
Cytotoxic activity of Styela plicata hemocytes against mammalian cell targets: II. Properties of the...
8
Cytotoxic activity of Styela plicata hemocytes against mammalian cell targets: I. Properties of the in vitro reaction against erythrocytes Loredana LIPARI, Matteo CAMMARATA, Vincenzo ARIZZA and Daniela PARRINELLO Laboratory oj Marine Immunobiology. Institute oj Zoology. University oj Palermo. Palermo. Italy ABSTRACT - Hemocytes of the ascidian Styela plicata showed a natural cytotoXiC capacity (HCA) when assayed in vitro against rabbit erythrocytes, whereas sheep erythrocytes were not sensitive to the cytotoxic cells. As expected for an osmo-conforming marine invertebrate, the effector cells functioned best in a medium closest to that of the aquarium sea water. HCA is a calcium dependent reaction, occurs rapidly (15-30 min), at 20-37 °C, at pH 8.0, in a medium isosmotic (1090 mOsm kg-I) for ascidian hemocytes, at a low effector:target ratios. Assays were carried out using appropriate controls of hemocyte mortality and erythrocyte spontaneous lysis. Individual hemolytic actiVity varied depending on the Styela donor and hemocyte numbers. Although soluble lytic factor(s) were not significantly present in the reac- tion medium, the low optimal effector:target cells ratios suggest a mechanism by which one effector hemocyte acts on several erythrocytes. Inhibition experiments with lipid components of erythrocyte membranes showed that sphingomyelin molecules may be involved in binding cytolysins. KEY WORDS: Invertebrate; Tunicate; Styela plicata; Hemocytes: Cytotoxicity; Rabbit eryth- rocytes INTRODUCTION annelids, echinoderms and molluscs have been compared to vertebrate natural killer cells fOr Cell killing by immunocytes represents a natural their anti - tumour activity (Decker et al., 1981), defence barrier against proliferation of transfor- the presence of specific surface markerS (France- med cells, virus-infeCted cells, parasites, and schi et al., 1991) Or pore-forming cytolytic other foreign invaders. In invertebrates, sponta- mechanisms (Porchet-Hennere et al., 1992; neous cytotoxic actiVity against allogeneic and Tyson and Jenkin, 1974; Luquet and Le Clerc, xenogeneic cells, including mammalian tumour 1983; S6derhall et al., 1985). Tunicates are fil- cells, has been demonstrated by in. vitro assays ter-feeding marine invertebrates considered to be (Parrinello and Arizza, 1992). The cell-killing the most primitive members of Chordata which mechanism involves soluble cytolysins or it exhibit features characteristic of the vertebrates reqUires contacts between effector and target (Berril, 1955). Recent studies of the molecular cells (reviewed in: Canicatti, 1990; Paninello and phylogeny of the animal kingdom by comparison Arizza, 1992; Lotzova, 1993). Cytotoxic cells from of sequences of 5S rRNA (Hori and Osawa, 1987) Results of this research have been presented at the meeting «Modulators of Immune Responses. The evolutionary traih spon- sored by USA BRDL, Breckenridge, Colorado. July 8-13. 1995.
Transcript of Cytotoxic activity of Styela plicata hemocytes against mammalian cell targets: II. Properties of the...

Cytotoxic activity of Styela plicata hemocytes against mammalian cell targets: I. Properties of the in vitro
reaction against erythrocytes
Loredana LIPARI, Matteo CAMMARATA, Vincenzo ARIZZA and Daniela PARRINELLO Laboratory ojMarine Immunobiology. Institute ojZoology. University ojPalermo. Palermo. Italy
ABSTRACT - Hemocytes of the ascidian Styela plicata showed a natural cytotoXiC capacity (HCA) when assayed in vitro against rabbit erythrocytes, whereas sheep erythrocytes were not sensitive to the cytotoxic cells. As expected for an osmo-conforming marine invertebrate, the effector cells functioned best in a medium closest to that of the aquarium sea water. HCA is a calcium dependent reaction, occurs rapidly (15-30 min), at 20-37 °C, at pH 8.0, in a
medium isosmotic (1090 mOsm kg-I) for ascidian hemocytes, at a low effector:target ratios. Assays were carried out using appropriate controls of hemocyte mortality and erythrocyte spontaneous lysis. Individual hemolytic actiVity varied depending on the Styela donor and hemocyte numbers. Although soluble lytic factor(s) were not significantly present in the reaction medium, the low optimal effector:target cells ratios suggest a mechanism by which one effector hemocyte acts on several erythrocytes. Inhibition experiments with lipid components of erythrocyte membranes showed that sphingomyelin molecules may be involved in binding
cytolysins.
