
10
15
20
*
A
Different groups
para
site
bur
den
of th
e ly
mph
nod
es
B
G1 G2 G3 G4 G50
20
40
60
80
100
**
GF
P e
xpre
ssio
n %
Fig. 2. Lymph nodes parasite quantification by microtitration analysis (A) and flowcytometry assay (B). Lymphatic cells were cultured in duplicate for 10 days at 26 °C inserial ten-fold dilutions. Results are expressed as mean±SD of three lymph nodes pergroup. "*", "**" reveal significant decrease in group 1 compared to unvaccinated controlgroups (pb0.05, pb0.005, respectively).
Table 2Parasite inhibition measured in terms of the decrease in intensity of green fluorescencein the footpad and lymph node of vaccinated animals by flow cytometry.
Vaccinated group Parasite Inhibition
Measured basedon footpad
Measured based onlymph node
cSLN-pcDNA-cpa/b/c 65.44% 86.3%pcDNA-cpa/b/c 55.78% 54.36%
158 D. Doroud et al. / Journal of Controlled Release 153 (2011) 154–162
GENEDELIVERY
5.2. SLN-pcDNA-cpa/b/c and pcDNA-cpa/b/c vaccination efficacy accordingto disease progression
The efficacy of SLN-pcDNA-cpa/b/c and pcDNA-cpa/b/c was evalu-ated by their capability to induce protection and specific immuneresponse to Leishmania infection. Footpad swelling of different groupsof mice was measured after challenge with EGFP-transfected L. major(Fig. 1A). Control groups showed significant lesions 7 weeks post-challenge. There was a significant difference between animalsreceiving SLN-pcDNA-cpa/b/c and pcDNA-cpa/b/c cocktail vaccineconfirming that the higher level of protection observed in group 1was due to cSLNs-pDNA formulation. This difference was significant(pb0.05) at the 9th week post-challenge. Vaccination with SLN-pcDNA-cpa/b/c and pcDNA-cpa/b/c delayed footpad swelling whencompared to control groups at this time point. For a certain time,immunization with pcDNA-cpa/b/c had also a significant effect indelaying footpad swelling. However this effect did not last following9 weeks after challenge. Vaccination with SLN-pcDNA-cpa/b/c andpcDNA-cpa/b/c (groups 1, 2) controlled the infection compared tocontrol groups. All the animals in group 1 developed significantlysmaller lesions at week 9.
To assess the reliability of this experiment at 9th week post-challenge, EGFP-expressing L. majorwas detected in mice footpads viafluorescence imaging-system that gives a precise two-dimensionalimage of the extent of infection. This enabled us to clearly detectparasite load in the infected footpads (Fig. 1B, D). As shown in Fig. 1D,in the vaccinatedmice GFP-fluorescence wasmore localized at the siteof inoculation compared to control animals in which parasites spreadto wider areas. Sum green intensity (pixel) from the imaging studieswere significantly higher in infected footpads of the control groups(pb0.005, Fig. 1B). Furthermore, group 1 of the tested animalsrevealed significantly lower pixel count compared to group 2 (pb0.05,Fig. 1B). The increasing thickness of the infected footpads was notcorrelated with the intensity of the detected GFPs in the testedanimals (Fig. 1D).
EGFP expression in L. major amastigotes resident in the footpadswas monitored by flow cytometry. High expression levels wereobserved in group 5 (Fig. 1C). The frequency of EGFP positive cells offootpads determined by fluorescence activated cell sorting (FACS)analysis using the appropriate gating are shown in Fig. 1C. Group 1had the lowest infection rate (12.50%) compared to group 2 (15.99%),3 and 4 (32.98%) and 5 (42.53%). Therefore detecting L. major directlyin the footpads could precisely give an idea about the parasite loadand thus vaccine efficacy.
Infection progression was also followed by determining totalparasite load in LNs of challenged mice at 9 weeks post-infection bythe microtitration method (Fig. 2A) and flow cytometry (Fig. 2B).Compared to that in control groups, LNs parasite load in vaccinatedgroups showed significant (pb0.05) decrease at 9 weeks afterchallenge. Although, parasite load in group 2 showed to besignificantly lower than in both control groups (pb0.05, Fig. 2A),there was nearly 2-fold decrease in group 1 compared to group 2.EGFP expression in 10−3 dilutionwas readily evident from the intensegreen fluorescence of the parasites (Fig. 2B). As shown in Fig. 2B,group 1 expressed significantly lower percent of EGFP that iscorrelated with the least promastigote parasite presence in the LNs.
