
Cytogenetic Analysis of 124 Prospectively Ascertained Male Germ Cell Tumors1

Fax +41 61 306 12 34E-Mail [email protected]
Chromosome Dynamics
Cytogenet Genome Res 2009;127:287–296 DOI: 10.1159/000304046
Cytogenetic Characterization and Fluorescence in situ Hybridization of (GATA) 10 Repeats on Established Primary Cell Cultures from Indian Water Snake (Natrix piscator) and Indian Mugger (Crocodylus palustris) Embryos
L. Rao R. Turlapati M. Patel B. Panda D. Tosh S. Mangalipalli A. Tiwari V.P. Orunganti D. Rose A. Anand M.K. Kulashekaran S.R. Priya R.K. Mishra K. Majumdar R.K. Aggarwal L. Singh
Centre for Cellular and Molecular Biology, Hyderabad , India
ed with fetal bovine serum and subcultured for a period of 8–10 months. The morphology of cell types was kept under constant observation microscopically. Interestingly, at a sub-sequent passage of the cells sporadically scattered neuro-nal-like and beating cells were observed. The suitable tem-perature for growth of these cell cultures was 28–30 ° C.Chromosome analysis was performed from the actively pro-liferating cells, which revealed 5 pairs of macrochromo-somes and 15 pairs of microchromosomes in Natrix piscator, and 15 pairs of only macrochromosomes in Crocodylus palus-tris . (GATA) n repeats are well known to be associated with sex chromosomes. Fluorescence in situ hybridization performed with (GATA) 10 repeats delineated the W chromosome in the cells of Natrix piscator which has so far not been reported. This cell culture method has presently only been applied to water snakes and crocodile embryos in the current study, but it will be employed in other reptilian species and could go a long way to being a sustainable source of primary cells. This would eventually serve as an important tool for molecular studies in reptiles and other species in the future.
Copyright © 2010 S. Karger AG, Basel
Key Words Fluorescence in situ hybridization � (GATA) 10 � Indian mugger � Primary cultures � W chromosome � Water snake
Abstract Sex determination among reptiles has continued to draw the attention of geneticists and the mechanisms involved have been extensively studied and documented in the past 3 de-cades. The setting up of primary cell lines of reptilian tissues is an important tool in the present study which is a unique aspect not applied in earlier studies. Establishing the cell lines from various species of reptiles would help in our un-derstanding of the mechanisms of evolution and differentia-tion of sex chromosomes. Therefore, in the present study, we have established for the first time primary cell cultures from Indian water snake (Natrix piscator) and Indian mugger (Cro-codylus palustris) embryos. In the preliminary growth stage, 2 types of cells, fibroblast- and epithelial-like, were found to be attached and proliferating in vitro. These fibroblast-like cell cultures were later overtaken by epithelial cells. The cell lines were grown in minimal essential medium supplement-
Published online: March 25, 2010
Dr. Lalji Singh Centre for Cellular and Molecular Biology Uppal Road, Hyderabad 500 007 (India) Tel. +91 40 2716 0789, Fax +91 40 2716 0252E-Mail lalji @ ccmb.res.in
© 2010 S. Karger AG, Basel1424–8581/09/1274–0287$26.00/0
Accessible online at:www.karger.com/cgr
Dow
nloa
ded
by:
Uni
v.-u
nd L
ande
sbib
lioth
ek S
achs
en-A
nhal
t
14
1.48
.162
.113
- 3
/28/
2014
3:2
9:49
PM

Rao et al. Cytogenet Genome Res 2009;127:287–296288
Reptiles display an unusual variability in sex chromo-some constitution and sex determination systems [Mar-shall Graves and Shetty, 2001; Miller et al., 2004; Quinn et al., 2009] having both male and female heterogametic genotypic sex determination (GSD). Among reptiles, tur-tles, crocodiles and some lizards exhibit sexual differen-tiation of gonads that is sensitive to temperature (TSD) during a critical period of embryonic development and which is most evident during egg incubation rather than at conception [Bull, 1983; Pieau et al., 2001; Valenzuela and Lance, 2004]. Indian mugger is one of such species used in the present study. However, this mechanism is not observed in snakes [Richard-Mercier et al., 1995]. Thus the reptiles of the TSD group do not exhibit heteromor-phic sex chromosomes and have no karyotypic differenc-es between males and females [Ferguson and Joanen, 1982; Head et al., 1987; Lang and Andrews, 1994; Creeet al. 1995; Ciofi and Swingland, 1997; Valenzuela and Lance, 2004]. However, recently Quinn et al. [2009] re-ported isolation of Y-chromosome DNA sequence from a lizard (Bassiana duperreyi) having TSD. They used am-plified fragment-length polymorphism PCR and con-nected that amplified sequence to a single locus assay for chromosomal sex. Some lizards and some turtles show TSD and GSD systems with both male and female hetero-gametes [Olmo and Signorino, 2005].
Snakes have female heterogamety with ZZ - /ZW U chromosome composition [Beçak et al., 1964; Beçak and Beçak, 1969; Singh, 1972]. Singh et al. [1980] isolated W chromosome-specific satellite DNA Bkm (banded krait minor satellite DNA). It is highly conserved and can iden-tify the W chromosome in all species of snakes in which the W chromosome has undergone heterochromatiniza-tion. In the primitive family of snakes such as Python, Boa, etc., the Z and W chromosomes are homomorphic and W does not show heterochromatinization. This rais-es fundamental questions: How and when did the diver-gence leading to functional specialization of the W chro-mosome originate? How did it progress? And why did it skip some species? It is possible to answer these questions by making cell lines of female snakes representing the primitive, intermediate and highly evolved states of dif-ferentiation of sex chromosomes, sorting out their re-spective W chromosomes by fluorescence-activated cell sorter (FACS), making chromosome-specific libraries and by completely sequencing the W chromosome of the respective species. The present study is a big step forward in this direction. Indian mugger, which has temperature-dependent sex determination and absence of morpholog-ically differentiated sex chromosomes, has been used to
obtain insight into the molecular mechanisms involved in TSD.
