
Childhood Behavior Precursors of Schizotypal Personality ...
Upload
independentCategory
view
0download
0
Delayed Maturation of C D 4 - C D 8 - Fc~/RII/III+ T and Natural Killer Cell Precursors in FceRI~/Transgenic Mice ByV&onique Flamand,* Elizabeth W. Shores,* Tom Tran,a Kun Huang,~ Eric Lee,~ Alexander Grinberg,~ Jean-pierre Kinet,* and Paul E. Love~
From the *Laboratory of MolecularAllergy & Immunology, National h;stitute of Allergy and Infectious Diseases, National h;stitutes of Health, Rockville, Maryland 20852; ~;Division of Hematologic Products, Center for Biologics Evaluation and Research, Food and Dn,g Administration, Bethesda, Maryland 20892; and the ~Laboratory of Mammalian Genes and Development, National Institute of Child Health and Human Development, National Institutes of Health, Bethesda, Maryland 20892
S u m m a r y
FceP,.Iy (y) is a member of a group of related proteins (the ~-family dimers) that function as signal-transducing components of both Fc receptors and the T cell antigen receptor (TC1K). Analysis of y expression during fetal thymus ontogeny revealed that it is expressed in early thy- mocytes, before the initiation of clonotypic TC1K-e¢ and TC1K-]3 gene rearrangement but is down-regulated in most adult thymocytes. To explore a possible role for y in thymocyte de- velopment, we generated transgenic mice in which this protein was overexpressed at all stages of ontogeny. Overexpression of 3' inhibited the maturation of T cells as well as natural killer (NK) cells. The developmental effects were transgene dose related and correlated with mark- edly delayed maturation of fetal C D 4 - C D 8 - FcRII/III + thymocytes, cells thought to include the progenitors of both T and NK cells. These results suggest that the ~ and y chains serve dis- tinctive functions in thymocyte development and indicate that Fc receptor(s) may play an im- portant role in regulating the differentiation of early progenitor cells within the thymus.
S ignals transduced by the T C R regulate the stage-spe- cific development and selection (positive and negative)
of thymocytes (1, 2). The signal-transducing potential of the T C R complex is conferred by multiple subunits (the CD3-y, -8, and-e , and the {, ~1, or Fc~RIy chains) that share a conserved structural element, the immunoreceptor tyrosine-based activation motif (ITAM) 1 (3). A consider- able body of evidence supports the current notion that ITAMs, through their ability to facilitate activation of cyto- plasmic protein tyrosine kinases, mediate all known T C R effector functions (4).
The signals that regulate the earliest stages of thymocyte maturation are less understood. Precursor C D 4 - C D 8 - T C R - thymocytes populate the murine thymus on or about day 12 of gestation (2). These early thymocytes, which are thought to include the precursors of both T cells and NK ceils (5, 6) progress through multiple stages of differentia- tion before expressing CD4, CD8, and 0t/]3TCR (2, 7).
l Abbrevtanons used m this paper ADCC, antibody-dependent cellular cyto- toxlc~ry; DN, double neganve; DP, double posmve; FCM, multicolor flow cytometry: fd, fetal day; "yTg, 2; transgemc, ITAM, mlmunoreceptor Wrosme-based activation mot~f~ poly(IC), polymosimc-polycytidyhc acid.
Although the molecular events that regulate their transit through these various stages remain uncharacterized, recent evidence suggests that they may receive activating signals through cell surface structures (8). The majori W of early fe- tal thymocytes express low affinity receptors for lgG (Fcy- iKII and/or FcylKIlI) (5). FcylKIII is also expressed on ma- ture NK cells where it functions as a signal-transducing complex to mediate antibody-dependent cellular cytotox- icity (ADCC), and on a subset of mature T cells (9). These observations have led to speculation that FcRIII may rep- resent a developmentally important signaling structure in early C D 4 - C D 8 - T C R - thymocytes (5).
In mice, the FcyRIII e~ chain forms a complex with a dimer composed of FcelKIy chains (10, 11). Initially identi- fied as a subunit of the high affinity lgE receptor, FcelKIy is also a subunit of the high affinity IgG receptor (Fc'ylKI), and in a subset of T cells, functions as a subunit of the TClK complex (12-17). The structural and functional simi- larity between Fc~lKIy and ~ suggests that these proteins are members of a family of signal-transducing proteins whose genes were likely generated by duplication (10, 18). Although FcelKIy is reportedly expressed in early thy- mocytes, its potential role in thymocyte development re- mains controversial. The observation that T cell develop- ment is unaffected in mice lacking FcelKI'y argues that its
1725 The Journal of Experimental Medicine • Volume 184 November 1996 1725-1735
on April 20, 2014
jem.rupress.org
Dow
nloaded from
Published November 1, 1996

function is not critical for T cell maturat ion (19). However , recent experiments indicate that Fc~ receptors on fetal thy- rnocytes may transduce developmental ly important signals (20). In this study, we document that F c e R I y is expressed early in fetal thymic ontogeny, before the rearrangement and expression o f the clonotypic TCI~-(x and TCR-[3 chains. To investigate the role o f F c e R I - / i n thymocyte de- velopment , we generated transgenic mice that express ele- vated levels o f FceI~I',/ at all stages o f fetal ontogeny. W e found that overexpression o f FceRI2~ inhibited the matura- t ion o f both T cells and N K cells. The developmental ef- fects were transgene dose related and were correlated with delayed maturation o f fetal F c y R I I / I I I + thymocytes into F c y P d I / I I I - thymocytes. Collectively, these results show that FceRI',/ can regulate the differentiation o f an early progenitor cell populat ion within the thymus.
Materials and Methods
DNA Constructs and Microinjection. A 4.8-kb BclI genormc frag- ment containing the human FceRI',/("/) promoter and coding se- quences (18) was cloned into the BamHI site of plasrnxd CD2- B-N2 (21) after removal of the SalI-BamHI fragment containing the human CD2 promoter and minigene. A 9.8-kb fragment composed of the ",/gene and the human CD2 3-pnme enhancer was then generated by digestion with NotI, separated from vector sequences by gel electrophoresis, punfied as described (22), and used for pronuclear mjecnon. Transgenlc mice were identified by Southern blotting of tail DNA using the human CD2 enhancer sequences as a probe.
Mice. Transgemc mice were generated by injection of zy- gotes obtained from matings of inbred FVB/N mice. Timed mat- ings were performed with either non Tg FVB/N males × non Tg C57BL/6J females or homozygous 2,551Tg FVB/N males × non Tg C57BL/6J females. The first day of mating was desig- nated as day 0.5. FVB/N × C57BL/6J matings were performed because staining with mAbs 2.4G2 (FcyRII/III) and PK136 (NKI.1) were determined to be superior m the C57BL/6J back- ground; however, the developmental effects of the "yTg were Identical regardless of the background.
RNA Extraction and Analysis. Purification of total thymocyte RNA and Northern blot analysis were performed as described previously (22). cDNA probes for TCR-~x (Co~; 23) and TCP,,-[3 (C~; 24) were provided by E. Shevach (National Institutes of Health [NIH]). ~ cDNA was provided by R. Klausner (NIH). Human (18) and murine (10) FceRI 3, cDNAs were isolated as described. Human GAPDH cDNA was purchased from Clontech Laboratones, Inc. (Palo Alto, CA).
Antibodies and Reagents. mAbs used for flow cytometric anal- ysis included: FITC conjugated antl-Thy-l.1 (OX-7), anti-Thy 1.2 (53-2.1), antl-CD4 (RM4.5), anti-TCR-[3 (H57-597), anti- CD8c¢ (53-6.7), anti-CD3e (145-2Cll), anti-CD2 (RNI2.5), anti-CD25 (7D4), and anti-HSA (M1/69); PE-conjugated anti- CD4 (RM4.5), anti-CD8~x (53-6.7), ann-CD3e (145-2Cll), anti-B220 (R, A3-6B2), and anti-NKl.1 (PK136); and biotiny- lated antl-Fc3,RII/III (2.4G2), ann-CD8o~ (53-6.7), antl-CD4 (RM4.5), anti-B220 (RA3-6B2), ann-CD3e (145-2Cll), ann- CD5 (53-7.3). Unconjugated mAb 2.4G2 was additionally used to block nonspecific binding and for immunopreclpitanon. All of the aforementioned antibodies were purchased from PharMingen (San Diego, CA). Streptavxdln-PE (PharMmgen) and streptaw-
din-Red 670 conjugate (GIBCO-BRL, Galthersburg, MD) were used as second step reagents. Bmtin or FITC-conjugated anti- human CD3 (mAb SK7; Becton Dickinson and Co., San Jose, CA) were used as control antibodies at a concentration of 10 Ixl/ 10 6 cells. Anti-FceRI-/was generated m rabbit to a pepnde corre- sponding to amino acids 80-86 of human FceRI'v (25). Rabbit anti-rat sera were purchased from Jackson lmmunoR, esearch Laboratories (West Grove, PA).
MulticolorFlow Cytometry (FCM). Single cell suspensions (106 cells/ml) of thymi or LN were washed twice in PBS containing 0.5% BSA and 0.05% sodium azide before and after each incuba- tion with antibody. Cell suspensions were premcubated with un- conjugated mAb 2.4G2 to inhibit nonspeclfiC staining unless cells were to be stained for Fc3~RII/III. Incubations with labeled mAb (10 txg/ml), streptavidin-PE or streptavidin-Red 670 were per- formed at 4°C for 30 rain. FCM was performed using a FACScan ® flow cytometer (Becton Dickinson Immunocytometry Systems, Mountain View, CA) and data were analyzed using FACScan ® research software. Unless stated otherwise, fluorescence data were collected on 5 × 104 cells, gating on viable cells as assessed by for- ward and side scatter profiles. In Fig. 6, B and C, gates were set on specific thymocyte subpopnlations at the time of collecnon and data were collected on 4-5 × 104 viable cells. Data were &s- played as logarithmic overlay histograms or dot plots.