It is noteworthy that in all groups, the rate of infection was inconcordance with the delay in the appearance of lesions andthickening of the footpads as well as the amount of parasite load inthe LNs and footpads that was determined by flow cytometry, imagingand microtitration methods.
The ability of the formulations to inhibit infection progress andparasite load (parasite inhibition percent or PI%) was predictedaccording to flow cytometric methods. These rates were calculated bycomparing the intensity of fluorescence in the footpads and LN ofvaccinated and non-vaccinated groups (Table 2). The results revealed
that vaccination with the SLN-pcDNA-cpa/b/c significantly (pb0.05)prevented leishmaniasis progression in mice (86.3%) compared topcDNA-cpa/b/c (54.36%).
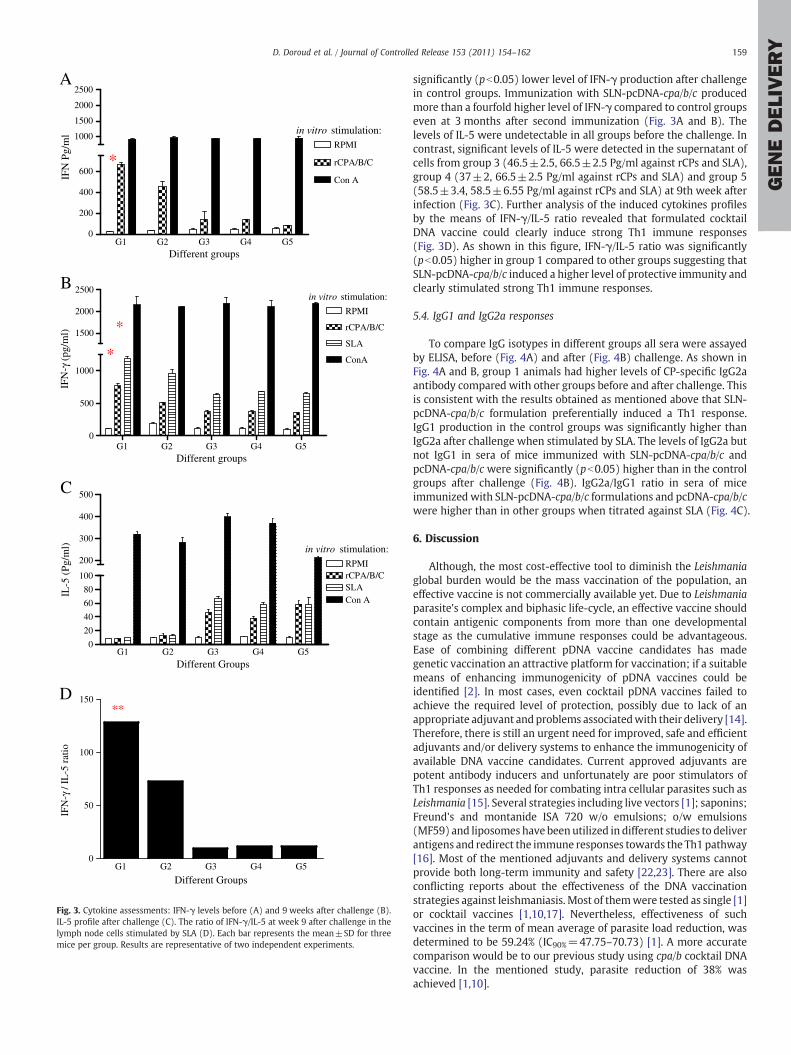
5.3. SLN-pcDNA-cpa/b/c enhances IFN-γ production
In order to compare the induced immune responses by vaccinationregimens and protective effects against L. major challenge, IFN-γ andIL-5 production were assessed before and after challenge. Accordingto IFN-γ production, cells from mice of groups 1 and 2 produced highamounts of IFN-γ before (668±14.7 and 452±39 Pg/ml, respective-ly) and after infection (766±28 and 507±7.9 Pg/ml) in response torCPS and SLA (1192±22 and 953±45 Pg/ml), respectively (Fig. 3Aand B). The significant difference in IFN-γ production could be one ofthe major explanations for the disease attenuation in group 1 at 9thweek following vaccination. Animals in group 2 showed lower level ofIFN-γ production against rCPs and SLA before and after infectiontherefore protection was not achieved as much as for mice in group 1.Low IFN-γ productionwas detected in supernatants of LN cells of micein all of the three control groups before challenge. Also there was
0
200
400
600
1000
1500
2000
2500
RPMI
rCPA/B/C
Con A
A
*
in vitro stimulation:
Different groups
IFN
Pg/
ml
G1 G2 G3 G4 G5
G1 G2 G3 G4 G5
G1 G2 G3 G4 G5
0
500
1000
1500
2000
2500
RPMI
rCPA/B/C
SLA
ConA
B
*
*
in vitro stimulation:
Different groups
IFN
-γ (
pg/m
l)
0
20
40
60
80
100
200
300
400
500
RPMIrCPA/B/C
Con ASLA
in vitro stimulation:
C
Different Groups
IL-5
(Pg
/ml)
G1 G2 G3 G4 G5
0
50
100
150**
D
Different Groups
IFN
-γ /
IL-5
rat
io
