
Polly Pocket & Ninja Turtles: A Content Analysis of Gender ...
Upload
independentCategory
view
2download
0
This article was downloaded by: [Igor G. Danilov]On: 21 February 2014, At: 11:45Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Journal of Systematic PalaeontologyPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/tjsp20
Cretaceous soft-shelled turtles (Trionychidae) ofMongolia: new diversity, records and a revisionIgor G. Danilova, Ren Hirayamab, Vladimir B. Sukhanovc, Shigeru Suzukid, Mahito Watabed &Natasha S. Viteke
a Zoological Institute of the Russian Academy of Sciences, Universitetskaya Emb. 1, St.Petersburg, 199034, Russiab Waseda University, Nishiwaseda 1-7-14, Shinjuku-ku, Tokyo, 169-0051, Japanc Borisyak Paleontological Institute of the Russian Academy of Sciences, Profsoyuznaya 123,Moscow, 117997, Russiad Hayashibara Natural Sciences Museum, Okayama, 700-0907, Japane Department of Geological Sciences, The University of Texas at Austin, EPS RM 1.130, 1University Station C9000, Austin, TX 78712-0254, USAPublished online: 19 Feb 2014.
To cite this article: Igor G. Danilov, Ren Hirayama, Vladimir B. Sukhanov, Shigeru Suzuki, Mahito Watabe & Natasha S. Vitek ,Journal of Systematic Palaeontology (2014): Cretaceous soft-shelled turtles (Trionychidae) of Mongolia: new diversity, recordsand a revision, Journal of Systematic Palaeontology, DOI: 10.1080/14772019.2013.847870
To link to this article: http://dx.doi.org/10.1080/14772019.2013.847870
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of theContent. Any opinions and views expressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon andshould be independently verified with primary sources of information. Taylor and Francis shall not be liable forany losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use ofthe Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

Cretaceous soft-shelled turtles (Trionychidae) of Mongolia: new diversity,records and a revision
Igor G. Danilova*, Ren Hirayamab, Vladimir B. Sukhanovc, Shigeru Suzukid, Mahito Watabed and Natasha S. Viteke
aZoological Institute of the Russian Academy of Sciences, Universitetskaya Emb. 1, St. Petersburg, 199034, Russia; bWaseda University,Nishiwaseda 1-7-14, Shinjuku-ku, Tokyo, 169-0051, Japan; cBorisyak Paleontological Institute of the Russian Academy of Sciences,Profsoyuznaya 123, Moscow, 117997, Russia; dHayashibara Natural Sciences Museum, Okayama, 700�0907, Japan; eDepartment ofGeological Sciences, The University of Texas at Austin, EPS RM 1.130, 1 University Station C9000, Austin, TX 78712-0254, USA
(Received 10 April 2013; accepted 2 June 2013)
This paper is devoted to the description and revision of material of Cretaceous soft-shelled turtles (Trionychidae) ofMongolia. It includes the description of seven trionychid species, six of which are new, and two new genera: thecyclanorbine Nemegtemys conflata gen. et sp. nov. from the Nemegt Formation (Maastrichtian), and the trionychinesGobiapalone breviplastra gen. et sp. nov. from the Nemegt and Barungoyot (Campanian) formations, G. orlovi from theBaynshire Formation (Cenomanian–Santonian), ‘Trionyx’ baynshirensis sp. nov. from the Baynshire Formation, ‘T.’gilbentuensis sp. nov. from the Nemegt Formation, ‘T.’ gobiensis sp. nov. from the Nemegt Formation, and ‘T.’shiluutulensis sp. nov. from an unknown formation (Campanian). In addition, one shell from the ?Baynshire Formation ofKhermin Tsav is assigned to Gobiapalone sp. The type material of Amyda menneri is considered to be Trionychidae indet.and Amyda menneri to be a nomen dubium. Finally, we revise other available materials of Cretaceous trionychids from 45localities in Mongolia. Nemegtemys conflata, if correctly assigned, is the earliest known member of Cyclanorbinae. Thetwo species of the new genus Gobiapalone are included in two phylogenetic analyses of Trionychidae. In both analysesGobiapalone is monophyletic. In the first analysis, Gobiapalone is placed within Apalonina. In the second analysis,Gobiapalone is sister to Apalonina. Thus, the results of both analyses show that Apalonina, which is a rather advanced andwell-supported trionychid clade, or its closest sister taxon (stem-Apalonina), were present in the Late Cretaceous of Asia.These results suggest that most other supra-generic clades of modern trionychids had been established in Asia by the LateCretaceous. That suggestion is supported by the discovery of a cyclanorbine Nemegtemys conflata in the Late Cretaceousof Mongolia. Finally we summarize the latest data on temporal and geographical distributions of Cretaceous Trionychidaeof Asia and North America.
http://zoobank.org/urn:lsid:zoobank.org:pub:921DA1C5-C4B2-463D-A49D-608024C6036A
Keywords: Testudines; Trionychidae; Upper Cretaceous; Asia; Mongolia
Introduction
Trionychidae Grey, 1825, or soft-shelled turtles, are a
group of aquatic cryptodires (Meylan 1987). The phylog-
eny and taxonomy of extinct species within this group are
still not well understood (Meylan 1987; Gardner et al.
1995; Karl 1998; Joyce & Lyson 2011; Vitek 2011,
2012). The lack of understanding is especially problem-
atic for Cretaceous trionychids, which are important for
understanding the early diversification and evolution of
the family (Danilov & Vitek 2012a provided a review of
Cretaceous trionychids of Asia).
This paper continues a series of publications on Creta-
ceous trionychids of Asia (Danilov & Vitek 2009; Vitek
& Danilov 2010, 2012; Danilov & Vitek 2012a, b) and is
devoted to Cretaceous trionychids of Mongolia.
Mongolian specimens from that time are particularly
important because of their geographical and temporal loca-
tion in relation to specimens from North America and the
rest of Asia. That location and the perspective it provides
may help resolve standing questions of trionychid evolu-
tion. Long ghost lineages near the base of the trionychid
tree, especially in regard to the Cyclanorbinae Hummel,
1929, remain problematic. The gap in the cyclanorbine fos-
sil record has recently been filled to some extent by the
discovery that Plastomenidae Hay, 1902 are stem-
cyclanorbines (Joyce & Lyson 2010). Cretaceous plasto-
menids as they are currently known are restricted to North
America, but trionychids probably first evolved in Asia
and extant species of Cyclanorbinae are restricted to Africa
and Asia (Ernst & Barbour 1989; Hutchison 2000). Creta-
ceous representatives of Cyclanorbinae and Plastomenidae
*Corresponding author. Email: [email protected]
� The Trustees of the Natural History Museum, London 2014. All Rights Reserved.
Journal of Systematic Palaeontology, 2014
http://dx.doi.org/10.1080/14772019.2013.847870
Dow
nloa
ded
by [
Igor
G. D
anilo
v] a
t 11:
45 2
1 Fe
brua
ry 2
014

are still unknown from the Cretaceous of Asia. The origin
of cyclanorbines remains unclear, as does the relationship
of plastomenids to the rest of cyclanorbine evolution in
Asia, for which the 80 million year long ghost lineage still
remains.
The example of cylanorbines and plastomenids can be
considered a specific example of the broader problem of
unknown biogeographical patterns within Trionychidae.
Current phylogenetic hypotheses for recently revised
taxa of Trionychidae support two potential hypotheses.
The hypothesis is that trionychids dispersed between
North America and Asia multiple times between the Cre-
taceous and today (Hutchison 2000; Vitek 2011). The
second hypothesis is that multiple species of trionychid
dispersed only once from Asia to North America, but
that most of the diversification leading to the extant gen-
era of trionychids had already occurred (Vitek 2011).
The composition and identity of the Mongolian triony-
chids may help support or refute those hypotheses,
because of the regions that are known to preserve rela-
tively complete Cretaceous fossil material, Mongolia is
closest to any potential land bridge between North
America and Asia.
The first report on Cretaceous trionychids of Mongolia
was published by Khosatzky & M»ynarski (1971), whodescribed ‘Trionyx sp. a’ based on an incomplete carapace
from the Nemegt Formation (Maastrichtian) of Tsagan
Khushu (see Fig. 1 for the position of this and other local-
ities of Cretaceous trionychids of Mongolia). M»ynarski &Narmandach (1972) mentioned a second carapace of
‘Trionyx sp. a’ and shell fragments of ‘Trionyx sp. b’, all
from the Nemegt Formation of Nemegt. Khosatzky
(1976) described Amyda orlovi Khosatzky, 1976 based on
an incomplete carapace from the lower part of the Bayn-
shire Formation (Cenomanian–early Turonian) of Bayn
Shire; the description of this species lacked illustrations.
Earlier, Sochava (1975) mentioned undescribed remains
attributed to Amyda orlovi (according to identifications by
L.I. Khosatzky) from the Baynshire Formation of Khara
Khutul. Undescribed records of trionychids from several
localities of the Baynshire (Cenomanian–Santonian) and
Nemegt formations of Mongolia were reported by
Figure 1. Map showing all known occurrences of Cretaceous Trionychidae in Mongolia: 1, Abdarain Nuru; 2, Abdrant Nuru; 3, AlakShand Khuduk; 4, Altan Ula, Altan Ula I, III and IV; 5, Amtgai; 6, Bayn Shire; 7, Bamba Khuduk; 8, Bayn Dzak; 9, Baishin Tsav; 10,Bugin Tsav and Bugin Tsav II; 11, Burkhant; 12, Char Teeg; 14, Dzun Shakhai; 16, Erdeni Ula; 17, Gilbentu; 18, Gurilin Tsav; 19,Ingeni Khobur and Ingeni Khobur III; 20, Ingeni Tsav; 22, Khara Khutul; 23, Khermin Tsav; 26, Khongil Tsav; 27, Khuren Tsav; 28,Nemegt; 29, Nogon Tsav and Nogon Tsav II; 30, Ongon Ulan Ula; 31, Shiluut Ula; 32, Shine Us Khuduk; 33, Shine Usny Tolgod; 34,Shiregin Gashun; 35, Tel Ulan Shalcha; 36, Tsagan Khushu; 37, Tsagan Teg; 39, Ulan Khushu; 40, Ulan Tsab Ula; 43, Ushyin Khuduk;44, Yagaan Khovil. Localities with uncertain position (13, 15, 21, 24, 25, 38, 41, 42, and 45) are not shown. See Online SupplementaryMaterial Appendix 1 for data on geology, age and material.
2 I. G. Danilov et al.
Dow
nloa
ded
by [
Igor
G. D
anilo
v] a
t 11:
45 2
1 Fe
brua
ry 2
014

Shuvalov & Chkhikvadze (1975, 1979). Chkhikvadze &
Shuvalov (1988) described Amyda menneri Chkhikvadze
in Chkhikvadze & Shuvalov, 1988 based on shell frag-
ments from the Nemegt Formation of Gurilin Tsav and
other localities. Khosatzky (1999) reported new triony-
chid shell material from several localities of the Lower
Cretaceous, and from the Baynshire, Barungoyot (Campa-
nian) and Nemegt formations of the Upper Cretaceous.
Sukhanov (2000) published illustrations of the holotype
of Amyda orlovi. Suzuki & Narmandakh (2004) reported
additional trionychid material from several Cretaceous
localities of Mongolia. Suzuki (2005) reported an almost
complete skeleton of Apalonina from an unreported local-
ity and formation of the Upper Cretaceous of Mongolia.
Data on Cretaceous trionychids of Mongolia were
included in reviews of published and/or figured material
of Cretaceous trionychids of Asia (Danilov & Vitek 2009,
2012a). Finally, Danilov et al. (2011a, b) reported prelim-
inary results of a revision of Cretaceous trionychids of
Mongolia.
Gilmore (1931) in his article entitled ‘Fossil turtles of
Mongolia’ mentioned fragmentary shell remains of trio-
nychids from the Cretaceous of the On Gong (Aptian–
Albian) and Iren Dabasu (?Campanian) formations.
Actually, these remains come from the territory of Inner
Mongolia, China, and are not considered in our study.
Here we revise, describe and illustrate material of Cre-
taceous trionychids of Mongolia (Figs 3–16), including
seven trionychid species, six of which are new, and two
new genera: the cyclanorbine Nemegtemys conflata gen.
et sp. nov. and the trionychines Gobiapalone breviplastra
sp. nov., G. orlovi, ‘Trionyx’ baynshirensis sp. nov., ‘T.’
gilbentuensis sp. nov., ‘T.’ gobiensis sp. nov. and ‘T.’ shi-
luutulensis sp. nov. In addition, one shell from the ?Bayn-
shire Formation of Khermin Tsav is assigned to
Gobiapalone sp. The type material of Amyda menneri is
considered to be Trionychinae indet. and Amyda menneri
to be a nomen dubium. Finally, we revised other available
material of Cretaceous trionychids from 45 localities of
Mongolia (Fig. 1, Table 1, Online Supplementary Mate-
rial Appendix 1).
The taxonomy of Trionychidae used here generally fol-
lows Meylan (1987), who presented the first comprehen-
sive phylogenetic analysis for living trionychids based on
skeletal morphology and provided diagnoses for the fam-
ily and all less inclusive taxa. Because Meylan’s (1987)
work relied on skeletal characters, it is especially useful
for dealing with fossil specimens and taxa. More recent
attempts to develop a phylogenetic taxonomy of triony-
chids (Engstrom et al. 2004; Joyce et al. 2004) are less
useful for Cretaceous taxa, because these studies also use
non-skeletal features and because few specimens of
Cretaceous trionychids have been included in cladistic
analyses. Anatomical terminology follows Meylan
(1987), Gardner & Russell (1994) and Karl (1999). Table1.ComparisonofshellcharactersofsomespeciesofCretaceoustrionychids.
Characters
‘Aspideretes’
maortuensis
Aspideretoides
riabinini
Gilmoremys
lancensis
Gobiapalone
breviplastra
Gobiapalone
orlovi
‘Trionyx’
gobiensis
‘Trionyx’
kansaiensis
‘Trionyx’
kyrgyzensis
‘Trionyx’
shiluutulensis
Maxim
um
carapacelength
(mm)
300�
500�
340
260�
335
130�
750�
150�
225
Nuchalem
argination
?Weak
Absent
Weakorabsent
Weakorabsent
Weakorabsent
Strong
Absent
Weak
Preneural
?Present
Present
Absent
Absent
Absent
Absent
?Present
Number
ofneurals
87
77or8or9
7or8
7or8
7or8
88
Neuralreversal
55or6
65
55
55
6Costals8
Small
Small
Large
Smallorabsent
Small
Large
Large
Large
Small
Epiplastralnotchonhyoplastron
Absent
Absent
Present
Absent
Absent
?Absent
Absent
?Medialprocesses
ofhyoplastron
Present
Present
Absent
Present
Present
?Present
Present
?Lateralhyoplastronlobein
relation
tomedialhyoplastronlobe
?Shorter
Alm
ostequal
Shorter
Shorter
?Longer
Longer
?
Ratio
ofminim
albridgelength
tomaxim
alhypoplastronlength
?About50%
About100%
About50%
About50%
?50–60%
About50%
?
Extensivemedialcontactofhyo-and
hypoplastra
No
No
No
No
No
?No
No
?
Medialhypoplastralprocesses
Clustered
Divided
?Divided
Divided
?Clustered
Clustered
?Number
ofplastralcallosities
Atleast2
Atleast4
Atleast4
55
?Atleast2
Absent
?
� Estim
ation
Cretaceous soft-shelled turtles (Trionychidae) of Mongolia 3
Dow
nloa
ded
by [
Igor
G. D
anilo
v] a
t 11:
45 2
1 Fe
brua
ry 2
014

Material and methods
We examined, either first hand or based on photographs,
most of the available material of Cretaceous trionychids
of Mongolia collected by the Mongolian Palaeontological
Expedition of Academy of Sciences of the USSR (MPE,
1946, 1948, 1949), the Polish-Mongolian Palaeontologi-
cal Expedition (PMPE, 1963–1971), the Joint Soviet-
Mongolian Palaeontological Expedition (JSMPE, 1969–
1991), and the Japan-Mongolia Joint Palaeontological
Expedition (JMJPE, 1993–2008), as well as material col-
lected by Russian and Mongolian geologists and biolo-
gists working in Mongolia in the twentieth century: V. N.
Chaykovskiy, R. Barsbold, L. Ya. Borkin, G. G. Martin-
son, I. Yu. Neustroeva, V. F. Shuvalov, A. V. Sochava
and N. N. Verzilin. However, we did not manage to exam-
ine material reported and described by Shuvalov &
Chkhikvadze (1975, 1979) and Chkhikvadze & Shuvalov
(1988), including type material of Amyda menneri, because
this material is not accessible for study (V. M. Chkhik-
vadze pers. comm. to I.G.D., 2012). Specimens collected
by JMJPE belong to the MPC, but are temporary stored in
the HMNS, and have both MPC and HMNS numbers.
The shell material described in our paper demonstrates
two main types of sculpturing on the shell surface, which
are referred to as type A and type B. The sculpturing of
type A is a pattern of thin, connected ridges forming a
honeycomb or netlike pattern. The sculpturing of type B
is a pattern of wide disconnected ridges and tubercles
raised above the plate. These types of sculpturing were
previously noted for Aspideretoides riabinini (Kuznetsov
& Chkhikvadze, 1987) (type A) and ‘Trionyx’ kansaiensis
Vitek & Danilov, 2010 (type B) from the Late Cretaceous
of Kazakhstan and Tajikistan (Fig. 2; see Vitek & Danilov
2010).
In order to determine the phylogenetic position of spe-
cies of Gobiapalone we performed two phylogenetic anal-
yses of trionychids, using the character/taxon matrices of
Joyce & Lyson (2011), and Vitek (2012), hereinafter anal-
yses 1 and 2 respectively. Characters coded for Gobiap-
alone breviplastra sp. nov. and G. orlovi in each of the
analyses are given in Online Supplementary Material
Appendices 2 and 3. The final data matrices include 83
characters for 28 taxa in analysis 1, and 88 characters for
41 taxa in analysis 2. In each analysis, Gobiapalone brevi-
plastra is 46% (analysis 1) and 49% (analysis 2) com-
plete, and G. orlovi is 80% (analysis 1) and 81% (analysis
2) complete. Each matrix was assembled using NDE 0.5.0
(Page 2001) and analysed with NONA ver. 2 and
Winclada ver. 1.00.08 by Ratchet algorithm with 1000
iterations. All characters were run unordered, unweighted,
and with no topological constraints. Support for each
node was measured by calculating Bremer supports
(Eriksson 1998).
Institutional abbreviationsHMNS: Hayashibara Museum of Natural Sciences,
Okayama, Japan; IPGAS: Institute of Paleobiology, Geor-
gian Academy of Sciences, Tbilisi, Georgia; MNHN:
Mus�eum national d’Histoire naturelle, Paris, France;
MPC: Mongolian Palaeontological Centre, Ulan Bator,
Mongolia; PIN: Borissyak Palaeontological Institute of
the Russian Academy of Sciences, Moscow, Russia; ZIN
PH: Paleoherpetological collection, Zoological Institute
of the Russian Academy of Sciences, St. Petersburg, Rus-
sia; ZPAL: Institute of Palaeobiology, Polish Academy of
Sciences, Warsaw, Poland. The term ‘Collection’ immedi-
ately preceding an institutional catalogue number
(e.g. Collection ZIN PH 156) indicates that multiple
specimens are accessioned under that number.
Systematic palaeontology
Testudines Batsch, 1788
Cryptodira Cope, 1868
Trionychidae Gray, 1825
Cyclanorbinae Hummel, 1929
Nemegtemys gen. nov.
Derivation of name. Nemegt, for the Nemegt locality,
and emys, Greek for turtle.
Type species. Nemegtemys conflata sp. nov.
Diagnosis. Same as for the type species.
Nemegtemys conflata sp. nov.
(Fig. 3)
1999 ‘Plastomenus’?; Khosatzky: 147.
2009 Trionychinae indet. 6; Danilov & Vitek: 54 (part.).
2012a Trionychidae indet. 5; Danilov & Vitek: 429
(part.).
Figure 2. Two main types of shell sculpturing of Cretaceoustrionychids of Asia. A, type A (ZIN PH 822/64, Aspideretoidesriabinini, costal 1); B, type B (ZIN PH 917/64, ‘Trionyx’kansaiensis, partial costal).
4 I. G. Danilov et al.
Dow
nloa
ded
by [
Igor
G. D
anilo
v] a
t 11:
45 2
1 Fe
brua
ry 2
014

