
Dr. Jekyll and Mr. Hyde - Distinctiveness and plasticity of mononuclear phagocytes in the mouse skin
Upload
independentCategory
view
0download
0
Journal of Neuroimmunology
ELSEVIER Journal of Neuroimmunology 56 (1995) 143-153
Cytokine secretion and eicosanoid production in the peripheral blood mononuclear cells of MS patients undergoing dietary supplementation
with n-3 polyunsaturated fatty acids
Virgilio Gallai a, * , Paola Sarchielli a, Albert0 Trequattrini a, Maristella Franceschini ‘, Ardesio Floridi b, Caterina Firenze a, Andrea Alberti a, Daniela Di Benedetto a,
Eduardo Stragliotto ’ a Neurological Clinic, Universi@ of Perugia, Via E. Dal Pozzo, Casella Postale 1427 Succ. 3, Policlinico Monteluce, 06126 Perugia, Italy
b Department of Cellular and Molecular Biology, University of Perugia, Perugia, Italy ’ Medical Department Pharmacia Farmitalia Carlo Erba, Milan, Italy
Received 24 March 1994; revised 13 September 1994; accepted 13 September 1994
Abstract
To demonstrate the influence of n-3 PUFA supplementation on cytokine and eicosanoid production in peripheral blood mononuclear cells (PBMCS) of MS patients (MSP), we investigated the impact of a g-month dietary supplementation with these fatty acids on the levels of interleukin-lbeta (IL-l@), IL-2, interferon-gamma (IFN-y) and tumor necrosis factor-alpha (TNF-~Y) in the supernatants of stimulated PBMCs and serum soluble IL-2 receptors in a group of 20 relapsing-remitting (R-R) MSP and a group of 15 age-matched control individuals (CI). The production of PGE2 and LTB4 in the stimulated PBMCs was also assessed in patient and control groups supplemented with n-3 PUFAs. In both groups, n-3 PUFA supplementation led to a significant decrease in the levels of IL-l/? and TNFIu, and this reduction was more pronounced in the 3rd and 6th month of supplementation. An analogous decrease was observed in the levels of IL-2 and IFN-y produced by stimulated PBMCs, and in the levels of serum soluble IL-2 receptors. n-3 PUFA supplementation also appeared to significantly affect prostaglandin E2 (PGE2) and leukotriene B4 (LTB4) production in PBMCs, both in MSP and the control group. The reduced production of these proinflammatory eicosanoids, and the decrease of some cytokines with an immunohenancing effect as a consequence of n-3
PUFA supplementation, could modulate some immune functions which have been demonstrated to be altered in MSP.
Keywords: n-3 PUFA supplementation; Multiple sclerosis; Cytokines; Leukotriene B4; Prostaglandin E2; Mononuclear cells; Soluble interleukin-2 receptors
1. Introduction
A great number of investigations have demonstrated that n-3 PUFAs have a profound effect on the immune function, both in animals and humans (Chandra et al., 1989). They exert a great influence both on cell-media- ted immunity (inhibition of in vitro transformation to T or B cell mitogens and antigens, cellular cytotoxicity against allogeneic cells, the monocyte function and
* Corresponding author. Fax ( + 39-75) 578 3568
01655728/95/$09.50 0 1995 Elsevier Science B.V. All rights reserved SSDI 0165-5728(94)00140-5
antigen-presenting cell function) and humoral immu- nity (decreased production of autoantibodies) (Hwang, 1989; Kelley et al., 1991). The marked anti-inflamma- tory action of n-3 PUFAs might explain the beneficial effect that they have on various inflammatory condi- tions and on pathologies characterized by an immune dysregulation (arthritis, autoimmune nephritis, psoria- sis, ulcerative colitis) (Chandra et al., 1989).
Studies performed on experimental animals and hu- mans have suggested that n-3-PUFAs exert at least part of their immunoregulatory effect by a direct influ- ence on immune cell membranes, inducing changes in the structural components of cell membrane phospho- lipids. This leads to alterations in the binding site

144 V. Gallai et al. /Journal of Neuroimmunology 56 (1995) 143-153
environment and the mechanisms of signal transduc- tion during cell activation (Salem et al., 1986). More- over, n-3 PUFAs compete with arachidonic acid (AA), thereby influencing leukotriene (LTB4) and prosta- glandin (PG2) synthesis. (Needelman et al., 1979; Lee et al., 1988).
Due to their influence on both phospholipid mem- brane content and the metabolization of AA, these fatty acids could also profoundly affect the synthesis of the cytokines, both directly and indirectly, by influenc- ing eicosanoid formation (Endres et al., 1989; Santoli and Zurier, 1989; Virella et al., 1989; Molvig et al., 1991; Urkaze et al., 1991; Meydani, 1992; Sierra et al., 1992).
The down-regulation of the production of some proinflammatory cytokines may be useful for the treat- ment of many autoimmune diseases or diseases charac- terized by an immune dysregulation, including multiple sclerosis (MS).
An insufficient dietary PUFA intake has been sug- gested as a factor influencing the incidence, clinical outcome and progression of MS (Swank and Dugan, 1990).
This hypothesis gave a theoretical basis for succes- sive controlled supplementation studies with n-6 and n-3 fatty acids.
Some of these studies demonstrated that patients treated with n-6 PUFAs had a less pronounced in- crease of disability and a lower relapse rate than the control group (treated with a placebo), regardless of the disability grade; others, on the contrary, showed a failure of a therapeutic effect of these fatty acids (Millar et al., 1973; Bates et al., 1978; Paty et al., 1978; Dworkin et al., 1984). However, in a recent double- blind study on the effects of n-3 fatty acid supplemen- tation, acute relapsing MSP underwent a mild im- provement after treatment (Bates et al., 1989).