Fig. 3. Cytokine assessments: IFN-γ levels before (A) and 9 weeks after challenge (B).IL-5 profile after challenge (C). The ratio of IFN-γ/IL-5 at week 9 after challenge in thelymph node cells stimulated by SLA (D). Each bar represents the mean±SD for threemice per group. Results are representative of two independent experiments.
159D. Doroud et al. / Journal of Controlled Release 153 (2011) 154–162
GENEDELIVERY
significantly (pb0.05) lower level of IFN-γ production after challengein control groups. Immunization with SLN-pcDNA-cpa/b/c producedmore than a fourfold higher level of IFN-γ compared to control groupseven at 3 months after second immunization (Fig. 3A and B). Thelevels of IL-5 were undetectable in all groups before the challenge. Incontrast, significant levels of IL-5 were detected in the supernatant ofcells from group 3 (46.5±2.5, 66.5±2.5 Pg/ml against rCPs and SLA),group 4 (37±2, 66.5±2.5 Pg/ml against rCPs and SLA) and group 5(58.5±3.4, 58.5±6.55 Pg/ml against rCPs and SLA) at 9th week afterinfection (Fig. 3C). Further analysis of the induced cytokines profilesby the means of IFN-γ/IL-5 ratio revealed that formulated cocktailDNA vaccine could clearly induce strong Th1 immune responses(Fig. 3D). As shown in this figure, IFN-γ/IL-5 ratio was significantly(pb0.05) higher in group 1 compared to other groups suggesting thatSLN-pcDNA-cpa/b/c induced a higher level of protective immunity andclearly stimulated strong Th1 immune responses.
5.4. IgG1 and IgG2a responses
To compare IgG isotypes in different groups all sera were assayedby ELISA, before (Fig. 4A) and after (Fig. 4B) challenge. As shown inFig. 4A and B, group 1 animals had higher levels of CP-specific IgG2aantibody compared with other groups before and after challenge. Thisis consistent with the results obtained as mentioned above that SLN-pcDNA-cpa/b/c formulation preferentially induced a Th1 response.IgG1 production in the control groups was significantly higher thanIgG2a after challenge when stimulated by SLA. The levels of IgG2a butnot IgG1 in sera of mice immunized with SLN-pcDNA-cpa/b/c andpcDNA-cpa/b/c were significantly (pb0.05) higher than in the controlgroups after challenge (Fig. 4B). IgG2a/IgG1 ratio in sera of miceimmunized with SLN-pcDNA-cpa/b/c formulations and pcDNA-cpa/b/cwere higher than in other groups when titrated against SLA (Fig. 4C).