Derivation of name. Species name, conflata, Latin for
fused, in reference to fused hyo- and hypoplastra.
Holotype. ZIN PH 1/157, lateral fragment of left hyo-
and hypoplastra, from Nemegt, Umunugovi Aimag, Mon-
golia, Nemegt Formation, Maastrichtian (collected by
MPE in 1948).
Referred material. MPC 25/161 (HMNS 2006-4-472),
lateral fragment of right hypoplastron, from Bugin Tsav,
Umunugovi Aimag, Mongolia, Nemegt Formation, Maas-
trichtian (collected by JMJPE in 2006).
Diagnosis. A cyclanorbine which can be differentiated
from all other cyclanorbines in that, at the stage when the
hyo- and hypoplastra are completely ossified (i.e. when
all their processes are covered by metaplastic bone), the
hyoplastral lappet is small, forming an almost straight
anterior margin of the hyoplastron. In all other cyclanor-
bines, at the similar stage of ossification, the hyoplastral
lappet is larger (more extensive anteroposteriorly), form-
ing the rounded anterior margin (shoulder) of the
hyoplastron.
Description. ZIN PH 1/157 is a lateral fragment of the
left hyo- and hypoplastra. It is 4.5 cm long (from the ante-
rior margin of the lateral hyoplastron to the posterior mar-
gin of the lateral hypoplastron). The estimated length of
the plastron (based on comparison with Cycloderma
aubryi, MNHN 1930-362) is about 130 mm. The hyo-
and hypoplastra are fused and well-ossified. There are two
lateral processes on both the hyo- and hypoplastra (visible
on the upper surface of the fragment). These processes are
almost completely covered by metaplastic bone and proj-
ect only slightly beyond it (this is visible on the processes
of the hypoplastron and reconstructed for the processes of
the hyoplastron, which are broken off). The lateral
margins of the hyo- and hypoplastra are almost straight
and were probably parallel to the midline of the plastron.
The anterior margin of the hyoplastron (and hyoplastral
lappet) is straight and perpendicular to the lateral margin
of the hyo- and hypoplastra. The hyoplastral lappet makes
up about half of the bridge length and is shorter anteropos-
teriorly than in other cyclanorbines. The plastral callosity
covers most of the external surface of the fragment, except
for a narrow band along the anterior and lateral margins of
the specimen. The sculpturing on the external surface is
made of a fine net of ridges and tubercles; the latter are
strongly expressed at the posterior margin of the
specimen.
MPC 25/161 is a lateral fragment of the right hypoplas-
tron from a slightly larger individual than ZIN PH 1/157.
The fragment is well ossified, probably had a straight lat-
eral margin. It has a pair of lateral processes, which pro-
trude only slightly beyond the metaplastic bone. The
plastral callosity completely covers the external surface of
the specimen. The sculpturing of the external surface is
similar to ZIN PH 1/157, although tubercles are absent in
MPC 25/161.
Remarks. ZIN PH 1/157 was mentioned by Khosatzky
(1999, p. 147) as a “plastron fragment of a young triony-
chid individual, characterized by a rapid extension of the
plastron in the ontogenesis, and by a sculpturing of the
external surface with tubercles (‘Plastomenus’?)”. Dani-
lov & Vitek (2009, 2012a) erroneously listed
‘Plastomenus’? amongst the synonymies of their Triony-
chinae indet. 6 and 5 respectively, which are herein con-
sidered to be synonymies of ‘Trionyx’ gilbentuensis sp.
nov. (see below). The most peculiar character of ZIN PH
1/157 is the fusion of the hyo- and hypoplastra at a small
size. According to Meylan (1987), such a fusion (just after
hatching) is a synapomorphy of recent cyclanorbines. In
Figure 3. Shell material of Nemegtemys conflata gen. et sp. nov. A, B, ZIN PH 1/157 (holotype), a lateral fragment of hyo- andhypoplastra from the Nemegt Formation of Nemegt in A, dorsal and B, ventral views; C, D, MPC 25/161, a lateral fragment of the righthypoplastron from Bugin Tsav in C, ventral and D, dorsal views. Reconstructed outlines are indicated by dashed lines.
Cretaceous soft-shelled turtles (Trionychidae) of Mongolia 5
Dow
nloa
ded
by [
Igor
G. D
anilo
v] a
t 11:
45 2
1 Fe
brua
ry 2
014

trionychines, the fusion of the hyo- and hypoplastra, if
present, occurs only in adults (at large size) of some spe-
cies. In the clade Plastomenidae, which is considered to
be stem Cyclanorbinae, the fusion of the hyo- and hypo-
plastra is not known (Hutchison 2009; Joyce et al. 2009;
Joyce & Lyson 2010, 2011).
Trionychinae Gray, 1825
Trionychini Gray, 1825
Gobiapalone gen. nov.
Derivation of name. Gobi-, for the Gobi Desert, and
Apalone Rafinesque, 1832, a genus of North American
trionychids.
Type species. Gobiapalone orlovi (Khosatzky, 1976)
comb. nov.
Other species. Gobiapalone breviplastra sp. nov.
Diagnosis. A member of Trionychini which is most simi-
lar to members of the clade Apalonina and to Oliveremys
uintaensis (Leidy, 1872) in the presence of a vomer that
divides the maxillae and reaches the intermaxillary fora-
men, and small or absent costals 8. It can be differentiated
from all other members of Trionychini and other Creta-
ceous trionychids with known shell morphology by the
presence of five plastral callosities. In addition, it can be
differentiated from Apalone spp. by an apertura narium
externa weakly emarginated laterally and an unfused hyo-
and hypoplastra; from Aspideretoides spp. by a narrower
and longer crista supraoccipitalis, a wider dentary sym-
physis, absence of a dentary pocket and absence of a pre-
neural; from Axestemys spp. by smaller size, a vomer that
does not divide the maxillae and reach the intermaxillary
foramen, absence of a dentary pocket, absence of a pre-
neural and two lateral hyoplastral processes; from
Oliveremys uintaensis by a larger intermaxillary foramen,
a smaller carapace size, less ossified hyo- and hypoplastra,
and xiphiplastra with sculptured callosities; from Trionyx
triunguis (Forska�l, 1775) by a larger intermaxillary fora-
men, and smaller costals 8. See Table 1 for further differ-
ences between Gobiapalone spp. and Cretaceous
trionychids.
Gobiapalone orlovi (Khosatzky, 1976), comb. nov.
(Figs 4–9)
1975 Amyda orlovi Khos.; Sochava: 115 (nomen
nudum).
1976 Amyda orlovi Khosatzky; Khosatzky: 6.
1988 Amyda orlovi Khosatzky; Chkhikvadze & Shuvalov:
200.
1999 Amyda sp. cf. A. orlovi; Khosatzky: fig. 1.
1999 Trionychina indet.; Khosatzky: 143, fig. 2.
1999 Trionychidae indet.; Khosatzky: 144, fig. 3.
2000 Amyda orlovi Khosatzky; Sukhanov: 345,
fig. 17.27.
2000 ‘Amyda’ orlovi Khosatzky; Sukhanov: 345.
2005 Apalonina; Suzuki: 27.
2009 ‘Amyda’ orlovi Khosatzky; Danilov & Vitek: 54.
2009 Trionychinae indet. 3; Danilov & Vitek: 54
2009 Trionychinae indet 4; Danilov & Vitek: 54.
2010 ‘Amyda’ orlovi Khosatzky; Vitek & Danilov: table 1,
fig. 1.
2011a Amyda orlovi Khosatzky; Danilov et al.: 15.
2011b ‘Amyda’ orlovi Khosatzky; Danilov et al.: 96.
2011b Trionychinae indet.; Danilov et al.: 96.
2012a Apalonina indet. 3; Danilov & Vitek: 421.
2012a ‘Amyda’ orlovi Khosatzky; Danilov & Vitek: 423,
fig. 23.2b.
2012a Trionychinae indet. 3; Danilov & Vitek: 429,
fig. 23.3e.
2012a Trionychinae indet 4; Danilov & Vitek: 429,
fig. 23.3f.
Holotype. PIN 557-132/1 (formerly PIN 557-1/1),
incomplete carapace, from Bayn Shire, Dornogovi Aimag,
Mongolia, lower part of the Baynshire Formation,
Cenomanian–early Turonian (collected by MPE in 1948).
Referred material. MPC 25/162 (HMNS 94-10-9), car-
apace (collected by JMJPE in 1994); MPC 25/160
(HMNS 96-12-21-25), partial skeleton (collected by
JMJPE in 1996), all from Burkhant, Dornogovi Aimag,
Mongolia, Baynshire Formation, Cenomanian–Santonian;
PIN 557-133, fragments of carapace on plaster, from
Bayn Shire, Dornogovi Aimag, Mongolia, lower part of
Baynshire Formation, Cenomanian–early Turonian (col-
lected by MPE); ZIN PH T/M46-1, partial hyo- and
hypoplastra, from Bayn Shire, Dornogovi Aimag, Mon-
golia, lower part of Baynshire Formation, Cenomanian–
early Turonian (collected by MPE in 1946); ZIN PH T/
M67-5, anterior part of carapace, from Unegetu Ula,
Dornogovi Aimag, Mongolia, Baynshire Formation, Cen-
omanian–Santonian (collected by A.V. Sochava in
1967); ZIN PH T/M71-1, incomplete carapace, from
Khongil, Dornogovi Aimag, Mongolia, Baynshire For-
mation, Cenomanian–Santonian (collected by V. F.
Shuvalov in 1971).
Diagnosis. A species of Gobiapalone with a carapace
length up to 335 mm, which can be differentiated from
G. breviplastra by the closed suprascapular
fontanelles in specimens larger than 275 mm in cara-
pace length, slender epiplastra and entoplastron, a
smaller (about 80�) angle between arms of entoplastron,
more numerous (five) medial processes of hyoplastron,
a bigger gap between anterior and posterior groups of
medial processes of hypoplastron, and slender
xiphiplastra.
6 I. G. Danilov et al.
Dow
nloa
ded
by [
Igor
G. D
anilo
v] a
t 11:
45 2
1 Fe
brua
ry 2
014

Figure 4. Skull of Gobiapalone orlovi (Khosatzky, 1976) (MPC 25/160) from the Baynshire Formation of Burkhant. A, dorsal view; B,ventral view; C, right lateral view; D, left lateral view; E, partial right ventrolateral view; F, anterior view; G, posterior view; H, partialposteroventral view; I, partial posterodorsal view.
Cretaceous soft-shelled turtles (Trionychidae) of Mongolia 7
Dow
nloa
ded
by [
Igor
G. D
anilo
v] a
t 11:
45 2
1 Fe
brua
ry 2
014

Figure 5. Skull of Gobiapalone orlovi (Khosatzky, 1976) (MPC 25/160) from the Baynshire Formation of Burkhant, explanation draw-ings: A, dorsal view; B, ventral view; C, right lateral view; D, left lateral view; E, partial right ventrolateral view; F, anterior view;G, posterior view; H, partial posteroventral view; I, partial posterodorsal view. Bones are filled with grey (foreground) and dark grey(background). Matrix or gypsum is stippled. Broken surfaces are hatched. Reconstructed sutures and outlines are indicated by dashedlines. Abbreviations: ane, apertura narium externa; bo, basioccipital; bs, basisphenoid; co, condylus occipitalis; ex, exoccipital; fi,foramen intermaxillaris; fjp, foramen jugulare posterius; fm, foramen magnum; fnh, foramen nervi hypoglossi; fpcci, foramen posteriuscanalis carotici interni; fpo, fenestra postotica; fpp, foramen palatinum posterius; fr, frontal; fst, foramen stapedio-temporalis; ica,incisura columellae auris; ina, internal narial opening; ju, jugal; mx, maxilla; op, opisthotic; pal, palatine; pf, prefrontal; po, postorbital;pro, prootic; pt, pterygoid; qj, quadratojugal; qu, quadrate; so, supraoccipital; sq, squamosal; vo, vomer.
8 I. G. Danilov et al.
Dow
nloa
ded
by [
Igor
G. D
anilo
v] a
t 11:
45 2
1 Fe
brua
ry 2
014

Description
Skull. The skull is known from a single specimen (MPC
25/160; Figs 4, 5). It is almost complete, but damaged due
to partial deformation and poor preservation. As a result, the
skull is covered with numerous cracks filled with matrix;
some breaks were restored with gypsum. Many sutures are
not distinct or barely visible. The premaxillae are absent, as
well as part of the left jugal arch. From the anteriormost tip
of the maxillae to the posteriormost end of the supraoccipi-
tal, the skull is about 14 cm long. The condylobasal length
of the skull is about 10 cm. The width of the skull at the
Figure 6. A–F, Lower jaw, and G, left cornu branchiale I of Gobiapalone orlovi (Khosatzky, 1976) (MPC 25/160) from the BaynshireFormation of Burkhant. A, D, left lateral view; B, E, left dorsolateral view; C, F, dorsal view; G, dorsal view. Bones are filled withgrey. Matrix or gypsum is stippled. Broken surfaces are hatched. Reconstructed sutures and outlines are indicated by dashed lines.Abbreviations: an, angular; ar, articular; cor, coronoid; den, dentary; fai, foramen alveolare inferius; fme, fossa meckelii; pra, prearticu-lar; sur, surangular.
Cretaceous soft-shelled turtles (Trionychidae) of Mongolia 9
Dow
nloa
ded
by [
Igor
G. D
anilo
v] a
t 11:
45 2
1 Fe
brua
ry 2
014

level of the quadrates is about 7.5 cm. Thus, the width/con-
dylobasal length ratio is 0.75. The face of the skull is rela-
tively wide and short. The orbits are relatively large and
face mainly dorsally. The ratio of the interorbital distance to
the length of the orbit is about 0.30.
The prefrontal contacts the frontal posteriorly and the
maxilla laterally, makes up the anterodorsal part of the
margin of the orbit between the frontal and maxilla, and
forms the dorsal edge of the apertura narium externa. A
descending process of the prefrontal was probably
Figure 7. Shell material of Gobiapalone orlovi (Khosatzky, 1976). A–D,MPC 25/160 from the Baynshire Formation of Burkhant: A, B,carapace in A, dorsal and B, ventral views; C, D, plastron in C, dorsal and D, ventral views. E, MPC 25/162, carapace from the Bayn-shire Formation of Burkhant in dorsal view. F, PIN 557-132/1, carapace (holotype) from the lower part of the Baynshire Formation ofBayn Shire in dorsal view. G, PIN 557-133, fragments of carapace on plaster from the lower part of the Baynshire Formation of BaynShire in dorsal view. H, ZIN PH T/M46-1, partial hyo- and hypoplastra from the lower part of the Baynshire Formation of Bayn Shire inventral view. I, ZIN PH T/M71-1, carapace from the Baynshire Formation of Khongil in dorsal view. J, ZIN PH T/M67-5, anterior partof the carapace from the Baynshire Formation of Unegetu Ula in dorsal view. Reconstructed sutures and outlines are indicated by dashedlines.
10 I. G. Danilov et al.
Dow
nloa
ded
by [
Igor
G. D
anilo
v] a
t 11:
45 2
1 Fe
brua
ry 2
014

present within the orbit, but it is not discernible from the
surrounding bones due to poor preservation. The
anterior margin of the prefrontal (i.e. the dorsal edge of
the apertura narium externa) is weakly emarginated
laterally as in many trionychids, but not at all medially
emarginated.
The frontal contacts the prefrontal anteriorly, the parie-
tal posteriorly, and the postorbital posterolaterally, and
forms a majority of the dorsal border of the orbit between
the prefrontal and postorbital. The suture between the
frontal and prefrontal is slightly oblique and the prefrontal
therefore projects slightly between the frontals. The suture
between the frontal and parietal is perpendicular to the
midline and partially interdigitated. The suture between
the frontal and postorbital is clearly visible only on the
right side of the skull, whereas on the left side that area is
damaged. The ventral surface of the frontals is not visible
and its condition (fused or unfused; see Joyce & Lyson
2011) cannot be established.
The dorsal plate of the parietal contacts the frontal ante-
riorly, the postorbital anterolaterally on the skull roof,
the jugal anterolaterally within the fossa temporalis, and
the supraoccipital posteriorly. The parietal does not con-
tribute to the orbital margin or walls. The anterior contacts
of the ventral plate of the parietal are unclear, whereas
posteriorly the parietal contacts the prootic and supraocci-
pital. The contribution of the ventral plate of the parietal
to the margin of the foramen trigemini is not clear. The
contribution of the ventral plate of the parietal to the
formation of the processus trochlearis oticum is about
one-fourth of the total width.
The postorbital contacts the frontal medially, the parie-
tal posteriorly and the jugal laterally. The postorbital con-
tributes to the posterior rim of the orbit and to the anterior
rim of the upper temporal emargination. The postorbital
bar is about a quarter of the orbit diameter.
On the external surface of the skull, the jugal contacts
the maxilla anteriorly and the postorbital anterodorsally,
and the quadratojugal posteriorly. The ventral contact
between the jugal and the parietal is clearly visible within
the fossa temporalis. Whether the jugal contributes to the
orbital rim is not clear.
The quadratojugal contacts the jugal anteriorly, and the
squamosal and quadrate posteriorly, and contributes to the
anterior rim of the cavum tympani. The quadratojugal
does not participate in the processus trochlearis oticum.
The squamosal contacts the quadratojugal anteriorly,
the quadrate ventrally and anteromedially, and the
Figure 8. Cervical vertebrae of Gobiapalone orlovi (Khosatzky, 1976) (MPC 25/160) from the Baynshire Formation of Burkhant, in A,ventral, B, left lateral and C, dorsal views. Reconstructed outlines are indicated by dashed lines.
Figure 9. Girdles of Gobiapalone orlovi (Khosatzky, 1976)(MPC 25/160) from the Baynshire Formation of Burkhant. A,right pectoral girdle in posterodorsal view; B, right pelvic girdlein dorsal view. Reconstructed outlines are indicated by dashedlines.
Cretaceous soft-shelled turtles (Trionychidae) of Mongolia 11
Dow
nloa
ded
by [
Igor
G. D
anilo
v] a
t 11:
45 2
1 Fe
brua
ry 2
014

opisthotic posteromedially. In lateral view, the squamosal
forms the roof of the cavum tympani as well as much of
the antrum postoticum.
The premaxillae are not preserved.
In dorsal view, the maxilla contacts the prefrontal ante-
romedially and the jugal posteriorly, and forms the lateral
margin of the apertura narium externa and the ventral mar-
gin of the orbit. The maxilla does not contact the frontal or
the quadratojugal. In ventral view, the maxilla forms the
primary palate and probably contacted the premaxilla ante-
riorly. The maxilla contacts the vomer anterior and the pal-
atine and pterygoid posterior to the choana. The foramen
intermaxillaris is present and makes up about 60% of the
length of the primary palate. The maxilla forms most of the
margin of that foramen. The maxilla does not participate in
the formation of the foramen palatinum posterius. The trit-
urating surface formed by the maxilla is triangular-shaped,
narrowed anteriorly and expanded posteriorly.
The vomer is shaped peculiarly, with an expanded ante-
rior plate and a narrow posterior process. The vomer con-
tacts the maxillae laterally and the palatines posteriorly. It
fully divides the maxillae and contributes to the formation
of the foramen intermaxillaris anteriorly and internal
narial openings posteriorly.
The palatines are only observable in ventral view. Each
palatine contacts its counterpart medially, the vomer ante-
romedially, the maxilla anterolaterally, the pterygoid lat-
erally and the basisphenoid posteriorly. The palatine
forms the posterior rim of the choana and, together with
the pterygoid, contributes to the formation of the small
foramen palatinum posterius. Other contacts and contribu-
tions of the palatine are not observable.
In ventral view, each pterygoid contacts the maxilla
anteriorly, the palatine, basisphenoid and basioccipital
medially, and the quadrate posterolaterally. The foramen
palatinum posterius is positioned between the anterior
portion of the pterygoid and the palatine. The foramen
posterius canalis carotici interni is positioned below the
lateral crest of the basioccipital tubercle and is completely
enclosed within the pterygoid. In posterior view, the pter-
ygoid contacts the opisthotic and quadrate and contributes
to the formation of the fenestra postotica. The contact of
the pterygoid with the exoccipital is not clear. The con-
tacts and contributions of the pterygoid to the lateral wall
of the braincase are not observable.
The epipterygoid and the trigeminal foramen are not
discernable due to poor preservation of the specimen.
The basisphenoid is trapezoidal in shape, narrowed ante-
riorly and widened posteriorly. It is not medially con-
stricted. The basisphenoid contacts the palatines anteriorly,
the pterygoids laterally and the basioccipital posteriorly.
The prootic forms much of the processus trochlearis oti-
cum. It contacts the parietal medially, the supraoccipital
posteromedially, the quadrate laterally and the opisthotic
posteriorly. The foramen stapedio-temporalis is
positioned between the prootic and the quadrate. Other
contacts and contributions of the prootic are not
observable.
In dorsal view, the opisthotic forms the posterior mar-
gin of the otic capsule and contacts the prootic anteriorly,
the quadrate and squamosal laterally and the supraoccipi-
tal posteromedially. In posterior view, the opisthotic con-
tacts the supraoccipital and exoccipital medially, and the
squamosal and quadrate laterally. The descending process
of the opisthotic excludes the foramen jugulare posterius
from the fenestra postotica only on the right side of the
skull.
In dorsal view the quadrate contacts the quadratojugal
anteriorly, the prootic anteromedially, the opisthotic post-
eromedially and the squamosal posteriorly. The quadrate
contributes minimally to the formation of the processus
trochlearis oticum and to the lateral rim of the foramen
stapedio-temporalis. In lateral view, the quadrate forms
most of the subtriangular cavum tympani and the articular
surface for the lower jaw. Within the cavum tympani, the
quadrate contacts the squamosal dorsally. Anterior to the
cavum tympani, the quadrate contacts the quadratojugal.
In ventral view, the quadrate contacts the pterygoid medi-
ally. In posterior view, the quadrate contacts the squamo-
sal dorsally, the opisthotic dorsomedially, and the
pterygoid ventromedially.
In ventral view, the basioccipital contacts the basisphe-
noid anteriorly and the pterygoids laterally. The basiocci-
pital forms a pair of distinct tubercula basioccipitale. In
posterior view, the basioccipital forms part of the condy-
lus occipitalis and contacts the exoccipitals laterally.
The exoccipitals form the dorsal part of the condylus
occipitalis. In posterior view, they contact the supraocci-
pital and opisthotic dorsally and the basioccipital ven-
trally. The single foramen nervi hypoglossi is visible on
the left exoccipital.
The supraoccipital projects from the posterior end of
the skull in a long process (crista occipitalis). The crista
occipitalis is T-shaped in cross section. In dorsal view, the
supraoccipital contacts the parietals anteriorly, the prootic
anterolaterally and the opisthotic laterally. In posterior
view, the supraoccipital makes up the dorsal margin of
the foramen magnum and contacts the exoccipitals ven-
trally and the opisthotic ventrolaterally.
Mandible. The mandible is almost complete, but like the
skull, is covered with a net of cracks. In spite of these
cracks, most sutures are clearly visible.
The anterior margin of the dentary is rounded in dorsal
view. The triturating surface of the dentary is flat, lacks
lingual or labial ridges, and is expanded (the length of its
symphyseal region is about 25% of the total length of the
mandible). In dorsal view, the dentary contacts the coro-
noid posterolaterally. In lateral view, the dentary produces
an ascending process that covers the anterior aspect of the
12 I. G. Danilov et al.
Dow
nloa
ded
by [
Igor
G. D
anilo
v] a
t 11:
45 2
1 Fe
brua
ry 2
014