It has been suggested that an alteration in PUFA metabolism could contribute to the development of MS lesions by altering the myelin composition, the immune function and the platelet function. PUFA supplemen- tation could affect all these pathogenetic mechanisms: in particular it may induce modifications of some of the immunological functions which have been consid- ered to be related to the activity and the progression of the disease, and also by influencing eicosanoid produc- tion and cytokine secretion in PBMCs.
The aim of this study was to evidence whether a 6-month period of n-3 PUFA supplementation influ- ences the PBMCs’ production of certain cytokines with proinflammatory effects (IL-lp, TNF-a, IL-2, IFN-y), and eicosanoids with immunoenhancing properties (LTB4 and PG2), and also whether it influences the levels of soluble IL-2 receptors @IL-2R) in serum, both in MS patients with the R-R form, and age-matched control individuals.
2. Materials and methods
2.1. Patient and control groups
20 MSP (age range: 20-50 years) were recruited from the Neurology Out-Patient Department of Peru- gia University, Italy.
All patients had clinically definite multiple sclerosis according to the Criteria of Poser. They had multiple sclerosis in the R-R form and Kurtzke Expanded Dis- ability Status Scale scores of 5 or less (Schumacher et al., 1965; Kurtzke, 1983; Poser et al., 1983).
All patients had suffered at least three definite relapses of the disease and one of the relapses had occurred during the 24 months prior, but not in the last 3 months before, the admission to the trial. They had had no steroid supplementation, ACTH or immuno- suppressive treatment for a minimum of 6 months before recruitment, and they showed no changes in their dietary habits as a consequence of developing MS. All patients were in remission and had no inter- current illnesses.
The patients were examined at the beginning of the study, and in the lst, 3rd, and 6th months of supple- mentation, and in the 1st and 3rd months after the supplementation was suspended. Treatment with ACTH or prednisone was permitted when clinical ex- acerbation occurred and patients were withdrawn from the immunological assessment (but not from the clini- cal evaluation) following steroid treatment for relapses.
Fifteen healthy control individuals (CI) (8 males and 7 females) were also selected from the friends and spouses of the MSP, and they were assessed for the same parameters at each stage of the study.
More details on inclusion and exclusion criteria and other clinical information are given in an other paper (Gallai, V., Sarchielli, P., Franceschini, M., Trequat- trini, A., Paciaroni, M., Usai, F., Murasecco, D., Tognellini, R., Malagigi, B., Floridi, A. and Stragliotto, E., submitted for publication).
Informed consent was obtained from all subjects and approval of the study was obtained from the local ethics committee.
2.2. Oral fatty acid supplementation
All patients and control individuals were treated for 6 months with six l-g capsules a day of a fish body oil (ESAPENT supplied by Carlo Erba, Milan, Italy) with an EPA and DHA content of at least 86% (51% EPA; 31% DHA) and 0.40 g of D,r_-a-tocopherols. The six capsules could be taken all at once or in several doses.
2.3. Dietary advice
Patients and control individuals were provided with advice and a booklet setting out a diet designed to

V. Gallai et al. /Journal of Neuroimmunology 56 (1995) 143-153 145
avoid a high intake of animal fats and to ensure a plentiful intake of n-6 polyunsaturated fatty acids.
2.4. Assessment of cytokine and eicosanoid production in PBMCs and determination of soluble IL-2 receptors in serum
The following parameters were assessed at the basal time, after 1 month, 3 months and 6 months of supple- mentation, and 1 month and 3 months after the end of the treatment:
2.4.1. Production of ILl-/3, TNF-(Y and IL-2 in PBMCs PBMCs were prepared by gradient centrifugation.
10 ml of peripheral blood were diluted in a 1: 1 ratio with saline and stratified on 4 ml Ficoll-Hypaque (Nyegard, Oslo, Norway) in a 15ml conical tube. The tubes were centrifuged at 400 x g for 30 min at 20°C. Recovered PBMCs were washed twice with Hank’s Balanced Salt Solution (HBSS) (Flow Lab. MacLean, VI> by centrifugation at 100 X g for 10 min at 10°C. After the second centrifugation, the pelleted PBMCs were resuspended in RPMI-1640 tissue culture medium (Flow Lab.) containing 100 kU penicillin (Flow Lab.) and 100 mg streptomycin (Flow Lab.). Viable PBMCs were determined by Trypan blue exclusion.
PBMCs were cultured in 96-well flat-bottom plates at a concentration of 1 x lo6 cells per ml in RPM1 1640 and supplemented with 20% heat-inactivated (56°C 30 min) fetal calf serum (FCS), penicillin (100 U/ml), streptomycin (100 pg/ml), 2 mM L-glutamine and polymixin B (12.5 pg/ml) (all provided by Flow Lab.), with and without lipopolysaccharide (LPS) (Sigma Co., St. Louis, MO) (2.5 pg/ml) for IL-l@ and TNF-a assays, and with and without phytohaemoagglu- tinin (PHA) (Sigma Co.) (10 pg/ml) for IL-2 assays. After 24 h of incubation at 37°C the medium was replaced with a fresh medium that had been equili- brated overnight in an incubator in order to bring it to a pH of 7.2. After 24 h, 100 ~1 of the supernatant from each well were pooled (3 wells/sample) and stored in cryotubes at -70°C until analysis.
For determination of IL-lp in PBMC supernatants, the detection limit of the assay was 0.01 ng/ml and the working range 0.01-5 ng/ml. The inter- and intra-as- say coefficients of variation were 12% and 8%, respec- tively. No cross-reactivity was demonstrated with re- combinant IL-l/3, IL-2, INF-y or TNF-CX.
