
Deficits in inhibitory control and conflict resolution on cognitive and motor tasks in Parkinson’s...
14
RESEARCH ARTICLE Deficits in inhibitory control and conflict resolution on cognitive and motor tasks in Parkinson’s disease Ignacio Obeso • Leonora Wilkinson • Enrique Casabona • Maria Luisa Bringas • Mario A ´ lvarez • La ´zaro A ´ lvarez • Nancy Pavo ´n • Maria-Cruz Rodrı ´guez-Oroz • Rau ´l Macı ´as • Jose A. Obeso • Marjan Jahanshahi Received: 28 January 2011 / Accepted: 16 May 2011 / Published online: 4 June 2011 Ó Springer-Verlag 2011 Abstract Recent imaging studies in healthy controls with a conditional stop signal reaction time (RT) task have implicated the subthalamic nucleus (STN) in response inhibition and the pre-supplementary motor area (pre-SMA) in conflict resolution. Parkinson’s disease (PD) is charac- terized by striatal dopamine deficiency and overactivity of the STN and underactivation of the pre-SMA during movement. We used the conditional stop signal RT task to investigate whether PD produced similar or dissociable effects on response initiation, response inhibition and response initiation under conflict. In addition, we also examined inhibition of prepotent responses on three cognitive tasks: the Stroop, random number generation and Hayling sentence completion. PD patients were impaired on the conditional stop signal reaction time task, with response initiation both in situations with or without conflict and response inhibition all being significantly delayed, and had significantly greater difficulty in suppressing prepotent or habitual responses on the Stroop, Hayling and random number generation tasks relative to controls. These results demonstrate the existence of a generalized inhibitory deficit in PD, which suggest that PD is a disorder of inhibition as well as activation and that in situations of conflict, execu- tive control over responses is compromised. Keywords Fronto-striatal circuits Á Parkinson’s disease Á Subthalamic nucleus Á Inhibition Á Stop signal task Introduction The basal ganglia and the frontal cortex are intimately connected via the fronto-striatal circuits, and direct, indi- rect and hyperdirect pathways between these structures have been delineated (Alexander et al. 1986; Middleton and Strick 2000). The direct and indirect pathways have been proposed to constitute an ideal system for response selection and initiation under competition or conflict, with the indirect pathway via the subthalamic nucleus (STN) inhibiting inappropriate responses to allow selection and initiation of the appropriate response through the direct pathway (e.g. Chevalier and Deniau 1990; Redgrave et al. 1999). In Parkinson’s disease (PD), degeneration of dopa- minergic neurons in the pars compacta of the substantia nigra alters the balance of activity in the direct and indirect pathways, the net effect of which increased inhibitory outflow from the internal segment of the globus pallidus I. Obeso Á L. Wilkinson Á M. Jahanshahi (&) Cognitive-Motor Neuroscience Group, Sobell Department of Motor Neuroscience and Movement Disorders, UCL Institute of Neurology, The National Hospital for Neurology and Neurosurgery, 33 Queen Square, London WC1N 3BG, UK e-mail: [email protected] E. Casabona Á M. L. Bringas Á M. A ´ lvarez Á L. A ´ lvarez Á N. Pavo ´n Á R. Macı ´as Movement Disorders and Neurophysiology Units, Centro Internacional de Restauracio ´n Neurolo ´gica (CIREN), La Habana, Cuba M.-C. Rodrı ´guez-Oroz Á J. A. Obeso Department of Neurology, Clı ´nica Universitaria and Medical School of Navarra, Neuroscience Centre, CIMA, University of Navarra, Pamplona, Spain M.-C. Rodrı ´guez-Oroz Á J. A. Obeso Centro de Investigacio ´n Biome ´dica en Red Sobre Enfermedades Neurodegenerativas (CIBERNED), Instituto Carlos III, Ministerio de Investigacio ´n y Ciencias, Barcelona, Spain Present Address: L. Wilkinson Brain Stimulation Unit, National Institute of Neurological Disorders and Stroke, Bethesda, MD 20892-1430, USA 123 Exp Brain Res (2011) 212:371–384 DOI 10.1007/s00221-011-2736-6
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Deficits in inhibitory control and conflict resolution on cognitive and motor tasks in Parkinson’s...

RESEARCH ARTICLE
Deficits in inhibitory control and conflict resolution on cognitiveand motor tasks in Parkinson’s disease
Ignacio Obeso • Leonora Wilkinson • Enrique Casabona • Maria Luisa Bringas •
Mario Alvarez • Lazaro Alvarez • Nancy Pavon • Maria-Cruz Rodrıguez-Oroz •
Raul Macıas • Jose A. Obeso • Marjan Jahanshahi
Received: 28 January 2011 / Accepted: 16 May 2011 / Published online: 4 June 2011
� Springer-Verlag 2011
Abstract Recent imaging studies in healthy controls with
a conditional stop signal reaction time (RT) task have
implicated the subthalamic nucleus (STN) in response
inhibition and the pre-supplementary motor area (pre-SMA)
in conflict resolution. Parkinson’s disease (PD) is charac-
terized by striatal dopamine deficiency and overactivity of
the STN and underactivation of the pre-SMA during
movement. We used the conditional stop signal RT task to
investigate whether PD produced similar or dissociable
effects on response initiation, response inhibition and
response initiation under conflict. In addition, we also
examined inhibition of prepotent responses on three
cognitive tasks: the Stroop, random number generation and
Hayling sentence completion. PD patients were impaired on
the conditional stop signal reaction time task, with response
initiation both in situations with or without conflict and
response inhibition all being significantly delayed, and had
significantly greater difficulty in suppressing prepotent or
habitual responses on the Stroop, Hayling and random
number generation tasks relative to controls. These results
demonstrate the existence of a generalized inhibitory deficit
in PD, which suggest that PD is a disorder of inhibition as
well as activation and that in situations of conflict, execu-
tive control over responses is compromised.
Keywords Fronto-striatal circuits � Parkinson’s disease �Subthalamic nucleus � Inhibition � Stop signal task
Introduction
The basal ganglia and the frontal cortex are intimately
connected via the fronto-striatal circuits, and direct, indi-
rect and hyperdirect pathways between these structures
have been delineated (Alexander et al. 1986; Middleton
and Strick 2000). The direct and indirect pathways have
been proposed to constitute an ideal system for response
selection and initiation under competition or conflict, with
the indirect pathway via the subthalamic nucleus (STN)
inhibiting inappropriate responses to allow selection and
initiation of the appropriate response through the direct
pathway (e.g. Chevalier and Deniau 1990; Redgrave et al.
1999). In Parkinson’s disease (PD), degeneration of dopa-
minergic neurons in the pars compacta of the substantia
nigra alters the balance of activity in the direct and indirect
pathways, the net effect of which increased inhibitory
outflow from the internal segment of the globus pallidus
I. Obeso � L. Wilkinson � M. Jahanshahi (&)
Cognitive-Motor Neuroscience Group, Sobell Department
of Motor Neuroscience and Movement Disorders, UCL Institute
of Neurology, The National Hospital for Neurology and
Neurosurgery, 33 Queen Square, London WC1N 3BG, UK
e-mail: [email protected]
E. Casabona � M. L. Bringas � M. Alvarez � L. Alvarez �N. Pavon � R. Macıas
Movement Disorders and Neurophysiology Units, Centro
Internacional de Restauracion Neurologica (CIREN),
La Habana, Cuba
M.-C. Rodrıguez-Oroz � J. A. Obeso
Department of Neurology, Clınica Universitaria and Medical
School of Navarra, Neuroscience Centre, CIMA,
University of Navarra, Pamplona, Spain
M.-C. Rodrıguez-Oroz � J. A. Obeso
Centro de Investigacion Biomedica en Red Sobre Enfermedades
Neurodegenerativas (CIBERNED), Instituto Carlos III,
Ministerio de Investigacion y Ciencias, Barcelona, Spain
Present Address:L. Wilkinson
Brain Stimulation Unit, National Institute of Neurological
Disorders and Stroke, Bethesda, MD 20892-1430, USA
123
Exp Brain Res (2011) 212:371–384
DOI 10.1007/s00221-011-2736-6

(GPi), and a resultant reduced activation of cortical pro-
jection sites via the thalamus (DeLong 1990). Conse-
quently, frontal areas such as the pre-supplementary motor
area (pre-SMA) are underactivated in PD during self-gen-
erated or self-initiated movement (Playford et al. 1992;
Jahanshahi et al. 1995). In the indirect pathway, reduced
external segment of the globus pallidus (GPe) output is
associated with increased and abnormally synchronized
STN neuronal activity (e.g. Vila et al. 1997; Kuhn et al.
2009), and this abnormal activity is considered a key
pathophysiological feature of PD and the rationale for
surgical treatment of the disorder with deep brain stimu-
lation (DBS) of the STN.
The stop signal reaction time (RT) task (Logan and
Cowan 1984) has been widely used for the assessment of
inhibition of overt motor responses. A conditional version of
the stop signal RT task (Aron et al. 2007) has the advantage
that it allows concurrent assessment of response initiation,
response inhibition and response initiation under conflict.
The conditional stop signal RT is a two-choice RT requiring
responses to left or right pointing arrows. For each partici-
pant, one of the arrows is assigned as the ‘critical’ and the
other as the ‘non-critical’ direction, with the instruction to
inhibit responses when a stop signal is presented after a
‘critical’ go stimulus but to respond as usual when a stop
signal follows a go stimulus in the ‘non-critical’ direction.