Earlier chromosomes from lymphocytes of Natrix pis-cator were reported by Singh et al. [1968]. The current study attempted to establish for the first time primary cultures from the embryos of water snake (Natrix pis-cator) and Indian mugger (Crocodylus palustris), which have not been reported before. The methodology was de-veloped with an objective to identify various cell types in the growing cultures of these species. Subsequently, chro-mosomes were made from these proliferating cells and further characterization of the W chromosome was per-formed using (GATA) 10 repeats with the fluorescence in situ hybridization (FISH) technique.
The application of cell culture in the study of reptilian species assumes significance in view of the fact that the cell types are endowed with unique abilities of extensive multipotency and morphogenetic potential. They display high plasticity of shapes and structures, rate of cell re-placement and proliferation rate. Thus they could serve as potentially rich sources of cell and tissue types.
Materials and Methods
Dissecting Natrix piscator and Crocodylus palustris Embryos A total of 9 water snake embryos of approximately 2–3 weeks
gestation and 5 crocodile embryos (25th tail somites) were dis-sected. In the case of water snakes, the embryos were too small and were dissected into 3 sections as head, mid and tail. Thus, in all there were 27 sections from 9 embryos that were set for pri-mary cultures. From the crocodile embryos, heart, pancreas, go-nads, brain and kidney organs were dissected and subsequently set for tissue specific cultures.
Establishment of Primary Cultures from Natrix piscator and Crocodylus palustris In the case of Natrix piscator embryonic cell cultures, each of
the head, mid and tail sections were gently cut into less than 1-mm fragments. These fragments were dissociated with Trypsin-EDTA for about 20 min at room temperature. The fragments were spun and subsequently seeded with DMEM supplemented with 20% fetal bovine serum and antibiotics in separate flasks for head, mid and tail sections. Hence, in all there were 27 flasks seeded and kept at 28 ° C with 5% CO 2 atmosphere. After adhesion (1–2 days after seeding) fresh medium was added to these flasks. Similarly, from Crocodylus palustris embryos the dissected organs that in-clude heart, pancreas, gonads, brain and kidney were cut into small fragments followed by treatment with Trypsin-EDTA. These organ-specific fragments were further seeded in individu-al flasks at 30 ° C with CO 2 atmosphere. The growing cultures from both snake and crocodile embryos were further microscop-ically monitored for various types of dividing cells.
Dow
nloa
ded
by:
Uni
v.-u
nd L
ande
sbib
lioth
ek S
achs
en-A
nhal
t
14
1.48
.162
.113
- 3
/28/
2014
3:2
9:49
PM

Cytogenet Genome Res 2009;127:287–296 289Characterization of W Chromosome from Indian Water Snake Embryos
Chromosome Preparations Approximately 7 h after subculturing the 60–80% confluent
cultures from Natrix piscator and Crocodylus palustris embryos, the dividing cells were treated for 4–6 h with colcemid. Metaphase chromosomes were prepared following standard procedures [Schmid et al., 1989]. Conventional karytoype analyses in cells from Natrix piscator embryos (for Z and W chromosome identi-fication) and for cells from Crocodylus palustris embryos were performed on Giemsa-stained metaphase spreads. Constitutive heterochromatin was visualized by C-banding using the standard BSG (barium hydroxide, saline, Giemsa) technique [Sumner, 1972] with slight modification.
Fluorescence in situ Hybridization Single-color FISH was performed on fixed chromosomes from
Natrix piscator embryos using the (GATA) 10 clone. (GATA) 10 was labeled with digoxigenin by nick translation according to the manufacturer’s instructions (Roche Molecular Biochemicals). FISH was performed according to Cross et al. [2005]. For digoxi-genin-labeled probes, hybridization sites were detected using mouse anti-digoxigenin, rabbit anti-mouse FITC (fluorescein iso-thiocyanate) and goat anti-rabbit FITC immunological reagents (Roche Diagnostics). The chromosomes were counterstained with DAPI (4�,6-diamidino-2-phenylindole) (Vector Labs) in Vectashield antifade. Images were recorded using a Zeiss Axio-skop fluorescence microscope equipped with a digital camera.
Results
Morphology of the Cells Derived from Natrix piscator and Crocodylus palustris Embryos The head, mid and tail fragments from 9 Natrix pisca-
tor embryos started growing with a diversity of cell types. Of all the cell cultures set from the 9 embryos, 2 did not survive because of contamination. The cells tend to form clumps in 2–3 days after setting of the initial cultures. Af-ter 3 passages of the cultures, 4 types of cells were noted to be predominant, i.e. filamentous fibroblast-like cells, epithelial cells ( fig. 1 a, b), neuronal cells with dendrite ap-pearance ( fig. 2 a, b) and beating cells of heart ( fig. 2 c, d).
