
Cretaceous methane-seep deposits from New Zealand and their fauna
18
Cretaceous methane-seep deposits from New Zealand and their fauna Steffen Kiel a, ⁎, Daniel Birgel b , Kathleen A. Campbell c , James S. Crampton d , Poul Schiøler d , Jörn Peckmann b a Georg-August Universität Göttingen, Geowissenschaftliches Zentrum, Abteilung Geobiologie and Courant Research Center Geobiology, Goldschmidtstr. 3, 37077 Göttingen, Germany b Universität Wien, Erdwissenschaftliches Zentrum, Department für Geodynamik und Sedimentologie, Althanstr. 14, 1090 Wien, Austria c Geology Programme, School of Environment, University of Auckland, Private Bag 92019, Auckland 1142, New Zealand d GNS Science, Post Office Box 30368, Lower Hutt, New Zealand abstract article info Article history: Received 2 June 2012 Received in revised form 22 August 2012 Accepted 8 October 2012 Available online xxxx Keywords: Cretaceous New Zealand Deep-sea Methane seepage Biogeography Endemism Six Cretaceous methane-seep deposits are reported from the Raukumara Peninsula, eastern North Island, New Zealand. Dinoflagellate dating indicates a Late Albian to mid-Cenomanian age for three deposits from Port Awanui, and a mid-Campanian age for two deposits from Waipiro Bay and for one from Owhena Stream. The dominant petrographic fabric of the carbonates is detrital micrite, and the numerous fenestrae and vugs are filled with various cements including two types of non-detrital micrite, and botryoidal and banded fibrous calcites. These petrographic features, in combination with negative δ 13 C values of early diagenetic micrites (as low as -29.2‰ vs. V-PDB) and 13 C-depleted molecular fossils such as the archaeal biomarkers pentamethylicosane (-97‰ vs. V-PDB) and biphytane (-99‰), and the bacterial biomarker anteiso-C 15 fatty acid (-47‰), reveal that sulfate-dependent anaerobic oxidation of methane (AOM) induced the formation of these deposits. The carbonates preserve a typical mollusk dominated, Late Mesozoic, deep-water seep fauna including the large modiomorphid bivalve Caspiconcha, the lucinid bivalve Ezolucina, and limpets, hokkaidoconchids and a large abyssochrysoid among the gastropods, with close biogeographic relationships to North Pacific seep faunas of Cretaceous age. In contrast to the general marine mollusk fauna of New Zealand, which shows a high degree of endemism in both the Cenozoic and Recent, New Zealand's Cretaceous to present-day seep faunas consist largely of taxa known from coeval seeps around the world. Thus while deep-water seeps in New Zealand were repeatedly or continuously colonized by members of the global seep fauna, the general mollusk fauna of the New Zealand region developed a considerable degree of endemism during this time. © 2012 Elsevier B.V. All rights reserved. 1. Introduction Methane seeps and hydrothermal vents in the deep sea harbor rich faunal communities dominated by animals living in symbiosis with chemotrophic bacteria that use reduced chemical compounds of the emitting fluids for nutrition (Felbeck, 1981; Paull et al., 1985; Dubilier et al., 2008). Many of these animals belong to groups that are restricted (endemic) to these environments and have wide geo- graphic distributions within and between ocean basins (Sibuet and Olu, 1998; Van Dover et al., 2002; Kojima et al., 2004; Olu et al., 2010). The geologic record of vent and seep faunas ranges back into Paleozoic time (Campbell, 2006) but the early faunas were taxonom- ically very different from those of today: among the dominant taxa were large brachiopods, a group that has largely vanished from these environments today (Campbell and Bottjer, 1995a; Peckmann et al., 2007). The rise of the modern, mollusk-dominated seep fauna began in the Cretaceous (Campbell and Bottjer, 1995a) and although several lineages of extant seep taxa can be traced back into Cretaceous time (Kiel and Little, 2006; Campbell et al., 2008b; Kaim et al., 2008, 2009), most of the dominant taxa at Cretaceous seeps belong to differ- ent taxonomic groups than those of today and these taxa have been termed the ‘Late Mesozoic Seep Fauna’ (Kiel, 2010). This fauna includes the modiomorphid bivalve Caspiconcha, the abyssochrysoid gastropods Paskentana and Hokkaidoconcha (Kiel and Peckmann, 2008; Kiel, 2010; Kiel et al., 2010) and, during the Early Cretaceous, the brachiopod Peregrinella (Campbell and Bottjer, 1995b; Sandy, 1995). Here we report the first Cretaceous seep deposits from the Southern Hemisphere. They are derived from an in situ deposit and several alloch- thonous boulder occurrences recovered from Raukumara Peninsula, eastern North Island, New Zealand. Petrographic thin sections, stable carbon and oxygen isotopes, and biomarker analyses are used to charac- terize the seep deposits; palynomorphs are used to determine their geologic ages; and finally the macrofauna is identified and its ecologic and biogeographic implications are discussed. 2. Geological setting and material The three localities described herein are (1) “Owhena stream” (informal name), a tributary of Waitahaia River in the central part of Raukumara Peninsula, and (2) Port Awanui and (3) Waipiro Bay on the east coast of the peninsula (Fig. 1). Palaeogeography, Palaeoclimatology, Palaeoecology xxx (2012) xxx–xxx ⁎ Corresponding author. Tel.: +49 551 3910954; fax: +49 551 397918. E-mail address: [email protected] (S. Kiel). PALAEO-06313; No of Pages 18 0031-0182/$ – see front matter © 2012 Elsevier B.V. All rights reserved. http://dx.doi.org/10.1016/j.palaeo.2012.10.033 Contents lists available at SciVerse ScienceDirect Palaeogeography, Palaeoclimatology, Palaeoecology journal homepage: www.elsevier.com/locate/palaeo Please cite this article as: Kiel, S., et al., Cretaceous methane-seep deposits from New Zealand and their fauna, Palaeogeography, Palaeoclimatology, Palaeoecology (2012), http://dx.doi.org/10.1016/j.palaeo.2012.10.033
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Cretaceous methane-seep deposits from New Zealand and their fauna

Palaeogeography, Palaeoclimatology, Palaeoecology xxx (2012) xxx–xxx
PALAEO-06313; No of Pages 18
Contents lists available at SciVerse ScienceDirect
Palaeogeography, Palaeoclimatology, Palaeoecology
j ourna l homepage: www.e lsev ie r .com/ locate /pa laeo
Cretaceous methane-seep deposits from New Zealand and their fauna
Steffen Kiel a,⁎, Daniel Birgel b, Kathleen A. Campbell c, James S. Crampton d, Poul Schiøler d, Jörn Peckmann b
a Georg-August Universität Göttingen, Geowissenschaftliches Zentrum, Abteilung Geobiologie and Courant Research Center Geobiology, Goldschmidtstr. 3, 37077 Göttingen, Germanyb Universität Wien, Erdwissenschaftliches Zentrum, Department für Geodynamik und Sedimentologie, Althanstr. 14, 1090 Wien, Austriac Geology Programme, School of Environment, University of Auckland, Private Bag 92019, Auckland 1142, New Zealandd GNS Science, Post Office Box 30368, Lower Hutt, New Zealand
⁎ Corresponding author. Tel.: +49 551 3910954; fax:E-mail address: [email protected] (S. Kiel).
