
29, Philadelphia, PA - American Crystallographic Association

Crystallographic Structure of the K Intermediate ofBacteriorhodopsin: Conservation of Free Energy afterPhotoisomerization of the Retinal
Brigitte Schobert1, Jill Cupp-Vickery1, Viktor Hornak2
Steven O. Smith3 and Janos K. Lanyi1*
1Department of Physiologyand Biophysics, University ofCalifornia, Irvine, CA 92697USA
2Center for Structural BiologyState University of New Yorkat Stony Brook, Stony BrookNY 11794, USA
3Department of Biochemistryand Cell BiologyState University of New Yorkat Stony Brook, Stony BrookNY 11794, USA
The K state, an early intermediate of the bacteriorhodopsin photocycle,contains the excess free energy used for light-driven proton transport.The energy gain must reside in or near the photoisomerized retinal,but in what form has long been an open question. We produced the Kintermediate in bacteriorhodopsin crystals in a photostationary state at100 K, with 40% yield, and determined its X-ray diffraction structure to1.43 A resolution. In independent refinements of data from four crystals,the changes are confined mainly to the photoisomerized retinal. Theretinal is 13-cis,15-anti, as known from vibrational spectroscopy. TheC13vC14 bond is rotated nearly fully to cis from the initial trans configu-ration, but the C14–C15 and C15vNZ bonds are partially counter-rotated.This strained geometry keeps the direction of the Schiff base N–H bondvector roughly in the extracellular direction, but the angle of its hydrogenbond with water 402, that connects it to the anionic Asp85 and Asp212, isnot optimal. Weakening of this hydrogen bond may account for many ofthe reported features of the infrared spectrum of K, and for its photoelec-tric signal, as well as the deprotonation of the Schiff base later in thecycle. Importantly, although 13-cis, the retinal does not assume theexpected bent shape of this configuration. Comparison of the calculatedenergy of the increased angle of C12–C13vC14, that allows this distortion,with the earlier reported calorimetric measurement of the enthalpy gainof the K state indicates that a significant part of the excess energy is con-served in the bond strain at C13.
q 2002 Elsevier Science Ltd. All rights reserved
Keywords: bacteriorhodopsin; retinal; K intermediate; photoisomerization;membrane proteins*Corresponding author
Introduction
Light causes the all-trans to 13-cis isomerizationof the retinal of bacteriorhodopsin. During the“photocycle” that ensues, the 13-cis,15-anti isomerreturns thermally to all-trans, and a series of struc-tural changes in the protein result in the trans-location of a proton from the cytoplasmic to theextracellular side of the membrane. The steps inthis sequence consist, approximately, of a linearchain of the reactions of intermediates K, L, M, Nand O. The changes in the retinal and the protein
during the photocycle, and why they cause proton(or perhaps hydroxyl ion) transfers, have beenstudied extensively by various kinds of spectro-scopy, site-specific mutagenesis, and diffraction of2D and 3D crystals. The results have given a fairlydetailed, although still incomplete, description ofthe mechanism of this light-driven pump.1 – 7
A less well understood but interesting questionis how energy is conserved in the K intermediate,i.e. after decay of the excited state of the retinalbut before any proton transfers. From a calori-metric measurement at 77 K, the enthalpy gain inthe K state was reported to be 67(^13) kJ/mol,8 avalue revised later to 49(^14) kJ/mol.9 If theprotein on the time-scale of the formation of the Kstate (picoseconds at ambient temperature,10,11 andvery rapid even at cryogenic temperatures12), isregarded as a rigid, non-compliant matrix, the
0022-2836/02/$ - see front matter q 2002 Elsevier Science Ltd. All rights reserved
E-mail address of the corresponding author:[email protected]
Abbreviations used: BR, bacteriorhodopsin; K, L, M,N, and O, photointermediates of bacteriorhodopsin;FTIR, Fourier transform infrared.
doi:10.1016/S0022-2836(02)00681-2 available online at http://www.idealibrary.com onBw
J. Mol. Biol. (2002) 321, 715–726

entropic contribution to the excess free energy of Kwill be only from the retinal. Thus, the gain offree energy in K might be larger than 49 kJ/mol,but probably not by much. Once the photocycle iscomplete, about 20 kJ/mol of free energy willhave been retained in the electrochemical protongradient created across the membrane barrier, andthe rest dissipated.
Studies of the K state have focused on theimmediate neighborhood of the Schiff base ofthe photoisomerized retinal. In non-illuminatedbacteriorhodopsin (the BR state), the Schiff baseN–H bond vector points in the extracellulardirection.13,14 As shown in Figure 1, the positivelycharged retinylidene nitrogen atom donates ahydrogen bond to water 402, which is in turn thehydrogen bond donor to the anionic carboxylategroups of Asp85 and Asp212.15,16 The 3D hydro-gen-bonded network contains also OD1 of Asp85,hydrogen-bonded to OG1 of Thr89, as well asOD1 of Asp212, hydrogen-bonded to the phenolicOH group of Tyr185. OD2 of Asp212 is, in turn,hydrogen-bonded to the phenolic OH group ofTyr57. The positive charge of Arg82 and the hydro-gen bonds of water molecules 401 and 406 providefurther stabilization to the negative charges nearwater 402. According to Raman and infraredspectra,17 – 20 the configuration of the retinal ischanged from all-trans in BR to a twisted 13-cis,15-anti in the K intermediate. As might beexpected, the distortion of the polyene chain isbetween C13 and the Schiff base. The prominentbands between 811 and 985 cm21 in the Ramanand Fourier transform infrared (FTIR) spectra of Koriginate from out-of-plane hydrogen vibrations,which increase in intensity upon displacement ofthe C14H, C15H and NZH hydrogen atoms out ofthe plane of the retinal. These distortions of thephotoisomerized retinal must upset the hydrogen-bonded network in Figure 1. Indeed, frequencyshifts of O–H stretching bands assigned to bound
water,21,22 Thr89,23,24 and Tyr18525,26 indicatedchanges in the strengths of numerous hydrogenbonds.
In what form, then, is energy conserved in the Kintermediate? The first suggestion27,28 was that theK intermediate (as bathorhodopsin in the case ofvisual rhodopsin) is a high-energy state becausethe positively charged retinal Schiff base hasmoved away from its counter-ion, i.e. Asp85 (laterconsidered to include Asp212). In a more recentversion of this model,29 the N–H bond of theSchiff base changes its direction, perforce, uponrotation of the C13vC14 double bond in K. In thiscase, the electrostatic cost of moving the N–Hbond from the highly polar extracellular region tothe hydrophobic cytoplasmic region would corre-spond, partly or fully, to the gain of free energy inK. The second model assumed that, as in batho-rhodopsin of visual rhodopsin, the gain of freeenergy in K resides in bond distortions in theretinal that arise from steric conflict with the bind-ing site after photoisomerization.8,30,31 Relaxationof the strained retinal, as the binding-site geometrychanges with time to accommodate it, wouldtransfer free energy to the protein. In the third, the“electrostatic steering” model,32 the retinal is dis-torted also, but its distortion arises because electro-static interaction with the counter-ion keeps theN–H bond in its original extracellular direction.This strained orientation would persist until theSchiff base is deprotonated later in the cycle.Deciding among these alternatives can be doneonly on the basis of a full description of thegeometry of the retinal and its surroundings inK. Electron diffraction of a 2D bacteriorhodopsincrystal, at 3.5 A resolution and in projection,detected no changes in the K state,33 but there is areport on the structure of K from X-raydiffraction.34 At 30% occupancy of K (produced byillumination with green laser at 100 K) and 2.1 Aresolution, the retinal chain appeared disorderedand the Schiff base N–H bond was shown asreoriented to the cytoplasmic direction. Water 402was not present (or disordered), and the peptideCvO of Lys216, to which the retinal is connected,had moved noticeably. Displacement of the car-boxyl group of Asp85 was suggested to be relatedto a rise of its pKa, in preparation for protonationby the Schiff base later in the cycle. Displace-ments were described also at Asp212, Tyr57, andMet20.