Protein Analysis. Thymocytes were lysed in buffer (0.15 M NaC1, 50 mM Tris-HC1, pH 7.4, 20 txg/ml leupeptin, 40 Ixg/rnl aprotmin) containing 1% NP-40 (adult thymocytes) or 1% digito- nm (fetal thymocytes), precleared with protein A-Sepharose plus normal rabbit serum, then incubated with protein A-Sepharose plus 10 txl antisera. Immunopreclpitates were washed, solublllzed in loading buffer + 2ME, and resolved by 14% SDS-PAGE. Sep- arated proteins were then transferred to nitrocellulose and blotted with a 1:1,000 dilution of antl-'y. Detection of immunoreactive proteins was done by chermluminescence using the ECL system (Amersham Corp., Arlington Heights, IL).
Cytotoxicity Assays. Mice were injected with 100 Ixg ofpoly- inosmlc-polycytldyhc acid (poly[IC]) (Sigma Chemical Co., St. Louis, MO). Cytotoxicity assays were performed with spleno- cytes obtained 36 h after injectmn. Target cells were labeled with SlCr (Amersham Corp.) at 100 IxCi/106 cells for 1 h at 37°C, washed twine, and used in a standard 6 h S*Cr-release assay with 2,500 target cells/well m 96-well V-bottom plates. Lysis was per- formed in RPMI 1640 medium containing 10% heat-inactivated FCS, 10 mM Hepes, and 500 U/ml human IL-2 (Hoffman- LaRoche, Nutley, NJ). NK activity was evaluated on the NK- sensitive target cell line, YAC-1. ADCC assays were performed essennally as described (5) with the exception that 3 mM rather than 30 mM 2,4,6-tnmtrobenzyl sulfonlc acid was used.
Results
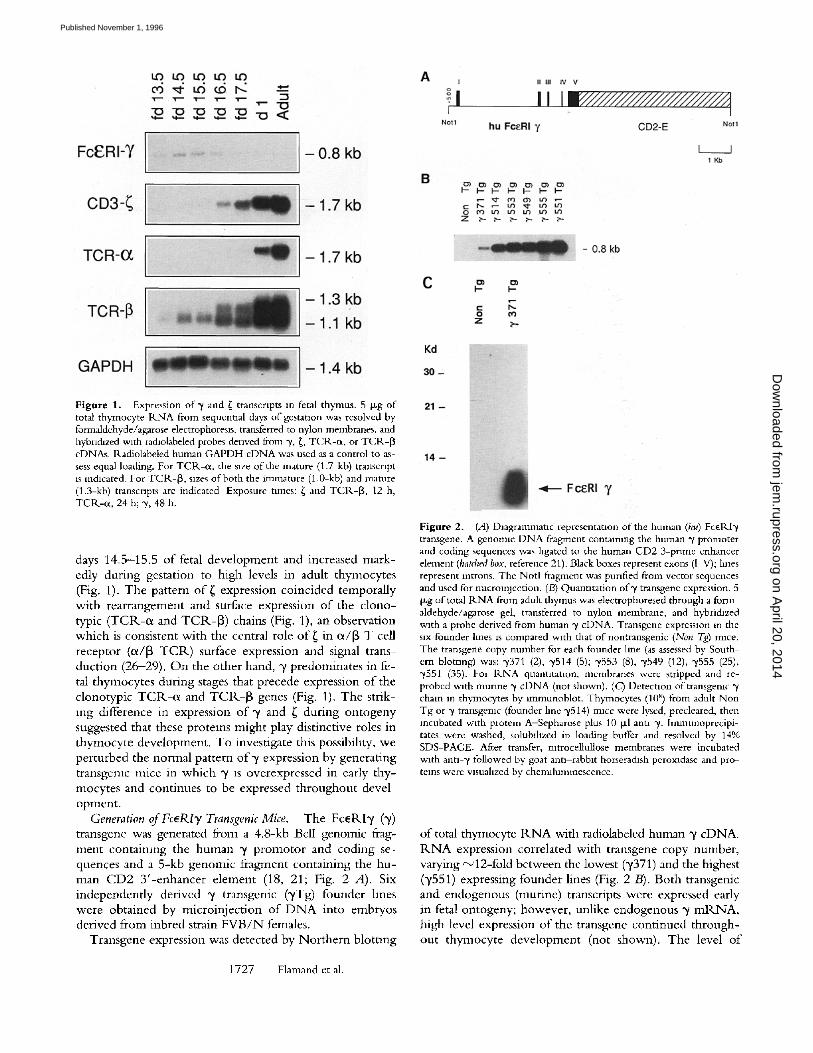
Differential Expression of ~ and FcERI T during Fetal Ontog- eny. To examine the normal pattern o f F c e R I y (~/) ex- pression during ontogeny, Nor thern blot analysis was per- formed on R N A obtained from thymi on sequential days o f fetal development. "/transcripts were present on fetal day 13.5, the earliest stage in which the thymus could clearly be identified in situ (Fig. 1). ~/ m R N A levels were highest at early stages o f development (fetal days 13.5-16.5), de- creased progressively during late gestation, and were barely detectable in adult thymocytes (Fig. 1). In contrast, low levels o f ~ m R N A were first detectable in the thymus on
1726 Delayed T and NK Cell Development in FceRD/Transgenic Mice
on April 20, 2014
jem.rupress.org
Dow
nloaded from
Published November 1, 1996
FcF_,RI-T
LO LO I..O i.O I.O
" - - " O " O - O ' 1 0 " O " O . . . . . " O ':I~
- 0.8 kb
CD3-(~
TCR-O(,
TCR-~
G A P D H
l - 1.7 kb
, ,e
I - 1 . 3 k b
1.1 kb
- 1.4 kb
Figure 1. Expression of 3' and ~ transcripts in fetal thymus. 5 ~g of total thymocyte R.NA from sequential days of gestation was resolved by formaldehyde/agarose electrophoresls, transferred to nylon membranes, and hybn&zed with racholabeled probes denved fi'om 3', ~, TCR-ct, or TCR-[3 cDNAs. Radiolabeled human GAPDH cDNA was used as a control to as- sess equal loading. For TClk-ot, the size of the mature (1.7-kb) transcript is indicated. For TCR-{3, sizes of both the immature (1.0-kb) and mature (1.3-kb) transcripts are indicated Exposure tunes: ~ and TCR.-[3, 12 h, TCP,-0t, 24 h; ",/, 48 h.
days 14.5-15.5 of fetal development and increased mark- edly during gestation to high levels in adult thymocytes (Fig. 1). The pattern of ~ expression coincided temporally with rearrangement and surface expression of the clono- typic (TCR-ot and TCR-[3) chains (Fig. 1), an observation which is consistent with the central role o f{ in ot/[3 T cell receptor (o~/[3 T C R ) surface expression and signal trans- duction (26-29). O n the other hand, "/predominates in fe- tal thymocytes during stages that precede expression of the clonotypic TCR-ot and TCR-[3 genes (Fig. 1). The strik- ing difference in expression of y and ~ during ontogeny suggested that these proteins might play distinctive roles in thymocyte development. To investigate this possibihty, we perturbed the normal pattern of~/expression by generating transgemc mice in which ~/ is overexpressed in early thy- mocytes and continues to be expressed throughout devel- opment.
Generation of FcERI y Transgenic Mice. The Fc~P, Iy (y) transgene was generated from a 4.8-kb BclI genomic frag- ment contaimng the human y promotor and coding se- quences and a 5-kb genomic fragment containing the hu- man CD2 3 ' -enhancer element (18, 21; Fig. 2 A). Six independently derived y transgenic (yTg) founder lines were obtained by microinjection of D N A into embryos derived from inbred strain FVB/N females.
Transgene expression was detected by Northern blotting
A I o
J Not1
B
C
Kd
3 0 -
2 1 -
t l JII
II hu FceRI y
0 no ~ L~ t~ U~
I- I--
z ~
IV V
GD2-E Not1
I I 1Kb
- 0 .8 kb
1 4 -
F c~.RI T
Figure 2. (A) Diagrammatic representation of the human (hu) FceRI3' transgene. A genomlc DNA fragment containing the human 3" promoter and coding sequences was hgated to the human CD2 3-prime enhancer element (hatched box, reference 21). Black boxes represent exons (l-V); hnes represent lntrons. The NotI fragment was punfied from vector sequences and used for rmcrolnlection. (B) Quanntation ofy transgene expression. 5 ~g of total P,.NA from adult thymus was electrophoresed through a form- aldehyde/agarose gel, transferred to nylon membrane, and hybri&zed with a probe derived from human 31 cDNA. Transgene expression m the six founder hnes is compared with that of nontransgenic (Non Tg) mace. The transgene copy number for each founder hne (as assessed by South- em blotting) was: 3'371 (2), 3"514 (5); "/553 (8), 3"549 (12), 3'555 (25), 3'551 (35). For P, NA quantltatlon, membranes were stripped and re- probed with munne 3' cDNA (not shown). (C) Detecnon oftransgenlc 3' chain m thymocytes by lmmunoblot. Thymocytes (10 s) from adult Non Tg or 3' transgemc (founder hne 3'514) mace were lysed, precleared, then incubated with protein A-Sepharose plus 10 ~1 anti-3'. Inmaunoprecipi- tates were washed, solublhzed in loading buffer and resolved by 14% SDS-PAGE. After transfer, mtrocellullose membranes were incubated with ant1-3' followed by goat ann-rabbit horsera&sh peromdase and pro- reins were visualized by chemdurmnescence.
of total thymocyte R N A with radiolabeled human "y cDNA. R N A expression correlated with transgene copy number, varying ~12-fold between the lowest (y371) and the highest (y551) expressing founder lines (Fig. 2 B). Both transgenic and endogenous (murine) transcripts were expressed early in fetal ontogeny; however, unlike endogenous ~ / m R N A , high level expression of the transgene continued through- out thymocyte development (not shown). The level of
1727 Flamand et al.
on April 20, 2014
jem.rupress.org
Dow
nloaded from
Published November 1, 1996
TOTAL THYMOCYTE NUMBER (x IO -6)
THYMUS LYMPH NODES
122
88
34
16
0o
O
'2"::'.i _ ~ . _ _ ~ . . . . .