0031-0182/$ – see front matter © 2012 Elsevier B.V. Allhttp://dx.doi.org/10.1016/j.palaeo.2012.10.033
Please cite this article as: Kiel, S., et alPalaeoclimatology, Palaeoecology (2012), ht
a b s t r a c t
a r t i c l e i n f oArticle history:Received 2 June 2012Received in revised form 22 August 2012Accepted 8 October 2012Available online xxxx
Keywords:CretaceousNew ZealandDeep-seaMethane seepageBiogeographyEndemism
Six Cretaceous methane-seep deposits are reported from the Raukumara Peninsula, eastern North Island, NewZealand. Dinoflagellate dating indicates a Late Albian to mid-Cenomanian age for three deposits from PortAwanui, and a mid-Campanian age for two deposits from Waipiro Bay and for one from Owhena Stream. Thedominant petrographic fabric of the carbonates is detritalmicrite, and the numerous fenestrae and vugs are filledwith various cements including two types of non-detrital micrite, and botryoidal and banded fibrous calcites.These petrographic features, in combination with negative δ13C values of early diagenetic micrites (as low as−29.2‰ vs. V-PDB) and 13C-depleted molecular fossils such as the archaeal biomarkers pentamethylicosane(−97‰ vs. V-PDB) and biphytane (−99‰), and the bacterial biomarker anteiso-C15 fatty acid (−47‰), revealthat sulfate-dependent anaerobic oxidation of methane (AOM) induced the formation of these deposits. Thecarbonates preserve a typical mollusk dominated, Late Mesozoic, deep-water seep fauna including the largemodiomorphid bivalve Caspiconcha, the lucinid bivalve Ezolucina, and limpets, hokkaidoconchids and a largeabyssochrysoid among the gastropods, with close biogeographic relationships to North Pacific seep faunas ofCretaceous age. In contrast to the general marine mollusk fauna of New Zealand, which shows a high degreeof endemism in both the Cenozoic and Recent, New Zealand's Cretaceous to present-day seep faunas consistlargely of taxa known from coeval seeps around the world. Thus while deep-water seeps in New Zealand wererepeatedly or continuously colonized by members of the global seep fauna, the general mollusk fauna of theNew Zealand region developed a considerable degree of endemism during this time.
© 2012 Elsevier B.V. All rights reserved.
1. Introduction
Methane seeps and hydrothermal vents in the deep sea harborrich faunal communities dominated by animals living in symbiosiswith chemotrophic bacteria that use reduced chemical compoundsof the emitting fluids for nutrition (Felbeck, 1981; Paull et al., 1985;Dubilier et al., 2008). Many of these animals belong to groups thatare restricted (endemic) to these environments and have wide geo-graphic distributions within and between ocean basins (Sibuet andOlu, 1998; Van Dover et al., 2002; Kojima et al., 2004; Olu et al.,2010). The geologic record of vent and seep faunas ranges back intoPaleozoic time (Campbell, 2006) but the early faunas were taxonom-ically very different from those of today: among the dominant taxawere large brachiopods, a group that has largely vanished fromthese environments today (Campbell and Bottjer, 1995a; Peckmannet al., 2007). The rise of the modern, mollusk-dominated seep faunabegan in the Cretaceous (Campbell and Bottjer, 1995a) and althoughseveral lineages of extant seep taxa can be traced back into Cretaceoustime (Kiel and Little, 2006; Campbell et al., 2008b; Kaim et al., 2008,
+49 551 397918.
rights reserved.
., Cretaceous methane-seeptp://dx.doi.org/10.1016/j.pala
2009), most of the dominant taxa at Cretaceous seeps belong to differ-ent taxonomic groups than those of today and these taxa have beentermed the ‘Late Mesozoic Seep Fauna’ (Kiel, 2010). This fauna includesthe modiomorphid bivalve Caspiconcha, the abyssochrysoid gastropodsPaskentana and Hokkaidoconcha (Kiel and Peckmann, 2008; Kiel, 2010;Kiel et al., 2010) and, during the Early Cretaceous, the brachiopodPeregrinella (Campbell and Bottjer, 1995b; Sandy, 1995).
Here we report the first Cretaceous seep deposits from the SouthernHemisphere. They are derived froman in situ deposit and several alloch-thonous boulder occurrences recovered from Raukumara Peninsula,eastern North Island, New Zealand. Petrographic thin sections, stablecarbon and oxygen isotopes, and biomarker analyses are used to charac-terize the seep deposits; palynomorphs are used to determine theirgeologic ages; and finally the macrofauna is identified and its ecologicand biogeographic implications are discussed.
2. Geological setting and material
The three localities described herein are (1) “Owhena stream”
(informal name), a tributary of Waitahaia River in the central part ofRaukumara Peninsula, and (2) Port Awanui and (3) Waipiro Bay onthe east coast of the peninsula (Fig. 1).
deposits from New Zealand and their fauna, Palaeogeography,eo.2012.10.033
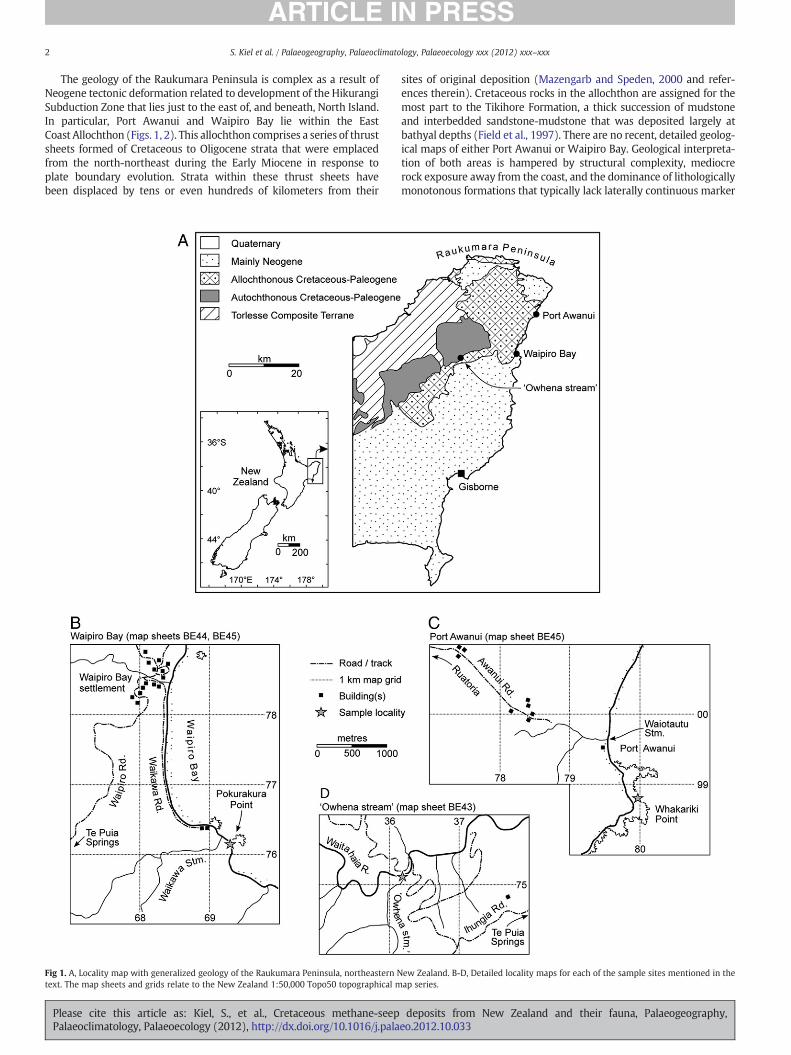
2 S. Kiel et al. / Palaeogeography, Palaeoclimatology, Palaeoecology xxx (2012) xxx–xxx
The geology of the Raukumara Peninsula is complex as a result ofNeogene tectonic deformation related to development of the HikurangiSubduction Zone that lies just to the east of, and beneath, North Island.In particular, Port Awanui and Waipiro Bay lie within the EastCoast Allochthon (Figs. 1, 2). This allochthon comprises a series of thrustsheets formed of Cretaceous to Oligocene strata that were emplacedfrom the north-northeast during the Early Miocene in response toplate boundary evolution. Strata within these thrust sheets havebeen displaced by tens or even hundreds of kilometers from their
Fig 1. A, Locality map with generalized geology of the Raukumara Peninsula, northeastern Ntext. The map sheets and grids relate to the New Zealand 1:50,000 Topo50 topographical m
Please cite this article as: Kiel, S., et al., Cretaceous methane-seepPalaeoclimatology, Palaeoecology (2012), http://dx.doi.org/10.1016/j.pala
sites of original deposition (Mazengarb and Speden, 2000 and refer-ences therein). Cretaceous rocks in the allochthon are assigned for themost part to the Tikihore Formation, a thick succession of mudstoneand interbedded sandstone-mudstone that was deposited largely atbathyal depths (Field et al., 1997). There are no recent, detailed geolog-ical maps of either Port Awanui or Waipiro Bay. Geological interpreta-tion of both areas is hampered by structural complexity, mediocrerock exposure away from the coast, and the dominance of lithologicallymonotonous formations that typically lack laterally continuous marker
ew Zealand. B-D, Detailed locality maps for each of the sample sites mentioned in theap series.
deposits from New Zealand and their fauna, Palaeogeography,eo.2012.10.033

Fig. 2. Summary of stratigraphic correlations and relationships of the seep deposits studied here, showing also the biostratigraphic ranges of key dinoflagellate species used todetermine the ages of deposits. Correlations of the New Zealand time scale to the international time scale follow Hollis et al. (2010).