6. Discussion
Although, the most cost-effective tool to diminish the Leishmaniaglobal burden would be the mass vaccination of the population, aneffective vaccine is not commercially available yet. Due to Leishmaniaparasite's complex and biphasic life-cycle, an effective vaccine shouldcontain antigenic components from more than one developmentalstage as the cumulative immune responses could be advantageous.Ease of combining different pDNA vaccine candidates has madegenetic vaccination an attractive platform for vaccination; if a suitablemeans of enhancing immunogenicity of pDNA vaccines could beidentified [2]. In most cases, even cocktail pDNA vaccines failed toachieve the required level of protection, possibly due to lack of anappropriate adjuvant andproblems associatedwith their delivery [14].Therefore, there is still an urgent need for improved, safe and efficientadjuvants and/or delivery systems to enhance the immunogenicity ofavailable DNA vaccine candidates. Current approved adjuvants arepotent antibody inducers and unfortunately are poor stimulators ofTh1 responses as needed for combating intra cellular parasites such asLeishmania [15]. Several strategies including live vectors [1]; saponins;Freund's and montanide ISA 720 w/o emulsions; o/w emulsions(MF59) and liposomes have been utilized in different studies to deliverantigens and redirect the immune responses towards the Th1 pathway[16]. Most of the mentioned adjuvants and delivery systems cannotprovide both long-term immunity and safety [22,23]. There are alsoconflicting reports about the effectiveness of the DNA vaccinationstrategies against leishmaniasis. Most of themwere tested as single [1]or cocktail vaccines [1,10,17]. Nevertheless, effectiveness of suchvaccines in the term of mean average of parasite load reduction, wasdetermined to be 59.24% (IC90%=47.75–70.73) [1]. A more accuratecomparison would be to our previous study using cpa/b cocktail DNAvaccine. In the mentioned study, parasite reduction of 38% wasachieved [1,10].

A B C A B C A B C A B C A B C0
1
2
3
IgG1
IgG2a
A: rCPAB: rCPBC: rCPC
A
G1 G2 G3 G4 G5
G1 G2 G3 G4 G5
O. D
. 492
nm
A B C S A B C S A B C S A B C S A B C S0
1
2
3
4
5IgG1
IgG2a
A: rCPAB: rCPBC: rCPCS: SLA
B
O. D
. 492
nm
G1 G2 G3 G4 G50.0
0.5
1.0
1.5
Different Groups
IgG
2a/I
gG1
Rat
io
C
Fig. 4. CP-specific IgG1 and IgG2a antibodies sub classes, induced by immunization withcps DNA with or without SLN in the vaccinated and control groups. Mice were bled atweek 9 after challenge and pooled sera/groupwas analyzed for presence of CPs-reactiveIgG1 and IgG2a. Shown data represent mean±SD for thirteen (n=13) mice in eachgroup. Results are representative of two independent experiments, each performed intriplicate. Antibody responses before (A) and after (B) challenge at 1:100 dilutions. Anaverage of duplicate absorbance values of each pooled sera sample was recorded. Theratio of IgG2a/IgG2a at 8 weeks after challenge in the different groups determinedagainst SLA (C).
160 D. Doroud et al. / Journal of Controlled Release 153 (2011) 154–162
GENEDELIVERY
These relatively low rates could be due to the reason that deliverysystems were rarely used in the formulation of anti leishmaniasis DNAvaccine candidates [1]. Therefore, in favor of such DNA vaccineapproaches; it is crucial to develop a potentiated delivery systemtoward Th1 responses. To address this unmet need, we have previouslydeveloped anovel cationic SLN-basedDNAvaccinedelivery system [13].The rationale behind this work was to create a delivery system topromote pDNA payload condensation and protection against extracel-lular degradation by DNAses [13] and lysosomes which diminish pDNAexpression and mammalian cells transfection enhancement [13]. Inregard to increasing the magnitude or duration of local pDNA targetingto antigen presenting cells (APCs) such as dendritic cells (DCs, tomodulate cellular and humoral immune responses) it is potentially
useful to have an effective adjuvant for developing vaccines againstleishmaniasis.
Optimal vaccine formulations are expected to contain promisingantigen(s) as well as immunostimulatory (IS) adjuvants and a suitablesystem to deliver both antigen and adjuvant. cSLN as a delivery system(DS) offers advantages including safety, storage stability, relativelyeasy large scale production, steam sterilization and lyophilizationpossibility [12,18].