3 1
I ""~'-
3>2
• ." ' - r ; .
14 .
" 4 ~ _ 1 2
, ;,rl, I "r v[I~IIF~.~II
8"/
, , , , I , , I I ' , ' " " 1 1 , , , , , , i i , , , , , , . _
i
5 i 7 6
I
1 3 -`~'~ r " ' 7
l '~";';"rl ' '""I' i l,n,, I "b ,.,,..
1 2 4 0
~~~~l; - _ 4 1
• ,, nl.q , • , , , , ,1 • | ~-Ul ., r.,,
1 1 1 1 1 1 1 1 II IIIpIII I I I l l l l I 1 5 I
1 1 1 1 1 1 | I | I | I I I I I I I I I I I I I I
co
O
1 7 " ! ' "
' Ii II I~i , r1,1,,l"'Hqrlnl
~ - ' & e ! ~ ' " " 5 3
i. , , , . . I , , . . 1 , , , , m t l n n n ~
, , if|., I
17
' '""I ' ' '""I ' ' '""
i ,nlq ~""I -' ' nnT
, , , , , , , i
I I I ) 'P l? l I '[ II1111 1 1 I I I I I I I I
5 8 2
f k
5 4 6
, n | , 1 , , l r . r |
i i 5 6 1
i
I
i
Non Tg
7514 Tg
y 549 Tg
y 551 Tg
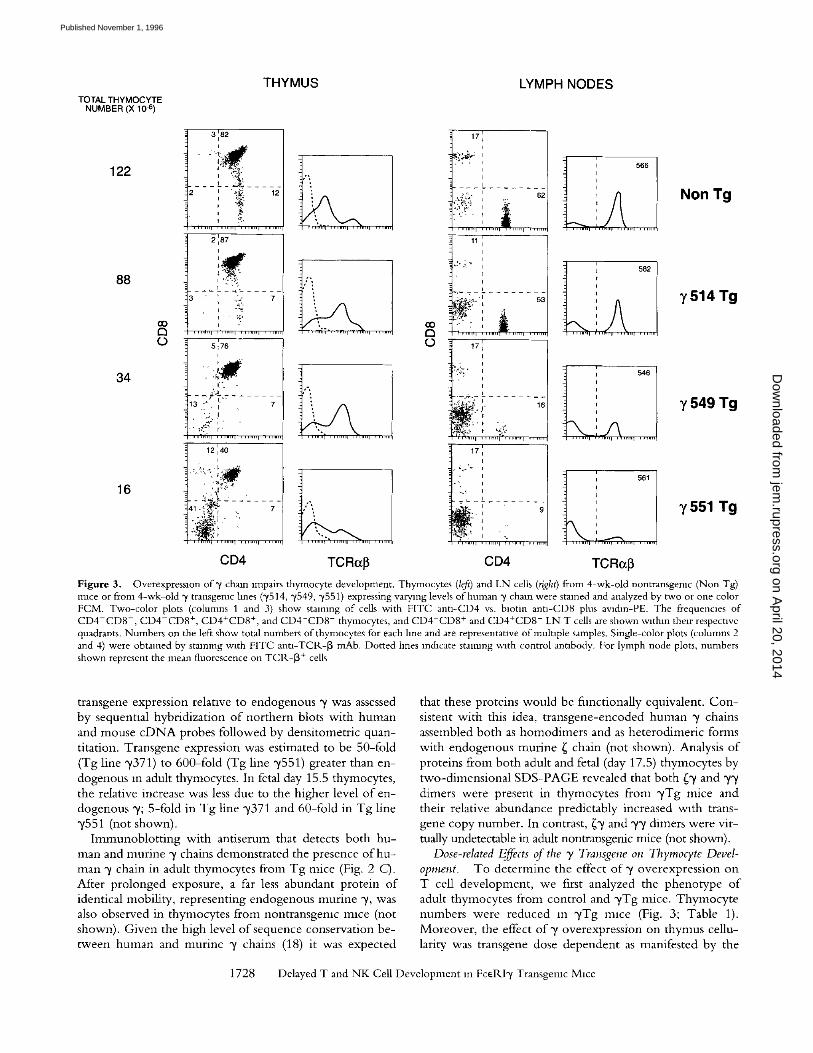
CD4 TCRotI8 CD4 TCRo~I3 Figure 3. Overexpresslon of y chain impairs thymocyte development. Thymocytes (left) and LN cells (right) from 4-wk-old nontransgemc (Non Tg) mice or from 4-wk-old ~/transgemc lines (~/514, y549, ~/551) expressing varymg levels of human 3' chain were stained and analyzed by two or one color FCM. Two-color plots (columns 1 and 3) show staining of cells with FITC ann-CD4 vs. biotin antl-CD8 plus avlchn-PE. The frequencies of CD4-CD8-, CD4-CD8 +, CD4+CD8 +, and CD4+CD8 - thymocytes, and CD4-CD8 + and CD4+CD8 - LN T cells are shown within their respective quadrants. Numbers on the left show total numbers of thymocytes for each line and are representanve of multiple samples. Single-color plots (columns 2 and 4) were obtamed by staining wtth FITC anu-TCP,,-[3 mAb. Dotted hnes indicate staining with control antibody. For lymph node plots, numbers shown represent the mean fluorescence on TCR-I 3+ cells
transgene expression relative to endogenous y was assessed by sequentlai hybridization o f nor thern blots with human and mouse c D N A probes followed by densitometric quan- titation. Transgene expression was estimated to be 50-fold (Tg line y371) to 600-fold (Tg line 3'551) greater than en- dogenous m adult thymocytes. In fetal day 15.5 thymocytes, the relative increase was less due to the higher level o f en- dogenous y; 5-fold in Tg line y371 and 60-fold in Tg line y551 (not shown).
Immunob lo t t i ng with antiserum that detects both hu- man and mur ine 3' chains demonstra ted the presence of hu- man 3' chain in adult thymocytes from Tg mice (Fig. 2 C-). After prolonged exposure, a far less abundant protein of identical mobility, representing endogenous mur ine y, was also observed in thymocytes f rom nontransgemc mice (not shown). Given the high level o f sequence conservation be- tween human and murine y chains (18) it was expected
that these proteins would be functionally equivalent. C o n - sistent with this idea, t ransgene-encoded human 3' chains assembled both as homodimers and as heterodimeric forms with endogenous mur ine { chain (not shown). Analysis o f proteins from both adult and fetal (day 17.5) thymocytes by two-dimensional SDS-PAGE revealed that both {y and yy dimers were present in thymocytes from 3'Tg mice and their relative abundance predictably increased with trans- gene copy number . In contrast, ~3' and y3" dimers were vir- tually undetectable in adult nontransgenic mice (not shown).
Dose-related Effects of the T Transgene on Thymocyte Devel- opment. To de te rmine the effect o f 3" overexpress ion on T cell development , we first analyzed the pheno type o f adult thymocytes from control and 3'Tg mice. T h y m o c y t e numbers were reduced in y T g mice (Fig. 3; Table 1). Moreover , the effect o f y overexpressi'on on thymus cellu- lari W was transgene dose dependent as manifested by the
1728 Delayed T and NK Cell Development in FceRI3, Transgenlc Mice
on April 20, 2014
jem.rupress.org
Dow
nloaded from
Published November 1, 1996
Non Tg ~/514 Tg y551
~>, "E.