Here, we report on our attempts to refine acrystallographic model of K in the same way as inthe earlier report,34 but from crystals that diffractedto 1.4–1.5 A. We find that the structural changesare restricted mainly to the retinal, which assumesa distorted 13-cis, 15-anti conformation consistentwith the interpretation of vibrational spectra inmany publications. As in the BR state, the Schiffbase N–H vector points approximately to theextracellular direction, but a decrease in theNZ–H–O angle makes its hydrogen bond towater 402 weaker. The results suggest that K is a
Figure 1. Region of the hydrogen-bonded network ofbacteriorhodopsin near the retinal Schiff base andwater 402. Only the side-chains and hydrogen bondsdiscussed in the text are included. Hydrogen bonds areshown in green. The Figure was prepared from thePDB coordinate set 1C3W,15 with the graphics programSetor.56
716 K Intermediate of Bacteriorhodopsin
high-energy state to a large extent because theangle of C12–C13vC14 is distorted, a consequenceof the fact that the retinal retains its initialextended shape in its binding site, in spite ofrotation of the C13vC14 double bond.
Results
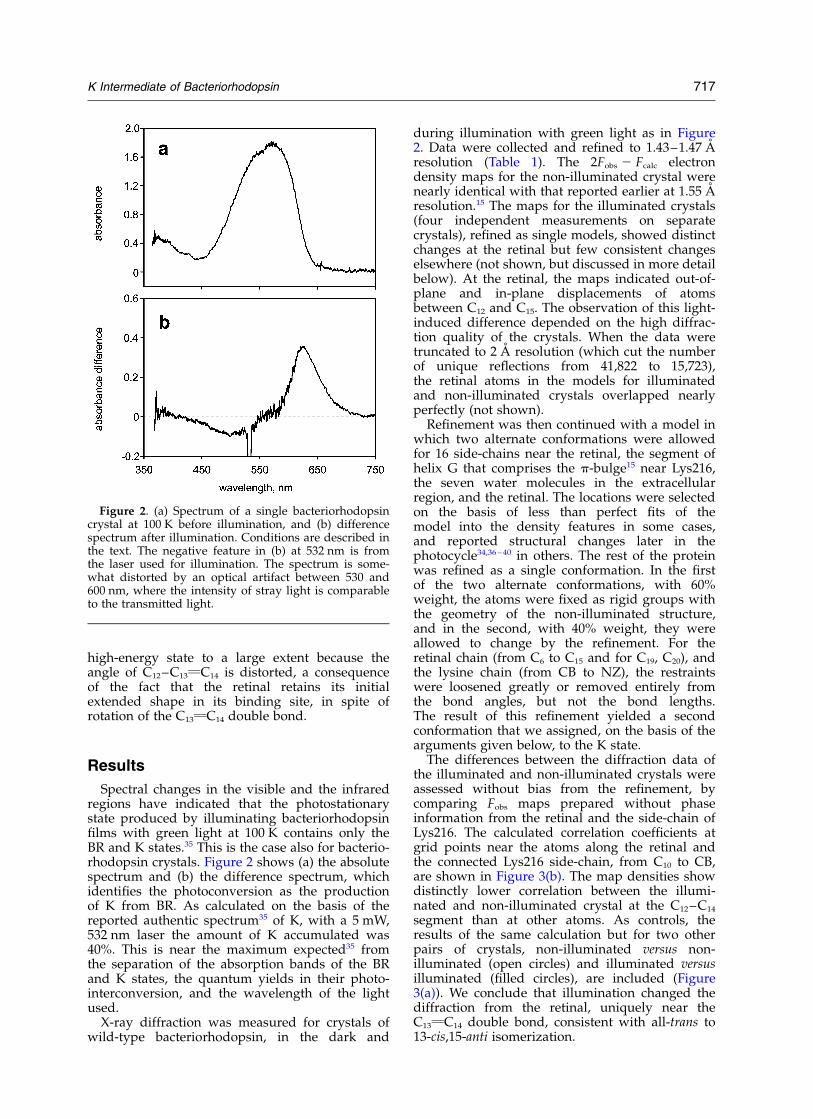
Spectral changes in the visible and the infraredregions have indicated that the photostationarystate produced by illuminating bacteriorhodopsinfilms with green light at 100 K contains only theBR and K states.35 This is the case also for bacterio-rhodopsin crystals. Figure 2 shows (a) the absolutespectrum and (b) the difference spectrum, whichidentifies the photoconversion as the productionof K from BR. As calculated on the basis of thereported authentic spectrum35 of K, with a 5 mW,532 nm laser the amount of K accumulated was40%. This is near the maximum expected35 fromthe separation of the absorption bands of the BRand K states, the quantum yields in their photo-interconversion, and the wavelength of the lightused.
X-ray diffraction was measured for crystals ofwild-type bacteriorhodopsin, in the dark and
during illumination with green light as in Figure2. Data were collected and refined to 1.43–1.47 Aresolution (Table 1). The 2Fobs 2 Fcalc electrondensity maps for the non-illuminated crystal werenearly identical with that reported earlier at 1.55 Aresolution.15 The maps for the illuminated crystals(four independent measurements on separatecrystals), refined as single models, showed distinctchanges at the retinal but few consistent changeselsewhere (not shown, but discussed in more detailbelow). At the retinal, the maps indicated out-of-plane and in-plane displacements of atomsbetween C12 and C15. The observation of this light-induced difference depended on the high diffrac-tion quality of the crystals. When the data weretruncated to 2 A resolution (which cut the numberof unique reflections from 41,822 to 15,723),the retinal atoms in the models for illuminatedand non-illuminated crystals overlapped nearlyperfectly (not shown).
Refinement was then continued with a model inwhich two alternate conformations were allowedfor 16 side-chains near the retinal, the segment ofhelix G that comprises the p-bulge15 near Lys216,the seven water molecules in the extracellularregion, and the retinal. The locations were selectedon the basis of less than perfect fits of themodel into the density features in some cases,and reported structural changes later in thephotocycle34,36 – 40 in others. The rest of the proteinwas refined as a single conformation. In the firstof the two alternate conformations, with 60%weight, the atoms were fixed as rigid groups withthe geometry of the non-illuminated structure,and in the second, with 40% weight, they wereallowed to change by the refinement. For theretinal chain (from C6 to C15 and for C19, C20), andthe lysine chain (from CB to NZ), the restraintswere loosened greatly or removed entirely fromthe bond angles, but not the bond lengths.The result of this refinement yielded a secondconformation that we assigned, on the basis of thearguments given below, to the K state.
The differences between the diffraction data ofthe illuminated and non-illuminated crystals wereassessed without bias from the refinement, bycomparing Fobs maps prepared without phaseinformation from the retinal and the side-chain ofLys216. The calculated correlation coefficients atgrid points near the atoms along the retinal andthe connected Lys216 side-chain, from C10 to CB,are shown in Figure 3(b). The map densities showdistinctly lower correlation between the illumi-nated and non-illuminated crystal at the C12–C14
segment than at other atoms. As controls, theresults of the same calculation but for two otherpairs of crystals, non-illuminated versus non-illuminated (open circles) and illuminated versusilluminated (filled circles), are included (Figure3(a)). We conclude that illumination changed thediffraction from the retinal, uniquely near theC13vC14 double bond, consistent with all-trans to13-cis,15-anti isomerization.