coronoid process. Posterior to the coronoid process, the
dentary contacts the coronoid anterodorsally, the surangu-
lar posterodorsally and the articular posteriorly. In medial
view, the dentary contacts the prearticular and angular
below the coronoid process. There are a number of rather
small foramina penetrating the dentary below the coro-
noid process, one of which probably represents the fora-
men dentofaciale majus.
The angular, as seen in the medial view of the right
ramus of the mandible, is broken and represented only by
its anterior half. It contacts the dentary ventrally, and
probably had a dorsal contact with the prearticular.
The surangular is clearly visible on the lateral side of
the mandible. It contacts the dentary anteriorly and the
coronoid anterodorsally. Contact between the surangular
and the angular is not observable due to breakage of the
latter. The foramen nervi auriculotemporalis is visible on
the surangular just anterior to the area articularis mandibu-
laris. In dorsal view, the surangular frames the lateral mar-
gin of the fossa Meckelii and forms the lateral part of the
area articularis mandibularis and the retroarticular process.
The coronoid forms most of the coronoid process, but
does not contribute to the triturating surface. It is over-
lapped anteriorly by the ascending process of the dentary.
The coronoid contacts the surangular lateral to and the
prearticular medial to the fossa Meckelii, respectively.
The prearticular is broken along the probable ventral
contact with the angular. It contacts the coronoid antero-
dorsally, the dentary anteriorly and the articular posteri-
orly, and forms the medial wall of the fossa Meckelii. The
contribution of the prearticular in the formation of the
foramen intermandibularis caudalis is unclear.
The articular contacts the surangular and dentary anteri-
orly and forms the medial part of the area articularis man-
dibularis and retroarticular process.
Hyoid. The hyoid is represented by a single element, the
left cornu branchiale I. In its outline it is most similar to
those of Aspideretes hurum (Gray, 1831) (see Meylan
1987, fig. 18C).
Carapace. The disc of the carapace is slightly longer
than wide in PIN 557-132/1 and about as long as wide in
other specimens. The anterior border of the carapace is
rounded, either with a weak nuchal emargination (MPC
25/160 and ZIN PH T/M71-1) or with no emargination
(MPC 25/162). The lateral borders of the carapace are
either smooth, or sometimes slightly scalloped. The poste-
rior margin of the carapace can be rounded (MPC 25/162
and MPC 25/160), straight (ZIN PH T/M71-1), or slightly
emarginated (PIN 557-133). The free end of the ribs,
when preserved, project far beyond the carapace disc even
in large specimens. Suprascapular fontanelles are present
in small and medium specimens (MPC 25/160, PIN
557-132/1 and PIN 557-133), and absent in the large
specimens (MPC 25/162, ZIN PH T/M71-1 and probably
ZIN PH T/M67-5). All specimens have type A sculptur-
ing. See Table 2 for variation in some carapace characters
of Gobiapalone orlovi.
The nuchal is more than four times wider than long (the
nuchal length/width ratio varies from about 7.0 in MPC
25/160 to about 5.0 in MPC 25/162). Its anterior margin is
not emarginated, and more (MPC 25/162) or less (MPC
25/160) rounded. As is visible in ventral view of MPC 25/
160, the costiform processes are united. The posterolateral
aspects of the nuchal are covered by the expansions of the
free ribs of costals 1.
The neural series is fully preserved in all specimens
except PIN 557-133, in which only the presence and shape
of neurals 1, 3, 5–7 can be reconstructed based on the
shape of adjacent costals. The preneural is absent in all
specimens. In most specimens neurals 1–4 are hexagonal
short-sided posteriorly, neural 5 is tetragonal (isometric),
neural 6 is hexagonal short-sided anteriorly, and neural 7
is pentagonal. A small diamond-shaped neural 8 is present
in MPC 25/162, where it is separated from neural 7 by the
short midline contact between costals 7. In MPC 25/160,
neural 9 is located at the posterior margin of the carapace.
It is semilunate in shape (rounded anteriorly) and is sepa-
rated from neural 7 by the midline contact of costals 7. In
ZIN PH T/M67-5, only neurals 1–3 are hexagonal short-
sided posteriorly, and neural 4 is isometric. In MPC 25/
160 and MPC 25/162, the neural series seems to be
slightly wider than in the holotype. In all specimens,
where observable, neural 1 is the longest. Its lateral bor-
ders are almost straight in the holotype and MPC 25/162,
but they are distinctly concave in MPC 25/160 and ZIN
PH T/M67-5. Neural 7 varies in size and shape, being the
largest and the longest in MPC 25/160, relatively small
Table 2. Variation of some carapace characters in Gobiapalone orlovi.
Characters PIN 557-133 PIN 557-132/1 ZIN PH T/M67-5 MPC 25/160 MPC 25/163 ZIN PH T/M71-1
Carapace length (mm) 155� 240� ? 275 290 335Suprascapular fontanelles Present Present Probably absent Separated Absent AbsentNumber of neurals 7 At least 7 ? 8 8 8Costals 8 Present Present ? Present Present PresentIsometric neural 5 5 4 5 5 5
�Estimation.
Cretaceous soft-shelled turtles (Trionychidae) of Mongolia 13
Dow
nloa
ded
by [
Igor
G. D
anilo
v] a
t 11:
45 2
1 Fe
brua
ry 2
014

and short in MPC 25/162, and even smaller in the
holotype.
Eight pairs of costals are present in specimens where
the posterior part of the carapace is preserved. The poste-
rior borders of costals 1 are almost perpendicular to the
midline in the holotype and MPC 25/162, but they are
curved anterolaterally in MPC 25/160 and ZIN PH T/
M67-5. Costals 6 are the longest laterally (at the free mar-
gin) of the costals. In the holotype, only the left costal 8 is
present. It is small, wider than long and probably con-
tacted its counterpart at the midline. In MPC 25/160 cos-
tals 8 are much smaller than in the holotype and are
separated from each other by the midline contact of cos-
tals 7. In MPC 25/162, costals 8 are larger than in the
holotype, and almost as long as wide. Depressions on cos-
tals 8 for contact of ilia are absent.
Plastron. Plastral remains consist of an almost complete
plastron (with a broken left hyo- and hypoplastra) of MPC
25/160, and a fragment of the left hyo- and hypoplastra
(ZIN PH T/M46-1). The bridge is short (the ratio of mini-
mal bridge length to maximal hypoplastron length is about
50%). The sculpturing of the plastron is less distinct than
those of the carapace. In MPC 25/160, it is most strongly
expressed in the lateral part of the hyo- and hypoplastra
and fades away in other parts of the plastron. In ZIN PH
T/M46-1, the sculpturing is similar to that on the carapace
in the lateral part of the specimen, but changes and con-
sists of tubercles and ridges in its central and medial parts.
There are five plastral callosities: one on the entoplastron,
one on each hyo- and hypoplastra, and one on each xiphi-
plastra (MPC 25/160).
The epiplastron is J-shaped and slender. The length of
the anterior process is about 50% of the length of the pos-
terior process and about 0.2 times the total width of the
hypoplastron. The epiplastron bears no callosity.
The entoplastron is V-shaped and slender. Each arm of
the entoplastron contacts the medial processes of the hyo-
plastron, but they are not sutured to the hyoplastron and
there is no hyoplastral shoulder to accommodate any
extensive entoplastron contact. The arms meet at about an
80� angle at a rounded anterior tip. The entoplastral cal-
losity is very small, rounded, and restricted to the middle
part of the plate.
The hyoplastron and hypoplastron are connected by a
suture and are not fused. The medial hypoplastral pro-
cesses could have met at the midline, but there was no
extensive midline contact between the hyoplastra or hypo-
plastra. The callosity covers part of the external surface of
the hyoplastron and hypoplastron. The medial, lateral and
posterior processes are left exposed. As is visible in MPC
25/160, the lateral hyoplastral lobe is almost equal to the
medial hyoplastral lobe in length. There are two lateral
hyoplastral processes in both specimens and five medial
hyoplastral processes (visible in MPC 25/160). The
hypoplastron has two lateral processes in both specimens.
As is visible in MPC 25/160, the medial hypoplastral pro-
cesses are divided into two groups (anterior and posterior)
separated by a short gap. The anterior group includes two
processes on the right hypoplastron and probably had
three processes on the left one. The anterior processes are
orientated roughly perpendicular to the midline. The pos-
terior group includes four processes directed posterome-
dially. The hyoplastron and hypoplastron of MPC 25/160
are different from those of ZIN PH T/M 46-1 in their
more emarginated lateral margin (between lateral pro-
cesses) and, as a result, in their slender lateral lobes, and
sculpturing (see above).
At the hypo-xiphiplastral contact, the xiphiplastral pro-
cess is lateralmost. There is no extensive midline contact
between the xiphiplastra. The right xiphiplastron of MPC
25/160 has three anteromedial processes, whereas the left
one has only two such processes. The posteromedial pro-
cesses of the xiphiplastra are singular. The lateral margin
of the xiphiplastra has a slight emargination. Most of the
external surfaces of both xiphiplastra, except for the pro-
cesses, are covered by callosities.
Nonshell postcranials. MPC 25/160 has preserved most
cervical, thoracic, sacral and caudal vertebrae, pectoral
and pelvic girdles, and limb bones.
The cervical vertebrae of MPC 25/160 are represented
by isolated fragments of cervical 1, complete cervicals
2–4, a partially preserved cervical 6 and complete cervi-
cals 7 and 8. Similar to other trionychids (see Meylan
1987), cervical centra 2–7 are opisthocoelous, cervical
centrum 8 has no posterior articular surface for contact
with thoracic vertebra 1 and a ventral process on cervical
8 is absent. There are no strong dorsal processes on the
cervicals, although there are small ridges along the central
axis. A ventral keel on cervical 8 is absent.
The first nine thoracic vertebrae are firmly attached to
the carapace in MPC 25/160. One more vertebral centrum
(10), connected with neural 9 (see above) is preserved
posterior to thoracic vertebra 9. There are two sacral cen-
tra and two pairs of isolated sacral ribs that became
detached during preparation of the specimen.
MPC 25/160 has a continuous series of caudal vertebrae
1–13 from the anterior part of the tail. They are procoe-
lous and steadily decrease in size towards the posterior
end of the tail.
The pectoral girdle of MPC 25/160 is represented by an
articulated right scapula (with the acromion process bro-
ken off) and partial coracoid, and fragments of the left
scapula and coracoid. It is not clear if the coracoid is the
longest of three pectoral processes, because the coracoid
is missing its distal end and the acromion process is bro-
ken off. The coracoid is flat and wide, as in other triony-
chids, with a slightly concave anterior margin and convex
posterior margin. The angle between the acromion process
14 I. G. Danilov et al.
Dow
nloa
ded
by [
Igor
G. D
anilo
v] a
t 11:
45 2
1 Fe
brua
ry 2
014

and the main body of the scapula and the angle between
the coracoid and the acromion process cannot be mea-
sured because the acromion process is broken off.
Only the right humerus is preserved in MPC 25/160. It
has a morphology typical for trionychids, including an
open ectepicondylar groove and the lesser trochanter of
the humerus lying in an anteroposterior plane that runs
through the main axis of the humerus (see Meylan 1987).
A single radius and ulna are preserved in MPC 25/160. It
is unclear whether they are from the right or the left forelimb.
The pelvic girdle is represented in MPC 25/160 by a
complete right half of the girdle and an isolated left ilium.
The epipubic region is not present and presumably did not
ossify. The pubis and ischium do not divide the thyroid
fenestra. The pectineal process is about the same width as
the medial border of the pubis (interpubic suture); they lie
in a single plane. The ilia curve posteriorly but not medi-
ally. The distal ends are not expanded. Both the thelial
process and the ilioischial notch are absent. The ischium
does not extend into the thyroid fenestra. A metischial
process is present.
Both femora are preserved in MPC 25/160, although
the distal part is broken off the left femora. Their mor-
phology is typical for trionychids in that both trochanters
lie at a wide angle to the anteroposterior plane that runs
through the main axis of the femur (Meylan 1987).
A single tibia and fibula are preserved in MPC 25/160.
It is unclear whether they are from the right or the left
hind limb.
MPC 25/160 has a number of disarticulated elements of
the autopodium of the fore- and hind limbs. It is unclear
how many clawed digits were in the manus or pes, or if
manus or pes digits display hyperphalangy.
Remarks. All specimens referred here to Gobiapalone
orlovi are similar to the holotype and/or to the most com-
plete specimen (MPC 25/160) in the following shell char-
acters: type A shell sculpturing, carapace length up to
335 mm, suprascapular fontanelles closed in specimens
larger than 275 mm in carapace length, and general mor-
phology of the hyo- and hypoplastra. In addition, all these
specimens come from the Baynshire Formation. Differen-
ces between these specimens can be explained through
intraspecific (number of neurals, position of an isometric
neural, shape of costals 8), and ontogenetic (presence/
absence of the suprascapular fontanelles) variation that
characterizes other species of Trionychidae (Gardner &
Russell 1994). In addition to ZIN PH T/M46-1 (consisting
of the lateral portion of the hyo- and hypoplastra and two
cervical vertebrae), which was primarily reported as
Amyda sp. cf. A. orlovi (Khosatzky 1999), and later
assigned to ‘Amyda’ orlovi (Danilov & Vitek 2009,
2012a), here we attribute several other specimens to this
species. Some of those specimens (MPC 25/162, PIN
557-133 and carapace fragment of ZIN PH T/M46-1)
have not been reported previously, whereas others (MPC
25/160, ZIN PH T/M67-5 and ZIN PH T/M71-1), have
been reported under different names. MPC 25/160 was
reported as Apalonina (Suzuki 2005) and Apalonina indet
3. (Danilov & Vitek 2012a). ZIN PH T/M67-5 was origi-
nally reported as “Trionychidae possibly representing a
new species and characterized by having a rather long
neural 1 that is widened anteriorly and posteriorly, costals
1 that considerably turn distally to the anterior, and an iso-
metric neural 4” (Khosatzky 1999, pp. 144–5). Later it
was mentioned as Trionychinae indet. 4 (Danilov & Vitek
2009; 2012a; Danilov et al. 2011a) or Trionychinae indet.
(Danilov et al. 2011b). ZIN PH T/M71-1 was originally
reported as Trionychina indet. (Khosatzky 1999), and later
mentioned as Trionychinae indet. 3 (Danilov & Vitek
2009, 2012a).
Gobiapalone breviplastra sp. nov.
(Figs 10–12)
1972 Trionyx sp. b; M»ynarski & Narmandach: 100.
1978 Platypeltis sp.; Merkulova: 156.
1999 Trionychidae indet.; Khosatzky: 145, fig. 4.
1999 Apalonini; Khosatzky: 147, fig. 5.
2009 Trionychinae indet. 5; Danilov & Vitek: 54.
2009 Trionychinae indet. 6 (part.); Danilov &
Vitek: 54.
2009 Apalonina indet.; Danilov & Vitek: 55.
2011a ‘Amyda’ menneri: Danilov et al.: 15 (part.).
2011b ‘Amyda’ menneri: Danilov et al.: 96 (part.).
2012a Apalonina indet. 1; Danilov & Vitek: 421,
fig. 23.2a.
2012a Apalonina indet. 2; Danilov & Vitek: 421.
2012a Trionychini indet. 2; Danilov & Vitek: 425,
fig. 23.2f.
2012a Trionychinae indet. 5; Danilov & Vitek: 429
(part.).
Derivation of name. Brevis, Greek for short, and
plastra, in reference to short anterior processes of
epiplastra.
Holotype. PIN 4694-3, partial shell, from Ulan Khushu
(¼ Ulan Bulak), Umunugovi Aimag, Mongolia, Nemegt
Formation, Maastrichtian (collected by JSMPE).
Referred material. MPC 25/163 (HMNS 94-10-24), par-
tial disarticulated skeleton, from Bugin Tsav, Umunugovi
Aimag, Mongolia, Nemegt Formation, Maastrichtian (col-
lected by JMJPE in 1994); MPC 25/164 (HMNS 98-16-
499), partial skeleton, from Bugin Tsav II, Umunugovi
Aimag, Mongolia, Nemegt Formation, Maastrichtian (col-
lected by JMJPE in 1998); PIN 551-461, partial juvenile
shell with fragments of non-shell postcrania, from Nem-
egt, Umunugovi Aimag, Mongolia, Nemegt Formation,
Maastrichtian (collected by MPE in 1949); PIN 4692-301,
Cretaceous soft-shelled turtles (Trionychidae) of Mongolia 15
Dow
nloa
ded
by [
Igor
G. D
anilo
v] a
t 11:
45 2
1 Fe
brua
ry 2
014

partial juvenile skeleton (collected by JSMPE in 1991),
PIN 4692-302, partial disarticulated skeleton (collected
by JSMPE in 1974), PIN 4692-303, partial shell (collected
by A. V. Sochava in 1968), all from Bugin Tsav, Umunu-
govi Aimag, Mongolia, Nemegt Formation, Maastrich-
tian; PIN 4694-1, partial entoplastron (collected by
JSMPE in 1970), PIN 4694-2, partial carapace (collected
by MPE), all from Ulan Khushu ( ¼ Ulan Bulak), Umunu-
govi Aimag, Mongolia, Nemegt Formation, Maastrich-
tian; PIN 5505-1, partial carapace, from Alak Shand
Khuduk, Mongolia, Nemegt Formation, Maastrichtian
(collected by JSMPE in 1974); ZIN PH 1/158, partial car-
apace on a cast of the internal cavity of the shell, from
Altan Ula I, Umunugovi Aimag, Mongolia, lower part of
the Nemegt Formation, Maastrichtian (collected by N. N.
Verzilin in 1976); ZIN PH T/M67-6, partial carapace,
from Bugin Tsav, Umunugovi Aimag, Mongolia, Barun-
goyot Formation, Campanian (collected by A. V. Sochava
in 1967); ZIN PH T/M68-1, a partial carapace, from
Nogon Tsav, Bayankhongor Aimag, Mongolia, upper part
Figure 10. Shell material of Gobiapalone breviplastra gen. et sp. nov. from the Nemegt Formation of: A–E, Nemegt; F–I, Ulan Khushu(¼ Ulan Bulak); J, Altan Ula; I, K, Alak Shand Khuduk; and L, from the upper part of the Barungoyot Formation of Nogon Tsav. A, B,PIN 551-461, a partial juvenile shell with fragments of non-shell postcrania in: A, dorsal and B, ventral views. C, ZPAL MgCh/80, pos-terior carapace fragment in dorsal view. D, ZPAL MgCh/81, left hypoplastron in ventral view. E, ZPAL MgCh/83, partial left hyo- andhypoplastra in ventral view. F, G, PIN 4694-3, a partial shell (holotype): F, carapace in dorsal view; G, plastron in ventral view. H, PIN4694-1, partial entoplastron in ventral view. I, PIN 4694-2, carapace in dorsal view. J, ZIN PH 1/158, partial carapace on a cast of theinternal cavity of the shell. K, PIN 5505-1, partial carapace in dorsal view. L, ZIN PH T/M68-1, partial carapace in dorsal view.Reconstructed sutures and outlines are indicated by dashed lines.
16 I. G. Danilov et al.
Dow
nloa
ded
by [
Igor
G. D
anilo
v] a
t 11:
45 2
1 Fe
brua
ry 2
014

Figure 11. Shell material of Gobiapalone breviplastra gen. et sp. nov. from the: A–D, F–M, Nemegt; and E, Barungoyot formations ofBugin Tsav. A–D, MPC 25/163; A, B, carapace in A, dorsal and B, ventral views; C, D, plastron in C, ventral and D, dorsal views. E, ZINPH T/M67-6, carapace in dorsal view. F, G, MPC 25/164; F, carapace in dorsal view; G, plastron in ventral view. H, I, PIN 4692-301,juvenile shell with fragments of non-shell postcrania in H, dorsal and I, ventral views; J, K, PIN 4692-302; J, carapace in dorsal view; K,plastron in ventral view. L, M, PIN 4692-303: L, carapace in dorsal view; M, right hyo- and hypoplastra in ventral view. Reconstructedsutures and outlines are indicated by dashed lines.
Cretaceous soft-shelled turtles (Trionychidae) of Mongolia 17
Dow
nloa
ded
by [
Igor
G. D
anilo
v] a
t 11:
45 2
1 Fe
brua
ry 2
014

of Barungoyot Formation, Campanian (collected by A. V.
Sochava in 1968); ZPAL MgCh/80, posterior carapace
fragment, including neurals 6 and 8 and medial parts of
costals 6 and 7 and additional costal fragment, ZPAL
MgCh/81, left hypoplastron, ZPAL MgCh/83, partial left
hyo- and hypoplastra missing most of their medial por-
tions, additional shell fragments and bones of non-shell
postcrania, all from Nemegt, Umunugovi Aimag, Mongo-
lia, Nemegt Formation, Maastrichtian (collected by
PMPE).
Diagnosis. A species of Gobiapalone with estimated car-
apace length up to 260 mm, which can be differentiated
from G. orlovi by the closed suprascapular fontanelles in
specimens larger than 224 mm in carapace length, more
robust epiplastra and entoplastron, a larger (about 100�)angle between the arms of the entoplastron, fewer (two or
three) medial processes of the hyoplastron, a smaller gap
between the anterior and posterior groups of the medial
processes of hypoplastron, and more robust xiphiplastra.
Description
Carapace. The carapace disc is longer than wide in MPC
25/164 and PIN 4692-301, as long as wide in PIN 4694-3
and, probably, PIN 551-461 and ZIN PH T/M68-1, and
wider than long in MPC 25/163, PIN 4692-302, PIN
4694-2 and ZIN PH T/M67-6. The anterior border of the
carapace is rounded and some specimens have a weak
nuchal emargination. The lateral borders of the carapace
are straight, or sometimes slightly scalloped. The poste-
rior margin of the carapace is rounded (PIN 551-461),
straight (MPC 25/163, PIN 4692-301, PIN 4692-302, PIN
4694-3, ZIN PH T/M67-6 and ZIN PH T/M68-1) or more
or less emarginated (PIN 4694-2 and PIN 5505-1). The
free ribs of the costals, when preserved, project far beyond
the carapace disc even in large specimens. The suprascap-
ular fontanelles are present and confluent in smaller speci-
mens (PIN 551-461, PIN 4692-301, PIN 4694-2, PIN
4694-3 and PIN 5505-1), present but separated from each
other in a larger one (MPC 25/164) and absent in the larg-
est specimens (MPC 25/163 and PIN 4692-303). The
exception is PIN 4692-302, which has a medium carapace
length, but has no suprascapular fontanelles. This may
indicate some variation in rates of shell growth potentially
connected to sexual dimorphism. All specimens have type
A shell sculpturing. See Table 3 for variation of some car-
apace characters of Gobiapalone breviplastra.
The nuchal is more than four times wider than long (the
nuchal length/width ratio varies from about 7.0 in MPC
25/163 to about 5.0 in PIN 5505-1). Its anterior margin is
either not emarginated (MPC 25/164, PIN 4692-303, PIN
4694-3, and ZIN PH T/M67-6) or weakly emarginated
(PIN 4692-302 and PIN 5505-1). As is visible in ventral
view of MPC 25/163, the costiform processes are united.
The posterolateral aspects of the nuchal are covered by
the expansions of costals 1, and free ribs of costals 1 lie
posterior to the nuchal/costals 1 suture. In other speci-
mens, the posterolateral aspects of the nuchal are covered
by the expansions of the free ribs of costals 1.
A complete neural series is present in MPC 25/163, PIN
551-461, PIN 4692-301, PIN 4692-302, PIN 4694-2, PIN
4694-3, ZIN PH T/M67-6 and ZIN PH T/M68-1. In MPC
25/164, only neurals 1–7 are preserved, and the shape of
neural 8 can be reconstructed based on adjacent elements.
In PIN 5505-1, all neurals are preserved, except 4 and 5,
Figure 12. Non-shell postcrania of Gobiapalone breviplastra gen. et sp. nov., MPC 25/163, from the Nemegt Formation of Bugin Tsav.A, cervical vertebrae in dorsal view; B, left scapula in lateral view; C, left coracoid in dorsal view; D, pelvic girdle in dorsal view.Reconstructed outlines are indicated by dashed lines.
18 I. G. Danilov et al.
Dow
nloa
ded
by [
Igor
G. D
anilo
v] a
t 11:
45 2
1 Fe
brua
ry 2
014