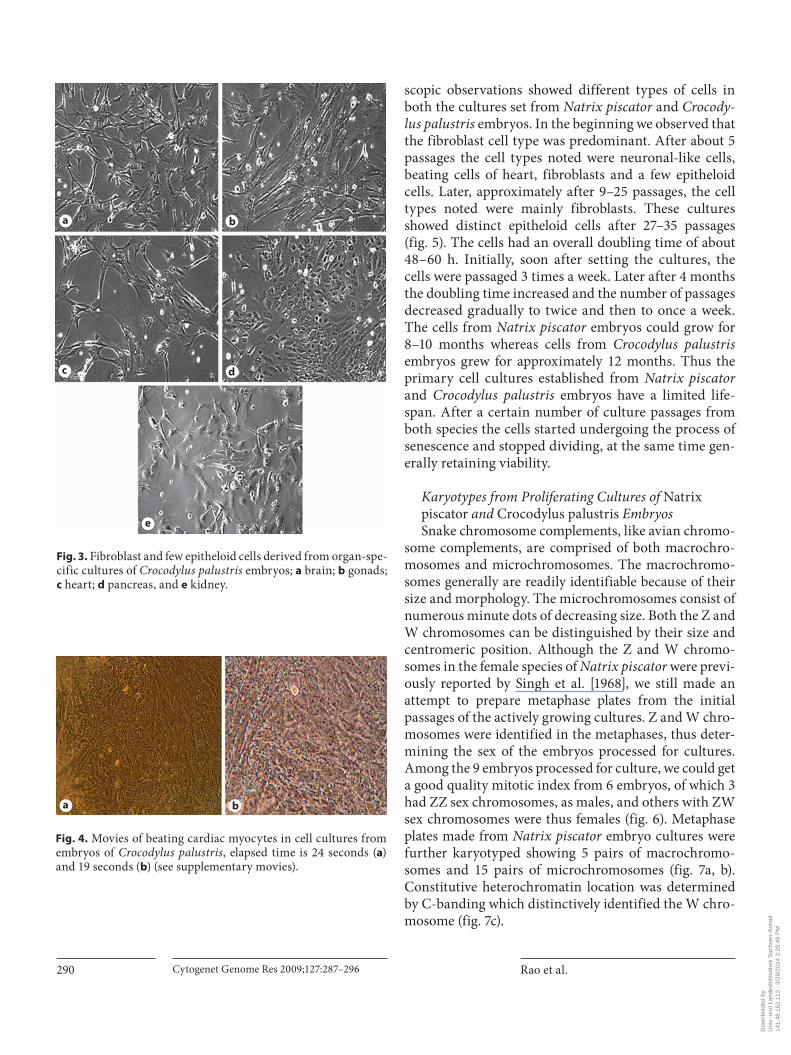
Similarly, the cells migrating from the heart, pancreas, gonads, brain and kidney cultures of Crocodylus palustris covered the bottom of the flask in 5–8 days mainly as fibroblast-like cells and with a few epitheloid cells ( fig. 3 a–e). Noted among them were slow growing pancreatic cells which covered the bottom of the flask within 10 days. Beating cells of cardiac myocytes were also noted in the cultures that were set from the dissected hearts of the embryos ( fig. 4 a, b).
The functional characteristics of cultured cells are sig-nificant from both their origin (brain, heart, etc.) and how well they adapt to the culture conditions. Biochemi-cal markers can be used in the future to determine if cells
are still carrying on specialized functions that they per-formed in vivo (e.g. brain cells). Morphology or ultra-structural markers can also be examined (e.g. beating heart cells). Frequently, these characteristics are either lost or changed as a result of the cells being placed in an artificial environment.
Growth Characteristics of the Primary Cultures from Natrix piscator and Crocodylus palustris Embryos Growth was assessed in the proliferating primary cul-
tures from initial stages after setting the cultures. Micro-
a b
Fig. 1. Photomicrographs of the cell types obtained in primary cultures of Natrix piscator embryos. a Fibroblast-like cells. b Epi-thelial cells.
a b
c d
Fig. 2. a Filamentous neuronal-like cells obtained in primary cul-tures from embryos of Natrix piscator . b Greater magnification rep-resenting the appearance of dendrite-like cells. c , d Movies of beat-ing cardiac myocytes in cultures from embryos of Natrix piscator; elapsed time is 32 seconds, and 19 seconds, respectively (for supple-mentary movies, see www.karger.com/doi/ 10.1159/ 000304046).
Dow
nloa
ded
by:
Uni
v.-u
nd L
ande
sbib
lioth
ek S
achs
en-A
nhal
t
14
1.48
.162
.113
- 3
/28/
2014
3:2
9:49
PM
Rao et al. Cytogenet Genome Res 2009;127:287–296290
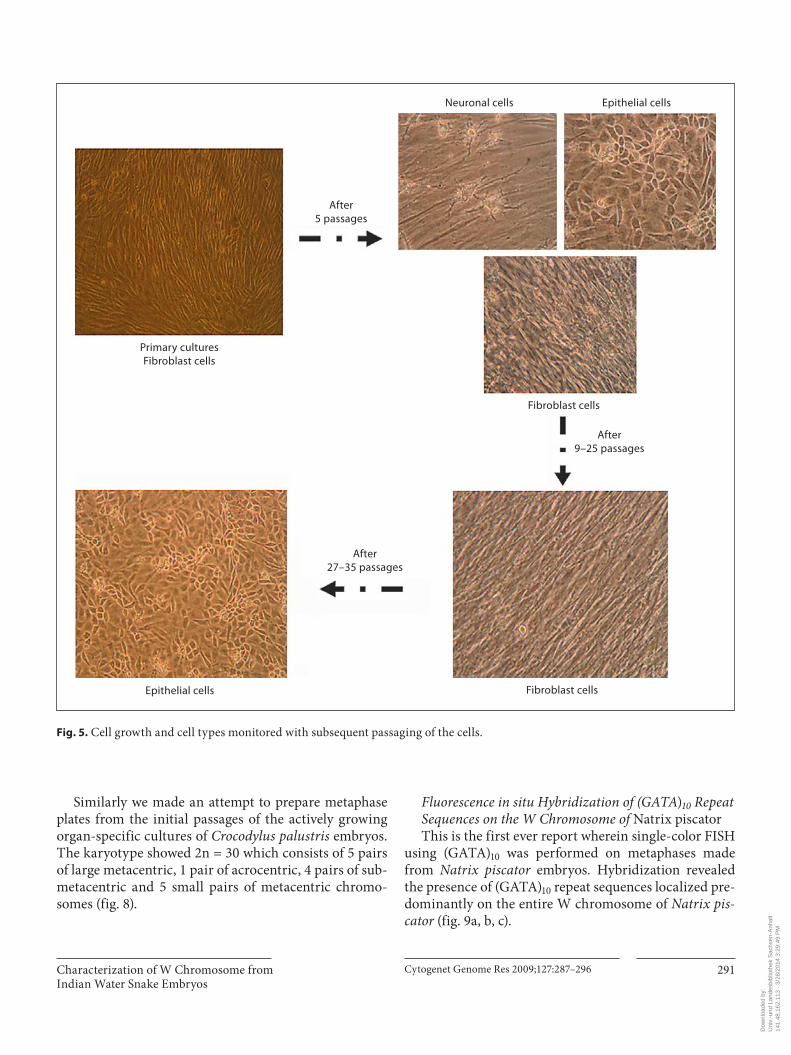
scopic observations showed different types of cells in both the cultures set from Natrix piscator and Crocody-lus palustris embryos. In the beginning we observed that the fibroblast cell type was predominant. After about 5 passages the cell types noted were neuronal-like cells, beating cells of heart, fibroblasts and a few epitheloid cells. Later, approximately after 9–25 passages, the cell types noted were mainly fibroblasts. These cultures showed distinct epitheloid cells after 27–35 passages ( fig. 5 ). The cells had an overall doubling time of about 48–60 h. Initially, soon after setting the cultures, the cells were passaged 3 times a week. Later after 4 months the doubling time increased and the number of passages decreased gradually to twice and then to once a week. The cells from Natrix piscator embryos could grow for 8–10 months whereas cells from Crocodylus palustris embryos grew for approximately 12 months. Thus the primary cell cultures established from Natrix piscator and Crocodylus palustris embryos have a limited life-span. After a certain number of culture passages from both species the cells started undergoing the process of senescence and stopped dividing, at the same time gen-erally retaining viability.
Karyotypes from Proliferating Cultures of Natrix piscator and Crocodylus palustris Embryos Snake chromosome complements, like avian chromo-
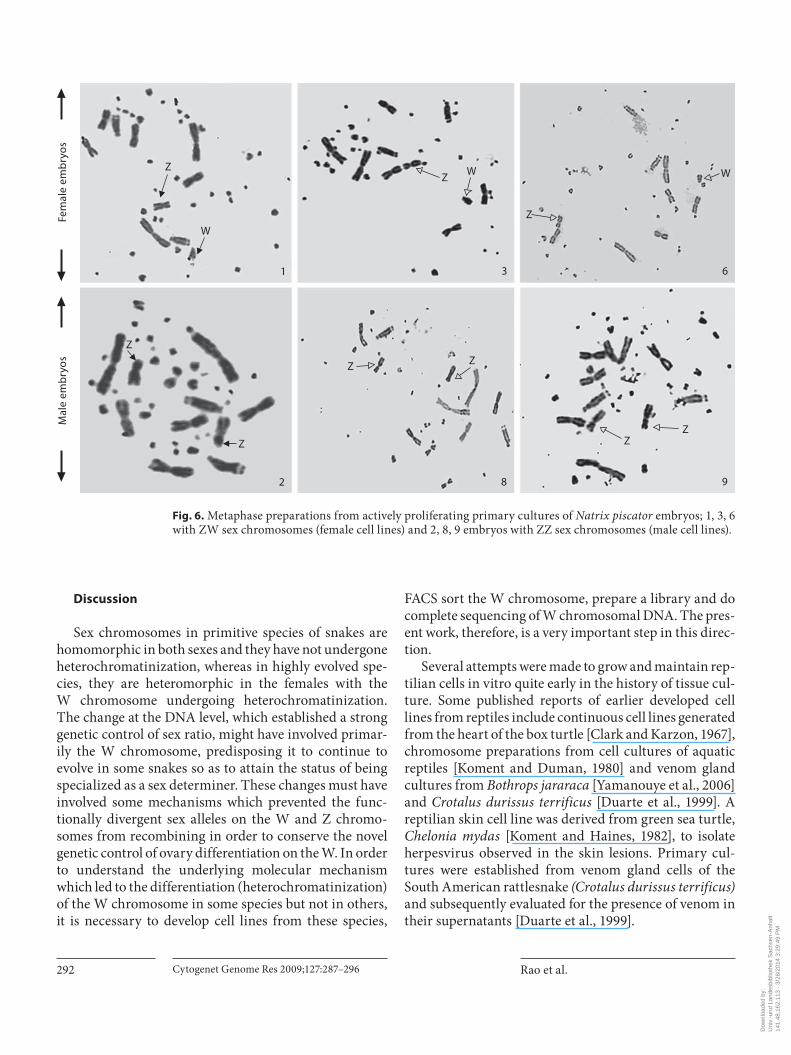
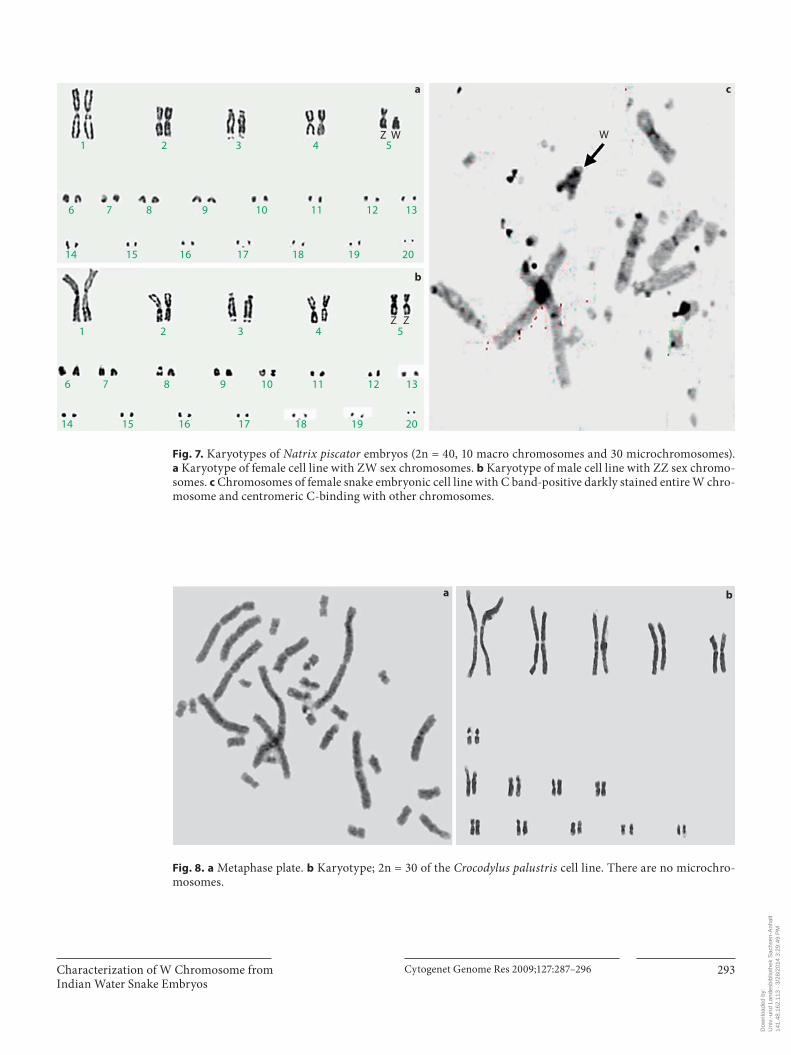
some complements, are comprised of both macrochro-mosomes and microchromosomes. The macrochromo-somes generally are readily identifiable because of their size and morphology. The microchromosomes consist of numerous minute dots of decreasing size. Both the Z and W chromosomes can be distinguished by their size and centromeric position. Although the Z and W chromo-somes in the female species of Natrix piscator were previ-ously reported by Singh et al. [1968], we still made anattempt to prepare metaphase plates from the initialpassages of the actively growing cultures. Z and W chro-mosomes were identified in the metaphases, thus deter-mining the sex of the embryos processed for cultures. Among the 9 embryos processed for culture, we could get a good quality mitotic index from 6 embryos, of which 3 had ZZ sex chromosomes, as males, and others with ZW sex chromosomes were thus females ( fig. 6 ). Metaphase plates made from Natrix piscator embryo cultures were further karyotyped showing 5 pairs of macrochromo-somes and 15 pairs of microchromosomes ( fig. 7 a, b). Constitutive heterochromatin location was determined by C-banding which distinctively identified the W chro-mosome ( fig. 7 c).
a b
c d
e
Fig. 3. Fibroblast and few epitheloid cells derived from organ-spe-cific cultures of Crocodylus palustris embryos; a brain; b gonads; c heart; d pancreas, and e kidney.
a b
Fig. 4. Movies of beating cardiac myocytes in cell cultures from embryos of Crocodylus palustris , elapsed time is 24 seconds ( a ) and 19 seconds ( b ) (see supplementary movies).
Dow