-5 (9 O
E . E
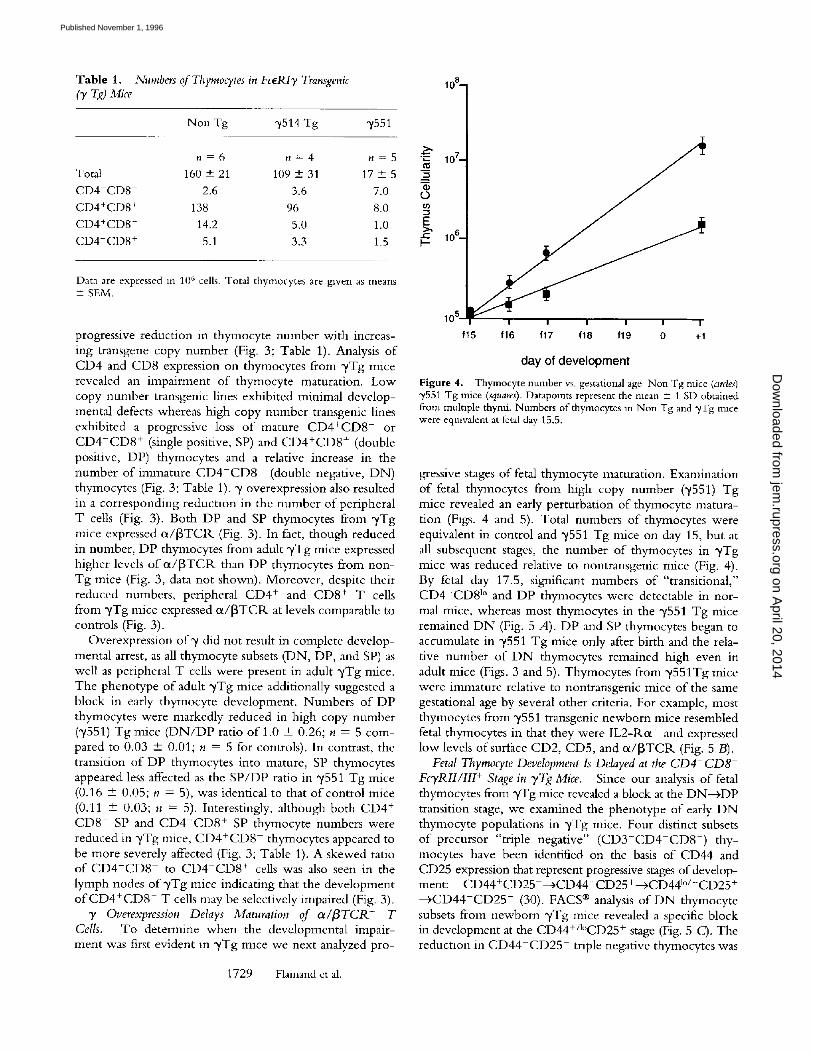
n = 6 n = 4 n = 5 Total 160 + 21 109 + 31 17 + 5 CD4-CD8 2.6 3.6 7.0 CD4+CD8 + 138 96 8.0 CD4+CD8 - 14.2 5.0 1.0 CD4-CD8 + 5.1 3.3 1.5
Data are expressed i n 1 0 6 cells. Total thymocytes are gwen as means + SEM.
progressive reduction in thymocyte number with increas- ing transgene copy number (Fig. 3; Table 1). Analysis o f CD4 and CD8 expression on thymocytes from ~/Tg mice revealed an impairment o f thymocyte maturation. Low copy number transgenic lines exhibited minimal develop- mental defects whereas high copy number transgenic lines exhibited a progressive loss o f mature CD4+CD8 - or C D 4 - C D 8 + (single positive, SP) and C D 4 + C D 8 + (double positive, DP) thymocytes and a relative increase in the number of immature C D 4 - C D 8 - (double negative, DN) thymocytes (Fig. 3; Table 1). y overexpression also resulted in a corresponding reduction in the number o f peripheral T cells (Fig. 3). Both DP and SP thymocytes from y T g mice expressed ot/[3TC1L (Fig. 3). In fact, though reduced in number, DP thymocytes from adult y T g mice expressed higher levels o f ot/[3TClK than DP thymocytes from non- Tg mice (Fig. 3, data not shown). Moreover, despite their reduced numbers, peripheral CD4 + and CD8 + T cells from y T g mice expressed ot/[3TCIL at levels comparable to controls (Fig. 3).
Overexpression of~/d id not result in complete develop- mental arrest, as all thymocyte subsets (DN, DP, and SP) as well as peripheral T cells were present in adult ~Tg mice. The phenotype o f adult y T g mice additionally suggested a block in early thymocyte development. Numbers o f DP thymocytes were markedly reduced in high copy number ("/551) Tg mice ( D N / D P ratio o f 1.0 + 0.26; n = 5 com- pared to 0.03 + 0.01; n = 5 for controls). In contrast, the transition o f DP thymocytes into mature, SP thymocytes appeared less affected as the SP /DP ratio in "y551 Tg mice (0.16 + 0.05; n = 5), was identical to that o f control mice (0.11 --- 0.03; n = 5). Interestingly, although both CD4 + C D 8 - SP and C D 4 - C D 8 + SP thymocyte numbers were reduced in ~Tg mice, C D 4 + C D 8 - thymocytes appeared to be more severely affected (Fig. 3; Table 1). A skewed ratio o f CD4+CD8 to C D 4 - C D 8 + cells was also seen in the lymph nodes o f y T g mice indicating that the development o f C D 4 + C D 8 - T cells may be selectively impaired (Fig. 3).
y Overexpression Delays Maturation of c~/IgTCR + T Cells. To determine when the developmental impair- ment was first evident in "/Tg mice we next analyzed pro-
10 7 _
~o~
10 5 I I I I I I
f15 f16 117 f18 f19 0 +1
Table 1. Numbers of Thymocytes in FcERIT Transgenic 10 8_ ('y Tg) Mice
day of d e v e l o p m e n t
Figure 4. Thymocyte number vs. gestational age Non Tg mice (ardes) "/551 Tg mice (squares). Datapolnts represent the mean + 1 SD obtained from multiple thymi. Numbers of thymocytes m Non Tg and ~/Tg mace were eqmvalent at fetal day 15.5.
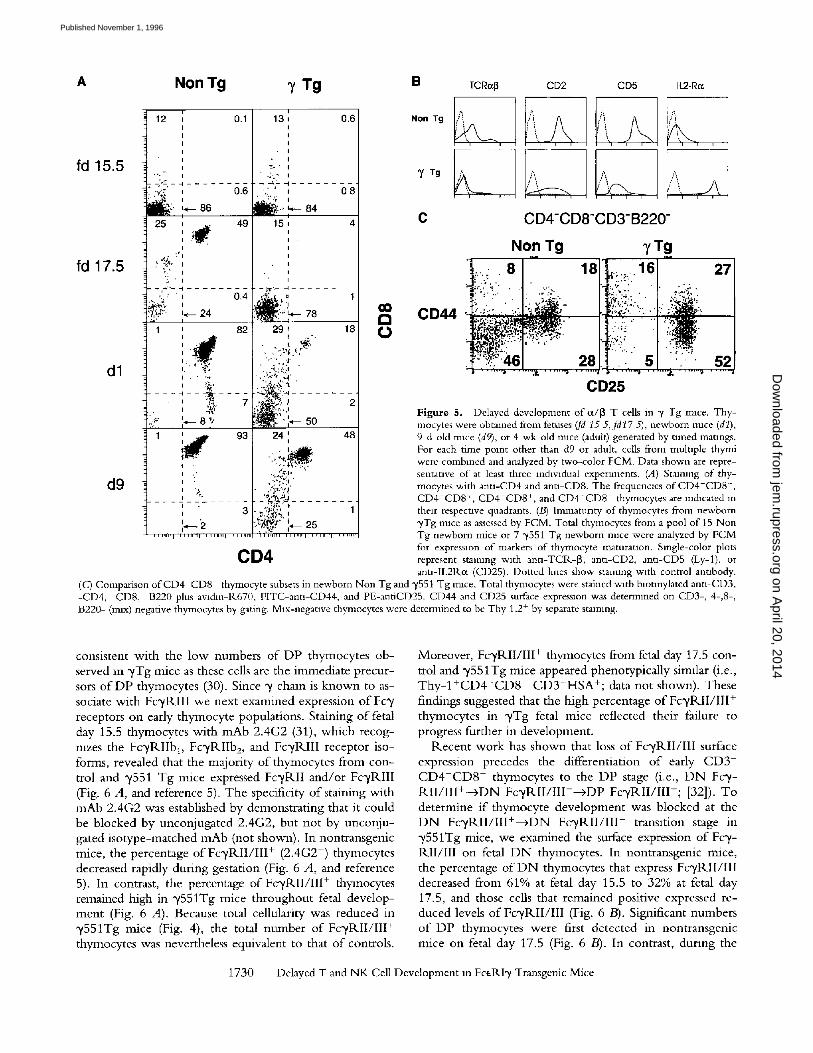
gressive stages o f fetal thymocyte maturation. Examination o f fetal thymocytes from high copy number ("/551) Tg mice revealed an early perturbation o f thymocyte matura- tion (Figs. 4 and 5). Total numbers o f thymocytes were equivalent in control and "/551 Tg mice on day 15, but at all subsequent stages, the number of thymocytes in ~/Tg mice was reduced relative to nontransgenic mice (Fig. 4). By fetal day 17.5, significant numbers o f "transitional," C D 4 - C D 8 l° and DP thymocytes were detectable in nor- mal mice, whereas most thymocytes in the y551 Tg mice remained D N (Fig. 5 A). DP and SP thymocytes began to accumulate in ",/551 Tg mice only after birth and the rela- tive number o f D N thymocytes remained high even in adult mice (Figs. 3 and 5). Thymocytes from y551Tg mice were immature relative to nontransgenic mice o f the same gestational age by several other criteria. For example, most thymocytes from "/551 transgenic newborn mice resembled fetal thymocytes in that they were IL2-1Kot + and expressed low levels o f surface CD2, CD5, and ot/[3TC1K (Fig. 5 B).
Fetal Thymocyte Development Is Delayed at the CD4-CD8- FcTRII/III + Stage in yTg Mice. Since our analysis o f fetal thymocytes 8om ~/Tg mice revealed a block at the DN--+DP transition stage, we examined the phenotype o f early D N thymocyte populations in y T g mice. Four distinct subsets o f precursor "triple negative" ( C D 3 - C D 4 - C D 8 - ) thy- mocytes have been identified on the basis of CD44 and CD25 expression that represent progressive stages of develop- ment: CD44+ CD25---+CD44+ CD25 +-->CD441°/- CD25+ - -+CD44-CD25- (30). FACS ® analysis o f D N thymocyte subsets from newborn ~/Tg mice revealed a specific block in development at the CD44+/l°CD25 + stage (Fig. 5 C-'). The reduction in C D 4 4 - C D 2 5 triple negative thymocytes was
1729 Flamand et al.
on April 20, 2014
jem.rupress.org
Dow
nloaded from
Published November 1, 1996
A
fd 15.5
fd 17.5
d l
d9
Non Tg y Tg
i i
12 , 0.1 13 I I I I I I
I I I , I I • I
.-:,a ' 0.6 ',; " ,
25 , 49 15 , , ~ , I I
I
' " Q i . ,
' ~ / " ,' 0.4 .~,.....it,~_.{=:" ~,,, " :,~'L?> '~-- 24 " 78
i
1 ' 82 29
' # ,,.£.:, ,~]. , " . ,t .'~3':.."~'" • ~ ".. '=: . ; , '
: % - ' , , ,1-
. . . . . . . = : ~ - - - r', - C ~ ~'--. . . . .