whose shape can be reconstructed based on adjacent ele-
ments. In PIN 4692-303, only neurals 1–4 are preserved.
In ZPAL MgCh/80, part of neural 6 and complete neural
8 are preserved, and the shape of neural 7 is reconstructed
based on adjacent elements. In total, specimens may have
seven (PIN 4692-302, PIN 4694-2, ZIN PH T/M67-6 and
ZIN PH T/M68-1), eight (MPC 25/163, MPC 25/164, PIN
4692-301, PIN 4694-3, PIN 5505-1 and ZPAL MgCh/80)
or nine (PIN 551-461) neurals. The preneural is absent in
all specimens. In all specimens, where observable, neurals
1–4 are hexagonal short-sided posteriorly, neural 5 is
tetragonal (isometric), and neural 6 is hexagonal short-
sided anteriorly. Neural 1 is longer than the other neurals
in most specimens, except PIN 4692-301 and ZIN PH T/
M68-1. The lateral borders of neural 1 are straight (PIN
4692-301, PIN 4694-3 and ZIN PH T/M68-1) or more or
less concave (other specimens). Neural 1 is either nar-
rowed anteriorly (most specimens) or as wide anteriorly
as posteriorly (largest specimens: MPC 25/163 and PIN
4692-303). Neural 7 is either pentagonal (MPC 25/163,
PIN 551-461, PIN 4692-301, PIN 4692-302, PIN 4694-2,
PIN 4694-3, ZIN PH T/M68-1) or hexagonal short-sided
anteriorly (MPC 25/164, PIN 5505-1, ZPAL MgCh/80).
Neural 8 is the most variable in shape and position. In
PIN 551-461, neural 8 is hexagonal and located between
costals 7 and 8, separated from neural 7 by short midline
contact of costals 7, and contacts neural 9 posteriorly. In
MPC 25/163, neural 8 is small and rounded, located
between costals 7 and 8, separated from neural 7 by long
midline contact between costals 7 and contributes to the
free margin of the carapace posteriorly. In PIN 4694-3,
neural 8 is large and almost triangular, located between
costals 7, separated from neural 7 by short midline contact
between costals 7 and contributes to the free margin of the
carapace posteriorly. In PIN 5505-1, neural 8 is irregularly
tetragonal in shape, located between costals 7, contacts
neural 7 anteriorly and was probably separated from the
free margin of the carapace by the midline contact of cos-
tals 7 or 8. In ZPAL MgCh/80, neural 8 is elongated and
tetragonal in shape, located between costals 7, contacts
neural 7 anteriorly and contributes to the free margin of the
carapace posteriorly. In PIN 4692-301, neural 8 is very
small and rounded, located between costals 7,
contacts neural 7 anteriorly and is separated from the
free margin of the carapace by the short midline contact of
costals 7. In MPC 25/164, neural 8 is not preserved, but
was probably tetragonal, located between costals 7, and
contacted neural 7 anteriorly and costals 8 posteriorly.
Seven (PIN 4692-301, PIN 4692-302, PIN 4694-3 and
ZIN PH T/M68-1) or eight (MPC 25/163, PIN 551-461
and ZIN PH T/M67-6) pairs of costals are present. Costals
6 and 7 make up the posterior margin of the carapace of
ZPAL MgCh/80. Costals 8 are reconstructed in MPC 25/
164, PIN 4694-2 and PIN 5505-1. The posterior border of
costals 1 curve anterolaterally in all specimens. Costal 1 is
either almost as long laterally as it is medially (most speci-
mens) or considerably shorter laterally than it is medially
(MPC 25/164 and PIN 4694-2). All other costals are usu-
ally longer laterally than medially. However, in PIN 4692-
303, costal 2 is very long laterally, whereas costal 3 is as
long laterally as medially. Costals 6 are much longer lat-
erally than medially, form the posterolateral edge of the
carapace and usually contribute to its posterior edge. Cos-
tals 7 are either large triangular-shaped elements which
form most of the posterior carapace margin (PIN 4692-
301, PIN 4692-302, PIN 4694-3, ZIN PH T/M68-1 and
ZPALMgCh/80), or smaller elements that are notched pos-
teriorly by costals 8, and make a smaller contribution to the
posterior carapace margin (MPC 25/163, MPC 25/164,
PIN 551-461, PIN 4694-2, PIN 5505-1 and ZIN PH T/
M67-6). Costals 7 usually contact each other along the
midline posterior to neurals 7 or 8, except MPC 25/164
and ZIN PH T/M67-6, in which costals 7 are separated by
neural 8 and costals 8 respectively. Costals 8, when pres-
ent, are always small, triangular-shaped elements, which
contribute to the posterior carapace margin. They are either
separated from each other by neurals 8 and 9 (PIN 551-
461), or only by neural 8 (MPC 25/163), or by costals 7
(PIN 5505-1 and PIN 4694-2), or contact each other along
the midline (MPC 25/164 and ZIN PH T/M67-6).
Table 3. Variation of some carapace characters in Gobiapalone breviplastra gen. et sp. nov.
CharactersPIN
4692-301PIN
551-461PIN
4692-302PIN
5505-1ZIN PHT/M67-6
PIN4694-3
PIN4694-2
MPC25/164
ZIN PHT/M68-1
MPC25/163
ZIN PH4692-303
Carapacelength (mm)
115 147� 185 193 196 197 207� 224� 230� 241 260�
Suprascapularfontanelles
Confluent ? Absent Confluent ? Confluent Confluent Separated ? Absent Absent
Number ofneurals
8 9 7 8 7 8 7 Probably 8 7 8 ?
Costals 8 Absent Present Absent Present Present Absent Probablypresent
Present Absent Present ?
�Estimation.
Cretaceous soft-shelled turtles (Trionychidae) of Mongolia 19
Dow
nloa
ded
by [
Igor
G. D
anilo
v] a
t 11:
45 2
1 Fe
brua
ry 2
014

Plastron. Plastral remains are represented by an almost
complete plastron missing the left epiplastron and lateral
portions of the entoplastron (MPC 25/163), a partial plas-
tron represented by the left hyo- and hypoplastra, a medial
portion the of right hyo- and hypoplastra, and central parts
of both xiphiplastra (MPC 25/164), a partial plastron rep-
resented by the left epiplastron, the entoplastron, medial
portions of both hyo- and hypoplastra, and fragments of
both xiphiplastra (PIN 4694-3), a partial plastron repre-
sented by the entoplastron, medial portions of both hyo-
and hypoplastra, and a fragment of the left xiphiplastron
(PIN 4692-301), a partial plastron represented by the left
epiplastron, a partial entoplastron, the right hyoplastron
and medial portion of the left hyo- and hypoplastra (PIN
4692-302), a partial plastron represented by the posterior
portion of the right epiplastron, the entoplastron, both
hyo- and hypoplastra and both xiphiplastra (PIN 551-
461), a partial entoplastron missing the left lateral portion
(PIN 4694-1), a partial right hyo- and hypoplastra (PIN
4692-303), a partial left hypoplastron (ZPAL MgCh/81),
and a partial right hyo-hypoplastron (ZPAL MgCh/83).
The bridge is short (the ratio of minimal bridge length to
maximal hypoplastron length is about 50%). The sculptur-
ing of the plastron is less distinct than that of the carapace
and is most strongly expressed on the hyo- and hypoplas-
tra. There are five plastral callosities: one on the entoplas-
tron, two on the hyo- and hypoplastra and two on the
xiphiplastra.
The epiplastron is J-shaped and more robust than in
Gobiapalone orlovi. The length of the anterior process is
about 45% of the length of the posterior process and about
0.2 times the total width of the hypoplastron. The epiplas-
tron bears no callosity.
The entoplastron is V-shaped, and more robust than in
Gobiapalone orlovi. Each arm of the entoplastron contacts
the medial processes of the hyoplastron, but they are not
sutured to the hyoplastron and there is no hyoplastral
shoulder to accommodate any extensive entoplastron con-
tact. The arms meet at about a 100� angle with a rounded
(PIN 551-461 and PIN 4694-3), pointed (PIN 4692-302
and PIN 4694-1) or truncated (MPC 25/163) anterior tip.
The entoplastral callosity is very small, rounded, and
restricted to the middle part of the bone.
The hyoplastron and hypoplastron are connected by a
suture and are not fused. The medial hypoplastral pro-
cesses could have met at the midline, but there was no
extensive midline contact between the hyoplastra or hypo-
plastra. The callosity covers part of the external surface of
the hyoplastron and hypoplastron, except for the medial,
lateral and posterior processes, as is visible in MPC 25/
163. In small specimens (PIN 551-461 and PIN 4692-
301), the callosity is relatively small and covers only the
central part of the hyoplastron and hypoplastron. As is vis-
ible in MPC 25/163, the lateral hyoplastral lobe is slightly
shorter than the medial hyoplastral lobe. There are two
lateral hyoplastral processes and two or three medial
hyoplastral processes. The hypoplastron has two lateral
processes. As is visible in MPC 25/163 and PIN 4694-3,
the medial hypoplastral processes are divided into two
groups (anterior and posterior) separated by a very short
gap (shorter than in Gobiapalone orlovi). The anterior
group includes two (MPC 25/163 and PIN 4694-3) or
three (PIN 551-461) processes, which are almost perpen-
dicular to the midline. The posterior group includes from
three to four processes directed posteromedially. One
hypoplastron (ZPAL MgCh/81; Fig. 10D), about the same
size as MPC 25/163, demonstrates more complete cover-
age of its medial and lateral lobes by the callosity.
At the hypo-xiphiplastral contact, the xiphiplastron is
lateralmost. There is no extensive midline contact
between the xiphiplastra. The right xiphiplastron of MPC
25/163 has one anteromedial processes, whereas the left
one has two such processes. The posteromedial processes
of the xiphiplastra are singular. The lateral margin of the
xiphiplastra has a very slight emargination (smaller than
in Gobiapalone orlovi). The majority of the xiphiplastra,
except the processes, is covered by a callosity.
Nonshell postcranials. MPC 25/163 has complete cervi-
cal vertebrae IV–VI and an anterior fragment of cervical
vertebra VIII. It also has thoracic, sacral and several cau-
dal vertebrae, the pectoral and pelvic girdles, both humeri
and part of the right(?) hind limb in articulation in matrix.
MPC 25/164 has cervical vertebrae IV–VI and VIII, frag-
ments of pectoral and pelvic girdles, a complete left
humerus and fragments of other limb bones. PIN 4692-
301 has the pectoral and pelvic girdles and a fragment of
the right humerus. PIN 4692-302 has cervical vertebra IV,
the pectoral girdle, and two humeri. PIN 551-461 has the
anterior caudal vertebrae, the pectoral and pelvic girdles,
and bones of the hind limb autopodium preserved.
The morphology of the available cervical vertebrae is
similar to that of Gobiapalone orlovi.
The first 11 thoracic vertebrae are attached to the cara-
pace in MPC 25/163. There is one sacral centrum with a
pair of sacral ribs that became detached during the prepa-
ration of the specimen.
MPC 25/163 has five anterior caudal vertebrae pre-
served, one of which is isolated and four of which are in
articulation; four more caudal vertebrae of the same speci-
men have been preserved in the piece of matrix.
As is visible in MPC 25/163, the coracoid and the main
body of the scapula are broken off distally and it is not
clear what pectoral process is the longest. The coracoid is
flat and wide, as in other trionychids, with a slightly con-
cave anterior margin and convex posterior margin. The
angle between the acromion process and the main body of
the scapula is about 60�, but the angle between the cora-
coid and the acromion process cannot be measured.
The morphology of the available humeri is similar to
that of Gobiapalone orlovi and other trionychids.
20 I. G. Danilov et al.
Dow
nloa
ded
by [
Igor
G. D
anilo
v] a
t 11:
45 2
1 Fe
brua
ry 2
014

The morphology of the pelvic girdle and hind limb
(as seen in MPC 25/163) is similar to that of Gobiapalone
orlovi, as well as most other trionychids.
Remarks. All specimens referred here to Gobiapalone
breviplastra are similar to the holotype and the most com-
plete specimen (MPC 25/163) in the following shell char-
acters: type A shell sculpturing, carapace length up to
260 mm, suprascapular fontanelles usually closed in
specimens larger than 224 mm in carapace length, and
morphology of the plastral elements (see Diagnosis). In
addition, all these specimens come from the Barungoyot
and Nemegt formations. Differences between the speci-
mens are explained by intraspecific (shape of the cara-
pace, number of neurals, shape of neurals 1, 7 and 8,
number of costals, shape and contacts of costals 7 and 8,
shape of the anterior tip of entoplastron, number of medial
processes of the hyo- and hypoplastra) and ontogenetic
(presence/absence of the suprascapular fontanelles and
degree of ossification of hyo- and hypoplastra) variation
noted previously (Gardner & Russell 1994). Some of
these specimens (MPC 25/163, MPC 25/164, PIN 4692-
301, PIN 4692-302, PIN 4692-303, PIN 4694-1, PIN
4694-2, PIN 5505-1, ZIN PH 1/158) have not been
reported previously, whereas others (PIN 551-461, ZIN
PH T/M67-6, ZIN PH T/M68-1, ZPAL MgCh/80, ZPAL
MgCh/81 and ZPAL MgCh/83) have been reported under
different names. PIN 551-461 was originally reported as
Platypeltis sp. (Merkulova 1978), and later mentioned as
Apalonina indet. (Danilov & Vitek 2009) and Apalonina
indet. 2 (Danilov & Vitek 2012a). ZIN PH T/M67-6 was
originally reported as Trionychidae indet. (Khosatzky
1999), and later mentioned as Trionychinae indet. 5 (Dan-
ilov & Vitek 2009) and Trionychini indet. 2 (Danilov &
Vitek 2012a). ZIN PH T/M68-1 was originally reported
as Apalonini (Khosatzky 1999), and later mentioned as
Apalonina indet. (Danilov & Vitek 2009) and Apalonina
indet. 1 (Danilov & Vitek 2012a). ZPAL MgCh/80,
ZPAL MgCh/81 and ZPAL MgCh/83 were originally
reported as ‘Trionyx sp. b’ (M»ynarski & Narmandach
1972), and later erroneously considered to be part of Trio-
nychinae indet. 6 (Danilov & Vitek 2009) and Trionychi-
nae indet 5 (Danilov & Vitek 2012a). Other specimens
assigned to ‘Trionyx sp. b’ (ZPAL MgCh/88, about 20
fragments of neurals and costals; ZPAL MgCh/89, six
plastral fragments; M»ynarski & Narmandach 1972) can-
not be identified below the family level and should be
considered Trionychidae indet.
Gobiapalone sp.
(Fig. 16A)
Referred material. MPC 25/165 (HMNS 97-21-55),
incomplete carapace, from Khermin Tsav, Umunugovi
Aimag, Mongolia, Lower White Beds, ?Baynshire Forma-
tion, Cenomanian–Santonian.
Description. This carapace is poorly preserved and dam-
aged. Its length is estimated to be about 230 mm. The car-
apace is slightly wider than long. Its anterior border is
rounded and has no nuchal emargination. Suprascapular
fontanelles are present and separated by contact between
the nuchal and neural 1. The number and shape of the neu-
rals is unclear. Seven pairs of costals are observable, but
the presence or absence of costals 8 is unclear. The speci-
men has type A shell sculpturing.
Remarks. This specimen is assigned to Gobiapalone
based on its similar size, carapace shape, presence of
suprascapular fontanelles and sculpture pattern. The spec-
imen is missing characters that would allow for identifica-
tion to species level.
Trionychinae incertae sedis
‘Trionyx’ baynshirensis sp. nov.
(Fig. 13A, B)
Derivation of name. Species name is from Bayn Shire
locality.
Holotype. PIN 557-134 (formerly PIN 557-130), medial
fragment of right hyo- and hypoplastra, from Bayn Shire,
Dornogovi Aimag, Mongolia, Baynshire Formation, Cen-
omanian–Santonian (collected by MPE in 1948).
Diagnosis. A trionychine, with an estimated carapace
length of about 500 mm, that is most similar amongst Cre-
taceous trionychids to ‘Trionyx’ kansaiensis in the shape
of the hyo- and hypoplastra and the shell sculpturing of
type B. It can be differentiated from ‘Trionyx’ kansaiensis
in that its medial hyoplastral and hypoplastral processes
are completely covered by a callosity at smaller size, and
the hyo- and hypoplastra has straight medial borders. It
can be differentiated from Gobiapalone orlovi (another
trionychid known from the Baynshire Formation) by its
larger size, shell sculpturing, callosity that completely
covers the medial hyoplastral and hypoplastral processes,
and straight medial borders of the hyo- and hypoplastra.
Description. PIN 557-134 belongs to an individual with
an estimated carapace length of about 500 mm, based on
comparison with ‘Trionyx’ kansaiensis. The specimen has
type B sculpturing. The suture between hyo- and hypo-
plastra is not discernable due to poor preservation, and
may be absent. The hyo- and hypoplastra have straight
medial borders. They could probably contact their coun-
terparts across the midline, but there is no evidence of a
midline suture. There were two medial fontanelles
between the hyo-and hypoplastra: the anterior one
between anterior halves of the hyoplastra, and the poste-
rior one between posterior halves of the hypoplastra. The
medial hyoplastral and hypoplastral processes are almost
completely covered by a callosity and their precise num-
ber is unclear.
Cretaceous soft-shelled turtles (Trionychidae) of Mongolia 21
Dow
nloa
ded
by [
Igor
G. D
anilo
v] a
t 11:
45 2
1 Fe
brua
ry 2
014

‘Trionyx’ gilbentuensis sp. nov.
(Fig. 13C, D)
1999 ‘Plastomenus’; Khosatzky: 147, fig. 6.
2009 Trionychinae indet. 6; Danilov & Vitek: 54 (part.).
2011b Trionychinae indet.; Danilov et al.: 96.
2012a Trionychidae indet. 5; Danilov & Vitek: 429 (part.),
fig. 23.3g.
Derivation of name. Species name is from Gilbentu
locality.
Holotype. ZIN PH T/M46-2, incomplete left hyo- and
hypoplastra, from Gilbentu, Umunugovi Aimag, Mongo-
lia, Nemegt Formation, Maastrichtian (collected by MPE
in 1946).
Diagnosis. A trionychine, with an estimated carapace
length of about 500 mm, that is most similar, amongst
Cretaceous trionychids, to ‘Trionyx’ baynshirensis in the
shape of the hyo- and hypoplastra, shell sculpturing and in
that its medial hyoplastral and hypoplastral processes are
completely covered by a callosity. It can be differentiated
from ‘T.’ baynshirensis by the convex medial borders of
Figure 13. A, B, ‘Trionyx’ baynshirensis sp. nov., PIN 557-134 (holotype), a medial fragment of right hyo- and hypoplastra from theBaynshire Formation of Bayn Shire in: A, ventral and B, dorsal views. C, D, ‘Trionyx’ gilbentuensis sp. nov., ZIN PH T/M46-2(holotype), incomplete left hyo- and hypoplastra from the Nemegt Formation of Gilbentu in: C, ventral and D, dorsal views.Reconstructed outlines are indicated by dashed lines.
22 I. G. Danilov et al.
Dow
nloa
ded
by [
Igor
G. D
anilo
v] a
t 11:
45 2
1 Fe
brua
ry 2
014

the hyo-and hypoplastra. It can be differentiated from
other trionychids of the Nemegt Formation by its larger
size and shell sculpturing. In addition, it can be differenti-
ated from Nemegtemys conflata by a sutural connection
between the hyo- and hypoplastra and from Gobiapalone
breviplastra by a callosity that completely covers the
medial hyoplastral and hypoplastral processes, a more
acute angle between the medial and lateral lobes of the
hypoplastron and a more complete covering of the lateral
hypoplastral lobe by a callosity.
Description. ZIN PH T/M46-2 (previously ZIN PHT
M46-2; Khosatzky 1999) consists of the medial part of a
left hyo- and hypoplastra and an additional posterolateral
fragment of the hypoplastron (not reported previously);
the posteromedial part of the hypoplastron is broken off.
The specimen belongs to an individual with an estimated
carapace length of about 500 mm, based on comparison
with ‘Trionyx’ kansaiensis. The specimen has type B
sculpturing. The hyo- and hypoplastra are connected by a
suture and are not fused. They could contact their counter-
parts across the midline only at the medialmost border of
the hyoplastron. The medial fontanelle between the hyo-
and hypoplastra was not hourglass-shaped, as opposed to
most other trionychines. The ratio of the minimum bridge
length to the maximum hypoplastron length is impossible
to estimate due to incompleteness of the specimen. The
medial hyoplastral processes are completely covered by a
callosity and their number is unclear. In ‘Trionyx’ kan-
saiensis, there are three large medial hyoplastral processes,
which are not covered by callosities in both small and large
sizes (Vitek & Danilov 2010). The medial hypoplastral
processes are completely covered by a callosity. Only
three processes, which project slightly beyond the medial
margin of the hypoplastron, are observable. The angle
between the medial and lateral hypoplastral lobes is acute
(75–80�). The posterolateral fragment of the hypoplastron,
if correctly identified, indicates that the lateral hypoplastral
lobe was almost entirely covered in sculpturing.
Remarks. Khosatzky (1999) described this specimen as
a relatively large trionychid with tuberculate sculpture
and assigned it to ‘Plastomenus’. Danilov & Vitek
(2009, 2012a) assigned this specimen to Trionychinae
(as Trionychinae indet. 6 and 5 respectively) based on its
similarities with the trionychine ‘Trionyx’ kansaiensis.
Those authors also noted that ‘Trionyx sp. b’, based on
shell fragments of a large trionychid that came from the
Nemegt Formation of the Nemegt locality (M»ynarski &Narmandach 1972), probably belonged to the same taxon
as ZIN PH T/M46-2. Here, this taxon (‘Trionyx’ gilben-
tuensis) is restricted to the holotype, whereas material of
‘Trionyx sp. b’ is referred to Gobiapalone breviplastra
and Trionychidae indet. (see account of G. breviplastra).
Danilov & Vitek (2012a) mentioned ‘Plastomenus’?
of Khosatzky (1999) amongst the synonymies of
Trionychinae indet. 5. Here, the material on which this
determination was based (a plastron fragment) is consid-
ered as the holotype of Nemegtemys conflata gen. et sp.
nov. (see above).
‘Trionyx’ gobiensis sp. nov.
(Fig. 14)
1971 Trionyx sp. a; Khosatzky & M»ynarski: 141,
pl. 24, fig. 2, text-fig. 7.
1972 Trionyx sp. a; M»ynarski & Narmandach: 100.
1999 ‘Trionyx’ sp.; Khosatzky: 144.
2009 Trionychinae indet. 2; Danilov & Vitek: 54.
2012a Trionychinae indet. 2; Danilov & Vitek: 429,
fig. 23.3d.
Derivation of name. Species name is from Gobi Desert.
Holotype. PIN 4064-2, incomplete carapace, from
Bamba Khuduk (Eastern Sayr), Umunugovi Aimag, Mon-
golia, Nemegt Formation, Maastrichtian (collected by
JSMPE in 1976).
Referred material. ZPAL MgCh/59 (formerly ZPAL
MgCh/52) and ZPAL MgCh/76, two incomplete carapa-
ces, from Tsagan Khushu and Nemegt, respectively;
Umunugovi Aimag, Mongolia, Nemegt Formation, Maas-
trichtian; collection ZPAL MgCh/121, shell fragment,
consisting of the lateral parts of the nuchal and both cos-
tals 1, from Altan Ula III, Umunugovi Aimag, Mongolia,
Nemegt Formation, Maastrichtian (all specimens col-
lected by PMPE); ZIN PH 2/24, left costals 1 and 2, from
Tsagan Khushu, Umunugovi Aimag, Mongolia, Nemegt
Formation, Maastrichtian (collected by MPE in 1948; as
“two very small costals with tuberculate sculpturing”;
Khosatzky, 1999, p. 147).
Diagnosis. A trionychine that can be differentiated from
all other Cretaceous trionychids with known shells
(Table 1) by its smaller size (up to 130 mm). In addition,
it can be differentiated from ‘Aspideretes’ maortuensis
Yeh, 1965 by large costals 8; from Aspideretoides spp. by
the absence of a preneural and the presence of large cos-
tals 8; from Gilmoremys lancensis (Gilmore, 1928) by the
absence of a preneural and the presence of neural reversal
at neural 5; from Gobiapalone spp. by the absence of
suprascapular fontanelles and large costals 8; from
‘Trionyx’ kansaiensis by weak or absent nuchal emargina-
tion; from ‘T.’ shiluutulensis sp. nov. by neural reversal at
neural 5 and large costals 8.
Description. The carapace length is up to 130 mm long.
The carapaces are roughly circular, almost as wide as
long, with a weak or absent nuchal emargination and a
small posterior caudal emargination (visible in ZPAL
MgCh/59). The carapace is widest at costals 4. The free
ribs project far beyond the carapace disc in the smallest
Cretaceous soft-shelled turtles (Trionychidae) of Mongolia 23
Dow
nloa
ded
by [
Igor
G. D
anilo
v] a
t 11:
45 2
1 Fe
brua
ry 2
014