nloa
ded
by:
Uni
v.-u
nd L
ande
sbib
lioth
ek S
achs
en-A
nhal
t
14
1.48
.162
.113
- 3
/28/
2014
3:2
9:49
PM
Cytogenet Genome Res 2009;127:287–296 291Characterization of W Chromosome from Indian Water Snake Embryos
Similarly we made an attempt to prepare metaphase plates from the initial passages of the actively growing organ-specific cultures of Crocodylus palustris embryos. The karyotype showed 2n = 30 which consists of 5 pairs of large metacentric, 1 pair of acrocentric, 4 pairs of sub-metacentric and 5 small pairs of metacentric chromo-somes ( fig. 8 ).
Fluorescence in situ Hybridization of (GATA) 10 Repeat Sequences on the W Chromosome of Natrix piscator This is the first ever report wherein single-color FISH
using (GATA) 10 was performed on metaphases made from Natrix piscator embryos . Hybridization revealed the presence of (GATA) 10 repeat sequences localized pre-dominantly on the entire W chromosome of Natrix pis-cator ( fig. 9 a, b, c).
Primary culturesFibroblast cells
Epithelial cells
After5 passages
After27–35 passages
After9–25 passages
Neuronal cells Epithelial cells
Fibroblast cells
Fibroblast cells
Fig. 5. Cell growth and cell types monitored with subsequent passaging of the cells.
Dow
nloa
ded
by:
Uni
v.-u
nd L
ande
sbib
lioth
ek S
achs
en-A
nhal
t
14
1.48
.162
.113
- 3
/28/
2014
3:2
9:49
PM
Rao et al. Cytogenet Genome Res 2009;127:287–296292
Discussion
Sex chromosomes in primitive species of snakes are homomorphic in both sexes and they have not undergone heterochromatinization, whereas in highly evolved spe-cies, they are heteromorphic in the females with theW chromosome undergoing heterochromatinization. The change at the DNA level, which established a strong genetic control of sex ratio, might have involved primar-ily the W chromosome, predisposing it to continue to evolve in some snakes so as to attain the status of being specialized as a sex determiner. These changes must have involved some mechanisms which prevented the func-tionally divergent sex alleles on the W and Z chromo-somes from recombining in order to conserve the novel genetic control of ovary differentiation on the W. In order to understand the underlying molecular mechanism which led to the differentiation (heterochromatinization) of the W chromosome in some species but not in others, it is necessary to develop cell lines from these species,
FACS sort the W chromosome, prepare a library and do complete sequencing of W chromosomal DNA. The pres-ent work, therefore, is a very important step in this direc-tion.