Previously, we have demonstrated that co-immunization of cpa/bgenes induced a partial protective response whereas the immuniza-tion with separate mentioned genes was not fully protective. Inaddition, immunization of BALB/c mice with combination of DNA andprotein (prime-boost) was more effective than DNA immunization[8–10]. However, complex logistics and high cost associated withmanufacturing a second vaccine platform is the main drawback forthe feasibility of this kind of vaccination strategy especially indeveloping countries. Therefore, we used cSLN for pDNA delivery tooptimize immune response priming that could potentially eliminatethe need for rprotein booster injection. cSLNs have been used aspDNAs delivery systems in vaccination studies and significantenhancement for both T-helper types of immune responses, reported.As against the Freund's incomplete adjuvant (FIA), SLN formulationswere well tolerated and exhibited higher adjuvant activity when theirsizes were greater than 100 nm [18]. J.M. Brewer et al. also reportedthat lipid vesicles with a mean diameter N225 nm preferentiallyinduces Th1 responses in BALB/c mice [21].
Therefore, it seems that two major design parameters havesignificant importance for developing DSs. These include size andcomposition ofmaterials used in the fabrication ofDS. Thefinecontrol ofthese parameters allows the exploitation of the beneficial aspects ofmononuclear phagocytic system (MPS) cell targeting leading to inducedinnate immunity.
In this study, the pcDNA-cps were formulated into cationic lipidparticles with nanometer range (~240–250 nm). It is expected thatthe particulate formulation in this size range facilitates the uptake bydendritic cells and macrophages, thereby enhancing the expression,processing and presentation of the payloaded antigens and theseresult in the stronger immune effects, as observed in this study.Besides the size parameter, facilitating intracellular delivery could bealso explained by the vigilant selection of the formulation composi-tion. The presence of cholesterol domains in cSLN might enhancetransfection efficiency by facilitating membrane fusion [13,22]. Inaddition, utilization of Tween 80 in the formulation could alsoenhance SLN-pDNAs phagocytosis by APC as well as localization indraining LNs following subcutaneous administration [23]. Since LNscontain a greater number of cells that express toll-like receptor 9(TLR-9), localization of pDNAs in the draining LNs possibly will be animportant mechanism by which cSLN formulation can enhance theimmunological activity of pDNAs. It is worth mentioning that theplasmids used to deliver these DNA immunogens has also the intrinsicability to induce the expression of co-stimulatory molecules on APCs,probably because the mentioned vaccine plasmids contain CpGmotifsthat signal through TLR-9 to activate APCs. Therefore, pcDNA-cpscould have intrinsic adjuvant properties due to the presence of theseCpG motifs, known as IS sequences. This is well demonstrated bycomparing the humoral and cellular responses acquired from G1 andG2. These results further support that CpG motif could not beconsidered as a standalone immune response modifier and therefore,needs a DS to enhance subsequent innate immune responses that canprovide generalized protection against Leishmaniasis. Nevertheless,stimulation of TLR-9, signaling pathways and potential adjuvanteffects of these SLN based formulations should be further tested by theassessment of the generation of proinflammatory cytokines (i.e. IL-6)and/or type I IFNs (i.e. IFN-α).
Another important aspect of this DS might be the presence ofDOTAP in particulate formulation. This cationic surface active lipid is

161D. Doroud et al. / Journal of Controlled Release 153 (2011) 154–162
GENEDELIVERY
able to activate the dendritic cells through a common binding partnerwith lipo polysaccharide (LPS) [24]. It also stimulates DC maturationand induces down-regulation of IL-1 signal that could suppress theTh2 response. Therefore, DOTAP is considered as an active stimulatorfor DC and poses known adjuvant activity [25]. Utilizing such a DCstimulator in pDNA vaccine formulation, suggests cSLN might not bejust a carrier for pDNA delivery into cells, but can modify cellularpathways and stimulate immune responses.
In this experiment, we utilized GFP-transfected Leishmania forgenerating experimental cutaneous leishmaniasis in BALB/c micewhich was reported as a novel tool allowing visualization andcorrelation of fluorescence intensity with parasite burden [19]. Thisnovel tool has been also developed in our laboratory, recently [20].The results of footpad fluorescence measurements by imaging andflow cytometry techniques correlate with LN parasite burden results.These methods were more perceptive, precise and less time-consuming than the standard caliper-based method for followingLeishmania infection progress in vivo.
