
Current perspectives on the darker beef often reported from extensively-managed cattle: Does...
22
Review article Current perspectives on the darker beef often reported from extensively-managed cattle: Does physical activity play a significant role? P.G. Dunne a, c, ⁎, F.J. Monahan b , A.P. Moloney a, c a Teagasc, Food Research Centre Ashtown, Ashtown, Dublin 15, Ireland b School of Agriculture, Food Science & Veterinary Medicine, University College Dublin, Dublin 4, Ireland c Teagasc, Animal & Grassland Research & Innovation Centre, Grange, Dunsany, Co. Meath, Ireland article info abstract Article history: Received 12 October 2010 Received in revised form 15 June 2011 Accepted 20 June 2011 Although frequently confounded by many factors, cattle raised outdoors on pasture are consistently reported to have darker muscle than cattle raised indoors. Exercise may play a key role. Colour differences apparently caused by nutrition are often confounded by environmental factors which may provide cattle with increased opportunities or requirements for activity. Furthermore, when slaughtered at similar ages and levels of fatness, diet composition (as opposed to energy level) does not appear to effect statistically significant variation in all muscle colour attributes in adult bovines or is at best inconsistent. The increased opportunity for pastured cattle to engage in activity, including grazing, rather than their grass diet per se may contribute to physiological and metabolic adaptation in muscle sufficient to alter qualitative and quantitative biochemical aspects of muscle metabolism, manifested as differences in post- mortem muscle colour. Concomitant changes in associated muscle fibre types and biochemical pathways usually occur, notably a shift towards oxidative metabolism. However, effects may depend on the age of the animal and duration, amount and intensity of activity and may be muscle-specific. Any effects of exercise on muscle characteristics, including colour, are likely to involve complex adaptive and interactive responses perhaps under all or some combination of genetic, nutritional and endocrinological control so that effects of dietary composition and energy, growth rate and growth path, amongst others, reflected in carcass and meat quality differences between extensively- and intensively-managed bovines, cannot be conveniently overlooked. © 2011 Elsevier B.V. All rights reserved. Keywords: Cattle Beef Exercise Meat Colour Review Contents 1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2 2. The basis of meat colour . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3 2.1. The phenomenon of meat colour and its measurement . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3 2.2. Muscle fibre types and meat colour . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4 3. Comparison of muscle colour of forage-finished and concentrate-finished cattle . . . . . . . . . . . . . . . . . . . . . . . . 4 3.1. Relevant comparisons: are they confounded by physical activity? . . . . . . . . . . . . . . . . . . . . . . . . . . . 4 3.2. Do dietary ingredients affect beef colour, i.e. is diet/nutrition always the obvious cause of differences in beef colour between pasture- and grain-fed cattle? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8 3.3. Could the method of colour evaluation affect colour differences between pasture/forage-fed and grain-fed cattle? . . . . 9 Livestock Science 142 (2011) 1–22 ⁎ Corresponding author at: Teagasc, Food Research Centre Ashtown, Ashtown, Dublin 15, Ireland. Tel.: + 353 1 8059500; fax: + 353 1 8059550. E-mail address: [email protected] (P.G. Dunne). 1871-1413/$ – see front matter © 2011 Elsevier B.V. All rights reserved. doi:10.1016/j.livsci.2011.06.018 Contents lists available at ScienceDirect Livestock Science journal homepage: www.elsevier.com/locate/livsci
Transcript of Current perspectives on the darker beef often reported from extensively-managed cattle: Does...

Livestock Science 142 (2011) 1–22
Contents lists available at ScienceDirect
Livestock Science
j ourna l homepage: www.e lsev ie r.com/ locate / l ivsc i
Review article
Current perspectives on the darker beef often reported fromextensively-managed cattle: Does physical activity play a significant role?
P.G. Dunne a,c,⁎, F.J. Monahan b, A.P. Moloney a,c
a Teagasc, Food Research Centre Ashtown, Ashtown, Dublin 15, Irelandb School of Agriculture, Food Science & Veterinary Medicine, University College Dublin, Dublin 4, Irelandc Teagasc, Animal & Grassland Research & Innovation Centre, Grange, Dunsany, Co. Meath, Ireland
a r t i c l e i n f o
⁎ Corresponding author at: Teagasc, Food ResearchE-mail address: [email protected] (P.G. Dunn
1871-1413/$ – see front matter © 2011 Elsevier B.V.doi:10.1016/j.livsci.2011.06.018
a b s t r a c t
Article history:Received 12 October 2010Received in revised form 15 June 2011Accepted 20 June 2011
Although frequently confounded by many factors, cattle raised outdoors on pasture areconsistently reported to have darker muscle than cattle raised indoors. Exercise may play a keyrole. Colour differences apparently caused by nutrition are often confounded by environmentalfactors which may provide cattle with increased opportunities or requirements for activity.Furthermore, when slaughtered at similar ages and levels of fatness, diet composition (asopposed to energy level) does not appear to effect statistically significant variation in all musclecolour attributes in adult bovines or is at best inconsistent. The increased opportunity forpastured cattle to engage in activity, including grazing, rather than their grass diet per se maycontribute to physiological and metabolic adaptation in muscle sufficient to alter qualitativeand quantitative biochemical aspects of muscle metabolism, manifested as differences in post-mortem muscle colour. Concomitant changes in associated muscle fibre types and biochemicalpathways usually occur, notably a shift towards oxidative metabolism. However, effects maydepend on the age of the animal and duration, amount and intensity of activity and may bemuscle-specific. Any effects of exercise on muscle characteristics, including colour, are likely toinvolve complex adaptive and interactive responses perhaps under all or some combination ofgenetic, nutritional and endocrinological control so that effects of dietary composition andenergy, growth rate and growth path, amongst others, reflected in carcass and meat qualitydifferences between extensively- and intensively-managed bovines, cannot be convenientlyoverlooked.
© 2011 Elsevier B.V. All rights reserved.
Keywords:CattleBeefExerciseMeatColourReview
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 22. The basis of meat colour . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3
2.1. The phenomenon of meat colour and its measurement . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32.2. Muscle fibre types and meat colour . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4
3. Comparison of muscle colour of forage-finished and concentrate-finished cattle . . . . . . . . . . . . . . . . . . . . . . . . 43.1. Relevant comparisons: are they confounded by physical activity? . . . . . . . . . . . . . . . . . . . . . . . . . . . 43.2. Do dietary ingredients affect beef colour, i.e. is diet/nutrition always the obvious cause of differences in beef colour
between pasture- and grain-fed cattle?. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 83.3. Could the method of colour evaluation affect colour differences between pasture/forage-fed and grain-fed cattle? . . . . 9
Centre Ashtown, Ashtown, Dublin 15, Ireland. Tel.: +353 1 8059500; fax: +353 1 8059550.e).
All rights reserved.

2 P.G. Dunne et al. / Livestock Science 142 (2011) 1–22
3.4. Age at slaughter can confound muscle colour comparisons based on nutrition . . . . . . . . . . . . . . . . . . . . . . 93.5. Confounding arising from differing nutritional planes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10
4. Effect of space allowance, confinement and accommodation type on muscle colour of cattle . . . . . . . . . . . . . . . . . 114.1. Grassland management and grazing systems . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 114.2. Spontaneous physical activity undertaken by grazing cattle . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 124.3. Measurement of physical activity engaged in by cattle . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 124.4. Energy expenditure by cattle . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 124.5. Effects on meat colour . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 134.6. Effects of accommodation type . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 14
5. Differential response of muscles to management practises under experimental conditions: effects on meat colour putativelydue to different activity levels . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 14
6. Relationship between muscle colour and associated biochemical characteristics: effects of exercise/activity on biochemicalcharacteristics and colour . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15
7. Effects of physical activity on muscle glycogen concentrations, pH and their potential effect on muscle colour . . . . . . . . . 168. Putative combined effect on muscle colour of increased pH and enhanced oxidative potential both caused by physical activity . 179. Summary and conclusions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 17Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18
1. Introduction
Colour, together with tenderness and flavour, is one of thecharacteristics that define sensory quality of beef. However,as is generally the case with food products, colour is often themost important of these characteristics at the point of sale,since it is the first quality characteristic which consumersexperience. Colour has a critical influence on beef purchasedecisions and thus is of fundamental importance to the beefindustry (Cornforth, 1994).
It is recognised that the practises involved in cattleproduction have a profound impact on carcass and meatquality, including meat colour, of beef cattle (Barker et al.,1995). These practises, such as the animal type chosen andmanagement practises undertaken may depend largely onclimatic, socio-economic, financial and regulatory factorsprevailing in a particular country or region (Keane andDrennan, 2007). In this regard, beef production ‘systems’constitute a composite of the bovine type chosen and the seriesof events and conditions which are followed from birth untilslaughter and hence, represent the combined and interactingeffects of genetic and environmental factors such as genotype,gender, age at slaughter and nutrition before slaughter(Moloney et al., 2001b) but perhaps also, and critically in thecontext of the present review, opportunities for and pre-dilections to physical activity by livestock under given environ-mental conditions.
Typically, management decisions dictate that systems arecomposed either partially orwholly of periods of housed and/orpasture feeding, with attendant changes in management andfeeding (Keane et al., 2009). Consequently, two broad catego-ries of beef production system can be identified — ‘extensive’and ‘intensive’. Although nuance is inherent where, forexample, even predominantly outdoor pastoral systems canbe regarded as ‘intensive’ or ‘extensive’ depending on output ofcarcass weight per unit area (stocking rate) or per unit ofnitrogen fertiliser application (Drennan andMcGee, 2009) andwhether animals are seasonally housed and fed concentrates(Keane et al., 2009), ‘extensive’ and ‘intensive’ productionsystems are synonymous with predominantly ‘outdoor’ and‘indoor’ rearing. The terms ‘extensive’, ‘free range’, ‘outdoor-
reared’ and ‘grass-fed’ have similar connotations and areoccasionally used interchangeably.
Production system intensity, indexed by daily growth rate(daily liveweight gains), has been demonstrated to havesignificant effects on meat quality, primarily on composition(% lipid) and colour (Keane and Allen, 1998) although, ifapplication of fertiliser nitrogen (in so far as this can be usedas an index of intensity within grassland production systems)was reduced there were no effects on meat quality, despite asignificant reduction in herbage production (Keane and Allen,1999). In practise, although the majority of beef productionsystemswill have substantial periods of outdoor, interspersedwith seasonal indoor management, at least in regions oftemperate grassland abundance such as North-WesternEurope, a definition of what exactly constitutes ‘grass-fed’beef has been difficult to encapsulate and therefore regulate;consequently, some official definitions are sufficiently pre-scriptive (e.g. USDA, 2007) to effectively exclude most cattlewhich, although having grazed forage at various stages duringtheir production, have necessarily received supplementaryconcentrates as nutritional augmentation to overcome sea-sonal deficiencies in pastures or forages.
In North-Western Europe, Ireland and the UK a temperatemaritime climate ensures conditions conducive to grassproduction and grassland utilisation (Lee, 1988) and in thisregion these form the basis for beef production (Bowman andWright, 1995; Thénard et al., 2006). Since feed costs are often amajor proportion of total variable costs in beef productionsystems, this favourable environment for abundant grassgrowth ensures that grazed grass is often the cheapest feedstuff(Balcaen et al., 2002; O'Riordan and O'Kiely, 1996). In Ireland,the majority of calves are Spring-born (Drennan et al., 1995).The most common beef production system involves finishingsteers from about 24 months of age with a grass silage/concentrate indoor finishing diet (Moloney et al., 2001a),after two grazing seasons that last from about March to earlyNovember (Drennanet al., 1995). Thus, by the timeof slaughterat 28 to 30 months cattle may have spent up to two-thirds oftheir lifespan grazing. In contrast, beef production in SouthernEurope does not rely to such a large extent on grassland,typically resulting in more intensively-managed, housed cattle

3P.G. Dunne et al. / Livestock Science 142 (2011) 1–22
which are fed concentrates and slaughtered at younger agesthan animals in North-Western Europe (Gigli and Iacurto,1995; Monserrat et al., 2001).
There is awidespread perception that animals reared under‘free-range’ or ‘extensive’ conditions produce darkermeat thanthose reared intensively due to greater levels of exercise/physical activity (Varnam and Sutherland, 1995). This concepthas been advanced as an explanation for observed differencesbetween the colour of grass and grain-fed beef (Muir et al.,1998b; Priolo et al., 2001). However, although exercise isrecognised as a factor which can cause muscle fibres to changewith possible consequences for meat quality (Lefaucheur andGerrard, 2000), until relatively recently very few studies havesought to confirm any such effect.
A key aspect of any ‘free-range’ or ‘extensive’ animalproduction system is likely to be the amount of space whichanimals are afforded to inhabit, which in any commercialcontext usually has some realistic upper limit, in markedcontrast to wild, free-living species, e.g. deer, but alsorepresenting a larger space allowance than is usually permittedto intensively-reared livestock. Another concept, synonymouswith ‘free-range’ rearing of livestock, is the provision of “theopportunity to livestock to perform their natural behaviouralpattern” (Jonsäll et al., 2002). For cattle on pasture, this is likelyto include significant periods in foraging or food-searchbehaviour, as demonstrated by O'Sullivan (1983). However,manipulation of housing conditions has also been shown tohave an effect onmeat colour,with cattle housed ondeep strawbedding producing darker meat than counterparts that wereconfined in stalls (Jensen and Oksama, 1996; Vestergaard et al.,1997) although, more recently Dunne et al. (2008) reported nodifferences in meat colour in different, although less divergentand extreme accommodation options.
In recent years, consumers have apparently become in-creasingly concerned about the origin of meat and how it isproduced, with respect to rearing conditions of meat-producing livestock, which has become an issue in the overallperception of meat quality (Jonsäll et al., 2001; Moloney et al.,2001b; Sather et al., 1997). Consumers also place greateremphasis on meat products that are perceived as healthy and‘natural’ (Morrissey et al., 1998). In both these regards, itseems that ‘grass-fed’ beef from cattle reared in a ‘free-range’or ‘extensive’ environment may conform to consumers'perceptions of ‘ethical’ quality or what constitutes a ‘natural’production system and is gaining popularity (Roosevelt,2006). However, therein lies a dichotomy since althoughbeef produced from pasture may be perceived to be morehealthy and natural, it is also perceived to be darker, which isgenerally less favoured by consumers in many markets.
The aimsof this revieware (i) to present current evidence insupport of the hypothesis that physical activity/exercise ratherthan diet/nutrition could be the primary contributor to theoften-reported darker muscle of pastured (or extensively-managed) cattle and (ii) to outline putative mechanisms bywhich such an effect may be caused. It could be hypothesisedthat ‘physical activity’ or ‘exercise’ (appropriately and contex-tually referred to as ‘grazing mobility’ by Jurie et al., 2006) is afactor implicated in production of ‘darker’ beef by extensively-managed cattle. Therefore, colour differences apparentlycaused by diet (grass/forage vs. grain) may in fact be primarilydue to physical activity or perhaps other, thus far unidentified
factors. To be specific, we hypothesised that the metabolictendencies of bovine muscle can be affected sufficiently by the‘exercise’ involved in grazing (perhaps in addition to, or incombinationwith other factors, including diet) to cause the cutsurface of pasture-fed beef to appear ‘darker’. Currently, fromour reviewing of the literature, presented herein, we feel thishypothesis may merit further investigation.
2. The basis of meat colour
2.1. The phenomenon of meat colour and its measurement
The phenomenon that is experienced as muscle colourdepends on the interaction of the incident radiation from thevisible region of the electromagnetic spectrum with thepigments (chiefly myoglobin but also putatively, to somesmall extent, haemoglobin and perhaps cytochromes). Percep-tion of colour also depends on optical phenomena such as lightscattering which is influenced by the interaction of incidentradiation with the ultrastructure of the muscle, since a cutmuscle surface is not a solid but a translucent surface. Thecolour of muscle depends both on quantitative and qualitativeaspects of the myoglobin present, i.e. on how much totalmyoglobin is present and onwhat proportion of themyoglobinpresent is oxygenated (oxymyoglobin), deoxygenated(‘deoxy'myoglobin ormyoglobin) or oxidised (metmyoglobin).A more comprehensive review of meat colour is beyond thescope of the present review but for convenience, the reader isreferred to many previous reviews of meat colour andmyoglobin chemistry which provide a more thorough, com-prehensive discussion of the subject (Bekhit and Faustman,2005; Cornforth, 1994; Govindarajan, 1973; Hunt and Kropf,1987; Lawrie, 1998a; Ledward, 1984, 1992; Livingston andBrown, 1982; MacDougall, 1982, 1986, 1994; Mancini, 2009a,2009b;Mancini andHunt, 2005; Pérez-Alvarez and Fernández-López, 2006; Varnam and Sutherland, 1995; Wallace et al.,1982; Young and West, 2001) whilst several reviews havefocused on, or included commentary on, the effects ofproduction methods (Daly et al., 1999; Geay et al., 2001;Mancini and Hunt, 2005; Muir et al., 1998a; Oddy et al., 2001;Owens and Gardner, 1999; Priolo et al., 2001; Young et al.,1999) or have discussed meat colour from the perspective ofpackaging systems influence on meat colour (Gill, 1996;Mancini and Hunt, 2005; McMillin, 2008).
In the context of the present review, it is useful to recognisethat there are many factors which have or are likely to have aneffect onpost-mortemmuscle biochemistry, includingeffects onpigments. These factors can be categorised as either extrinsic orintrinsic. Extrinsic factors can be subdivided into (i) environ-mental (e.g. climate, nutrition, accommodation, stocking rates/densities/space allowance, floor type/underfoot conditions,social behaviours and acute ante-mortem stress) and (ii) post-mortem technological factors (e.g. electrical stimulation, carcasssuspension techniques, chilling rate, ageing, packaging, freez-ing and further processing) (Dunne et al., 2008). Intrinsicfactors relate to the biology of the animal, e.g.breed, gender, ageat slaughter, live- and carcassweight and fatness.Manyof theseintrinsic factors are themselves artefacts of the particularproduction system used, and are thus dictated by climatic,socio-economic, cultural and regulatory factors.