This conditional task requires response initiation on some
trials (‘critical’ or ‘non-critical’ go trials), suppression of a
prepared response on other trials (‘critical’ stop signal trials)
and response initiation under conflict (‘non-critical’ stop
signal trials) on yet another set of trials. Of interest in rela-
tion to the pathophysiology of PD is the demonstration in an
imaging study with healthy participants that on the condi-
tional stop signal task, relative to the Go trials, both suc-
cessful stopping on ‘critical’ stop trials and conflict induced
slowing on ‘non-critical’ stop trials are associated with
significant activation of the ‘braking’ network of right
inferior frontal cortex (IFC), STN and pre-SMA (Aron et al.
2007). Based on correlations between activation in the IFC
and STN with each other and with the stop signal RT
(SSRT), it was proposed that these two areas formed an
inhibition network for stopping. In contrast, the pre-SMA
activation did not significantly correlate with SSRT but
increased in proportion to the conflict induced slowing of
RTs on non-critical stop signal trials and hence was con-
sidered to play a role in conflict monitoring/resolution.
In light of evidence for the overactivity of the STN and
underactivity of the pre-SMA as pathophysiological fea-
tures of PD and that PD patients exhibit deficits in inhibitory
control (Cooper et al. 1994; Gauggel et al. 2004; Bokura
et al. 2005; Baglio et al. 2009; Beste et al. 2009) and in
conflict resolution (Praamstra and Plat 2001; Seiss and
Praamstra 2004, 2006; Wylie et al. 2005, 2009a), the aim of
this study is to investigate both inhibition of a prepared
response and response initiation under conflict in the same
PD cohort of patients. We used the conditional stop signal
RT task to determine whether movement-related underac-
tivation of the pre-SMA and overactivity of the STN in PD
produce consistent or dissociable effects on response initi-
ation (Go RTs), response inhibition (SSRT) and response
initiation under conflict (conflict induced slowing, CIS).
Furthermore, to test the generality of inhibitory deficits in
PD across motor and cognitive domains, we also included
three cognitive tasks requiring executive control: the
Stroop, random number generation and the Hayling Sen-
tence Completion test in which selection of the correct
response necessitates inhibition of alternative competing,
prepotent or habitual responses. We predicted that (1)
inhibition of a prepared response as well as response initi-
ation under conflict would both be impaired on the condi-
tional stop signal RT in PD relative to age-matched healthy
controls and (2) relative to controls, patients with PD would
show concurrent deficits in volitional suppression of pre-
potent responses across both motor and cognitive tasks.
Methods
Participants
Eighteen right-handed individuals with a clinical diagnosis
of idiopathic PD (12 men, mean age = 55.72, SD = 6.7)
were recruited from the outpatients clinic at the Centro In-
ternacional de Restauracion Neurologica (CIREN), Havana,
Cuba. The patients were consecutive referrals for functional
neurosurgery and were assessed prior to surgery. The Uni-
fied Parkinson’s Disease Rating Scale (UPDRS) was used to
obtain a measure of disease severity when patients were off
their usual medication. All patients met the UK Brain Bank
diagnostic criteria for PD (Hughes et al. 1992), had a good
dopaminergic response, and absence of atypical symptoms.
The sample had advanced illness (UPDRS III M = 42.00,
SD = 12.4). All patients were non-demented as demon-
strated by scores[26 on the mini–mental state examination
(MMSE; Folstein et al. 1975). The patients were also
screened for clinical depression (scores [18) on the Beck
Depression Inventory (BDI; Beck et al. 1961). While three
patients had a score above the cut-off, these high scores were
related to the negative impact of living with PD, and there
was no evidence of clinical depression as established during
neurological examination and the NeuroPsychiatric Inven-
tory (NPI) interview. The patients were treated with levo-
dopa and dopamine agonists and were assessed on their
usual medication. For each patient, the levodopa equivalent
dose (LEDD) was calculated following the procedures of
Williams-Gray et al. (2007).
372 Exp Brain Res (2011) 212:371–384
123

Twenty-nine volunteers (16 men, 23 right-handed) aged
between 45 and 68 (M = 57.13, SD = 6.8) took part in the
study. The control group was recruited via advertisement
and was assessed at the UCL Institute of Neurology, UK,
by the same investigator (IO) who completed the patient
assessments in Havana. None of the controls had any
neurological disorder or history of psychiatric illness or
drug or alcohol abuse, and none were taking any medica-
tion. Information about the patients with PD and the con-
trols is presented in Table 1.
The study was approved by The National Hospital for
Neurology and Neurosurgery and Institute of Neurology
Joint Research Ethics Committee as well as the Cuban
National Ethical Committee. Informed consent was
obtained prior to participation in the study.
Conditional stop signal task
The conditional stop signal task employed here was—with
the exception of the use of a visual rather than auditory stop
signal—otherwise identical to that used by Aron et al.
(2007). It allowed measurement of both (a) how well a
participant can inhibit an already initiated response and
(b) the time it takes to initiate a response under conditions
of conflict or ‘conflict induced slowing’. The stop signal
task consisted of a combination of Go and Stop trials. On
Go trials, a left (or right) pointing green arrow was pre-
sented 500 ms after presentation of a black circular fixation
point in the centre of a computer screen, and participants
had to respond as fast as possible using their index and
middle fingers of their dominant hand to press a left or right
key. On Stop trials (25% of all trials), a stop signal (red
cross) was presented after a variable stop signal delay
(SSD) following the green arrow. For each participant,
either the left or right pointing arrows were designated as
the ‘critical’ direction. When a stop signal was presented
following an arrow/go signal in the ‘critical’ direction, the
participants had to stop their response. In contrast, when a
stop signal was presented following an arrow/go signal in
the ‘non-critical’ direction, participants were instructed to
ignore the stop signal and respond to the ‘non-critical’ go
signal. For 10/18 PD patients and 14/29 controls, the
‘critical’ direction was left, and for 8/18 PD patients and
15/29 of the controls, it was right. There were three blocks
of trials consisting of 32 Stop and 96 Go trials per block
(128 total trials per block) presented in a randomized order.
In each block, the number of left and right pointing arrows
was equal. Durations from 0.5 to 4 s were inserted as null
events between the Stop or Go trials.
A trial began with the presentation of a black circular
fixation point in the centre of the screen. This was replaced
after 500 ms by the presentation of a green arrow in the
centre of the screen. The green arrow remained on the
screen a maximum of two seconds (limited hold), followed
by the background screen during the null period. If a par-
ticipant responded within the limited hold length, the arrow
disappeared, leaving the background screen and the null
period.
On Stop trials, the green arrows were replaced by a red
cross at some stop signal delay (SSD) after the green arrow.
The SSD value for the Stop trials was sampled from one of
four staircases, changing dynamically throughout the task
based on the participant’s behaviour. Initially, the four
staircases started with SSD values of 100, 150, 200 and
250 ms, respectively. For each SSD, successful inhibition
of a response on a Stop trial made inhibition more difficult
on the next Stop trial by increasing the SSD by 50 ms. In
contrast, if the response was not successfully inhibited,
then inhibition became easier by decreasing the SSD by
50 ms. Staircases of four step-up and step-down algorithms
were used in this way to ensure convergence to P(inhibit)
of 50% by the end of the three blocks. This allowed us to
obtain measures of each individual’s mean SSD when the
probability of them successfully inhibiting their behaviour
is at 50%. This is required to estimate participants’ SSRT.
As each block had 32 Stop trials, 16 of these Stop trials
were for the ‘critical’ direction. Therefore, each staircase
moved four times within each block for the Stop trials of
the ‘critical’ direction. SSDs for the Stop trials of the ‘non-
critical’ direction were yoked to the ‘critical’ direction
values. The staircases were independent but were randomly
mixed in a block of trials.
Participants were instructed that the most important
aspect of the task was to respond to the green arrows by
pressing the correct response key as fast and as accurately
as possible; while at the same time, they should also look
Table 1 Demographic and clinical characteristics of the patients with Parkinson’s disease (PD) and healthy controls
Group Age MMSE BDI UPDRS III (off med) Disease duration (years)
Parkinson’s disease (n = 18) 55.72 (6.7) 28.11 (1.3) 14.00 (8.2) 42.00 (12.4) 9.39 (2.9)
Controls (n = 29) 57.03 (6.9) 29.59 (.6) 6.07 (5.1)
P .41 .001 .001
Mean values are shown; the numbers in parentheses are standard deviations. MMSE mini–mental state examination, BDI Beck Depression
Inventory, UPDRS III Unified Parkinson’s Disease Rating Scale part III
Exp Brain Res (2011) 212:371–384 373
123

out for the appearance of the red cross and try to withhold
their response to the green arrow on the occasions when the
red cross followed a green arrow pointing in the ‘critical’
direction. In addition, participants were informed that due
to the variable nature of the SSD, it would not always be
possible to stop their response on ‘critical’ stop trials.
Finally, participants were specifically instructed not to let
their performance on the stopping task interfere with their
performance on the Go task, and in particular, they were
asked not to delay their performance on the Go task in
order to improve their chances of stopping after green
arrows pointing in the ‘critical’ direction. Following these
instructions, 20 practice trials were completed. Each block
was preceded by an instruction screen and ended with
presentation of the mean correct reaction time (RT) and the
number of discrimination errors on the Go trials.