Several attempts were made to grow and maintain rep-tilian cells in vitro quite early in the history of tissue cul-ture. Some published reports of earlier developed cell lines from reptiles include continuous cell lines generated from the heart of the box turtle [Clark and Karzon, 1967], chromosome preparations from cell cultures of aquatic reptiles [Koment and Duman, 1980] and venom gland cultures from Bothrops jararaca [Yamanouye et al., 2006] and Crotalus durissus terrificus [Duarte et al., 1999]. A reptilian skin cell line was derived from green sea turtle, Chelonia mydas [Koment and Haines, 1982], to isolate herpesvirus observed in the skin lesions. Primary cul-tures were established from venom gland cells of the South American rattlesnake ( Crotalus durissus terrificus ) and subsequently evaluated for the presence of venom in their supernatants [Duarte et al., 1999].
Z
W
1
Z W
3
Z
W
6
Z
Z
2
Z Z
8
ZZ
9
Mal
e em
bryo
sFe
mal
e em
bryo
s
Fig. 6. Metaphase preparations from actively proliferating primary cultures of Natrix piscator embryos; 1, 3, 6 with ZW sex chromosomes (female cell lines) and 2, 8, 9 embryos with ZZ sex chromosomes (male cell lines).
Dow
nloa
ded
by:
Uni
v.-u
nd L
ande
sbib
lioth
ek S
achs
en-A
nhal
t
14
1.48
.162
.113
- 3
/28/
2014
3:2
9:49
PM
Cytogenet Genome Res 2009;127:287–296 293Characterization of W Chromosome from Indian Water Snake Embryos
1
6
14 15 16 17 18 19 20
14 15 16 17 18 19 20
7 8 9 10 11 12 13
6 7 8 9 10 11 12 13
2 3 4 5
1 2 3 4 5
WZ W
Z Z
a c
b
Fig. 7. Karyotypes of Natrix piscator embryos (2n = 40, 10 macro chromosomes and 30 microchromosomes). a Karyotype of female cell line with ZW sex chromosomes. b Karyotype of male cell line with ZZ sex chromo-somes. c Chromosomes of female snake embryonic cell line with C band-positive darkly stained entire W chro-mosome and centromeric C-binding with other chromosomes.
a b
Fig. 8. a Metaphase plate. b Karyotype; 2n = 30 of the Crocodylus palustris cell line. There are no microchro-mosomes.
Dow
nloa
ded
by:
Uni
v.-u
nd L
ande
sbib
lioth
ek S
achs
en-A
nhal
t
14
1.48
.162
.113
- 3
/28/
2014
3:2
9:49
PM

Rao et al. Cytogenet Genome Res 2009;127:287–296294
In the present study, we established primary cultures from 2 Indian reptilian species, namely embryos of water snake ( Natrix piscator ) and Indian mugger (Crocodylus palustris) which have not been reported before. All the primary cultures of Natrix piscator and Crocodylus palus-tris created from the embryos were successful in the first attempt. We also attempted to establish primary cultures from the adult animals of these species, but these cell lines could grow only for a limited period of 2 months with a very slow proliferation rate as compared to those derived from the embryonic lines. The enzyme digestion procedure was deployed and the established primary cul-tures showed epithelial, fibroblast, dendritic-neuronal and beating heart cells. This entire process lasted for ap-proximately 10 months. The cell lines established from these species could have enormous utility in many fields of study, particularly phenotype and functional charac-teristics studies.
It has been a well-documented fact that sex chromo-somes in primitive species of snakes have not undergone heterochromatinization and thus are homomorphic in both sexes. In highly evolved species of snakes, the fe-males are heteromorphic with a W chromosome that un-dergoes heterochromatinization. Metaphase chromo-somes were made from embryonic primary proliferating cell lines established by us from Natrix piscator and Cro-codylus palustris . These cell lines were developed from 6 Natrix piscator embryos which showed 3 embryos as males with a ZZ chromosome complement and 3 embry-os as females with a ZW chromosome complement. The C band-positive W chromosome shows heterochromati-nization which is very prominently seen in the metaphas-es made from female embryos. In the light of the above it is evident that the W chromosome of female Natrix pisca-tor may have evolved from primitive species of snakes.
The Bkm (Banded krait minor) satellite DNA is a sim-ple sequence repeat, which is highly conserved in eukary-otes and predominantly consists of tetranucleotide repeat GATA [Ray-Chaudhuri et al., 1971]. The Bkm probe was used to identify the W chromosome of different species of snakes using Southern blots as well as in situ hybridiza-tion. This technique, however, did not allow the identi-fication of W chromosomes in primitive species since they have undifferentiated sex chromosomes [Singh et al., 1980]. The GATA repeat studies have initially been cru-cial in studies of the evolution and differentiation of sex chromosomes in snakes. Later GATA repeat studies were documented in mouse wherein the Bkm sequences are mainly hybridized in the p arm of the Y chromosome [Singh and Jones, 1982]. Thereafter the GATA repeat studies have branched out, determining the sex chromo-somes of various other eukaryotes [Subramanian et al., 2003].
In the present study we performed single-color FISH on female metaphases of Natrix piscator embryos which showed the presence of GATA 10 repeats all along the length of the W chromosome. The GATA sequences which form clusters on the W chromosome could have played a significant role during the course of evolution and also in the evolution and differentiation of the sex chromosomes. The W chromosome remains condensed at the metaphase stage whereas at the interphase stage it forms a W-chromatin body. We are presently working on procedures to immortalize the cell lines with the objec-tive to expand the lifespan of the cells to grow them in large quantities. In the future, advanced techniques like FACS of these cells would be employed to enable us to flow-sort W and Z chromosomes and subsequently gen-erate W and Z chromosome libraries. Thus using con-temporary genomic technologies, mainly FACS and
a cb
Fig. 9. FISH using (GATA) 10 probe paint-ing the W chromosome from cell cultures of Natrix piscator embryos. a Inverted im-age of female metaphase plate. b W chro-mosome painted with (GATA) 10 probe. c Painting of W chromosome in interphase nuclei from female embryos with (GATA) 10 probe.