,, I , , ' "
] , . , , , , % .
I
I
~ 3 I •
0.6
0 8
4
1 0 0
18 ¢j
2
48
, ' ] r , p
V .~ . ; i
' ',,~ , , , , , - i , , H , , , I , , , , , H
CD4
B
Non Tg
y T g
T C R ~ CD2 CD5 IL2-Ro~
i ~ , , , , , , , ,
A C CD4-CD8"CD3-B220-
CD44
Non Tg
. .."~-- ;~ . ; . . . . .
~:.. .-2 ' . " " : ' ~ ...... .....,..:.;~_, f ; * ~ ' . . . . . . . . :
28
0D25
7Tg
. : ' % :
' 52
Figure 5. Delayed development of c¢/~ T cells in y Tg mace. Tby- mocytes were obtained from fetuses (fd 15 5,fd17 5), newborn mice (dl), 9-d-old rmce (d9), or 4-wk-old rmce (adult) generated by timed matmgs. For each time point other than d9 or adult, cells from mulnple thymi were combined and analyzed by two-color FCM. Data shown are repre- sentative of at least three mdwldual experiments. (A) Staining of thy- mocytes with anu-CD4 and ann-CD8. The frequencms of CD4-CD8-, CD4-CD8 +, CD4+CD8 +, and CD4+CD8 - thyrnocytes are indicated m their respective quadrants. (B) Immatunty of thymocytes from newborn ~/Tg mice as assessed by FCM. Total thymocytes from a pool of 15 Non Tg newborn mice or 7 "y551 Tg newborn mice were analyzed by FCM for expression of markers of thymocyte maturanon. Single-color plots represent staining with antl-TCR-[3, antl-CD2, ann-CD5 (Ly-1), or ann-IL2Rc((CD25). Dotted hnes show staining wzth control annbody.
(c) Comparison ofCD4-CD8- thymocyte subsets in newborn Non Tg and y551 Tg mice. Total thymocytes were stained with blotmylated antl-CD3, -CD4, -CD8, -B220 plus avldm-R670, FITC-ann-CD44, and PE-antiCD25. CD44 and CD25 surface expression was determined on CD3-, 4-,8-, B220- (max) negative thymocytes by gating. Mix-negative thymocytes were determined to be Thy 1.2 + by separate staining.
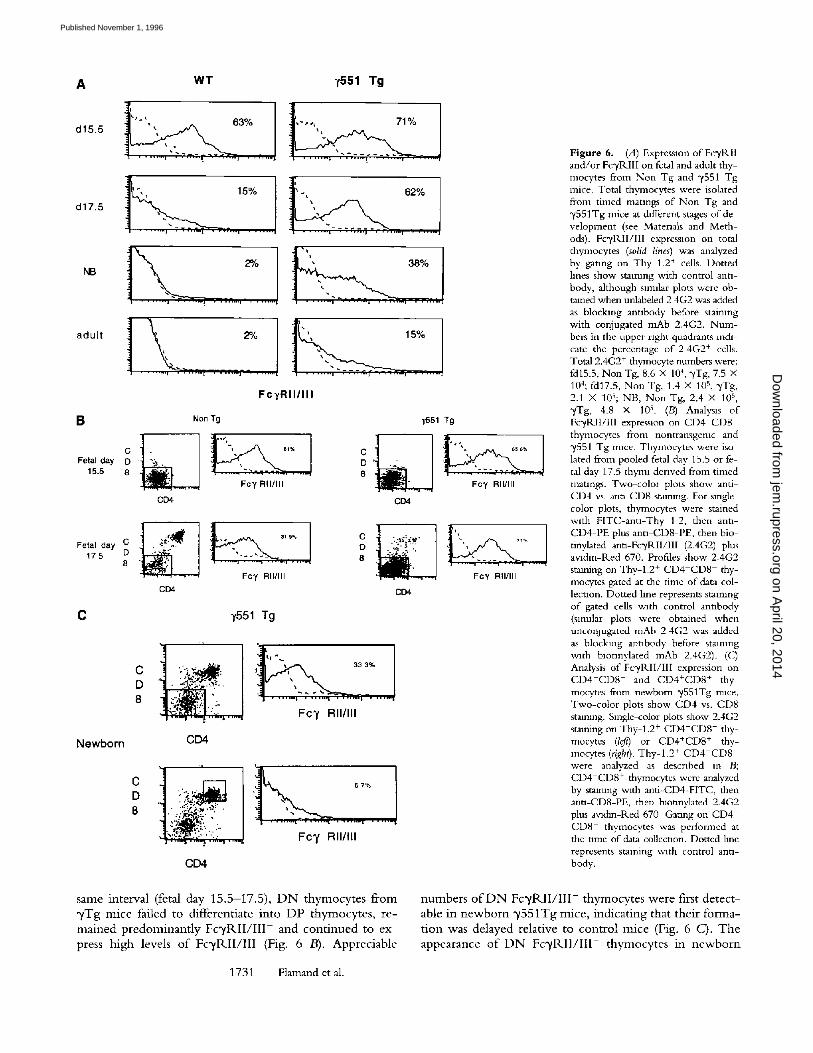
consistent with the low numbers o f DP thymocytes ob- served m ",/Tg mice as these cells are the immediate precur- sors o f DP thymocytes (30). Since ",/chain is known to as- sociate with FcyRI I I we next examined expression o f Fc',/ receptors on early thymocyte populations. Staining of fetal day 15.5 thymocytes wi th mAb 2.4G2 (31), which recog- nizes the F c y R I I b D Fc 'yRIIb 2, and Fc 'yRIII receptor iso- forms, revealed that the majority o f thymocytes from con- trol and ~/551 Tg mice expressed FcyRI I and/or Fc'yRIII (Fig. 6 A, and reference 5). The specificity of staining with mAb 2.4G2 was established by demonstrating that it could be blocked by unconjugated 2.4G2, but not by unconju- gated isotype-matched mAb (not shown). In nontransgenic mice, the percentage o f F o / R I I / I I I + (2.4G2 +) thymocytes decreased rapidly during gestation (Fig. 6 A, and reference 5). In contrast, the percentage of FcyRII / I I I + thymocytes remained high in "y551Tg mice throughout fetal develop- ment (Fig. 6 A). Because total cellularity was reduced in "y551Tg mice (Fig. 4), the total number o f FcyRI I / I I I + thymocytes was nevertheless equivalent to that o f controls.
Moreover, FcyRII / I I I + thymocytes from fetal day 17.5 con- trol and "y551Tg mice appeared phenotypically sirmlar (i.e., T h y - I + C D 4 - C D 8 C D 3 - H S A + ; data not shown). These findings suggested that the high percentage o f Fc 'yRII / I I I + thymocytes in "yTg fetal mice reflected their failure to progress further in development.
Recen t work has shown that loss o f FcI, R I I / I I I surface expression precedes the differentiation o f early C D 3 - C D 4 - C D 8 - thymocytes to the DP stage (i.e., D N Fcy- R I I / I I I + - + D N F c ~ / R I I / I I I - - + D P F c y R I I / I I I - ; [32]). To determine i f thymocyte development was blocked at the D N F o / R I I / I I I + - + D N F c y R I I / I I I - transition stage in y551Tg mice, we examined the surface expression of Fcy- RI I / I I I on fetal D N thymocytes. In nontransgenic mice, the percentage o f D N thymocytes that express Fc~/RII/III decreased from 61% at fetal day 15.5 to 32% at fetal day 17.5, and those cells that remained positive expressed re- duced levels o f Fc~/RII/III (Fig. 6 B). Significant numbers o f DP thymocytes were first detected in nontransgenic mice on fetal day 17.5 (Fig. 6 B). In contrast, dunng the
1730 Delayed T and NK Cell Development m FceRIy Transgenic Mice
on April 20, 2014
jem.rupress.org
Dow
nloaded from
Published November 1, 1996
A
d15.5
d17.5
NB
adult
B
c Fetal day D
15.5 8
Fetal day C 175 D
8
C
Newborn
WT 7551 Tg
t . . . . . . . . . . . . . j a , . . ~ . \ , , , | = . . . . . . . .
. . . . . . . . l . . . . . . . . i . . . . . . . . J
, - , ,, 71%
7.:.:_J
I, ", ', 62%
~ _ . , 3 8 % .,
~ 15%
FcyRI I / I I I
Ncn Tg
Fc 7 RII/III CD4
7551 Tg
C
° .L 8
. . . . .