Several RT measures were computed to the nearest ms:
mean Go RT for both ‘critical’ and ‘non-critical’ trials,
percentage of Stop trials with successful inhibition (Stop-
Inhibit) for the ‘critical’ direction, number of Stop trials
resulting in inhibition for the ‘non-critical’ direction and
mean RT on Stop trials without successful inhibition
(StopRespond) for both ‘critical’ and ‘non-critical’ direc-
tions. Similar to Aron et al. (2007), one of the main
comparisons of interest was ‘non-critical’ Stop trials minus
‘non-critical’ Go trials, which is the measure of ‘conflict-
induced slowing’. Additionally, we also looked at suc-
cessful inhibition using the standard Race Model (Logan
and Cowan 1984) to compute the stop signal reaction time
(SSRT). We estimated SSRT by subtracting the average
SSD from the mean correct ‘critical’ Go RT. Due to the
dynamic adjusting of the SSD, we computed the average
SSD for each participant, using the values of the four
staircases after the participant had converged on 50%
P (inhibit) which similar to Aron et al. (2007) was aver-
aged from the mean values for the last six moves in each
staircase. Discrimination errors (using the incorrect finger
to press the wrong key for the right or left stimulus pre-
sented) and omission errors (failure to respond to a go
signal in the ‘critical’ or ‘non-critical’ direction) and
commission errors (responding to a stop signal presented
after a ‘critical’ go trial which resulted in the ‘critical’
StopRespond RTs) were recorded.
Other tests of inhibition
Hayling sentence completion test (Burgess and Shallice
1997)
In the Hayling sentence completion test, participants
complete a series of sentences from which the last word is
missing. For the assessment of the patients in Cuba, we
produced a Spanish version of the Hayling test using the
same sentences where possible or constructing closely
related sentences with a high-frequency missing word in
Spanish, which were pre-tested, on 50 Cuban healthy
participants. The test has two sections: (1) Section A:
response initiation: participants were instructed to provide
an appropriate word to complete a sentence from which the
last word is missing. For example, ‘the captain wanted to
stay with the sinking…’ for which the word ‘ship’ would
be an appropriate response and (2) Section B: response
suppression: For each sentence read out, participants were
required to provide a word that made no sense at all in the
context of the sentence. For example, ‘Most cats see very
well at …’ for which ‘night’ is the high-frequency word
that has to be suppressed and ‘banana’ would be an
appropriate and unrelated response. The response time (i.e.
mean response latency across all items measured with a
stopwatch) was measured separately for sections A and B.
In addition, in section B, for each item, an error score was
obtained by classifying responses as belonging to category
A (connected to the meaning of the sentence) or category B
(somewhat related to the meaning of the sentence). Using
the scoring guidelines and normative data provided in the
Hayling manual, RTs for sections A and B were converted
using the normative Hayling table, and scaled scores for
the combined Type A and B errors in section B were
obtained. The higher the converted scores are, the better
performance in the test.
Stroop interference test (Stroop 1935)
The Stroop test assesses the ability to inhibit the prepotent
response of reading words in order to produce the alter-
native response of naming the colour of ink the words are
printed in. Based on the Delis–Kaplan battery (2001), we
used the colour naming as the control condition along with
the Stroop interference condition. In the control condition,
the participant had to name aloud the colour of 100 col-
oured rectangles printed in blue, red or green. In the Stroop
interference condition, the participant had to name the
colour of ink of the colour words blue, red or green printed
in an incongruent colour. A Spanish version of the task was
used for the interference condition where colour words
were written in Spanish. For each condition, the total
completion time, the number of self-corrected and uncor-
rected errors was recorded.
Random number generation (Jahanshahi et al. 1998, 2000)
Participants were asked to generate numbers from 1 to 9 in
a random fashion for 100 trials in synchrony with a pacing
visual stimulus presented at 1 Hz. The concept of ran-
domness was explained with the analogy of picking out
numbers out of a hat with replacement. The 100 numbers
374 Exp Brain Res (2011) 212:371–384
123

generated by the participant were recorded as well as the
total time taken to generate them. Participants generated
numbers in their mother tongue. To measure randomness,
we obtained count scores that are measures of seriation
during random number generation (RNG), based on the
method of Spatt and Goldenberg (Spatt and Goldenberg
1993). Count Score 1 (CS1) and Count Score 2 (CS2)
measure the number of times the participant counts in
ascending or descending series, respectively, in steps of
one or steps of two. In calculating the count scores, the
sequence length is squared to give higher weights to runs of
longer sequences. High count scores indicate an inability to
suppress habitual counting tendencies during RNG.
Results
Patient and control groups did not differ significantly in
terms of age [t(45) = -.82, P = .41], sex distribution
[v(1) = 1.65, P = .19] or handedness [t(21.3) = -1.47,
P = .15]. Although none of the PD patients were demented
or clinically depressed, nevertheless as a group they scored
significantly lower than controls on the MMSE and signifi-
cantly higher on the BDI (see Table 1). One patient and one
control were excluded as they failed to reach the criterion of
successful inhibition on approximately 50% of trials.
Conditional stop signal task
Critical trials
The data relating to the conditional stop signal task are
presented in Table 2. The dynamic adjustment of SSD
converged on probabilities of inhibition that were close to
50% and these percentages did not differ significantly
between groups (Table 2), which is a prerequisite for cor-
rect interpretation of the results. For the patients, the mean
SSD (M = 183.69, SD = 98.2) was shorter than the mean
SSD for the controls (M = 209.15, SD = 95.2), but the
difference was not significant [t(45) = -.88, P = .38].
Using the Race Model, SSRT was estimated at
289.86 ms (SD = 48.1) for controls and 382.41 ms
(SD = 94.1) for PD patients, with the patients having
significantly longer SSRTs than the controls [t(22.5) = 3.87,
Table 2 Mean reaction time (RT, in milliseconds) and error data for the conditional stop signal reaction time task for the patients with
Parkinson’s disease (PD) and the healthy controls
Trial description Measure PD patients Controls P
‘Critical’ direction
Goa RT 566.10 (124.0) 495.69 (89.7) .02*
StopInhibit % Correct inhibition 53.56 (10.9)% 52.59 (10.1)% .76§
StopRespond RT 568.96 (118.3) 465.85 (79.6) .003**
Go errors Number of omissions 5.06 (6.8) .62 (1.0) .001§**
‘Non-critical’ direction
Go RT 513.32 (96.8) 433.14 (44.3) .003**
StopInhibit % Incorrect inhibition 7.32 (5.7)% 3.10 (4.4)% .01*
StopRespondb RT 678.61 (164.7) 552.30 (78.9) .006*
Go errors Number of omissions 3.44 (4.4) .45 (.7) .001§*
Other variables
Go discrimination errors Number of errors 1.83 (3.4) .45 (.6) .11§
Mean SSD Time 183.69 (98.2) 209.15 (95.2) .38
SSRT RT 382.41 (94.1) 289.86 (48.1) .001**
Conflict induced slowing (CIS) Time 165.29 (93.3) ms 119.15 (67.6) ms .05*
Standard deviations are given in brackets
SSD the average stop signal delay, computed from four staircases (see ‘Methods’) at the point at which P(inhibit) = *50%
SSRT is computed for each participant by subtracting the mean SSD from the mean critical Go RT
Go omission errors failure to respond on a go trial
Go discrimination errors pressing the response key in the opposite direction indicated by the stimulus
CIS the cognitive induced slowing measured by subtracting mean non-critical Go RTs from mean non-critical StopRespond RTs
** Ps \ .005; * Ps \ .05; § Mann–Whitney test used. The rest were independent samples t testsa Mean ‘critical’ Go RTs are significantly slower than mean ‘non-critical’ Go RTs (P \ .01) between groups comparisonb Mean ‘critical’ StopRespond RTs were significantly slower than mean ‘non-critical’ StopRespond RTs in controls (P \ .01) but not in the PD
group
Exp Brain Res (2011) 212:371–384 375
123

P = .001] (Fig. 1). In addition, as expected the StopRe-
spond RTs (M = 465.85, SD = 79.6) of the controls were
faster than their Go RTs (M = 495.69, SD = 89.7), a
difference that approached significance (t(28) = 1.9,
P = .06) by an average of 29 ms; whereas for the PD
patients, StopRespond RTs (M = 568.96, SD = 118.3) did
not differ from Go RTs (M = 566.10, SD = 124.0,
t(17) = -.11, P = .90).
On ‘critical’ Go trials, controls responded quickly and
accurately (Table 2; Fig. 1). In contrast, PD patients
responded significantly slower on ‘critical’ Go trials than
controls, and they made significantly more omission errors
(Table 2). Furthermore, PD patients made more discrimina-
tion errors than controls, albeit non-significantly so (Table 2).