KEY WORDS: Invertebrate; Tunicate; Styela plicata; Hemocytes: Cytotoxicity; Rabbit eryth
rocytes
INTRODUCTION annelids, echinoderms and molluscs have been compared to vertebrate natural killer cells fOr
Cell killing by immunocytes represents a natural their anti -tumour activity (Decker et al., 1981),
defence barrier against proliferation of transfor the presence of specific surface markerS (Francemed cells, virus-infeCted cells, parasites, and schi et al., 1991) Or pore-forming cytolyticother foreign invaders. In invertebrates, sponta mechanisms (Porchet-Hennere et al., 1992; neous cytotoxic actiVity against allogeneic and Tyson and Jenkin, 1974; Luquet and Le Clerc, xenogeneic cells, including mammalian tumour 1983; S6derhall et al., 1985). Tunicates are filcells, has been demonstrated by in. vitro assays ter-feeding marine invertebrates considered to be (Parrinello and Arizza, 1992). The cell-killing the most primitive members of Chordata which mechanism involves soluble cytolysins or it exhibit features characteristic of the vertebrates reqUires contacts between effector and target (Berril, 1955). Recent studies of the molecular cells (reviewed in: Canicatti, 1990; Paninello and phylogeny of the animal kingdom by comparison Arizza, 1992; Lotzova, 1993). Cytotoxic cells from of sequences of 5S rRNA (Hori and Osawa, 1987)
Results of this research have been presented at the meeting «Modulators of Immune Responses. The evolutionary traih sponsored by USA BRDL, Breckenridge, Colorado. July 8-13. 1995.
io
Text Box
15

132 Lipari L.. Cammarata M.. Arizza V. and Parrinello D.
and 18S rRNA (Field et ai.. 1988; Wada and Satoh. 1994), as well as studies of the structure of genes for muscle actin (Kusakabe et al.. 1992) support this view. In tunicates. hemocytes display cytotoxic activity in vitro when examined in xenogeneic or allogeneic combinations (Fuke. 1980; Kelly et ai.. 1992), are also involved in non-fusion reactions between allogeneic colonies of BotryLLus (Saito and Watanabe. 1982), in tunic allograft rejections (Raftos et al.. 1987a. 1987b), and tunic disruptive inflammatory reaction (Parrinello et al.. 1977. 1984). In the solitary ascidian Ciona intestinaLis. in vitro spontaneous cytotoxic activity against mammalian target cells has been reported. Erythrocytes from rabbit, human. guinea pig and sheep (Parri nello and Arizza. 1992; Parrinello et ai.. 1993; Parrinello. 1995), mouse and human tumour cell lines (Peddie and Smith. 1993; 1994) were the targets. Differences for effector: target cell ratios so far reported. show that anti-erythrocytes (Parri nello et ai.. 1995) and anti-tumour (Peddie and Smith. 1993) HCAs present distinguishing characteristics in their mechanisms. In addition. Parrinello et al.. (1995) showed that sphingomyelin (choline-containing lipid), which is primarily contained in the outer layer of sheep erythrocytes (see reviews: Shiga and Maeda. 1980; Chapman. 1983), inhibits the cytotoxic mechanism of C. intestinaLis hemocytes against this type of target. In the present paper we show that hemocytes from the solitary ascidian StyeLa pLicata express cytotoxic activity. Some properties of the in vitro reaction against erythrocytes are reported.
MATERIALS AND METHODS
Bleeding and hemocyte preparation and quantification The ascidians (4-10 cm in length) were collected from Palermo harbour. kept in filtered. aerated sea water at 15 ce. and fed with a marine invertebrate diet (Hawaiian Marine Imports. Inc.. Houston. Texas) until used. Before bleeding the tunic was cleaned of epiphytes. and the sample area sterilized with alcohol. The hemolymph. collected by inserting a needle throughout the tunic into the body cavity. was mixed (v/v) FSW-EDTA (artificial sea water without CaClz and MgClz but enriched with 10 mM EDTA, pH 7.2) to prevent clotting. Finally the cells were suspended in marine solution (MS) (12 mM CaClz .6HzO: 11 mM KCI: 26 mM MgClz.6HzO: 43 mM Tris: 38 mM HCl: 0.4 M NaCI. pH 8) which is isosmotic with hemocytes (1090 mOsm Kg-I). Assays in Tris buffered saline TBSCa (50 mM Tris-HCl. 150 mM NaCI. 10 mM CaCIz.6HzO. pH 8. 450 mOsm Kg-I) which is optimal for with erythrocytes were also performed. Cell counts
were determined using an improved Neubauer hemocytometer. Hemocyte suspenSions containing 5 x 106 cells per mI were prepared.
Chemicals
Unless otherwise reported. all chemicals were purchased from Sigma Chemicals Co. St. Luis
Preparation ofErythrocyte Targets
Rabbit erythrocytes (RE) were from Sclavo (Siena. Italy), sheep erythrocytes (SE) were proVided by the Zooprophylaxis Institute (Palermo. Italy). The erythrocytes were washed three times in phosphate buffered saline (PBS), and. after centrifugation for 10 min at 400 g. resuspended in 10 ml MS at the required concentration. In spite of the high osmolality of the medium. significant cell lysis was not found when the erythrocytes were suspended in MS.
E"-']JenmentaL cytotoxic assay qf the hemocytes (HCA).
The cytotoxic assay has been previously described (Parrinello et aL. 1993: 1995). In brief. a 200 pi of hemocyte suspenSion (1 x 106 cells) in MS or TBS-Ca was mixed with an equal volume of freshly prepared SE or RE suspenSion (8 x 106 cells) in the same medium. Control erythrocyte suspenSions showed that the spontaneous hemoglobin release never exceeded 5% of total release. Hemocyte counts were determined in the final volume of the reaction mixture. The mixture was incubated with continuous and moderate shaking at 20°C for 30 min. At the end of the incubation. the supernatant was separated and the amount of the released hemoglobin (Hb) was estimated by reading the absorbance at 541 nm. Experimental controls and hemolysis degree was determined according to Parrinello et aL. (1995). HCA from each of 60 S. pLicata specimens was evaluated. For each experiment three samples were prepared. the values representing the means ± S.D.