TNF-cr in the mononuclear cell culture supernatants was measured with a commercially available Biokine TNF-a Test kit (T cell Science Inc., Cambridge, MA) as recommended by the manufacturer. The detection limit of the assay was 0.010 ng/ml and the working range 0.01-3 ng/ml. The intra-assay and inter-assay coefficients of variation were 6% and lo%, respec- tively. Levels of immunoreactive IL-2 in the super- natants of mononuclear cell cultures, frozen at -70°C
until determination, were measured using a commer- cial kit (Intertest 2R, Genzyme Corps, Boston, MA). The kit was a solid phase enzyme-linked immunosor- bent test (ELISA), consisting of monoclonal mouse anti-rabbit Ig (absorbed to mouse tissue and alkaline phosphate linked) and recombinant human IL-2. This kit was able to detect concentrations of IL-2 ranging from 0.05 to 500 U/ml.
2.4.2. sIL-2R assay Serum sIL-2R levels were measured by ELISA using
a commercial kit (Cell free, T cell Sciences, Cam- bridge, MA) according to the manufacturer’s instruc- tions. An internal standard supplied by the manufac- turer and a serum with high values of sIL2-2R, diluted 5-fold, were tested together with the samples under study.
The detection limit of the assay was 18 U/ml. The intra-assay and inter-assay coefficients of variation were 8% and lo%, respectively.
2.4.3. Interferon-y production in concanavalin A-stitnu - lated cultures
This technique was performed following the sugges- tions of Beck et al. (1990). The whole blood technique was described in detail by Kirchner et al. (19821.
Blood was collected in preheparinized syringes and mixed at a ratio of 1: 1.5 with RPM1 1640 medium supplemented with 2 mM glutamine, 100 III/ml peni- cillin and 100 pg/ml streptomycin (all from Flow Laboratories) without any additional serum. The white cell blood count was performed routinely and, in the range obtained, the cell numbers did not influence the level of IFN-y production. The cultures were incu- bated with concanavalin A (ConA) (final concentra- tion: 5 kg/ml) (both from Sigma Co.). Unstimulated cells were used as a control. All cultures were incu- bated for 48 h in round-bottom well plates (Nunc, Roskilde, Denmark). The supernatants were harvested and stored at -70°C until assayed.
IFN-7 production in the supernatants was measured by a two-site immunoradiometric assay according to the manufacturer’s instructions (Sucrosep IFN--y IRMA, Boots-cell Tech. Diagnostic Ltd., UK). Linear- ity of the standard curve was obtained for IFN-y con- centrations of up to 500 III/ml. The intra-assay coeffi- cients of variation (CV) were less than 5% over a range of 50-1000 IU/ml, but less than 2% over the range 230-460 IU/ ml.
2.4.4. LTB4 and PGE2 assays in the supernatants of PBMCs
PBMCs were cultured with and without LPS (25 pg/ml) in 96-well flat-bottom plates at a concentration of 1 X lo6 cells per ml in RPM1 1640, and supple- mented with 5% heat-inactivated (56°C 30 min) fetal bovine serum, 20% penicillin (100 U/ml), strepto-

146 V. Gal& et al./Joumal of Neuroimmunology 56 (1995) 143-153
mycin (100 pg/ml), 2 mM L-glutamine and polymixin 2.4.6. PGE, assay B (12.5 pg/ml) (all from Flow Lab.). After 24 h of The PGE, concentration in PBMCs was determined incubation at 37°C the medium was replaced with by radioimmunoassay (RIA). The detection limit of the fresh medium that had been equilibrated overnight in assay was approximately 2 pg 0.002 ng/tube. The the incubator to bring it to a pH of 7.2. After another intra- and inter-assay coefficients, in the range of 24 h, 100 ~1 of the supernatants from each well were 0.002-0.15 ng, were 4% and 6% respectively. There pooled (three wells/ sample) and stored in cryotubes at was no cross-reactivity with PGE,. - 70°C before analysis.
2.4.5. LTB, assay 2.5. Statistical analysis
The LTB4 in the supernatants of PBMCs were Data were expressed as mean +SEM. Statistical measured with a commercially available kit (Amersham, analysis included one-way analysis of variance and Cat NO, TRK 9809). The detection limit of the assay paired and unpaired Student’s t-test. The analysis of was approximately 0.016 ng/tube. The intra- and in- variance (ANOVA) with Tukey’s test was performed to ter-assay coefficients of variation were respectively, in the range 0.016-0.1 ng.
4 ILi 6 WnL)
7% and 9% compare the mean values of each of the evaluated variables at the various stages of the study: at the basal
without LPS
0 MSP
r-l A Cl
W IL1 6 (nglml)
5
4
3 ii 2
1
with LPS
0 MSP L-l A Cl
0 basal lslmonth 3rdnnnth 6thmmth lstrnonth period
3rdmonth SUP@. SUppI. SUppI. aftw SUSp. after Sup.
Fig. 1. IL-lp levels (ng/ml) in the supernatants of PBMC cultures with (a) and without (b) LPS in patients with MS (MSP) and in healthy
individuals (CI), at the basal time, in the lst, 3rd, 6th month of n-3 PUFA supplementation (suppl.) and in the 1st and 3rd month after suspension (susp.) of supplementation.

V. Gallai et al. /Journal of Neuroimmunology 56 (1995) 143-153 147
time, at the end of the lst, 3rd and 6th month of n-3 fatty acid supplementation, and at the end of the 1st and 3rd month of suspension of the treatment. Fisher’s least significant difference (LSD) was also performed to compare the main effect means in ANOVA in the case of equal cell sizes @AS User’s Guide: Statistics, 1986).
Five per cent for one-sided tests was chosen as the level of statistical significance unless otherwise stated.