‘Non-critical’ trials
For ‘non-critical’ trials, one of the main measures of
interest is the difference between mean ‘non-critical’ Go
RTs and mean ‘non-critical’ StopRespond RTs as this
provides a measure of ‘conflict induced slowing’ (CIS). To
establish whether RTs differed significantly in the two
groups across trial types, an ANOVA was performed on
mean ‘non-critical’ RT with Trial Type (Go vs. StopRe-
spond) as a within-subject variable and Group as a
between-groups variable. This analysis revealed a signifi-
cant main effect of Trial Type [F(1,45) = 146.26, P \ .001]
because, overall, ‘non-critical’ StopRespond RTs were
slower than the non-critical Go RTs, which is indicative of
the CIS effect due to the conditional nature of stopping.
There was also a significant main effect of Group
[F(1,45) = 14.96, P \ .001], showing that, overall, the
Control group had faster ‘non-critical’ RTs relative to the
PD group. Most importantly, there was a significant
Group 9 Trial Type interaction [F(1,45) = 3.84, P = .05],
indicating that the magnitude of the CIS effect was sig-
nificantly larger in the PD than in the Control group.
In light of the significant Group 9 Trial Type interac-
tion, post hoc analysis of the group differences on the
‘non-critical’ direction was undertaken. Similar to critical
Go trials, PD patients responded significantly slower on
‘non-critical’ Go trials than controls (Table 2). In addition,
the ‘non-critical’ StopRespond RT, [recall in this instance
is the RT for correct responses following a ‘non-critical’
stop signal, (i.e. a ‘to be ignored’ stop signal)], was also
significantly longer in PD patients relative to controls
(Table 2). A difference score measure of CIS was calcu-
lated for each participant by subtracting mean ‘non-critical’
Go RTs from the mean ‘non-critical’ StopRespond RT.
Therefore, a positive CIS difference score is indicative of
higher CIS or greater slowing under conflict. The mean CIS
in the PD group (M = 165.29, SD = 93.3) was signifi-
cantly greater for the controls (M = 119.15, SD = 67.6,
t(45) = 1.96, P = .05) (Fig. 2).
Patients made significantly more errors of omission on
‘non-critical’ Go trials than controls (Table 2). Further-
more, PD patients incorrectly inhibited their responses and
failed to respond on ‘non-critical’ Stop trials significantly
more often than controls (Table 2).
In both groups, Go RTs in the ‘critical’ direction were
slower than Go RTs in the ‘non-critical’ direction, with the
differences being significant for the controls (t(28) = 5.69,
P \ .001) and the PD patients (t(17) = 2.52, P = .02).
Controlling for group differences in Go RTs, depression
and global cognitive ability
To control for group differences in Go RTs, ANCOVAs were
performed to compare group differences in SSRT and CIS
after co-varying out the critical Go RTs. Group differences in
the SSRT, [F(1,44) = 13.22, P \ .01] and the CIS, [F(1,44) =
3.83, P = .05] remained significant even after co-varying out
group differences in ‘critical’ Go RTs. These results suggest
that the delayed inhibition (SSRT) and delayed response
initiation under conflict (CIS) of the PD patients were not
solely attributable to their slowed Go RTs.
The two groups also differed significantly in MMSE and
BDI scores. Therefore, to control for the contribution of
Fig. 1 a Mean RTs in
milliseconds in the ‘critical’
direction for the Go and
StopRespond (trials with a stop
signal on which participants
failed to stop and responded)
trials for PD patients and
healthy controls. b The mean
stop signal reaction times
(SSRT) for the patients (whitebars) and the healthy controls
(black bars). An asteriskindicates a significant difference
between groups. Error barsdepict standard errors
376 Exp Brain Res (2011) 212:371–384
123

cognitive impairment and self-reported depression to SSRT
and CIS, the group differences were re-examined using
these scores as covariates. SSRT differences between
groups remained significant after co-varying out the effect
of MMSE [F(1,44) = 10.49, P \ .01] and BDI [F(1,44) =
17.21, P \ .001]. The group differences in CIS did not
remain significant after co-varying out MMSE [F(1,44) =
4.48, P = .47] and approached significance after co-vary-
ing out BDI [F(1,44) = 3.59, P = .06].
Other tests of inhibition
Hayling sentence completion test
To compare the mean converted response times for Hay-
ling sections A (initiation) and B (inhibition) for the PD
(Section A converted response times: M = 5.24,
SD = 1.1, Section B converted response times: M = 5.24,
SD = 1.0) and controls (Section A converted response
times: M = 6.07, SD = .9, Section B converted response
times: M = 5.48, SD = .8), an ANOVA was performed on
Hayling Section (A vs. B) as a within-subject variable and
Group as a between-groups variable. This analysis revealed
a significant Group 9 Hayling Section interaction
[F(1,45) = 4.59, P = .03], indicating that the degree of
difference between performance on sections A and B (or
the ‘Hayling effect’) differed significantly between the
groups. Similarly, the main effects of Hayling Section
[F(1,45) = 4.59, P = .03] and Group were significant
[F(1,45) = 4.80, P = .03]. In light of the significant inter-
action, we compared the difference score in response times
between sections A and B of the Hayling (Hayling effect)
for the patients and controls. This difference score was
significantly higher/better for the controls than the PD
patients (t(45) = 2.96, P = .03), due to the fact that the
patients failed to modulate their response times as a func-
tion of Hayling section whereas the controls did so.
PD patients made significantly more Type A errors rela-
tive to controls (PD M = 14.69, SD = 14.7, Control
M = 5.18, SD = 6.5, Mann–Whitney U = -2.89,
P \ .01). The Type B errors were again significantly
greater in PD patients (M = 9.75, SD = 9.5) compared to
Controls (M = 1.39, SD = 1.4, Mann–Whitney U =
-4.01, P \ .001). These clearly indicate that PD patients
found it difficult to withhold the prepotent response in sec-
tion B of the Hayling test (Fig. 3).
The mean global scaled score on the Hayling test was
significantly better for the controls (Control M = 5.90,
SD = 1.1) than the PD patients (PD M = 4.06, SD = 1.6,
t(45) = -4.29, P \ .001), indicating impairment of
suppression of prepotent response for the PD patients rel-
ative to the controls. Using the MMSE and BDI as
covariates, the differences in Hayling global score between
the groups were still significant after co-varying out MMSE
[F(1,45) = 5.00, P = .03] and BDI [F(1,45) = 13.57,
P = .001].
Stroop interference task
Figure 4 shows the mean total completion time for the
control and inhibition conditions of the Stroop test, plotted
separately for the two groups. An ANOVA was performed
on mean total completion time with Condition (control vs.
interference) as a within-subject variable and Group as a
between-groups variable. This analysis revealed significant
main effects of Group [F(1,45) = 40.43, P \ .001]—
showing that PD patients were significantly slower than
controls across both Stroop conditions and Condition
[F(1,45) = 200.69, P \ .001]. The Group 9 Condition
interaction [F(1,45) = 6.15, P \ .01] was also significant,
Fig. 2 a Mean RTs in milliseconds in the ‘non-critical’ direction for
the Go and StopRespond trials (trials on which a stop signal was
presented which participants were instructed to ignore and respond)
for PD patients and the healthy controls. b Mean conflict induced
slowness (CIS) in milliseconds plotted for PD patients (white bars)
and healthy controls (black bars). An asterisk indicates a significant
difference between groups. Error bars depict standard errors
Exp Brain Res (2011) 212:371–384 377
123
indicating that the magnitude of the PD versus control
differences was differentially greater for the Stroop inter-
ference than the control task. In light of the significant
Group 9 Condition interaction, we examined group dif-
ferences in the difference score (interference - control),
which was significantly larger for the PD patients (M =
33.16, SD = 16.5) than controls (M = 20.66, SD = 13.0,
t(44) = 2.85, P \ .01), confirming a significantly greater
‘Stroop effect’ and conflict induced slowing for the patients
(Fig. 4). Furthermore, the Group 9 Condition interaction
remained significant after co-varying out MMSE [F(1,45) =
3.9, P = .05] and BDI [F(1,45) = 6.29, P = .01], showing
that the differences in the magnitude of the Stroop effect
between groups was significant above and beyond any
effects of cognitive impairment or depression.
For the interference condition, PD patients (M = 2.31,
SD = 4.1) made significantly more uncorrected errors than
controls (M = .18, SD = .6, Mann–Whitney U = -3.03,
P \ .01). For the control condition, there was a significant
difference between the number of uncorrected errors made
by PD patients (M = 1.28, SD = 1.2) and controls (M =
.03, SD = .18, Mann–Whitney U = -4.75, P \ .001). For
the interference condition, PD patients (M = 1.88,
SD = 1.5) made significantly more self-corrected errors
than controls (M = .50, SD = .9, Mann–Whitney U =
-3.28, P = .001). For the control condition, there was a
significant difference between the number of self-corrected
errors made by PD patients (M = 1.31, SD = 1.3) and
controls (M = .03, SD = .1, Mann–Whitney U = -4.42,
P \ .001).