Inhibition experiment
All chemicals (Sigma) tested for inhibitory actiVity. sphingomyelin (from bovine brain), phosphatidylcholine. phosphatidylserine. cholesterol. and phosphatidylethanolamine were dissolved in MS. Phosphatidylcholine. phosphatidylserine. sphingomyelin phosphatidylethanolamine and cholesterol were relatively insoluble. For these compounds. stock solutions were briefly sonicated (Vibra-Cell. Sonics & Materials Inc.. Dambury. CT). Hemocyte suspensions were prepared in the medium containing the compounds at the various concentrations. After 20 min preincubation. the erythrocyte suspenSion (prepared in reaction medium containing the same amount of inhibitor), was added at various E:T ratios for cytotoxic assay. Lysis due to chemicals alone was measured as control.
133 Cytotoxicity by Styela plicata hemocytes: I.
Preparation of lysate from hemocytes
Supernatants from homogenated or sonicated hemocytes were used. Briefly, hemocytes were separated from hemolymph -FSW- EDTA, immediately suspended in ice cold FSW-EDTA. After centrifugation, 3 x 106 mP hemocytes were resuspended in MS and sonicated at 4 DC for 20 sec (Branson, model B15, Danbury, CT, U.S.A) or homogenated for 3 min at 4 DC. The lysate was then centrifuged at 27,000 g for 20 min at 4 DC and cytotoxic activity of the supernatant was assayed.
Statistical analyses
Functional relationship between two variables were expressed in terms of linear regression equation. Statistical significance was determined by reference to the t-distribution, using the standard error of the correlation coefficient (Sokal and Rholf, 1981). The statistical significance of differences between experimental groups was determined by Student's two tailed t-test.
To perform statistical analyses, the Statistica for Windows (Statsoft Inc.) package was used.
RESULTS
Properties of the in vitro reaction against rabbit erythrocytes
Eosin Y exclusion test showed that MS is a suitable medium for Styela hemocyte suspenSion. The hemocyte mortality after 1 hour at 20°C in MS ranged from 0.2% to 5%. Table I presents hemocyte cytotoxiC activity assayed in MS. The hemocytes, in absence of Caz+
ions only presented a residual level of lysis versus RE which were the most sensitive target. A 12 mM CaClz appeared to be the lower salt concentration which produced a degree of hemolysis ranging from 35% t073 %, whereas 100 mM CaClz did not
Table I Hemocyte cytotoxic activity (HCA) obtained from reaction mixture in which Styela plicata hemocytes (5 x 106 ml- l )
had reacted against rabbit erithrocytes (45x106 mJ-l) in MS with or without Ca2+.
- - - - MS - - - - - Ms - - - - - - - - MS
~p~~i!Jlt:Il __ ~13Il1rvt <::~2+! J\Vitl1()1lt.c;~2~1 J~() mM EDTA)
1 35 12 2 28 11 3 64 26 4 33 9 5 62 2 6 77 1
Hemolysys degree (%)
100
80
;? ~ Q) 60 ~ Cl CII "C (/)
'iii>- 40 '0 E CII I
20
O...L..L.-----'- RE SE
MS
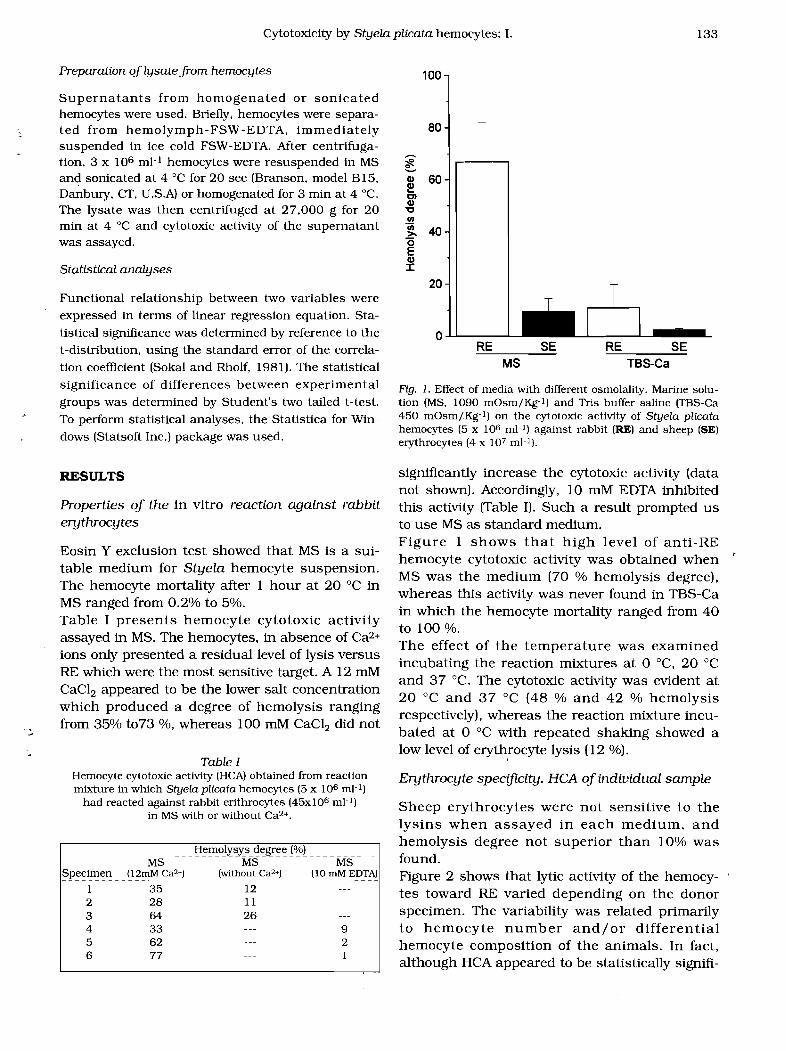
Fig. 1. Effect of media with different osmolality. Marine solution (MS, 1090 mOsm/Kg-l) and Tris buffer saline (TBS-Ca 450 mOsm/Kg-l) on the cytotoxic activity of Styela plicata hemocytes (5 x 106 mJ-l) against rabbit (RE) and sheep (SE) erythrocytes (4 x 107 mJ-l).