3. Results
3.1. Production of IL-lp, TNF-(Y, IL-2 and IFN-y in PBh4Cs
The basal production of IL-lp and TNF-(Y in the PBMCs of MSP, regardless of stimulation with LPS (25
a) TNF CX (n@mL)
41
Ops
006
004 /
002
i i
b) TNF o( (n@mL)
3 I
pg/ml), and that of IL-2 in PBMCs, whether sponta- neous or PHA-induced, did not differ significantly from production of the control group. Additionally, at the beginning of the study, no statistically significant differ- ences were found between the levels of IFN-y pro- duced by PBMCs in the presence of ConA in MSP and CI. The values of these four cytokines in the super- natants of PBMCs (whether stimulated or not) were found to be significantly decreased in the 1st month of supplementation with respect to the basal period, both in MSP and CI. The reduction in the content of all the cytokines in PBMC supernatants was more accentu- ated in the 3rd and 6th months of supplementation (suppl.) in both control and patient groups (MSP: ANOVA without stimulus IL-l& P < 0.03; TNF-cu, P < 0.02; IL-2, P < 0.002; and IFN-y, P < 0.01; with stimulus IL-l/?, P < 0.01; TNF-a, P < 0.02; IL-2, P <
without LPS
ri I
lstrwmth Jrdmorlth ahmonth 1StmOllth 3rdmonth
WM. SuppI. suppl. after-p. after susp.
with LPS
l MSP u A Cl
1st month SUPpl.
3rd month 6thmonth lstrnonth 3rd month
SUPpl. SuppI. after susp. after susp.
Fig. 2. TNF-(Y levels (ng/ml) in the supernatants of PBMC cultures with (a) and without (b) LPS in patients with MS (MSP) and in healthy
individuals (CI), at the basal time, in the lst, 3rd, 6th month of n-3 PUFA supplementation (suppl.) and in the 1st and 3rd month after
suspension (susp.) of supplementation.

148 VI Gallai et al. /Journal of Neuroimmunology 56 (1995) 143-153
0.003; and IFN-y, P < 0.005. CT: ANOVA without stimulus IL-l/?, P < 0.04; TNF-a, P < 0.03; IL-2, P < 0.005; and IFN-7, P < 0.03; with stimulus IL-lp, P < 0.02; TNF-a, P < 0.03; IL-2, P < 0.003; and IFN-y, P < 0.004) (Figs. 1-4). The cytokine values returned towards basal levels after suspension (susp.) of the n-3 PUFA supplementation, with the exception of IL-2 which rose progressively during the follow-up period but did not reach the basal values even after the 3rd month of suspension.
duction was already present from the 1st month of supplementation in MSP (ANOVA: P < 0.002). The values remained below the basal levels even in the 3rd month after suspension. A significant decrease in serum sIL-2R also occurred in control subjects. This reduc- tion, as in MSP, was most pronounced in the 6th month of supplementation (ANOVA: P < 0.004).
Significantly lower values of sIL-2R persisted in the serum even after suspension of the treatment in both patient and control groups (Fig. 5).
3.2. Serum sIL-2R 3.3. LPS-stimulated secretion of LTB4 and PGE2 in PBMCs
The MS patients displayed higher (albeit not signifi- cantly) values of serum sIL-2R than control subjects. During the treatment with n-3 PUFAs the sIL-2R levels tended to be significantly reduced, and this re-
The spontaneous and LPS-stimulated secretion of LTB, and PGE, in PBMCs did not differ between groups at the beginning of the study.
50
40
30
20
10
4
IL2 (UlmL)
without PHA
. MSP L-A A Cl
T
1: ii
lstmonth sdmonth 6thmorrth lstrmnth 3fdmonth
period SUppI. sup@. SUppl. -susp -swp
b)
IL2 (U/ml) with P HA
200 /T T
jrCI 1
6th month 11 month 3rd month SUpQl. @twwsp --rJ.
Fig. 3. IL-2 levels (U/ml) in the supernatants of PBMC cultures stimulated with PI-IA in patients with MS (MSP) and in healthy individuals (Cl), at the basal time, in the lst, 3rd, 6th month of n-3 PUFA supplementation (suppl.) and in the 1st and 3rd month after suspension (susp.) of supplementation.
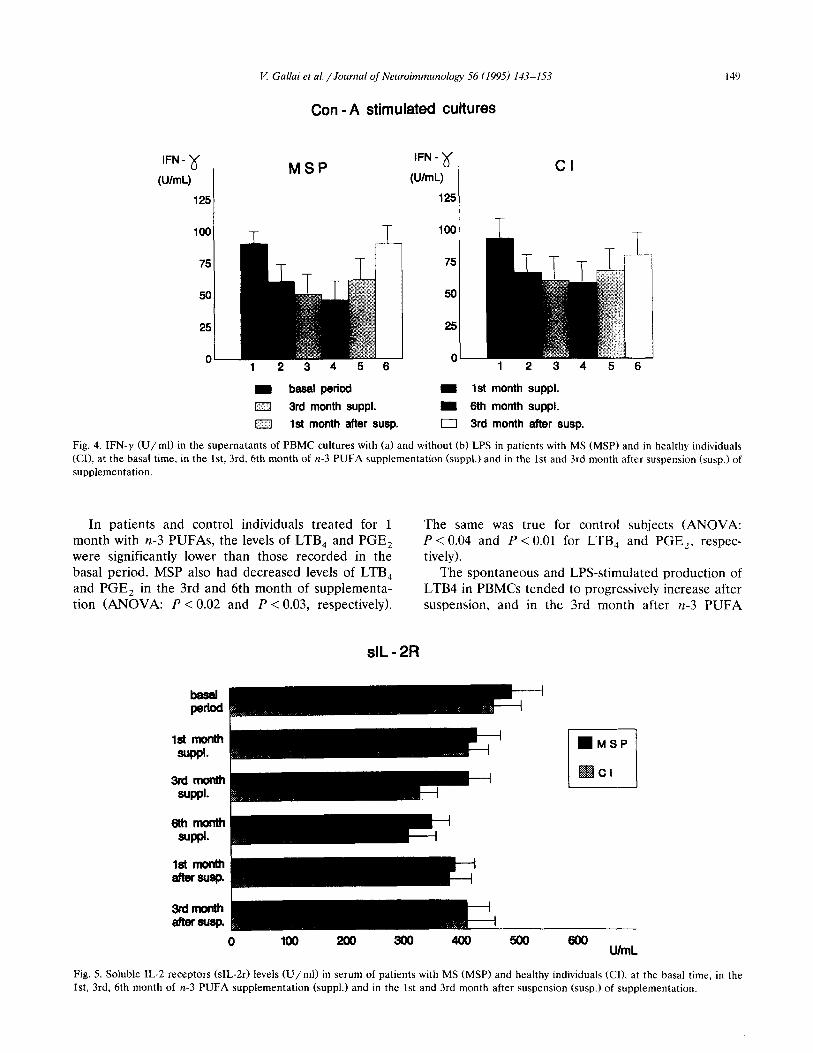
V. Gallai et al. /Journal of Neuroimmunology 56 (1995) 143-153 14’)
Con -A stimulated cultures
IFN-x
(UlmL)
125
MSP IFN-‘d
(U/ml)
125
Cl
T T 1 loo
25
0 123456
0 123456
w baaal period m 1st month suppl.