3S. Kiel et al. / Palaeogeography, Palaeoclimatology, Palaeoecology xxx (2012) xxx–xxx
beds. Geological mapping has typically been reliant, at least in part, onmicropaleontologial and palynological age determinations.
In contrast, strata exposed at Owhena stream are autochthonousand have been mapped in some detail (Phillips, 1985; Mazengarbet al., 1991; Schiøler et al., 2000); biostratigraphical analyses of thesestrata based on dinoflagellates also have been published (Schiøleret al., 2000; Crampton et al., 2006b). Autochthonous late Early to LateCretaceous strata on Raukumara Peninsula are exposed generallysouth and west of the East Coast Allochthon (Figs. 1, 2). They weredeposited during the earliest phases of thermal subsidence on the pas-sive paleo-Pacific margin of New Zealand, following separation of theNew Zealand crustal block from Gondwanaland (Crampton et al.,2006b, and references therein). The relevant part of the succession atOwhena stream comprises the Owhena and overlying Whangai forma-tions, and apparently records deposition in increasing water depths,from inner shelf to outer shelf or bathyal, during transgression(Schiøler et al., 2000; Crampton et al., 2006b).
2.1. Owhena stream
The Owhena stream section was logged and sampled in detail in1999 by two of us (JSC, PS). The samples described herein derive froma conspicuous concretionary mudstone exposed within the river bedat the mouth of Owhena stream (Fig. 1). This outcrop was revisited in2010 but found to be largely covered by river gravel, a situation that islikely to change periodically depending on river flow and movementof gravel during floods. In 1999, approximately 2 m of stratigraphicthickness of knobbly, concretionary, laminated, green-black, sandymudstone was exposed. Moore (1988) placed a gradational contactbetween the Owhena and overlying Whangai formations at the levelof the concretionary mudstone. New age constraints suggest that this
Please cite this article as: Kiel, S., et al., Cretaceous methane-seepPalaeoclimatology, Palaeoecology (2012), http://dx.doi.org/10.1016/j.pala
contact represents a time break and should be placed immediatelybelow the sample horizon or somewhere in the obscured, ~10 m thickstratigraphic interval beneath (see below). Although strata immediate-ly above the sampled outcrop also are obscured, distinctive, tube- andpipe-shaped concretions extend for at least 15 m or more up into thebase of the Whangai Formation. The Owhena stream locality has notyielded macrofossils, but shows clear petrographic and isotopic evi-dence for methane seepage at the sea floor (see below), contains a mi-crobial thrombolite, and is included here because it is the only known,in situ Cretaceous seep locality in New Zealand.
2.2. Port Awanui
The fossil faunas described here from Port Awanui that were col-lected by us (SK, JSC) were derived from two large (~1 m diameter)boulders on the shore platform close to the headland of WhakarikiPoint (Fig. 1). These boulders are here termed Awanui I and II, andhave New Zealand Fossil Record File (FRF) locality numbers Z15/f83and Z15/f82, respectively. A third fauna, collected by AlexanderMcKay in 1887, was described as coming from “Secondary beds,Awanui …” and is most likely to have been taken from a boulder onthe same beach. The significance of this find was noted by McKayhimself, who commented that “a remarkable shell, probably allied toUnio [Caspiconcha sp. herein], is found in these beds…” (McKay, 1887:214–215); the fauna was recorded also by Ongley and Macpherson(1928: 32–3). This collection is called Awanui GS 688 herein, based onits old New Zealand Geological Survey collection number; the FRF local-ity number is Z15/f7485.
The boulders fromPort Awanui comprise hard,mediumgrey, calcar-eous mudstone and almost certainly have been eroded from a well-exposed, calcareous mudstone unit that forms the shore platform on
deposits from New Zealand and their fauna, Palaeogeography,eo.2012.10.033

4 S. Kiel et al. / Palaeogeography, Palaeoclimatology, Palaeoecology xxx (2012) xxx–xxx
the point. This unit was included by Mazengarb and Speden (2000)within the undifferentiated Whangai–Wanstead formations, and hadbeen examined previously (Pick, 1962). An unpublished stratigraphiclog for this locality by P. R. Moore and H. E. G. Morgans (1983, pers.comm.) indicates that the unit is at least 50 m thick, contains pebblymudstone, minor conglomerate, numerous large concretionary clasts,and has been affected by ubiquitous, syn-sedimentary slumping. Our
Fig. 3. Selected Late Jurassic to mid-Campanian dinoflagellates from the six seep localities oscale bar in N. Specimen name is followed by GNS fossil record file number, palynoloA, Senoniasphaera edenensis, Z16/f83, L25837/1, J18/0. B, Diconodinium cristatum, Z15/f83,ingramii, Z15/f83, L25836/1, B46/0. E, Laciniadinium tenuistriatum, Z15/f7485, L26065/2, E4L25836/1, F28/4. H, Vozzhennikovia spinulosa, Z16/f83, L25837/1, B34/4. I, Heslertonia striaK, Leberidocysta chlamydata, Z15/f82, L25835/1, G47/0. L, Satyrodinium bengalense, Z16/f83,turneri, Z15/f82, L25835/1, D33/4.
Please cite this article as: Kiel, S., et al., Cretaceous methane-seepPalaeoclimatology, Palaeoecology (2012), http://dx.doi.org/10.1016/j.pala
observations confirm their interpretation that the unit is internallychaotic and is likely to represent some formof debrite deposit. Micropa-leontological age determinations from thematrix of the unit indicate anearly Ypresian age (earliest Eocene,Waipawan local stage, unpubl. data,H.E.G. Morgans, 1989); whereas, palynological ages indicate thatthe sampled large concretionary clasts are reworked Early Cretaceousboulders (see below). The debrite unit is overlain conformably by
n the Raukumara Peninsula, New Zealand. All specimens are shown to the same scale;gical laboratory preparation number/slide number and England Finder coordinates.L25836/1, L15/4. C, Yalkalpodinium scutum, Z15/f83, L25836/1, P32/1. D, Eurydinium
2/4. F, Sepispinula ambigua, Z15/f83, L25836/1, D20/0. G, Balcattia cirribarbata, Z15/f83,ta, Z15/f82, L25835/1, H23/1. J, Prolixosphaeridium conulum, Z15/f83, L25836/1, B37/4.L25837/1, C48/2. M, Wallodinium glaessneri, Z15/f83, L25836/1, J14/4. N, Endoceratium
deposits from New Zealand and their fauna, Palaeogeography,eo.2012.10.033

5S. Kiel et al. / Palaeogeography, Palaeoclimatology, Palaeoecology xxx (2012) xxx–xxx
well-bedded (though somewhat deformed), Early to Middle Eocenesandstone, glauconitic sandstone, mudstone, and minor conglomerateof the Wanstead Formation.
2.3. Waipiro Bay
The fossil faunas from Waipiro Bay described here were collectedby us (SK, JSC) from loose boulders on the beach of Pokurakura Point,immediately south of Waikawa Stream (Fig. 1). They are namedWaipiro I and III herein, with FRF locality numbers Z16/f83 and Z16/f84, respectively. As mapped by Mazengarb and Speden (2000), thisbeach lies adjacent to outcrops of undifferentiated mélange that wasformed in association with emplacement of the East Coast Allochthon.We did not observe any outcrop on the beach and can add nothing tothe published interpretation of Mazengarb and Speden (2000), butinfer here that the boulders are sourced from themélange. The sampledboulders comprise hard, medium grey, calcareous mudstone. Palyno-logical determinations for the two boulders indicate Late Cretaceousages (see below).