4 P.G. Dunne et al. / Livestock Science 142 (2011) 1–22
Apart from the earlier mentioned reviews of muscle colour,the phenomenon of discolouration in meat has also receivedattention in the literature (Bekhit andFaustman, 2005; Faustmanand Cassens, 1990; Jakobsen and Bertelsen, 2002), althoughusually in the context of its dependence on other factors, such asmetmyoglobin reducing activity (Bekhit and Faustman, 2005) ordietary vitamin E supplementation (Faustman et al., 1998; Liuet al., 1995); often, many of the reviews of factors affectingmeatcolour have included a fully integrated discussion on discoloura-tion. However, Dunne (2003) proposed separate definitions for‘meat colour’ and ‘meat colour stability’, recognising thatalthough colour and its stability (or discolouration) are influ-enced by many of the same factors, they are capable of separatedescription and definition. The present review will focus onputative effects of exercise onmeat colour rather than effects onmeat colour stability although there may also be scope todevelop hypotheses addressing the implication that exercisemay affect meat colour stability (i.e. colour shelf-life), perhapsdue to generation of reactive oxygenmetabolites, lipid oxidationintermediates or unsaturated aldehydes.
Meat colour was defined by Dunne (2003) as “the colourattained by a freshor aged slice ofmuscle tissue of a pre-definedthickness, usually 2.5 cm, with its cut surface perpendicular tothe direction of the muscle fibres. This colour will usually havebeen attained after a user-defined duration of exposure tonormal atmospheric concentrations of oxygen, in darkness,such that blooming occurs”. Meat colour stability was definedby Dunne (2003) as “the ability of bloomed meat to retain itsbloomed colour, or at least a desirable colour [however thatmight be defined in a particular set of circumstances], eitherunder normal atmospheric concentrations of oxygen, or in anatmosphere containing an elevated concentration of oxygen,typically 80%O2: 20%CO2 (referred to as a ‘modified atmo-sphere’) and typically under [simulated] retail display condi-tions.” Where reference is made in these definitions to pre-definedphysical characteristics ofmeat slices andmeasurementconditions the reader is recommended to refer to Boccard et al.(1981), Honikel (1998) or the American Meat Science Associ-ation Guidelines for Meat Colour Evaluation (1991) forstandardised methods of meat colour measurement (currentlybeing revised and updated).
Colour of meat, in common with other foods, is charac-terised by its lightness or luminosity (‘L’ value), its redness/greenness (‘a’ value) and its yellowness/blueness (‘b’ value).Saturation or chromaticity (‘C’ value) and hue angle (‘H’ value)describe the intensity of a particular colour value and the actualperceived colour, respectively. The annotation used for thesecolour descriptors also depends on the colour space that is usedwith the most common colour spaces being the Hunter Labcolour space and the CIE (1976) L* a*b* (CIELAB) colour space ofthe Commission Internationale de l'Éclairage. For a morecomprehensive and detailed explanation of the theory under-lying meat colour measurement, the reader is referred toCornforth (1994), Francis and Clydesdale (1975), Hunt andKropf (1987), MacDougall (1982), MacDougall (2002), andPomeranz and Meloan (1994).
2.2. Muscle fibre types and meat colour
Although beyond the scope of this review, some under-standing of muscle fibre types, how they are defined and
identified is necessary to be able to fully understand howphysical activity or exercise may affect meat colour. One ofthe original reasons for studying muscle fibre types in a meatquality context was to gain a greater understanding of thebasis for the colour of different muscles within and betweenspecies (Aalhus et al., 2009). Muscle fibre types and meatquality have been reviewed by Aalhus et al. (2009), Choi andKim (2009), Klont et al. (1998) and Lefaucheur and Gerrard(2000). According to Aalhus et al. (2009) a number of majorclassification systems have evolved for muscle fibre typing, sothat depending on whether the context for muscle fibretyping and identification is in human medicine, agriculturaland food science or physical education and athletic perfor-mance the nomenclature is likely to be different. However, allof these classification systems share some commonalities sothat classifications based on combinations of colour (red orwhite; based on myoglobin and mitochondrial content,capillarisation), contractile speed (slow or fast; based onmyosin ATPase activity) or metabolic oxidative and anaerobiccapacity (based on enzyme activities) are possible (seeAalhus et al., 2009; Greaser et al., 2001). A key concept inunderstanding the likely muscle fibre response to exercise isthat muscle fibres exhibit plasticity, i.e. the ability to changefrom one fibre type to another, a process which in relation tothe effects of exercise, at least, appears to be under acombination of environmental but also genetic control(Booth and Neufer, 2005; Wackerhage and Woods, 2002).
There is also a continuum of muscle fibres within muscles,such that it is rare for a muscle to have exclusively ‘red’ or‘white’ fibres — most muscles have a mixture of both fibretypes together with a substantial proportion of intermediatefibres to permit flexibility to adapt to particular environmen-tal stimuli or to increased demands for exercise, such asincreased grazing mobility. Moreover, different muscles arelikely to respond differently to exercise, depending onwhether they are predominantly postural or propulsive/locomotive (Aalhus et al., 2009), which depends on theiranatomical location but is perhaps not surprising given thatdifferences in global gene expression patterns betweenmuscle types continue to emerge (Campbell et al., 2001).However, Ordway and Garry (2004) stated, in relation tomyoglobin content, that “although genetics plays an impor-tant role in establishing inherent levels of muscle myoglobincontent, developmental programmes and/or environmentalcues and stresses such as physical activity, temperature andO2 availability play at least equally important roles in deter-mining functional levels of this protein”.
3. Comparison of muscle colour of forage-finished andconcentrate-finished cattle
3.1. Relevant comparisons: are they confounded by physicalactivity?
Many muscle colour comparisons of forage-fed or grazinganimals with concentrate-fed animals have been made, withthe latter typically being accommodated in a feedlot or drylotsituation (Table 1). Cattle raised in these contrasting situationshave different diets but data in the literature remain equivocalwith regard to any effect of diet composition on bovinemusclecolour, when freshly cut meat is assessed (as opposed to the

Table 1Experiments which have compared meat colour of forage-fed (predominantly pasture) and grain-fed cattle.
Reference Comparison Colour difference Potential confounding factors Assessment (subjective/instrumental)
Huuskonen et al. (2010) 62 day grazing season (1 June–1 August)(PAS) v. uninsulated barn (UB)
UB bulls were lighter, more red and moreyellow than PAS bulls (all Pb0.05) atequivalent carcass weights
Carcass fatness Instrumental (Minolta CR200 chromameter)L value of 41.0 and 39.2, a value of 27.8 and26.6 and b value of 10.4 and 9.6 for UB andPAS bulls, respectively
Blanco et al. (2010) Concentrates(CON) v lucernegrazing+1.8 kg DM barley/day (LUC)v same management as LUC for2 months then CON for 2 months
No difference Instrumental (Minolta CM-2600dspectrophotometer)
DelCampo et al. (2008) T1: pasture, offered at 4% of liveweight (LW); T2: pasture, offered at 3%of LW and grain (0.6% of LW); T3: pasture,offered at 3% of LW and grain (1.2% LW);T4: concentrate and alfalfa hay, offeredad libitum.
No difference in muscle lightness after20 days ageing but T4 significantlyless red than other T
Age at slaughter, carcass weight Instrumental
Duckett et al. (2007) Angus cross steers assigned to eitherconc (CONC) or pasture (PAST) finishingsystems after the stocker phase
PAST darker and less red than CONC Carcass fatness Instrumental (Minolta CR310 chromameter)L* value 42.06 and 38.93 for CONC and PAST(Pb0.001) and a* value 24.96 and 23.17 forCONC and PAST (P=0.002).
Dunne et al. (2006) Permanently-housed plus concentrates(PH-CON) v grazing (PAS – 90 d)+ concentrates (PAS-CON), grasssilage (GS) at 20% of dietary drymatter (DM) (PAS-GS20), 50% ofdietary DM (PAS-GS50) or zero-grazedgrass (PAS-GRA). All treatments (T)receiving supplementary concentratesto ensure similar growth rates.Heifers slaughtered at days (D) 28,56, 91 and 120 after housing.
Tendency for PH-CON heifers to havelighter muscle and lower pigmentationthan other heifers
Instrumental (Minolta CR300 chromameter)
Razminowicz et al.(2006)a
Pasture-derived label beef (suckler beef(PS) and steers/heifers(PF)), other labelbeef (young bulls, LB) and conventionalbeef (heifers (CH) and young bulls (CB))
LB was lightest (significantly differentfrom PF, CH and CB by t-test, Pb0.05).PF tended to be most red but wassignificantly more red than PS (Pb0.05).
Gender, probable age at slaughter, carcassweight, carcass fatness as these were retailsamples not generated under controlledexperimental conditions, though notdocumented
Instrumental (Minolta CR300)L* values of 39.8, 38.8, 41.0, 37.5 and 38.7(SEM=0.88) for PS, PF, LB, CH and CB.a* values of 20.3, 23.6, 22.6, 22.2 and 21.4 for PS,PF, LB, CH and CB.
Gatellier et al. (2005) Pasture (during summer) or mixed-diet (maize silage and hay) finishing
At day 0 of simulated aerobic retaildisplay, L* values were higher in LDfrom pasture-fed animals
Animal age, as these animals were fromcommercial farms
Instrumental
Nuernberg et al. (2005) Grass-based v concentrate for each ofGerman Holstein (GH) and GermanSimmental (GS) bulls
Grass-based darker (see also Fig. 3) Age at slaughter Instrumental
Realini et al. (2004) Pasture v concentrate v concentrateplus 1000 I.U. vitamin E
Pasture darker Carcass fatness Instrumental (Minolta CR210 chromameter)L* value 33.80 and 35.56 for pasture and concentratesteers, respectively (Pb0.05). No difference in thea* value (20.45 and 20.42, respectively)
Moloney et al. (2004) 24-week-old Holstein Friesian bull calveseither housed (H, 2.5 m2/animal) ormaintained at pasture (P, 133m2/animal)for 180 days prior to slaughter
No difference in colour, either at 2 or 7 days(‘. . .exercise/environment sufficient to alterbody composition did not affect LTmuscle colour in bulls’)
Instrumental (Hunter lab UltraScan XEspectrocolorimeter)
(continued on next page) 5P.G
.Dunne
etal./
LivestockScience
142(2011)
1–22
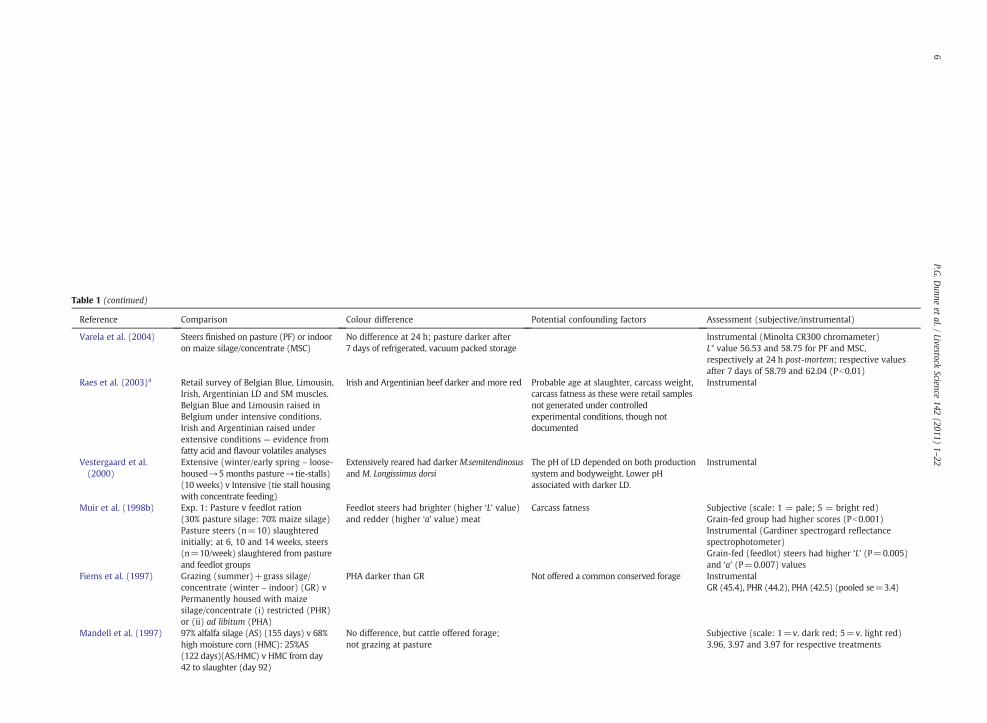
Table 1 (continued)
Reference Comparison Colour difference Potential confounding factors Assessment (subjective/instrumental)
Varela et al. (2004) Steers finished on pasture (PF) or indooron maize silage/concentrate (MSC)
No difference at 24 h; pasture darker after7 days of refrigerated, vacuum packed storage
Instrumental (Minolta CR300 chromameter)L* value 56.53 and 58.75 for PF and MSC,respectively at 24 h post-mortem; respective valuesafter 7 days of 58.79 and 62.04 (Pb0.01)
Raes et al. (2003)a Retail survey of Belgian Blue, Limousin,Irish, Argentinian LD and SM muscles.Belgian Blue and Limousin raised inBelgium under intensive conditions.Irish and Argentinian raised underextensive conditions — evidence fromfatty acid and flavour volatiles analyses
Irish and Argentinian beef darker and more red Probable age at slaughter, carcass weight,carcass fatness as these were retail samplesnot generated under controlledexperimental conditions, though notdocumented
Instrumental
Vestergaard et al.(2000)
Extensive (winter/early spring – loose-housed→5 months pasture→tie-stalls)(10 weeks) v Intensive (tie stall housingwith concentrate feeding)
Extensively reared had darkerM.semitendinosusand M. Longissimus dorsi
The pH of LD depended on both productionsystem and bodyweight. Lower pHassociated with darker LD.
Instrumental
Muir et al. (1998b) Exp. 1: Pasture v feedlot ration(30% pasture silage: 70% maize silage)Pasture steers (n=10) slaughteredinitially; at 6, 10 and 14 weeks, steers(n=10/week) slaughtered from pastureand feedlot groups
Feedlot steers had brighter (higher ‘L’ value)and redder (higher ‘a’ value) meat
Carcass fatness Subjective (scale: 1 = pale; 5 = bright red)Grain-fed group had higher scores (Pb0.001)Instrumental (Gardiner spectrogard reflectancespectrophotometer)Grain-fed (feedlot) steers had higher ‘L’ (P=0.005)and ‘a’ (P=0.007) values
Fiems et al. (1997) Grazing (summer)+grass silage/concentrate (winter – indoor) (GR) vPermanently housed with maizesilage/concentrate (i) restricted (PHR)or (ii) ad libitum (PHA)
PHA darker than GR Not offered a common conserved forage InstrumentalGR (45.4), PHR (44.2), PHA (42.5) (pooled se=3.4)
Mandell et al. (1997) 97% alfalfa silage (AS) (155 days) v 68%high moisture corn (HMC): 25%AS(122 days)(AS/HMC) v HMC from day42 to slaughter (day 92)
No difference, but cattle offered forage;not grazing at pasture
Subjective (scale: 1=v. dark red; 5=v. light red)3.96, 3.97 and 3.97 for respective treatments
6P.G
.Dunne
etal./
LivestockScience
142(2011)
1–22
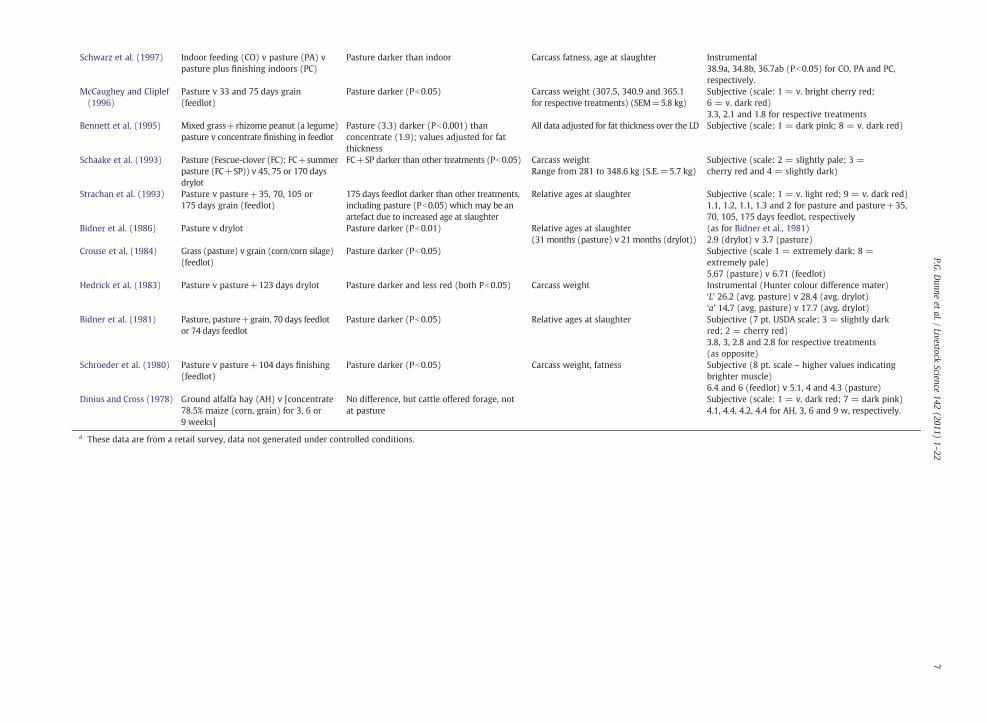
Schwarz et al. (1997) Indoor feeding (CO) v pasture (PA) vpasture plus finishing indoors (PC)
Pasture darker than indoor Carcass fatness, age at slaughter Instrumental38.9a, 34.8b, 36.7ab (Pb0.05) for CO, PA and PC,respectively.
McCaughey and Cliplef(1996)
Pasture v 33 and 75 days grain(feedlot)
Pasture darker (Pb0.05) Carcass weight (307.5, 340.9 and 365.1for respective treatments) (SEM=5.8 kg)
Subjective (scale: 1 = v. bright cherry red;6 = v. dark red)3.3, 2.1 and 1.8 for respective treatments
Bennett et al. (1995) Mixed grass+rhizome peanut (a legume)pasture v concentrate finishing in feedlot
Pasture (3.3) darker (Pb0.001) thanconcentrate (1.9); values adjusted for fatthickness
All data adjusted for fat thickness over the LD Subjective (scale: 1 = dark pink; 8 = v. dark red)
Schaake et al. (1993) Pasture (Fescue-clover (FC); FC+summerpasture (FC+SP)) v 45, 75 or 170 daysdrylot
FC+SP darker than other treatments (Pb0.05) Carcass weightRange from 281 to 348.6 kg (S.E.=5.7 kg)
Subjective (scale: 2 = slightly pale; 3 =cherry red and 4 = slightly dark)
Strachan et al. (1993) Pasture v pasture+35, 70, 105 or175 days grain (feedlot)
175 days feedlot darker than other treatments,including pasture (Pb0.05) which may be anartefact due to increased age at slaughter
Relative ages at slaughter Subjective (scale: 1 = v. light red; 9 = v. dark red)1.1, 1.2, 1.1, 1.3 and 2 for pasture and pasture+35,70, 105, 175 days feedlot, respectively
Bidner et al. (1986) Pasture v drylot Pasture darker (Pb0.01) Relative ages at slaughter(31 months (pasture) v 21 months (drylot))
(as for Bidner et al., 1981)2.9 (drylot) v 3.7 (pasture)
Crouse et al. (1984) Grass (pasture) v grain (corn/corn silage)(feedlot)
Pasture darker (Pb0.05) Subjective (scale 1 = extremely dark; 8 =extremely pale)5.67 (pasture) v 6.71 (feedlot)
Hedrick et al. (1983) Pasture v pasture+123 days drylot Pasture darker and less red (both Pb0.05) Carcass weight Instrumental (Hunter colour difference mater)‘L’ 26.2 (avg. pasture) v 28.4 (avg. drylot)‘a’ 14.7 (avg. pasture) v 17.7 (avg. drylot)
Bidner et al. (1981) Pasture, pasture+grain, 70 days feedlotor 74 days feedlot
Pasture darker (Pb0.05) Relative ages at slaughter Subjective (7 pt. USDA scale; 3 = slightly darkred; 2 = cherry red)3.8, 3, 2.8 and 2.8 for respective treatments(as opposite)
Schroeder et al. (1980) Pasture v pasture+104 days finishing(feedlot)
Pasture darker (Pb0.05) Carcass weight, fatness Subjective (8 pt. scale – higher values indicatingbrighter muscle)6.4 and 6 (feedlot) v 5.1, 4 and 4.3 (pasture)
Dinius and Cross (1978) Ground alfalfa hay (AH) v [concentrate78.5% maize (corn, grain) for 3, 6 or9 weeks]
No difference, but cattle offered forage, notat pasture
Subjective (scale: 1 = v. dark red; 7 = dark pink)4.1, 4.4, 4.2, 4.4 for AH, 3, 6 and 9 w, respectively.
a These data are from a retail survey, data not generated under controlled conditions.
7P.G
.Dunne
etal./
LivestockScience
142(2011)
1–22