1=3 3rd month suppl. I 6th month suppl.
El 1st month after susp. 0 3rd month after susp.
Fig. 4. IFN-y W/ml) in the supernatants of PBMC cultures with (a) and without (b) LPS in patients with MS (MSP) and in healthy individuals
(CI), at the basal time, in the lst, 3rd, 6th month of n-3 PUFA supplementation (suppl.) and in the 1st and 3rd month after suspension (susp.) of
supplementation.
In patients and control individuals treated for 1 The same was true for control subjects (ANOVA: month with n-3 PUFAs, the levels of LTB, and PGE, P < 0.04 and P < 0.01 for LTB, and PGE2, respec- were significantly lower than those recorded in the tively). basal period. MSP also had decreased levels of LTB, The spontaneous and LPS-stimulated production of and PGE, in the 3rd and 6th month of supplementa- LTB4 in PBMCs tended to progressively increase after tion (ANOVA: P < 0.02 and P < 0.03, respectively). suspension, and in the 3rd month after n-3 PUFA
slL - 2R
0 100 zoo 300 400 500 600 WmL
Fig. 5. Soluble IL-2 receptors &IL-2r) levels (U/ml) in serum of patients with MS (MSP) and healthy individuals (Cl), at the basal time, in the
1st 3rd, 6th month of n-3 PUFA supplementation (suppl.) and in the 1st and 3rd month after suspension (susp.) of supplementation.

Tab
le
1 PG
E2
(ng/
ml)
A
ND
L
TB
4 (n
g/m
l)
in t
he
supe
rnat
ants
of
PB
MC
cu
lture
s w
ith
(a)
and
with
out
(b)
LPS
jn
pat
ient
s w
ith
MS
and
in h
ealth
y in
divi
dual
s (C
D,
at
the
basa
l tim
e,
in t
he
1st
3rd,
6t
h
mon
th
of
n-3
PUFA
su
pple
men
tatio
n an
d in
th
e 1s
t an
d 3r
d m
onth
af
ter
susp
ensi
on
of
supp
lem
enta
tion
Bas
al
peri
od
1st
mon
th
supp
l. 3r
d m
onth
su
ppl.
6th
mon
th
supp
l. 1s
t m
onth
af
ter
susp
. 3r
d m
onth
af
ter
susp
.
Non
e L
PS
Non
e L
PS
Non
e L
PS
Non
e L
PS
Non
e L
PS
Non
e L
PS
PG
E2
hg/m
l)
Con
trol
s 0.
19
f0.0
3 1.
62kO
.16
0.13
kO
.04
1.34
kO.2
3 0.
12
kO.0
2 1.
25+0
.14
0.12
kO
.02
1.16
kO.1
5 0.
14
kO.0
3 1.
33dz
o.14
0.
15
kO.0
2 1.
41 k
O.1
6
MS
patie
nts
0.17
kO
.02
1.36
+0.1
7 0.
12
kO.0
3 1.
24kO
.15
0.13
f0
.03
1.13
*0.1
4 0.
11
kO.0
3 1.
05*0
.18
0.12
kO
.02
1.34
kO.1
4 0.
13
+0.0
2 1.
53kO
.17
LT
B4
(ng/
mB
Con
trol
s 0.
O72
fO.0
07
O.l1
+0.0
7 0.
063+
0.00
9 0.
08+0
.02
0.03
5+0.
004
0.07
+0.0
06
0.01
5+0.
006
0.07
+0.0
3 0.
032+
0.00
7 0.
05f0
.02
0.04
6+0.
008
0.04
+0.0
6
MS
patie
nts
0.06
1 f
0.00
5 0.
12 f
0.
05
0.05
2 +
0.00
7 0.
09 +
0.0
3 0.
034
f 0.
005
0.08
0.
02
+ 0.
023
+ 0.
007
0.08
0.
02
k 0.
041+
0.
008
0.12
kO.0
3 0.
052z
kO.0
06
0.17
+0.0
5

V. Gallai et al./Journal of Neuroimmunology 56 (1995) 143-153 151
suspension the values of both control and patient groups reached the levels recorded at the beginning of the study.
A progressive rise in the production of PGE2 was found in both groups after suspension of the treatment, and the levels returned to the range of basal values after the 3rd month of suspension (Table 1).
4. Discussion
There has been controversy over the implications of clinical results of supplementation studies with PUFAs on the course and activity of MS. We therefore at- tempted to verify the effects of n-3 PUFA supplemen- tation on some immunological parameters in MSP which have been considered to be markers of disease activity in MS.