Figure 2. (a) Spectrum of a single bacteriorhodopsincrystal at 100 K before illumination, and (b) differencespectrum after illumination. Conditions are described inthe text. The negative feature in (b) at 532 nm is fromthe laser used for illumination. The spectrum is some-what distorted by an optical artifact between 530 and600 nm, where the intensity of stray light is comparableto the transmitted light.
K Intermediate of Bacteriorhodopsin 717

Figure 4(a) and (b) show 2Fobs 2 Fcalc densitymaps and models at the retinal for an illuminatedand a non-illuminated crystal. The maps differonly slightly (indicated by an arrow), as expectedfrom the relatively small displacements of atomsin the C13vC14 isomerization. The models for bothdata sets were refined with two conformations,
and in the same way. As expected for a homo-geneous sample, in the case of the non-illuminatedcrystal the second conformation refined as all-trans,and only slightly different from the first con-formation (Figure 4(b)). In the model for theilluminated crystal (Figure 4(a)), the second confor-mation (i.e. the putative K state) differs from the
Table 1. X-ray data collection and refinement statistics for data sets from a non-illuminated crystal and a crystal illumi-nated with green laser at 100 K
Non-illuminated crystal Illuminated crystal
Data resolution range (A) 1.47–25.0 1.43–25.0Total observations 599,134 608,447Unique reflections 38,667 41,822Rmerge
a,b (%) 3.9 (63.9) 3.7 (66.4)Average I/s(I )b,c 35.5 (2.9) 34.9 (2.4)Completenessb (%) 97.3 (92.5)c 96.2 (89.4)Mosaicity (deg.) 0.50 0.55
Refinement range (A) 1.47–25.0 1.43–25.0Structure factors 38,646 41,809Restraints 8145 8447Parameters 8296 8359Twin ratio 54:46 51:49Protein atoms 1720 1745Retinal atoms 20 40Water molecules 23 30Lipid atoms 310 310R-factord,e (%) 14.9 (13.6) 13.4 (12.7)Rfree
e,f (%) 17.9 (16.5) 17.6 (16.6)Average protein B (A2) 26.7 26.2Average retinal B (A2) 17.5 17.3Average water B (A2) 32.4 33.8Average lipid B (A2) 60.8 60.0Deviation from ideal bond lengths (A) 0.015 0.012Deviation from ideal bond angle distances (A) 0.029 0.029
The illuminated crystal data were refined as two conformations at some locations in the protein, as described in the text.a RmergeðIÞ ¼
Phkl
PilIhkl;i 2 kIhklll=
Phkl
PilIhkl;il; where kIhkll is the average intensity of the multiple Ihkl,i observations for symmetry-
related reflections.b Values in parentheses for the 1.47–1.51 A resolution shell in the case of the non-illuminated crystal, and for the 1.43–1.48 A resol-
ution shell in the case of the illuminated one.c I/s(I ), average of the diffraction intensities, divided by their standard deviations.d R-factor ¼
PhkllFobs 2 Fcalcl=
PhkllFobsl; where Fobs and Fcalc are observed and calculated structure factors, respectively.
e Values are for all data, those in parentheses for F . 4s(F ).f Rfree ¼
Phkl1TlFobs 2 Fcalcl=
Phkl1TlFobsl; where a test set (5% of the data) is omitted from the refinement in such a way that all
structure factors in each of several thin resolution shells were selected to avoid bias from merohedral twinning.
Figure 3. Correlation coefficientsbetween pairs of Fobs maps, pre-pared with intensities from thereflection files and phases fromcoordinate sets with the retinal andthe side-chain of Lys216 removed.The correlations for these maps,unbiased by any model at theretinal, were calculated for gridpoints near the atoms shown, fromC10 of the retinal to CB of Lys216.(a) Comparison of a pair ofmaps for non-illuminated crystals(present data and 1C3W,15 opencircles) and illuminated crystals(present data, filled circles).(b) Comparison of maps for anilluminated and a non-illuminatedcrystal (present data, open circles).
718 K Intermediate of Bacteriorhodopsin

first, and the C13vC14 double bond is clearly 13-cis.Because the differences in the maps are small andthe model for the K state is derived from theminority conformation, confidence in the structurerequires multiple independent measurements. In
the following, we compare the models fromdiffraction determined for four illuminated crystals.
Figure 5(a) shows the refined models of theretinal and Lys216 in the non-illuminated crystal(BR state), with atom notations and numbering.Figure 5(b) shows overlapped models for K fromfour illuminated crystals (blue, green, yellow, andred), in two views, and the model from thehighest-resolution data (red). In the latter, theretinal in BR is underlaid to illustrate the changesin the K state. The changes appear to be restrictedmainly to the retinal atoms between C12 and C15.Much less displacement, if any, is detected else-where in the retinal, and in the lysine side-chain.The variability among the four K structures illus-trates the magnitude of uncertainty concerning thegeometry of the retinal in this state, with data ofthe quality currently available.
Table 2 contains selected retinal bond angles forthe BR and K state. A large increase in the bondangle at C13 is seen, from 1128 to 1138 in BR withall-trans retinal to 145(^12)8 in K. This angle in BRis near the value in retinal crystals. The increasedangle in K is consistent with the overall distortionof the retinal, which keeps it linear as in all-transin spite of rotation of the C13vC14 bond thatwould otherwise introduce a sharp bend at C13.The ideal values for the torsion angles of theC13vC14, C14–C15, and C15vNZ bonds would be1808, 1808, and 1808 in all-trans, and 08, 1808, and1808 in 13-cis,15-anti. In the BR state, the torsionangles are well defined: at C13vC14 it is somewhatless, by about 258, than expected, and the othersare near their ideal values. In K, the standarderrors of these angles are much greater, but thereis little doubt that the C13vC14 bond has rotated tonear its ideal torsion angle for 13-cis, whileC15vNZ, and probably C14–C15, rotated also anddeviate from the expected torsion angles of 1808.For one crystal, illumination was not throughoutthe diffraction measurement as before34 but for tenminutes before data collection only; the structuralchanges were the same as in the other three modelsfor K.
In view of the importance of water 402 and itsproximity to the retinal Schiff base,14 – 16 and theearlier report that it was not observable in the Kstate,34 we paid special attention to its presence inK. Whether the occupancy of this water moleculeis 100% or 60% was difficult to decide, as it couldbe judged only from the temperature factors ofthe atoms connected to water 402. For the non-illuminated crystal, the B-factor of water 402refined to 23 A2. The B-factors of atoms in itsimmediate vicinity, NZ of Lys216, OD2 of Asp85,and OD2 of Asp212, were somewhat lower, at20 A2, 18 A2, and 20 A2, respectively, forming apattern. In the illuminated crystal, with a singleconformation model and full occupancy of water402, the water oxygen atom had a B-factor of24 A2. This value forms the same pattern withthe B-factors of NZ of Lys216, OD2 of Asp85,and OD2 of Asp212 (22 A2, 20 A2, and 19 A2,
Figure 4. The 2Fobs 2 Fcalc maps for (a) an illuminatedcrystal and (b) a non-illuminated crystal near the retinalSchiff base region. In both cases, refinement assumedtwo conformations, with occupancies of 60% (unchangedBR state) and 40% (K state), respectively, as discussed inthe text. The model of the first conformation is shown ingreen, the second in gray. The Figure was prepared withgraphics program Setor.56
Figure 5. Conformation of the retinal and the Lys216side-chain in the (a) BR and (b) K states. In (a) all-transretinal from the PDB coordinates 1C3W,15 the atomdesignation and numbering are given. In (b) thedistorted 13-cis,15-anti retinal is shown in various ways.In descending order, the models from four independentdeterminations (blue, green, yellow, and red) are shownoverlapped, in two views at right-angles to one another,and the model from the highest-resolution data set (red,cf. Table 1) is shown from the same two directions.For the latter, the model for all-trans retinal (gray, from(a)) is underlaid to illustrate the changes.