specimen (ZPAL MgCh/59) and project very little in
larger specimens (PIN 4064-2, ZPAL MgCh/76). The
suprascapular fontanelles are not present in any specimen.
The sculpturing on the surface of all specimens is similar
to those of Gobiapalone spp. and Aspideretoides spp.
(see above).
The nuchal is more than four times wider than long
(slightly more than four times in PIN 4064-2 and ZPAL
MgCh/59; more than five times in ZPAL MgCh/76). Its
anterior margin is either straight or has a weak emargina-
tion. As is visible in the ventral view of PIN 4064-2, the
costiform processes are united. The posterolateral aspects
of the nuchal are covered by the expansions of costal ribs 1.
The neural series is complete in all three relatively
complete carapaces and consists of seven neurals in PIN
4064-2 and ZPAL MgCh/59 and eight neurals in ZPAL
MgCh/76. The preneural is absent. Neurals 1–4 are hexag-
onal short-sided posteriorly, neural 5 is tetragonal
(isometric), and neural 6 is hexagonal short-sided anteri-
orly. Neural 7 is pentagonal in PIN 4064-2 and ZPAL
MgCh/59 and hexagonal short-sided anteriorly in ZPAL
MgCh/76. Neural 8 is pentagonal and present only in
ZPAL MgCh/76. Neural 1 is variable in shape, either nar-
rowed anteriorly (PIN 4064-2) or not (ZPAL MgCh/76).
Eight pairs of costals are present in three specimens,
although some of them are broken off or damaged. The
costals which are longest at the lateral margin are costals
2 and 5 in PIN 4064-2 and, probably, also in ZPAL
MgCh/76, and costals 3 and 5 in ZPAL MgCh/59. In PIN
4064-2 and ZPAL MgCh/59, costals 7 and 8 contacted
each other at the midline, whereas in ZPAL MgCh/76
only costals 8 have midline contact. Costals 8 are rela-
tively large and are wider than long. As is visible in
ZPAL MgCh/59, they formed most of the posterior cara-
pace margin.
Remarks. Khosatzky & M»ynarski (1971) described
ZPAL MgCh/59 (as ZPAL MgCh/52) as ‘Trionyx sp. a’.
In the schematic drawing of this specimen (Khosatzky &
M»ynarski 1971, text-fig. 7) the carapace had no preneu-
ral, eight neurals and eight pairs of costals. Other speci-
mens attributed to ‘Trionyx sp. a’ (ZPAL MgCh/60,
Figure 14. Shell material of ‘Trionyx’ gobiensis sp. nov. A, B, PIN 4064-2, an incomplete carapace from the Nemegt Formation ofBamba Khuduk in A, dorsal and B, ventral views. C, ZPAL MgCh/59, incomplete carapace from the Nemegt Formation of TsaganKhushu in dorsal view. D, ZPAL MgCh/76, incomplete carapace from the Nemegt Formation of Nemegt in dorsal view. E, ZPALMgCh/121, shell fragment, consisting of lateral parts of the nuchal and both costals 1, from the Nemegt Formation of Altan Ula III indorsal view. F, ZIN PH 2/24, left costals 1 and 2 in dorsal view from the Nemegt Formation of Tsagan Khushu. Reconstructed suturesand outlines are indicated by dashed lines.
24 I. G. Danilov et al.
Dow
nloa
ded
by [
Igor
G. D
anilo
v] a
t 11:
45 2
1 Fe
brua
ry 2
014

ZPAL MgCh/76 and unnumbered) have never been
described (Khosatzky & M»ynarski 1971; M»ynarski &
Narmandach 1972). Later, Danilov & Vitek (2012a)
reported that judging from a photograph of ZPAL MgCh/
59 (as ZPAL MgCh/52) it is impossible to confirm the
presence of a preneural and neural 8. These authors fur-
ther suggested that if this specimen has no preneural and
only seven neurals, it could be attributed to ‘Amyda’ men-
neri. However, new observations of this and other speci-
mens with similar morphology show that there was no
preneural, and the number of neurals varies between seven
and eight. All these specimens differ from other Creta-
ceous trionychids in multiple features (see Diagnosis
above) and, for that reason, we identify them as a new
species within Trionyx Geoffroy Saint Hillaire, 1809
sensu lato. Isolated costal fragments (ZPAL MgCh/60 and
unnumbered) from Tsagan Khushu, also attributed to
‘Trionyx sp. a’ by Khosatzky & M»ynarski (1971), cannotbe identified below the family level. The statement of
Khosatzky & M»ynarski (1971, p. 142) that “the greatest
number of the remains of these small soft-shelled turtles
are in the collections of Zoological Institute in Leningrad”
(ZIN PH) cannot be confirmed.
‘Trionyx’ shiluutulensis sp. nov.
(Fig. 15)
2011a Aspideretoides; Danilov et al.: 15.
2011b Aspideretoides; Danilov et al.: 96.
Derivation of name. Species name is from the Shiluut
Ula locality.
Holotype. MPC 25/166 (HMNS 97-21-56), carapace,
from Shiluut Ula, Umunugovi Aimag, Mongolia;
unknown formation, Campanian (collected by JMJPE in
1997; for age see Watabe et al. 2010).
Diagnosis. A trionychine with a carapace length of
225 mm that can be differentiated from all other Creta-
ceous trionychids with known carapaces (Table 1) by a
combination of two characters: the presence of both a pre-
neural and eight neurals. In addition, it can be differenti-
ated from ‘Aspideretes’ maortuensis by smaller size and
neural reversal at neural 6; from Aspideretoides spp. by
smaller size; from Gilmoremys lancensis by smaller size
and reduced costals 8; from Gobiapalone spp. by neural
reversal at neural 6; from ‘T.’ gobiensis by larger size,
neural reversal at neural 6 and small costals 8; and from
‘T.’ kyrgyzensis Nessov, 1995 by larger size, neural rever-
sal at neural 6 and small costals 8.
Description. The carapace length is 225 mm long. It is
almost as wide as long, and narrowed anteriorly due to the
strong emargination of its lateral borders between the free
ribs of costals 1 and 2. Posterior to these emarginations,
the lateral borders of the carapace are scalloped due to
smaller emarginations between the free ribs of costals 2
and 3, 3 and 4, 4 and 5, and 5 and 6. The free ribs of cos-
tals project beyond the carapace disc. Suprascapular fon-
tanelles are absent.
The nuchal is more than seven times wider than long.
Its anterior margin is not emarginated. As is visible in
ventral view, the costiform processes are united. The pos-
terolateral aspects of the nuchal are covered by the expan-
sions of the ribs of costals 1.
The neural series included a preneural and eight neurals,
of which the preneural and neurals 1, 2 and 7 are complete,
and neurals 3–6 are partially preserved. Neural 8 is not pre-
served, but its shape can be reconstructed based on adja-
cent bones. The preneural is roughly pentagonal and wider
than long. Its anterior point extends into the nuchal. Neu-
rals 1–5 are hexagonal short-sided posteriorly, neural 6 is
reconstructed as isometric (tetragonal), neural 7 is hexago-
nal short-sided anteriorly, and neural 8 is reconstructed as
pentagonal. Neurals 1 to 7 steadily decrease in width
towards the posterior end of the carapace.
Eight pairs of costals are present. The medial border of
costal 1 is divided into two parts, a short one for contact
with the preneural and a long one for contact with neural
1. The suture between costals 1 and 2 curves anterolater-
ally. The lateral border of costal 2 is much longer on the
right side of the carapace than on the left side. The lateral
margins of costals 6 are as long as the free, lateral margin
of left costal 2. Costals 8 are small and are wider than
long. Depressions on costals 8 for contact of the ilia are
absent.
Remarks. Previously, this specimen was attributed to the
genus Aspideretoides Gardner et al., 1995 (Danilov et al.
2011a, b), based on the presence of a preneural. However,
a preneural is characteristic of other Cretaceous triony-
chids such as the genus Gilmoremys Joyce & Lyson,
2011, which is considered to be a member of the Plasto-
menidae (Joyce & Lyson 2011). In addition, new observa-
tion shows that this specimen had eight neurals, a
character that is also observed in some Cretaceous triony-
chines, but is absent in Aspideretoides spp. and Gil-
moremys, which have seven neurals. For this reason, we
decide to establish a new species within Trionyx sensu
lato based on this specimen.
Trionychidae nomen dubium
Amyda menneri Chkhikvadze in Chkhikvadze &
Shuvalov, 1988
1988 Amyda menneri Chkhikvadze in Chkhikvadze &
Shuvalov; Chkhikvadze & Shuvalov: 200, fig. 1.
1999 Amyda menneri Chkhikvadze in Chkhikvadze &
Shuvalov; Khosatzky: 147.
Cretaceous soft-shelled turtles (Trionychidae) of Mongolia 25
Dow
nloa
ded
by [
Igor
G. D
anilo
v] a
t 11:
45 2
1 Fe
brua
ry 2
014

2000 ‘Amyda’ menenri (sic.) Chkhikvadze in
Chkhikvadze & Shuvalov; Sukhanov: 345.
2009 ‘Amyda’ menneri; Chkhikvadze in Chkhikvadze &
Shuvalov; Danilov & Vitek: 54.
2010 ‘Amyda’ menneri; Chkhikvadze in Chkhikvadze &
Shuvalov; Vitek & Danilov: table 1, fig. 1.
2011a Amyda menneri Chkhikvadze in Chkhikvadze &
Shuvalov; Danilov et al.: 14, 15 (part).
2011b Amyda menneri Chkhikvadze in Chkhikvadze &
Shuvalov; Danilov et al.: 95, 96 (part).
2012a ‘Amyda’ menneri; Chkhikvadze in Chkhikvadze &
Shuvalov; Danilov & Vitek: 421.
Holotype. IPGAS 11-5-1, incomplete postcrania of one
individual, including a nuchal, the proximal part of costal
1, a fragment of the right hyo- and hypoplastron, a frag-
ment of a xiphiplastron, and assorted non-shell bones,
from Gurilin Tsav, Umunugovi Aimag, Mongolia, Nem-
egt Formation, Maastrichtian.
Paratypes. IPGAS 11-5-2, nuchal fragment, IPGAS 11-
5-3, anterior part of carapace, IPGAS 11-5-4, distal part
of right hyoplastron, IPGAS 11-5-5, right costal 7, and
IPGAS 11-5-6, right hyoplastron, all from holotype
locality; IPGAS 11-13-11, medial part of right hyoplas-
tron, from Bugin Tsav, Umunugovi Aimag, Mongolia,
Nemegt Formation, Maastrichtian; IPGAS 11-14-2, left
posterior part of carapace, IPGAS 11-14-3, medial part
of left hypoplastron, and IPGAS 11-14-4, posterior part
of carapace, all from Ingeni Khobur, Umunugovi Aimag,
Mongolia, Nemegt Formation, Maastrichtian; IPGAS 11-
17-1, left half of carapace, from Bamba Khuduk, Umunu-
govi Aimag, Mongolia, Nemegt Formation, Maastrichtian.
Remarks. Amyda menneri was described based on iso-
lated shell fragments from several localities of the
Nemegt Formation of Mongolia (Chkhikvadze & Shu-
valov 1988; see Danilov & Vitek 2012a for the English
diagnosis of this species). Chkhikvadze & Shuvalov
(1988) suggested that material called ‘Trionyx sp. a’ by
Khosatzky & M»ynarski (1971) from a different locality
(Tsagan Khushu) in the Nemegt Formation might
belong to A. menneri. Danilov et al. (2011a, b) sug-
gested that most forms of trionychids known from the
Barungoyot and Nemegt formations of Mongolia are
attributable to Amyda menneri. Danilov & Vitek
(2012a, p. 423) reported that “figures in the published
type description [of Amyda menneri] are of such low
quality that we have found it difficult to meaningfully
compare the above-listed specimens [see Referred mate-
rial] with those of other trionychids”. Those authors
mentioned some similarities between A. menneri and
Apalonina (in the reduced costal 8), but conservatively
assigned the species to Trionychini. They also sug-
gested that some of the materials mentioned by Kho-
satzky (1999, p. 147) from the Nemegt Formation may
also belong to this species. However, results of the
present study show that the diversity of trionychids of
the Nemegt Formation is higher than considered previ-
ously. Given that material of Amyda menneri is very
fragmentary and comes from different localities, it may
belong to multiple trionychid taxa. Some of this mate-
rial (IPGAS 11-14-4; Chkhikvadze & Shuvalov 1988,
fig. 1E) shows similarities with Gobiapalone spp. in the
shape of costals 6–8, whereas other figured specimens,
including the holotype (IPGAS 11-5-1, IPGAS 11-5-6,
IPGAS 11-14-3; Chkhikvadze & Shuvalov 1988, fig.
1“A–)”), can be identified only as Trionychidae indet.
For this reason, we consider all material of Amyda men-
neri to be Trionychidae indet. and Amyda menneri to be
a nomen dubium.
Figure 15. Carapace of ‘Trionyx’ shiluutulensis sp. nov., MPC 25/166, from the unknown formation (Campanian) of Shiluut Ula in: A,dorsal and B, ventral views. Reconstructed outlines are indicated by dashed lines.
26 I. G. Danilov et al.
Dow
nloa
ded
by [
Igor
G. D
anilo
v] a
t 11:
45 2
1 Fe
brua
ry 2
014

Figure 16. Shell material of Late Cretaceous Trionychidae of Mongolia from different localities. A, Gobiapalone sp., MPC 25/165,incomplete carapace from the Lower White Beds of Khermin Tsav in dorsal view. B–X, Trionychidae indet: B, C, material from theDjadokhta Formation of Abdarain Nuru; B, ZIN PH 1/159, partial nuchal in dorsal view; C, ZIN PH 2/159, partial right hyo- andhypoplastra in ventral view; D, ZPAL MgCh, unnumbered, costal fragment from the Djadokhta Formation of Bayn Dzak in dorsal view;E, PIN 3458-4, partial carapace from the upper part of the Baynshire Formation of Bayshin Tsav in dorsal view; F, ZPAL MgCh/71,fragment of the carapace on a cast of the internal cavity of the shell from the ?Nemegt Formation of Char Teeg in dorsal view; G, ZINPH 1/163, neural from the Nemegt Formation of Gilbentu; H–J, material from unknown formation of Khamdin Khural; H, ZIN PH 1/162, shell fragment; I, ZIN PH 2/162, lateral fragment of left hyoplastron in ventral view; J, middle part of hyoplastron in ventral view;K, L, material from the Baynshire Formation of Khara-Khutul; K, ZIN PH 52/80, medial part of right hypoplastron in ventral view; L,ZIN PH 53/80, middle part of hypoplastron in ventral view; M, MPC 25/167 (HMNS 2006-04-477), shell fragment from the UpperWhite Bed of Khermin Tsav II; N, ZIN PH 1/164, lateral costal fragment from the Nemegt Formation of Khermin Tsav in dorsal view;O, ZIN PH 2/88, partial nuchal from the Baynshire Formation of Khongil Tsav in dorsal view; P, ZIN PH 1/166, lateral hyoplastronfragment from the lower part of the Baynshire Formation of Ongon Ulan Ula in ventral view; Q, ZIN PH 2/81, partial costal from theBaynshire Formation of Shine Us Khuduk in dorsal view; R, ZIN PH 1/167, lateral costal fragment from the upper part of the BaynshireFormation of Shiregin Gashun; S, ZIN PH 1/168, costal fragment from the unknown formation of Tel Ulan Shalcha in dorsal view; T,ZIN PH 3/24, partial right hyo- and hypoplastra from the Nemegt Formation of Tsagan Khushu in ventral view; U, V, material from theBaynshire Formation of Tsagan Teg; U, ZIN PH 2/91, costal fragment in dorsal view; V, ZIN PH 3/91, posterior neural in dorsal view;W, ZIN PH 1/169, lateral costal fragment from the lower part of the Baynshire Formation of Tugrikin Ula in dorsal view; X, ZIN PH 8/97, lateral fragment of hyo- or hypoplastron from the unknown locality in ventral view. Reconstructed outlines are indicated by dashedlines.
Cretaceous soft-shelled turtles (Trionychidae) of Mongolia 27
Dow
nloa
ded
by [
Igor
G. D
anilo
v] a
t 11:
45 2
1 Fe
brua
ry 2
014