Dow
nloa
ded
by:
Uni
v.-u
nd L
ande
sbib
lioth
ek S
achs
en-A
nhal
t
14
1.48
.162
.113
- 3
/28/
2014
3:2
9:49
PM

Cytogenet Genome Res 2009;127:287–296 295Characterization of W Chromosome from Indian Water Snake Embryos
high-throughput genomic sequencing, may well help to unravel the mystery related to sex chromosome evolu-tion, the early events in sex determination and dosage compensation. This information, it is felt, would contrib-ute extensively towards a general understanding of the genetic control of development in reptiles.
In addition to the Natrix piscator cell lines, we have also established, for the first time, organ-specific primary cell cultures from embryos of the Indian mugger (Croco-dylus palustris) . Crocodylus palustris possesses a TSD mechanism of sex determination, in which the incuba-tion temperature of the egg determines the sex of the hatchling. It has been suggested that changes in tempera-ture affect the expression of sex-determining genes in the embryo during gonadal differentiation [Deeming and Ferguson, 1988]. This hypothesis is supported by the ob-servation that DMRT1 expression in the genital ridge of turtle embryos is increased at the low temperatures that promote male rather than female differentiation [Ket-tlewell et al., 2000]. DMRT1 maps to chromosome 6 in2 species, red-eared slider (Trachemys scripta) and Nile crocodile (Crocodylus niloticus) [Kasai and Ferguson-Smith, unpublished] and also to chromosome 6 in the Chinese soft-shelled turtle [Matsuda et al., 2005]. Chro-mosome 6 in these reptiles thus seems to represent the ancestral pair of chromosomes from which the ZZ/ZW pair of chromosomes in birds has evolved. Our previous study showed isolation of the DMRT1 homologue from Crocodylus palustris , along with its partial genomic locus. DMRT1 exhibits transcriptional diversity as seen in GSD species [Anand et al., 2008]. The karyotype analysis from the early passages of actively proliferating cultures showed only 30 homomorphic chromosomes without any distinguishable sex chromosome pair as reported earlier [Singh and Ray-Chaudhuri, 1973]. We are current-ly engaged in the process of mapping DMRT1 on chromo-some preparations made from organ-specific cell lines established from Crocodylus palustris embryos. The pres-ent study’s techniques could be employed in the future to map other genes as well that have an essential role in the TSD mechanism of sex determination in Crocodylus palustris .
Turtles, crocodiles and birds are known to have high-ly conserved karyotypes, and in their comparison with snakes, the snakes show considerable rearrangements. The most significant differences are seen in the snake Z and W sex chromosomes which are not homologous to the avian sex chromosomes and must have evolved inde-pendently from a different pair of ancestral autosomes. The orthologues of Z genes in the chicken, including
DMRT1, map to chromosome 2 in both the Japanese 4-striped rat snake, Elaphe quadrivirgata [Matsuda et al., 2005], and the Australian tiger snake, Notechis scutatus [Kasai and Ferguson-Smith, unpublished]. These map-ping studies indicate that the snake Z shares homology with chicken chromosome 2 and human chromosomes 7 and 10. The primary sex-determining switch and the role of DMRT1 in sex differentiation in snakes are not fully understood. These genomic differences support the molecular phylogenetic data which suggest that snakes are more distantly related to birds and to other reptiles than was previously thought [Kumar and Hedges, 1998]. In the light of this information the cell lines derived from the Natrix piscator embryos could serve to unravel the evolutionary aspects of Z and W chromosomes and sub-sequently facilitate finding the homology of the respec-tive sex chromosome-specific sequences with chicken and human chromosomes.
It may thus be concluded that this study has paved a way to establish primary cell lines from 2 reptilian spe-cies, Natrix piscator and Crocodylus palustris embryos. The derived primary cell lines were useful for studying chromosomes in both species. The characterization of the W chromosome was further performed by hybridiza-tion with GATA 10 repeats in cells from Natrix piscator which helps us to understand the evolution of Natrix pis-cator from primitive species of snakes.
This study could also open a window to the vast po-tential of establishing primary cell lines from the embry-os of different species of snakes representing various states of differentiation of sex chromosomes [Singh et al., 1976, 1980]. This would make FACS sorting of the W chromosomes at different states of differentiation possi-ble. Construction of their respective libraries and com-plete sequencing would unravel the underlying molecu-lar mechanisms involved in evolution and differentiation of the W chromosome which is mysterious so far. This would also help in our understanding of evolution of sex chromosomes in other species.
Acknowledgment
The study was supported by a grant from the Council of Sci-entific and Industrial Research (CSIR), New Delhi, Government of India.
Dow
nloa
ded
by:
Uni
v.-u
nd L
ande
sbib
lioth
ek S
achs
en-A
nhal
t
14
1.48
.162
.113
- 3
/28/
2014
3:2
9:49
PM

Rao et al. Cytogenet Genome Res 2009;127:287–296296
References
Anand A, Patel M, Lalremruata A, Singh AP, Agrawal R, et al: Multiple alternative splicing of Dmrt1 during gonadogenesis in Indian mugger, a species exhibiting temperature-dependent sex determination. Gene 425: 56–63 (2008).
Beçak W, Beçak ML: Cytotaxonomy and chro-mosomal evolution in Serpentes. Cytogenet-ics 8: 247–262 (1969).
Beçak W, Beçak ML, Nazareth HR, Ohno S: Close karyological kinship between the rep-tilian suborder Serpents and the class Aves. Chromosoma 15: 606–617 (1964).
Bull JJ: Evolution of Sex Determining Mecha-nisms. (Benjamin-Cummings, Menlo Park 1983).
Ciofi C, Swingland IR: Environmental sex deter-mination in reptiles. Appl Anim Behav Sci 51: 251–265 (1997).