2.4. Provenance of the fossiliferous boulders at Port Awanui andWaipiro Bay
The fossiliferous boulders sampled at Port Awanui and WaipiroBay have been reworked from mid- to Late Cretaceous strata. It isnot possible to identify provenance with any confidence: similar, con-cretionary and age-correlative rocks occur within both the East CoastAllochthon and within autochthonous formations further to the west(e.g., Tikihore and Karekare formations, respectively; see Field et al.,1997). Although many of the Cretaceous units on Raukumara Penin-sula are richly fossiliferous, similar seep faunas have not been identi-fied in situ. It is possible that the boulders are derived from areas ofthe allochthon lying to the northeast and offshore, or from autochtho-nous rocks that are buried beneath the allochthon or buried beneathextensive, post-Early Miocene strata.
3. Methods
3.1. Petrography and stable isotope analysis
Thin sections of 30 or 60 μm thickness were examined by planepolarized, cross polarized and reflected light microscopy.
Samples for stable carbon and oxygen isotope analysis wereextracted from rock chips from which thin sections were preparedusing a hand-held microdrill. Carbonate powders were reacted with100% phosphoric acid at 75 °C using a Kiel IV online carbonate prepara-tion line connected to a Finnigan DeltaPlus triple collector mass spec-trometer. All values are reported in per mil relative to Vienna Peedeebelemnite (V-PDB) standard by assigning a δ13C value of +1.95‰ anda δ18O value of −2.20‰ to NBS19. Reproducibility was checked byreplicate analysis of laboratory standards and is better than ±0.05‰for δ13C and ±0.06‰ for δ18O values.
3.2. Palynology
Five matrix samples from the collected carbonate rocks wereprocessed for palynological analysis with the aim of dating the seepfaunas. The samples were processed following standard palynologicaltechniques for pre-Quaternary samples (cf. Batten, 1999), includingtreatment with 5 N hydrochloric acid, 40% hydrofluoric acid andtwo minutes oxidation in 36% nitric acid followed by heavy liquidseparation using sodium polytungstate. Treatment with diluted alkaliafter the oxidation and ultrasonic treatment was not used. The sampleresiduewas filtered on 6 and 15 μm filter cloths andmounted in glycer-ine jelly. The samples were studied qualitatively for dinoflagellates andacritarchs. Dinoflagellate taxamentioned in the text are fully referencedin the Lentin and Williams index (Fensome et al., 2008).
Please cite this article as: Kiel, S., et al., Cretaceous methane-seepPalaeoclimatology, Palaeoecology (2012), http://dx.doi.org/10.1016/j.pala
3.3. Biomarkers
Limestone samples from Awanui II and Waipiro III were preparedand decalcified after a method described previously (Birgel et al.,2006). After a saponification procedure with 6% KOH in methanol, thesamples were extracted with a microwave extraction system (CEMDiscovery) at 80 °C and 250 W with a dichloromethane–methanol(3:1) mixture. The total extracts were pre-cleaned by a separationinto an n-hexane soluble and a dichloromethane-soluble fraction. Then-hexane fraction was further separated by column chromatographyinto four fractions of increasing polarity (cf. Birgel et al., 2008a). Onlythe hydrocarbons and carboxylic acid fractions were found to containcompounds that help identify the process of carbonate precipitation.The alcohol fractions were found to be strongly affected by thermalmaturation and biodegradation and did not yield genuine lipidbiomarker signatures. Both the hydrocarbon and the carboxylic acidfractions were analyzed by coupled gas chromatography–mass spec-trometry (GC-MS) with an Agilent 7890 A GC system coupled to anAgilent 5975 C inert MSD mass spectrometer at the Department forGeodynamics and Sedimentology, University of Vienna. Compoundswere quantified by gas chromatography-flame ionization detection(GC-FID) with an Agilent 7820 A GC system. Both GC-MS and GC-FIDwere equipped with a 30 m HP-5 MS UI fused silica capillary column(0.25 mm i.d., 0.25 μm film thickness). The carrier gas was helium.The GC temperature program used for both fractions was as follows:60 °C (1 min); from 60 °C to 150 °C at 10 °C/min; from 150 °C to320 °C at 4 °C/min, 25 min isothermal. Identification of compoundswas based on retention times and published mass spectral data.Compound-specific carbon isotope analysis was performed with aThermo Fisher Trace GC Ultra connected via a Thermo Fisher GC Isolinkinterface to a Thermo Fisher Delta V Advantage spectrometer at theDepartment of Terrestrial Ecosystem Research, University of Vienna.Conditions chosen for gas chromatography were identical to thosedescribed above. Carbon isotopes are given as δ values in per mil rela-tive to the V-PDB standard. Each measurement was calibrated usingseveral pulses of carbon dioxide with known isotopic composition atthe beginning and end of the run. Instrument precision was checkedwith amixture of n-alkanes (C14 to C38) of known isotopic composition.Analytical standard deviation was below 0.5‰.
3.4. Degree of endemism
The degree of endemism of the macrofauna at each seep depositwas calculated for paleoecologic analyses. Only the mollusks are con-sidered in this context and the criteria of endemic vs. non-endemictaxa are those outlined by Kiel (2010). In brief, the following taxaare considered as endemic in these Cretaceous seep deposits: thelarge bivalve Caspiconcha, the lucinid bivalve Nipponothracia (butnot the other lucinid genera), and all small limpets and skeneiformgastropods. All other taxa recognized in this study (Table 1) areconsidered non-endemic. The degree of endemism is the percentageof endemic mollusks species of the total number of mollusk speciesat each seep deposit.
4. Results
4.1. Age of the deposits
The six carbonate deposits contain rich, well-preserved palyno-morph assemblages dominated by dinoflagellates. The presence of anumber of key dinoflagellate taxa can be used to date the samplesusing published biostratigraphic ranges from New Zealand, Australiaand Antarctica (Fig. 2). The dinoflagellate assemblages from two of thesamples indicate mixing of two or more ages and, therefore, reworkingof dinoflagellate floras. Many of the key taxa are shown in Fig. 3. Theseep carbonates found at Owhena stream are of mid-Campanian age,
deposits from New Zealand and their fauna, Palaeogeography,eo.2012.10.033

Fig. 4. Ammonites from Cretaceous seep deposits on the Raukumara Peninsula, New Zealand. A, Scaphites equalis coverhamensis Henderson 1973 from Awanui II (CE2925). B,smooth, undetermined species from Waipiro III (CE2926). C, Kossmaticeratidae gen. et sp. indet. from Waipiro III (CE2927). D, E, Neograhamites cf. transitorius Henderson 1970from Waipiro III (CE2928).
6 S. Kiel et al. / Palaeogeography, Palaeoclimatology, Palaeoecology xxx (2012) xxx–xxx
those from Port Awanui are of Late Albian to mid-Cenomanian age, andthose from Waipiro Bay are of mid-Campanian age. The ammonitesfound at Awanui II and Waipiro III (Fig. 4) support these conclusions(cf., Henderson, 1970, 1973).
Table 1Fossil fauna of Cretaceous seep deposits on Raukumara Peninsula, North Island, New Zealan
Taxon/Seep site Awanui I Awanui II GS 688
Bivalvessolemyid xNucinella sp. ? x
nuculanid x xthyasirid x xinoceramids x x xAucellina aff. euglypha xcarditiid xCaspiconcha sp.* x x
venerid xEzolucina sp. “Awanui” x x
Ezolucina? sp. “ridge” xEzolucina sp. “Waipiro”
Ezolucina? sp. “tall” xNipponothracia? sp.* ? x
GastropodsSerradonta sp.*
Retiskenea? sp.* x
Hokkaidoconcha sp. 1* x
Hokkaidoconcha sp. 2* xfusiform gastropod xindet gastropods x“low spired”* x“small globular”* xaporrhaid xabyssochryssoid*
naticid?