CD4
8 D ~ C ::~.~::.~.
CD4
y551 Tg
D 8
CD4
Fc 7 RII/III
i
Fc 7 RII/III
CD4
s a m e in t e rva l (fetal day 1 5 . 5 - 1 7 . 5 ) , D N t h y m o c y t e s f r o m
y T g m i c e fa i led to d i f f e r en t i a t e i n t o D P t h y m o c y t e s , r e -
m a i n e d p r e d o m i n a n t l y F c ' y R I I / I I I + a n d c o n t i n u e d to e x -
press h i g h levels o f F c ' , / R I I / I I I (Fig. 6 B). A p p r e c i a b l e
1731 Flamand et al.
Fc 7 RII/III
Fc7 RII/III
Figure 6. (A) Expression of Fc~/RII and/or Fc~/RIII on fetal and adult thy- mocytes from Non Tg and ~/551 Tg mice. Total thymocytes were isolated from timed matmgs of Non Tg and "y551Tg mice at different stages of de- velopment (see Matenals and Meth- ods). Fc~RII/III expression on total thymocytes (solid lines) was analyzed by gating on Thy 1.2 + cells. Dotted hnes show staining with control anti- body, although slrmlar plots were ob- tained when unlabeled 2 4G2 was added as blocking antibody before staimng with conjugated mAb 2.4G2. Num- bers in the upper nght quadrants re&- care the percentage of 2 4G2 + cells. Total 2.4G2 + thymocyte numbers were: fd15.5, Non Tg, 8.6 X 10 4, 3'Tg, 7.5 X 104; fd17.5, Non Tg, 1.4 X 105, "yTg, 2.1 X 10s; NB, Non Tg, 2.4 X 105 , 3'Tg, 4.8 X 105 . (B) Analysis of Fc'yRII/llI expression on CD4 CD8 thymocytes from nontransgemc and 3~551 Tg mice. Thymocytes were iso- lated from pooled fetal day 15.5 or fe- tal day 17.5 thyrm derived from timed matmgs. Two-color plots show anti- CD4 vs. ann-CD8 stmmng. For single- color plots, thymocytes were stained with FITC-antl-Thy 1 2, then anti- CD4-PE plus ann-CD8-PE, then bio- tinylated anu-Fc~RIl/III (2.4G2) plus avldm-Red 670. Profiles show 2.4G2 staining on Thy-l.2 + CD4 CD8 thy- mocytes gated at the time of data col- lection. Dotted hne represents staining of gated cells with control antibody (slmdar plots were obtained when uncon3ugated mAb 2 4G2 was added as blocking antibody before staining with blotinylated mAb 2.4G2). (C) Analysis of Fc'yPdI/III expression on C D 4 - C D 8 - and CD4+CD8 + thy- mocytes from newborn "/551Tg mace. Two-color plots show CD4 vs. CD8 stalmng. Single-color plots show 2.4G2 staining on Thy-l.2 + CD4-CD8- thy- mocytes (left) or CD4+CD8 + thy- mocytes (right). Thy-l .2 + CD4-CD8- were analyzed as described in B; CD4+CD8 + thymocytes were analyzed by stalmng v~th anfi-CD4-FITC, then anti-CD8-PE, then blounylated 2.4G2 plus avldin-Red 670 Gating on CD4 + CD8 + thymocytes was performed at the tmle of data collection. Dotted hne represents staining with control anti- body.
n u m b e r s o f D N F c ~ / R I I / I I I - t h y m o c y t e s w e r e first d e t e c t -
able in n e w b o r n y 5 5 1 T g m i c e , i n d i c a t i n g tha t t h e i r f o r m a -
t i o n was d e l a y e d re la t ive to c o n t r o l m i c e (Fig. 6 C). T h e
a p p e a r a n c e o f D N F c ~ / R I I / I I I - t h y m o c y t e s in n e w b o r n
on April 20, 2014
jem.rupress.org
Dow
nloaded from
Published November 1, 1996

I - t - o I-.--
Z 2"-,.
1 2 1 2
14 kD
-'-3'
Figure 7. Colmmunoprecipitation of "/ chain with mAb 2.4G2. Fetal day 17.5 thymocytes from Non Tg (15 × 106) or y Tg (8 × 106) mace were lysed in buffer containing 1% digltonin, precleared with protein A-Sepharose plus normal rabblt serum, then incubated with protein A-Sepharose plus rabbit anti-rat anubody (lane 1), or rabbit anti-rat anti- body plus mAb 2.4G2 (lane 2). Immunopreclpltates were washed, solubl- lized in reducing SDS loading buffer, and separated by SDS-PAGE (14%). Proteins were transferred to nitrocellulose and blotted with a 1:1,000 di- lution of ann-"/sera Immunoreactxve proteins were detected by chemilu- minescence. Exposure Umes were 30 s for Non Tg samples and 3 s for "/Tg samples
~/551Tg mice coincided with the appearance o f DP thy- mocytes, which were also F c y R I I / I I I - (Fig. 6 C). Collec- tively, these observations indicate that y overexpression de- lays the maturation o f D N Fc~/R.II/III + thymocytes.
Expression of FcTRllI in Fetal 77zymocytes. mAb 2.4G2 re- acts with three distinct F c y R isoforms (tklIbl, RI Ib 2, and Rill) only one o f which (Fc',/RIII) is known to associate with y chain. Consequently, we were interested in deter- mining whether the 2.4G2 staining on fetal thymocytes from 3'551Tg mice reflected expression o f Fc',/tLIII. Re - verse transcription-PCR analysis demonstrated that tran- scripts for both FcyP, IIb and FcyRI I I (5; data not shown) are expressed in both fetal and newborn thymocytes from control and ~/551Tg mice. The presence of FcyRIII (ix + y) complexes was then assessed by immunopreclpitation with m_Ab 2.4G2 followed by immunoblotting with anti-y chain sera. As shown in Fig. 7, y chain could be coprecipitated with mAb 2.4G2 from both nonTg and y551Tg fetal day 17.5 thymocyte lysates. However, the amount o f c o i m m u - nopreclpitated y chain was significantly greater in thy- mocyte lysates from fetal day 17.5 y551Tg mice.
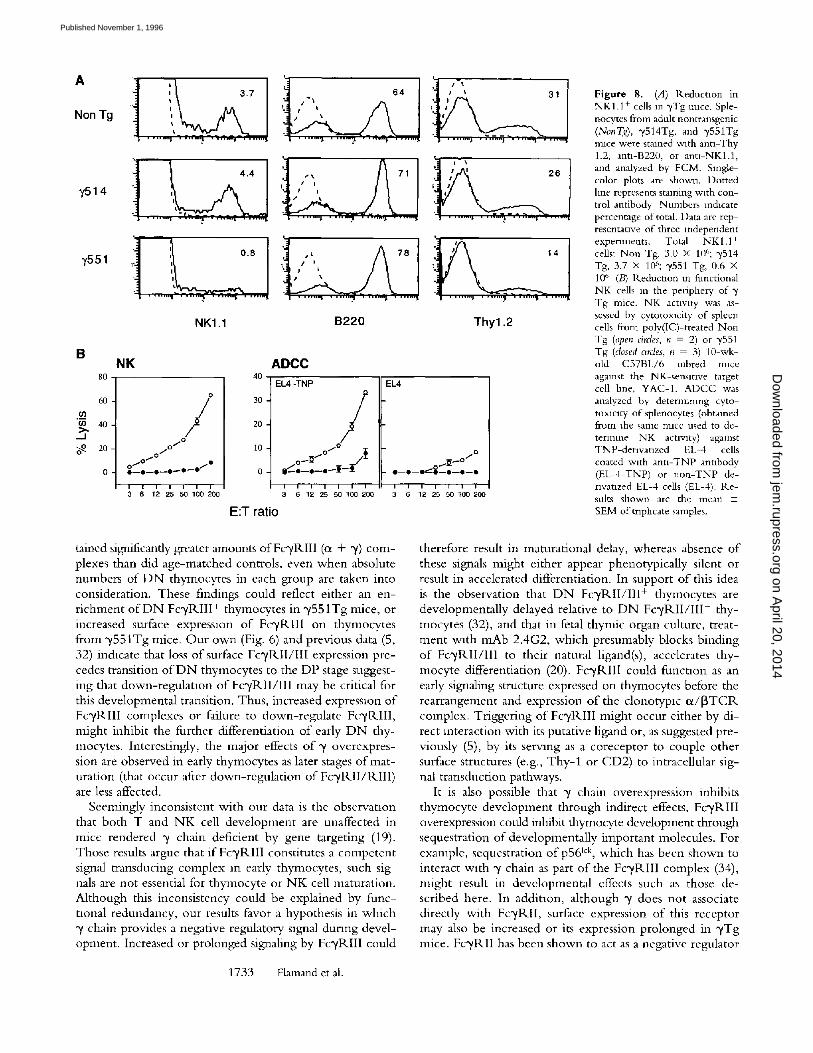
Impaired NK Cell Development in yTg Mice. In view of recent data indicating that at least some N K cells may orig- inate from early Fc~/RIII + thymocyte progenitors (5, 6), we assessed whether N K cell development was compro- mised in ~/Tg mice. Analysis o f splenocytes (Fig. 8 A) and
PBL (not shown) demonstrated that the number o f N K 1.1 + cells was reduced in y551Tg mice relative to non- transgenic controls. In contrast, B cell numbers were nor- mal or elevated in these mice (Fig. 8 A). As with T cells (indicated by staining with anti-Thy 1.2), the reduction in NKI .1 + cells was transgene dose dependent (Fig. 8 A). To assess N K activity, nontransgenic and ~/551 Tg mice were injected with poly(IC) and spleen cells were assayed for cy- totoxicxty against the NK-sensitive target cell, YAC-1, and for ADCC. Consistent with the FACS ® analysis, NK activ- ity and A D C C were negligible in y551 Tg mice (Fig. 8 B). Taken together, these results demonstrate that the number o f N K cells is markedly reduced in y551 Tg mice.