K Intermediate of Bacteriorhodopsin 719

respectively). However, when the occupancy ofwater 402 in the illuminated crystal was set to60%, its B-factor refined to 16 A2. This is muchlower than what the pattern in the non-illuminatedcrystal would predict, and suggests that water 402is not absent or disordered in the second confor-mation, i.e. in K. The FTIR difference spectra ofthe K intermediate, measured with polarizationand in 2H2O to separate the O– 2H stretch bandsfrom interfering spectral features,21,22 lead to thesame conclusion. The O– 2H depletion bandsobserved at 2171 and 2292 cm21 very likely origi-nate both from water 402, because they are thebands with greatest downshift relative to theO– 2H stretch of liquid 2H2O (near 2600 cm21), andtherefore represent the water molecule(s) moststrongly hydrogen-bonded in the protein. Water402 is unique in BR, being hydrogen-bonded totwo anionic carboxylate groups and polarizedfurther by a third hydrogen-bond, to the Schiffbase (Figure 1). In the K state, both O– 2H bandsshift to higher frequency, but by only 70–90 cm21,i.e. their hydrogen bonds are weaker but notnearly as much as what might be expected fordisordered water.
Inspection of the 2Fobs 2 Fcalc maps, and Fobs 2Fobs difference maps, revealed little that was signifi-cantly changed in K elsewhere in the protein. Com-parison of the maps for non-illuminated andilluminated crystals, as in Figure 4, revealed nomovement, within error, of the side-chains ofVal49, Tyr57, Tyr83, Asp85, Trp86, Thr89, Leu93,Thr178, Trp182, the p-bulge of helix G,15 andwater molecules 401–404, and 406. There wassome apparent movement (in a few cases up to1 A) of the side-chains of Arg82, Leu181, Glu194,Glu204, Asp212, and of water molecule 407, butthe same kind of displacements were seen in thesecond conformations of both illuminated andnon-illuminated data sets. These discrepanciesmust reflect, therefore, local disorder instead ofchanges in K. The only displacement of possiblesignificance, observed in all three illuminatedcrystals, was for water 405 which moved by0.7–1.2 A in the second conformation (i.e. in K),
but only 0.3 A in the equivalent conformation ofthe non-illuminated crystal. This movementwould break the hydrogen bond of water 405 toNE of Arg82, although not to OE1 of Glu194. Inthe final refinement, only the retinal with all ofLys216 including its peptide segment, the mainchain of helix G with the p-bulge, the side-chainsof Asp85 and Arg82, and the seven watermolecules, 401–407, were modeled as two confor-mations. There may be structural changes in theprotein additional to those in the retinal and atwater 405, but their magnitudes are too small tobe detected or assigned uniquely at the resolutionof the present data.
Discussion
The torsion angle for C14–C15, (dihedral angle forC13vC14–C15–NZ) of 177–1798 in non-illuminatedbacteriorhodopsin (Table 2) appears to be inconflict with the 164(^6)8 obtained for theH–C14–C15–H dihedral angle from solid-stateNMR.41 We found that removing all restraintsregarding planarity from the input model betweenC13 and NZ resulted in a C14–C15 torsion angledifferent from 1778 by only 18 (not shown). Thus,for non-illuminated bacteriorhodopsin, at theavailable resolution and 100% occupancy, theretinal geometry is very well defined. As pointedout,41 both angles may be right. In a quantummechanical/molecular dynamics calculation theC13vC14–C15–NZ and H–C14–C15–H dihedralangles were predicted42 to be different: 1798 and1658, respectively, which agree fully with the laterexperimental findings.
The retinal geometry in the K state is less welldefined (see Figure 5(b) and the statistics for fourdata sets in Table 2), as it is the result of dissectingthe contribution of a minority conformation to thediffraction. However, the results do answer themain questions concerning K. From the changes ofthe C13vC14 torsion angle (Table 2) we concludethat the C13vC14 double bond is rotated in Kfully, or nearly fully, from trans to cis. In a relaxed
Table 2. Retinal bond angles in non-illuminated bacteriorhodopsin (BR) and the K intermediate
Coordinate seta C12–C13vC14 angle (8) C13vC14 torsion (8) C14–C15 torsion (8) C15vNZ torsion (8)
BR (1C3W) 113 2157 179 2163BR (this study) 112 2154 177 2172
K (crystal 1) 143 11 154 67K (crystal 2) 160 25 123 79K (crystal 3) 130 256 173 131K (crystal 4) 148 25 94 119K (average)b 145 ^ 12 22 ^ 39 138 ^ 35 101 ^ 31
Positive torsion angles refer to clockwise rotation from 08 when viewed from the b-ionone ring of the retinal, negative torsion anglesrefer to counter-clockwise rotation. Thus, 1808 and 21808 coincide.
a The data for the second BR and crystal 3 are in Table 1. The crystallographic resolutions for crystals 1, 2, and 4 were 1.54 A, 1.50 A,and 1.48 A, respectively. Unlike crystals 1–3, which were illuminated throughout the data collection, crystal 4 was illuminated for tenminutes, and the diffraction data were collected in the dark.
b Averages weighted according to the number of structure factors in the refinement.
720 K Intermediate of Bacteriorhodopsin

13-cis,15-anti configuration, where the three torsionangles in Table 2 would assume values of 08, 1808,and 1808, respectively, this would reorient theSchiff base N–H bond from the extracellular tothe cytoplasmic direction. However, the torsionangle of the C15vNZ double bond (and probablyalso the C14–C15 single bond), deviates from 1808in the counterclockwise direction. The partialreverse rotation(s) allow the N–H bond vector toremain roughly in the extracellular direction inspite of the rotation of the C13vC14 bond (Figure6). The non-planar torsion angles about theC14–C15 and C15vNZ bonds are consistent withthe large amplitudes of the observed hydrogenout-of-plane (HOOP) modes in the K state.17 – 20
The new retinal configuration in K shouldchange the strengths of hydrogen bonds in thenetwork around water 402. This is evident in FTIRspectra, which have provided a fairly detaileddescription of these changes for K produced atcryogenic temperatures. From shifts of twostrongly hydrogen-bonded O– 2H stretch bands tohigher frequencies it could be concluded21,22 thattwo hydrogen bonds become weaker. The O– 2Hbands were assigned tentatively to watermolecules 402, 401 or 406, but most likely theyboth belong to water 402, as discussed above.Another O– 2H stretch, a more weakly hydrogen-bonded one, was shifted to lower frequency,leading to the suggestion that the hydrogen bondof a third water molecule becomes stronger.21 Theshift of the O–H and O–2H stretch bands of Thr89to lower frequency23,24 suggested that in K thehydrogen bond of OG1 of Thr89, with OD1 ofAsp85, becomes stronger also. Further, it wasargued25,26 from a negative band at 1277 cm21 in K,assigned to the phenolic C–O2 stretch of Tyr185,that in the hydrogen bond between the side-chainsof Asp212 and Tyr185 the hydrogen atom shifts to
Tyr185. Because Tyr185 is the hydrogen bonddonor, this would mean that the Tyr185 to Asp212hydrogen bond becomes weaker.