8 P.G. Dunne et al. / Livestock Science 142 (2011) 1–22
stability of the colour subsequently, which is a separate issue).In suchcomparisonsbetweengrass- andgrain-/concentrate-fedcattle, where pasture or grass-fed cattle have been consistentlyreported to have ‘darker’ muscle, most commonly in theM. longissimus dorsi (LD), direct inference regarding the effectof diet composition on muscle colour can be misleading.
Such studies are often confounded by some environmen-tal factor(s), inherent to comparisons of pasture or rangelandwith intensive feedlots or housing situations, such as averagespace allowance per animal and foraging opportunity.Therefore, it seems reasonable to hypothesise that pasturedcattle engage in more physical activity/exercise than cattle infeedlots or relatively confined indoor environments, but theamount of activity could also be contingent on grazingmanagement techniques employed, as demonstrated byO'Sullivan (1983). It could be further hypothesised that thisenhanced level of activity causes the frequently reported‘darker’ muscle of pasture cattle.
This hypothesis derives from a common perception,reinforced by anecdotal evidence, that muscles of wild, free-living species such as deer, or free range livestock, are ‘darker’than domesticated or intensively-managed counterparts(Essén-Gustavsson, 1996; Varnam and Sutherland, 1995).Craig et al. (1966) recognised exercise as a potentialcontributor to differences in beef colour. However, wherepasture and grain-fed feedlot cattle are slaughtered at thesame carcass weight or after the same duration of feeding,differences in age at slaughter or in carcass weight andfatness, respectively, confound the effect of diet compositionon meat colour, but also make it difficult to conclude thatphysical activity unequivocally caused muscle of pasture-fedcattle to be darker.
Many trials, outlined in Table 1, were concerned withexamining the effects of pasture versus grain-feeding onanimal performance and meat quality, but in relation to thelatter, the main focus was often not placed on meat colour,hence the presence of confounding alternative explanationsto observed differences in meat colour. Despite the presenceof confounding factors arising from dietary and environmen-tal differences in many trials, the balance of availableevidence, presented in Table 1, suggests strongly that cattlewhich engage in grazing activity or that are reared in a so-called ‘extensive’ environment, consistently produce darkermuscle. However, the question remains: is the ‘darker’ beef ofpasture-fed cattle caused by increased physical activity orgrazing mobility? Alternatively, the question could be ‘howmuch variation in muscle colour is caused by physicalactivity?’ (which accepts that physical activity may not besolely responsible for differences in meat colour betweengrass/forage-fed and grain-fed cattle, but is a component ofthat variation)..
Hocquette et al. (1998) recognised the potential effect ofexercise on meat quality. Muscle colour differences betweenpasture- and grain-fed cattle appear to have many causes, oneof which may plausibly be differences in physical activity.When allied to anecdotal evidence concerning meat colourdifferences between extensively- and intensively-managedanimals, it has led to speculation amongst animal and meatscientists that perhaps the increased opportunity for pasturedcattle to engage in voluntary and spontaneous activity, as aconsequence of their grazing environment, rather than their
grass diet per se may contribute to the production of darkermuscle (Muir et al., 1998b; Vestergaard et al., 2000). Jurie et al.(2006) recognised that it was “tempting to speculate that, incattle, a spontaneous, regular and low level of physical activity,such as that observed at pasture, may be sufficient to regulatemuscle metabolic properties” but did not specifically extendtheir hypothesis to include effects on meat colour, althoughthey made reference to it. Notably, however, Jurie et al. (2006)also recognised the central conundrum in comparing muscleproperties of grass-fedwith concentrate-fed cattle, i.e.whethereffects were due to diet per se or ‘grazing mobility’ as theyreferred to it; in a novel experiment they reported that theactivity of certain oxidative enzymeswas increasedwhen cattlewere offered grass in comparison with maize silage, perhapsrelated to compositional differences such as in polyunsaturatedfatty acids or trace minerals such as manganese and resultingmetabolic effects. Vestergaard et al. (2000) proposed that instudies utilising grazing any effect of physical activity on beefcolour may be confounded with the effect of feeding level and(or) feed ration but the converse may also apply; comparingmuscle colour of pasture-finished cattle with those finishedindoors on concentratesmay fail to take account of thedifferentenvironmental influences and opportunities for physicalactivity that exist.
3.2. Do dietary ingredients affect beef colour, i.e. is diet/nutritionalways the obvious cause of differences in beef colour betweenpasture- and grain-fed cattle?
A hypothesis that diet composition has little impact onmuscle colour, at least in adult bovines slaughtered at similarages, levels of fatness and (or) carcass weights, is supportedby data from Moloney et al. (2000a). Moloney et al. (2000b)reported no effect of diet composition, (grass silage versusearly and late wheat silages) on beef colour and both Frenchet al. (2000, 2001)and Walsh et al. (2007) also reported noeffect. Mandell et al. (1997) found no difference betweenforage- and grain-fed steers in terms of muscle colour,although their colour values were subjective grading scores.Varela et al. (2004) reported no difference in muscle colourbetween Rubia Gallega steers finished on pasture or maizesilage plus concentrates, when slaughtered at 30 months andequivalent carcass weights. Moloney et al. (2001a) did reporta diet effect (Pb0.05) on muscle ‘a’ (redness) and ‘b’(yellowness) values, as did Hoving-Bolink et al. (1999),who also reported a diet effect on ‘L’ (lightness) values.Kennedy et al. (2009) reported no difference in L* values ofcontinental cross steers offered diets including grass silage orlupins/triticale silage and vetch/barley silage offered eitheralone or in combination. The ultimate pH was not differentamongst treatments. Lee et al. (2009) reported no differencein bloomed colour between cull cows offered grass silage andred clover silage.
It is important to re-emphasise the subtle distinctionbetween ‘meat colour’ and ‘meat colour stability’ at this pointsince although evidence suggests, as outlined, that dietcomposition/ingredients (as opposed to dietary energy) donot cause significant variation in muscle colour afterblooming, there is abundant evidence in the literature thatthe ingredients in ruminant diets can play a significant role inthe colour and associated lipid stability (Lanari et al., 2002;

9P.G. Dunne et al. / Livestock Science 142 (2011) 1–22
O'Sullivan et al., 2004; Poulson et al., 2004; Realini et al.,2004; Warren et al., 2008), probably mediated by the dietarysupply of unsaturated fatty acids (see review by Woods andFearon, 2009) and antioxidants, particularly vitamin E (seereviews by Faustman et al., 1998; Liu et al., 1995).
It is important to note that dietary composition can differconsiderably between extensively-managed and intensively-managed cattle since, in general, extensively-managed cattlespend a greater time grazing pasture. In such situations,intakes of unsaturated fatty acids (particularly α-linolenicacid, C18:3n-3), antioxidants (particularly vitamins C and E)and minerals such as manganese (Jurie et al., 2006) areusually greater for grazing ruminants. However, althoughdietary composition, at equal energy intakes, can affectoxidative enzymes activity in bovine muscle (Jurie et al.,2006; Listrat et al., 1999) whether and by what mechanisms,colour might be affected, remains to be consistently demon-strated and thereby clarified. In this regard, both polyunsat-urated fatty acids (Clarke, 2000; Wahle et al., 2003) andvitamin E (Rimbach et al., 2002) can act as cell signallingmolecules and thereby may play a role, as yet undetermined,in processes affecting muscle colour. Elevated intakes of n-3polyunsaturated fatty acids are associated with increasedoxidative metabolism (Jurie et al., 2006; Maillet and Weber,2007).
3.3. Could the method of colour evaluation affect colourdifferences between pasture/forage-fed and grain-fed cattle?
Although a diet effect has been reported on muscle ‘a’ and‘b’ values, to achieve perceptible differences in visual colour,changes in the ‘L’ value of meat might also be necessary, sincesubjective assessments of colour differences between grassand grain-fed beef have most frequently been described interms of the lightness component of colour, i.e. pasture-fedbeef has traditionally been specifically described as ‘darker’where comparisons, usually subjective, have been made(Table 1). Consequently, it could be reasonably concludedthat the lightness of beef is themost easily perceived aspect ofcolour during subjective, visual assessment.
Hence, ambiguity regarding the effects of diet on meatcolour may also be partly contributed to by inconsistencies inhow meat colour is assessed, in some cases by instrumentalmeasurement (Dufrasne et al., 1995; French et al., 2001;Moloney et al., 2001a, 2000a, 2000b; Muir et al., 1998b;Vestergaard et al., 2000, 1997) and in other cases by subjectivevisual appraisal (Crouse et al., 1984; Harrison et al., 1978;Mandell et al., 1997; McCaughey and Cliplef, 1996; Schaakeet al., 1993). Priolo et al. (2001) reported that subjective visualassessment had several weaknesses including frequent omis-sion of a blooming time, undue influence of lighting conditions,lack of standardisation and the inherent bias associated withany subjective appraisal. Although guidelines exist whereinstandardised methods of meat colour measurement arerecommended (American Meat Science Association, 1991;Boccard et al., 1981), various authors have measured beefcolour under a variety of conditions (Table 2).
Furthermore, ‘colour’ as a concept is psychophysical innature, involving perception and subjective interpretation andcan be complicated by the involvement of psychologicalconcepts such as appearance (Sheppard, 1968) as well as
factors such as surface texture (Stevenson, 1990).Hence,whilsta description of beef from grass- or forage-fed cattle as being‘darker’ than that from feedlot counterparts may be adequatelydescriptive from a sensory perspective, it may lack sufficientdescriptive detail when it is replaced by the more extensiverange of instrumental colour co-ordinates, i.e. subjectiveassessment often fails to take account of the redness andyellowness, hue and chroma/colour intensity of beef. In thisregard, it is interesting to note that in studies where musclecolour has been instrumentally assessed, pasture-fed cattlehave been reported to producemuscle which is less red aswellas being darker (Muir et al., 1998b).
3.4. Age at slaughter can confound muscle colour comparisonsbased on nutrition
It is important to recognise that an effect of diet com-position or more general nutritional effects on muscle colourmay also be contingent on the age of the animals concerned,since for example, it is common practise to manipulate thediet of calves, principally with respect to iron content but alsoother ingredients, to effect a lighter colour to their muscle(Agboola et al., 1990; Pommier et al., 1995). In contrast, moststudies comparing pasture and feedlot cattle have originatedin North America, Australia or New Zealand and haveinvolved adult bovines of an age that is characteristic ofcattle preparing for commercial beef slaughter, as opposed toveal production.
Keane and Allen (1998) compared three beef productionsystems, two of which utilised steers; in a ‘conventional’system steers were finished on silage plus concentrates andslaughtered at about 24 months; in an ‘extensive’ modifica-tion of the conventional system steers were finished offpasture and slaughtered at about 29 months. The extensiveanimals had darker (lower ‘L’ value; 28.2 v 35.8 (S.E.=0.7))and less red meat although these animals were five monthsolder. A Danish study comparing an intensive with anextensive system reported darker meat from the extensive-ly-reared steers, which had been loose-housed throughoutthe winter and then put out to pasture from May to Octoberbut a ten week finishing period before slaughter minimiseddifferences (Anon, 2000).
The age range of these animals (Anonymous, 2000) wasrelatively narrow in comparison with those used by Bidner etal. (1986) who found that forage-fed steers produced darkercoloured meat. The average age of these forage-fed steers atslaughter was 31 months in comparison with 21 months forthe grain-fed feedlot steers. The age of the forage-fed steers islikely to have played a key role in producing darker meat,since as age at slaughter increases muscle myoglobinconcentrations increase and meat becomes darker (Boccardet al., 1979; Varnam and Sutherland, 1995).
Hedrick et al. (1983) found that steers slaughtered directlyoff pasture had darker coloured meat but that a 123 days grainfeeding period on drylot, post-grazing, resulted in paler meat(‘L’ value increased by an average of 2.2 L units) whichwas alsomore red (‘a’ value increased by 3 a units) (both Pb0.05).Similarly, Schroeder et al. (1980) reported that cattle slaugh-tered off pasture produced darker coloured muscle than theircounterparts that were finished for 104 days on a grain-baseddiet in a feedlot after pasture. In bothof these studies, the grain-

Table 2Colour measurement conditions used by various authors.
Reference Colour measurement conditions a
Huuskonen et al. (2010) Instrumental assessment after 14 days; blooming time not specified.Blanco et al. (2010) Instrumental assessment at 24 h post-mortem; blooming time of 4 h, in the dark at 4 °C.DelCampo et al. (2008) Instrumental assessment after 7 and 20 days vacuum packed storage at 2–4 °C.Duckett et al. (2007) Instrumental assessment after 14 days at 4 °C followed by 30 min blooming at 4 °C.Dunne et al. (2006) Instrumental assessment after 48 h post mortem followed by 3 h blooming at 4 °C.Nuernberg et al. (2005) Instrumental assessment after 24 h post mortem; bloom time not specifiedGatellier et al. (2005) Instrumental assessment of LD recovered at 24 h post mortem and aged for 14 days in vacuo; bloom time of 2 h.Realini et al. (2004) Instrumental assessment after 24 h post mortem followed by 1 h bloom time.Moloney et al. (2004) Instrumental assessment after 7 days vacuum packed storage at 4 °C.Varela et al. (2004) Instrumental assessment after 24 h and 7 days post mortemRaes et al. (2003) Sampling from retail outlets with post sampling 1 h bloom time.Vestergaard et al. (2000) Instrumental assessment after 8 days of storage under vacuum at 5 °CFiems et al. (1997) Instrumental assessment according to standard practise recommended by Boccard et al. (1981).Schwarz et al. (1997) Instrumental assessment, time post mortem not specifiedBennett et al. (1995) Subjective assessment; carcasses were chilled at 0–2 °C for 24 h, ribbed, and graded for USDA quality and yield factorsSchaake et al. (1993) Subjective assessment after 48 h post mortem at 1±2 °CDinius and Cross (1978) Subjective assessment after 48 h post mortem at 2–3 °CTherkildsen et al. (1998) Instrumental assessment after 8 days at 4 °C followed by 80 min blooming at 4 °C.Muir et al. (1998b) Subjective and instrumental assessment within 24 h following a 30 min blooming.Mandell et al. (1997) Subjective assessment following 24 to 48 h at chill temperature.Andersen et al. (1997) Instrumental assessment after 7 days at 4 °C followed by 80 min blooming at 4 °C.Strachan et al. (1993) Subjective assessment after chilled carcass was quartered at the 10th rib and permitted to bloom for 30 min.Bidner et al. (1986) Subjective assessment and instrumental measurement following 2 to 12 months of frozen storage at −29 °C and
thawing for 18 h at 2 °C.Crouse et al. (1984) Subjective assessment following 48 h at chill temperature.Hedrick et al. (1983) Instrumental measurement following 10 days storage at 2 to 4 °C.Bidner et al. (1981) Subjective assessment and instrumental measurement following 24 h at 3 °CSchroeder et al. (1980) Subjective assessment following 24 h at chill temperature
a Includes factors and conditions, where specified or provided, such as bloom time, storage conditions (temperature and duration) prior to measurement andwhether colour was subjectively assessed or instrumentally measured.
10 P.G. Dunne et al. / Livestock Science 142 (2011) 1–22
fed cattle produced lighter coloured meat (subjectivelyassessed by Schroeder et al. (1980)) despite being substantiallyolder at slaughter, i.e. 123 and 104 days, respectively, and alsoall cattle were grazed similarly up to the point where somewere slaughtereddirectly off pasture and those remainingwerefinished in feedlots.
Therefore, it can be assumed thatmuscle colour of the cattleultimatelyfinishedon feedlotswith grainwas the same comingoff pasture as the cattle slaughtered at that point. In otherwords, the cattle slaughtered off feedlots may have had darkermuscle earlier in their lives. If it is accepted that grass-feedingper se apparently has little effect, or is at least equivocal withregard to causing darker fresh beef colour, then it is feasible tohypothesise that any effect of pasture-feeding (or conversely,withdrawal from pasture) on beef colour is mediated throughthe effects of the physical activity, or its relative absence, onmuscle biochemistry and physiology.
Furthermore, the results of Schroeder et al. (1980) andHedrick et al. (1983) suggest that muscle colour, even if darkwhen cattle are at pasture, could be rendered paler followingwithdrawal from pasture. Perhaps this reversion to palermuscle is a consequence of the relative lack of activity of cattlewhen in feedlots/housing relative to pasture-feeding, althoughtransfer to a high-energy diet may confound any effect due todifferences in fatness (Section 2.2). However, this point is alsoequivocal since McCaughey and Cliplef (1996) concluded that“the rib eye colour. . . . . was darkest for animals slaughtereddirectly off pasture and became cherry red as the length of thegrain feeding period increased” but in contrast, Dinius andCross (1978) concluded that “the length of the period thatconcentrate was fed did not affect lean colour.”
The approach taken by Hedrick et al. (1983) and Schroederet al. (1980)was to permit all cattle to graze pasture and then toselect from those the cattle that would be grain-finished. BothCrouse et al. (1984) and Muir et al. (1998b) took a differentapproach whereby from the outset of the trials cattle wereplaced on either a grain or forage/grazing diet. In both cases,muscle colour of pasture-fed cattlewas darker. Pasture affectedboth the ‘L’ (P=0.005) and ‘a’ (P=0.007) values of the steersused by Muir et al. (1998b) whereby pasture-fed steersproduced darker (lower ‘L’ value) and less red (lower ‘a’value) meat. The authors speculated that this may have beendue to higher myoglobin concentrations in the muscles ofpastured cattle as a result of ‘free-range’ finishing.
Although Nuernberg et al. (2005) reported that grass-fedbulls had darker LD than concentrate-fed bulls, the grass-fedbulls were substantially older at slaughter. Similarly, Mahechaet al. (2009) reported a significant effect of finishing diet on L*and although weight at slaughter was not different, the steerswith darker beef on treatment 3 (ad libitum grass plus 1 kgconcentrate) were substantially older at slaughter, by almost100 days. Warren et al. (2008) also reported that, at 19 months,both Holstein-Friesian and Aberdeen Angus steers fed aconcentrate diet had darker, more red and more saturated (allPb0.001) loinmuscle thansteers fedagrass silagediet.However,no differences were observed at 24 months of age, which high-lights different responsiveness ofmuscle colour at different ages.
3.5. Confounding arising from differing nutritional planes
An alternative argument provided by Muir et al. (1998b)to explain the darker meat of the pasture-fed steers was that