To evaluate the protection rate, footpad swelling after challenge inimmunized mice was compared with control groups. The resultsdemonstrated that lesion size in mice immunized with SLN-pcDNA-cpa/b/c at week 9 post challenge was significantly (pb0.05) smallercompared to that in other groups, confirming the adjuvant effect of cSLNfor this genetic vaccination (Fig. 1A, B and C). As another indication ofthe protection rate, number of viable L. major parasites was quantitatedin the LNs of all the groups after challenge. Again, lowest number of liveparasites was seen in this group which was comparable to the footpadswelling results. It is worth mentioning that among the differentmethods performed on both footpads and lymph nodes of the testedmice for parasite inhibition assessments, only footpad flowcytometryassessmentdidn't showa significantdifference betweenG1 andG2. Thismight be due to the fact that footpad samples for flow cytometryanalysis could not be efficiently homogenized with the similar standardoperating procedures for all the groups, as we have recently described[26]. Therefore, the footpad results might not be sufficiently accurateand, in practice, the lymph node evaluation for parasite burdenassessments is more accurate and reproducible [26]. Despite thisexceptional result, it seems that pcDNA-cpa/b/c induced lower levelsof the protection and needs a suitable delivery system to enhance itsimmunoprotective activity. The cytokines, produced by antigen-specificT-cells, determined the profile of an elicited antibody response; IL-5 isassociated with high levels of IgG1, whereas production of IgG2a isdependent on IFN-γ. Our results demonstrated that the immuneresponse elicited by SLN-pcDNA-cpa/b/c was a dominant Th1 responsedenoted by the production of IFN-γ. The IFN-γ/IL-5 ratio afterstimulation with SLA antigen for group 1, 2 was 127.94 and 72.63,respectively. This ratio is 1.76 fold higher when SLN has been used as adelivery system and preceded as a Th1 stimulator adjuvant andtherefore up-regulated the vaccine efficiency in elicitation of protectiveresponses against leishmaniasis.
IgG1 and IgG2a antibody titers were also used as an indicator ofTh2 and Th1 immune responses, respectively. Significantly (pb0.05),the highest IgG2a was seen in group 1 both before and after challengecompared with other groups. This might be an indication that cSLN-pcDNA formulation induces a more potent durable immune responsecompared to unformulated pDNAs. The enhanced ratio of IgG2a/IgG1in mice particularly after challenge was correlated with a Th1response and further confirmed that SLN-pcDNA-cpa/b/c formulationcould induce a potent Th1 immune response and protection againstleishmaniasis at least in mice model.
Therefore, due to the careful selection of the composition, thisvaccine formulation could not only proceed as a delivery system dueto the particulate nature of cSLN promoting the uptake of associatedcps into the APCs, but also might act as an IS adjuvant for Leishmaniavaccine. In fact, the presence of CpGmotifs in the pDNA backbone andDOTAP in cSLN formulation, resulting in stimulating immune cells in
specific way. As a result of enhancing immune responses, the percentof parasite inhibition by pDNA vaccination technology could beincreased considerably from 38% [1] to 86.3% according to the LNparasite burden determination by precise technique such as flowcytometry, as discussed previously.
7. Conclusion
Vaccine formulations are often needed to have particulate naturewith suitable dimensions for stimulating the immune system,appropriately. However, up to now, only a few adjuvants and/ordelivery systems have been approved for routine and clinical use.Therefore, there is an urgent medical need for effective and safeadjuvant development capable of stimulating specific immunity.cSLNs are promising and adaptable delivery systems that can bemodified rationally towards specific vaccine targets by choosing theirsize and composition, accurately. In this work, we have primarilyassessed the possibility of cSLN utilization as an adjuvant (DS) forfuture vaccine development against leishmaniasis to improve inducedimmunogenicity. Simplicity, biocompatibility and scale up possibilityof the cSLN manufacturing process together with the appropriateimmunostimulatory effects of the formulation, due to the presence ofCpG motifs in pCDNA backbone and DOTAP in cSLN structure,suggested that this delivery system might be utilized to createstronger protective vaccines in combination with Leishmania cps. Thecurrent data, in murinemodel of L. major infection, showed promisingrole of cSLN as an adjuvant to enhance stronger immune responseagainst this infection. The mean average of parasite load reduction forDNA vaccination regimens is determined to be 59.24% (IC 95%: 47.75–70.73) [1]. Herein for the first time, we report that the percentage ofparasite inhibition by DNA vaccination technology could be increasedup to 86.3% according to the precise flow cytometry method ofparasite burden determination. These promising data warrant furtherinvestigations in this regard. Our next step is to consider thecombination of cps types I, II, and III formulated with cSLN as avaccine formulation against L. infantum infection. This work willanswer one of the main challenges to Leishmania vaccine develop-ment which is discovering a vaccine by which cross protectionbetween different Leishmania species can be properly created.