Discuss ion
The present study was directed at exploring a possible role for F c e R b / i n thymocyte development. Northern blot analysis o f fetal and adult thymocytes demonstrate that ~/is expressed in the fetal thymus: expression is highest at early stages (fetal day 13.5-15.5), and decreases progressively with gestational age. This pattern contrasts sharply with that o f the structurally and functionally related ~ chain, which is expressed later in development and functions pri- marily as a signal-transducing subumt o f the ix/[3TCtL complex. In view of these results, we generated transgenic mice that overexpress ",/, reasoning that a perturbation o f thymocyte maturation, if observed, might yield insight into its function in early thymocytes. Analysis o f multiple trans- genic founder lines revealed that y overexpression does in- deed impair early thymocyte development. The transgene effects are dose related and result in delayed maturation o f fetal D N thymocytes, with less severe effects on later stages o f development.
Analysis of fetal thymocytes from high copy number y551Tg mice revealed that ~/chain overexpression specifi- cally blocked thymocyte development at the D N CD25 + F c 3 ' R I I / R I I I + - + D N C D 2 5 - Fcytkl I /P , I I I - transition stage. As D N Fc~/R.II/RIII + fetal thymocytes are thought to include common precursors o f T lymphocytes and NK cells (5), it is notable that N K cell development was like- wise impaired in ~/Tg mice. Further support for the idea that T and N K cells share a common precursor is provided by the observation that perturbation o f both T cell and N K cell development was also observed in transgenic mice in which human CD3e was overexpressed in early thy- mocytes (33).
One interpretation of our observations is that the ~/trans- gene-induced effects are mediated by Fc2*tLIII (CD16) com- plexes on early thymocytes. Although the mAb used to assess F c y R I I / I I I surface expression does not distinguish between these isoforms and therefore does not enable specific quan- titative evaluation o f FcyRII I surface expression, Western blot analysis is consistent with the idea that y is associated with FcyRIIIix in both nontransgenic and ~/Tg fetal thy- mocytes. However, fetal day 17.5 thymocytes from y551Tg mice, which express high surface levels o f Fc2/RII/III, con-
1732 Delayed T and NK Cell Development in FceRIy Transgenic Mice
on April 20, 2014
jem.rupress.org
Dow
nloaded from
Published November 1, 1996
A
Non Tg
~,51 4
7551
B
80
6O
O3
"~ 40 _J
i
, t. t t
) . . . . . . . . I . . . . . . . . ~ . . . . . . . . It . . . . . . . ~ Jt 1~
i
! . ]., I I /
NKI.1 B220
i
' i i 3 1
' ; , 2 8
Thy1.2
NK
/ f o
j o
i ° 0 ~ , 0 . ~ 0 0 - - 0 - - 0 - - 0 ~ 0 - o
i I i / I i i 3 6 12 25 50 100 200
40
30
20
10
0
E:T ratio
ADCC EL4 -TNP
Q
/ / . o
o ~ I O A
i i i i i i i
3 6 12 25 50 100 200
EL4
Q - - O - - O ~ ' O - - O - - O - - Q
I I ! I I I I
3 6 12 25 50 100
Figure 8. (.4) Reductmn in NKI.1 + cells m ~vTg mice. Sple- nocytes from adult nontransgenic (NonT~a), y514Tg, and ~,551Tg rmce were stamed w~th ann-Thy 1.2, ann-B220, or ann-NKl.1, and analyzed by FCM. Single- color plots are shown. Dotted hne represents staining w~th con- trol antibody Numbers m&cate percentage of total. Data are rep- resentanve of three independent experiments. Total NK 1.1 + cells: Non Tg, 3.0 × 10~'; y514 Tg, 3.7 × 106; 3,551 Tg, 0.6 × 106 (B) R.educuon m functional NK cells m the periphery of Tg mice. NK acnvlty was as- sessed by cytotoxlcity of spleen cells from poly(IC)-treated Non Tg (open circles, n = 2) or y551 Tg (closed ames, n = 3) 10-wk- old C57BL/6 inbred mace against the NK-sensmve target cell hne, YAC-1. ADCC was analyzed by deterrmnmg cyto- toxicity of splenocytes (obtained from the same rmce used to de- termine NK actlwty) against TNP-denvanzed EL-4 cells coated with ann-TNP annbody (EL-4-TNP) or non-TNP de- nvanzed EL-4 cells (EL-4). R.e- suits shown are the mean 4- SEM of triplicate samples.
tained significantly greater amounts of FcyRII I (or + ~) com- plexes than did age-matched controls, even when absolute numbers o f D N thymocytes in each group are taken into consideration. These findings could reflect either an en- r ichment o f D N F c y R I I I + thymocytes in y551Tg mice, or increased surface expression o f Fc~RII I on thymocytes from y551Tg mice. O u r own (Fig. 6) and previous data (5, 32) indicate that loss of surface Fc',/lq.II/III expression pre- cedes transition o f D N thymocytes to the DP stage suggest- ing that down-regula t ion o f Fc'yP,.II/III may be critical for this developmental transition. Thus, increased expression of Fc 'yRIII complexes or Failure to down-regulate Fc'yR.III, might inhibit the further differentiation o f early D N thy- mocytes. Interestingly, the major effects o f "y overexpres- sion are observed in early thymocytes as later stages o f mat- uration (that occur after down-regula t ion o f Fc 'yRI I /RI I I ) are less affected.
Seemingly inconsistent wi th our data is the observation that both T and N K cell development are unaffected in mice rendered "g chain deficient by gene targeting (19). Those results argue that if Fc~RII I constitutes a competent signal-transducing complex m early thymocytes, such sig- nals are not essential for thymocyte or N K cell maturation. Al though this inconsistency could be explained by func- tional redundancy, our results favor a hypothesis in which y chain provides a negative regulatory signal during devel- opment. Increased or prolonged signaling by Fc',/RIII could
1733 Flamand et al.
therefore result in maturational delay, whereas absence o f these signals might either appear phenotypically silent or result in accelerated differentiation. In support o f this idea is the observation that D N F c y R I I / I I I + thymocytes are developmental ly delayed relative to D N F c ~ R I I / I I I - thy- mocytes (32), and that in fetal thymic organ culture, treat- ment with mAb 2.4G2, which presumably blocks binding of Fo /R . I I / I I I to their natural ligand(s), accelerates thy- mocyte differentiation (20). Fc',/RIII could function as an early signaling structure expressed on thymocytes before the rearrangement and expression o f the clonotyplc <x/I3TCP,. complex. Triggering o f Fo/P,.III might occur either by di- rect interaction with its putative ligand or, as suggested pre- viously (5), by its serving as a coreceptor to couple other surface structures (e.g., Thy-1 or CD2) to intracellular sig- nal transduction pathways.
It is also possible that "y chain overexpression inhibits thymocyte development through indirect effects. Fc' ,/RIII overexpression could inhibit thymocyte development through sequestration o f developmental ly important molecules. For example, sequestration o f p56 Ick, which has been shown to interact with "y chain as part o f the Fc~/RIII complex (34), might result in developmental effects such as those de- scribed here. In addit ion, a l though "y does not associate directly with Fc'yP,.II, surface expression o f this receptor may also be increased or its expression pro longed in "gTg mice. Fcyt<II has been shown to act as a negative regulator
on April 20, 2014
jem.rupress.org
Dow
nloaded from
Published November 1, 1996

of hematopoetic cell activation (35); therefore its failure to be down-regulated in early thymocytes could be inhibi- tory. Since mAb 2.4G2 does not distinguish between Fc3'RII and Fc3'R.III we were unable to discern the relative expression o f these receptors in non -Tg and 3' Tg mice. However, both we (data not shown) and others (5) have detected Fc3'P, IIb expression during fetal thymocyte devel- opment by reverse transcription-PCP,. Notwithstanding, two lines o f evidence indicate that the developmental ef- fects observed in 3'Tg mice are specific to y chain. First, we have previously shown that overexpression of both ~1 chain or a truncated form of ~ chain that, like y, contains only a single ITAM does not inhibit thymocyte maturation (22). Second, reconstitution of~-deficient (~-/-) mice with y trans- genes revealed that the phenotype o f y T g mice is identical regardless o f the presence o f absence o f endogenous ~ chain (data not shown). These results indicate that the develop- mental effects o f y overexpression are not the result o f its association with ~ chain or due to competition with ~ chain for association with other molecules.
The phenotype o f 3'Tg and ~Tg mice also provides In- sight into the function o f these molecules in developing thymocytes. Although T cell development is adversely af- fected in both ~ and 3' transgenic mice, their phenotypes are strikingly different, y chain overexpression delays, but does not arrest, thymocyte development at the D N Fc3'P, I I / I I I + stage, whereas ~ chain overexpression results in a near com- plete arrest at a later stage o f development (22). Fetal thy- mocytes from ~Tg mice but not 3'Tg mice are further dis- tinguished by their inappropriate expression of activation markers and premature down-regulation o f P, agl and II.ag2 (22). These data suggest that y and { may couple to different signaling pathways in early thymocytes. One pos- sibility is that y and ~ associate preferentially with different cell surface structures, e.g., ~ chain along with the CD3 subunits as part o f the p r e - T C R and oe/[3TCP, complexes, and 3' chain with the Fc3'RIII complex. Indeed, the dis- tinctive patterns o f 3' and ~ expression during gestation would be consistent with this idea.
We thank Drs. A. Singer, D. Welst, and B.J. Fowlkes for critical review of the manuscript.
Address correspondence to Paul E. Love, Bml&ng 6B, Room 2B-210, Laboratory of Mammalian Genes and Development, National Institutes of Health, Bethesda, MD 20892. J.-P. Kmet's present address is Harvard Medical School, Beth Israel Hospital, Research North Building, 99 Brookllne Avenue, Boston, MA 02215.
Received for publication 11June 1996 and in revised form 8 August 1996.
References 1. Teh, S.H., P. Kisielow, B. Scott, H. Kishi, Y. Uematsu, H.