11P.G. Dunne et al. / Livestock Science 142 (2011) 1–22
grain-fed steers had an increased ‘marbling’ or intramuscularfat (IMF) content. However, in only four of ten experimentsreviewed by them (Muir et al., 1998a) did improved (lighter,more red) lean colour result from an increase in marblingscores. Animals fed on a grain-based diet and so on acomparatively higher plane of nutrition than grazing animalsproduce fatter carcasses and such increases in fatness areseen in all fat depots, including IMF (Harrison et al., 1978).The influence of plane of nutrition per se on meat colour hasnot been the subject of much study (Shorthose and Harris,1991). Schroeder et al. (1980) found that subjective marblingscores were lower (less marbling) for steers slaughtered offpasture than for those finished in drylot. Similar results wereobtained by Hedrick et al. (1983) whereby steers that wereslaughtered off pasture gave lower subjectivemarbling scoresthan steers that were finished in drylots for 123 days afterwithdrawal from pasture. Therefore, any assertion that thepaler muscle of the older feedlot cattle in the experiments ofSchroeder et al. (1980) and Hedrick et al. (1983) was causedby their relative lack of activity can be countered by theargument in favour of paler muscle being due to highermarbling scores.
Shorthose and Harris (1991) stated that if myoglobinconcentration (and presumably meat colour) is influenced byage, rather than carcass weight, then slaughter of treatmentgroups at the same weight will result in animals on the lowerplane of nutrition having muscles with greater myoglobinconcentrations (because these animals will take longer toreach the same endpoint and so therefore will be older atslaughter). However, if weight is the important factor, thenmuscles of animals slaughtered at the same carcass weightwill have similar myoglobin concentrations, irrespective ofage. Thénard et al. (2006) reported that haeminic iron wasalways lowest in intensively-managed Holstein and Montbé-liard steers, which suggests that, perhaps the rate of increaseof haeminic iron concentration is lower than the rate ofaccretion of other cellular components.
Therefore, in experiments comparing cattle slaughtered offpasture and those finished on grain, the plane of nutrition maybe a confounding factor in relation to meat colour, due to thefact that cattle on the lower plane, typically those on pasture,are invariably older at slaughter if all animals are slaughtered atthe same carcass weight. Alternatively, if slaughtered at thesame age, such animals will have less IMF. According toKrzywicki (1979), marbling, together with pH, connectivetissue, muscle fibre size and denaturation of sarcoplasmicproteins, is one of the factors that affects the path of lightthrough meat and thus, affects the lightness component ofmuscle colour. It is thought that marbling affects both visualand instrumental evaluation of colour (Murray, 1995).
The combined effects of an increase inmyoglobin with ageand less marbling may cause darker coloured meat inpasture-fed cattle, although Craig et al. (1959) reported thatdifferences in colour were most likely due to differences in fatand moisture, not myoglobin. Priolo et al. (2001) mentionthat in several experiments where the carcasses from pasturewere fatter than carcasses from concentrates, the carcassesfrom pasture still produced darker meat. This supports theargument that increased physical activity of pasture-fedcattle and not reduced IMF may be the cause of their darkermeat.
There is another important phenomenon that must beconsideredwhencomparingpasture-fedwith grain-fed cattle—compensatory growth. There is some suggestion in theliterature that restriction of dietary energy may lead toincreased numbers of oxidative muscle fibres or conversion of‘white’ to ‘red’ fibres, a phenomenon acknowledged byVestergaard et al. (2000). Solomon and Lynch (1988) reportedthat lambs fed a low concentrate, high roughage diet(1.87 Mcal ME/kg diet) had fewer αW and more αR fibresthan those fed a high concentrate, low roughage diet(2.8 Mcal ME/kg diet). Johnston et al. (1981) reported thatthere were fewer (Pb0.05) αR and more αW fibres in LD ofgrain-fed compared with grain plus grass-fed cattle and that, ingeneral, as energy levels in the ration increased the percentageof αR fibres decreased and percentage of αW fibres increased.Thus, energy levels can easily confound any putative effects ofexercise on muscle fibre types, oxidative potential and colour.
4. Effect of space allowance, confinement andaccommodation type on muscle colour of cattle
Two recent studies in Finland each compared two indooraccommodation options with an outdoor pasture (Huuskonenet al., 2009; Tuomisto et al., 2009). In both studies the pasture-based animals, growing Ayrshire (Tuomisto et al., 2009) andHereford (Huuskonen et al., 2009) bulls (initial liveweights(across treatments) of 185and285 kg in the respective studies)were accommodated in 50 m×100 m paddocks, providing1000 m2 per bull. In contrast, the indoor bulls were accommo-datedeither in pensmeasuring4 m×8 m(6.4 m2per bull; bothstudies) or in individual tie-stalls (the same dimensions in bothstudies). Meat quality measurements were not performed. Ageneral assumption when comparing extensive/outdoor andintensive/indoor systemsof beef production is that cattle have agreater average space allowance in an extensive system (asdemonstrated by Tuomisto et al. (2009) and Huuskonen et al.(2009)) andwill opportunistically engage in a greater degree ofphysical activity or exercise (‘grazing mobility’), which mayhave physiological and metabolic consequences, and effects onmeat quality including colour. In a grazing situation, anincreased level of activity will also include substantial periodsof necessary foraging or food search behaviour, as reported byO'Sullivan (1983) but this may depend to some extent ongrassland management and grazing systems employed.
4.1. Grassland management and grazing systems
The objective of good grassland management is to strike anoptimal balance between growth, harvest (either by livestockor mechanically) and senescence (Parsons and Chapman,1998). “A grazing management system is defined as anintegrated combinationof animal, plant, soil and other environ-mental components, and the grazing methods by which thesystem is managed to achieve specific results or goals of aproducer” (Clark and Kanneganti, 1998). Ultimately, “grazingsystems and stocking rates are used to influence livestockgrazing behaviour with the intent of improving livestock andvegetation performance” (Hepworth et al., 1991). In anagricultural context, the two principal features of grasslandmanagement are grazing by livestock and conservation ofmechanically harvested swards in temperate grasslands where

12 P.G. Dunne et al. / Livestock Science 142 (2011) 1–22
sward productivity is seasonal. Grassland management fordairy production is generallymore sophisticated anddevelopedthan for sheep and beef cattle (including suckler cows), wherefarmers have traditionally and continue to graze livestock onnon-intensive permanent pasture and rough grazing (Rath andPeel, 2005). Grazing by livestock can be broadly categorised aseither continuous or rotational.
In general, cattle grazing in a continuous grazing systemwill be afforded more space than the same number and typeof cattle in a rotational grazing situation where more focus isplaced on efficient utilisation of the grassland resource byrestricting the available grazing area into smaller discreteareas or paddocks. Perhaps the most important factor toconsider in the context of potential exercise effects on meatquality, including colour, is not the herbage availability per se,but the average amount of space provided to animals ingrazing systems which is at least notionally related to theopportunity to forage and is often dictated by the productivecapacity of the pasture. Other factors such as stocking rateand social interactions between grazing animals (Sowellet al., 1999), nutritive quality and quantity of forage (Senft etal., 1985) and importantly, the accessibility and proximity ofwater sources (probably mostly but not exclusively in arid orsemi-arid regions; paddock/field size may be a factor) (Hartet al., 1993; Hepworth et al., 1991; Irving et al., 1995; Senftet al., 1985; Tomkins and Filmer, 2007) need to be borne inmind and emphasise the important interactive effectsbetween these factors on time spent grazing and/or distancetravelled (Hart et al., 1993; Hepworth et al., 1991; Irving et al.,1995). Therefore grazing management, in so far as it cancontrol the stocking rate or density and the available herbagemass, and therefore the distances that animals have to travelto secure adequate feed resources, probably has an effect onthe amount of exercise and energy expenditure in whichanimals engage.
4.2. Spontaneous physical activity undertaken by grazing cattle
Very few studies have yet been undertaken which attemptto isolate the effects of exercise on bovine muscle colour andassociated biochemical characteristics, althoughboth Jurie et al.(2006) and Dunne et al. (2005) provided steers with dailyexercise of a magnitude (in terms of distance travelled) similarto grazing cattle (5.2 km and 4.41 km, respectively, each day).Studies which have used exercise as a potential explanation forthe observed differences in muscle colour between treatmentgroups have done so in the context of a systems-basedapproach (Muir et al., 1998b; Vestergaard et al., 2000). Suchexperiments implicitly contain a number of componentsresponsible for variation in different production systems andhence, confounding factors can be easily introduced.
A valid question would be to ask howmuch activity grazingcattle undertake voluntarily during a givenperiod of time undergiven conditions. Cattle under ‘free range’ conditions in a widevariety of natural conditions in Australia, South Africa, NewGuinea and the USA spend from 30 to 110 min per 24 h inlocomotion, in addition to which between 500 and 600 min per24 h are spent in mobile grazing; cumulatively, this representsabout10 hofmovementper grazing cow ina typical 24 hperiod(Fraser and Broom, 1997). O'Sullivan (1983), reporting Irishdata, calculated that at a high stocking rate (2740 kg/hectare
(ha)) with a continuous grazing system, cattle would typicallywalk 5085 m, having spent 654 min (10.9 h) engaged ingrazing activity. However, when using a rotational grazingsystem at the same high stocking rate, cattle only walked1876 m and spent only 7.9 h engaged in grazing behaviour.This was probably due to the restriction in the area availablefor grazing and a consequent decrease in per capita herbagemass availability. When monitoring grazing behaviour of 9-month old steers Fraser et al. (2009) reported that 578 minwasspent eating in a 24 h period whilst at 20 months old, steersspent 571 and 488 min grazing on permanent pasture andsemi-natural rough grazing, respectively. Distance travelled,usuallymeasured by pedometer, has also beenused as an indexof activity in grazing cattle on pasture/rangeland (Andersonand Kothmann, 1980).
4.3. Measurement of physical activity engaged in by cattle
In a grazing situation,where strategies to adequately controlcattle activity levels may be limited, except for provision ofvarying space allowance, pedometers provide a means ofactivity measurement. Pedometers (including for example,most recently the ALT pedometer; Brehme et al., 2008) areroutinely used to monitor activity levels in dairy cows as ameans of detecting onset of oestrus (Maatje et al., 1997) andhave been used to monitor activity levels in horses (Holland etal., 1996) and grazing beef cattle (Anderson and Kothmann,1977; Walker et al., 1985). Behavioural observations areanother viable though time-consuming option, with activitylevels indexed by time spent engaged in different activities in agivenunit time (Hassoun, 2002;O'Sullivan, 1983) as opposed tothe more direct pedometer measurement of distance walkedper day. An indirect index of grazing activity is the time that hasbeen determined as ‘eating’ time recorded using GRAZE™[http://www.ultrasoundadvice.co.uk/pages/GRAZEhome.html]JawMovement analysis software, utilisedbyFraser et al. (2009).
Other technologies that have been utilised tomeasure cattleactivity in rangeland or pasture environments, in a rangelandmanagement context, are global positioning systems (GPS) andgeographical information systems (GIS) that may offer moreaccurate and precise measures of locomotive activity anddistances travelled, if not directly of grazing behaviour (Bailey,2001; Tomkins and Filmer, 2007;Ungar et al., 2005).GPS collarsare reported to be accurate to between 4 and 5 m horizontaldistance (Agouridis et al., 2004) and may aid in understanding“how spatial and temporal variability of animal, forage, soil andlandscape features affect grazing behaviour and forage utilisa-tion” so as to provide “potential to modify pasture manage-ment, improve efficiency of utilisation and maximise profits”(Turner et al., 2000) as well as improving sustainability of aridrangelands (Bailey, 2004). However, in the context of thepresent review, any technology or combination of technologiesthat can accurately and cost-effectively record either actualactivity (e.g. walking during foraging/grazing behaviour) orsome measurement which could indirectly index activitywould prove useful.
4.4. Energy expenditure by cattle
Osuji (1974) reviewed the physiology of eating and theenergy expenditure of ruminants at pasture and made the

13P.G. Dunne et al. / Livestock Science 142 (2011) 1–22
point that animals at pasture spend much more time eatingand in food search behaviour compared to indoor housedanimals and that consequently, due to greater muscularactivity and distances travelled, maintenance energy could be25–50% greater than housed counterparts. DiMarco and Aello(1998) reported that the energy expenditure of individually-corralled Angus steers (18–20 months of age; 258±14.9 kgbodyweight (BW) as small size and 327±6.4 kgw BW forlarger animals) was 82.6±3.1 kcal h−1∙100 kg BW−1 [equiv-alent to 650 mL CO2∙h−1∙(BW0.75)−1]. When the steers weretrained to follow a tractor at a constant speed over a pre-determined distance, which varied, the additional cost ofwalking on a level surface and on a 6° gradient was calculatedto be 9.0±1.14 and 16.4±2.18 kcal km−1∙100 kg BW−1,respectively. Ultimately, if pasture-reared or pasture-finishedanimals expend more energy to a substantial extent, musclephysiological and metabolic parameters may alter to such anextent that meat quality, and particularly colour may beaffected, although likely in amuscle-dependentmanner. Evenif cattle accommodated outdoors on pasture had the sameamount of exercise, in terms of distance walked, other factorsthat would likely affect energy expenditure, muscle physiol-ogy and metabolism and could therefore confound any datagenerated, would be altitude, topography and climate (Hays,1976).
4.5. Effects on meat colour
Vestergaard et al. (1997), when comparing two produc-tion systems (extensive and intensive)×two slaughterweights (360 kg and 460 kg), found that when comparingat 360 kg, LD pigmentation was 23% higher and meat colourwas darker by 5.4 L units in the extensive bulls. Similarly,Jensen and Oksama (1996) reported that lightness in LDmuscle was almost 4 L units lower and pigmentation 26.6%higher in deep-bedded compared with tie-stall housed youngbulls. Vestergaard et al. (1997) acknowledged that the darkercolour of the extensive bulls may have been due, in part, tothe significantly higher pH of the meat in addition to thehigher pigment content.
Dufrasneet al. (1995) reporteddarkermeat of bulls that hadbeen grazing for 140 days at bothmedium (n=6/ha) and high(n=8/ha) stocking rates in comparison with those fed aconcentrate diet indoors during the same period, despiteabsence of a difference in myoglobin content between groups(2.60, 2.68 and 2.67 mg/g for medium, high and indoor groups,respectively (S.E.D.=0.25; Pb0.05)). However, bulls whichhad been grazing had higher pH (5.53, 5.52 and 5.48 formedium, high and indoor groups, respectively (S.E.D.=0.02;Pb0.05)) and this is likely to have contributed to their darkermeat. Srinivasan et al. (1998) reported a significantly higheriron concentration in the semimembranosus muscle of grazingsteers compared with those fed grain at pasture.
Greater space allowance can be achieved by permittinganimals to graze but Wiklund et al. (1998) examined eightmuscles of each of ten Swedish dairy cows, four of whichwere constantly tied with a further six being permitted tograze for a period of five months. They concluded that theeffect of grazing did not change any of the biologicalparameters measured in the muscles. Changes in theseparameters (proportion of oxidative muscle fibres, cyto-
chrome oxidase activity and capillary density) might havebeen expected to coincide with a change in muscle colour.
However, two important points to note about this studyare (a) the animals used were all aged between 4 and 8 yearsand so perhaps these cows were too old for any effect ofexercise, such as an increase in muscle pigments, to manifestitself and (b) given the animals age and their probablepredisposition to activity, the intensity of any activity mayhave been insufficient to induce changes in muscle biology,certainly with regard to bioenergetic metabolism andconcomitant colour changes. Also, and notably, only fouranimals were used per treatment.
Ozutsumi and Okada (1981) in a comparison betweensteers reared on mountain grazing for about 200 days beforehousing and steers permanently housed before slaughter,concluded that some ‘white’ muscle fibres changed into ‘red’fibres during the mountainous grazing period. Presumablythe mountainous grazing was sufficient to impose appropri-ate exercise on the steers such that these changes occurred.The nature of the exercise, by implication undertakenvoluntarily by the steers, would probably be close to‘endurance training’ as described by Vestergaard et al.(2000). However, Ådnøy et al. (2005) reported no differencesin meat pH or colour between lambs reared in lowland andmountainous areas in western Norway, although lamb fromnorthern Norway was lighter and more yellow.
Vestergaard et al. (2000) also proposed that muscle fibresaremutable, i.e. can change fromone formto another. Ozutsumiand Okada (1981) reported that the percentage of red musclefibres decreased in the three muscles examined in themountainous grazing group during the fattening period thatfollowed mountainous grazing. It can be speculated that thiswas caused by the absence or lack of physical activity of thesesteers after housing. This reinforces the hypothesis that anyeffects of physical activity onmuscle colour are reversible sinceany effects are likely to be mere adaptations by the muscle to achanged series of stimuli imposed on it. Adaptation of muscletissue to physical activity iswidely recognised in contexts otherthan animal production and meat quality. According to Boothand Neufer (2005) “exercise studies have led Neufer tohypothesise that the adaptations associated with severalweeks of aerobic exercise training actually stem from thecumulative effects of the transient increases in gene expressionthat occur in response to each individual exercise bout. Onlythose proteins with a sufficiently long half-life will accumulatewith successive bouts of exercise, independent of the level ofactivation of the corresponding gene.” This rationale may wellprovide avenues for future research in this area.
In this regard, Solomon and West (1985) pointed out thatnatural selection, i.e. in wild, free-living species, probablyfavours higher muscle aerobic potential, as an adaptation toincreased space and involvement in activity, which enhancesthe ability of themuscles to resist fatigue. For wild bovines andother ruminants, this may also represent a survival-enhancingadaptation since in their natural habitats these herbivoreshave evolved as prey species. Although domestication involvedgenetic selection based on performance characteristics such asgrowth rate and conformation, Ashmore et al. (1972b)proposed that domestication of animals led to a progressivediminution in the importance of red muscle fibres as a naturalselection advantage since muscular endurance was less