Random number generation
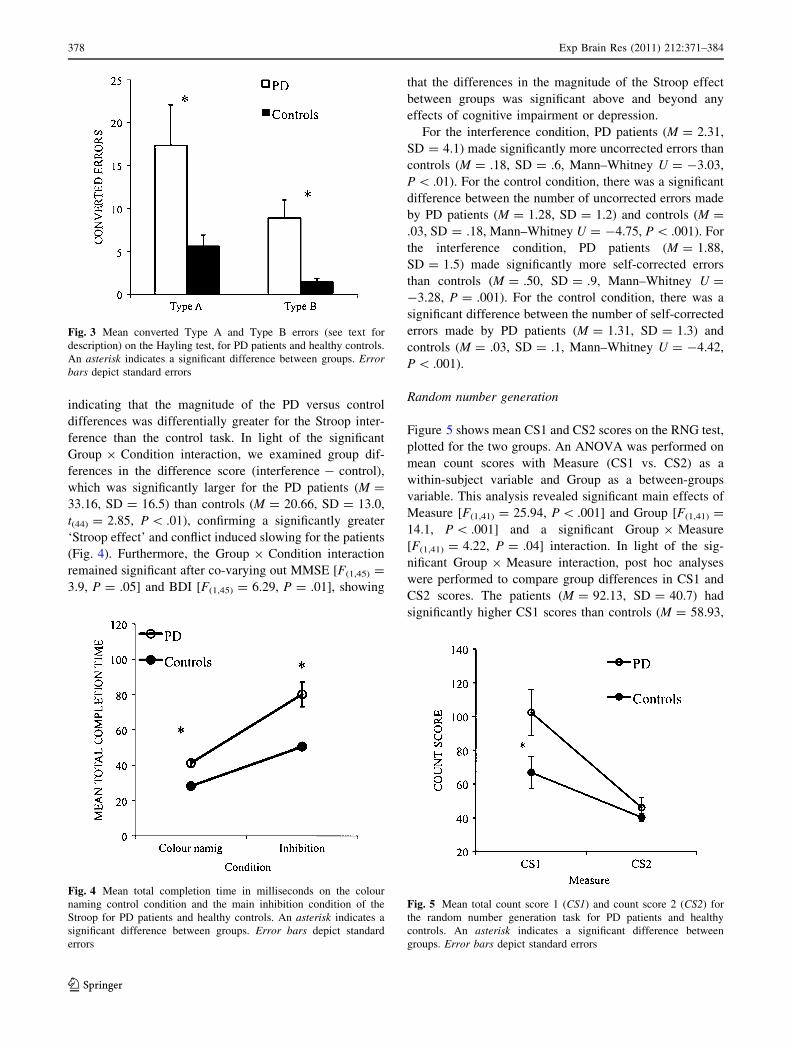
Figure 5 shows mean CS1 and CS2 scores on the RNG test,
plotted for the two groups. An ANOVA was performed on
mean count scores with Measure (CS1 vs. CS2) as a
within-subject variable and Group as a between-groups
variable. This analysis revealed significant main effects of
Measure [F(1,41) = 25.94, P \ .001] and Group [F(1,41) =
14.1, P \ .001] and a significant Group 9 Measure
[F(1,41) = 4.22, P = .04] interaction. In light of the sig-
nificant Group 9 Measure interaction, post hoc analyses
were performed to compare group differences in CS1 and
CS2 scores. The patients (M = 92.13, SD = 40.7) had
significantly higher CS1 scores than controls (M = 58.93,
Fig. 3 Mean converted Type A and Type B errors (see text for
description) on the Hayling test, for PD patients and healthy controls.
An asterisk indicates a significant difference between groups. Errorbars depict standard errors
Fig. 4 Mean total completion time in milliseconds on the colour
naming control condition and the main inhibition condition of the
Stroop for PD patients and healthy controls. An asterisk indicates a
significant difference between groups. Error bars depict standard
errors
Fig. 5 Mean total count score 1 (CS1) and count score 2 (CS2) for
the random number generation task for PD patients and healthy
controls. An asterisk indicates a significant difference between
groups. Error bars depict standard errors
378 Exp Brain Res (2011) 212:371–384
123

SD = 26.3 t(41) = 3.2, P \ .01), but the two groups did
not differ in terms of CS2 scores (PD: M = 47.06,
SD = 25.0; Controls M = 39.78, SD = 12.9, t(41) = 1.2,
P = .21). The Group 9 Measure interaction was no
longer significant when group differences in MMSE
[F(1,41) = 1.5, P = .22] and BDI scores were covaried out
[F(1,41) = 1.5, P = .26].
Correlational and regression analysis
For the PD group, we examined the association between
the two key variables from the stop signal task that is
SSRT and CIS and other variables of interest on this task
(omission errors on ‘critical’ and ‘non-critical’ direction,
discrimination errors) and demographic and clinical vari-
ables (age, UPDRS, LEDD, duration of illness, MMSE
and BDI) and cognitive measures of inhibition (Hayling
Type A, Type B errors and global score; Stroop control
and interference response times; and RNG CS1 scores).
Table 3 shows both significant and non-significant corre-
lations between either SSRT or CIS and the demographic,
clinical and cognitive inhibition measures. None of the
remaining correlations were of a sizeable magnitude
(above 0.47) or significant.
To identify the variables that contributed to the variance
of SSRT and CIS, two multiple regression analyses were
performed in the PD group. In the first regression analysis,
the dependent variable was SSRT. The independent vari-
ables entered as potential predictors were demographic or
clinical variables including age, MMSE, mean LEDD and
the Hayling section B Type A error score. This regression
analysis was significant [F(6,15) = 5.74, P = .05,
R2 = .74]. The Hayling Type A errors [b = .511, t(12) =
2.23, P = 0.043] predicted 26% of the variance of SSRT
(adjusted R2) in PD. In the second regression analysis, CIS
was the dependent variable and in addition to age, and
LEDD, MMSE and Stroop interference condition com-
pletion times, the two variables which had significant
correlations with CIS were the independent variables.
MMSE emerged as the only significant predictor [F(6,15) =
7.16, P = .018, R2 = .338, b = -.582, t(12) = 2.68,
P = .018], which accounted for 29% of the variance of
CIS (adjusted R2) in PD.
Discussion
We found impaired inhibition on the conditional stop
signal task in PD patients relative to age-matched controls.
This impaired motor inhibition in PD was also accompa-
nied by deficits in volitional suppression of habitual or
prepotent responses on three cognitive tests the Hayling
sentence completion test, the Stroop, and fast-paced Ta
ble
3C
orr
elat
ion
mat
rix
sho
win
gas
soci
atio
no
fd
emo
gra
ph
ican
dcl
inic
alfe
atu
res
of
Par
kin
son
’sd
isea
se(P
D)
and
cog
nit
ive
task
mea
sure
sw
ith
SS
RT
and
CIS
Ag
eU
PD
RS
III
LE
DD
Du
rati
on
of
illn
ess
MM
SE
BD
IG
oer
rors
crit
ical
Go
erro
rn
on
-
crit
ical
Dis
c
erro
r
SS
RT
Hay
lin
gT
yp
e
Aer
ror
Hay
lin
gT
yp
e
Ber
ror
Hay
lin
g
sco
re
Str
oo
p
I
Str
oo
p
II
RN
G
CS
1
CIS
.05
2.2
54
.01
3.3
69
-.6
61
-.3
26
-.0
07
.05
0.2
91
.00
4.0
96
-.0
26
-.2
75
.28
8.4
90
-.1
81
.83
.34
.96
.13
.01
.18
.97
.84
.24
.98
.70
.92
.26
.27
.05
.50
SS
RT
.16
0.2
18
-.2
78
.06
9-
.26
3-
.13
9.6
66
.52
0.5
65
.45
9-
.33
8-
.10
8.4
74
.45
4-
.38
1
.52
.41
.29
.78
.29
.58
.00
.02
.01
.05
.17
.66
.06
.07
.14
Pea
rso
n’s
corr
elat
ion
coef
fici
ent
inre
gu
lar
fon
t;P
val
ues
init
alic
fon
t.S
ign
ifica
nt
corr
elat
ion
sar
em
ark
edin
bo
ld.
UP
DR
SII
IU
nifi
edP
ark
inso
n’s
Dis
ease
Rat
ing
Sca
lem
oto
rp
art,
LE
DD
lev
od
op
aeq
uiv
alen
td
aily
do
se,
MM
SE
min
i–m
enta
lst
ate
exam
inat
ion
,B
DI
Bec
kD
epre
ssio
nIn
ven
tory
,C
ISco
nfl
ict
ind
uce
dsl
ow
nes
s,S
SR
Tst
op
sig
nal
reac
tio
nti
me,
Dis
cer
ror
dis
crim
-
inat
ion
erro
rs,
Str
oo
pI
con
tro
lco
nd
itio
n,
Str
oo
pII
inte
rfer
ence
con
dit
ion
,R
NG
CS
1ra
nd
om
nu
mb
erg
ener
atio
nco
un
tsc
ore
1
Exp Brain Res (2011) 212:371–384 379
123

random number generation relative to controls. The second
main finding is that PD patients had worse conflict reso-
lution than controls on the conditional stop signal task.
Finally, the prolonged SSRT in PD is not simply due to
motor slowness, reduced cognitive efficiency or depres-
sion, as the group differences from healthy controls
remained significant even after controlling for motor
slowness (Go RTs), MMSE or BDI scores.