significantly increase the cytotoxic activity (data not shown). Accordingly, 10 mM EDTA inhibited this activity (Table I). Such a result prompted us to use MS as standard medium. Figure 1 shows that high level of anti-RE hemocyte cytotoxic activity was obtained when MS was the medium (70 % hemolysis degree), whereas this activity was never found in TBS-Ca in which the hemocyte mortality ranged from 40 to 100 %. The effect of the temperature was examined incubating the reaction mixtures at 0 DC, 20°C and 37 DC. The cytotoxic activity was evident at 20°C and 37 °C (48 % and 42 % hemolysis respectively), whereas the reaction mixture incubated at 0 °C with repeated shaking showed a low level of erythrocyte lysis (12 %).
Erythrocyte specificity. HCA of individual sample
Sheep erythrocytes were not sensitive to the lysins when assayed in each medium, and hemolysis degree not superior than 10% was found. Figure 2 shows that lytic activity of the hemocytes toward RE varied depending on the donor specimen. The variability was related primarily to hemocyte number and/or differential hemocyte composition of the animals. In fact, although HCA appeared to be statistically signifi-
RE SE TBS-Ca
134
100
Lipari L.. Cammarata M .. Arizza V. and Parrinello D.
0 0 0 DO 0
80 0
0 0 0 0
....... ~ 0 0
0
Qlo 0 0
0 ~ Q) ~. 60
DO
0 0 0
0 0 0
Cl Q)
"C 0 0
0
0 0 0
0
0 0 0
0 0
.!!.! III >. "6 40
0 0
0
0 0 0
0
0
E Q) ::c 0
0 0
0 0 0
0
0
0
20 0
0
0
0 0
0 0
246 8 10
Cell number (1 06/ml)
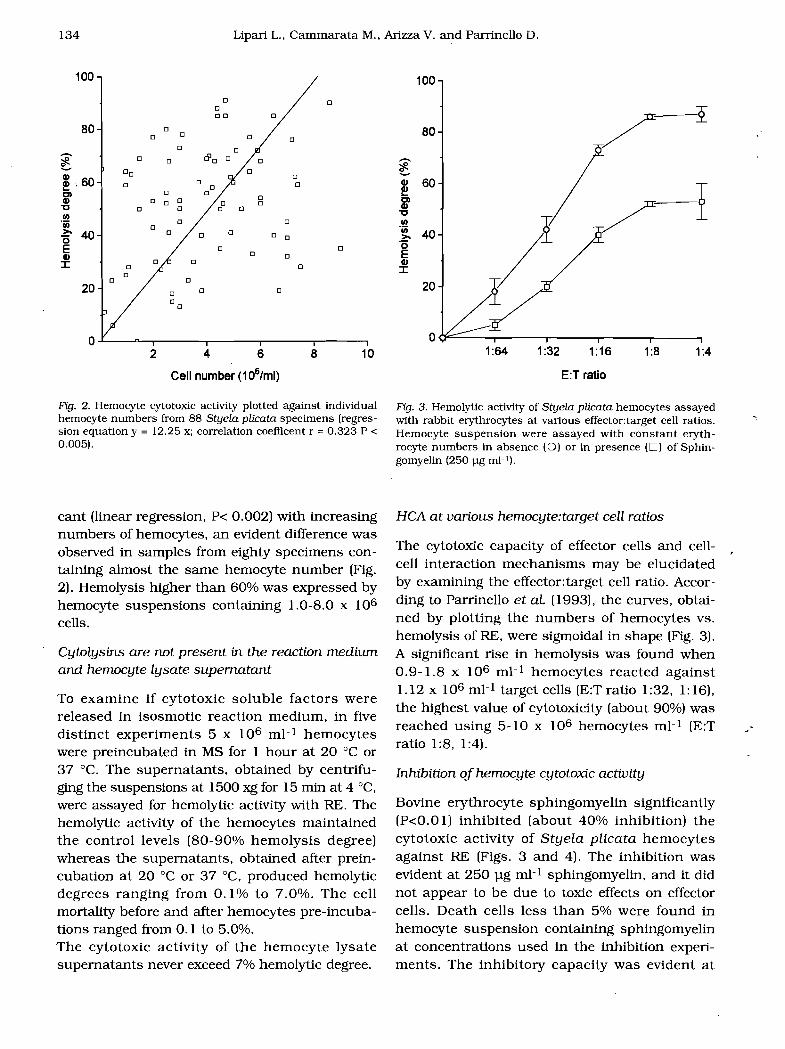
Fig. 2. Hemocyte cytotoxic activity plotted against individual hemocyte numbers from 88 Styela plicata specimens (regression equation y = 12.25 x; correlation coefficent r = 0.323 P < 0.005).