14 P.G. Dunne et al. / Livestock Science 142 (2011) 1–22
important because animalswere generally protected from theirenemies and from the requirement to locate food and water.Provision of such conditions was termed by Ashmore et al.(1972b) as “management practises which tend increasingly torestrict muscular activity of meat-producing animals”. Conse-quently, due to inherent metabolic differences, muscles of wildand physically active animals are often said to be darker thanmuscles from domesticated animals (Essén-Gustavsson, 1996).In contrast, Edwards (2005) concluded that exercise in outdoorpig production had little effect on pork colour. Why this mightbe the case is difficult to explain but in each case, activity wasnot measured and there may be behavioural differencesbetween species which could explain these observations.Alternatively, differences between porcine and bovine myo-globin may affect how the molecule itself or its synthesis isaffected by exercise or any of the other factors that differbetween outdoor and indoor management systems.
4.6. Effects of accommodation type
Anenhanced level of physical activitymayalso be replicatedin an indoor environment, though probably to a lesser extent,by manipulating the housing conditions of the animals. Meatquality differences between groups of bovines in differenthousing systems, including commentary on effects on meatcolour, have also been examined (Table 3). Vestergaard et al.(1997) compared tie-stall housed with loose-housed youngbulls. The loose-housed bulls were accommodated on slattedfloors and also on deep bedding The deep-bedded bulls hadhigher muscle pigmentation of LD muscle, (140 v 120 ppm,measured as acid haematin; Hornsey (1956)) as well as darker(lower ‘L’ value)muscle (35.2 v 38.2) and less intense (lower ‘C’value) colour (19.5 v 21.0) (all Pb0.05).
These results were not unexpected. It was anticipated thatthe tie-stall bulls would have engaged in less physical activityconsistent with being restrained. However, an alternative
Table 3Experiments which have compared meat colour of bovines in different housing or
Reference Comparison Colour d
Dunne et al. (2008) Steers on slatted concrete floors (SLA) v straw-bedding (STR) v out-wintering pads (OWP)
No overaon colou
Iacurto et al. (2005) Chianina young bulls on slatted floor (SF;n=10)v feedlot (FL; n=12) v pasture (PA; n=8)
PA darkeno differPA had aeither SFNote: nohousing
Xiccato et al. (2002) Holstein calves housed either in group pens(4 per 7.04 m2) or individual stalls (1 per 1.15 m2)
No differinstrume
Jensen and Oksama(1996)
Tie-stall (TS) v slatted floor (SF)† vdeep-bedded (DB)††: both designated as loose-housing
Meat coPb0.05)‘L’ valuesand DB,
Vestergaard et al.(1997)
TS v SF v DB (as above) Meat colin loose-‘L’ valuesand DB,
Andrighetto et al.(1999)
Veal calves in individual crates (IC) vgroup-pen housing (GP)
GP darkInstrumNo diffe54.5 (GPGP had
argument could be made that cattle restrained in stalls mightperhaps also experience chronic sub-clinical stress resultingfrom such restraint and lack of activity. This might affectmuscle metabolism in a manner having consequences for itscolour when a cut surface is established. In this regard,haematocrit and haemoglobin levels change following expo-sure to adverse conditions for short periods (Broom andJohnson, 1993) so that perhaps such change might also bewitnessed in myoglobin levels.
Dunne et al. (2008) concluded that accommodation typehad only “a minor transient effect” on meat colour. However,in that study the treatments were less divergent, in terms ofspace allowance and the commensurate potential for somephysical activity than in other such experiments, since steerson slatted floors, straw bedding or outdoor wood chip beddedpads had 2.5 m2/500 kg liveweight, 4 m2 per head and 18 m2
per head, respectively.
5. Differential response of muscles to managementpractises under experimental conditions: effects on meatcolour putatively due to different activity levels
It is clear that both grazing of cattle in a pasture situationand also the type of housing used in an indoor environmentcan affect meat colour and it is plausible to suggest that thiseffect may be mediated in both cases by the level of physicalactivity. The differentiation of muscles overall as ‘red’ or‘white’ reflects their relative proportions of individual red andwhitemyofibres (Lawrie, 1998b)which can be affected by thenature of stimuli, which presumably could be changed byexercise of sufficient intensity and duration (Pearson, 1990).
However, it is noteworthy that Vestergaard et al. (1997)mention that in relation to comparison of housing systems, theeffects of deep bedding seen on LD muscle colour andpigmentation were not so pronounced in M. supraspinatus(SU), which indicated that muscles were affected differentially
accommodation systems.
ifference Assessment
ll effect of accommodation systemr of LD or ECR muscles
Instrumental (Hunter Lab Ultra scanXE spectrocolorimeter)
r than SF but not different from FL;ence in chroma between treatments;higher (Pb0.05) hue angle thanor FL.reported interaction betweensystem andmuscle type.
Instrumental (Minolta CM-2600dcolorimeter-spectrometer)
ence, either by visual assessment orntal measurement
Instrumental (Minolta CR100) andsubjective (scale: 1 (pale) to 5 (dark)).
lour darker (lower ‘L’ value;in loose-housed compared with TS.of 39.1, 37.4 and 35.3 for TS, SF
respectively.
Instrumental (Data colour Dataflash2000)
our darker (lower ‘L’ value; Pb0.05)housed compared with TS.of 38.2, 36.1 and 35.2 for TS, SF
respectively.
Instrumental (as for Jensen andOksama)
er (subjective assessment)ental assessmentrence in ‘L’ value; 56.4 (IC) v) (rsd=3.5)higher ‘a’ and ‘b’ (both Pb0.001)
Subjective (scale 1 to 3; 1 = white;2 = pink; 3 = red)Instrumental (Minolta Chromameter,model CR100)

15P.G. Dunne et al. / Livestock Science 142 (2011) 1–22
byphysical activity. Vestergaard et al. (1997)proposed that thiswas due to the fact that the SU muscle is considered a ‘red’(oxidative) muscle whereas LD is considered to be a ‘mixed’ or‘white’ muscle and so has inherently less oxidative character.Based on this rationale, itwould be expected that stimuliwhichwould/could lead to a greater degree of oxidative character, e.g.physical activity, would have a greater impact on ‘white’muscle, so that a putative effect of physical activity on musclecolour is likely to be muscle-dependent.
Vestergaard et al. (2000) tested this concept in an experi-ment which followed a 2 (production systems – extensive andintensive)×2 (body weight (BW) – 360 kg and 460 kg)factorial arrangement and observed that with regard to colourvariables (‘L’, ‘a’, ‘b’, ‘H’ and ‘C’) and pigmentation no significantdifferences emerged between production system at either BWfor M. semitendinosus (ST) or SU. However, for all muscles, atboth BW, there was a tendency for ‘L’ values and ‘a’ values (LDmuscle only) to be lower for extensive relative to intensivebulls. The LD and ST muscles of extensive bulls tended to havehigher levels of pigmentation at both BWbut this effectwas notseen in the SU muscle.
Overall, significant effects were only observed in the LDmuscle but therewas a strong tendency for similar effects in theST muscle. The SU muscle remained largely unaffected withregard to pigmentation or colour variables. This highlightedthe differential response of muscles to the productionsystem. Vestergaard et al. (2000) speculated that differencesin physical activity could explain the colour differencesbetweenextensive and intensive bulls aswell as thedifferentialresponse of muscles.
Dunne et al. (2005) exercised steers daily over a distance of4.41 km for 18 weeks to determine whether steers that wereexercised produced darker muscle than non-exercised steersandwhether any effectwasmuscle-related; steerswere offereda common diet. They reported that muscle lightness was notaffected by exercise but that redness was, although the effectwas muscle-dependent (muscle×exercise, P=0.038). Whilstexercise tended to increase theM. extensor carpi radialis (ECR; aforeleg muscle) redness the opposite effect was observed withLD and SMmuscles. Muscle lightness was not affected.
Dunne et al. (2005) postulated that the differentialresponse of muscle surface colour to exercise may havebeen related to the depth of the oxymyoglobin layer. The ECRmuscle had a significantly (Pb0.001) higher proportion ofoxymyoglobin than either the LD or SM; the authors statedthat a putative increase in the respiratory enzyme capacity ofLD and SMmuscles with exercise, indicated by a tendency forhaem pigments to increase, may have lead to increasedcompetition for dissolved oxygen at the muscle cut surface.Thus, a thinner oxymyoglobin layer may have formed and alower ‘a’ value was measured. However, since the ECRmusclewas likely more oxidative in nature than either the LD or SMregardless of exercise, when exercise was applied itsresponse, in terms of oxidative enzyme capacity, may havebeen relatively less. This would mean that any potentialdisruptive influence of oxidative enzymes on oxygenation ofmyoglobin during blooming of ECR muscle was diminished.However, although not darker the response of LD redness toexercise in the study of Dunne et al. (2005) mirrors theresponse of the LD to grazing in other studies (Hedrick et al.,1983; Muir et al., 1998b).
6. Relationship between muscle colour and associatedbiochemical characteristics: effects of exercise/activity onbiochemical characteristics and colour
Although her work was conducted on rats, Gauthier(1969) concluded that there was a clear relationship betweenthe colour of the ST muscle and cytological features of itscomponent fibres. This was sufficient to justify use of theterms ‘red’, ‘white’ and ‘intermediate’ as simple descriptionsfor fibre types in mammalian skeletal muscle. Furthermore,measurement of the cross-sectional area contributed byeach fibre type to the total area indicated that both red andintermediate fibres may contribute to redness in mammalianskeletal muscle. The fibre diameter of ‘red’muscle fibres types(βR or ‘slow-oxidative’ (SO) and αR or ‘fast-oxidative-glycolytic’ (FOG) (Greaser et al., 2001)) is positively corre-lated with the ‘a’ value in LD muscle of Japanese Black steers(Ozawa et al., 2000). Gil et al. (2001) reported that haempigments were significantly positively correlated with ‘a’and ‘C’ values in five and four of the seven breeds studied,respectively.
Red muscle fibres tend to be relatively narrow and are so-called because they contain abundant myoglobin and mito-chondria, are extensively capillarised and have a large com-plementof respiratory enzymes (Lawrie, 1998b). Thus, they areequipped to perform oxidative metabolism whereas so-called‘white’ fibres are better equipped to perform glycolyticmetabolism (Cassens and Cooper, 1971) since they tend to bebroad, myoglobin-poor fibres, with only small amounts ofmitochondria and associated respiratory enzymes, with agreater activity of lactate dehydrogenase (LDH) (Lawrie,1998b).
White fibres have a greater buffering capacity than redfibres due to higher concentrations of the dipeptides carnosineand anserine. Rao and Gault (1989) reported that, in general,whitefibre-typemuscles had significantly (Pb0.05) higher acidbuffering capacities whereas red fibre-type muscles hadsignificantly (Pb0.05) higher total pigment concentrationsand pHu values. The observation that white fibres had higheracid buffering capacities seems plausible since a reliance onglycolytic metabolism would be expected to lead to increasedconcentrations of lactic acid in vivo which would require to bebuffered. This phenomenon would have implications for post-mortem pH attainment. Consequently, changes in muscle fibretype composition are usually reflected by concomitant changesin associated grossmuscle biochemical features suchasenzymeactivity and pH.
This metabolic differentiation of different muscle types ispossible at the level of enzymatic organisation (Pette, 1971).Gil et al. (2001) reported that the haem pigments, indicativeof oxidative metabolism and responsible for the red colour ofbeef, were significantly positively correlated with isocitratedehydrogenase in three of the seven breeds studied butsignificantly negatively correlated with LDH in four of theseven breeds. “The metabolic potential of a muscle is theexpression of an adjustment to the functional characteristicsof the muscle such as the quality and temporal pattern ofenergy expenditure” (Pette, 1971), i.e. whether the muscle isinvolved in steady and continuous activity, steady anddiscontinuous activity or is engaged in only discontinuousperformance of work.

16 P.G. Dunne et al. / Livestock Science 142 (2011) 1–22
In functional terms, muscles can be broadly categorised aseither postural or locomotive. This functional categorisationcoincides very well with the anatomical location andmetabolic profile of muscles. Muscles which are used tomaintain posture, or which are otherwise used frequently bythe animal, have a high proportion of ‘red’ fibres and are thusadapted for sustained activity, whilst muscles which arerelatively inactive have a high proportion of ‘white’ fibres(Ashmore et al., 1972b). Red muscles are thought to beinvolved in activities of a low-intensity, prolonged-durationnature whereas white muscles are thought to be involvedprimarily in high-intensity, short-duration types of exercise(Barnard et al., 1970). Muscles located along or adjacent tothe vertebral column have a dominant postural function, e.g.LD, whilst limb muscles are involved to a greater extent inlocomotion, e.g. M. biceps femoris (BF), SM, ST (hindlimbmuscles), SU or ECR (forelimb muscles), although largehindquarter muscles such as BF also play a role in mainte-nance of posture (Brocks et al., 2000) and thus can display aduality of function.
Muscle tissue has the ability to adapt to prolonged,submaximal or low intensity physical activity, such as cattleengaged in grazing undertake, by becoming more oxidative(Holloszy et al., 1971). This type or pattern of activityconstitutes what is referred to in the literature as ‘enduranceactivity’. Biochemical adaptations resulting from exposure to‘endurance exercise’ can result in as much as a two-foldincrease in the capacity of skeletal muscle for aerobicmetabolism (Holloszy et al., 1971). Both Jurie et al. (2006)and Vestergaard et al. (2000) reported increased activity ofcitrate synthase and 3-hydroxyacyl-CoA dehydrogenase, bothindicators of oxidative metabolism, and decreased activity ofLDH, an indicator of glycolytic metabolism, in differentmusclesfrom extensively-managed cattle, implying a somewhat con-sistent effect of ‘exercise’, ‘physical activity’ or ‘grazingmobility’across muscles, although to different extents. Hence, thecontention that muscles which have an inherently highoxidative character will be less affected by physical activity ifthe effect of this activity is merely to increase oxidativecharacter (Dunne et al., 2005).
This is corroborated by the findings of Aalhus and Price(1991). They found that muscles such as the M. psoas major(PM) hadmore SO fibres [23.3±2.74% (control group); 25.9±2.62% (exercised group)] thanmuscleswith amajor propulsivefunction such as the ST [14.1±1.94% (control group); 21.6±2.05% (exercised group)]. Exercise increased the percentage ofSOfibres in allmuscles studied except the PM, expected to havean inherently high oxidative character. This agrees with thefindings of Vestergaard et al. (2000) who found that the SUmuscle, primarily involved in forelimbmovementand alsowithan oxidative character,was less affected by the physical activityof grazing than the LD or ST.
However, in the experiment used by Aalhus and Price(1991) the ram lambs were exposed to steady, continuousexercise on a treadmill whereas the cattle used by Vestergaardet al. (2000) did actually graze on pasture. Therefore, the issueof the intensity and duration of exercise is relevant. This isprobably a reflection of whether exercise is imposed or volun-tary although the exercise model used by Aalhus and Price(1991) probably approximates the actual exercise in a grazingsituation with steady, continuous but low intensity activity.
Petersen et al. (1998) stated that exercise training studies maynot adequately replicate the effects of spontaneous exerciseoccurring in outdoor or free-range production systems.
7. Effects of physical activity on muscle glycogenconcentrations, pH and their potential effect onmuscle colour
The ultimate pH of meat is also critically important inrelation tomeat colour. ThepHofmeat affects its light scatteringproperties (MacDougall, 1982). The pH of muscle drops in thepost-mortem condition due to the accumulation of lactic acidresulting from anaerobic glycolysis. Lactic acid is ultimatelyproduced from muscle stores of glycogen. The glycogenconcentration at slaughter must exceed 1.03 mg/100 g muscleso that sufficient lactic acid is produced to reduce muscle pH toabout 5.5 (Tarrant, 1989).
Vestergaard et al. (1997) and Dufrasne et al. (1995) bothobserved that grazing bulls had darker coloured meat and bothacknowledged that the higher pH of the LD muscle could havecontributed to this. However, althoughVestergaard et al. (1997)found that the grazing bulls had higher levels of pigments in allmuscles studied, Dufrasne et al. (1995) did not— there were nodifferences in pigment content between grazed and indoorfinished bulls. This observation demonstrates the importance ofpH in relation to meat colour, since muscle of grazing bulls wasdarker yet had the same level of pigment as indoor bulls. Dalyet al. (1999) claimed that pasture-reared cattle may be moresusceptible to muscle glycogen depletion than feedlot-rearedcattle, proposing that this resulted from an increased sensitivityof pasture-reared cattle relative to feedlot cattle to the environ-mental stressors invariably present during transport andhandling prior to slaughter.
It can be hypothesised that if grazing animals exhibit highermuscle pH, it is caused either by (a) an acute shortage ofmuscleglycogen, insufficient to facilitate adequate pH drop, or perhaps(b) a lower activity of one of the glycogenolytic enzymes orglycogen synthetase. A plausible counter argument is that thehigher pH of grazing cattle results from a generally increasedoxidative potential across muscles, so that live muscles are lessreliant onglycogen and thus, less is available in thepost-mortemstate, regardless of post-mortem muscle conditions. The firstpresumption is also countered by anomalous observations inthe literature whereby exercise is reported to increase muscleglycogen stores and a concomitant increase in glycogensynthetase activity occurs (Piehl et al., 1974) both of whichmight therefore be expected in grazing animals as a result of theengagement in physical activity. However, unlike the observa-tions of Vestergaard et al. (1997) and Dufrasne et al. (1995) theend result of such a glycogen increase in grazing animalswouldbe a lowering of muscle pH post-mortem. Possibly, any positiveor beneficial effects of exercise with regard to activationof muscle glycogen stores of grazing animals may be negatedby the relatively low plane of nutrition in comparison toconcentrate-fed animals.
Pethick and Rowe (1996) found that feeding level andexercise significantly affected muscle glycogen levels in SMand ST of wethers. Increased feeding level increased muscleglycogen in SM and ST of sedentary and exercised wethers,both at biopsy and 10 min. post-mortem. Exercise resulted inlower levels of muscle glycogen at biopsy relative to