The structural model for K we report hereexplains many, although not all, of these findings.Figure 6 shows the region at the retinal Schiffbase in the BR and K states. Movements of theSchiff base nitrogen atom NZ, water 402, andAsp85 and Asp212 in K are no greater than0.1–0.2 A, i.e. within the estimated positionalerror in the map. However, if the Schiff baseN–H bond remains coplanar with C15vNZ–CE,as one must assume, the angle of the hydrogenbond NZ–H–O changes from a nearly perfectlinear 170–1768 in BR to 113(^13)8 in K. Indeed,polarized FTIR measurements of K in 2H2O indi-cate that the direction of the NZ– 2H bond changesby about this extent.43 Deviations from the optimalangle will make the hydrogen bond between theSchiff base and water 402 weaker, and increasethe partial negative charge of the oxygen atom. Byinference, water 402 will become a poorer hydro-gen donor, and therefore its hydrogen bonds withthe two aspartate carboxyl groups will becomeweaker. This would be consistent with theobserved upshift of the two lowest-frequencyO–2H stretch bands. Further, being the acceptor ofa weaker hydrogen bond, the carboxyl group ofAsp85 will become more negative and a betterhydrogen bond acceptor to Thr89, as indeedsuggested from FTIR.24 For the same reason, itshydrogen-bond with water 401 should becomestronger, consistent with the downshift of anO–2H stretch band frequency.21 Alternatively, thisband might originate from water 405. By the sameargument, however, the hydrogen bond betweenAsp212 and Tyr185 should become stronger, andthat is contrary to the conclusion made fromFTIR.25,26 The reason for the disagreement is notclear.
As for the Schiff base CvN stretch, itsfrequency is well defined in the Raman spectra ofboth BR (1640 cm21) and the L state (1644 cm21).Coupling of the CvN stretching internal coordi-nate to the N–H rock results in shifts of 220 cm21
(BR) and 224 cm21 (L) in 2H2O.44 These frequenciesand frequency shifts have been interpreted45,46 interms of a relatively weak hydrogen-bonding ofNZ–H. The CvN stretch in the K state is at ananomalously low frequency (1609 cm21), andappears to shift only 4 cm21 upon N-deuteration,suggesting further weakened hydrogen-bondingand thus lesser coupling with C15vNZ stretch.This is consistent with the structural change atwater 402 (Figure 5). The distorted C14–C15vNZ–CE geometry and the associated weakenedhydrogen bond of the Schiff base with water 402in K (Figure 5) is an unstable arrangement, notencountered in model compounds. We note thatremoving this hydrogen-bond may be sufficient,by itself, to destabilize the four buried chargesaround water 402 (Figure 1), and result in protona-tion of Asp85 by the Schiff base later in the cycle.
Figure 6. Structural changes in the K state at the retinalSchiff base and water 402. Oxygen atoms are red,nitrogen atoms are blue, and the Schiff base proton isshown in white, the hydrogen bonds are shown ingreen. (a) Model of the K state. The N–H–O angle inthe hydrogen-bond of the Schiff base NZ with water 402has an unfavorable value, at 113(^14)8 (average fromthe four models in Figure 5). (b) Model of the BR state(from the present data). The N–H–O atoms are nearlyco-linear, with an angle of 170–1768.
K Intermediate of Bacteriorhodopsin 721
The red-shifted maximum of K in the visible regionis explained, at least partly, by the weaker Schiffbase N–H hydrogen-bond (Figure 6) and theunusual C15vNZ torsion (Table 2).
There is a large electrical signal from the K statein oriented bacteriorhodopsin, with a sign oppositeto the direction of proton transport.47 Its originwas suggested to be either the displacement of thepositively charged Schiff base in the cytoplasmicdirection or the redistribution of electrons, uponthe isomerization of the retinal. In the crystallo-graphic structure of K we find very little changein the position of NZ of Lys216 and the surround-ing atoms (Figure 6). However, the redistributionof electrons between NZ and water 402, and in theconnected hydrogen-bonded network, is a goodcandidate for the observed electric signal, as itwould increase the magnitude of the Schiffbase-counterion dipole.
Table 2 compares the angle of C12–C13vC14 in BRand K. The increase of this bond angle from thenear-canonic value of 1128 in BR (all-trans retinal)to 145(^12)8 in K is related to the fact that theend-to-end length of the retinal is virtuallyunchanged in spite of the rotation of the C13vC14
double bond. We assume the reason is stiffness of
the retinal-binding site. Because in K the polyenechain cannot, as yet, assume the bent shape of therelaxed 13-cis,15-anti retinal, at least one bondangle is forced to increase. The angle that increasesis the one at C13. We had described a somewhatincreased angle at C13 (1258) in one of the M statesthat arises later in the photocycle,37 but thisdistortion is exaggerated in K.
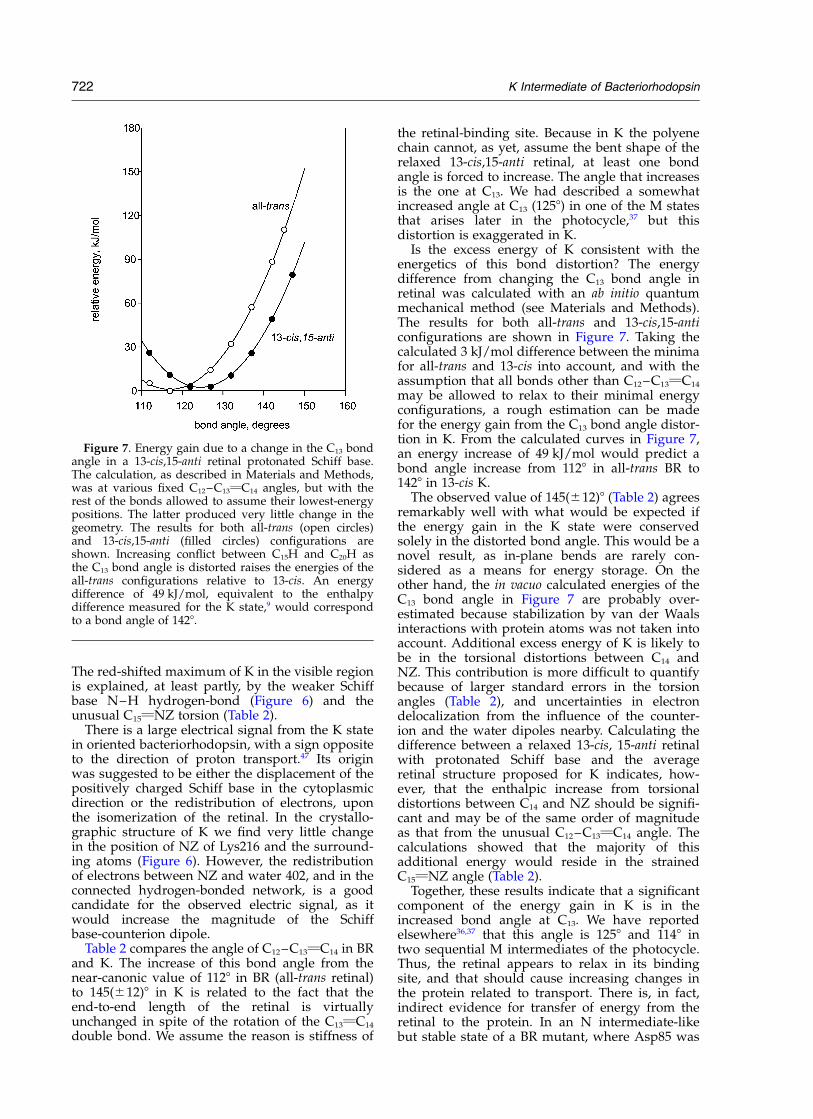
Is the excess energy of K consistent with theenergetics of this bond distortion? The energydifference from changing the C13 bond angle inretinal was calculated with an ab initio quantummechanical method (see Materials and Methods).The results for both all-trans and 13-cis,15-anticonfigurations are shown in Figure 7. Taking thecalculated 3 kJ/mol difference between the minimafor all-trans and 13-cis into account, and with theassumption that all bonds other than C12–C13vC14
may be allowed to relax to their minimal energyconfigurations, a rough estimation can be madefor the energy gain from the C13 bond angle distor-tion in K. From the calculated curves in Figure 7,an energy increase of 49 kJ/mol would predict abond angle increase from 1128 in all-trans BR to1428 in 13-cis K.