Acknowledgments
D. Doroud thanks Pasteur Institute of Iran and Tehran University ofMedical sciences for supporting her PhD studentship. The authorswish to thank Mr. A. Eravani and Mr. A. Javadi (Pasteur Institute ofIran, Department of Immunology) and also Mr. Sh. Alizadeh for theirtechnical assistance.
References
[1] C.B. Palatnik-de-Sousa, Vaccines for leishmaniasis in the fore coming 25 years,Vaccine 26 (2008) 1709–1724.
[2] J.A. Carvalho, J. Rodgers, J. Atouguia, D.M. Prazeres, G.A. Monteiro, DNA vaccines: arational design against parasitic diseases, Expert Rev. Vaccines 9 (2) (2010)175–191.
[3] G.H. Coombs, J.C. Mottram, Proteinases of trypanosomes and Leishmania,Trypanosomiasis and Leishmaniasis: Biology and Control, CAB International,Oxford, 1997, p. 177.
[4] J.A. Sakanari, S.A. Nadler, V.J. Chan, J.C. Engel, C. Leptak, J. Bouvier, Leishmaniamajor: comparison of the cathepsin L- and B-like cysteine protease genes withthose of other trypanosomatids, Exp. Parasitol. 85 (1997) 63–67.
[5] C.D. Robertson, G.H. Coombs, M.J. North, J.C. Mottram, Parasite cysteine pro-teinases, in: J.H. McKerrow, M.N.G. James (Eds.), Perspectives in Drug Discoveryand Design, 6, ESCOM Science Publisher, 1996, p. 99.
[6] S. Rafati, N. Fasel, S. Masina, Leishmania cysteine proteinases: from gene tosubunit vaccine, Curr. Genomics 4 (2003) 253–261.
[7] N. Khoshgoo, F. Zahedifard, H. Azizi, Y. Taslimi, M. Jiménez Alonso, S. Rafati,Cysteine proteinase type III is protective against Leishmania infantum infection inBALB/c mice and highly antigenic in visceral leishmaniasis in individuals, Vaccine26 (46) (2008) 5822–5829.

162 D. Doroud et al. / Journal of Controlled Release 153 (2011) 154–162
GENEDELIVERY
[8] S. Rafati, A. Nakhaee, T. Taheri, Y. Taslimi, H.Darabi, D. Eravani, Protective vaccinationagainst experimental canine visceral leishmaniasis using a combination of DNA andprotein immunization with cysteine proteinases type I and II of L. infantum, Vaccine23 (2005) 3716–3725.
[9] S. Rafati, F. Zahedifard, F. Nazgouee, Prime-boost vaccination using cysteineproteinases types I and II of Leishmania infantum confers protective immunity inmurine visceral leishmaniasis, Vaccine 24 (2006) 2169–2175.
[10] S. Rafati, A.H. Salmanian, T. Taheri, M. Vafa, N. Fasel, A protective cocktail vaccineagainst murine cutaneous leishmaniasis with DNA encoding cysteine proteinasesof Leishmania major, Vaccine 19 (2001) 3369–3375.
[11] A. Zadeh-Vakili, T. Taheri, Y. Taslimi, F. Doustdari, A.H. Salmanian, S. Rafati,Immunization with the hybrid protein vaccine, consisting of Leishmania majorcysteine proteinases Type I (CPB) and Type II (CPA), partially protects againstleishmaniasis, Vaccine 22 (2004) 1930–1940.