17P.G. Dunne et al. / Livestock Science 142 (2011) 1–22
sedentary wethers but caused an elevated level in muscles ofexercised wethers at 10 min. post-mortem although this wasmore apparent in SM than in ST. In this respect, because thepresence of muscle glycogen is more critical in the post-mortem condition from a meat quality perspective, it could beproposed that exercise had a positive effect, albeit muscle-specific. This contention is reinforced by the finding thatexercise training caused a significant reduction in pH of theST, although not in the SM or the LD. Anomalies arise in thestudy of Vestergaard et al. (2000) whereby at 460 kg theextensively-reared bulls displayed the highest meat pH (5.51v 5.46; S.E.D.=0.066) despite having the highest muscleglycogen level (62.5 μmol/g v 51.8 μmol/g; S.E.D.=1.6).
Furtherdifferences emergebetween the studyofVestergaardet al. (2000) and the studyof Pethick andRowe (1996) since theformer authors found that in ST and LDmuscles of bulls, muscleglycogen levels were higher in extensive than in intensivebulls at biopsy in the week prior to slaughter whereas Pethickand Rowe (1996) had found that exercise resulted in lowerbiopsy glycogen levels. Comparing these data assumes thatexercise was the factor driving differences in glycogen levelsbetween extensive and intensive bulls. Itwould be expected thatextensive bulls would engage in more activity and they mightalso have a lower plane of nutrition than intensive bulls andwould consequently have lower levels of muscle glycogen invivo. Dunne et al. (2005) reported that exercised steers hadsignificantly lower glycogen and glycolytic potential than non-exercised steers.
The occurrence of pre-slaughter stress in bovines and itsimpact on initial meat colour should not be overlooked in thiscontext. The phenomenon known as ‘dark-cutting’ (DC) or‘dark, firm and dry’ (DFD) beef is caused by muscle glycogendeficiency, usually resulting from glycogenolysis in skeletalmuscle induced by physiological stress, accompanied byactivity and “emotional arousal in response to a novel orthreatening situation” (McVeigh and Tarrant, 1982), e.g.herding and transport of pasture-fed cattle, prior to slaughter,which had had low exposure to human contact and activity(Daly et al., 1999). Physical activity or exercise is usuallyassociated with muscle glycogen depletion. However, thisphenomenon usually relates to physical activity engaged inby the animal in the final 24 h ante-mortem. Therefore, in thecontext of effects of physical activity/exercise on muscleglycogen concentrations, it is necessary to clearly distinguishbetween an ‘acute exercise event’, perhaps also stressful,immediately or shortly before slaughter and chronic orlonger-term exercise, of low intensity and sustained over aperiod of perhaps weeks or months, e.g. grazing mobility.
8. Putative combined effect on muscle colour of increasedpH and enhanced oxidative potential both caused byphysical activity
Even if the previous argument regarding a putativeprotective effect of physical activity on glycogen has somerelevance tomeat colour, theobservation thatmeat colour fromgrazing cattle is ‘darker’may be explained more easily, but in amanner still dependent on higher levels or markedly differentpatterns of activity by grazing cattle. However, this requiresthat the phenomenon of ‘darker’ colour consistently perceived
in pasture-fed beef is simply regarded as a less extreme form ofDC beef.
All beef will appear ‘dark’ if it is cut within 24 h ofslaughter, usually because the muscle has not reached itsultimate pH (Murray, 1995) and thus, the change from thedark, jelly-like appearance to a more opaque and paler colourhas not occurred (MacDougall, 1982). Ashmore et al. (1972a)stated that the DFD phenomenon in beef was primarily a cutsurface phenomenon and that unlike normal beef, DFD beefsimply did not brighten upon exposure to air. Ashmore et al.(1972a) reported that the failure of a bright-red colour todevelop was due to the maintenance of a relatively highoxygen consumption rate (OCR) at the cut surface whereoxygen is available, thereby competing for available oxygenand maintaining the myoglobin in the reduced rather thanthe oxygenated state. Further, this enhanced OCR was due toenhancedmitochondrial activity which was in turn facilitatedby the high pH. Beef from extensively-managed or pasture-fed cattle has been shown to display a higher pH and it is alsolikely that physical activity engaged in by such cattle wouldcause enhanced oxidative capacity and associated mitochon-drial activity. In pasture-fed beef, failure of pigments tocompletely oxygenate would also account for less red beefwhen measured instrumentally. In such a situation it is alsoconceivable that the colour would not depend absolutely onthe pigment content of the beef. Given the recognisedrelationship between meat colour and pH, the above aspectsof glycogen metabolism, their relation to DC beef and theirpossible links to the phenomenon of darker colour in pasture-fed or grazing cattle have been considered. However, it is alsoimportant to emphasise that any effect of pH or a tendencytowards the DC condition may only have a marginal influenceon darker pasture-fed beef and may only be relevant inextremis, e.g., where eccentric stressful ante-mortem situa-tions are encountered.
9. Summary and conclusions
Many factors contribute to the eventual colour of bovinemuscle, once it is converted to beef and its colour measured.Thesemay include exercise; exercise is frequently referred to instandard meat quality texts as contributing to darker colour.Exercise in cattle has not yet been shown to cause ‘darker’bovine muscle colour under experimental conditions designedto achieve this. However, limited data available in the literaturesuggest that ‘exercise’ or ‘physical activity’ or ‘grazing mobility’may be a contributory factor influencing beef colour. Manyearlier experiments comparing forage and grain-fed beef colourwere confounded either by differences in age at slaughter or incarcass weight and fatness. Hence, it is not clear whetherobserved colour differences between pasture- and grain-fedcattleweredue todifferences in age at slaughter, carcassweight,fatness, marbling or different activity levels. In such cases, it hasproved impossible to isolate any effect that activity levels mayhave had on muscle colour, although an effect is suggestedstrongly, but by no means conclusively. Another importantfactor to bear inmindmaybe the pattern of energy expenditureand energy intake during the life of the animal and any possibleasynchrony between them. Innovative studies are required toexamine these factorswhilst not confoundingmeasurements ofmeat colour with age or carcass characteristics.

18 P.G. Dunne et al. / Livestock Science 142 (2011) 1–22
What has been called ‘sustained, submaximal exercise’generally causes an increase in myoglobin levels and a shift inmuscle metabolic potential towards oxidative metabolism.Enhancedmuscle oxidative andmitochondrial activity result-ing from extensive management practises may also increaseOCR at the cut surface of pasture-fed beef, thus making itappear darker, although this remains to be experimentallyconfirmed.
It is important to bear inmind that ‘colour’ of bovinemuscleonly becomes an issuewhen a cut surface has been established.Therefore, in this review emphasis has been placed onmetabolic tendencies of the livemuscle and thus, by extension,those of the beef, i.e. themuscle when cut. These tendencies, asdiscussed, can be affected by exercise which in turn may affecthow the cut surface interactswith its environment (e.g. oxygenpartial pressure, light, temperature) to create the appearanceattributes of the beef. Given that much recent work hasdemonstrated that pasture-fed beef canbepotentially healthierthan beef from intensively-reared, concentrate-fed cattle(related to the fatty acid and antioxidant content; reviewedby Daley et al. (2010)), ensuring consistent and acceptablecolour of such ‘extensive’, ‘free range’, ‘pasture-fed’ or ‘grass-fed’ beef should also remain a priority.
In summary, the following points can be made.
(a) Although beef from extensively-managed cattle isoften described as ‘darker’, (i) many factors, includingexercise, may contribute and (ii) differences maydepend on how colour is assessed.
(b) Diet composition does not appear to play an importantrole in muscle colour of adult bovines but this requiresconfirmatory studies (although it is important indetermining colour stability, which relates to shelf-life (Geay et al., 2001) and is a separate issue).
(c) The pattern, duration and intensity of exercise oractivity may be important in effecting physiologicalchanges in muscle which manifest post-mortem asdarker meat. The exercise model used by Dunne et al.(2005), where intensity was indexed by distancewalked and time required to do so, did not causedarker beef. However, studies with younger animals oranimals varying widely in age may be required.
(d) Pasture-fed cattle may be more susceptible to dark-cutting (DC) due to (i) a lower ME intake and (ii) lessexposure and thus, less habituation to human contactand handling so that in situations where a certainstress level is unavoidably encountered such as duringherding and transport prior to slaughter, there isinsufficient muscle glycogen to compensate for thestress-induced glycolysis. However, the potential effectof DC and pH differences should not be overstated andis likely not as important as differences in carcassweight and fatness, reflecting differences in age atslaughter and growth rate, which can easily confoundobserved colour differences and limit interpretativeutility.
Acknowledgements
The authors acknowledge and greatly appreciate theassistance of Ms. Mary Weldon in typing this manuscript.
References
Aalhus, J.L., Price, M.A., 1991. Endurance-exercised growing sheep: I. Post-mortem and histological changes in skeletal muscles. Meat Sci. 29,43–56.
Aalhus, J.L., Robertson, W.M., Ye, J., 2009. Muscle fibre characteristics and theirrelation to meat quality. In: Du, M., McCormick, R.J. (Eds.), Applied MuscleBiology and Meat Science. CRC Press, Boca Raton, FL, USA, pp. 97–114.
Ådnøy, T., Haug, A., Sørheim, O., Thomassen, M.S., Varszegi, Z., Eik, L.O., 2005.Grazing on mountain pastures — does it affect meat quality in lambs?Livest. Prod. Sci. 94, 25–31.
Agboola, H.A., Cahill, V.R., Conrad, H.R., Ockerman, H.W., Parker, C.F., Parrett,N.A., Long, A.R., 1990. The effects of individual and combined feeding ofhigh monosodium phosphate and alpha-tocopherol supplemented milkreplacer diets and an alternate protein diet on muscle color, compositionand cholesterol content of veal. J. Anim. Sci. 68, 117–127.
Agouridis, C.T., Stombaugh, T.S., Workman, S.R., Koostra, B.K., Edwards, D.R.,Vanzant, E.S., 2004. Suitability of a GPS collar for grazing studies. Proc.Am. Soc. Agric. Eng. 47, 1321–1329.
American Meat Science Association, 1991. Guidelines for meat colourevaluation. Proceedings of the 44th Reciprocal Meat Conference, KansasState University, Manhattan, June 9–12, pp. 3–17.
Andersen, H.R., Jensen, L.R., Munksgaard, L., Ingvartsen, K.L., 1997. Influenceof floor space allowance and access sites to feed trough on theproduction of calves and young bulls and on the carcass and meatquality of young bulls. Acta Agric. Scand. Section A, Anim. Sci. 47, 48–56.
Anderson, D.M., Kothmann, M.M., 1977. Monitoring animal travel withdigital pedometers. J. Range Manag. 30, 316–317.
Anderson, D.M., Kothmann, M.M., 1980. Relationship of distance travelledwith diet andweather for Hereford heifers. J. Range. Manag. 33, 217–220.
Andrighetto, I., Gottardo, F., Andreoli, D., Cozzi, G., 1999. Effect of type ofhousing on veal calf growth performance, behaviour and meat quality.Livest. Prod. Sci. 57, 137–145.
Anon, 2000. Ungtyreproduktion på graes eller stald (Steer production onpasture or in housing). Nutr. Abstr. Rev. 70, 559.
Ashmore, C.R., Parker,W., Doerr, L., 1972a. Respiration of mitochondria isolatedfrom dark-cutting beef: postmortem changes. J. Anim. Sci. 34, 46–48.
Ashmore, C.R., Tompkins, G., Doerr, L., 1972b. Postnatal development ofmuscle fiber types in domestic animals. J. Anim. Sci. 34, 37–41.
Bailey, D.W., 2001. Evaluating new approaches to improve livestock grazingdistribution usingGPS andGIS technology. Proc. 1stNational ConferenceonGrazing Lands, December 5–8, 2000, Las Vegas, Nevada, US, pp. 91–99.
Bailey, D.W., 2004. Management strategies for optimal grazing distributionand use of arid rangelands. J. Anim. Sci. 82, E147–E153.
Balcaen, A., Raes, K., Claeys, E., DeSmet, S., 2002. Influence of finishingregimen on meat colour, colour and lipid stability in Belgian Blue bulls.In: Durand, J.L., Emile, J.C., Huyghe, C., Lemaire, G. (Eds.), EuropeanGrassland Federation — Multi-function Grasslands: Quality Forages,Animal Products and Landscapes, La Rochelle, France, p. 546.
Barker, B.P., Mies, W.L., Turner, J.W., Lunt, D.K., Smith, S.B., 1995. Influence ofproduction system on carcass characteristics of F1 Wagyu×Angus steersand heifers. Meat Sci. 41, 1–5.
Barnard,R.J., Edgerton,V.R., Peter, J.B., 1970.Effect of exerciseon skeletalmuscle.I. Biochemical and histochemical properties. J. Appl. Physiol. 28, 762–766.
Bekhit, A.E.D., Faustman, C., 2005.Metmyoglobin reducing activity.Meat Sci. 71,407–439.
Bennett, L.L., Hammond, A.C., Williams, M.J., Kunkle, W.E., Johnson, D.D.,Preston, R.L., Miller, M.F., 1995. Performance, carcass yield and carcassquality characteristics of steers finished on rhizome peanut (Arachisglabrata)-tropical grass pasture or concentrate. J. Anim. Sci. 73, 1881–1887.
Bidner, T.D., Schupp, A.R., Montgomery, R.E., Carpenter Jr., J.C., 1981.Acceptability of beef finished on all-forage, forage-plus-grain or highenergy diets. J. Anim. Sci. 53, 1181–1187.
Bidner, T.D., Schupp, A.R., Mohamad, A.B., Rumore, N.C., Montgomery, R.E.,Bagley, C.P., McMillin, K.W., 1986. Acceptability of beef from Angus-Hereford or Angus-Hereford-Brahman steers finished on all forage or ahigh energy diet. J. Anim. Sci. 62, 381–387.
Blanco, M., Casasús, I., Ripoll, G., Panea, B., Albertí, P., Joy, M., 2010. Lucernegrazing compared with concentrate-feeding slightly modifies carcaseand meat quality of young bulls. Meat Sci. 84, 545–552.
Boccard, R.L., Naude, R.T., Cronje, D.E., Smit, M.C., Venter, H.J., Rossouw, E.J.,1979. The influence of age, sex and breed of cattle on their musclecharacteristics. Meat Sci. 3, 261–280.
Boccard, R., Buchter, L., Casteels, E., Cosentino, E., Dransfield, E., Hood, D.E.,Joseph, R.L., MacDougall, D.B., Rhodes, D.N., Schon, I., Tinbergen, B.J.,Touraille, C., 1981. Procedures for measuring meat quality characteristicsin beef production experiments. Report of a working group in theCommission of the European Communities (CEC) Beef ProductionProgramme. Livest. Prod. Sci. 8, 385–397.