cant (linear regression. P< 0.002) with increasing numbers of hemocytes, an evident difference was observed in samples from eighty specimens containing almost the same hemocyte number (Fig. 2). Hemolysis higher than 60% was expressed by hemocyte suspensions containing 1.0-8.0 x 106
cells.
Cytolysins are not present in the reaction medium and hemocyte lysate supernatant
To examine if cytotoxic soluble factors were released in isosmotic reaction medium. in five distinct experiments 5 x 106 ml- 1 hemocytes were preincubated in MS for 1 hour at 20 cC or 37 cC. The supernatants, obtained by centrifuging the suspensions at 1500 xg for 15 min at 4 cC.
were assayed for hemolytic activity with RE. The hemolytic activity of the hemocytes maintained the control levels (80-90% hemolysis degree) whereas the supernatants. obtained after preincubation at 20 cC or 37 cC. produced hemolytic degrees ranging from 0.1 % to 7.0%. The cell mortality before and after hemocytes pre-incubations ranged from 0.1 to 5.0%. The cytotoxic activity of the hemocyte lysate supernatants never exceed 7% hemolytic degree.
100
80
....... ~ ~ Q) 60
I~ Cl Q)
"C III 'iii >. 40 "6 E Q)
::c 20
1:64 1:32 1:16 1:8 1:4
E:T ratio
Fig. 3. Hemolytic activity of Styela plicata hemocytes assayed with rabbit erythrocytes at various effector:target cell ratios. -. Hemocyte suspension were assayed with constant erythrocyte numbers in absence (0) or in presence (0) of Sphingomyelin (250 pg m)-l).
HCA at various hemocyte:target cell ratios
The cytotoxic capacity of effector cells and cellcell interaction mechanisms may be elucidated by examining the effector:target cell ratio. According to Parrinello et al. (1993), the curves, obtained by plotting the numbers of hemocytes vs. hemolysis of RE. were sigmoidal in shape (Fig. 3). A significant rise in hemolysis was found when 0.9-1.8 x 106 mP hemocytes reacted against 1.12 x 106 mI-l target cells (E:T ratio 1:32, 1: 16). the highest value of cytotoxicity (about 90%) was reached using 5-10 x 106 hemocytes mI-l (E:T ratio 1:8. 1:4).
Inhibition ofhemocyte cytotoxic activity
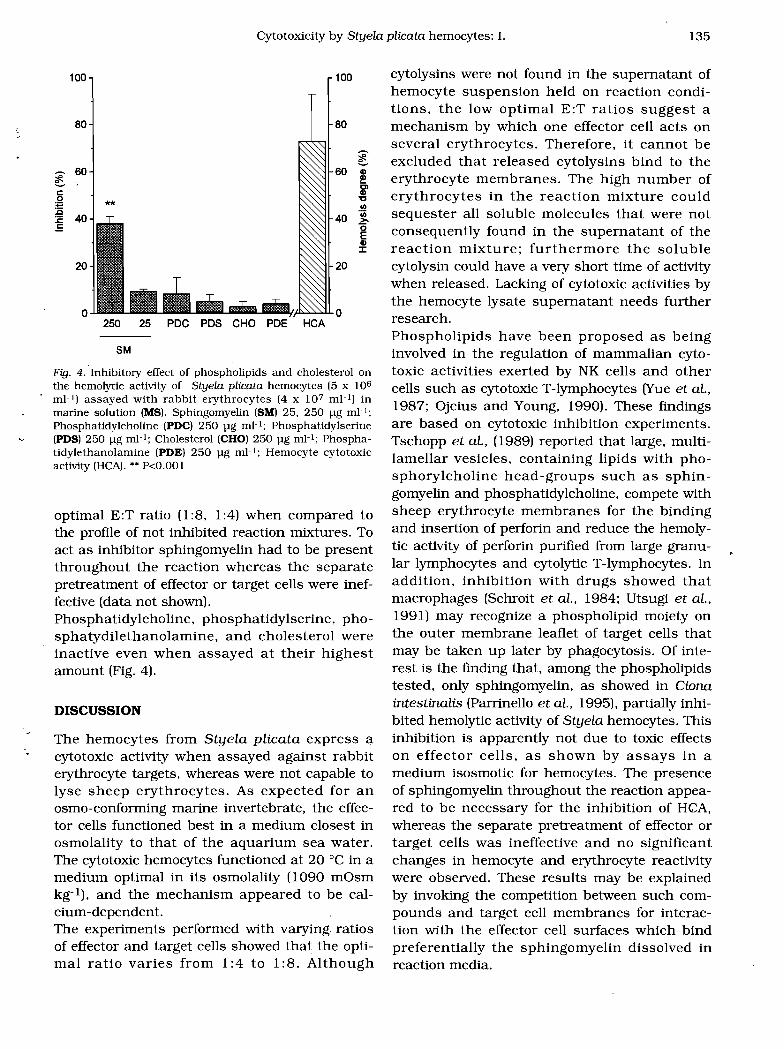
Bovine erythrocyte sphingomyelin significantly (P<O.O 1) inhibited (about 40% inhibition) the cytotoxic activity of Styela plicata hemocytes against RE (Figs. 3 and 4). The inhibition was evident at 250 pg mI-l sphingomyelin. and it did not appear to be due to toxic effects on effector cells. Death cells less than 5% were found in hemocyte suspension containing sphingomyelin at concentrations used in the inhibition experiments. The inhibitory capacity was evident at