We have shown, for the first time, deficits in both
response inhibition and conflict resolution in the same
sample of PD patients, supporting previous evidence on the
role of basal ganglia in executive control of action. Motor
inhibition and conflict resolution are two distinct but inter-
related executive processes, since response initiation under
conflict often requires inhibition of alternative competing
responses. In fact, as noted above, imaging has shown that
on the conditional stop signal RT task used here, both
successful inhibition of responses on ‘critical’ stop trials
and conflict induced slowing on ‘non-critical’ stop trials
are associated with activation of the ‘braking’ network of
right IFC, STN and pre-SMA (Aron et al. 2007). Our
results showing that PD patients have deficits on both
motor inhibition and conflict resolution are consistent with
this and also provide further evidence for a generalized
inhibitory deficit in PD patients and that in situations of
conflict, executive control over responses is compromised
in PD. Below, we will first discuss our findings in relation
to delayed motor inhibition and then conflict induced
slowing in PD.
Delayed inhibition in PD
The PD patients had significantly longer SSRTs than the
age-matched healthy controls, and group differences
remained significant even after co-varying out group dif-
ferences in Go RTs and MMSE, and BDI thus indicating
that the prolonged SSRTs in PD patients were not simply
due to motor slowness, reduced cognitive efficiency or
depression. Our findings on the conditional stop signal RT
task are consistent with the results of Gauggel et al. (2004)
who found significantly longer SSRTs in PD on the standard
stop signal RT task. Similar prolongation of the stop signal
RT relative to orthopaedic controls has been reported in 8
patients with focal lesions of the basal ganglia (Rieger et al.
2003). Evidence from other tasks requiring inhibition of
prepotent responses such as go no go RT (Cooper et al.
1994; Bokura et al. 2005; Baglio et al. 2009; Beste et al.
2009), anti-saccade (Chan et al. 2005; Rivaud-Pechoux
et al. 2007) and flanker (Praamstra and Plat 2001; Seiss and
Praamstra 2004, 2006; Wylie et al. 2005, 2009a) tasks also
supports impairment of inhibitory processes in PD.
The ‘critical’ StopRespond trials are of interest since
these are trials on which the participants failed to inhibit
the response when a stop signal was presented and thus
represent a failure of inhibition. The StopRespond RTs
were significantly slower for the PD patients than the
controls. Furthermore, for the healthy controls, the ‘criti-
cal’ StopRespond RTs were faster than their ‘critical’ Go
RTs by an average of 29 ms, suggesting that the failure to
inhibit the response on these stop signal trials was due to
their faster speed of responding to the Go signal on these
trials. In contrast, such a speed advantage for ‘critical’
StopRespond RTs over ‘critical’ Go RTs was not observed
for the PD patients. This implies that for the PD patients,
failure of inhibition is not simply due to faster RTs on
StopRespond trials, but that other processes such as a
failure to trigger the inhibitory process or lapses of atten-
tion may also be at play. This possibility requires further
investigation in future studies.
Different types of inhibition have been proposed, the
most common distinction being between ‘volitional’ and
‘reactive’ inhibition (e.g. Harnishfeger 1995; Nigg 2000).
Volitional inhibition is intentional and often effortful. It is
necessary for self-control of behaviour, such as stopping
oneself from having another glass of wine or eating another
sweet. Volitional inhibition is also often required for
executive control in conditions involving conflict or inter-
ference from competing responses (e.g. Harnishfeger 1995;
Nigg 2000). Our results provide evidence for impairment
of volitional inhibition in tasks involving interference from
competing responses and hence requiring executive control
in patients with PD. Relative to controls, PD patients were
significantly impaired on the Hayling and Stroop tasks,
similar to previous findings (Brown and Marsden 1988;
Brown and Marsden 1991; Bouquet et al. 2003). A parallel
inability to suppress prepotent and habitual responses and
to engage in strategic response selection is observed during
RNG, during which PD patients produce higher count
scores than matched controls (Brown et al. 1998; Jahan-
shahi et al. 2000; Dirnberger et al. 2005). Our aim in
including the Hayling, Stroop and RNG tasks in the present
study was to demonstrate the cross-task generality of def-
icits in suppression of prepotent responses across tasks
involving both motor (stop signal) and cognitive (Hayling,
Stroop, RNG) volitional inhibition in PD patients. Fur-
thermore, Type A errors on the Hayling task correlated
positively and significantly with SSRT, and the completion
time on the Stroop interference task correlated negatively
and significantly with CIS suggesting an association
between the impairments in motor and cognitive volitional
inhibition in PD.
Response initiation under conflict in PD
The magnitude of the conflict induced slowing/interference
was significantly greater in PD than controls, consistent
380 Exp Brain Res (2011) 212:371–384
123

with previous evidence of deficits in executive control in
PD on tasks involving conflict and requiring volitional
suppression of prepotent responses. While the error rate was
generally low, PD patients made significantly more errors of
omission and discrimination errors on the stop signal RT
task than the controls. Furthermore, instead of ignoring the
stop signal and responding on all ‘non-critical’ trials, PD
patients incorrectly inhibited their responses and failed to
respond on a significantly larger proportion of the ‘non-
critical’ StopRespond trials than the controls. These higher
error rates in PD also suggest the increased susceptibility of
their responses to situations of conflict. These higher error
rates in PD than in controls are also consistent with the
proposal that the basal ganglia together with the anterior
cingulate contribute to error monitoring and the error-rela-
ted negativity which is reduced in PD, thus indicating def-
icits in error detection (Falkenstein et al. 2001).
Inherent in the ‘non-critical’ StopRespond trials is a
conflict between stopping and responding. Thus, on these
trials, the conflict revolves around whether to stop or not,
that is whether to ignore the stop signal (as per experi-
mental instructions) and initiate the prepared response or to
inhibit it. The fact that the StopRespond RTs were the
slowest RTs in both the patient and control groups suggests
that this conflict was operational for all participants.
However, the impact of this conflict was significantly
greater for the PD patients than controls. In addition, as
noted above, the PD patients incorrectly failed to respond
on a significantly larger proportion of the ‘non-critical’
StopRespond trials than the controls.
Similarly, on the Eriksen flanker task, at least in some
studies PD patients were found to show greater interference
effects from the incongruent flanking stimuli (e.g. Praam-
stra and Plat 2001; Wylie et al. 2005, 2009a), particularly
under speed instructions (Wylie et al. 2009b) or when
patients had to inhibit distracters over extended delays
(Cagigas et al. 2007), although others have not found this
(Lee et al. 1999; Falkenstein et al. 2006). This exaggerated
flanker interference effect in PD has been interpreted to
reflect the patients’ reduced ability to suppress the auto-
matic activation of the conflicting response that then
interferes with selection of the correct response indicated
by the target stimulus (Praamstra et al. 1998; Praamstra and
Plat 2001; Wylie et al. 2009a).
In both the PD and control groups, Go RTs in the
‘critical’ direction were significantly slower than Go RTs
in the ‘non-critical’ direction. This indicates that in this
conditional stop signal RT task, the ‘context’ within which
the go signals were presented, whether on the ‘critical’ or
‘non-critical’ trials, influenced the participants’ strategy
and their speed of reaction to the stimuli. When presented
with an arrow in the ‘non-critical’ direction, participants
knew that they had to respond anyway even if a stop signal
was subsequently presented and, therefore, fast RTs were
appropriate. In contrast, on trials with an arrow presented
in the ‘critical’ direction, there was a possibility that it may
be followed by a stop signal, in which case the response
would need to be inhibited. Thus, on ‘critical’ trials when
participants were aware that they might have to stop a
response on a proportion of trials, their Go RTs were
slower. This ‘anticipatory’ or ‘strategic’ slowing of Go RTs
in the context of trials where a proportion of the responses
would have to be subsequently stopped, a sort of
responding with restraint, was present for both patients and
controls and is consistent with previous evidence of pro-
active adjustment of response strategies on the stop signal
RT task (Rieger and Gauggel 1999; Aron et al. 2007;
Verbruggen and Logan 2009a). Such proactive response
strategy adjustments become evident when stop signals are
expected, as a result of which response thresholds are
increased to slow down Go RTs and increase accuracy and
likelihood of stopping (Verbruggen and Logan 2009b).
Theoretical and clinical implications
In a series of elegant experiments in monkeys, Isoda and
Hikosaka (Isoda and Hikosaka 2007, 2008) have provided
evidence that the STN and pre-SMA play a role in
switching from automatic to controlled eye movements.
They proposed that the pre-SMA acts to suppress the
automatic saccade, resolve response conflict and facilitate
selection of the controlled response. The control signal for
switching originating from the pre-SMA is proposed to be
implemented by the STN through its connections with
other basal ganglia nuclei. Such a switch from automatic to
controlled processing captures the essence of executive
control, and the data from Isoda and Hikosaka (2007, 2008)
confirm the engagement of the fronto-striatal system in
higher-order control processes. The evidence from the
current study also suggests that the basal ganglia may be
involved in volitional inhibition necessary for suppression
of the prepotent response in the stop signal RT, Stroop,
Hayling or RNG tasks. There is also some evidence in
support of this proposal from DBS of the STN in PD which
has shown that acute manipulation of the output from the
STN affects performance on tasks involving executive
control such as the Stroop (Jahanshahi et al. 2000; Sch-
roeder et al. 2002; Witt et al. 2004), a go no go RT task
with high target frequency (Hershey et al. 2004), the stop
signal RT (van den Wildenberg et al. 2006; Ray et al.
2009), suppression of habitual counting during fast-paced
RNG (Thobois et al. 2007) and probabilistic decision-
making under conflict (Frank et al. 2007).