Discussion
Cretaceous trionychids of MongoliaThe material described in this study belongs to seven trio-
nychid species: the cyclanorbine Nemegtemys conflata,
and the trionychines Gobiapalone breviplastra, G. orlovi,
‘Trionyx’ baynshirensis, ‘T.’ gilbentuensis, ‘T.’ gobiensis
and ‘T.’ shiluutulensis.
Nemegtemys conflata is known from two plastral frag-
ments from the Nemegt Formation. This taxon is assigned
to Cyclanorbinae based on the fusion of the hyo- and hypo-
plastra at a small size (just after hatching) (see Meylan
1987). Nemegtemys conflata, if correctly assigned, is the
first Cretaceous and the earliest known cyclanorbine. Previ-
ously, fossil cyclanorbines were known from a series of
African and Arabian localities that date as far back as the
Early Miocene (see Lapparent de Broin (2000) for sum-
mary of cyclanorbine fossil record). Plastomenidae, known
from the Campanian to Eocene of North America, are now
considered to be stem-cyclanorbines (Joyce et al. 2009;
Joyce & Lyson 2010, 2011). According to Danilov & Vitek
(2012a), no diagnosable specimens of Plastomenidae are
known from the Cretaceous of Asia.
Gobiapalone breviplastra and G. orlovi are members of
the genus Gobiapalone. This genus is assigned to Triony-
chinae based on a combination of the following charac-
ters: the dorsal edge of the apertura narium externa is
slightly to strongly emarginate, the foramen jugulare pos-
terius is isolated from the fenestra postotica by a descend-
ing arch of the opisthotic, the nuchal bone is at least three
times wider than long, and the neural series always con-
tains at least one reversal in neural orientation (synapo-
morphies of Trionychinae sensu Meylan 1987). In
addition, it lacks the following synapomorphies of Plasto-
menidae (Joyce & Lyson 2011): the presence of a preneu-
ral, contact between the xiphiplastra along their entire
length, the formation of an extensive, infolded secondary
palate by the maxillae, and an extremely elongated mandi-
ble. Gobiapalone is assigned to Trionychini based on the
following characters: the parietal makes up nearly one-
quarter of the processus trochlearis oticum, and the pres-
ence of seven neurals (Meylan 1987).
Gobiapalone breviplastra is represented by abundant
shell material and some non-shell postcrania from the
Nemegt Formation. We refer historical specimens that
had been reported under different names to this species
(see Systematic palaeontology section for complete list of
synonymies). The material of this species demonstrates a
high degree of variation in shell morphology (see Descrip-
tion). This species differs from other trionychine species
of the Nemegt Formation (‘Trionyx’ gilbentuensis and
‘T.’ gobiensis) in having a medium carapace length (up to
260 mm), suprascapular fontanelles (at carapace length
up to 224 mm), and type A shell sculpturing, as well as
differing in the shape of the hyo- and hypoplastra.
Gobiapalone orlovi is the best known trionychid from
the Late Cretaceous of Mongolia. Remains include a com-
plete skeleton, several mostly complete carapaces, and a
plastral fragment from the Baynshire Formation. Gobiap-
alone orlovi has a long list of synonymies and material of
this species demonstrates a high degree of variation in the
shell morphology (see the account of this species), similar
to G. breviplastra. This species differs from ‘Trionyx’
baynshirensis (the second trionychid of the Baynshire
Formation) in its smaller size (up to 335 mm), type A shell
sculpturing and the shape of the hyo- and hypoplastra.
‘Trionyx’ baynshirensis is known from a single plastral
fragment from the Baynshire Formation. That species is
assigned to Trionychinae based on its similarity with the
trionychine ‘Trionyx’ kansaiensis. This species differs
from the other trionychid taxon known from the Baynshire
Formation, Gobiapalone orlovi, in its larger size (up to
500 mm), type B shell sculpturing, and the shape of the
hyo- and hypoplastra. These characters are also found in
‘Trionyx’ gilbentuensis from the Nemegt Formation.
‘Trionyx’ gilbentuensis is also known from a single
plastral fragment from the Nemegt Formation. This spe-
cies is assigned to Trionychinae based on its similarity
with the trionychine ‘Trionyx’ baynshirensis. It differs
from other trionychine species of the Nemegt Formation
(Gobiapalone breviplastra and ‘Trionyx’ gobiensis) in its
larger size (up to 500 mm), type B shell sculpturing, and
the shape of the hyo- and hypoplastra.
‘Trionyx’ gobiensis is known from three incomplete
carapaces and additional carapace fragments from the
Nemegt Formation. This species is assigned to Trionychi-
nae based on a combination of the following characters:
the nuchal bone is at least three times wider than long, the
neural series always contains at least one reversal in neu-
ral orientation (synapomorphies of Trionychinae sensu
Meylan 1987), and the preneural is absent (difference
from Plastomenidae which have a preneural; Joyce &
Lyson 2011). We refrain from assigning ‘T.’ gobiensis to
Trionychini based on the presence of seven neurals
(Meylan 1987), because this character is variable in ‘T.’
gobiensis and no other characters of Trionychini are
known in this species. ‘Trionyx’ gobiensis differs from
other trionychine species of the Nemegt Formation
(Gobiapalone breviplastra and ‘Trionyx’ gilbentuensis) in
its smaller size (up to 130 mm), absence of suprascapular
fontanelles, type A sculpturing, and large costals 8.
‘Trionyx’ shiluutulensis is known from a carapace from
an unknown formation of the Campanian. This species is
assigned to Trionychinae based on the combination of the
following characters: the nuchal is at least three times
wider than long, the neural series contains at least one
reversal in neural orientation (synapomorphies of Triony-
chinae sensu Meylan 1987), and eight neurals are present
(as compared to Plastomenidae which have seven neurals;
Joyce & Lyson 2011). This species differs from all other
28 I. G. Danilov et al.
Dow
nloa
ded
by [
Igor
G. D
anilo
v] a
t 11:
45 2
1 Fe
brua
ry 2
014
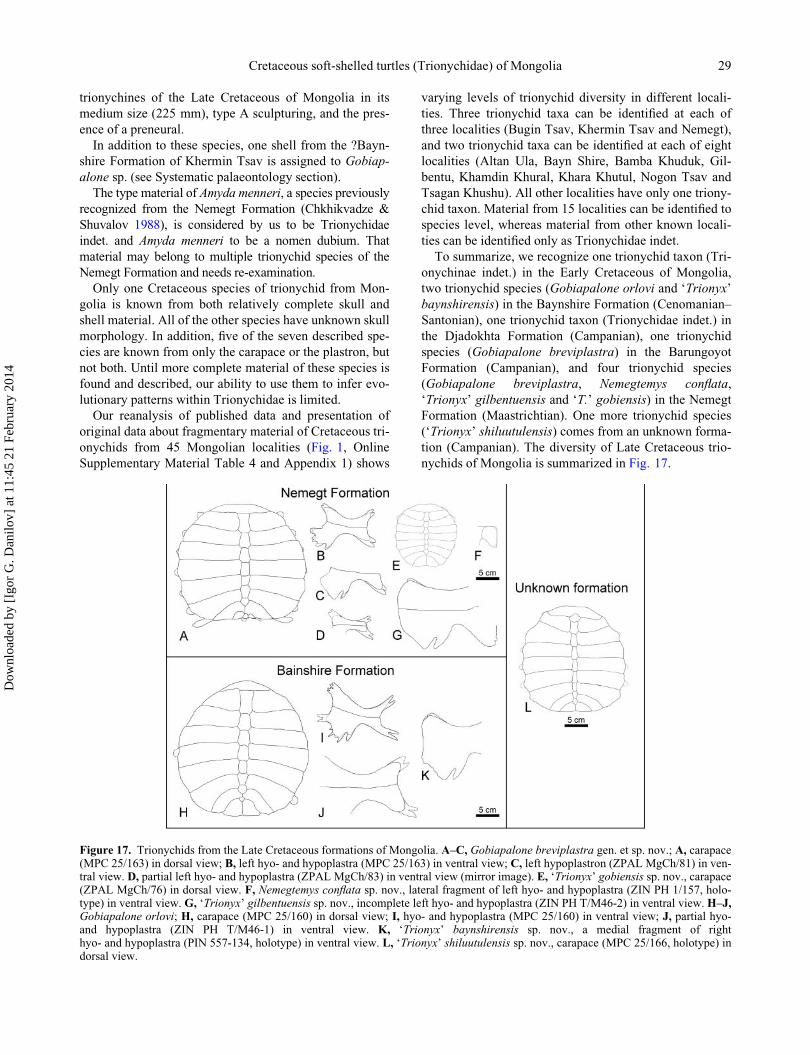
trionychines of the Late Cretaceous of Mongolia in its
medium size (225 mm), type A sculpturing, and the pres-
ence of a preneural.
In addition to these species, one shell from the ?Bayn-
shire Formation of Khermin Tsav is assigned to Gobiap-
alone sp. (see Systematic palaeontology section).
The type material of Amyda menneri, a species previously
recognized from the Nemegt Formation (Chkhikvadze &
Shuvalov 1988), is considered by us to be Trionychidae
indet. and Amyda menneri to be a nomen dubium. That
material may belong to multiple trionychid species of the
Nemegt Formation and needs re-examination.
Only one Cretaceous species of trionychid from Mon-
golia is known from both relatively complete skull and
shell material. All of the other species have unknown skull
morphology. In addition, five of the seven described spe-
cies are known from only the carapace or the plastron, but
not both. Until more complete material of these species is
found and described, our ability to use them to infer evo-
lutionary patterns within Trionychidae is limited.
Our reanalysis of published data and presentation of
original data about fragmentary material of Cretaceous tri-
onychids from 45 Mongolian localities (Fig. 1, Online
Supplementary Material Table 4 and Appendix 1) shows
varying levels of trionychid diversity in different locali-
ties. Three trionychid taxa can be identified at each of
three localities (Bugin Tsav, Khermin Tsav and Nemegt),
and two trionychid taxa can be identified at each of eight
localities (Altan Ula, Bayn Shire, Bamba Khuduk, Gil-
bentu, Khamdin Khural, Khara Khutul, Nogon Tsav and
Tsagan Khushu). All other localities have only one triony-
chid taxon. Material from 15 localities can be identified to
species level, whereas material from other known locali-
ties can be identified only as Trionychidae indet.
To summarize, we recognize one trionychid taxon (Tri-
onychinae indet.) in the Early Cretaceous of Mongolia,
two trionychid species (Gobiapalone orlovi and ‘Trionyx’
baynshirensis) in the Baynshire Formation (Cenomanian–
Santonian), one trionychid taxon (Trionychidae indet.) in
the Djadokhta Formation (Campanian), one trionychid
species (Gobiapalone breviplastra) in the Barungoyot
Formation (Campanian), and four trionychid species
(Gobiapalone breviplastra, Nemegtemys conflata,
‘Trionyx’ gilbentuensis and ‘T.’ gobiensis) in the Nemegt
Formation (Maastrichtian). One more trionychid species
(‘Trionyx’ shiluutulensis) comes from an unknown forma-
tion (Campanian). The diversity of Late Cretaceous trio-
nychids of Mongolia is summarized in Fig. 17.
Figure 17. Trionychids from the Late Cretaceous formations of Mongolia. A–C, Gobiapalone breviplastra gen. et sp. nov.; A, carapace(MPC 25/163) in dorsal view; B, left hyo- and hypoplastra (MPC 25/163) in ventral view; C, left hypoplastron (ZPAL MgCh/81) in ven-tral view. D, partial left hyo- and hypoplastra (ZPAL MgCh/83) in ventral view (mirror image). E, ‘Trionyx’ gobiensis sp. nov., carapace(ZPAL MgCh/76) in dorsal view. F, Nemegtemys conflata sp. nov., lateral fragment of left hyo- and hypoplastra (ZIN PH 1/157, holo-type) in ventral view. G, ‘Trionyx’ gilbentuensis sp. nov., incomplete left hyo- and hypoplastra (ZIN PH T/M46-2) in ventral view. H–J,Gobiapalone orlovi; H, carapace (MPC 25/160) in dorsal view; I, hyo- and hypoplastra (MPC 25/160) in ventral view; J, partial hyo-and hypoplastra (ZIN PH T/M46-1) in ventral view. K, ‘Trionyx’ baynshirensis sp. nov., a medial fragment of righthyo- and hypoplastra (PIN 557-134, holotype) in ventral view. L, ‘Trionyx’ shiluutulensis sp. nov., carapace (MPC 25/166, holotype) indorsal view.
Cretaceous soft-shelled turtles (Trionychidae) of Mongolia 29
Dow
nloa
ded
by [
Igor
G. D
anilo
v] a
t 11:
45 2
1 Fe
brua
ry 2
014

The presence of four species of trionychids in the
Nemegt Formation of Mongolia marks it as amongst the
most diverse localities in the entire known record of trio-
nychid evolution, including the modern day. The only
other known localities with comparable diversity are the
Campanian Judith River Formation in Canada (five spe-
cies), the Eocene Bridger Formation in the western USA
(at least four species), and the modern day fauna of
Java, Indonesia (four species: Pritchard 2001; Vitek
2012).
Phylogenetic position of GobiapalonePhylogenetic analysis 1 resulted in four shortest trees of
248 steps each (CI ¼ 0.45, RI ¼ 0.63). The strict consen-
sus tree is shown in Fig. 18A. Phylogenetic analysis 2
resulted in 24 most parsimonious trees of 294 steps each
(CI ¼ 0.39, RI ¼ 0.61). The strict consensus tree is shown
in Fig. 18B.
In both analyses Gobiapalone is monophyletic. In anal-
ysis 1, Gobiapalone is placed within Apalonina Meylan,
1987 and forms a polytomy with Rafetus euphraticus
(Daudin, 1802) and Apalone Rafinesque, 1832. The mono-
phyly of Gobiapalone is supported by three characters: a
nuchal width-to-length ratio greater than four, the pres-
ence of seven or eight neurals, and the presence of five
plastral callosities. Apalonina is supported by five charac-
ters: the vomer divides the maxillae, the vomer reaches
the intermaxillary foramen, the average ratio of the inter-
maxillary foramen length to the length of the primary pal-
ate is about 60%, seven or more ossifications in cornu
branchiale II are present, and costals 8 are reduced or
absent (Meylan 1987).
In analysis 2, Gobiapalone is united with Oliveremys
uintaensis (Leidy, 1872) in a clade that is sister to Apalo-
nina. The monophyly of Gobiapalone is supported only
by the presence of five plastral callosities. The clade
Gobiapalone þ Oliveremys is supported by two charac-
ters: a nuchal width-to-length ratio greater than four, and
a postorbital bar with a width about equal or a third of the
diameter of the orbit. This clade is united with Apalonina
by one character: the posterior edge of neural 5 (not
counting the preneural) lies anterior to the posterior edge
of costal 5. The clade Apalonina þ (Gobiapalone þ Oli-
veremys) forms a polytomy with ‘Trionyx’ egregius Hay,
1908 and a clade uniting extinct species of the genera
Aspideretoides and Axestemys Hay, 1899. The results of
analysis 2 are different from the strict consensus tree in
Figure 18. Strict consensus trees resulting from A, analysis 1, and B, analysis 2 of fossil and recent Trionychidae based on morphologicaldata (see Discussion for description of the trees). Numbers below nodes are Bremer support indices (0 indexes are not shown).
30 I. G. Danilov et al.
Dow
nloa
ded
by [
Igor
G. D
anilo
v] a
t 11:
45 2
1 Fe
brua
ry 2
014

the analysis of Vitek (2012, fig. 2.1) with respect to the
better resolution within Trionychinae, and are more simi-
lar to a tree resulting from the same data matrix but delet-
ing Nilssonia formosa (Grey, 1869) and the fossil
‘Trionyx’ egregius (see Vitek 2012, fig. 2.2). However,
unlike the analysis of Vitek (2012), analysis 2 does not
support monophyly of Axestemys.
Thus, the results of analyses 1 and 2 show that Apalo-
nina, which is a relatively derived and well-supported trio-
nychid clade (Meylan 1987; Gardner et al. 1995;
Engstrom et al. 2004; Joyce et al. 2009; Joyce & Lyson
2011; Vitek 2011, 2012; this study), or its closest sister
taxa (stem-Apalonina) were already present in the Late
Cretaceous of Asia. Their presence suggests that most
other supra-generic clades of modern trionychids had
already evolved in Asia by the Late Cretaceous (Danilov
et al. 2011a, b) and should be present in the fossil record.
This hypothesis is supported by the discovery of a cycla-
norbine, Nemegtemys conflata, in the Late Cretaceous of
Mongolia. Another trionychid clade, formed by the sister
relationship of the extinct genera Aspideretoides and
Axestemys (Vitek 2012; this study), is also present in the
Late Cretaceous of Asia and represented by Aspidere-
toides spp. (Vitek & Danilov 2010; Danilov & Vitek
2012a, b). Members of other crown-clades that should
have been present given the hypotheses discussed above,
such as Chitrini, Pelodiscini, Aspideretini and Gigantae-
suarochelys, are still unknown in the published
Cretaceous fossil record (for a review see Danilov &
Vitek 2012a). Their absence and long ghost lineages in
general highlight a need for further collection, description
and revision of Cretaceous–Holocene fossil material of
trionychids. In particular, it is possible that some stem- or
crown-clade members of these clades are already recorded
in the literature, but unidentified as members of modern
clades because of a general lack of recent re-examination
of known fossils from Europe and Asia.
Distribution of Cretaceous Trionychidae in AsiaSince the Cretaceous trionychids of North America and
Asia were last reviewed (Danilov & Vitek 2012a), newly
published data on the subject can refine or revise our
understanding of the biogeography and evolutionary pat-
terns of trionychids (Liu et al. 2011; Danilov & Vitek
2012b; Hirayama et al. 2012; Vitek 2012; Vitek &
Danilov 2012).
Liu et al. (2011) reported a new pan-trionychid (¼ stem-
trionychid) from the Jiufotang Formation (Aptian) of west-
ern Liaoning, China. Danilov & Vitek (2012b) described
trionychid shell material from the Bissekty Formation (late
Turonian) of Uzbekistan. This material was assigned to
two shell-based taxa, Aspideretoides cf. riabinini and
‘Trionyx’ cf. kansaiensis. Skull-based taxa from the same
formation are Khunnuchelys kizylkumensis and Trionychini
indet. Hirayama et al. (2012) described the stem-
Figure 19. Temporal and geographical distributions of Cretaceous Trionychidae of Asia and North America. Gaps in record are filledwith grey. �denotes skull-only taxa. See text for the Mongolian record. ‘Stem-trionychid’ denotes an occurrence from Liu et al. (2011).Other data are after Danilov & Vitek (2012a), with additions and modifications from Danilov & Vitek (2012b), Hirayama et al. (2012),Vitek (2012) and Vitek & Danilov (2012) (see Discussion for details).
Cretaceous soft-shelled turtles (Trionychidae) of Mongolia 31
Dow
nloa
ded
by [
Igor
G. D
anilo
v] a
t 11:
45 2
1 Fe
brua
ry 2
014

trionychid, Kappachelys okurai Hirayama et al., 2012,
based on two isolated carapacial elements from the Early
Cretaceous (?late Neocomian) Akaiwa Formation of west-
central Honshu, Japan. Vitek (2012) published a revision
of North American trionychids of the genus Axestemys.
According to this revision, Axestemys was present in the
Late Cretaceous (Campanian and Maastrichtian) of North
America. Vitek & Danilov (2012) described trionychid
shell material from the Santonian–?middle Campanian
Kyrkkuduk I locality in southern Kazakhstan. This material
was assigned to two taxa, ‘Trionyx’ kansaiensis and Trio-
nychidae indet. ‘Trionyx’ kansaiensis from Kyrkkuduk I
corresponds to Trionychinae indet. 1 of Danilov & Vitek
(2012a). An updated summary of the temporal and geo-
graphical distributions of Cretaceous Trionychidae of Asia
and North America is provided in Fig. 19.
In general, new data supports the recognition of two
patterns. First, the earliest and only known stem-triony-
chids are from the Early Cretaceous of Japan and China,
and support the hypothesis that trionychids evolved some-
where in Asia (Danilov 2005; Danilov & Vitek 2012a).
Second, two lineages of trionychids persisted in Middle
Asia and Kazakhstan throughout the Late Cretaceous.
Danilov & Vitek (2012a) identified three patterns of
distribution of Cretaceous trionychids in Asia: (1) repre-
sentatives of the genus Aspideretoides (Trionychini with a
preneural) are known only from Middle Asia and Kazakh-
stan; (2) Apalonina indet. and other Trionychini without a
preneural are known only from Mongolia; and (3) mem-
bers of Trionychinae with eight neurals have the broadest
geographical distribution and are present in Middle Asia,
Kazakhstan, Mongolia and China. Our study updates these
patterns. We show that Trionychini with a preneural had a
broader distribution and were also present in Mongolia
(‘Trionyx’ shiluutulensis).
Conclusions
A revision of the available material of Cretaceous triony-
chids of Mongolia allows us to recognize seven species,
six of which are new. Amongst these species are the oldest
known cyclanorbine, Nemegtemys conflata, two species of
the genus Gobiapalone, which is a close relative of mod-
ern Apalonina, and four other species (‘Trionyx’ baynshir-
ensis, ‘T.’ gilbentuensis, ‘T.’ gobiensis, and ‘T.’
shiluutulensis). Some of these species are based on speci-
mens collected by the Mongolian Palaeontological Expe-
dition (1946, 1948, 1949) that remained undescribed and
unrecognized for more than 60 years. Most of these spe-
cies are based on incomplete material. Some of the revised
material, including several skulls, needs additional prepara-
tion and study. In spite of the recent contributions to our
knowledge of Cretaceous trionychids of Asia and North
America, there are still many gaps in the record, and the
phylogenetic position of most taxa of Cretaceous triony-
chids remains poorly understood. In particular, the relation-
ship of Nemegtemys conflata to other cyclanorbines and
plastomenids remains an open question, as does the rela-
tionship of ‘Trionyx’ kansaiensis and similar Mongolian
taxa to other trionychids. Early representatives of crown
clades other than Apalonina are still missing from the
known fossil record. New discoveries of more complete
material of Cretaceous trionychids of Mongolia are needed
to further improve our knowledge about diversity of Creta-
ceous trionychids of Mongolia and Asia in general and
understanding of the early diversification of this family.
Acknowledgements
We thank Magdalena Borsuk-Bia»ynicka for access to tri-
onychid specimens in ZPAL and E. V. Syromyatnikova
(ZIN) for providing photographs of these specimens. We
thank Walter Joyce and an anonymous reviewer for useful
comments and suggestions. This study was supported by
the grants of the President of Russian Federation to the
Leading Scientific Schools [No. NSh-6560.2012.4],
Russian Foundation for Basic Research [No. 14-04-
00416_a], and Hirayama Ikuo Silk Road Museum to I.G.D.
Supplemental material
Supplemental material for this article can be accessed
here: http://dx.doi.org/[DOI: 10.1080/14772019.2013.
847870]
References
Batsch, A. J. G. C. 1788. Versuch einer Anleitung, zur Kenntnißund Geschichte der Thiere und Mineralien. AkademischeBuchhandlung, Jena, 528 pp.
Chkhikvadze, V. M. & Shuvalov, V. F. 1988. A new species ofa trionychid from the Upper Cretaceous deposits of Mongo-lia. Izvestiya Akademii Nauk Gruzinskoi SSR, 14, 198–204[In Russian].
Cope, E. D. 1868. On the origin of genera. Proceedings of theAcademy of Natural Sciences of Philadelphia, 20, 242–300.
Danilov, I. G. 2005. Die fossilen Schildkr€oten Europas. Pp.329–441 in U. Fritz (ed.) Handbuch der Reptilien undAmphibien Europas. Band 3/IIIB: Schildkr€oten (Testudines)II. Aula-Verlag, Wiebelsheim.
Danilov, I. G. & Syromyatnikova, E. V. 2008. New materialson turtles of the family Nanhsiungchelyidae from the Creta-ceous of Uzbekistan and Mongolia, with a review of thenanhsiungchelyid record in Asia. Proceedings of the Zoolog-ical Institute of the Russian Academy of Sciences, 312, 3–25.
Danilov, I. G. & Vitek, N. S. 2009. Cretaceous trionychids ofAsia: a review of record and biogeography. Pp. 52–58 inGaffney Turtle Symposium. Abstract Volume. Royal TyrrellMuseum, Drumheller.
32 I. G. Danilov et al.
Dow
nloa
ded
by [
Igor
G. D
anilo
v] a
t 11:
45 2
1 Fe
brua
ry 2
014