135 Cytotoxicity by Styela plicata hemocytes: I.
100 100
80 80
~ 0
60 60 Gl
~ ~ Cl
c: Gl
:g0 '0 III
:E 40 40 .~
(5.= E Gl J:
20 20
o 250 25 POC POS CHO POE HCA
8M
Fig. 4. Inhibitory effect of phospholipids and cholesterol on the hemolytic activity of Styela plicata hemocytes (5 x 106
mI-I) assayed with rabbit erythrocytes (4 x 107 mI-I) in marine solution (MS). Sphingomyelin (SM) 25. 250 llg mP; Phosphatidy1choline (PDC) 250 llg mP; Phosphatidylserine (PDS) 250 llg mP; Cholesterol (CHOj 250 llg mP; PhosphatidylethanoIamine (PDE) 250 llg mI-I; Hemocyte cytotoxic activity (HCA).•• P<O.OOI
optimal E:T ratio (1:8, 1:4) when compared to the profile of not inhibited reaction mixtures. To act as inhibitor sphingomyelin had to be present throughout the reaction whereas the separate pretreatment of effector or target cells were ineffective (data not shown). Phosphatidylcholine, phosphatidylserine, phosphatydilethanolamine, and cholesterol were inactive even when assayed at their highest amount (Fig. 4).
DISCUSSION
The hemocytes from Styela plicata express a cytotoxic activity when assayed against rabbit erythrocyte targets, whereas were not capable to lyse sheep erythrocytes. As expected for an osmo-conforming marine invertebrate, the effector cells functioned best in a medium closest in osmolality to that of the aquarium sea water. The cytotoxic hemocytes functioned at 20°C in a medium optimal in its osmolality (1090 mOsm kg-I), and the mechanism appeared to be calcium-dependent. The experiments performed with varying ratios of effector and target cells showed that the optimal ratio varies from 1:4 to 1:8. Although
cytolysins were not found in the supernatant of hemocyte suspension held on reaction conditions, the low optimal E:T ratios suggest a mechanism by which one effector cell acts on several erythrocytes. Therefore, it cannot be excluded that released cytolysins bind to the erythrocyte membranes. The high number of erythrocytes in the reaction mixture could sequester all soluble molecules that were not consequently found in the supernatant of the reaction mixture; furthermore the soluble cytolysin could have a very short time of activity when released. Lacking of cytotoxic activities by the hemocyte lysate supernatant needs further research. Phospholipids have been proposed as being involved in the regulation of mammalian cytotoxic activities exerted by NK cells and other cells such as cytotoxic T-Iymphocytes (Yue et al., 1987; Ojcius and Young, 1990). These findings are based on cytotoxiC inhibition experiments. Tschopp et al., (1989) reported that large, multilamellar vesicles, containing lipids with phosphorylcholine head-groups such as sphingomyelin and phosphatidylcholine, compete with sheep erythrocyte membranes for the binding and insertion of perforin and reduce the hemolytic activity of perforin pUrified from large granular lymphocytes and cytolytic T-Iymphocytes. In addition, inhibition with drugs showed that macrophages (Schroit et al., 1984; Utsugi et al., 1991) may recognize a phospholipid moiety on the outer membrane leaflet of target cells that may be taken up later by phagocytosis. Of interest is the finding that, among the phospholipids tested, only sphingomyelin, as showed in Ciona intestinalis (Parrinello et al., 1995), partially inhibited hemolytic activity of Styela hemocytes. This inhibition is apparently not due to toxic effects on effector cells, as shown by assays in a medium isosmotic for hemocytes. The presence of sphingomyelin throughout the reaction appeared to be necessary for the inhibition of HCA, whereas the separate pretreatment of effector or target cells was ineffective and no significant changes in hemocyte and erythrocyte reactivity were observed. These results may be explained by invoking the competition between such compounds and target cell membranes for interaction with the effector cell surfaces which bind preferentially the sphingomyelin dissolved in reaction media.