The observed value of 145(^12)8 (Table 2) agreesremarkably well with what would be expected ifthe energy gain in the K state were conservedsolely in the distorted bond angle. This would be anovel result, as in-plane bends are rarely con-sidered as a means for energy storage. On theother hand, the in vacuo calculated energies of theC13 bond angle in Figure 7 are probably over-estimated because stabilization by van der Waalsinteractions with protein atoms was not taken intoaccount. Additional excess energy of K is likely tobe in the torsional distortions between C14 andNZ. This contribution is more difficult to quantifybecause of larger standard errors in the torsionangles (Table 2), and uncertainties in electrondelocalization from the influence of the counter-ion and the water dipoles nearby. Calculating thedifference between a relaxed 13-cis, 15-anti retinalwith protonated Schiff base and the averageretinal structure proposed for K indicates, how-ever, that the enthalpic increase from torsionaldistortions between C14 and NZ should be signifi-cant and may be of the same order of magnitudeas that from the unusual C12–C13vC14 angle. Thecalculations showed that the majority of thisadditional energy would reside in the strainedC15vNZ angle (Table 2).
Together, these results indicate that a significantcomponent of the energy gain in K is in theincreased bond angle at C13. We have reportedelsewhere36,37 that this angle is 1258 and 1148 intwo sequential M intermediates of the photocycle.Thus, the retinal appears to relax in its bindingsite, and that should cause increasing changes inthe protein related to transport. There is, in fact,indirect evidence for transfer of energy from theretinal to the protein. In an N intermediate-likebut stable state of a BR mutant, where Asp85 was
Figure 7. Energy gain due to a change in the C13 bondangle in a 13-cis,15-anti retinal protonated Schiff base.The calculation, as described in Materials and Methods,was at various fixed C12–C13vC14 angles, but with therest of the bonds allowed to assume their lowest-energypositions. The latter produced very little change in thegeometry. The results for both all-trans (open circles)and 13-cis,15-anti (filled circles) configurations areshown. Increasing conflict between C15H and C20H asthe C13 bond angle is distorted raises the energies of theall-trans configurations relative to 13-cis. An energydifference of 49 kJ/mol, equivalent to the enthalpydifference measured for the K state,9 would correspondto a bond angle of 1428.
722 K Intermediate of Bacteriorhodopsin

replaced with Asn and Asp96 had a lowered pKa
and was unprotonated, FT-Raman spectra showedthat the retinal spontaneously (in the dark) iso-merized to 13-cis,15-anti.31 This suggested thatonce the proton acceptor and donor groups changetheir ionization states during the transport cycle,the retinal-binding site will favor the photo-isomerized configuration. A similar conclusionwas reached recently from interatomic distancesdetermined with solid-state NMR in the D85Nmutant at alkaline pH.48 Thus, while the excessfree energy of the unusually wide C13 bond angledisappears as the retinal gradually relaxes to anundistorted 13-cis configuration, in the processenergy will be conserved in structural changes inthe protein. These are the changes that result inprotonation and deprotonation of strategicallyplaced groups and/or water and the release of aproton to the extracellular side and its uptakefrom the cytoplasmic side, i.e. the transport. Theexcess energy initially contained in the retinal inthe K state, is thereby dissipated through creationof a transmembrane electrochemical gradient forthe transported proton, and recovery of the initialstate of the protein.
Finally, NMR studies on the primary photo-product of the visual pigment rhodopsin49 arrivedat a similar conclusion as above, namely thatelectrostatic interactions and out-of-plane distor-tions were not sufficient to explain the storedenergy. As in bacteriorhodopsin, the retinal ispacked tightly in the interior of this protein, and itwas concluded that a significant component of theenergy gained following retinal isomerizationmust reside in unusual in-plane bends of thepolyene chain.
Materials and Methods
Data collection
As before,14,15,36,37 the crystals, grown in cubic lipidphase as earlier described,50,51 are thin hexagonal platesabout 80 mm £ 80 mm £ 10–15 mm in size. Pieces of thecubic phase with encased crystals were soaked overnightin the mother liquor plus 0.1% (w/v) octylglucoside, andthe crystals were extracted and mounted by mechanicalmanipulation with a nylon loop. Spectra before andafter illumination with a green laser (532 nm, 5 mW) at100 K were taken with a fiber-optic microspectro-photometer.37
The crystals were light-adapted with white lightbefore freezing in liquid nitrogen. Diffraction data werecollected at 100 K, either while continuously illuminatingthe crystals with green laser, as reported earlier,34 or afterten minutes illumination and in the dark, at beamlines5.0.1 and 5.0.2 of ALS (Lawrence Berkeley Laboratory),using a 2 £ 2 array CCD detector (ADSC, San Diego).Typically, 90–150 images with 1–0.68 oscillation angleswere integrated and scaled with DENZO andScalepack,52 in the P63 space group, with a ¼ b ¼ 61:2 �A;c ¼ 111:1 �A:
Refinement
Refinement of the models with SHELX 9753 tookmerohedral twinning14 into account, and used as initialinput the model from 1.55 A resolution X-ray diffraction(PDB entry code 1C3W; Luecke et al.15). For the non-illuminated crystal, the 2Fobs 2 Fcalc maps, and thecoordinates of the refined model indicated no significantdifferences from the earlier structure. The refinement ofthe models for illuminated crystals allowed two alternateconformations, with weights of 60% and 40%, at variousside-chains, part of the main chain, the retinal and somewater molecules. Rfree was not affected significantly bychanging the weights of the two conformations, andthey were fixed on the basis of the composition of thephotostationary state in the crystals as estimated fromthe spectral changes. The groups in the two confor-mations were selected initially on the basis of possiblechanges in the density maps or changes reported inphotointermediates that follow K. The side-chains withtwo allowed conformations were Val49, Tyr57, Arg82,Tyr83, Asp85, Trp86, Leu93, Thr178, Leu181, Trp182,Tyr185, Glu94, Glu204, Asp212, and Lys216. The mainchain so refined included the segment between the CAof Ala215 and the N of Val217. Two conformations wereallowed also for all retinal atoms, and water molecules401–407. The groups in the majority conformation wereeach fixed as rigid fragments with geometries as in themodel of the non-illuminated crystal, and the groups inthe minority conformation, in the putative structure ofthe K state, were refined together with the rest of theprotein where the occupancies of atoms were set to100%. As a control, and in addition to the single confor-mation refinement, the model for the non-illuminatedcrystal was refined with two conformations in the sameway. Small displacements at the retinal, and largerdisplacements elsewhere in the protein in this data setplaced limits on what would be considered meaningfuldifferences between illuminated and non-illuminateddata sets. With the possible exception of water 405,as discussed in Results, significant and consistentdifferences between the models for the illuminated andnon-illuminated data sets were observed only at theretinal. Omit maps indicated that the input model hadlittle influence on the electron densities at the retinal.The coordinate set reported in Table 1 for the illuminatedcrystal is for the final refined model, in which only theretinal with the connected Lys216 and the main-chainatoms that comprise the p-bulge of helix G, Asp85,Arg82, and water molecules 401–407 were set as twoconformations.
Correlation coefficients were calculated with pro-grams in the CCP4 suite54 for Fobs maps, made from thestructure factor files with phases from coordinate filesfrom which the side-chain of Lys216 and the retinalwere removed. A tagged atom map was made withcoordinates from a chain of 11 retinal and Lys216 atomswith sfall, and the correlation coefficients between pairsof Fobs maps were calculated with overlapmap at thegrid-points near the desired atoms.