Bluthmann, and H. von Boehmer. 1988. Thymlc major his- tocompatibility complex antigens and the specificity of the ot[3 T cell receptor determine the CD4/CD8 phenotype of cells. Nature (Lond.). 335:229-233.
2. Fowlkes, B.J., and D.M. Pardoll. 1989. Molecular and cellu- lar events o fT cell development. Adv. Immunol. 44:207-265.
3. Reth, M. 1989. Antigen receptor tail clue. Nature (Lond.). 338:383-384.
4. Weiss, A., and D.R. Littman. 1994. Signal transductaon by lymphocyte antigen receptors. Cell. 76:263-274.
5. R.odewald, H-R., P. Momgeon, J.L. Luclch, C. Dosiou, P. Lopez, and E.L. Relnherz. 1992. A population of early fetal thymocytes expressing Fc3'RII/III contains precursors of T lymphocytes and natural killer cells. Cell. 69:139-150.
6. S~nchez, M.J., M.O. Muench, M.G. Roncarolo, L.L. Lanier, andJ.H. Phillips. 1994. Idenufication of a common T/natural killer cell progemtor in human fetal thymus. J. Exp. Med. 180:569-576.
7. Kruisbeek, A.M. 1993. Development of c~[3 T cells. Curr. Opin. Immunol. 5:227-234.
8. Zfifiiga-Pflficker, J.C., H. Schwartz, and MJ. Lenardo. 1993. Gene transcripnon in differentiating immature T cell recep- tor neg thymocytes resembles antigen-activated mature T cells. J. Exp. Med. 178:1139-1149.
9. Lanier, L.L., T.J. Klpps, and J.H. Phillips. 1985. Functional properties of a unique subset of cytotoxlc CD3 + T lympho-
cytes that express Fc receptors for IgG (CD16/Leu-11 anti- gen).J. Exp. Med. 162:2089-2106.
10. Ra, C., M-H. Jouvin, and J-P. Klnet. 1989. Complete struc- ture of the mouse mast cell receptor for IgE (FceR1) and sur- face expression of chimeric receptors (rat-mouse-human) on transfected cells. J. Biol. Chem. 264:15323-15327.
1 l. Kurosaki, T., I. Gander, andJ.V. Ravetch. 1991. A subunit common to an IgG Fc receptor and the T cell receptor medi- ates assembly through different interactions. Proc. Natl. Acad. Sci. USA. 88:3837-3841.
12. Orloff, D.G., C. Ra, S.J. Frank, R.D. Klausner, andJ.-P. Ki- net. 1990. Family of disulphide-linked dlmers containing the
and ~ chains of the T-cell receptor and the 3' chain of Fc receptors. Nature (Lond.). 347:189-191.
13. Vivier, E., N. Rochet, J.P. Kochan, D.H. Preski, and S.F. Schlossman. 1991. Structural similarity between Fc receptors and T cell receptors. Expression of the 3' subunit of FceRI in human T cells, natural killer cells and thymocytes.J. Immunol. 147:4263-4270.
14. Koyasu, S., L. D'Adamio, A.IL.N. Arulanandam, S. Abra- ham, L.K. Clayton, and E. Relnherz. 1992. T cell receptor complexes containing FcetLI3' homodlmers in lieu of CD3~ and CD3"q components: a novel lsoform expressed on large granular lymphocytes.J. Exp. Med. 175:203-209.
15. Ernst, L.K., A.-M. Duchemin, and C.L. Anderson 1993. As- sociation of the high-affinity receptor for IgG (Fc',/RI) with the 3' subumt of the IgE receptor. Proc. Natl. Acad. Sci. USA.
1734 Delayed T and NK Cell Development in FceRIy Transgenic Mice
on April 20, 2014
jem.rupress.org
Dow
nloaded from
Published November 1, 1996

90:6023-6027. 16. Scholl, P.R., and R.S. Geha. 1993. Physical association be-
tween the high-affinity IgG receptor (Fc gamma P,I) and the gamma subunit of the high-affinity IgE receptor (Fc epsilon P,.I gamma). Proc. Natl. Acad. Sci. USA. 90:8847-8850.
17. Qian, D., A. Sperhng, D.W. Lancki, Y. Tatsumi, T.A. Bar- rett, J.A. Bluestone, and F.W. Fitch. 1993. The 3' chain of the high-affinity receptor for IgE is a major functional sub- unit of the T-cell antigen receptor complex in ~ T lympho- cytes. Proc. Natl. Acad. Sci. USA. 90:11875-11879.
18. Kiister, H., H. Thompson, andJ.-P. Kmet. 1990. Character- lzation and expression of the gene for the human Fc receptor
subunit. Definition of a new gene family. J. Biol. Chem. 265:6448-6452.
19. Takai, T., M. Li, D. Sylvestre, R. Clynes, andJ.V. Ravetch. 1994. FcR',/ chain deletion results m pleiotropic effector cell defects. Cell. 76:519-529.
20. Sandor, M., J. Galon, L. Takacs, Y. Tatsumi, A. L. Mueller, C. Sautes, and 1L.G. Lynch. 1994. An alternative Fc'y-recep- tor hgand: potential role in T cell development. Proc. Natl. Acad. Sci. (USA). 91:12857-12861.
21. Greaves, D.R., F.D. Wilson, G. Lang, and D. gaoussis. 1989. Human CD2 3'-flanking sequences confer high-level, T cell- specific, position-independent gene expression in transgenic mice. Cell. 56:979-986.
22. Love, P.E., E.W. Shores, E.J. Lee, A. Gnnberg, T.I. Munitz, H. Westphal, and A. Singer. 1994. Differential effects of and "q transgenes on early or/J3 T cell development. J. Exp. Med. 179:1485-1494.
23. Fink, P.J., L.A. Matls, D.L. McElhgott, M. Bookman, and S.M. Hedrlck. 1986. Correlations between T-cell specificity and the structure of the antigen receptor. Nature (Lond.). 321: 219-226.
24. Hedrick, S.M., D.I. Cohen, E.A. Nielsen, and M.M. Davis. 1984. Isolation of cDNA clones encoding T cell-specific membrane-associated proteins. Nature (Lond.). 308:149-153.
25. Letourneur, O., I.C.S. Kennedy, A.T. Brini, J.P,. Ortaldo, J.J. O'Shea, and J.-p. Kinet. 1991. Characterization of the family of dimers associated with Fc Receptors (FcelkI and Fc~RIII).J. Immunol. 147:2652-2656.
26. Liu, C.-P., P,. Ueda, J. She, J. Sancho, B. Wang, G. Wed- dell, J. Loring, C. Kurahara, E.C. Dudley, A. Hayday et al.
1993. Abnormal T cell development in CD3-~ - / - mutant mice and identification of a novel T cell population in the in- testine. EMBO (Eur. Mol. Biol. Organ.) J. 12:4863-4875.
27. Love, P.E., E.W. Shores, M.D. Johnson, M.L. Tremblay, E.J. Lee, A. Grinberg, S.P. Huang, A. Singer, and H. Westphal. 1993. T cell development in mice that lack the ~ chain of the T cell antigen receptor complex. Science (Wash. DC). 261: 918-921.
28. Ohno, H., T. Aoe, S. Taki, D. Katamura, Y. Ishida, K. 1ka- jewsky, and T. Saito. 1993. Developmental and functional impairment of T cells in mice lacking CD3~ chains. EMBO (Eur. Mol. Biol. Organ.)J. 12:4357-4366.
29. Malissen, M., A. Gillet, B. Rocha, J. Tmcy, E.Viver, C. Boyer, F. Kontgen, N. Brun, G. Mazza, E. Spanopoulou et al. 1993. T cell development in mice lacking the CD3-~/'q gene. EMBO (Eur. Mol. Biol. Organ.)J. 12:4347-4355.
30. Godfrey, D.I., and A. Zlotnik. 1993. Control points in early T-cell development. Immunol. Today. 14:547-553.
31. Unkeless, J.C. 1979. Characterization of a monoclonal anti- body directed against mouse macrophage and lymphocyte Fc receptors.J. Exp. Med. 150:580-596.
32. Rodewald, H-P,., K. Awad, P. Moingeon, L. D'Adamio, D. Rabinowitz, Y. Shinkai, F.W. Alt, and E.L. R_einherz. 1993. Fc2tlklI/IlI and CD2 expression mark distinct subpopulations of immature CD4-CDS- munne thymocytes: in vlvo devel- opmental kinetics and T cell receptor [3 chain rearrangement status.J. Exp. Med. 177:1079-1092.
33. Wang, B., C. Blron, J. She, K. Higglns, M.-J. Sunshine, E. Lacy, N. Lonberg, and C. Terhorst. 1994. A block in early T lymphocyte and natural killer cell development in transgenic mice with high-copy numbers of the human CD3E gene. Proc. Natl. Acacl. Sci. USA. 91:9402-9406.
34. Salcedo, T.W., T. Kurosaki, P. Kanakaraj, J.V. Ravetch, and B. Perussla. 1993. Physical and functional association of p56 lck with Fc'yRIIIA (CD16) in natural killer cells. J. Exp. Med. 177:1475-1480.
35. D'Ambroslo, D., K.H. Hlppen, S.A. Minskoff, I. Mellman, K.A. Slminovitch, G. Pani, andJ.C. Cambier. 1995. PTP1C recruitment and activation by Fc',/R.IIBI: a molecular basis for negative signal cooperativlty with the B cell antigen re- ceptor. Science (Wash. DC). 268:263-264.
1735 Flamand et al.
on April 20, 2014
jem.rupress.org
Dow
nloaded from
Published November 1, 1996
Copyright © 2022 FDOKUMEN




