136 Lipari L., Cammarata M., Arizza V. and Parrinello D.
In contrast to Ciona intestinalis (Parrinello et at, 1995), where the sphingomyelin inhibition was complete at 25 pg mI-l, we observed a maximum of inhibition of 40 % when 250 pg mI-l of sphingomyelin was used. The residual activity expressed by S. plicata hemocytes following sphingO¥1yelin inhibition, suggests an additional cytotoxic mechanism independent from phospholipid membrane composition of the erythrocytes. However, in some way membrane components of the targets may be involved because we demonstrate that sheep erythrocytes were not suscepti ble to the lysis. In this respect cytotoxicity of Styela plicata hemocytes differs in specificity from that of Ciona intestinalis which partially lysed (40% hemolysis degree) sheep erythrocytes when assayed in marine solution medium (1090 mOsm kg-I). We are not able to propose a model that explains the lytic mechanism of ascidian hemocytes against erythrocyte targets. Since the sphingomyelin inhibition was not complete at higher concentration, apparently two lytic factors as well as different hemocytes could be involved into the lysis. One of them appear to be dependent from interactions of hemocyte lysin with membrane components which could cause changes in erythrocyte membrane permeability leading to lysis. It is known that the reaction of some soluble invertebrate hemolysins, present in the cell-free coelomic fluid, may be mediated by interactions with membrane targets through lipids. In Holothuria poW (Canicatti et at, 1987) and Marthasterias glacialis (Canicatti, 1989), sphingomyelin is an inhibitor of hemolytic reactions, whereas cholesterol, phosphatidylinositol and phosphatidylethanolamine were effective in inhibiting hemolysin from Paracentrotus lividus (Canicatti, 1991). The lipid-lysin interaction, in conjunction with mammalian models, suggests that an unrestricted, relatively simple mechanism, involving target membrane lipids may also be important in the expression of tunicate natural cytotoxicity, a process that has endured in the evolutionary development of immunity.
Acknowledgements
The work was supported by research grant from
C.N.R. (1994) and M.U.R.S.T. (1994). We thank Dr.
Benedetto Savona, Mr. Giuseppe Pitucco and Mr. Sal
vatore Manzella, for their helpfull assistance.
REFERENCES
BERRIL, N.J.. 1955. TI1e origin of vertebrates. Oxford University Press, New York
CANlcATn', C., PARRINELLO, N., and ARlZZA. V., 1987. Inhibitory activity of sphingomyelin on hemolytic activity of coelomic fluid of Holothuria poW (Echinodermata). Dev. Compo Immunol.. 11: 29-35.
CANICATTI'. C., 1989. Evolution of lytic system in echinoderms. II Naturally occurring hemolytic activity in Marthasterias glacialis (Asteroidea). Compo Biochem Physlol., 9SA: 587-591.
CANlcATn'. C.. 1990. Hemolysins: Pore-forming proteins in invertebrates. Experientia, 46: 239-244.
CANICATn', C., 1991. Binding properties of Paracentrotus lividus (Echinoidea) hemolysin. Compo Biochem Physiol.. 9SA: 463-468.
CHAPMAN, D.. 1983. Biomembrane fluidity in biology. Vol II Aloia, RC.. (Ed.). New York. Academic Press, pp. 5-39.
DECKER, J.M.. ELMHOLT. A, and MUCHMORE, AV., 1981. Spontaneous cytotoxicity mediated by invertebrate mononuclear cells toward normal and malignant vertebrate targets: inhibition by defmed mono- and disaccharides. Cell. Immunol., 59: 161-170.
FIELD, KG., OLSEN, G.J .. LANE., D.J. GIOVANNONI. S.J .. GHISELIN, M.T.. RAFF, E.C., PACE N.R, and RAFF, RA, 1988. Molecular phylogeny of the animal kingdom. Sciem:e, 239: 748-753.
FRANCESCHI, C., COSSARIZZA, A, MONTI, D., and OrrAVlANI, E., 1991. Cytotoxicity and immunocyte markers in cells from the freshwater snail Planorbartus comeus L. (Gastropoda Pulmonata): implication for the evolution of natural killer cells. Eur. J. Immunol., 21: 489-493.
FuKE, M.T., 1980. "Contact reaction" between xenogeneic or allogeneic coelomic cells of solitary ascidians. BioI. Bull., , 158: 304-315.
HORI, H., and OSAWA, S., 1987. Origin and evolution of organisms as deduced from 5S ribosomal RNA sequences. Mol. BioI. Evol., 4: 445-472.
KELLY, KL., COOPER, E.L., and RAFTos, D.A., 1992. In vitro allogeneic cytotOXicity in the solitary urochordate Styela clava. J. Exp. Zool., 262: 202-208.
KUSAKABE, T.. MAKABE, KW., and SATOH, N., 1992. Tunicate muscle actin genes: Structure and organization as a gene cluster. J. Mol. BioI., 227: 955-960.
LoTzOVA', E., 1993. Immune surveillance and natural immunity. In: Developmental Immunology. Cooper, E.L. and Nisbet-Brown, E. (Eds.). New York. Oxford University Press.
LuguET, G., and LECLERC, M., 1983. Spontaneous and induced cytotoxicity of axial organ cells from Asterias rubens (Asterid-Echinoderm). Immunol. Lett., 6: 339-342.
OJCIUS, D.M., and YOUNG, J.D.E., 1990. Characterization of the inhibitory effect of lysolipids on perforin-mediated hemolysis. Mol. Immunol., 27: 257-261.