When required to engage in conflictual decision-mak-
ing, patients with PD were more impulsive with DBS of the
STN on than off (Frank et al. 2007). With DBS of the STN,
Exp Brain Res (2011) 212:371–384 381
123

PD patients had problems switching from automatic
habitual counting to strategic response selection during
fast-paced RNG, a deficit associated with increased acti-
vation of the GPi and reduced activation of the prefrontal
cortex and anterior cingulate and reduced pallidal-pre-
frontal/cingulate coupling relative to when the stimulators
were off. This hypothesis that in concert with the prefrontal
cortex, the basal ganglia, particularly the STN, are
involved in inhibitory control to enable either switching
from automatic to controlled processing or response
selection under conflict requires direct examination in
future studies.
The two major clinical manifestations related to abnor-
mal dopaminergic activation in PD are levodopa-induced
dyskinesias and impulse control disorders such as patho-
logical gambling, shopping, binge eating and hypersexual-
ity, as well as punding and compulsive medication use
(Voon et al. 2009), which represent failures of inhibition. In
the cognitive domain, impaired set-shifting (Owen et al.
1991; Hayes et al. 1998; Cools et al. 2001, 2003) and
reversal learning (Cools et al. 2002) have been well docu-
mented in PD, both of which require suppressing responses
from one set of criteria guiding behaviour to respond to
another set of criteria. Therefore, some of the motor,
behavioural and cognitive deficits in PD may reflect the
impairment of inhibitory processes revealed in this study.
The study was undertaken with patients in the ‘on’
medication state because the baseline severity of PD pre-
vented adequate performance of the tests in the ‘off’
medication state. Neuropharmacological studies in experi-
mental animals, in healthy participants or in patients with
ADHD suggest that noradrenaline (NA) is a key neuro-
transmitter with an impact on inhibitory control on the stop
signal task (Chamberlain et al. 2006, 2007, 2009). In
contrast, the effect of dopamine on SSRT remains unclear
(for review see Eagle and Baunez 2010). Definitive
assessment of any dissociable effects of levodopa medi-
cation on Go RTs and SSRT in PD awaits assessment in
future on/off medication studies following overnight
withdrawal of medication or through comparison of
untreated (‘de novo’) and chronically medicated PD
patients. Our preliminary results from a study comparing
performance of PD patients on versus off medication on the
conditional stop signal RT task suggest that levodopa
medication does not significantly influence inhibition or
conflict induced slowing as, respectively, measured by the
SSRT and the CIS (Obeso et al., submitted).
Conclusions
We show here, for the first time, that PD patients are
impaired on a conditional version of the stop signal RT
task, with the initiation of motor responses both in situa-
tions with (‘non-critical’ StopRespond RTs and CIS) or
without conflict (‘critical’ Go RTs) and inhibition of a
prepared motor response (SSRT) all being significantly
delayed relative to age-matched controls. PD patients also
had significantly greater difficulty in suppressing prepotent
or habitual responses on the Stroop, Hayling sentence
completion and RNG tasks relative to controls. These
results demonstrate the existence of a generalized inhibi-
tory deficit in PD patients assessed on medication, which
suggests that PD is a disorder of inhibition as well as
activation and that in situations of conflict, executive
control over responses is compromised in PD.
Acknowledgments We are grateful to all the participants. This
work was supported by a PhD studentship from Fundacion Caja
Madrid (IO), a Career Development Fellowship from the Parkinson’s
disease Society (LW) and a Royal Society Travelling Fellowship.
References
Alexander GE, DeLong MR, Strick PL (1986) Parallel organization of
functionally segregated circuits linking basal ganglia and cortex.
Annu Rev Neurosci 9:357–381
Aron AR, Behrens TE, Smith S, Frank MJ, Poldrack RA (2007)
Triangulating a cognitive control network using diffusion-
weighted magnetic resonance imaging (MRI) and functional
MRI. J Neurosci 27(14):3743–3752
Baglio F, Blasi V, Falini A, et al. (2009) Functional brain changes in
early Parkinson’s disease during motor response and motor
inhibition. Neurobiol Aging. doi:10.1016/j.neurobiolaging.2008.
12.009
Beck AT, Ward CH, Mendelson M, Mock J, Erbaugh J (1961) An
inventory for measuring depression. Arch Gen Psychiatry
4:561–567
Beste C, Willemssen R, Saft C, Falkenstein M (2009) Response
inhibition subprocesses and dopaminergic pathways: basal
ganglia disease effects. Neuropsychologia. doi:10.1016/j.
neuropsychologia.2009.09.023
Bokura H, Yamaguchi S, Kobayashi S (2005) Event-related potentials
for response inhibition in Parkinson’s disease. Neuropsychologia
43:967–975. doi:10.1016/j.neuropsychologia.2004.08.010
Bouquet CA, Bonnaud V, Gil R (2003) Investigation of supervisory
attentional system functions in patients with Parkinson’s disease
using the Hayling task. J Clin Exp Neuropsychol 25:751–760
Brown RG, Marsden CD (1988) Internal versus external cues and the
control of attention in Parkinson’s disease. Brain 111(Pt 2):
323–345
Brown RG, Marsden CD (1991) Dual task performance and
processing resources in normal subjects and patients with
Parkinson’s disease. Brain 114(Pt 1A):215–231
Brown RG, Soliveri P, Jahanshahi M (1998) Executive processes in
Parkinson’s disease-random number generation and response
suppression. Neuropsychologia 36:1355–1362
Burgess PW, Shallice T (1997) The Hayling and Brixton tests.
Thames Valley Test Company Limited, Bury St Edmunds
Cagigas XE, Filoteo JV, Stricker JL, Rilling LM, Friedrich FJ (2007)
Flanker compatibility effects in patients with Parkinson’s
disease: impact of target onset delay and trial-by-trial stimulus
variation. Brain Cogn 63:247–259. doi:10.1016/j.bandc.2006.
09.002
382 Exp Brain Res (2011) 212:371–384
123

Chamberlain SR, Muller U, Blackwell AD, Clark L, Robbins TW,
Sahakian BJ (2006) Neurochemical modulation of response
inhibition and probabilistic learning in humans. Science
311:861–863. doi:10.1126/science.1121218
Chamberlain SR, Del Campo N, Dowson J, Muller U, Clark L,
Robbins TW, Sahakian BJ (2007) Atomoxetine improved
response inhibition in adults with attention deficit/hyperactivity
disorder. Biol Psychiatry 62:977–984. doi:10.1016/j.biopsych.
2007.03.003
Chamberlain SR, Hampshire A, Muller U et al (2009) Atomoxetine
modulates right inferior frontal activation during inhibitory
control: a pharmacological functional magnetic resonance
imaging study. Biol Psychiatry 65:550–555. doi:
10.1016/j.biopsych.2008.10.014
Chan F, Armstrong IT, Pari G, Riopelle RJ, Munoz DP (2005) Deficits in
saccadic eye-movement control in Parkinson’s disease. Neuro-
psychologia 43:784–796. doi:10.1016/j.neuropsychologia.
2004.06.026
Chevalier G, Deniau JM (1990) Disinhibition as a basic process in the
expression of striatal functions. Trends Neurosci 13:277–280
Cools R, Barker RA, Sahakian BJ, Robbins TW (2001) Enhanced or
impaired cognitive function in Parkinson’s disease as a function
of dopaminergic medication and task demands. Cereb Cortex
11:1136–1143
Cools R, Clark L, Owen AM, Robbins TW (2002) Defining the neural
mechanisms of probabilistic reversal learning using event-related
functional magnetic resonance imaging. J Neurosci
22:4563–4567
Cools R, Barker RA, Sahakian BJ, Robbins TW (2003) L-Dopa
medication remediates cognitive inflexibility, but increases
impulsivity in patients with Parkinson’s disease. Neuropsycho-
logia 41:1431–1441
Cooper JA, Sagar HJ, Tidswell P, Jordan N (1994) Slowed central
processing in simple and go/no-go reaction time tasks in
Parkinson’s disease. Brain 117(Pt 3):517–529
Delis DC, Kaplan E, Kramer JH (2001) Delis–Kaplan executive
function scale. The Psychological Corporation, San Antonio
DeLong MR (1990) Primate models of movement disorders of basal
ganglia origin. Trends Neurosci 13:281–285
Dirnberger G, Frith CD, Jahanshahi M (2005) Executive dysfunction
in Parkinson’s disease is associated with altered pallidal-frontal
processing. Neuroimage 25:588–599. doi:10.1016/j.neuroimage.
2004.11.023
Eagle DM, Baunez C (2010) Is there an inhibitory-response-control
system in the rat? Evidence from anatomical and pharmacolog-
ical studies of behavioral inhibition. Neurosci Biobehav Rev
34:50–72
Falkenstein M, Hielscher H, Dziobek I, Schwarzenau P, Hoormann J,
Sunderman B, Hohnsbein J (2001) Action monitoring, error
detection, and the basal ganglia: an ERP study. Neuroreport
12:157–161
Falkenstein M, Willemssen R, Hohnsbein J, Hielscher H (2006)
Effects of stimulus–response compatibility in Parkinson’s dis-
ease: a psychophysiological analysis. J Neural Transm 113:1449–
1462. doi:10.1007/s00702-005-0430-1
Folstein MF, Folstein SE, McHugh PR (1975) ‘‘Mini-mental state’’. A
practical method for grading the cognitive state of patients for
the clinician. J Psychiatr Res 12:189–198
Frank MJ, Samanta J, Moustafa AA, Sherman SJ (2007) Hold your
horses: impulsivity, deep brain stimulation, and medication in
Parkinsonism. Science 318:1309–1312. doi:10.1126/science.