19P.G. Dunne et al. / Livestock Science 142 (2011) 1–22
Booth, F.W., Neufer, P.D., 2005. Exercise controls gene expression. Am. Sci. 93,28–35.
Bowman, B.G., Wright, I.A., 1995. Description of the UK beef industry, structureand production systems. Extensification of beef and sheep production ongrassland. Occasional publication No.2. Proceedings of a workshop held inParis, 22–24 November for Concerted Action AIR 3-CT93-0947.
Brehme, U., Stollberg, U., Holz, R., Schleusener, T., 2008. ALT-pedometer —new sensor-aided measurement system for improvement in oestrusdetection. Comp. Electron. Agric. 62, 73–80.
Brocks, L., Klont, R.E., Buist, W., deGreef, K., Tieman, M., Engel, B., 2000. Theeffects of selection of pigs on growth rate vs leanness on histochemicalcharacteristics of different muscles. J. Anim. Sci. 78, 1247–1254.
Broom, D.M., Johnson, K.G., 1993. Stress and Animal Welfare. Chapman &Hall, London, p. 116.
Campbell, W.G., Gordon, S.E., Carlson, C.J., Pattison, J.S., Hamilton, M.T., Booth,F.W., 2001. Differential global gene expression in red and white skeletalmuscle. Am. J. Physiol. Cell Physiol. 280, C763–C768.
Cassens, R.G., Cooper, C.C., 1971. Red andwhitemuscle. Adv. FoodRes. 19, 1–74.Choi, Y.M., Kim, B.C., 2009. Muscle fibre characteristics, myofibrillar protein
isoforms, and meat quality. Livest. Sci. 122, 105–118.Clark, D.A., Kanneganti, V.R., 1998. Grazing management systems for dairy
cows. In: Cherney, J.H., Cherney, D.J.R. (Eds.), Grass for Dairy Cattle. CABIPublishing, Wallingford, Oxon, UK, pp. 311–334.
Clarke, S.D., 2000. Polyunsaturated fatty acid regulation of gene transcrip-tion: a mechanism to improve energy balance and insulin resistance. Br.J. Nutr. 83, S59–S66.
Cornforth, D., 1994. Colour — its basis and importance. In: Pearson, A.M.,Dutson, T.R. (Eds.), Quality Attributes and their Measurement in Meat,Poultry and Fish Products. Advances in Meat Research Series, Volume 9.Chapman & Hall, London, pp. 34–78.
Craig, H.B., Blumer, T.M., Barrick, E.R., 1959. Effect of several combinations ofgrass and grain in the ration of beef steers on the colour characteristics oflean and fat. J. Anim. Sci. 18, 241–248.
Craig, H.B., Blumer, T.N., Smart Jr., W.W.G., Wise, M.B., 1966. Evaluation ofhaemoglobin, myoglobin, blood oxygen content and organo lepticqualities of beef from steers fed grain on pasture or cut forage andgrain in drylot. J. Anim. Sci. 25, 1128–1132.
Crouse, J.D., Cross, H.R., Seideman, S.C., 1984. Effect of a grass or grain diet onthe quality of three beef muscles. J. Anim. Sci. 58, 619–625.
Daley, C.A., Abbott, A., Doyle, P.S., Nader, G.A., Larson, S., 2010. A review offatty acid profiles and antioxidant content in grass-fed and grain-fedbeef. Nutr. J. 9, 10 Available at http://www.nutritionj.com/content/pdf/1475-2891-9-10.pdf accessed April 28, 2010.
Daly, C.C., Young, O.A., Graafhuis, A.E., Moorhead, S.M., Easton, H.S., 1999. Someeffects of diet on beefmeat and fat attributes. N.Z. J. Agric. Res. 42, 279–287.
DelCampo, M., Brito, G., Saores de Lima, J.M., Vaz Martins, D., Sañudo, C., SanJulián, R., Hernández, P., Montossi, F., 2008. Effects of feeding strategiesincluding different proportions of pasture and concentrate, on carcassand meat quality traits in Uruguayan steers. Meat Sci. 80, 753–760.
DiMarco, O.N., Aello, M.S., 1998. Energy cost of cattle walking on the level andon a gradient. J. Range Manag. 51, 9–13.
Dinius, D.A., Cross, H.R., 1978. Feedlot performance, carcass characteristicsand meat palatability of steers fed concentrate for short periods. J. Anim.Sci. 47, 1109–1113.
Drennan, M.J., McGee, M., 2009. Performance of spring-calving beef sucklercows and their progeny to slaughter on intensive and extensivegrassland management systems. Livest. Sci. 120, 1–12.
Drennan, M.J., Keane, M.G., Dunne, L., 1995. Outline of beef production inIreland. Extensification of Beef and Sheep Production on Grassland:Occasional Publication No.2 of Concerted Action AIR3-CT93-0947, Paris,France, p. 87.
Duckett, S.K., Neel, J.P.S., Sonon Jr., R.N., Fontenot, J.P., Clapham, W.P., Scaglia,G., 2007. Effect of winter stocker growth rate and finisher system on: II.Ninth-tenth-eleventh rib composition, muscle colour and palatability.J. Anim. Sci. 85, 2691–2698.
Dufrasne, I., Gielen, M., Limbourg, P., van Eenaeme, C., Istasse, L., 1995. Effectsof a grazing period on performance of finishing bulls: comparison withan indoor finishing system. Anim. Sci. 60, 75–80.
Dunne, P.G. 2003. Colour of bovine adipose and muscle tissue: impact ofanimal type and management factors. PhD dissertation, NationalUniversity of Ireland, University College Dublin, 395 pp.
Dunne, P.G., O'Mara, F.P., Monahan, F.J., French, P., Moloney, A.P., 2005.Colour of muscle of 18-month-old steers given long term daily exercise.Meat Sci. 71, 219–229.
Dunne, P.G., O'Mara, F.P., Monahan, F.J., Moloney, A.P. 2006. Changes incolour characteristics and pigmentation of subcutaneous adipose tissueand M. longissimus dorsi of heifers fed grass, grass silage or concentrate-based diets. Meat Sci. 74, 231–241.
Dunne, P.G., Rogalski, J., Moreno, T., Monahan, F.J., French, P., Moloney, A.P.,2008. Colour, composition and quality of M. longissimus dorsi and
M. extensor carpi radialis of steers housed on straw or concrete slats oraccommodated outdoors on wood chips. Meat Sci. 79, 700–708.
Edwards, S.A., 2005. Product quality attributes associated with outdoor pigproduction. Livest. Prod. Sci. 94, 5–14.
Essén-Gustavsson, B., 1996. Comparative aspects between species. Skeletalmuscle adaptation with use & disuse Proc. 42nd ICoMST, Lillehammer,Norway.
Faustman, C., Cassens, R.G., 1990. The biochemical basis for discolouration infresh meat: a review. J. Muscle Foods 1, 217–243.
Faustman, C., Chan, W.K.M., Schaefer, D.M., Havens, A., 1998. Beef colourupdate: the role for vitamin E. J. Anim. Sci. 76, 1019–1026.
Fiems, L.O., Boucqué, Ch.V., Cottyn, B.G., DeCampeneere, S., DeBrabander, D.L.,1997. Use of grass and grass silage in comparisonwithmaize silage for bullbeef production. Effects of Extensification on Animal Performance, CarcassCompositionand ProductQuality:Occasional PublicationNo.4of ConcertedAction AIR3-CT93-0947, Melle-Gontrode, Belgium, pp. 108–119.
Francis, F.J., Clydesdale, F.M., 1975. Food Colorimetry: Theory and Applica-tions. AVI, Westport, CT, USA.
Fraser, A.F., Broom, D.M., 1997. Farm Animal Behaviour andWelfare, third ed.CAB International, Wallingford, Oxon, UK, p. 117.
Fraser, M.D., Davies, D.A., Vale, J.E., Nute, G.R., Hallett, K.G., Richardson, R.I.,Wright, I.A., 2009. Performance and meat quality of native andcontinental cross steers grazing improved upland pasture or semi-natural rough grazing. Livest. Sci. 123, 70–82.
French, P., O'Riordan, E.G., Monahan, F.J., Caffrey, P.J., Vidal, M., Mooney, M.T.,Troy, D.J., Moloney, A.P., 2000. Meat quality of steers finished on autumngrass, grass silage or concentrate-based diets. Meat Sci. 56, 173–180.
French, P., O'Riordan, E.G., Monahan, F.J., Caffrey, P., Mooney, M.T., Troy, D.J.,Moloney, A.P., 2001. The eating quality of meat of steers fed grass and/orconcentrates. Meat Sci. 57, 379–386.
Gatellier, P., Mercier, Y., Juin, H., Renerre, M., 2005. Effect of finishing mode(pasture- or mixed-diet) on lipid composition, colour stability and lipidoxidation in meat from Charolais cattle. Meat Sci. 69, 175–186.
Gauthier, G.F., 1969. On the relationship of ultrastructural and cytochemicalfeatures to colour inmammalian skeletal muscle. Z Zellforsch 95, 462–482.
Geay, Y., Bauchart, D., Hocquette, J.F., Culioli, J., 2001. Effect of nutritionalfactors on biochemical, structural and metabolic characterictics ofmuscles in ruminants, consequences on dietetic value and sensorialqualities of meat. Reprod. Nutr. Dev. 41, 1–26.
Gigli, S., Iacurto, M., 1995. Present and future cattle and sheep meatproduction systems in Italy. Extensification of beef and sheep productionon Grassland. Occasional Publication No. 2 of Concerted Action AIR3-CT93-0947, Paris, France, pp. 253–267.
Gil, M., Serra, X., Gispert, M., Àngels Oliver, M., Sañudo, C., Panea, B., Olleta,J.L., Campo, M., Oliván, M., Osoro, K., García-Cachán, M.D., Cruz-Sagredo,R., Izquierdo, M., Espejo, M., Martín, M., Piedrafita, J., 2001. The effect ofbreed-production systems on the myosin heavy chain 1, the biochemicalcharacteristics and the colour variables of Longissimus thoracis fromseven Spanish beef cattle breed. Meat Sci. 58, 181–188.
Gill, C.O., 1996. Extending the storage life of raw chilled meats. Meat Sci. 43,S99–S109.
Govindarajan, S., 1973. Fresh meat colour. CRC Crit. Rev. Food Technol. 4,117–140.
Greaser, M.L., Okochi, H., Sosnicki, A.A., 2001. Role of fibre types in meatquality. Proceedings of the 47th ICoMST, Kraków, Poland, p. 34.
Harrison, A.R., Smith, M.E., Allen, D.M., Hunt, M.C., Kastner, C.L., Kropf, D.H.,1978. Nutritional regime effects on quality and yield characteristics ofbeef. J. Anim. Sci. 47, 383–388.
Hart, R.H., Bissio, J., Samuel, M.J., Waggoner Jr., J.W., 1993. Grazing systems,pasture size, and cattle grazing behaviour, distribution and gains. J. RangeManag. 46, 81–87.
Hassoun, P., 2002. Cattle feeding behaviour at pasture: a methodologyrelated to on farm measurements. Anim. Res. 51, 35–41.
Hays, F.L., 1976. Alp and valley cattle: exercise in cold, hot and highenvironments. Pflügers Archiv – Europ. J. Physiol. 362, 185–192.
Hedrick, H.B., Paterson, J.A., Matches, A.G., Thomas, J.D., Morrow, R.E., Stringer,W.C., Lipsey, R.J., 1983. Carcass and palatability characteristics of beefproduced on pasture, corn silage and corn grain. J. Anim. Sci. 57, 791–801.
Hepworth, K.W., Test, P.S., Hart, R.H., Waggoner Jr., J.W., Smith, M.A., 1991.Grazing systems, stocking rates and cattle behaviour in southeasternWyoming. J. Range Manag. 44, 259–262.
Hocquette, J.F., Ortigues-Marty, I., Pethick, D., Herpin, P., Fernandez, X., 1998.Nutritional and hormonal regulation of energy metabolism in skeletalmuscles of meat producing animals. Livest. Prod. Sci. 56, 115–143.
Holland, J.L., Kronfeld, D.S., Meacham, T.N., 1996. Behaviour of horses is affectedby soy lecithin and corn oil in the diet. J. Anim. Sci. 74, 1252–1255.
Holloszy, J.O., Oscai, L.B., Molé, P.A., Don, I.J., 1971. Biochemical adaptations toendurance exercise in skeletal muscle. In: Pernow, B., Saltin, B. (Eds.),Muscle Metabolism during Exercise. Advances in Experimental Medicineand Biology, Volume 11. Plenum Press, New York, USA, pp. 51–61.

20 P.G. Dunne et al. / Livestock Science 142 (2011) 1–22
Honikel, K.O., 1998. References methods for the assessment of physicalcharacteristics of meat. Meat Sci. 49, 447–457.
Hornsey, H.C., 1956. The colour of cooked cured pork. I. Estimation of thenitric oxide-haem pigments. J. Sci. Food Agric. 7, 534–540.
Hoving-Bolink, A.H., Hanekamp, W.J.A., Walstra, P., 1999. Effects of dietcarcass, meat and eating quality of once-bred Piedmontese×Friesianheifers. Livest. Prod. Sci. 57, 267–272.
Hunt, M.C., Kropf, D.H., 1987. Colour and appearance. In: Pearson, A.M.,Dutson, T.R. (Eds.), Advances in Meat Research, Volume 3. AVIPublishers, New York, pp. 125–159.
Huuskonen, A., Tuomisto, L., Joki-Tokola, E., Kauppinen, R., 2009. Animalperformance and carcass characteristics of growing Hereford bulls underinsulated, uninsulated and outdoor housing conditions in NorthernFinland. Agric. Food Sci. 18, 16–26.
Huuskonen, A., Jansson, S., Honkavaara, M., Tuomisto, L., Kauppinen, R., Joki-Tokola, E., 2010. Meat colour, fatty acid profile and carcass characteristicsof Hereford bulls finished on grazed pasture or grass silage-based dietswith similar concentrate allowance. Livest. Sci. 131, 125–129.
Iacurto, M., Gaddini, A., Gigli, S., Cerbini, G.M., Ballico, S., DiGiacomo, D., 2005.Comparison of rearing systems at low environmental impact: qualitymeat of beef cattle in central Italy. In: Hocquette, J.F., Gigli, S. (Eds.),Indicators of Milk and Beef Quality. Wageningen Academic Publishers,The Netherlands, pp. 453–458.
Irving, B.D., Rutledge, P.L., Bailey, A.W., Naeth, M.A., Chanasyk, D.S., 1995.Grass utilisation and grazing distribution within intensively managedfields in Central Alberta. J. Range Manag. 48, 358–361.
Jakobsen, M., Bertelsen, G., 2002. Modelling colour stability inmeat. In: Kerry,J., Kerry, J., Ledward, D.A. (Eds.), Meat Processing — Improving Quality.Woodhead Publishing Ltd., Cambridge, UK, pp. 122–136.
Jensen, L.R., Oksama, M., 1996. Influence of different housing systems oncarcass and meat quality in young bulls. Proc. 42nd ICoMST, Lilleham-mer, Norway, p. 436.
Johnston, D.M., Moody, W.G., Boling, J.A., Bradley, N.W., 1981. Influence ofbreed type, sex, feeding systems and muscle bundle size on bovine fibretype characteristics. J. Food Sci. 46, 1760–1764.
Jonsäll, A., Johansson, L., Lundström, K., 2001. Sensory quality and cookingloss of ham muscle (M. biceps femoris) from pigs reared indoors andoutdoors. Meat Sci. 57, 245–250.
Jonsäll, A., Johansson, L., Lundström, K., Andersson, K.H., Nilsen, A.N., Risvik,E., 2002. Effects of genotype and rearing system on sensory character-istics and preference for pork (M. Longissimus dorsi). Food Qual. Pref. 13,73–80.
Jurie, C., Ortigues-Marty, I., Picard, B., Micol, D., Hocquette, J.F., 2006. Theseparate effects of the nature of diet and grazing mobility on metabolicpotential of muscle from Charolais steers. Livest. Sci. 104, 182–192.
Keane, M.G., Allen, P., 1998. Effects of production system intensity onperformance, carcass composition and meat quality of beef cattle. Livest.Prod. Sci. 56, 203–214.
Keane, M.G., Allen, P., 1999. Effects of fertiliser N level on herbagecomposition, animal performance and on carcass and meat qualitytraits. Livest. Prod. Sci. 61, 233–244.
Keane, M.G., Drennan, M.J., 2007. A comparison of Friesian, AberdeenAngus×Friesian and Belgian Blue×Friesian steers finished at pasture orindoors. Livest. Sci. 115, 268–278.
Keane, M.G., O'Riordan, E.G., O'Kiely, P., 2009. Dairy Calf-to-Beef ProductionSystems. Teagasc, Oak Park, Carlow Ireland. 56 pp.
Kennedy, P., Dawson, L., Moss, B.W., Fearon, A., 2009. Meat quality ofbeef cattle offered grass silage and legume/cereal wholecrop silage eitheralone or in combination with grass silage. Proc. Brit. Soc. Anim. Sci. , p. 50.
Klont, R., Brocks, L., Eikelenboom, G., 1998. Muscle fibre type and meatquality. Meat Sci. 49, S219–S229.
Krzywicki, K., 1979. Assessment of relative content of myoglobin, oxymyo-globin and metmyoglobin at the surface of beef. Meat Sci. 3, 1–10.
Lanari, M.C., Brewster, M., Yang, A., Tume, R.K., 2002. Pasture and grainfinishing affect the colour stability of beef. J. Food Sci. 67, 2467–2473.
Lawrie, R.A., 1998a. The eating quality of meat. Meat Science. PermagonPress, Oxford, UK, pp. 212–257.
Lawrie, R.A., 1998b. Chemical and biochemical constitution of muscle. MeatScience. Permagon Press, Oxford, UK, pp. 58–95.
Ledward, D.A., 1984. Haemoproteins in meat and meat products. In: Hudson,B.J.F. (Ed.), Developments in Food Proteins, 3. Elsevier Applied SciencePublishers, London, UK, pp. 33–68.
Ledward, D.A., 1992. Colour of raw and cooked meat. In: Ledward, D.A.,Johnston, D.E., Knight, M.K. (Eds.), The Chemistry of Muscle-based Foods.The Royal Society of Chemistry, Cambridge, UK, pp. 128–144.
Lee, J., 1988. Forages. Livest. Prod. Sci. 19, 13–46.Lee, M.R.F., Evans, P.R., Nute, G.R., Richardson, R.I., Scollan, N.D., 2009. A
comparison between red clover silage and grass silage feeding on fattyacid composition, meat stability and sensory quality of theM. longissimusmuscle of dairy cull cows. Meat Sci. 81, 738–744.
Lefaucheur, L., Gerrard, D., 2000. Muscle fibre plasticity in farm mammals.Proc. Am. Soc. Anim. Sci. 1–19.
Listrat, A., Rakadjiyski, N., Jurie, C., Picard, B., Touraille, C., Geay, Y., 1999.Effect of type of diet on muscle characteristics and meat palatability ofgrowing Salers bulls. Meat Sci. 53, 115–124.
Liu, Q., Lanari, M.C., Schaefer, D.M., 1995. A review of dietary vitamin Esupplementation for improvement of beef quality. J. Anim. Sci. 73,3131–3140.
Livingston, D.J., Brown, W.D., 1982. The chemistry of myoglobin and itsreactions. Food Technol. 35, 244–252.
Maatje, K., Loeffler, S.H., Engel, B., 1997. Predicting optimal time ofinsemination in cows that show visual signs of estrus by estimatingonset of estrus with pedometers. J. Dairy Sci. 80, 1098–1105.
MacDougall, D.B., 1982. Changes in the colour and opacity of meat. FoodChem. 9, 75–88.
MacDougall, D.B., 1986. The chemistry of colour and appearance. Food Chem.21, 283–299.
MacDougall, D.B., 1994. Colour of meat. In: Pearson, A.M., Dutson, T.R. (Eds.),Advances in Meat Research Series, Volume 9. Chapman & Hall, London,pp. 79–93.
MacDougall, D.B., 2002. Colour measurement of food: principles and practice.In: MacDougall, D.B. (Ed.), Colour in Food: Improving Quality. WoodheadPublishing Ltd., Cambridge, UK.
Mahecha, L., Nuernberg, K., Nuernberg, G., Ender, K., Hagemann, E.,Dannenberger, D., 2009. Effects of diet and storage on fatty acid profile,micronutrients and quality of muscle from German Simmental bulls.Meat Sci. 82, 365–371.
Maillet, D., Weber, J.M., 2007. Relationship between n-3PUFA content andenergy metabolism in the flight muscles of a migrating shorebird:evidence for natural doping. J. Exp. Biol. 210, 413–420.
Mancini, R.A., 2009a. Meat colour. In: Du, M., McCormick, R.J. (Eds.),Applied Muscle Biology and Meat Science. CRC Press, Boca Raton, FL,USA, pp. 217–226.
Mancini, R.A., 2009b. Meat colour. In: Kerry, J., Ledward, D.A. (Eds.),Improving the Sensory and Nutritional Quality of Fresh Meat. WoodheadPublishing Ltd., Cambridge, UK.
Mancini, R.A., Hunt, M.C., 2005. Current research in meat colour. Meat Sci. 71,100–121.
Mandell, I.B., Gullett, E.A., Buchanan-Smith, J.G., Campbell, C.P., 1997. Effectsof diet and slaughter endpoint on carcass composition and meat qualityin Charolais cross steers. Can. J. Anim. Sci. 77, 403–413.
McCaughey, W.P., Cliplef, R.L., 1996. Carcass and organoleptic characteristicsof meat from steers grazed on alfalfa/grass pastures and finished ongrain. Can. J. Anim. Sci. 76, 149–152.
McMillin, K.W., 2008. Where is MAP going? A review and future potential ofmodified atmosphere packaging for meat. Meat Sci. 80, 43–65.
McVeigh, J.M., Tarrant, P.V., 1982. Behavioural stress and skeletal muscleglycogen metabolism in young bulls. J. Anim. Sci. 54, 790–795.
Moloney, A.P., Drennan, M.J., O'Neill, B.J., Mooney, M.T., Troy, D.J., 2000a. Fatcolour and the quality of meat from beef cattle offered grass silage orconcentrate-based diets. Ir. J. Agric. Food Res. 39, 152 (abs.).
Moloney, A.P., O'Kiely, P., Vidal, M., Troy, D.J., 2000b. Fat colour and quality ofmeat from heifers offered grass silage or whole-crop silage based rations.Teagasc, Grange Research Centre Research Report, pp. 128–129.
Moloney, A.P., Keane, M.G., Dunne, P.G., Mooney, M.T., Troy, D.J., 2001a.Delayed concentrate feeding in a grass silage/concentrate beef finishingsystem: effects on fat colour and meat quality. Proc. 47th ICoMST,Kraków, Poland, p. 188.
Moloney, A.P., Mooney, M.T., Kerry, J.P., Troy, D.J., 2001b. Producing tenderand flavoursome beef with enhanced nutritional characteristics. Proc.Nutr. Soc. 60, 221–229.
Moloney, A.P., Fallon, R.J., Mooney, M.T., Troy, D.J., 2004. The quality of meatand fatness of bulls offered ad libitum concentrates, indoors or at pasture.Livest. Prod. Sci. 87, 271–276.
Monserrat, L., Varela, A., Sánchez, L., Carballo, J.A., 2001. Extensification oftypical “gallego” calf beef production: effect on growth, carcass and meatcharacteristics. Proc. 52nd Annual Meeting of the European Associationof Animal Production, Budapest, p. 239.
Morrissey, P.A., Sheehy, P.J.A., Galvin, K., Kerry, J.P., Buckley, D.J., 1998. Lipidstability in meat and meat products. Meat Sci. 49, S73–S86.
Muir, P.D., Deaker, J.M., Bown, M.D., 1998a. Effects of forage and grain-basedfeeding systems on beef quality: a review. N.Z. J. Agric. Res. 41, 623–635.
Muir, P.D., Smith, N.B., Wallace, G.J., Cruickshank, G.J., Smith, D.R., 1998b. Theeffect of short-term grain feeding on liveweight gain and beef quality.N.Z. J. Agric. Res. 41, 517–526.
Murray, A.C., 1995. The evaluation of muscle quality. In: Morgan Jones,S.D. (Ed.), Quality and Grading of Carcasses of Meat Animals. CRC Press,pp. 83–108.
Nuernberg, K., Dannenberger, D., Nuernberg, G., Ender, K., Voigt, J., Scollan,N.D., Wood, J.D., Nute, G.R., Richardson, R.I., 2005. Effect of a grass-based