Clark HF, Karzon DT: Terrapene heart (TH-1), a continuous cell line from the heart of the box turtle Terrapene carolina . Exp Cell Res 28: 263–268 (1967).
Cree A, Thompson MB, Daugherty CH: Tuatara sex determination. Nature 375: 543 (1995).
Cross, I, Díaz E, Sánchez I, Rebordinos L: Mo-lecular and cytogenetic characterization of Crassostrea angulata chromosomes. Aqua-culture 247: 135–144 (2005).
Deeming DC, Ferguson MW: Environmental regulation of sex determination in reptiles. Philos Trans R Soc Lond B Biol Sci 322: 19–39 (1988).
Duarte MM, Montes de Oca H, Diniz CR, Fortes-Dias CL: Primary culture of venom gland cells from the South American rattlesnake ( Crotalus durissus terrificus ). Toxicon 37: 1673–1682 (1999).
Ferguson MW, Joanen T: Temperature of egg in-cubation determines sex in Alligator missis-sippiensis . Nature 296: 850–853 (1982).
Head G, May RM, Pendleton L: Environmental determination of sex in the reptiles. Nature 329: 198–199 (1987).
Kettlewell JR, Raymond CS, Zarkower D: Tem-perature-dependent expression of turtle Dmrt1 prior to sexual differentiation. Gen-esis 26: 174–178 (2000).
Koment RW, Duman RG: Preparation of meta-phase chromosomes in cell cultures derived from an aquatic reptile. J Tissue Cult Meth 6: 25–28 (1980).
Koment RW, Haines H: Characterization of a reptilian epithelioid skin cell line derived from the green sea turtle, Chelonia mydas . In Vitro18: 227–232 (1982).
Kumar S, Hedges SB: A molecular timescale for vertebrate evolution. Nature 392: 917–920 (1998).
Lang JW, Andrews HV: Temperature-dependent sex determination in crocodilians. J Exp Zoo 270: 28–44 (1994).
Marshall Graves JA, Shetty S: Sex from W to Z: evolution of vertebrate sex chromosomes and sex determining genes. J Exp Zool 290: 449–462 (2001).
Matsuda Y, Nishida-Umehara C, Tarui H, Ku-roiwa A, Yamada K, et al: Highly conserved linkage homology between birds and turtles: birds and turtle chromosomes are precise counterparts of each other. Chromosome Res 13: 601–615 (2005).
Miller D, Summers J, Silber S: Environmental versus genetic sex determination: a possible factor in dinosaur extinction? Fertil Steril 81: 954–964 (2004).
Olmo E, Signorino G: Chromorep: a reptile chromosomes database. (http://193.206.118.100/professori/chromorep.pdf) (2005).
Pieau C, Dorizzi M, Richard-Mercier N: Tem-perature-dependent sex determination and gonadal differentiation in reptiles. EXS 91: 117–141(2001).
Quinn AE, Radder RS, Sarre SD, Georges A, Ezaz T: Isolation and development of a mo-lecular sex marker for Bassiana duperreyi , a lizard with XX/XY chromosomes and tem-perature-induced sex reversal. Mol Genet Genomics 281: 665–672 (2009).
Ray-Chaudhuri SP, Singh L, Sharma T: Evolu-tion of sex chromosomes and formation ofW chromatin in snakes. Chromosoma 33: 239–251 (1971).
Richard-Mercier N, Dorizzi M, Desvages G, Gi-rondot M, Pieau C: Endocrine sex reversal of gonads by the aromatase inhibitor Letrozole (CGS 20267) in Emys orbicularis , a turtle with temperature-dependent sex determina-tion. Gen Comp Endocrinol 100: 314–326 (1995).
Schmid M, Enderle E, Schindler D, Schempp W: Chromosome banding and DNA replication patterns in bird karyotypes. Cytogenet Cell Genet 52: 139–146 (1989).
Singh L: Evolution of karyotypes of snakes. Chromosoma 38: 185–236 (1972).
Singh L, Jones KW: Sex reversal in mouse ( Mus musculus ) is caused by a recurrent non-re-ciprocal crossover involving the X and anaberrant Y chromosome. Cell 28: 205–216 (1982).
Singh L, Ray-Chaudhuri SP: DNA replication pattern in the chromosomes of Crocodylus palustris (Lesson). Nucleus 16:33–37 (1973).
Singh L, Sharma T, Ray-Chaudhuri SP: W chro-mosome in the Indian water snake (Check-ered keel back) Natrix piscator (Colubridae). Experientia 24: 79–80 (1968).
Singh L, Purdom IF, Jones KW: Satellite DNA and evolution of sex chromosomes. Chro-mosoma 59: 43–62 (1976).
Singh L, Purdom IF, Jones KW: Sex chromosome associated satellite DNA: evolution and con-servation. Chromosoma 79: 137–157 (1980).
Subramanian S, Mishra RK, Singh L: Genome-wide analysis of Bkm sequences (GATA re-peats): predominant association with sex chromosomes and potential role in higher order chromatin organization and function. Bioinformatics 19: 681–685 (2003).
Sumner AT: A simple technique for demonstrat-ing centromeric heterochromatin. Exp Cell Res 75: 304–306 (1972).
Valenzuela N, Lance VA: Temperature Depen-dent Sex Determination in Vertebrates. (Smithsonian Books, Washington DC 2004).
Yamanouye N, Kerchove CM, Moura-da-Silva AM, Carneiro SM, Markus RP: Long-term primary culture of secretory cells of Bothrops jararaca venom gland for venom production in vitro. Nat Protoc 1: 2763–2766 (2006).
Dow
nloa
ded
by:
Uni
v.-u
nd L
ande
sbib
lioth
ek S
achs
en-A
nhal
t
14
1.48
.162
.113
- 3
/28/
2014
3:2
9:49
PM
Copyright © 2022 FDOKUMEN




