Energy calculations
Ab initio energy calculations for different confor-mations of the retinal with protonated Schiff basewere carried out in vacuo, using the Gaussian 98program55 with the 6-31Gp basis set. The effects of theenvironment, including the protein, counter-ions, andwater molecules, was not taken into account. The energy
K Intermediate of Bacteriorhodopsin 723

differences calculated result only from conformationaldistortions in the retinal itself. The side-chain of Lys216was truncated after the CE carbon atom. Full optimi-zation of all-trans retinal from BR resulted in a planarconjugated double-bond system with bond angles closeto those observed in retinal model compounds. Fulloptimization starting from the average experimental Kstructure ended in 13-cis, 14-s-trans,15-anti-retinal withan overall energy only ,3 kJ/mol higher than theoptimized retinal structure in BR. The optimized Kstructure was used to calculate how the energy changesas a function of the C12–C13vC14 angle. The angle wasfixed within the range of 112–1528 in 58 increments, andthe rest of the retinal was allowed to relax to its lowest-energy state. Calculations to determine the contributionof torsion angle changes to the overall energy werecarried out in a similar fashion. In this case, constrainedoptimization of the retinal was performed with one ormore of the pertinent torsion angles fixed at the averagevalues for K listed in Table 2, while the rest of the retinalwas allowed to relax.
Protein Data Bank accession codes
The coordinates of the two structures in Table 1 havebeen deposited in the Protein Data Bank under entrycodes 1M0L and 1M0K, for the non-illuminated and theilluminated crystals, respectively.
Acknowledgments
We thank the beamline staff at the Advanced LightSource (ALS) in Berkeley, particularly K. Hendersonand G. McDermott. We are grateful to T. Poulos,D. Teller, C. Hill, H. Luecke, and L. S. Brown for valuablediscussions and suggestions. The work was supported,in part, by grants from NIH (R01-GM29498) and fromDOE (DEFG03-86ER13525).
References
1. Lanyi, J. K. & Varo, G. (1995). The photocycles ofbacteriorhodopsin. Isr. J. Chem. 35, 365–386.
2. Oesterhelt, D. (1998). The structure and mechanismof the family of retinal proteins from halophilicarchaea. Curr. Opin. Struct. Biol. 8, 489–500.
3. Haupts, U., Tittor, J. & Oesterhelt, D. (1999). Closingin on bacteriorhodopsin: progress in understandingthe molecule. Annu. Rev. Biophys. Biomol. Struct. 28,367–399.
4. Lanyi, J. K. (1999). Progress toward an explicitmechanistic model for the light-driven pump,bacteriorhodopsin. FEBS Letters, 464, 103–107.
5. Lanyi, J. K. & Luecke, H. (2001). Bacteriorhodopsin.Curr. Opin. Struct. Biol. 11, 415–419.
6. Lanyi, J. K. (2001). X-ray crystallography of bacterio-rhodopsin and its photointermediates: insights intothe mechanism of proton transport. Biochemistry(Moscow), 66, 1192–1196.
7. Luecke, H. (2000). Atomic resolution structures ofbacteriorhodopsin photocycle intermediates: the roleof discrete water molecules in the function of thislight-driven ion pump. Biochim. Biophys. Acta, 1460,133–156.
8. Birge, R. R. & Cooper, T. M. (1983). Energy storage inthe primary step of the photocycle of bacterio-rhodopsin. Biophys. J. 42, 61–69.
9. Birge, R. R., Cooper, T. M., Lawrence, A. F., Masthay,M. B., Zhang, C. F. & Zidovetzki, R. (1991). Revisedassignment of energy storage in the primary photo-chemical event in bacteriorhodopsin. J. Am. Chem.Soc. 113, 4327–4328.
10. Polland, H. J., Franz, M. A., Zinth, W., Kaiser, W.,Kolling, E. & Oesterhelt, D. (1986). Early picosecondevents in the photocycle of bacteriorhodopsin. Bio-phys. J. 49, 651–662.
11. Doig, S. J., Reid, P. J. & Mathies, R. A. (1991). Pico-second time-resolved resonance Raman spectroscopyof bacteriorhodopsin: structure and kinetics of theJ, K, and KL intermediates. Springer Proc. Phys. 68,45–48.
12. Chamorovsky, S. K., Kononenko, A. A., Rubin, A. B.& Chernavskii, D. S. (1987). Temperature depen-dence of the fast negative phase in the bacterio-rhodopsin cycle. Biofizika, 32, 601–605.
13. Lin, S. W. & Mathies, R. A. (1989). Orientation of theprotonated retinal Schiff base group in bacterio-rhodopsin from absorption linear dichroism. Biophys.J. 56, 653–660.
14. Luecke, H., Richter, H. T. & Lanyi, J. K. (1998). Protontransfer pathways in bacteriorhodopsin at 2.3 Aresolution. Science, 280, 1934–1937.
15. Luecke, H., Schobert, B., Richter, H. T., Cartailler, J. P.& Lanyi, J. K. (1999). Structure of bacteriorhodopsinat 1.55 A resolution. J. Mol. Biol. 291, 899–911.
16. Belrhali, H., Nollert, P., Royant, A., Menzel, C.,Rosenbusch, J. P., Landau, E. M. et al. (1999). Protein,lipid and water organization in bacteriorhodopsincrystals: a molecular view of the purple membraneat 1.9 A resolution. Struct. Fold. Des. 7, 909–917.
17. Pande, J., Callender, R. H. & Ebrey, T. G. (1981). Reso-nance Raman study of the primary photochemistryof bacteriorhodopsin. Proc. Natl Acad. Sci. USA, 78,7379–7382.
18. Braiman, M. & Mathies, R. (1982). Resonance Ramanspectra of bacteriorhodopsin’s primary photo-product: evidence for a distorted 13-cis retinalchromophore. Proc. Natl Acad. Sci. USA, 79, 403–407.
19. Rothschild, K. J., Roepe, P., Lugtenburg, J. &Pardoen, J. A. (1984). Fourier transform infraredevidence for Schiff base alteration in the first step ofthe bacteriorhodopsin photocycle. Biochemistry, 23,6103–6109.
20. Maeda, A., Sasaki, J., Pfefferle, J. M., Shichida, Y. &Yoshizawa, T. (1991). Fourier transform infraredspectral studies on the Schiff base mode of all-transbacteriorhodopsin and its photointermediates, Kand L. Photochem. Photobiol. 54, 911–921.
21. Kandori, H. & Shichida, Y. (2000). Direct observationof the bridged water stretching vibrations inside aprotein. J. Am. Chem. Soc. 122, 11745–11746.
22. Kandori, H., Furutani, Y., Shimono, K., Shichida, Y. &Kamo, N. (2001). Internal water molecules ofpharaonis phoborhodopsin studied by low-tempera-ture infrared spectroscopy. Biochemistry, 40,15693–15698.
23. Kandori, H., Kinoshita, N., Yamazaki, Y., Maeda, A.,Shichida, Y., Needleman, R. et al. (1999). Structuralchange of threonine 89 upon photoisomerization inbacteriorhodopsin as revealed by polarized FTIRspectroscopy. Biochemistry, 38, 9676–9683.
24. Kandori, H., Kinoshita, N., Yamazaki, Y., Maeda, A.,Shichida, Y., Needleman, R. et al. (2000). Local and
724 K Intermediate of Bacteriorhodopsin

distant protein structural changes on photoisomeri-zation of the retinal in bacteriorhodopsin. Proc. NatlAcad. Sci. USA, 97, 4643–4648.
25. Rothschild, K. J., Roepe, P., Ahl, P. L., Earnest, T. N.,Bogomolni, R. A., Das Gupta, S. K. et al. (1986).Evidence for a tyrosine protonation change duringthe primary phototransition of bacteriorhodopsinat low temperature. Proc. Natl Acad. Sci. USA, 83,347–351.