CrustaceaCallianassa spp. xsolitary coral x
Fishindet. scales xArgentinidae (otolith) xendemic mollusks 1 of 9 4 of 10 4 of 10
Please cite this article as: Kiel, S., et al., Cretaceous methane-seepPalaeoclimatology, Palaeoecology (2012), http://dx.doi.org/10.1016/j.pala
4.1.1. Awanui I (Z15/f83)This block contains Prolixosphaeridium conulum, Balcattia cirribarbata
and Wrewittia helicoidea, together indicating a broad Late Albian tomid-Cenomanian age (Morgan, 1980; Backhouse, 2006). The presence
d. Asterisks indicate taxa considered as endemic to seeps.
Waipiro I Waipiro III related taxa
xN. gigantea (Late Cretaceous, Hokkaido),Amano et al. (2007) and Kiel et al. (2008a)
xx
Caspiconcha sp. (Campanian, Hokkaido)and C. major (Tithonian-Albian,California), Jenkins et al. (in press)
small E. inflata (Cenomanian, Hokkaido),Amano et al. (2008)
x E. inflata (Cenomanian, Hokkaido) andE. colusaensis (Tithonian-Hauterivian,California), Amano et al. (2008)
N. elassodyseides (Eocene, Washington),Saul et al. (1996)
x S. omagariensis (Campanian, Hokkaido),Kaim et al. (2008)R.? kieli (Aptian-Albian, California),Campbell et al. (2008b)Hokkaidoconcha sp. (Albian, Spain),Agirrezabala et al. (this volume)
?
x Early Cretaceous A. giganteum (California)and A.? eucosmeta (Poland), Kiel et al.(2008b, 2010)
x
x
0 of 1 3 of 7
deposits from New Zealand and their fauna, Palaeogeography,eo.2012.10.033

7S. Kiel et al. / Palaeogeography, Palaeoclimatology, Palaeoecology xxx (2012) xxx–xxx
of Diconodinium cristatum and Yalkopodinium scutum may furtherrestrict the age of the sample to Late Albian (Morgan, 1980; Partridge,2006). The presence of Wallodinium glaessneri, not previously reportedfrom New Zealand, may indicate reworking of Jurassic sediments asthis species has not been recorded from strata younger than Late Jurassicin Australia (R. Helby, pers. comm., 2011).
4.1.2. Awanui II (Z15/f82)This block contains Endoceratium turneri and Endoceratium
ludbrookiae, both having range tops in the mid-Cenomanian, andthe latter having range base in Late Albian (Wilson, 1984; Helby et al.,1987; Riding and Crame, 2002). The two taxa coexist in the samplewith Heslertonia striata and W. helicoidea, which do not rangeabove the Mid-Cenomanian (Morgan, 1980). A Late Albian to mid-Cenomanian age range may be assigned to the sample.
4.1.3. Awanui GS 688 (Z15/f7485)In the material collected by Alexander McKay (1887) at Port
Awanui, the joint presence of E. turneri and H. striata indicates abroad Late Aptian to mid-Cenomanian age range. The presence ofIsabelidinium acuminatum and Eurydinium ingramii, both recordedwith range base in the mid- to Late Cenomanian (cf. Cookson andEisenack, 1970; Morgan, 1980), further refines the age of the sampleto mid-Cenomanian. It is not mentioned in McKay's (1887) reportwhether all specimens with the collection number GS 688 werecollected from a single block. However, we assume this to be thecase because such seep limestone blocks are extremely rare at PortAwanui: we surveyed the area extensively for two days and onlyfound the two blocks reported herein. Furthermore, the limestonematrix adhering to most fossils from GS 688 is nearly identical onvisual inspection and these fossils are therefore assumed to be derivedfrom a single deposit. Only a few lucinid bivalve specimens have slightlyless indurated and sandier sediment adhering to them, and these werenot considered further. The dinoflagellates reported here are fromcarbonate matrix adhering to a Caspiconcha bivalve, the characteristicmacrofaunal taxon of this locality.
4.1.4. Waipiro I (Z16/f83)This block contains Satyrodinium haumuriense, Satyrodinium
bengalense, Vozzhennikovia spinulosa and Senoniasphaera edenensisthat together indicate a mid-Campanian age for the sample (Roncagliaet al., 1999; Cooper, 2004).
4.1.5. Waipiro III (Z16/f84)This boulder contains an age diagnostic assemblage similar to that of
Waipiro I and can be assigned a mid-Campanian age. Older, pre-Campanian (probably Aptian–Albian) reworking is indicated by thejoint presence of poorly preserved specimens of Hapsocysta peridictya,Circulodinium attadalicum and Sepispinula ambigua (cf. Helby et al.,1987).
4.1.6. Owhena stream (Y16/f945)The age of this sample has been previously reported by Schiøler et al.
(2000) as middle Early Haumurian in the New Zealand chrono-stratigraphic scheme, equivalent to amid-Campanian age in the interna-tional timescale. This was based on the joint occurrence of Satyrodiniumbengalense, Vozzhennikovia spinulosa and an undescribed species ofTrichodinium. A mid-Campanian age of the sample is further supportedby the presence of Satyrodinium haumuriense and Senoniasphaeraedenensis (Roncaglia et al., 1999; Cooper, 2004). Based on a quantitativebiostratigraphic analysis, Crampton et al. (2006a, fig. 6 and DR5)inferred that the contact between the Owhena andWhangai formationsat Owhena stream corresponds to a condensed horizon or depositionalhiatus representing part of the early to middle Campanian. Althoughthe exact placement of this hiatus is not constrained by samples, allavailable evidence suggests that it lies immediately below the level of
Please cite this article as: Kiel, S., et al., Cretaceous methane-seepPalaeoclimatology, Palaeoecology (2012), http://dx.doi.org/10.1016/j.pala
the sample studied here or within the ~10 m of unexposed strata lyingbeneath.
4.2. Petrography and stable isotopes
Representative photomicrographs of petrographic fabrics from stud-ied samples are shown in Figs. 5–9. As withmost fossil seep carbonates,the New Zealand Cretaceous boulders comprise dominantly microcrys-talline calcite (micrite) with volumetrically minor but ubiquitous pyriteframboids scattered throughout. Three types of micrites are preserved,with the detrital rich type (m1, containing angular, silt- to sand-sizedquartz grains, shell fragments and some articulated bivalves, ±peloids)being by far the most dominant volumetrically. Two detrital poor typesalso are present: m2 an authigenic micrite displaying various microbialtextures including clotted, spheroidal, and laminated coated grains, andm3, a late diagenetic, homogeneousmicrite. Unusual amongmodern orfossil seep carbonates from elsewhere in the world, these New ZealandCretaceous deposits contain commonmicritic spheroidal textures, bothsmall (approximately 0.5 mm diameter) and internally structureless(e.g. Fig. 7), and large and concentrically laminated (approximately5 mm diameter, Waipiro I, but also showing a few that measure100 μm to slightly more than 1 mm in diameter) (e.g. Figs. 7B, C, 8B,D, 9B–D). The micritic spheroids, large and small, are inferred to be ofmicrobial origin. In the studied carbonates, H2S corrosion is evidentfrom the common and irregularly etched surfaces in m1, some ofwhich are pyrite coated, and from the occurrence of numerous fenes-trae and vugs. These are filled with botryoidal and banded fibrouscalcites (bbc), equant calcite spar and/or m3. Minor scalenohedralcalcite (sCc) lins vugs in some Awanui I samples (Fig. 5C). Somesamples, particularly bbc fabrics, show partial silica replacement(s) ofpreviously formed carbonate cements (Figs. 5D, 7A), or late diageneticfill of remaining vug porosity by silica (Fig. 8A). Clear equant calcite(ec) fills veins cross-cutting many samples.