PARRINELLO, N., PATRICOLO, E., and CANICATTI', C., 1984. Inflammatory-like reactions in the tunic of Ciona intestinalis (Tunicata). I Encapsulation and tissue injury. BioI. Bull., 167: 229-273.
PARRINELLO, N., 1995. Cytotoxic actiVity of tunicate hemocytes. In: Progress in Molecular and Subcellular Biology. Invertebrate Immunology. B. Rinkevich and Muller, (Ed.]. Vol. 15, pp. 190-217.

137 Cytotoxicity by Styela plicata hemocytes: 1.
PARRINELLO, N., and ARIZZA, V., 1992. Cytotoxic activity of invertebrate hemocytes with preliminary findings on the tunicate Ciona intestinalis. Boll. Zool., 59: 183-190.
PARRINELLO, N., PATRICOLO, E., and CANICATTI, C., 1977. Tunicate immunobiology. Tunic reaction of Ciona intestinalis L. to erythrocyte injection. Boll. Zool., 44: 373-381
PARRINELLO, N., ARIZZA, V., CAMMARATA, M., and PARRINELLO D.M., 1993. Cytotoxic activity of Ciona intestinalis (Tunicata) hemocytes: properties of the in vitro reaction against erythrocyte targets. Dev. Compo Immunol., 17: 19-27.
PARRINELLO, N.. CAMMARATA, M., LIPARI, L., and ARIZZA, V., 1995. Sphingomyelin inhibition of Ciona intestinalis (Tunicata) citotoxic hemocytes assayed against sheep erytrocytes. Dev. Compo Immunol., 19: 31-41.
PEDDIE, C.M., and SMllli, V.J., 1993. In vitro cytotoxic activity against mammalian target cells by the hemocytes of the solitary ascidian, Ciona intestinalis. J. Exp. Zool., 267: 616-623.
PEDDIE, C.M., and SMIlli, V.J., 1994. Mechanism of cytotoxic activity by hemocytes of the solitary ascidian, Ciona intestina!is. J. Exp. Zool., 270: 335-342.
PORCHET-HENNERE', E., DUGIMONT, T., and FISCHER, A., 1992. Natural killer cells in a lower invertebrate, Nereis diversicolor. Eur. J. Immunol., 58: 99-107.
RAITos, D.A., TAIT, N.N., and BRISCOE, D.A., 1987a. Allograft rejection and alloimmune memory in the solitary urochordate, Styelaplicata. Dev. Compo Immunol.. 11: 343-351.
RAITos, D.A., TAlT, N.N., and BRISCOE, D.A., 1987b. Cellular basis of allograft rejection in the solitary urochordate Styela pUcata. Dev. Compo Immunol., 11: 713-725.
SAITO, Y., and WATANABE, H., 1982. Colony specificity in the compound ascidian, Botryllus scalaris. Proc. Jpn. Acad., IBI 58: 105-108.
Received April 1995; Accepted July 1995
Address and correspondence:
Matteo Cammarata Laboratory of Marine Immunobiology Institute of Zoology Via Archirafi, 18 1-90123 PALERMO ITALY
SCHROlT, A.J., TANAKA, Y., MADSEN, J., and FIDLER, I.J., 1984. The recognition of red blood cells by macrophages: role of phosphatidyIserine and possible implications of membrane phospholipid asymmetry. BioI. Cell., 51: 227-238.
SHIGA, T., and MAEDA, N., 1980. Influence of membrane fluidity on erythrocyte functions. Biorheology, 17: 485-494.
SODERHALL , K., WINGREN, A., JOHANSSON, M.W., and BERlliEUSSEN, K., 1985. The cytotoxic reaction of hemocytes from the freshwater crayfish Astacus astacus. Ceil. Immunol, 94: 326-332.
SOKAL, RR, and ROHLF, F.J., 1981. Biometry. W.H. Freeman and Company, New York, p.859.
TSCHOPP, J., SCHAFER, S., MASSON, D., PEITSCH, M., and HEUSSER, C., 1989. PhosphoryIcholine acts as Ca2+-dependent receptor molecule for lymphocyte perforin. Nature, 337: 272-274.
TYSON, C.J., and JENKIN, C.R, 1974. The cytotoxic effect of haemocytes from the crayfish (Paracentrotus bicarinatus) on tumor cells of vertebrates. Aust. J. Exp. BioI. Med. Sci., 52: 915-923.
WADA, H., and SATOH, N., 1994. Details of the evolutionary history from invertebrates to vertebrates, as deduced from the sequences of 18S rDNA. Proc. Natl. Acad. Sci., 91: 1801-1804.
UTSUGI, T., SCHROlT, A.J., CONNOR, J., BUCANA, C.D., and FIDLER, I.J., 1991. Elevated expression of phosphatidylserine in the outer membrane leaflet of human tumor cells and recognition by activated human blood monocytes. Cancer Res., 51: 3062-3066.
YUE, C.C., REYNOLDS, C.W., and HENKART, P.A., 1987. Inhibition of cytolysin activity in large granular lymphocyte granules by lipids: evidence for a membrane insertion mechanism oflysis. Mol. Immunol., 24: 647-653.

1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
" 1
1
1
1
. 1
1
1
1
1
1
1
1
1




