26. Rothschild, K. J., Braiman, M. S., He, Y. W., Marti, T.& Khorana, H. G. (1990). Vibrational spectroscopy ofbacteriorhodopsin mutants. Evidence for the inter-action of aspartic acid 212 with tyrosine 185 andpossible role in the proton pump mechanism. J. Biol.Chem. 265, 16985–16991.
27. Schulten, K. & Tavan, P. (1978). A mechanism for thelight-driven proton pump of Halobacterium halobium.Nature, 272, 85–86.
28. Honig, B., Ebrey, T., Callender, R. H., Dinur, U. &Ottolenghi, M. (1979). Photoisomerization, energystorage, and charge separation: a model for lightenergy transduction in visual pigments andbacteriorhodopsin. Proc. Natl Acad. Sci. USA, 76,2503–2507.
29. Kolbe, M., Besir, H., Essen, L. O. & Oesterhelt, D.(2000). Structure of the light-driven chloride pumphalorhodopsin at 1.8 A resolution. Science, 288,1390–1396.
30. Warshel, A. & Barboy, N. (1982). Energy storage andreaction pathway in the first step of the visionprocess. J. Am. Chem. Soc. 104, 1469–1476.
31. Dioumaev, A. K., Brown, L. S., Needleman, R. &Lanyi, J. K. (1998). Partitioning of free energy gainbetween the photoisomerized retinal and theprotein in bacteriorhodopsin. Biochemistry, 37,9889–9893.
32. Herzfeld, J. & Tounge, B. (2000). NMR probes ofvectoriality in the proton-motive photocycle ofbacteriorhodopsin: evidence for an electrostaticsteering mechanism. Biochim. Biophys. Acta, 1460,95–105.
33. Bullough, P. A. & Henderson, R. (1999). The projec-tion structure of the low temperature K intermediateof the bacteriorhodopsin photocycle determined byelectron diffraction. J. Mol. Biol. 286, 1663–1671.
34. Edman, K., Nollert, P., Royant, A., Belrhali, H.,Pebay-Peyroula, E., Hajdu, J. et al. (1999). High-resolution X-ray structure of an early intermediatein the bacteriorhodopsin photocycle. Nature, 401,822–826.
35. Balashov, S. P. & Ebrey, T. G. (2001). Trapping andspectroscopic identification of the photointer-mediates of bacteriorhodopsin at low temperatures.Photochem. Photobiol. 73, 453–462.
36. Luecke, H., Schobert, B., Richter, H. T., Cartailler, J. P.& Lanyi, J. K. (1999). Structural changes in bacterio-rhodopsin during ion transport at 2 A resolution.Science, 286, 255–261.
37. Luecke, H., Schobert, B., Richter, H. T., Cartailler,J.-P., Rosengarth, A., Needleman, R. et al. (2000).Coupling photoisomerization of the retinal inbacteriorhodopsin to directional transport. J. Mol.Biol. 300, 1237–1255.
38. Royant, A., Edman, K., Ursby, T., Pebay-Peyroula, E.,Landau, E. M. & Neutze, R. (2000). Helix defor-mation is coupled to vectorial proton transport inthe photocycle of bacteriorhodopsin. Nature, 406,645–648.
39. Sass, H. J., Bueldt, G., Gessenich, R., Hehn, D., Neff,D., Schlesinger, R. et al. (2000). Structural alterationsfor proton translocation in the M state of wild-typebacteriorhodopsin. Nature, 406, 649–653.
40. Facciotti, M. T., Rouhani, S., Burkard, F. T.,Betancourt, F. M., Downing, K. H., Rose, R. B. et al.(2001). Structure of an early intermediate in theM-state phase of the bacteriorhodopsin photocycle.Biophys. J. 81, 3442–3455.
41. Lansing, J. C., Hohwy, M., Jaroniec, C. P., Creemers,A. F., Lugtenburg, J., Herzfeld, J. et al. (2002).Chromophore distortions in the bacteriorhodopsinphotocycle: evolution of the H–C14–C15–Hdihedral angle measured by solid-state NMR. Bio-chemistry, 41, 431–438.
42. Hayashi, S. & Ohmine, I. (2000). Proton transferin bacteriorhodopsin: structure, excitation, IRspectra, and potential energy surface analyses by anab initio QM/MM method. J. Phys. Chem. B, 104,10678–10691.
43. Kandori, H., Belenky, M. & Herzfeld, J. (2002).Vibrational frequency and dipolar orientation ofthe protonated Schiff base in bacteriorhodopsinbefore and after photoisomerization. Biochemistry,141, 6026–6031.
44. Smith, S. O., Lugtenburg, J. & Mathies, R. A. (1985).Determination of retinal chromophore structurein bacteriorhodopsin with resonance Raman spectro-scopy. J. Membr. Biol. 85, 95–109.
45. Smith, S. O., Marvin, M. J., Bogomolni, R. A. &Mathies, R. A. (1984). Structure of the retinal chromo-phore in the hR578 form of halorhodopsin. J. Biol.Chem. 259, 12326–12329.
46. Smith, S. O., Braiman, M. S., Myers, A. B., Pardoen,J. A., Courtin, J. M. L., Winkel, C. et al. (1987).Vibrational analysis of the all-trans-retinal chromo-phore in light-adapted bacteriorhodopsin. J. Am.Chem. Soc. 109, 3108–3125.
47. Keszthelyi, L. (1988). Primary charge motions andlight-energy transduction in bacteriorhodopsin. Bio-phys. Chem. 29, 127–136.
48. Hatcher, M. E., Hu, J. G., Belenky, M., Verdegem, P.,Lugtenburg, J., Griffin, R. G. et al. (2002). Control ofthe pump cycle in bacteriorhodopsin: mechanismselucidated by solid-state NMR of the D85 N mutant.Biophys. J. 82, 1017–1029.
49. Smith, S. O., Courtin, J., de Groot, H., Gebhard, R. &Lugtenburg, J. (1991). 13C magic-angle spinningNMR studies of bathorhodopsin, the primary photo-product of rhodopsin. Biochemistry, 30, 7409–7415.
50. Landau, E. M. & Rosenbusch, J. P. (1996). Lipidiccubic phases: a novel concept for the crystallizationof membrane proteins. Proc. Natl Acad. Sci. USA, 93,14532–14535.
51. Rummel, G., Hardmeyer, A., Widmer, C., Chiu, M. L.,Nollert, P., Locher, K. P. et al. (1998). Lipidic cubicphases: new matrices for the three-dimensionalcrystallization of membrane proteins. J. Struct. Biol.121, 82–91.
52. Otwinowski, T. (1993). SCALEPACK data collectionand processing. In Data Collection and Processing(Sawyer, L., Isaacs, N. & Bailey, S., eds), pp. 56–62,SERC Daresbury Laboratory, Warrington, UK.
53. Sheldrick, G. M. & Schneider, T. (1997). High resolu-tion refinement. Methods Enzymol. 277, 319–343.
54. Collaborative Computational Project Number 4(1994). The CCP4 suite: programs for protein crystal-lography. Acta Crystallog. sect. D, 50, 760–763.
K Intermediate of Bacteriorhodopsin 725

55. Frisch, M. J., Trucks, G. W., Schlegel, H. B., Scuseria,G. E., Robb, M. A., Cheeseman, J. R., et al. (1998).Gaussian 98, Revision A.7, Gaussian, Inc, Pittsburgh,PA.
56. Evans, S. V. (1993). SETOR: hardware-lighted three-dimensional solid model representations of macro-molecules. J. Mol. Graph. 11, 134–138.
Edited by D. Rees
(Received 4 March 2002; received in revised form 18 June 2002; accepted 21 June 2002)
726 K Intermediate of Bacteriorhodopsin
Copyright © 2022 FDOKUMEN




