The composite paragenetic sequence combined for all four sites is asfollows: m1→corrosion→m2→bbc→s→sCc→ec/m3→m3/s. Notall sites contain all textures. For example scalenohedral, or dogtoothcalcite (sCs) was only found at Awanui I, and silica replacement/fillfabrics are preserved at Awanui I, Waipiro I and Waipiro III sites. Thisparagenetic sequence is interpreted to represent a transition fromearly diagenetic (seafloor cementation, silica replacement) to latediagenetic (burial cements, late silica vug fill) events associated withmethane seepage (see Section 5.1).
An isotopic cross-plot of inorganic carbonate carbon and oxygenvalues (δ13C and δ18O, ‰ V-PDB) from cement phases and shells ofthe New Zealand Cretaceous seep carbonates evaluated in this study(Fig. 10) reveals several site- and cement-specific patterns. Awanui Iand Waipiro I cement phases (all comprising what were originallyearly diagenetic m1 and m2 micrites) record the lowest carbonate car-bon values of all samples, to−29.2‰ V-PDB, and occur in a distinctiveisotopic grouping (depleted in 13C, relatively enriched in 18O). Awanui Isandy micrites, in particular, exuded a strong petroliferous odor whenpowdered with the micro-drill. Awanui II and Waipiro III samples arethe least 13C depleted of all sites, showing a wide isotopic range, withsome micrites indicating near seawater values and another samplefrom Awanui II showing the most 13C enriched signature (δ13C of+7.1‰ V-PDB). Owhena samples yielded intermediate δ13C values.Both late diagenetic (burial) equant spar samples from Waipiro III andOwhena are depleted in 13C compared to earlier cement phases fromthe same sites. The bivalve shells indicate precipitation in isotopic equi-librium with seawater bicarbonate. With respect to oxygen isotopicsignatures, most samples constitute early micrites (m1 and m2 fabrics)and suggest relatively minor diagenetic modification. Both early andlate cements from Waipiro III and Owhena carbonates are the mostdiagenetically modified, with late diagenetic equant calcite sparsamples showing the greatest depletion in 18O (to −14.7‰ V-PDB). A
deposits from New Zealand and their fauna, Palaeogeography,eo.2012.10.033
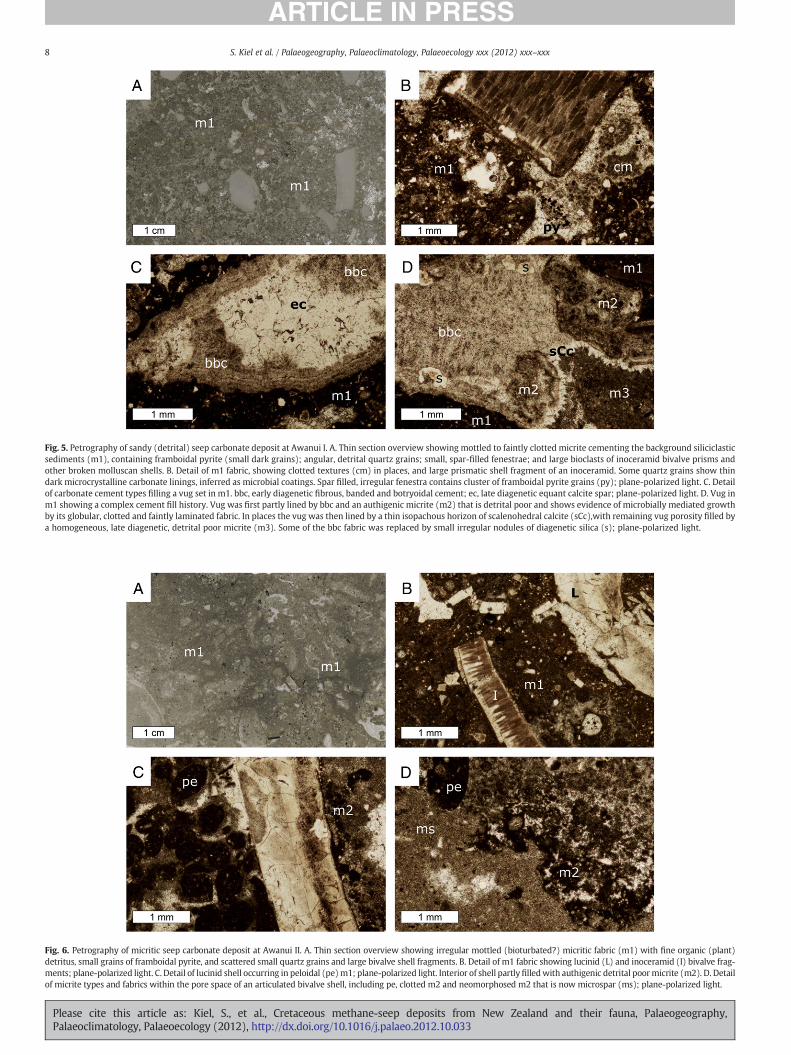
Fig. 6. Petrography of micritic seep carbonate deposit at Awanui II. A. Thin section overview showing irregular mottled (bioturbated?) micritic fabric (m1) with fine organic (plant)detritus, small grains of framboidal pyrite, and scattered small quartz grains and large bivalve shell fragments. B. Detail of m1 fabric showing lucinid (L) and inoceramid (I) bivalve frag-ments; plane-polarized light. C. Detail of lucinid shell occurring in peloidal (pe)m1; plane-polarized light. Interior of shell partlyfilledwith authigenic detrital poormicrite (m2). D. Detailof micrite types and fabrics within the pore space of an articulated bivalve shell, including pe, clotted m2 and neomorphosed m2 that is now microspar (ms); plane-polarized light.
Fig. 5. Petrography of sandy (detrital) seep carbonate deposit at Awanui I. A. Thin section overview showing mottled to faintly clotted micrite cementing the background siliciclasticsediments (m1), containing framboidal pyrite (small dark grains); angular, detrital quartz grains; small, spar-filled fenestrae; and large bioclasts of inoceramid bivalve prisms andother broken molluscan shells. B. Detail of m1 fabric, showing clotted textures (cm) in places, and large prismatic shell fragment of an inoceramid. Some quartz grains show thindark microcrystalline carbonate linings, inferred as microbial coatings. Spar filled, irregular fenestra contains cluster of framboidal pyrite grains (py); plane-polarized light. C. Detailof carbonate cement types filling a vug set in m1. bbc, early diagenetic fibrous, banded and botryoidal cement; ec, late diagenetic equant calcite spar; plane-polarized light. D. Vug inm1 showing a complex cement fill history. Vug was first partly lined by bbc and an authigenic micrite (m2) that is detrital poor and shows evidence of microbially mediated growthby its globular, clotted and faintly laminated fabric. In places the vug was then lined by a thin isopachous horizon of scalenohedral calcite (sCc),with remaining vug porosity filled bya homogeneous, late diagenetic, detrital poor micrite (m3). Some of the bbc fabric was replaced by small irregular nodules of diagenetic silica (s); plane-polarized light.
8 S. Kiel et al. / Palaeogeography, Palaeoclimatology, Palaeoecology xxx (2012) xxx–xxx
Please cite this article as: Kiel, S., et al., Cretaceous methane-seep deposits from New Zealand and their fauna, Palaeogeography,Palaeoclimatology, Palaeoecology (2012), http://dx.doi.org/10.1016/j.palaeo.2012.10.033

Fig. 7. Petrography of micritic seep carbonate deposit at Waipiro I; all crossed-polarized light. A. Vug within spheroidal micrite (m2) filled with fibrous, botryoidal and banded cal-cite (bbc) and homogeneous, late diagenetic micrite (m3). Part of bbc replaced by diagenetic silica (s). B. Coated grains of putative microbial origin enclosed by spheroidal micrite(m2) and microspar (ms); the coated grain on the lower right is partially silicified (s). C. Coated grains (cg) in a matrix of spheroidal micrite (m2); coated grain is partially silicified(s). D. Spheroidal micrite detail.