We demonstrated that the release of IL-l& TNF-a and IL-2 by PBMCs is profoundly affected by a 6-month supplementation with n-3 PUFAs, both in MSP and in control subjects. This can be of relevance in treatment of MS, in which the central role of these cytokines in the pathogenesis of demyelinating lesions has been clearly demonstrated.
Patients with chronic P and R-R forms of MS are sometimes found to have elevated levels of IL-2, TNF- cy, IFN-y and sIL-2R in their serum and cerebrospinal fluid compared to healthy subjects and sufferers from other neurological diseases (Beck et al., 1988; Adachi et al., 1989; Papiha et al., 1989; Capra et al., 19901. Few reports have been devoted to IL-l, a cytokine potentially involved in the pathogenesis of MS lesions because it can act synergistically with TNF-a in proin- flammatory reactions, potentiating the synthesis of en- dogenous IL-2 (Peter et al., 1991). Like IL-2, it can increase the expression of IL-2R. In any case, measure- ment of the levels of these cytokines in serum should be carefully used for therapeutic and prognostic pur- poses with MSP. More information could be obtained by determining the production of these cytokines in the supernatants of mitogen-stimulated PBMCs. However, few studies have been performed to verify the varia- tions in this production in relation to disease activity (Selmaj et al., 1988).
In our research no significant variations were found between the levels of IL-l, TNF-cw, IFN-7 and IL-2 in the supernatants of PBMCs in MSP (with and without LPS, PHA or ConA as a stimulus) and those in CI at the beginning of the study. It must be taken into account that none of our MSP were in relapse, and this could explain the finding that the PBMC production of these proinflammatory cytokines did not significantly differ from that in control subjects. In both MSP and CI groups, n-3 PUFA supplementation induced a sig- nificant suppression of the production of all these
cytokines after only 4 weeks of supplementation. 12 weeks after the end of supplementation all cytokine levels returned to the pre-supplementation values, with the exception of IL-2, the levels of which remained significantly lower even 3 months after suspension of PUFA supplementation. The decreased production of IL-l and TNF-(Y in the PBMCs of MSP in our study confirms the results of previous research performed on healthy subjects, insulin-dependent diabetic patients, and patients with rheumatoid arthritis supplemented with n-3 PUFAs (Endres et al., 1989; Meydani et al., 1991; Molvig et al., 19911.
Since IL-lp and TNF-(r are the principal mediators of many inflammatory responses, the reduced produc- tion of these cytokines in LPS-stimulated PBMCs evi- denced by our study may contribute to the modulation of the immune function in MSP supplemented with n-3 PUFAs. It must be taken into consideration that im- munosuppression similar to that induced by n-3 PU- FAs can only be achieved by the administration of glucocorticoids or cyclosporine A, which have well- known adverse side effects, particularly after long-term administration.
In our study we also observed a reduction in the production of IL-2 in PBMCs supernatants and in the serum levels of sIL-2R in both patients and control subjects supplemented with n-3 PUFAs. IL-2 plays a key role in promoting the growth and proliferation of T lymphocytes by interacting with its specific IL-2 recep- tors (Smith and Cantrell, 198.5). High amounts of sIL- 2R in serum might indicate a systemic immune activa- tion, leading to a persistent trafficking of activated T cells between the blood and the CNS, particularly in the P form but also in the R-R form of MS. Lowering the peripheral levels of IL-2 and sIL-2R in the course of MS could be useful for modulating and reducing in vivo immune system activation. The results of our study, which evidenced a decreased production of IL-2 in the PBMCs of MSP and CI, concur with our finding of reduced IL-2 receptor expression in the PBMCs of MSP supplemented with n-3 fatty acids (Gallai, V., Sarchielli, P., Franceschini, M., Trequattrini, A., Pacia- roni, M., Usai, F., Murasecco, D., Tognellini, R., Malagigi, B., Floridi, A. and Stragliotto, E., submitted for publication). Moreover, these results confirm those of in vitro research which demonstrate that the addi- tion of PUFAs to human PBMCs induces a lowering in IL-2 receptor expression and IL-2 production (Santoli and Zurier, 19891.
In the present study, n-3 PUFA supplementation appeared to significantly affect PGE2 and LTB4 pro- duction in PBMCs both in MSP and the control group.
PGEs of the 2-series (in particular PGE2) are im- portant mediators of inflammatory responses and influ- ence many immunological functions (lymphocyte prolif- eration, natural killer activity and other lymphocyte

1.52 K Gallai et al. /Journal of Neuroimmunology 56 (1995) 143-153
functions mediated by the increase of CAMP levels) (Hwang, 1988). On the other hand, LTB4 is an impor- tant stimulator of leukocyte function (Fisher et al., 1986). The implication of some proinflammatory eicosanoids (PGE2 and LTB4) in the pathogenesis of MS has been prospected, and in part demonstrated, by studies performed on peripheral blood immune cells (Prosiegel et al., 1987; Aberg et al., 1990, Dore-Duffy and Donovan, 1991; Reder et al., 1991; Sierra et al., 1992).
Eicosanoids derived from n-3 PUFAs, particularly EPA, are of the trienoic series because of the presence of three double bonds in their structures. As men- tioned before, the competitive effect of n-3 and n-6 PUFAs on various cellular functions and clinical dis- eases can depend, at least in part, on the differing eicosanoid production. In particular higher doses of n-3 fatty acids induced an increased production of PG13, TX3 and LTBS. The suppressed synthesis of LTB4 observed in our study during n-3 PUFA treat- ment, both in MSP and control subjects, confirmed the previous findings of a reduced leukocyte production in this eicosanoid with chemotactic and inflammatory ac- tivity, after fish oil supplementation (Fisher et al., 1987; Lee et al., 1988). The substitution of LTB4 with a lipoxygenase product, characterized by much lower in- flammatory activity (LTBS), could also contribute to the modulation of the monocyte and lymphocyte func- tion by reducing the production of cytokines involved in inflammatory processes, such as IL-1 and TNF. This could be of importance in the prevention of the im- munological alterations involved in the pathogenesis of MS lesions. The inhibition of PGE2 and the induction of PGE3 could also be implicated in the regulation of some immune responses, including T-cell proliferation, IL-2 secretion and cytotoxicity.