1146157
Gauggel S, Rieger M, Feghoff TA (2004) Inhibition of ongoing
responses in patients with Parkinson’s disease. J Neurol Neuro-
surg Psychiatry 75:539–544
Harnishfeger KK (1995) The development of cognitive inhibition:
Theories, definitions, and research evidence. In: Dempster FN,
Brainerd CJ (eds) Interference and inhibition in cognition.
Academic Press, San Diego
Hayes AE, Davidson MC, Keele SW, Rafal RD (1998) Toward a
functional analysis of the basal ganglia. J Cogn Neurosci
10:178–198
Hershey T, Revilla FJ, Wernle A, Gibson PS, Dowling JL, Perlmutter
JS (2004) Stimulation of STN impairs aspects of cognitive
control in PD. Neurology 62:1110–1114
Hughes AJ, Daniel SE, Kilford L, Lees AJ (1992) Accuracy of
clinical diagnosis of idiopathic Parkinson’s disease: a clinico-
pathological study of 100 cases. J Neurol Neurosurg Psychiatry
55:181–184
Isoda M, Hikosaka O (2007) Switching from automatic to controlled
action by monkey medial frontal cortex. Nat Neurosci
10:240–248. doi:10.1038/nn1830
Isoda M, Hikosaka O (2008) Role for subthalamic nucleus neurons in
switching from automatic to controlled eye movement. J Neuro-
sci 28:7209–7218. doi:10.1523/JNEUROSCI.0487-08.2008
Jahanshahi M, Jenkins IH, Brown RG, Marsden CD, Passingham RE,
Brooks DJ (1995) Self-initiated versus externally triggered move-
ments. I. An investigation using measurement of regional cerebral
blood flow with PET and movement-related potentials in normal
and Parkinson’s disease subjects. Brain 118(Pt 4):913–933
Jahanshahi M, Profice P, Brown RG, Ridding MC, Dirnberger G,
Rothwell JC (1998) The effects of transcranial magnetic
stimulation over the dorsolateral prefrontal cortex on suppression
of habitual counting during random number generation. Brain
121:1533–1544
Jahanshahi M, Ardouin CM, Brown RG et al (2000) The impact of
deep brain stimulation on executive function in Parkinson’s
disease. Brain 123(Pt 6):1142–1154
Kuhn AA, Tsui A, Aziz T et al (2009) Pathological synchronisation in
the subthalamic nucleus of patients with Parkinson’s disease
relates to both bradykinesia and rigidity. Exp Neurol
215:380–387. doi:10.1016/j.expneurol.2008.11.008
Lee SS, Wild K, Hollnagel C, Grafman J (1999) Selective visual
attention in patients with frontal lobe lesions or Parkinson’s
disease. Neuropsychologia 37:595–604
Logan GD, Cowan WB (1984) On the ability to inhibit thought and
action: a theory of an act of control. Psychol Rev 91:295–327
Middleton FA, Strick PL (2000) Basal ganglia output and cognition:
evidence from anatomical, behavioral, and clinical studies. Brain
Cogn 42:183–200
Nigg JT (2000) On inhibition/disinhibition in developmental psycho-
pathology: views from cognitive and personality psychology and
a working inhibition taxonomy. Psychol Bull 126:220–246
Owen AM, Roberts AC, Polkey CE, Sahakian BJ, Robbins TW
(1991) Extra-dimensional versus intra-dimensional set shifting
performance following frontal lobe excisions, temporal lobe
excisions or amygdalo-hippocampectomy in man. Neuropsych-
ologia 29:993–1006
Playford ED, Jenkins IH, Passingham RE, Nutt J, Frackowiak RS,
Brooks DJ (1992) Impaired mesial frontal and putamen activa-
tion in Parkinson’s disease: a positron emission tomography
study. Ann Neurol 32:151–161. doi:10.1002/ana.410320206
Praamstra P, Plat FM (2001) Failed suppression of direct visuomotor
activation in Parkinson’s disease. J Cogn Neurosci 13:31–43
Praamstra P, Stegeman DF, Cools AR, Horstink MW (1998) Reliance
on external cues for movement initiation in Parkinson’s disease.
Evidence from movement-related potentials. Brain 121(Pt
1):167–177
Ray NJ, Jenkinson N, Brittain J et al (2009) The role of the
subthalamic nucleus in response inhibition: evidence from deep
Exp Brain Res (2011) 212:371–384 383
123

brain stimulation for Parkinson’s disease. Neuropsychologia
47:2828–2834. doi:10.1016/j.neuropsychologia.2009.06.011
Redgrave P, Prescott TJ, Gurney K (1999) The basal ganglia: a
vertebrate solution to the selection problem? Neuroscience
89:1009–1023
Rieger M, Gauggel S (1999) Inhibitory after-effects in the stop-signal
paradigm. Br J Psychol 90:509–518
Rieger M, Gauggel S, Burmeister K (2003) Inhibition of ongoing
responses following frontal, nonfrontal, and basal ganglia
lesions. Neuropsychology 17:272–282
Rivaud-Pechoux S, Vidailhet M, Brandel JP, Gaymard B (2007)
Mixing pro- and antisaccades in patients with parkinsonian
syndromes. Brain 130:256–264. doi:10.1093/brain/awl315
Schroeder U, Kuehler A, Haslinger B et al (2002) Subthalamic
nucleus stimulation affects striato-anterior cingulate cortex
circuit in a response conflict task: a PET study. Brain
125:1995–2004
Seiss E, Praamstra P (2004) The basal ganglia and inhibitory
mechanisms in response selection: evidence from subliminal
priming of motor responses in Parkinson’s disease. Brain
127:330–339. doi:10.1093/brain/awh043
Seiss E, Praamstra P (2006) Time-course of masked response priming
and inhibition in Parkinson’s disease. Neuropsychologia
44:869–875
Spatt J, Goldenberg G (1993) Components of random generation by
normal subjects and patients with dysexecutive syndrome. Brain
Cogn 23:231–242
Stroop JR (1935) Studies of interference in serial verbal reactions.
J Exp Psychol 18:643–662
Thobois S, Hotton GR, Pinto S, Wilkinson L, Limousin-Dowsey P,
Brooks DJ, Jahanshahi M (2007) STN stimulation alters pallidal-
frontal coupling during response selection under competition.
J Cereb Blood Flow Metab 27:1173–1184
van den Wildenberg WP, van Boxtel GJ, van der Molen MW, Bosch
DA, Speelman JD, Brunia CH (2006) Stimulation of the
subthalamic region facilitates the selection and inhibition of
motor responses in Parkinson’s disease. J Cogn Neurosci
18:626–636
Verbruggen F, Logan GD (2009a) Automaticity of cognitive control:
goal priming in response-inhibition paradigms. J Exp Psychol
Learn Mem Cogn 35:1381–1388. doi:10.1037/a0016645
Verbruggen F, Logan GD (2009b) Proactive adjustments of response
strategies in the stop-signal paradigm. J Exp Psychol Hum
Percept Perform 35:835–854. doi:10.1037/a0012726
Vila M, Levy R, Herrero MT et al (1997) Consequences of
nigrostriatal denervation on the functioning of the basal ganglia
in human and nonhuman primates: an in situ hybridization study
of cytochrome oxidase subunit I mRNA. J Neurosci 17:765–773
Voon V, Fernagut PO, Wickens J et al (2009) Chronic dopaminergic
stimulation in Parkinson’s disease: from dyskinesias to impulse
control disorders. Lancet Neurol 8:1140–1149. doi:10.1016/
S1474-4422(09)70287-X
Williams-Gray CH, Foltynie T, Brayne CE, Robbins TW, Barker RA
(2007) Evolution of cognitive dysfunction in an incident
Parkinson’s disease cohort. Brain 130:1787–1798. doi:
10.1093/brain/awm111
Witt K, Pulkowski U, Herzog J, Lorenz D, Hamel W, Deuschl G,
Krack P (2004) Deep brain stimulation of the subthalamic
nucleus improves cognitive flexibility but impairs response
inhibition in Parkinson disease. Arch Neurol 61:697–700. doi:
10.1001/archneur.61.5.697
Wylie SA, Stout JC, Bashore TR (2005) Activation of conflicting
responses in Parkinson’s disease: evidence for degrading and
facilitating effects on response time. Neuropsychologia
43:1033–1043
Wylie SA, van den Wildenberg WP, Ridderinkhof KR, Bashore TR,
Powell VD, Manning CA, Wooten GF (2009a) The effect of
Parkinson’s disease on interference control during action selec-
tion. Neuropsychologia 47:145–157. doi:10.1016/j.neuropsy
chologia.2008.08.001
Wylie SA, van den Wildenberg WP, Ridderinkhof KR, Bashore TR,
Powell VD, Manning CA, Wooten GF (2009b) The effect of speed-
accuracy strategy on response interference control in Parkinson’s
disease. Neuropsychologia 47:1844–1853. doi:10.1016/j.
neuropsychologia.2009.02.025
384 Exp Brain Res (2011) 212:371–384
123




