Fig. 8. Petrography of micritic seep carbonate deposit at Waipiro III. A. Hand sample of typical Waipiro III rock fabric showing irregular, mottled to clotted brownmicrite interruptedby numerous large irregular vugs filled with carbonate cements and silica (s). B. Detail of detrital poor, clotted micrite fabric (m2) with irregular pore spaces filled with translucentfibrous calcite in a common carbonate fabric known as ‘structure grumeleuse’. Small coated grain (cg) also preserved; plane-polarized light. C. Clotted to spheroidal micrite (m2) cutby thin and thick veins filled with late diagenetic equant calcite spar (ec). Possible recrystallized radiolarian (arrow); plane-polarized light. D. Coated grain in a matrix of peloidal(m1) and clotted micrite (m2); plane-polarized light. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)
9S. Kiel et al. / Palaeogeography, Palaeoclimatology, Palaeoecology xxx (2012) xxx–xxx
Please cite this article as: Kiel, S., et al., Cretaceous methane-seep deposits from New Zealand and their fauna, Palaeogeography,Palaeoclimatology, Palaeoecology (2012), http://dx.doi.org/10.1016/j.palaeo.2012.10.033

Fig. 9. Lithologic and petrographic characteristics of the Owhena deposit. A. Field photograph of concretionary carbonate (grey band, nodules) in fissile mudstone (dark grey, ho-mogeneous background sediments). B. Hand sample of grey spheroidal micrite nodule, inferred as a microbial thrombolite, with beige fibrous cement-filled vugs, and all cross-cutby thick, jagged veins filled with white calcite spar. C. Detail of spheroidal micrite patch demarcated on its outer margin by a pyrite-coated corrosion surface (arrow) and cross-cutby late diagenetic sparry calcite veins (white). Nodular sphere-rich regions are surrounded by sphere- and detrital-poor, clotted micrites (m2); plane-polarized light. D. Close upview of spheroidal micrite, showing subangular to subrounded, concentrically laminated nature of the fabric; plane-polarized light.
10 S. Kiel et al. / Palaeogeography, Palaeoclimatology, Palaeoecology xxx (2012) xxx–xxx
biogeochemical interpretation of these isotopic values is discussed inSection 5.1.
4.3. Biomarkers
The Awanui II and Waipiro III limestones revealed different degreesof preservation of lipid biomarkers. Whereas the Awanui II limestoneshows a moderate preservation of molecular fossils of prokaryotesinvolved in methane consumption at this location, the lipid pattern ofthe Waipiro III limestone apparently has been altered. Apart fromn-alkanes, isoprenoids are the dominant compounds of the hydrocar-bon fraction of the Awanui II limestone (Fig. 11A). Among those,phytane yielded a δ13C value of −59‰, pristane a value of −37‰.Biphytane (−99‰) and 2,6,10,15,19-pentamethylicosane (PMI;−97‰) were found to be even more 13C-depleted, revealing δ13Cvalues that are typical of prokaryotes involved in anaerobic oxidationof methane. The n-alkanes, on the other hand, with an average δ13Cvalue of −30‰ for C17 to C22 are much less 13C-depleted with theexception of n-tricosane (C23), which is slightly more 13C-depleted(−36‰) than the other n-alkanes. Due to substantial biodegradation,reflected in two humps of unresolved complex mixtures, it was notpossible to obtain δ13C values of the compounds in the hydrocarbonfraction of the Waipiro III limestone. The isoprenoids identified includepristane, phytane, and PMI. The only other compounds present in thisfraction are n-alkanes and traces of steranes.
The carboxylic acid fraction of the Awanui II limestone is character-ized by an even-over-odd dominance of n-fatty acids (Fig. 11B). Apartfrom n-fatty acids, terminally-branched C15 fatty acids have been recog-nized. Anteiso-C15 fatty acid revealed a lower δ13C value (−47‰) thaniso-C15 fatty acid (−35‰). The δ13C value of iso-C15 fatty acid is onlyslightly more negative than the values of short-chain, mid-chain, andlong chain n-fatty acids ranging from −34 to −32‰. The δ13C valueof ββ-C32 hopanoic acid of −42‰ falls between the values of the two
Please cite this article as: Kiel, S., et al., Cretaceous methane-seepPalaeoclimatology, Palaeoecology (2012), http://dx.doi.org/10.1016/j.pala
terminally-branched fatty acids. The n-fatty acids extracted from theWaipiro III limestone show similar δ13C values (−35 to −30‰) likethose of the Awanui II limestone. However, the amount of fatty acidsin the Waipiro III limestone is much lower than in the Awanui II lime-stone and no compounds other than n-fatty acids were detected.
4.4. Fauna
The most conspicuous taxa of the macrofauna are lucinid andmodiomorphid bivalves, and at Waipiro III a large abyssochrysoid gas-tropod. Small hokkaidoconchid gastropods, solemyid, nucinellid, andthyasirid clams, as well as inoceramid bivalves also are present atmost seep deposits. Minor components include poorly preservednuculanid bivalves and a diversity of small gastropods, including aSerradonta limpet at Waipiro III, crustaceans, and a solitary coral atAwanui GS 688. A list of the taxa and their distribution among sites isgiven in Table 1.
The modiomorphid bivalve found at Awanui II and Awanui GS 688reaches 160 mm in length and resembles certain species of the seep-restricted genus Caspiconcha (Fig. 13D,E). Similar are C. major fromCalifornia (Stanton, 1895; Jenkins et al., in press) and a species reportedas “Calyptogena sp.” from the Omagari seep site in Japan (Hikida et al.,2003) which is now considered as belonging to Caspiconcha(cf. Amano and Kiel, 2007; Jenkins et al., in press). Both of these specieshave a similar broad and sometimes curved posterior side. In contrast,Caspiconcha whithami from Greenland (Kelly et al., 2000), C. rubanifrom the Crimean peninsula (Kiel et al., 2010), and Caspiconchasp. from an Albian seep locality near Ispaster in northern Spain(Agirrezabala et al., this volume) are more elongate and have an almostlinear or sub-lineate ventral margin.
With at least five species, the lucinids are very diverse, but are dif-ficult to characterize taxonomically due to poor preservation. Four ofthe five species might belong to Ezolucina. Firstly the most common
deposits from New Zealand and their fauna, Palaeogeography,eo.2012.10.033

Fig. 10. Isotopic cross plot of δ13C vs δ18O values (‰ PDB) for the Cretaceous seep carbon-ates and fossil shells evaluated in this study. Awanui I andWaipiro I cement phases recordthe lowest carbonate carbon values of all samples. Awanui II and Waipiro III samples arerelatively most enriched in 13C, with the Owhena samples yielding intermediate δ13Cvalues. The bivalve shells indicate precipitation in isotopic equilibrium with seawaterbicarbonate. With respect to oxygen isotopic signatures, most samples are early micrites(m1 and m2 fabrics) and indicate relatively minor diagenetic modification. Early andlate cements from Waipiro III and Owhena carbonates are the most diagenetically modi-fied, with late diagenetic equant calcite spar samples showing the greatest depletion in18O. See text for further discussion.
11S. Kiel et al. / Palaeogeography, Palaeoclimatology, Palaeoecology xxx (2012) xxx–xxx
lucinid at the Awanui II and Awanui GS 688 sites, here referred to asEzolucina sp. “Awanui” (Fig. 13G–J), reaches 50 mm in length, resem-bles small specimens of Ezolucina inflata from the Cenomanian ofHokkaido, Japan (Amano et al., 2008), and appears to have strong car-dinal teeth. Secondly the large (length 120 mm) species fromWaipiro, here named Ezolucina sp. “Waipiro” (Fig. 15A, B) resemblesthe large, veneriform specimens of the Japanese E. inflata (Amanoet al., 2008). Thirdly the species from Awanui GS 688, which reaches
Please cite this article as: Kiel, S., et al., Cretaceous methane-seepPalaeoclimatology, Palaeoecology (2012), http://dx.doi.org/10.1016/j.pala
37 mm in length, is similar to Ezolucina sp. “Awanui” but differs inpossessing a strong posterior radial ridge and is here namedEzolucina? sp. “ridge” (Fig. 13L). The fourth species from Awanui I,reaching 27 mm in length, is also similar to Ezolucina sp. “Awanui”but differs by being taller; it is here designated as Ezolucina? sp.“tall” (Fig. 12B). Finally, the fifth lucinid resembles Nipponothracia,was found at Awanui GS 688 (Fig. 13M) where it reaches 80 mm inlength, and possibly occurs also at Awanui I (Fig. 12C) where itreaches 32 mm in length. This genus is known from many fossilseep sites of the Pacific realm from mid-Cretaceous (Albian) to Plio-cene time (Kanie and Sakai, 1997; Kase et al., 2007; Kiel et al.,2008a). The New Zealand species differs markedly from all other Cre-taceous Nipponothracia species by being relatively flat and elongate,and having the umbo in a comparatively anterior position. Withthese characters it is more similar to the younger N. elassodyseides(Saul et al., 1996) from the Eocene of western Washington. All livinglucinids examined to date have sulfide-oxidizing symbionts (Dandoet al., 1986; Taylor and Glover, 2006) and this mode of life is also in-ferred for their fossil counterparts.