Danilov, I. G., Hirayama, R., Sukhanov, V. B.,Watabe, M. &Suzuki, S. 2011a. Turtles-trionychids of the Cretaceous ofMongolia: new data and a revision. Pp. 14–16 in Otchetnayanauchnaya sessiya po itogam rabot 2010 g. Tezisy dokladov.Zoological Institute of the Russian Academy of Sciences, St.Petersburg. [In Russian].
Danilov, I., Hirayama, R., Sukhanov, V., Watabe, M. &Suzuki, S. 2011b. Cretaceous softshelled turtles (Trionychi-dae) of Mongolia: new data and a revision. Journal of Verte-brate Paleontology, 31 (Supplement 2), 95–96.
Danilov, I. G. & Vitek, N. S. 2012a. Chapter 23. Cretaceous tri-onychids of Asia: an expanded review of their record andbiogeography. Pp. 419–438 in D. B. Brinkman, P. A.Holroyd & J. D. Gardner (eds)Morphology and Evolution ofTurtles. Springer, Berlin.
Danilov, I. G. & Vitek, N. S. 2012b. Soft-shelled turtles (Trio-nychidae) from the Bissekty Formation (Late Cretaceous:late Turonian) of Uzbekistan: Shell-based taxa. CretaceousResearch, 41, 55–64.
Daudin, F. M. 1802. Histoire Naturelle, G�enerale et Partic-uli�ere des Reptiles (Vol. 2). F. Dufart, Paris, 432 pp.
Engstrom, T. N., Shaffer, H. B. & McCord, W. P. 2004. Mul-tiple data sets, high homoplasy, and the phylogeny of soft-shell turtles (Testudines: Trionychidae). Systematic Biology,53, 693–710.
Eriksson, T. 1998. AutoDecay. 4.0.1. Department of Botany,Stockholm University, Stockholm.
Ernst, C. H. & Barbour, R. W. 1989. Turtles of the World.Smithsonian Institution Press, Washington, D.C., 313 pp.
Forska�l, O. 1775. Descriptiones Animalium. Avium, amphibio-
rum, piscium, insectorum, vermium. Quae in itinere orientalobservavit. Hauniae, M€olleri, 164 pp.
Gardner, J. D. & Russell, A. P. 1994. Carapacial variationamong soft-shelled turtles (Testudines: Trionychidae), andits relevance to taxonomic and systematic studies of fossiltaxa. Neues Jahrbuch f€ur Geologie und Pal€aontologie, 193(2), 209–244.
Gardner, J. D., Russell, A. P. & Brinkman, D. B. 1995. Sys-tematics and taxonomy of soft-shelled turtles (Family Trio-nychidae) from the Judith River Group (mid-Campanian) ofNorth America. Canadian Journal of Earth Sciences, 32,631–643.
Geoffroy Saint Hillaire, M. 1809. Sur les tortues molles,nouveau genre sous le nom de Trionyx et sur la formationdes carapaces. Annales du Mus�eum d’Histoire Naturelle, 14,1–20.
Gilmore, C. W. 1928. Description of two new species of fossilturtles, from the Lance Formation of Wyoming. Proceedingsof the US National Museum, 50, 641–646.
Gilmore, C. W. 1931. Fossil turtles of Mongolia. Bulletin of theAmerican Museum of Natural History, 59, 213–257.
Gray, J. E. 1825. A synopsis of the genera of reptiles andamphibia, with a description of some new species. Annals ofPhilosophy, 10, 193–217.
Gray, J. E. 1831. Synopsis Reptilium; or short descriptions ofthe species of reptiles. Part I. Cataphracta. Tortoises, croco-diles, and enaliosaurians, London: Treuitel, Wiirtz and Co.,78 pp.
Gray, J. E. 1869. Notes on the families and genera of tortoises(Testudinata), and on the characters afforded by the study oftheir skulls. Proceedings of the Zoological Society of Lon-don, 1869, 165–225.
Hay, O. P. 1899. On the nomenclature of certain American fos-sil vertebrates. American Geologist, 24, 345–349.
Hay, O. P. 1902. Bibliography and catalogue of fossil verte-brates of North America. United States Geological SurveyBulletin, 179, 1–449.
Hay, O. P. 1908. The fossil turtles of North America. CarnegieInstitute of Washington Publications, 75, 1–568.
Hirayama, R., Isaji, S. & Hibino, T. 2012. Kappachelys okuraigen. et sp. nov., a new stem soft-shelled turtle from the EarlyCretaceous of Japan. Pp. 179–185 in D. B. Brinkman, P. A.Holroyd & J. D. Gardner (eds) Morphology and evolution ofturtles. Springer, Dordrecht.
Hummel, K. 1929. Die fossilen Weichschildkr€oten (Triony-chia). Eine morphologisch-systematische und stammesge-schichtliche studie. Geologische und Palaeontologische,Abhandlungen, 16, 359–487.
Hutchison, J. H. 2000. Diversity of Cretaceous turtle faunas ofeastern Asia and their contribution to the turtle faunas ofNorth America. Paleontological Society of Korea SpecialPublication, 4, 27–38.
Hutchison, J. H. 2009. New soft-shelled turtles (Plastomeninae:Trionychidae: Testudines) from the Late Cretaceous andPaleocene of North America. Paleobios, 29, 36–47.
Joyce, W. G., Parham, J. F. & Gauthier, J. A. 2004. Develop-ing a protocol for the conversion of rank-based taxon namesto phylogenetically defined clade names, as exemplified byturtles. Journal of Paleontology, 78, 989–1013.
Joyce, W. G., Revan, A., Lyson, T. R. & Danilov, I. G. 2009.Two new plastomenine softshell turtles from the Paleoceneof Montana and Wyoming. Bulletin of the Peabody Museumof Natural History, 50, 307–325.
Joyce, W. G. & Lyson, T. R. 2010. A neglected lineage ofNorth American turtles fills a major gap in the fossil record.Palaeontology, 53, 241–248.
Joyce, W. G. & Lyson, T. R. 2011. New material of Gil-moremys lancensis nov. comb. (Testudines: Trionychi-dae) from the Hell Creek Formation and the diagnosisof plastomenid turtles. Journal of Paleontology, 85,442–459.
Karl, H.-V. 1998. Zur Taxonomie der k€anozoischen Weich-schildkr€oten €Osterreichs und Deutschlands (Trionychidae:Trionychinae). Mitteilungen der Abteilungs f€ur Geologieund Pal€aontologie der Landesmuseum Joanneum, 56, 273–328.
Karl, H.-V. 1999. Die Zoogeographie der k€anozoischenWeichschildkr€ote Trionyx triunguis Forsk€al, 1775. JoanneaGeoogie und Pal€aontologie, 1, 27–60.
Khosatzky, L. I. & M»ynarski, M. 1971. Chelonians from theUpper Cretaceous of the Gobi Desert, Mongolia. Palaeonto-logia Polonica, 25, 131–144.
Khosatzky, L. I. 1976. A new representative of trionychids fromthe Late Cretaceous of Mongolia. Pp. 3–19 in Gerpetolo-giya. Kuban State University, Krasnodar. [In Russian].
Khosatzky, L. I. 1999. Turtles – trionychids of the Cretaceousof Mongolia. Pp. 141–149 in Voprosy Paleontologii. T. 11.St. Petersburg State University, St. Petersburg. [In Russian].
Kuznetsov, V. V., & Chkhikvadze, V. M. 1987. The Late Cre-taceous trionychids from Shakh-Shakh locality in Kazakh-stan. Materialy po Istorii Fauny i Flory Kazakhstana, 9, 33–39. [In Russian].
Lapparent de Broin, F. 2000. African chelonians from theJurassic to the Present. A preliminary catalog of the Africanfossil chelonians. Palaeontologica Africana, 36, 43–82.
Leidy, J. 1872. Notice of donation of fossils etc. fromWyoming.Proceedings of the Academy of Natural Sciences of Phila-delphia, 1872, 267–268.
Cretaceous soft-shelled turtles (Trionychidae) of Mongolia 33
Dow
nloa
ded
by [
Igor
G. D
anilo
v] a
t 11:
45 2
1 Fe
brua
ry 2
014

Liu, J., Joyce, W. & Tong, H. 2011. The oldest morphologi-cally modern soft-shelled turtle (Testudines, Pan-Trionychi-dae) from the Jehol biota of China. Journal of VertebratePaleontology, 31 (Supplement 2), 144.
Merkulova, N. N. 1978. A new Trionyx from the Nemegt(MPR). Byulleten’ Moskovskogo Obshchestva IspytateleyPrirody. Otdel Geologicheskiy, 53, 156. [In Russian].
Meylan, P. A. 1987. The phylogenetic relationships of soft-shelled turtles (Family Trionychidae). Bulletin of the Ameri-can Museum of Natural History, 186, 1–101.
M»ynarski, M. & Narmandach, P. 1972. New turtle remainsfrom the Upper Cretaceous of the Gobi Desert, Mongolia.Palaeontologia Polonica, 27, 95–102.
Page, R. D. M. 2001. NDE: NEXUSData Editor 0.5.0.Univer-sity of Glasgow, Glasgow, U.K.
Pritchard, P. C. H. 2001. Observations on body size, sympatry,and niche divergence in softshell turtles (Trionychidae).Chelonian Conservation and Biology, 4, 5–27.
Rafinesque, C. S. 1832. Description of two new genera of softshell turtles of North America. Atlantic Journal and Friendof Knowledge, 1, 64–65.
Sochava, A. V. 1975. Stratigraphy and lithology of the UpperCretaceous deposits of southern Mongolia. Trudy Sovmest-noy Sovetsko-Mongol’skoy Nauchno-Issledovatelskoy Geo-logicheskoy Ekspeditsii, 13, 113–182. [In Russian].
Shuvalov, V. F. & Chkhikvadze, V. M. 1975. New data on theLate Cretaceous turtles of South Mongolia. Trudy Sovmest-noy Sovetsko-Mongol’skoy Paleontologicheskoy Ekspeditsii,2, 214–229. [In Russian].
Shuvalov, V. F. & Chkhikvadze, V. M. 1979. On stratigraphicand systematic position of some freshwater turtles from newCretaceous localities in Mongolia. Trudy Sovmestnoy Sovet-sko-Mongol’skoy Paleontologicheskoy Ekspeditsii, 8, 58–76. [In Russian].
Sukhanov, V. B. 2000. Mesozoic turtles of Middle and CentralAsia. Pp. 309–367 in M. J. Benton, M. A. Shishkin, D. M.Unwin & E. N. Kurochkin (eds) The Age of Dinosaurs in Rus-sia and Mongolia. Cambridge University Press, Cambridge.
Suzuki, S. & Narmandakh, P. 2004. Change of the Cretaceousturtle faunas in Mongolia. Hayashibara Museum of NaturalSciences Research Bulletin, 2, 7–14.
Suzuki, S. 2005. An almost complete skeleton of the trionychidturtle (Trionychidae: Trionychinae) from the Upper Creta-ceous of Mongolia. P. 27 in Abstracts with Programs of the154th Regular Meeting of the Palaeontological Society ofJapan. Yamagata, Japan. [In Japanese].
Vitek, N. S.& Danilov, I. G. 2010. New material and a reassess-ment of soft-shelled turtles (Trionychidae) from the LateCretaceous of Middle Asia and Kazakhstan. Journal of Ver-tebrate Paleontology, 30, 383–393.
Vitek, N. S. 2011. Insights into the taxonomy and systematics ofNorth American Eocene soft-shelled turtles from a well-pre-served specimen. Bulletin of the Peabody Museum of Natu-ral History, 52(2), 189–208.
Vitek, N. S. 2012. Giant fossil soft-shelled turtles of NorthAmerica. Palaeontologia Electronica, 15(1), 1–43.
Vitek, N. S. & Danilov, I. G. 2012. New data on the soft-shelledturtles from the Upper Cretaceous of Kyrkkuduk I localityof Southern Kazakhstan. Proceeding of the Zoological Insti-tute of the Russian Academy of Sciences, 316(1), 50–56.
Watabe, M., Tsogtbaatar, Kh., Suzuki, S. & Saneyoshi, M.2010. Geology of dinosaur-fossil-bearing localities (Jurassicand Cretaceous: Mesozoic) in the Gobi Desert: Results ofthe HMNS-MPC Joint Paleontological Expedition. Hayashi-bara Museum of Natural Sciences Research Bulletin, 3, 41–118.
Yeh, H.-K. 1965. New materials of fossil turtles of Inner Mon-golia. Vertebrata PalAsiatica, 9, 47–69.
34 I. G. Danilov et al.
Dow
nloa
ded
by [
Igor
G. D
anilo
v] a
t 11:
45 2
1 Fe
brua
ry 2
014

Table 4. Localities, formations and identifications of Cretaceous Trionychidae of Mongolia. “*” denotes identifications based on published data. No. Locality Formation(s) Identification(s) 1 Abdarain Nuru Djadokhta Trionychidae indet. 2 Abdrant Nuru Djadokhta Trionychidae indet.* 3 Alak Shand Khuduk Nemegt Gobiapalone breviplastra 4 Altan Ula, Altan Ula I, III
and IV Nemegt Gobiapalone breviplastra, ‘Trionyx’
gobiensis 5 Amtgai Baynshire Trionychidae indet.* 6 Bayn Shire Baynshire Gobiapalone orlovi, ‘Trionyx’
baynshirensis 7 Bamba Khuduk Nemegt ‘Trionyx’ gobiensis, Trionychinae indet. 8 Bayn Dzak Djadokhta Trionychidae indet. 9 Bayshin Tsav Baynshire (upper part) Trionychidae indet.
10 Bugin Tsav Barungoyot and Nemegt Gobiapalone breviplastra (Barungoyot), G. breviplastra, Nemegtemys conflata
(Nemegt) 11 Burkhant Baynshire Gobiapalone orlovi 12 Char Teeg ?Nemegt Trionychidae indet. 13 Dersnii Khuduk Baynshire (lower part) Trionychidae indet.* 14 Dzun Shakhai Unknown (Early Cretaceous) Trionychidae indet.* 15 Ekhin Tukhum Baynshire (lower part) Trionychidae indet.* 16 Erdeni Ula Khulsangol Trionychidae indet.* 17 Gilbentu Nemegt ‘Trionyx’ gilbentuensis, Trionychidae
indet. 18 Gurilin Tsav Nemegt Trionychidae indet.* 19 Ingeni Khobur and Ingeni
Khobur III Nemegt Trionychidae indet.*
20 Ingeni Tsav Barungoyot and Nemegt Trionychidae indet.* 21 Khamdin Khural Unknown (Late Cretaceous) Trionychidae indet. (two forms) 22 Khara Khutul Baynshire Trionychidae indet. (two forms) 23 Khermin Tsav and
Khermin Tsav II ?Baynshire and Nemegt Gobiapalone sp. (?Baynshire),
Trionychidae indet (two forms; Nemegt) 24 Khongil Baynshire Gobiapalone orlovi 25 Khongil Obo Baynshire (upper part) Trionychidae indet.* 26 Knongil Tsav Baynshire Trionychidae indet. 27 Khuren Tsav Barungoyot (lower part) Trionychidae indet.* 28 Nemegt Nemegt Gobiapalone breviplastra, Nemegtemys
conflata, ‘Trionyx’ gobiensis 29 Nogon Tsav and Nogon
Tsav II Barungoyot (upper part) and
Nemegt Gobiapalone breviplastra (Barungoyot),
Trionychidae indet. (Nemegt) 30 Ongon Ulan Ula Baynshire (lower part) Trionychidae indet. 31 Shiluut Ula Unknown (Campanian) ‘Trionyx’ shiluutulensis 32 Shine Us Khuduk Baynshire Trionychidae indet. 33 Shine Usny Tolgod Unknown (Early Cretaceous) Trionychidae indet.* 34 Shiregin Gashun Baynshire (upper part) Trionychidae indet. 35 Tel Ulan Shalcha Unknown (probably, early Late
Cretaceous) Trionychidae indet.
36 Tsagan Khushu Nemegt ‘Trionyx’ gobiensis, Trionychidae indet. 37 Tsagan Teg Baynshire Formation Trionychidae indet. 38 Tugrikin Ula Baynshire (lower part) Trionychidae indet. 39 Ulan Khushu Nemegt Gobiapalone breviplastra 40 Ulan Tsab Ula Probably, Baynshire Trionychidae indet.* 41 Unegetu Ula Baynshire Gobiapalone orlovi 42 Uryl’b Usu Baynshire (lower part) Trionychidae indet.* 43 Ushyin Khuduk Baynshire (upper part) Trionychidae indet.* 44 Yagaan Khovil Djadokhta Trionychidae indet.* 45 Unknown locality Unknown (?Late Cretaceous) Trionychidae indet.

Appendix 1. Review of records of Cretaceous Trionychidae in Mongolia. See Fig. 1 for map of
occurrences.
1. Abdarain Nuru, Northern Gobi.
Geology and age. Djadokhta Formation, Campanian.
Material and references. Partial nuchal, partial right hyo- and hypoplastra, and other shell
fragments of Trionychidae indet. (Fig. 16B, C; collection ZIN PH 159; collected by A.V.
Sochava in 1970; as “part of plastron”; Khosatzky 1999: 145).
Remarks. The proportions of the nuchal (that was at least four times wider than long)
correspond to Trionychinae or Plastomenidae. The outline of the preserved part of the hyo- and
hypoplastra and type A sculpturing are similar to those of Gobiapalone spp. However, because
plastral morphology is not known in most other Mongolian trionychids of the Cretaceous, this
material is identified as Trionychidae indet.
2. Abdrant Nuru, Umunugovi Aimag.
Geology and age. Djadokhta Formation, Campanian.
Material and references. Disarticulated shell fragments and several skulls of Trionychidae
indet. (as Trionychidae; Suzuki & Narmandakh 2004: 9).
3. Alak Shand Khuduk (SW from Alak Shand Khuduk spring), Umunugovi Aimag.
Geology and age. Nemegt Formation, Maastrichtian.
Material and references. Partial carapace of Gobiapalone breviplastra (see Systematic
palaeontology section).
Remarks. Alak Shand Khuduk spring is situated 30 km W from Bugin Tsav.
4. Altan Ula, Altan Ula I, III and IV, Umunugovi Aimag.
Geology and age. Nemegt Formation, Maastrichtian.
Material and references. Partial carapace on a cast of the internal cavity of the shell of
Gobiapalone breviplastra from Altan Ula I (see Systematic palaeontology section); shell
fragment of ‘Trionyx’ gobiensis from Altan Ula III (see Systematic palaeontology section); shell
fragments of Trionychidae indet. from Altan Ula (PIN 553-66, collected by MPE in 1948; as
“shell fragments of trionychids”; Khosatzky 1999: 147), Altan Ula III (collection ZPAL
MgCh/121, collected by PMPE) and Altan Ula IV (HMNS 2006-04-474, collected by JMJPE in
2006; as Trionychidae; Suzuki & Narmandakh 2004: 10).

Remarks. All shell fragments of Trionychidae indet. from Altan Ula, Altan Ula III and IV have
type A sculpturing.
5. Amtgai, Umunugovi Aimag.
Geology and age. Baynshire Formation, Cenomanian – Santonian.
Material and references. Shell fragments of Trionychidae indet. (as Trionychidae; Suzuki &
Narmandakh 2004: 8).
6. Bayn Shire (= Bayn Shire), Dornogovi Aimag.
Geology and age. Baynshire Formation, Cenomanian – Santonian.
Material and references. Two incomplete carapaces and hyo- and hypoplastra of Gobiapalone
orlovi from the lower part of the Baynshire Formation, Cenomanian – early Turonian (see
Systematic palaeontology section); medial fragment of hyo-and hypoplastron of ‘Trionyx’
baynshirensis (see Systematic palaeontology section); shell fragments and two cervical vertebrae
of Trionychidae indet. (collection ZIN PH 156; collected by MPE in 1948 and G.G. Martinson in
1967; Khosatzky 1999: 142); shell fragments of Trionychidae indet. (collection PIN 557,
collected by MPE in 1948; collection ZPAL MgCh/149, collected by PMPE in 1963).
Remarks. The shell fragments of Trionychidae indet. from Bayn Shire have type A sculpturing.
7. Bamba Khuduk (= Bambu Khuduk), Umunugovi Aimag.
Geology and age. Nemegt Formation, Maastrichtian.
Material and references. Incomplete carapace of ‘Trionyx’ gobiensis (see Systematic
palaeontology section); left half of carapace of Trionychidae indet. (IPGAS 11-17-1, paratype of
Amyda menneri; see Systematic palaeontology section); unprepared skull of Trionychidae indet.
in matrix (PIN 4064-3); numerous shell fragments of Trionychidae indet. (collection PIN 4064,
collected by JSMPE in 1976).
Remarks. The shell fragments of Trionychidae indet. from Bamba Khuduk have type A
sculpturing. All these specimens may belong to a single taxon. The skull specimen needs a
preparation.
8. Bayn Dzak (= Shabarakh Usu), Umunugovi Aimag.
Geology and age. Djadokhta Formation, Campanian.
Material and references. Costal fragment of Trionychidae indet. (Fig. 16D; ZPAL MgCh,
unnumbered, collected by PMPE in 1970).
Remarks. The fragment has type A sculpturing.

9. Bayshin Tsav, Umunugovi Aimag.
Geology and age. Upper part of the Baynshire Formation, late Turonian – Santonian.
Material and references. Partial carapace of Trionychidae indet. (Fig. 16E; PIN 3458-4,
collected by JSMPE in 1972); shell fragments of Trionychidae indet. (as Trionychidae; Suzuki &
Narmandakh 2004: 8).
Remarks. PIN 3458-4 is represented by the anterior two-thirds of a carapace, which needs
additional preparation. The length of the preserved part of the carapace is about 22 cm, and its
estimated total length was about 25 cm. The outline of the carapace is slightly rounded laterally
and weakly scalloped, similar to Gobiapalone spp. The proportions of the nuchal (about five
times wider than long) correspond to Trionychinae or Plastomenidae. The neurals are not
observable. Only parts of costals 1–6 are preserved, and their total number is not clear. The free
ribs project slightly from the costals, similar to Gobiapalone spp. The specimen has type A
sculpturing. In spite the similarities of PIN 3458-4 with Gobiapalone spp. some important
characters of this specimen are not observable and, for this reason, it is determined only as
Trionychidae indet.
10. Bugin Tsav and Bugin Tsav II, Umunugovi Aimag.
Geology and age. Barungoyot Formation, Campanian, and Nemegt Formation, Maastrichtian.
Material and references. Four partial skeletons and a partial shell of Gobiapalone breviplastra
from the Nemegt Formation of Bugin Tsav and Bugin Tsav II, and partial carapace of
Gobiapalone breviplastra from the Barungoyot Formation of Bugin Tsav (see Systematic
palaeontology section); lateral fragment of right hypoplastron of Nemegtemys conflata from the
Nemegt Formation of Bugin Tsav (see Systematic palaeontology section); partial carapace and
skull of Trionychidae indet., which need a preparation, from the Nemegt Formation of Bugin
Tsav II (HMNS 2006-04-470, collected by JMJPE in 2006); fragmentary shell remains of
Trionychidae indet. from the Nemegt Formation of Bugin Tsav II (HMNS 2006-04-471,
collected by JMJPE in 2006); nuchal of Trionychidae indet. from the Nemegt Formation of
Bugin Tsav (HMNS 2006-04-473, collected by JMJPE in 2006; as Trionychidae; Suzuki &
Narmandakh, 2004); medial part of right hyoplastron of Trionychidae indet. from the Nemegt
Formation of Bugin Tsav (IPGAS 11-13-11, paratype of Amyda menneri; see Systematic
palaeontology section); shell fragments of Trionychidae indet. from the Nemegt Formation of
Bugin Tsav (collection ZIN PH 160, collected by A.V. Sochava in 1967).
11. Burkhant, Dornogovi Aimag.

Geology and age. Baynshire Formation, Cenomanian – Santonian.
Material and references. Partial skeleton and carapace of Gobiapalone orlovi (see Systematic
palaeontology section).
12. Char Teeg (= Khar Teg), Dornogovi Aimag.
Geology and age. ?Nemegt Formation, ?Maastrichtian.
Material and references. Fragment of the carapace on a cast of the internal cavity of the shell of
Trionychidae indet. (Fig. 16F; ZPAL MgCh/71, collected by PMPE in 1963; Khosatzky and
Młynarski 1971).
Remarks. This specimen was mentioned by Khosatzky & Młynarski (1971) as “an imprint of a
large fragment of carapace together with pleural plate”. Actually, it consists of the complete left
costal 5 and the medial fragment of the left costal 6. In addition, imprints of neurals 5-7 and left
costals 6-8 are visible on a cast of the internal cavity of the shell. The estimated length of the
shell was about 40 cm. Type A sculpturing. There were seven neurals and costals 7 and 8
contacted at the midline. Neural 5 was isometric, neural 6 was hexagonal short-sided anteriorly,
and neural 7 was pentagonal. Costal 5 was longer laterally than medially. The free rib of costal 5
slightly projects from the border. Given the incompleteness of this specimen and the uncertainty
with the formation, we can identify it only as Trionychidae indet.
13. Dersnii Khuduk, Dornogovi Aimag.
Geology and age. Lower part of the Baynshire Formation, Cenomanian – early Turonian.
Material and references. Shell fragments of Trionychidae indet. (as Trionyx sp.; Shuvalov &
Chkhikvadze 1975: 217, 222, 229).
14. Dzun Shakhai, Dornogovi Aimag.
Geology and age. Early Cretaceous.
Material and references. Incomplete nuchal and costals of Trionychidae indet. (as
Trionychidae; Suzuki & Narmandakh 2004: 8, pl. 1, pl.-fig. 2; as Trionychinae indet. 7; Danilov
& Vitek, 2009: 54; Trionychinae indet. 6; Danilov & Vitek 2012: 430).
Remarks. Danilov & Vitek (2009, 2012) identified this material as Trionychinae indet. based on
the nuchal which is is at least three times wider than long. However, recently Joyce & Lyson
(2011) demonstrated that the presence of a nuchal at least four times wider than long is also
characteristic of Plastomenidae. For this reason, we reidentify the material from Dzun Shakhai
as Trionychidae indet.