In conclusion, these data add further evidence of the modulatory effects of n-3 PUFA supplementation on some aspects of the immune function, both in MSP and in CI. The effects can be ascribed not only to the modification of the immune cell membrane composi- tion but also to the modification of the metabolic pathways involved in immune activation, particularly cytokine production and eicosanoid formation. n-3 PU- FAs may also contribute to the inhibition of leuko- cyte/ endothelium interaction, which plays a central role in the induction and maintenance of brain tissue damage in demyelinating diseases. This effect might be mediated by lowering of the LTB4 production which is implied in the up-regulation of CDllb/C18 adhesion receptors and by the reduction of platelet-activating factor production (Fisher et al., 1986; Fox and Di Corleto, 1988).
The fatty acid composition in PBMC membranes before, during and after n-3 supplementation, both in MSP and CI, will be the subject of a successive study.
Acknowledgement
We thank Carlo Erba Farmitalia, LTD, Milan, Italy for supplying the capsules containing n-3 PUFAs free of charge.
References
Aberg, J.A., Demers, L.M., Romano, P.J. and Tenser, R.B. (1990) Prostaglandin production in chronic progressive multiple sclero- sis. J. Clin. Lab. Anal. 4, 246-250.
Adachi, K., Kumamoto, T. and Araki S. (1989) Interleukin-2 recep- tor levels indicating relapse in multiple sclerosis. Lancet i, 559- 560.
Bates, D., Fawcett, P.R.W., Shaw, D.A. and Weightman D. (1978) Polyunsaturated fatty acids in treatment of acute remitting multi- ple sclerosis. Br. Med. J. 2, 1390-1392.
Bates, D., Cartlidge, N.E.F., French, J.M., Jackson, M.J., Nightin- gale, S., Shaw, D.A., Smith, S., Woo, E., Hawkins, S.A. and Millar, J.H. (1989) A double blind controlled trial of long chain n-3 polyunsaturated fatty acids in the treatment of multiple sclerosis. J. Neurol. Neurosurg. Psychiatr. 52, 18-22.
Beck, J., Rondot, P., Catinot, L., Falcoff, E., Kirschner, H. and Wietzerbin J. (1988) Increased production of interferon gamma and tumor necrosis factor precedes clinical manifestation in mul- tiple sclerosis: do cytokines trigger off exacerbations? Acta Neu- rol. Stand. 78, 318-323.
Capra, R., Mattioli, F., Marciano, N., Vignolo, L.A., Bettinzioli, M., Airo, P. and Cattaneo R. (1990) Significantly higher levels of soluble interleukin 2 in patients with relapsing-remitting multiple sclerosis compared with healthy subjects. Arch. Neurol. 47, 254.
Chandra, R.K. (1989) Health Effects of Fish and Fish Oils. ARTS Biomedical, St John’s, New-Foundland, Canada.
Dore-Duffy, P. and Donovan, C. (1991) Functional desensitization of monocytes from patients with multiple sclerosis due to the prostaglandin E. Clin. Immunol. Immunopathol. 61, 119-128.
Dworkin, R.H., Bates, D., Millar, J.H.D. and Paty, D.W. (1984) A re-analysis of three double-blind trials. Neurology 34, 1441-1445.
Endres, S., Ghorbani, R., Kelley, V., Georgilis, K., Lonnemann, G., van der Meer, J.W., Cannon, J.G., Rogers, T.S., Klempner, M.S. and Weber, P.C. (1989) The effect of dietary supplementation on the synthesis of interleukin-1 and tumor necrosis factor by mononuclear cells. N. Engl. J. Med. 320, 265-271.
Fisher, M., Upchurch, K.S., Levine, P.H., Johnson, M.H., Vaudreuil, C.H., Natale A. and Hoogasian, J.J. (1986) Effect of dietary fish oil supplementation on polymorphonuclear leukocyte inflamma- tory potential. Inflammation 10, 387-392.
Fisher, S., Preac-Mursic, V. and Weber, P.C. (1987) Dietary docosa- hexaenoic acid is retroconverted in man in eicosapentaenoic acid, which can quickly be transformed to prostaglandin 13. Prostaglan- dins 34, 367-375.
Fox, P.L. and Di Corleto, P.E. (1988) Fish oils inhibit endothehal production of platelet-derived growth factor-like protein. Science 241, 453-456.
Hwang, D.H. (1989) Essential fatty acids and immune response. FASEB J. 3,2052-2061.
Hwang, D.H., Bouderau, M. and Chanmugan, P. (1988) Dietary linoleic acid and longer chain n-3 fatty acids: comparison effects on arachidonic acid metabolism in rats. J. Nutr. 118, 427-437.
Kelley, D.S., Branch, L.B., Love, J.E., Taylor, PC., Rivera, Y.M. and Iacono, J.M. (1991) Dietary alpha-linoleic acid and immunocom- petence in humans. Am. J. Nutr. 53, 40-46.
Kirschner, H., Kleinicke, C. and Digel, W. (1982) A whole-blood

I/: Gallai et al. /Journal of Neuroimmunology 56 (1995) 143-153 153
technique for testing production of human interferon by leuko- sclerosis: guidelines for research protocols. Ann. Neurol. 13,
cytes. J. Immunol. Methods 48, 213-219. 227-231.
Kurtzke, J.F. (1983) Rating neurologic impairment in multiple scle-
rosis. Neurology 33, 1444-1452.