21P.G. Dunne et al. / Livestock Science 142 (2011) 1–22
and a concentrate feeding system on meat quality characteristics andfatty acid composition of longissimus muscle in different cattle breeds.Livest. Prod. Sci. 94, 137–147.
O'Riordan, E.G., O'Kiely, P., 1996. Potential of beef production systems basedon grass. Ir. Grassland Anim. Prod. Assoc. 30, 185–217.
O'Sullivan, M., 1983. Measurement of grazing behaviour and herbage intakeon two different grazing management systems for beef production.Grassland Beef Production: A seminar in the CEC Program of Co-ordination of Research on Beef Production, Martinus Nijhoff, The Hague,Netherlands, pp. 141–150.
O'Sullivan, A., O'Sullivan, K., Galvin, K., Moloney, A.P., Troy, D.J., Kerry, J.P.,2004. Influence of concentrate composition and forage type on retailpackaged beef quality. J. Anim. Sci. 82, 2384–2391.
Oddy, V.H., Harper, G.S., Greenwood, P.L., McDonagh, M.B., 2001. Nutritionaland developmental effects on the intrinsic properties of muscles as theyrelate to the eating quality of beef. Aust. J. Exp. Agric. 41, 921–942.
Ordway, G.A., Garry, D.J., 2004. Myoglobin: an essential hemoprotein instriated muscle. J. Exp. Biol. 207, 3441–3446.
Osuji, P.O., 1974. The physiology of eating and the energy expenditure of theruminant at pasture. J. Range Manag. 27, 437–443.
Owens, F.N., Gardner, B.A., 1999. Ruminant nutrition and meat quality.Proc.52nd Ann. Recip. Meat Conf, pp. 25–36.
Ozawa, S., Mitsuhashi, T., Mitsumoto, M., Matsumoto, S., Itoh, N., Itagaki, K.,Kohno, K., Dohgo, T., 2000. The characteristics of muscle fibre types oflongissimus thoracis muscle and their influences on the quantity andquality of meat form Japanese Black steers. Meat Sci. 54, 65–70.
Ozutsumi, K., Okada, M., 1981. The effect of mountainous grazing on type andcross-sectional area of muscle fibre in Holstein steers. Jap. J. Zootech. Sci.52, 741–748.
Parsons, A.J., Chapman, D.F., 1998. Principles of grass growth and pastureutilisation. In: Cherney, J.H., Cherney, D.J.R. (Eds.), Grass for Dairy Cattle.CABI Publishing, Wallingford, Oxon, UK, pp. 283–309.
Pearson, A.M., 1990. Muscle growth and exercise. Crit. Rev. Food Sci. Nutr. 29,167–196.
Pérez-Alvarez, J.A., Fernández-López, J., 2006. Chemistry and biochemistry ofcolour in muscle foods. In: Hui, Y.H. (Ed.), Food Biochemistry and FoodProcessing. Blackwell Publishing, Oxford, UK, pp. 337–350.
Petersen, J.S., Henckel, P., Oksbjerg, N., Sorensen, M.T., 1998. Adaptations inmuscle fibre characteristics induced by physical activity in pigs. Anim.Sci. 66, 733–740.
Pethick, D.W., Rowe, J.B., 1996. The effect of nutrition and exercise on carcassparameters and the level of glycogen in skeletal muscle of Merino sheep.Aust. J. Agric. Res. 47, 525–537.
Pette, D., 1971. Metabolic differentiation of distinct muscle types at the levelof enzymatic organisation. In: Pernow, B., Saltin, B. (Eds.), MuscleMetabolism during Exercise. Advances in Experimental Medicine andBiology, Volume 11. Plenum Press, New York, pp. 33–49.
Piehl, K., Adolfsson, S., Nazar, K., 1974. Glycogen storage and glycogensynthetase activity in trained and untrained muscle of man. Acta Physiol.Scand. 90, 779.
Pomeranz, Y., Meloan, C.E., 1994. Measurement of colour, Food Analysis:Theory and Practice, 3rd edn. Chapman & Hall, New York, pp. 87–98.
Pommier, S.A., Lapierre, H., de Pasillé, A.M., Gariépy, C., 1995. Control of thebioavailability of iron in heavy veal production by different feedingmanagement systems: use of Ca-EDTA as an iron chelating agent. Can. J.Anim. Sci. 75, 37–44.
Poulson, C.S., Dhiman, T.R., Ure, A.L., Cornforth, D., Olson, K.C., 2004.Conjugated linoleic acid content of beef from cattle fed diets containinghigh grain, CLA or raised on forages. Livest. Prod. Sci. 91, 117–128.
Priolo, A., Micol, D., Agabriel, J., 2001. Effects of grass feeding systems onruminant meat colour and flavour. Anim. Res. 50, 185–200.
Raes, K., Balcaen, A., Dirinck, P., DeWinne, A., Claeys, E., Demeyer, D., DeSmet,S., 2003. Meat quality, fatty acid composition and flavour analysis inBelgian retail beef. Meat Sci. 65, 1237–1246.
Rao, M.V., Gault, N.F.S., 1989. The influence of fibre type composition andassociated biochemical characteristics on the acid buffering capacities ofseveral beef muscles. Meat Sci. 26, 5–18.
Rath, M., Peel, S., 2005. Grassland in Ireland and the UK. In: McGilloway, D.A.(Ed.), Grassland — A Global Resource. Wageningen Academic Publishers,The Netherlands, p. 13.27.
Razminowicz, R.H., Kreuzer, M., Scheeder, M.R.L., 2006. Quality of beef fromtwo grass-based production systems in comparison with conventionalbeef. Meat Sci. 73, 351–361.
Realini, C.E., Duckett, S.K., Brito, G.W., Dalla Rizza, M., DeMattos, D., 2004.Effect of pasture vs. concentrate feeding with or without antioxidants oncarcass characteristics, fatty acid composition, and quality of Uruguayanbeef. Meat Sci. 66, 567–577.
Rimbach, G., Minihane, A.M., Majewicz, J., Fischer, A., Pallauf, J., Virgli, F.,Weinberg, P.D., 2002. Regulation of cell signalling by vitamin E. Proc.Nutr. Soc. 61, 415–425.
Roosevelt, M., 2006. The Grass-fed Revolution. Time Magazine.Sather, A.P., Jones, S.D.M., Schaefer, A.L., Colyn Robertson, W.M., 1997.
Feedlot performance, carcass composition andmeat quality of free-rangereared pigs. Can. J. Anim. Sci. 77, 225–232.
Schaake, S.L., Skelley, G.C., Halpin, E., Grimes, L.W., Brown, R.B., Cross, D.L.,Thompson, C.E., 1993. Carcass and meat sensory traits of steers finishedon fescue and clover, summer forage or for different periods in drylot.J. Anim. Sci. 71, 3199–3205.
Schroeder, J.W., Cramer, D.A., Bowling, R.A., Cook, C.W., 1980. Palatability,shelf-life and chemical differences between forage- and grain-finishedbeef. J. Anim. Sci. 50, 852–859.
Schwarz, F.J., Augustini, C., Kirchgessner, M., 1997. Effect of grazing or indoorfeeding on animal performance and carcass and meat quality ofSimmental or Angus × Simmental heifers. Effects of Extensification onAnimal Performance, Carcass Composition and Product Quality: Occa-sional Publication No.4 of Concerted Action AIR3-CT93-0947, Melle-Gontrode, Belgium, pp. 238–247.
Senft, R.L., Rittenhouse, L.R., Woodmansee, R.G., 1985. Factors influencingpatterns of cattle grazing behaviour on shortgrass steppe. J. RangeManag. 38, 82–87.
Sheppard Jr., J.J., 1968. Human Color Perception. A Critical Study of theExperimental Foundation. Elsevier Applied Science, New York, pp. 33–34.
Shorthose, W.R., Harris, P.V., 1991. Effects of growth and composition onmeat quality. In: Pearson, A.M., Dutson, T.R. (Eds.), Growth Regulation inFarm Animals. Advances in Meat Research, Volume 7. Elsevier AppliedScience, Amsterdam, Netherlands, p. 515.
Solomon, M.B., Lynch, G.P., 1988. Biochemical, histochemical and palatabilitycharacteristics of young ram lambs as affected by diet and electricalstimulation. J. Anim. Sci. 66, 1955–1962.
Solomon, M.B., West, R.L., 1985. Profile of fibre types in muscles from wildpigs native to the United States. Meat Sci. 13, 247–254.
Sowell, B.F., Mosley, J.C., Bowman, J.G.P., 1999. Social behaviour of grazing beefcattle: implications for management. Proc. Amer. Soc. Anim. Sci, pp. 1–5.
Srinivasan, S., Xiong, Y.L., Blanchard, S.P., Moody, W.G., 1998. Proximate,mineral and fatty acid composition of semimembranosus and cardiacmuscles from grass- and grain-fed and zeranol-implanted cattle. FoodChem. 63, 543–547.
Stevenson, J.M., 1990. Colour measurement in meat and meat products.Proc. 26thMeat Industry Research Conference, Hamilton, New Zealand,pp. 306–312.
Strachan, D.B., Yang, A., Dillon, R.D., 1993. Effect of grain-feeding on fat colourand other carcass characteristics in previously grass-fed Bos indicussteers. Aust. J. Exp. Agric. 33, 269–273.
Tarrant, P.V., 1989. Animal behaviour and environment in the dark-cuttingcondition in beef — a review. Ir. J. Food Sci. Technol. 13, 1–21.
Thénard, V., Dumont, R., Grosse, M., Trommenschlager, J.M., Fiorelli, J.L.,Roux, M., 2006. Grass steer production system to improve carcass andmeat quality. Livest. Sci. 105, 185–197.
Therkildsen, M., Vestergaard, M., Jensen, L.R., Andersen, H.R., Sejrsen, K.,1998. Effect of feeding level, grazing and finishing on growth and carcassquality of young Friesian bulls. Acta Agric. Scand. Section A, Anim. Sci. 48,193–201.
Tomkins, N., Filmer, M., 2007. GPS tracking to boost sustainability. FarmingAhead 185, 68–71.
Tuomisto, L., Huuskonen, A., Ahola, L., Kauppinen, R., 2009. Different housingsystems for growing dairy bulls in Northern Finland — effects onperformance, behaviour and immune response. Acta Agric. Scand.Section A, Anim. Sci. 59, 35–47.
Turner, L.W., Udal, M.C., Larson, B.T., Shearer, S.A., 2000. Monitoring cattlebehaviour and pasture use with GPS and GIS. Can. J. Anim. Sci. 80,405–413.
Ungar, E.D., Henkin, Z., Gutman, M., Dolev, A., Genizi, A., Ganskopp, D., 2005.Inference of animal activity from GPSA collar data on free-ranging cattle.Rangel. Ecol. Manag. 58, 256–266.
USDA, 2007. hhttp://www.ams.usda.gov/AMSv1.0/getfile?dDocName=STELPRDC5063842 [Accessed 28 April, 2010].
Varela, A., Oliete, B., Moreno, T., Portela, C., Monserrat, L., Carballo, J.A.,Sánchez, L., 2004. Effect of pasture finishing on the meat characteristicsand intramuscular fatty acid profile of steers of the Rubia Gallega breed.Meat Sci. 67, 515–522.
Varnam, A.H., Sutherland, J.P., 1995. The Colour of Meat. In: Meat and MeatProducts: Technology, Chemistry and Microbiology. Chapman and Hall,London, UK, pp. 26–30. 92–97.
Vestergaard,M., Jensen, L.R., Andersen, H.R., 1997. Influence of housing system,floor space allowance and grazing for young bulls on meat and eatingquality. Effects of Extensification on Animal Performance, Carcass Compo-sition and Product Quality:Occasional PublicationNo.4 of ConcertedActionAIR3-CT93-0947, Melle-Gontrode, Belgium, pp. 268–277.
Vestergaard, M., Oksbjerg, N., Henckel, P., 2000. Influence of feedingintensity, grazing and finishing feeding on muscle fibre characteristics

22 P.G. Dunne et al. / Livestock Science 142 (2011) 1–22
and meat colour of semitendinosus, longissimus dorsi and supraspinatusmuscles of young bulls. Meat Sci. 54, 177–185.
Wackerhage, H., Woods, N.M., 2002. Exercise-induced signal transductionand gene regulation in skeletal muscle. J. Sports Sci. Med. 1, 103–114.
Wahle, K.W.J., Rotondo, D., Heys, S.D., 2003. Polyunsaturated fatty acids andgene expression in mammalian systems. Proc. Nutr. Soc. 62, 349–360.
Walker, J.W., Heitschmidt, R.K., Dowhower, S.L., 1985. Evaluation ofpedometers for measuring distance travelled by cattle on two grazingsystems. J. Range Manag. 38, 90–93.
Wallace, W.J., Houtchens, R.A., Maxwell, J.C., Caughey, S., 1982. Mechanism ofautoxidation for haemoglobins and myoglobins. J. Biol. Chem. 257,4966–4977.
Walsh, K., O'Kiely, P., Moloney, A.P., Boland, T.M., 2007. Intake, performanceand carcass characteristics of beef cattle offered diets based on whole-crop wheat or forage maize relative to grass silage or ad libitumconcentrates. Livest. Sci. 116, 223–236.
Warren, H.E., Scollan, N.D., Nute, G.R., Hughes, S.I., Wood, J.D., Richardson,R.I., 2008. Effects of breed and a concentrate or grass silage diet on beef
quality in cattle of 3 ages. II: meat stability and flavour. Meat Sci. 78,270–278.
Wiklund, E., Malmfors, G., Lundström, K., 1998. The effects of exercise onmuscle fibre composition and oxidative capacity in eight bovine skeletalmuscles. Swed. J. Agric. Res. 28, 111–116.
Woods, V.B., Fearon, A.M., 2009. Dietary sources of unsaturated fatty acids foranimals and their transfer into meat, milk and eggs. Livest. Sci. 126, 1–20.
Xiccato, G., Trocino, A., Queaque, P.I., Sartori, A., Carazzolo, A., 2002. Rearingveal calves with respect to animal welfare: effects of group housing andsolid feed supplementation on growth performance and meat quality.Livest. Prod. Sci. 75, 269–280.
Young, O.A., West, J., 2001. Meat colour. In: Hui, Y.H., Nip, W.K., Rogers, R.W.,Young, O.A. (Eds.), Meat Science & Applications. Markel Dekker, Inc.,New York, pp. 39–70.
Young, O.A., Braggins, T.J., West, J., Lane, G.A., 1999. Animal productionorigins of some meat colour and flavour attributes. In: Xiong, Y.L., Ho,C.T., Shahidi, F. (Eds.), Quality Attributes of Muscle Foods. KluwerAcademic/Plenum Publishers, New York, pp. 11–29.




