15. Ekhin Tukhum, Dornogovi Aimag.
Geology and age. Lower part of the Baynshire Formation, Cenomanian – early Turonian.
Material and references. Shell fragments of Trionychidae indet. (as Trionyx sp.; Shuvalov &
Chkhikvadze 1979: 70).
16. Erdeni Ula (= Erdene Uul), Dundgov Aimag.
Geology and age. Khulsangol Svita, Aptian–Albian.
Material and references. Shell fragments of Trionychidae indet. (as trionychid; Khosatzky
1999: 141).
Remarks. We did not manage to find any trionychid specimens from Erdeni Ula in ZIN PH
collections studied by L.I. Khosatzky. It is probable, that specimens which Khosatzky identified
as trionychids are actually specimens of Nanhsiungchelyidae, which were reported from this
locality by Danilov & Syromyatnikova (2008).
17. Gilbentu, Umunugovi Aimag.
Geology and age. Nemegt Formation, Maastrichtian.
Material and references. A partial hyo- and hypoplastra of ‘Trionyx’ gilbentuensis (see
Systematic palaeontology section); shell fragments of Trionychidae indet. (Fig. 16G; collection
ZIN PH 163, collected by MPE in 1946).
Remarks. The shell fragments of Trionychidae indet. from Gilbentu are characterized by type A
sculpturing, whereas ‘Trionyx’ gilbentuensis has type B sculpturing.
18. Gurilin Tsav, Umunugovi Aimag.
Geology and age. Nemegt Formation, Maastrichtian.
Material and references. Incomplete postcranium of one individual, including nuchal, proximal
part of costal 1, fragment of right hyo- and hypoplastron, fragment of xiphiplastron, and assorted
non-shell bones of Trionychidae indet. (IPGAS 11-5-1, holotype of Amyda menneri; see
Systematic palaeontology section).
19. Ingeni Khobur and Ingeni Khobur III, Umunugovi Aimag.
Geology and age. Nemegt Formation, Maastrichtian.
Material and references. Shell fragments of Trionychidae indet. from Ingeni Khobur (IPGAS
11-14-2, IPGAS 11-14-3, and IPGAS 11-14-4, paratypes of Amyda menneri; see Systematic
palaeontology section); shell fragments of Trionychidae indet. from Ingeni Khobur (Khosatzky,

1999: 147); shell fragments of Trionychidae indet from Ingeni Khobur III (as Trionyx sp.;
Shuvalov & Chkhikvadze 1979: 66).
20. Ingeni Tsav, Bayankhongor Aimag.
Geology and age. Barungoyot Formation, Campanian, and Nemegt Formation, Maastrichtian.
Material and references. Shell fragments of Trionychidae indet. (as Trionyx sp.; Shuvalov &
Chkhikvadze 1979: 63, 64).
21. Khamdin Khural.
Geology and age. Unknown formation, Late Cretaceous.
Material and references. Shell fragments of Trionychidae indet. (Fig. 16H–J; collection ZIN
PH 162; collected by G.G. Martinson in 1971).
Remarks. The shell fragments of Trionychidae indet. belong to two forms, which demonstrate
two types of sculpturing: the first form (Fig. 16H) has type A sculptruing, whereas the second
one (Fig. 16I, J) has shell sculpturing which is a pattern of more dense net of ridges and smaller
pits. The second type of sculpturing is present on two plastral fragments. One of them (Fig. 16I),
probably, from a lateral part of hyoplastron, demonstrates high degree of ossification.
22. Khara Khutul (= Khara Khutul Ula; = Khar Hötöl Uul), Dornogovi Aimag.
Geology and age. Baynshire Formation, Cenomanian – Santonian.
Material and references. Shell fragments of Trionychidae indet. (Fig. 16K, L; collection ZIN
PH 80, collected by G.G. Martinson in 1971 and 1978, V.F. Shuvalov in 1971 and N.N. Verzilin
in 1978); one fragmentary shell remain of Trionychidae indet. (as Trionychidae; Suzuki &
Narmandakh 2004: 8).
Remarks. The shell fragments of Trionychidae indet. from Khara Khutul belong to two forms
wich demonstrate different types of shell sculpturing. The first form, represented by the medial
part of a hypoplastron and other fragments, has type A sculpturing and is most similar to
Gobiapalone orlovi. It is probable, that this material was mentioned as Amyda orlovi by Sochava
(1975: 115) and Khosatzky (1999: 142). The second form, represented by a fragment of a
hypoplastron has type B sculpturing and, most probably, was mentioned by Khosatzky (1999:
142) as “middle-sized trionychids with tuberculated relief of the plastron between axillary and
inguinal notches”.
23. Khermin Tsav (= Khermeen Tsav) and Khermin Tsav II, Umunugovi Aimag.

Geology and age. Lower White Beds, ?Baynshire Formation, Cenomanian – Santonian, and
Upper White Beds, Nemegt Formation, Maastrichtian.
Material and references. Shell fragments of Trionychidae indet. from the Nemegt Formation of
Khermin Tsav (as Trionyx sp.; Shuvalov & Chkhikvadze 1975: 219, 222, 229); MPC 25/165,
nearly complete carapace of Gobiapalone sp. from the Lower White Beds of Khermin Tsav (see
Systematic palaeontology section); shell fragments of Trionychidae indet. from the Upper White
Beds of Khermin Tsav, shell fragments of Trionychidae indet. from the Lower White Bed of
Khermin Tsav II and ten specimens of Trionychidae indet. from the Upper White Bed of
Khermin Tsav II (Fig. 16M; MPC 25/167, collected by JMJPE in 2006; as Trionychidae; Suzuki
& Narmandakh 2004: 9); shell fragments of Trionychidae indet from Khermin Tsav II
(collection ZPAL MgCh/160, collected by PMPE in 1971); shell fragments of Trionychidae
indet. from the Nemegt Formation of Khermin Tsav (Fig. 16N; collection ZIN PH 164, collected
by I. Yu. Neustroeva in 1977).
Remarks. The ten specimens of Trionychidae indet. from the Upper White Bed of Khermin
Tsav II are shell fragments with sculpturing similar to the type B. Other shell fragments from
Khermin Tsav and Khermin Tsav II that we examined have type A sculpturing. Therefore,
trionychids of Khermin Tsav are represented by one form (Gobiapalone sp.) in the ?Baynshire
Formation and probably by two forms (Trionychidae indet.) in the Nemegt Formation.
24. Khongil, Dornogovi Aimag.
Geology and age. Baynshire Formation, Cenomanian – Santonian.
Material and references. Incomplete carapace of Gobiapalone orlovi (see Systematic
palaeontology section).
Remarks. Khosatzky (1999) mentioned that Khongil is situated 2 km N from Tsagan Teg Obo
(=Tsagan Teg), which is very close to Khongil Tsav, whereas Sochava (1975, fig. 24) indicated
that Khongil was about 50 km SW from Khongil Tsav.
25. Khongil Obo, Dornogovi Aimag.
Geology and age. Upper part of the Baynshire Formation, late Turonian – Santonian.
Material and references. Trionychidae indet. (as Trionyx sp.; Shuvalov & Chkhikvadze 1979:
67).
Remarks. According to Shuvalov & Chkhikvadze (1979), Khongil Obo is situated in the
northern part of Bayshin Tsav Depression, 3 km E from the Khongil Obo Mountain.
26. Khongil Tsav (= Khangil Tsav), Dornogovi Aimag.

Geology and age. Baynshire Formation, Cenomanian – Santonian.
Material and references. Shell fragments of Trionychidae indet. (Fig. 16O; collection ZIN PH
88, collected by R. Barsbold and G.G. Martinson in 1967; as “a fragment of the nuchal of the
trionychid of the Turonian-Santonian type and a lateral part of the plastron, which can be
assigned to “Plastomenus””; Khosatzky 1999: 144); five specimens of Trionychidae indet. (as
Trionychidae; Suzuki & Narmandakh 2004: 8).
Remarks. The shell fragments of Trionychidae indet. from Khongil Tsav have type A
sculpturing.
27. Khuren Tsav, Bayankhongor Aimag.
Geology and age. Lower part of the Barungoyot Formation, Campanian.
Material and references. Trionychidae indet. (as Trionyx sp.; Shuvalov & Chkhikvadze 1975:
221, 222, 229).
28. Nemegt, Umunugovi Aimag.
Geology and age. Nemegt Formation, Maastrichtian.
Material and references. Lateral fragment of hyo- and hypoplastra of Nemegtemys conflata (see
Systematic palaeontology section); partial juvenile shell with fragments of non-shell postcrania,
posterior carapace fragment, left hypoplastron, and partial left hyo- and hypoplastra of
Gobiapalone breviplastra (see Systematic palaeontology section); incomplete carapace of
‘Trionyx’ gobiensis (see Systematic palaeontology section); shell fragments of Trionychidae
indet. (ZPAL MgCh/88 and ZPAL MgCh/89, collected by PMPE; as Trionyx sp. b; Młynarski &
Narmandach 1972: 100; collection ZIN PH 157, collected by MPE in 1948; Khosatzky 1999:
147); two specimens of Trionychidae indet. (as Trionychidae; Suzuki & Narmandakh 2004: 10).
Remarks. The shell fragments of Trionychidae indet. from Nemegt have type A sculpturing.
29. Nogon Tsav and Nogon Tsav II, Bayankhongor Aimag.
Geology and age. Upper part of the Barungoyot Formation, Campanian, and Nemegt Formation,
Maastrichtian.
Material and references. Partial carapace of Gobiapalone breviplastra from the upper part of
the Barungoyot Formation of Nogon Tsav (see Systematic palaeontology section); shell
fragments of Trionychidae indet. from the Nemegt Formation of Nogon Tsav (collection ZIN PH
165, collected by MPE, by A.V. Sochava in 1966 and 1968, and by L.Ya. Borkin in 1980 and
1981; as “bones of trionychids”; Khosatzky 1999: 147); shell fragments of Trionychidae indet.

from the Nemegt Formation of Nogon Tsav II (as Trionyx sp.; Shuvalov & Chkhikvadze 1979:
66).
Remarks. The shell fragments of Trionychidae indet. from the Nemegt Formation of Nogon
Tsav have type A sculpturing.
30. Ongon Ulan Ula, Umunugovi Aimag.
Geology and age. Lower part of the Baishire Formation, Cenomanian – early Turonian.
Material and references. Shell fragments of Trionychidae indet. (Fig. 16P; collection ZIN PH
166).
Remarks. The shell fragments of Trionychidae indet from Ongon Ulan Ula have type A
sculpturing.
31. Shiluut Ula, Umunugovi Aimag.
Geology and age. Unknown formation, Campanian (see Watabe et al., 2010).
Material and references. Carapace of ‘Trionyx’ shiluutulensis (see Systematic palaeontology
section).
32. Shine Us Khuduk (=Shine Us Khudag), Dornogovi Aimag.
Geology and age. Baynshire Formation, Cenomanian – Santonian.
Material and references. Shell fragments of Trionychidae indet. (Fig. 16Q; collection ZIN PH
81; collected by G.G. Martinson in 1968 and 1971, and by N.N. Verzilin in 1978; Khosatzky,
1999: 141; three specimens of Trionychidae indet. (as Trionychidae; Suzuki & Narmandakh
2004: 8).
Remarks. The shell fragments of Trionychidae indet from Shine Us Khuduk have type A
sculpturing.
33. Shine Usny Tolgod, Dornogovi Aimag.
Geology and age. Early Cretaceous.
Material and references. One specimen of Trionychidae indet. (as Trionychidae; Suzuki &
Narmandakh 2004: 8).
34. Shiregin Gashun, Bayankhongor Aimag.
Geology and age. Upper part of the Baynshire Formation, late Turonian – Santonian.

Material and references. Trionychidae indet. (as Trionyx sp.; Shuvalov & Chkhikvadze 1975:
220, 222, 229); shell fragments of Trionychidae indet. (Fig. 16R; collection ZIN PH 167,
collected by G.G. Martinson and N.N. Verzilin in 1978).
Remarks. The shell fragments of Trionychidae indet from Shiregin Gashun have type A
sculpturing.
35. Tel Ulan Shalcha (= Tel Ulan Chaltsai), Dornogovi Aimag.
Geology and age. Unknown formation, probably, early Late Cretaceous.
Material and references. Shell fragments of Trionychidae indet. (Fig. 16S; collection ZIN PH
168, collected by V.F. Shuvalov in 1971; as “shell fragments of rather large trionychids”;
Khosatzky 1999: 144).
Remarks. The shell fragments of Trionychidae indet. from Tel Ulan Shalcha have type A
sculpturing.
36. Tsagan Khushu (= Tsagan Ula), Umunugovi Aimag.
Geology and age. Nemegt Formation, Maastrichtian.
Material and references. Incomplete carapace and left costals 1 and 2 of ‘Trionyx’ gobiensis
(see Systematic palaeontology section); shell fragments of Trionychidae indet. (Fig. 16T;
collection ZIN PH 24, collected by MPE in 1948; as “parts of the plastron with considerably
projecting gastralia and vermiform details of the external relief”; Khosatzky 1999: 147); shell
fragments of Trionychidae indet. (ZPAL MgCh/60, collected by PMPE in 1964); four specimens
of Trionychidae indet. (as Trionychidae; Suzuki & Narmandakh 2004: 10).
Remarks. The shell fragments of Trionychidae indet from Tsagan Khushu (collection ZIN PH
24) have type A sculpturing and correspond in size to Gobiapalone spp. Among these fragments,
a partial right hyo- and hypoplastra (Fig. 16T; ZIN PH 3/24) is similar in its outline to
Gobiapalone spp. That specimen was probably mentioned by Khosatzky (1999) as “parts of the
plastron with considerably projecting gastralia and vermiform details of the external relief”.
Thus, we can recognize two forms of trionychids in Tsagan Khushu: ‘Trionyx’ gobiensis and
Trionychidae indet. (similar to Gobiapalone spp.).
37. Tsagan Teg (= Tsagan Teg Obo), Dornogovi Aimag.
Geology and age. Baynshire Formation, Cenomanian – Santonian.
Material and references. Shell fragments of Trionychidae indet. (Fig. 16U, V; collection ZIN
PH 91, collected by V.F. Shuvalov in 1971, G.G. Martinson and N.N. Verzilin in 1978; as “large

trionychid of the Turonian-Santonian type”; Khosatzky 1999: 144); one specimen of
Trionychidae indet. (as Trionychidae; Suzuki & Narmandakh 2004: 8).
Remarks. The shell fragments of Trionychidae indet. from Tsagan Teg demonstrate type A
sculpturing (Fig. 16U; ZIN PH 2/91) and B (Fig. 16V; ZIN PH 3/91). Thus, probably, there are
two forms of trionychids in this locality.
38. Tugrikin Ula (near Khamarin Khural), Dornogovi Aimag.
Geology and age. Lower part of the Baynshire Formation, Cenomanian – early Turonian.
Material and references. Shell fragments of Trionychidae indet. (Fig. 16W; collection ZIN PH
169, collected by V.F. Shuvalov in 1971; as “remains of medium-sized trionychids”; Khosatzky
1999: 144).
Remarks. The shell fragments of Trionychidae indet. from Tugrikin Ula have type A
sculpturing.
39. Ulan Khushu (= Ulan Bulak), Umunugovi Aimag.
Geology and age. Nemegt Formation, Maastrichtian.
Material and references. Partial shell, partial carapace and partial entoplastron of Gobiapalone
breviplastra (see Systematic palaeontology section); one specimen of Trionychidae indet. (as
Trionychidae; Suzuki & Narmandakh 2004: 9); shell fragments of Trionychidae indet.
(collection PIN 4694, collected by JSMPE); carapace fragment of Trionychidae indet. (ZIN PH
1/170); shell fragments of Trionychidae indet. (collection ZPAL MgCh/69, collected by PMPE
in 1963).
Remarks. The shell fragments of Trionychidae indet. from Ulan Khushu have type A
sculpturing.
40. Ulan Tsab Ula (9 km NW from Ulan Tsab Ula), Bayankhongor Aimag.
Geology and age. Probably, Baynshire Formation, Cenomanian – Santonian.
Material and references. Shell fragments of Trionychidae indet. (as “fragments of medium-
sized trionychids”; Khosatzky 1999: 144).
41. Unegetu Ula, Dornogovi Aimag.
Geology and age. Baynshire Formation, Cenomanian – Santonian.
Material and references. Anterior part of carapace of Gobiapalone orlovi (see Systematic
palaeontology section).

42. Uryl’b Usu, Dornogovi Aimag.
Geology and age. Lower part of the Baynshire Formation, Cenomanian – early Turonian.
Material and references. Trionychidae indet. (as Trionyx sp.; Shuvalov & Chkhikvadze 1979:
69).
43. Ushyin Khuduk, Dornogovi Aimag.
Geology and age. Upper part of the Baynshire Formation, late Turonian – Santonian.
Material and references. Trionychidae indet. (as Trionyx sp.; Shuvalov & Chkhikvadze 1979:
71).
44. Yagaan Khovil, Umunugovi Aimag.
Geology and age. Djadokhta Formation, Campanian.
Material and references. Six specimens of Trionychidae indet. (as Trionychidae; Suzuki &
Narmandakh 2004: 10).
45. Unknown locality, probably, north-eastern regions of Mongolia.
Geology and age. ?Late Cretaceous.
Material and references. Shell fragments of Trionychidae indet. (Fig. 16X; collection ZIN PH
97, collected by V.N. Chaykovskiy in 1936 from unknown locality, probably, in north-eastern
regions of Mongolia, where he worked as a geologist (Khosatzky 1976)).
Remarks. The probable Late Cretaceous age of the trionychid remains is confirmed by presence
of Nanhsiungchelyidae in the same collection (Danilov & Syromyatnikova 2008). The shell
fragments of Trionychidae indet. from this locality have type A sculpturing.
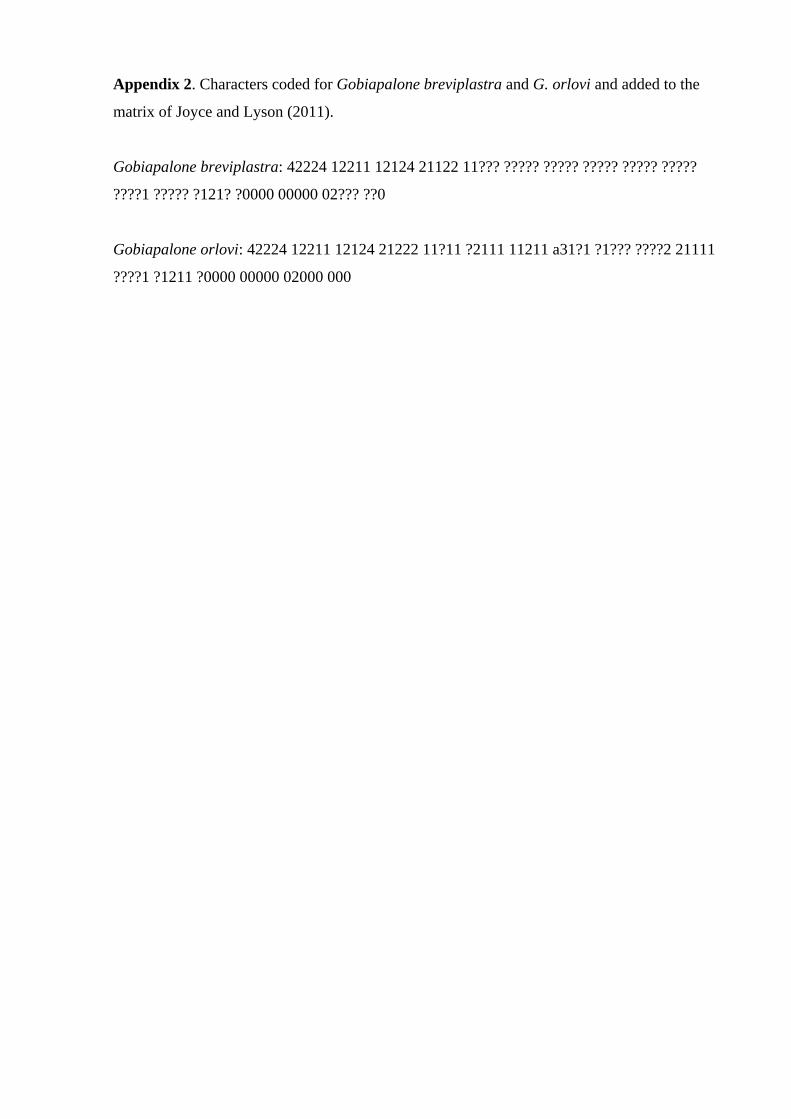
Appendix 2. Characters coded for Gobiapalone breviplastra and G. orlovi and added to the
matrix of Joyce and Lyson (2011).
Gobiapalone breviplastra: 42224 12211 12124 21122 11??? ????? ????? ????? ????? ?????
????1 ????? ?121? ?0000 00000 02??? ??0
Gobiapalone orlovi: 42224 12211 12124 21222 11?11 ?2111 11211 a31?1 ?1??? ????2 21111
????1 ?1211 ?0000 00000 02000 000

Appendix 3. Characters coded for Gobiapalone breviplastra and G. orlovi and added to the
matrix of Vitek (2012).
Gobiapalone breviplastra: 31113 01200 0a0b1 00110 0???? ????? ????? ????? ????? ????? ???0?
????? 010?? 00000 00000 2???? ?0110 000
Gobiapalone orlovi: 31113 01200 0a0b1 01110 0?00? 10000 0100a 20?0? 0???? ???11 0000?
???0? 0100? 00000 00000 20000 00110 000
Copyright © 2022 FDOKUMEN



