[12] R.H. Muller, K. Mader, S. Gohla, Solid lipid nanoparticles (SLN) for controlled drugdelivery—a review of the state of the art, Eur. J. Pharm. Biopharm. 50 (2000)161–177.
[13] D. Doroud, A. Vatanara, F. Zahedifard, E. Gholami, R. Vahabpour, A.H. RouholaminiNajafabadi, S. Rafati, Cationic solid lipid nanoparticles loaded by cysteine proteinasegenes as a novel anti-leishmaniasis DNA vaccine delivery system: characterizationand in vitro evaluations, J. Pharm. Pharm. Sci. 13 (3) (2010) 320–335.
[14] J.M. Mutiso, J.C. Macharia, M.M. Gicheru, A review of adjuvants for Leishmaniavaccine candidates, J. Biomed. Res. 24 (1) (2010) 16–25.
[15] S.G. Reed, S. Bertholet, R.N. Coler, M. Friede, New horizons in adjuvants for vaccinedevelopment, Trends Immunol. (30) (2009) 23–32.
[16] S. Bhowmick, N. Ali, Recent developments in leishmaniasis vaccine deliverysystems, Expert Opin. Drug Deliv. 5 (7) (2008) 789–803.
[17] S. Ben Hadj Ahmed, L. Touihri, Y. Chtourou, K. Dellagi, C. Bahloul, DNA basedvaccination with a cocktail of plasmids encoding immunodominant Leishmania(Leishmania) major antigens confers full protection in BALB/c mice, Vaccine 27(2009) 99–106.
[18] M.D. Joshi, R.H. Müller, Lipid nanoparticles for parenteral delivery of actives, Eur. J.Pharm. Biopharm. 71 (2009) 161–172.
[19] R.S. Mehta, R. Huang, M. Yang, X.Q. Zhang, B. Kolli, K.P. Chang, R.M. Hoffman, Y.Goto, R. Badaro, R.T. Schooley, Real-time in vivo green fluorescent protein imagingof a murine leishmaniasis model as a new tool for Leishmania vaccine and drugdiscovery, Clin. Vaccine Immunol. 15 (2008) 1764–1770.
[20] A. Bolhassani, T. Taheri, Y. Taslimi, S. Zamanilui, F. Zahedifard, N. Seyed, F.Torkashvand, B. Vaziri, S. Rafati, Fluorescent Leishmania species: Development ofstable GFP expression and its application for in vitro and in vivo studies,Exp. Parasitol. 127 (2011) 637–645.
[21] J.M. Brewer, L. Tetley, J. Richmond, F.Y. Liew, J. Alexander, Cellular immunologyand immune regulation: lipid vesicle size determines the Th1 or Th2 response toentrapped antigen, J. Immunol. 161 (1998) 4000–4007.
[22] L. Xu, T.J. Anchordoquy,Cholesterol domains in cationic lipid/DNAcomplexes improvetransfection, Biochim. Biophys. Acta (BBA) Biomembr. 1778 (10) (2008) 2177–2181.
[23] F. Seeballuck, E. Lawless, M.B. Ashford, C.M. O'Driscoll, Stimulation of triglyceride-rich lipoprotein secretion by polysorbate 80: in vitro and in vivo correlation usingcaco-2 cells and a cannulated rat intestinal lymphatic model, Pharm. Res. 21(2004) 2320–2326.
[24] D.P. Vangasseri, Z. Cui, W. Chen, D.A. Hokey, L.D. Falo, L. Huang, Immunostimulation of dendritic cells by cationic liposomes, Mol. Membr. Biol. 23 (5)(2006) 385–395.
[25] W. Yan, W. Chen, L. Huang, Mechanism of adjuvant activity of cationic liposome:phosphorylation of a MAP kinase, ERK and induction of chemokines, Mol.Immunol. 44 (15) (2007) 3672–3681.
[26] D. Doroud, F. Zahedifard, A. Vatanara, A.R. Najafabadi, S. Rafati, Cysteineproteinase type I, encapsulated in Solid Lipid Nanoparticles induces substantialprotection against L. major infection in C57BL/6 mice, Parasite Immunol. 33 (6)(2011) 335–348.




