Lee, T.H., Sethi, T., Crea, A.E., Peters, W., Arm, J.P., Horton, C.E.,
Walport, M.J. and Spur, B.W. (1988) Characterization of
leukotriene B,: comparison of its biological activity with
leukotriene B, and leukotriene B, in complement receptor en-
hancement, lysozyme release and chemotaxis of human neu-
trophils. Clin. Sci. 74, 467-475.
Prosiegel, M., Neu, I., Wildfeur, A., Mehlber, L., Malhnger, J. and
Ruhenstroth-Bauer, G. (1987) Leukotrienes B4 and C4 in MS.
Acta Neural. Stand. 75, 361-363.
Meydani, S.N. (1992) Modulation of cytokine production by dietary
polyunsaturated fatty acids. PSEBM 200, 189-193.
Millar, J.H., Zilkha, K.J., Langman, M.J., Wright, H.P., Smith, A.D.,
Belin, J. and Thompson, R.H. (1973) Double blind trial of
linoleate supplementation of the diet in multiple sclerosis. Br.
Med. J. 31, 765-768.
Reder, A.T., Amason, B.G., Maimoneone, D. and Rohwer-Nutter,
D. (1991) The function of the CD2 protein is abnormal in
multiple sclerosis. J. Autoimmun. 4, 479-491.
Salem, N., Kim, H.Y. and Yergey, J.A. (1986) Docosahexaenoic acid:
membrane function and metabolism. In: A.P. Simopulous, R.R.
Kifer and R.E. Martin (Eds.), Health Effects of Polyunsaturated
Fatty Acids in Seafoods. Academic Press Inc, Orlando, FL, pp.
263-318.
Molvig, J., Pociot, F., Worsaae, H., Wogensen, L.D., Baek, L.,
Christensen, P., Mandrup-Poulsen, T., Andersen, K., Madsen, P.,
Dyerberg, J. and Nerup, J. (1991) Dietary supplementation with
w-3 polyunsaturated fatty acids decreases mononuclear cell pro-
liferation and interleukin-lp content but not monokine secretion
in healthy and insulin-dependent diabetic individuals. Stand. J.
Immunol. 34, 399-410.
Santoh, D. and Zurier, R.B. (1989) Prostaglandin E precursor fatty
acids inhibit human IL-2 production by a prostaglandin E-inde-
pendent mechanism. J. Immunol. 143, 1303-1309.
SAS Institute Inc. SAS/SAT user’s guide, relase 6.03. Edition Gary
NC SAS Institute Inc, 1986.
Needeleman, P., Raz, A., Minkes, M.S., Ferrendelli, J.A. and
Sprecher, H. (1979) Triene prostaglandins: prostacychn and
thromboxane biosynthesis and unique biological properties. Proc.
Natl. Acad. Sci. USA 76, 944-948.
Papiha, S.S., Boddy, J., Roberts, D.F. and Bates, D. (1989) PHA-in-
duced interferon in multiple sclerosis: association between gamma
interferon and clinical and genetical variables. Acta Neural.
Stand. 80, 145-150.
Paty, D.W., Cousin, H.K., Read, S. and Adlakha, K. (1978) Linoleic
acid in multiple sclerosis: failure to show therapeutic benefit.
Acta Neurol. Stand. 58, 53-58.
Peter, J.B., Boctor, F.N. and Tourtellotte, W.W. (1991) Serum and
CSF levels of IL-2, sIL2-R, TNF-(Y, and IL-1 p in chronic
progressive Multiple Sclerosis: expected lack of clinical utility.
Neurology 41, 121-123.
Schumacher, G.A., Beebe, G., Kibler, R.F., Kurland, L.T., Kurtzke,
J.F., McDowell, F., Nafgler, B., Sibley, W.A., Tourtellotte, W.W.
and Williman, J. (1965) Problems of experimental trials of thera-
phy in multiple sclerosis. Ann. N.Y. Acad. Sci. 122, 552-568.
Selmaj, K., Nowak, Z. and Tchorzewski, H. (1988) Interleukin-1 and
interleukin-2 production by peripheral blood mononuclear cells
in multiple sclerosis patients. J. Neural. Sci. 85, 67-76.
Sierra, A., Ferrer, I., Arbizu, T. and Buendia, E. (1992) Multiple
sclerosis: IL-2, IFN-y and PGE2 secretion in mononuclear cell
cultures stimulated with PHA and the effect of these cytokines on
oligodendroglial cells. Neurologia 7, 247-253.
Smith, K.A. and Cantrell, D.A. (1985) Interleukin 2 regulates its own
receptors. Proc. Nat]. Acad. Sci. USA 82, 864-868.
Swank, R.L. and Dugan, B.B. (1990) Effect of low saturated fat diet
in early and late cases of multiple sclerosis. Lancet 336, 37-39.
Urakaze, M., Sugiyama, E., Xu, L., Auron, P., Yeh, E. and Robin-
son, D. (1991) Dietary marine lipids suppress IL-lb mRNA levels
in lipopolysaccharide stimulated monocytes. Clin. Res. 3, 475
(abstract).
Poser, C.M., Paty, D., Scheinberg, L., McDonald, W.I., Davis, F.A.,
Ebers, G.C., Johnson, K.P., Sibley, W.A., Silberberg, D.H. and
Tourtellotte, W.W. (1983) New diagnostic criteria for multiple
Virella, G., Kilpatrick, J.M., Rugeles, M.T., Hyman, B. and Russell,
R. (1989) Depression of humoral responses and phagocytic func-
tions in vivo and in vitro fish oil and eicosapentanoic acid. Clin.
Immunol. Immunopathol. 52, 257-270.
Copyright © 2022 FDOKUMEN




