Solemyids at Awanui II (Fig. 13A) and Waipiro III are too poorlypreserved for generic identification and biogeographic interpreta-tions. Nucinella sp. at Awanui II (Fig. 13B, C), and possibly at AwanuiI (Fig. 12A), is rare and reaches no more than 5 mm in length. It isthus remarkably smaller than the Nucinella species found at other fos-sil seep sites (Amano et al., 2007; Kiel et al., 2008a; Hammer et al.,2011; Peckmann et al., 2011). The few thyasirid bivalves found atAwanui II and GS688 (Fig. 13F) and Waipiro III never exceed 10 mmin length and thus also are small compared to other thyasirids at Cre-taceous seep deposits, which can reach up to 80 mm in length(Nobuhara et al., 2008). Solemyids, Nucinella, and most largethyasirid bivalves live in symbiosis with sulfide-oxidizing bacteria(Dando and Southward, 1986; Fisher and Childress, 1986; Dufour,2005; Taylor and Glover, 2010; Oliver and Taylor, 2012). Thus we as-sume this mode of life for the species reported here as well.
The Waipiro III seep deposit is dominated numerically by a largeabyssochrysoid gastropod that reaches to at least 34 mm in height(Fig. 15I-L). Extant abyssochrysoids include characteristic vent andseep taxa such as the ubiquitous Provanna and the large, vent-inhabiting Ifremeria and Alviniconcha (Johnson et al., 2010), but thenominate genus Abyssochrysos is a rare deep-water taxon that is notrestricted to seeps or other chemosynthesis-based ecosystems. Largeabyssochrysoid species similar to the one from Waipiro III are knownfromEarly Cretaceous seep deposits in California and possibly CzechRe-public (Kiel et al., 2010). However, the feeding strategy of these large,seep-inhabiting gastropods is unknown.
There are two or possibly three species of Hokkaidoconcha in theCretaceous seep deposits of New Zealand. The slender Hokkaidoconchasp. 1 from Awanui II (Fig. 14C) is similar to the Campanian speciesH. tanabei from Japan and the Early Cretaceous H. occidentalis fromCalifornia (Kaim et al., 2008; Kiel et al., 2008b). However, themost sim-ilar species isHokkaidoconcha sp. from the Albian of Spain (Agirrezabalaet al., this volume). The other specimens found at Awanui GS688(Fig. 14D) and Waipiro III (Fig. 15H) have more numerous and densercollabral ribs andmay belong to one or two species. These small gastro-pods were likely bacteria- and detritus-feeders, as are their modernrelatives, the provannids (Kaim et al., 2008). A single, small, fusiformgastropod found at Awanui I (Fig. 12F) resembles certain neogastropodssuch as columbellids, but also is similar to the seep endemic genusAtresius known from Early Cretaceous seep deposits in California (Kielet al., 2008b) and the extant provannid Rubyspira (Johnson et al.,2010). Due to the lack of diagnostic characters, however, its identityremains uncertain.
The tall, slender limpet Serradonta sp. found atWaipiro III (Fig. 15C–E)is 4 mm high and is very similar to the Late Cretaceous Serradontaomagariensis from northern Japan (Jenkins et al., 2007; Kaim et al.,2009). The small globular gastropod from Awanui GS688 (Fig. 14A, B)
deposits from New Zealand and their fauna, Palaeogeography,eo.2012.10.033

Fig. 11. Lipid biomarker results. (A) Gas chromatograms (total ion currents) of the hydro-carbon fractions of Awanui II andWaipiro III limestones. Circles: n-alkanes; white triangles:head-to-tail linked isoprenoids; black triangles: tail-to-tail linked isoprenoids; graytriangles: head-to-head linked isoprenoids; black stars: contaminants; pr: pristane;ph: phytane; istd: internal standard; PMI: 2,6,10,15,19-pentamethylicosane; UCM:unresolved complex mixture; LCAs: long-chain n-alkanes. Compound-specific δ13Cvalues are indicated for isoprenoids, n-C23 alkane, as well as an average value forshort-chain n-alkanes from the Awanui II sample. The δ13C values of compoundsassigned to methane-oxidizing archaea are indicated by bold numbers. (B) Gas chro-matograms (total ion currents) of the carboxylic acid fractions of Awanui II andWaipiro IIIlimestones. Circles:n-fatty acids; stars: contaminants; i-/ai-15: iso-/anteiso-C15 fatty acids;istd: internal standard. Compound-specific δ13C values are indicated for i-/ai-C15 fattyacids and the ββ-C32 hopanoic acid from the Awanui II sample, as well as average valuesfor short-chain fatty acids (C14–C18), mid-chain fatty acids (C19–C25), and long-chain fattyacids (C26–C29) for Awanui II andWaipiro III samples. The δ13C values of compounds ten-tatively assigned to sulfate-reducing bacteria are indicated by bold numbers.
12 S. Kiel et al. / Palaeogeography, Palaeoclimatology, Palaeoecology xxx (2012) xxx–xxx
resembles the (presumed) neomphalin Retiskenea? kieli from anAptian-Albian seep deposit at Cold Fork of Cottonwood Creek in Califor-nia (Campbell et al., 2008b). However, the poor state of preservation ofthe New Zealand specimens does not allow a definitive identification as
Please cite this article as: Kiel, S., et al., Cretaceous methane-seepPalaeoclimatology, Palaeoecology (2012), http://dx.doi.org/10.1016/j.pala
Retiskenea. Finally the crustacean fragments (Fig. 15G)most likely belongto a mud-dwelling, deposit-feeding callianassid shrimp.
5. Discussion
5.1. Nature of the carbonate blocks
The biomarker pattern obtained for the Awanui II limestone con-firms that the biogeochemical process of sulfate-dependent anaerobicoxidation of methane (AOM) shaped the paleoenvironment, whichboth sustained the observed endemic fauna and favored carbonateprecipitation. Molecular fossils of archaea involved in AOM include13C-depleted PMI and biphytane (cf. Peckmann and Thiel, 2004;Birgel et al., 2008b). Based on its intermediate δ13C value of −59‰,falling between the values of the archaeal isoprenoids and the associatedn-alkanes, phytane likely derived frommixed sources, including lipids ofmethanotrophic archaea and marine phototrophs (cf. Peckmann et al.,2009). A lower δ13C value of n-tricosane compared to other n-alkanesagrees with observations made in other seep limestones and microbialmats at modern seeps. This compound derives from n-tricos-10-ene,which is produced by yet unknown prokaryotes involved in AOM(Thiel et al., 2001b). Sulfate-reducing bacteria of the Desulfosarcina/Desulfococcus branch are commonly the syntrophic partners ofmethanotrophic archaea in AOM (e.g., Knittel and Boetius, 2009).Anteiso-C15 fatty acid is particularly abundant in sulfate reducers,which accompany the so-called ANME-1 group of methanotrophic ar-chaea (Thiel et al., 2001a; Niemann and Elvert, 2008). A derivation ofthe Awanui II anteiso-C15 fatty acid from sulfate reducers involved inAOM agrees with its low δ13C value of −47‰. A dominance ofANME-1 archaea over other groups of archaeal methanotrophs at theAwanui II seep is also indicated by the occurrence of biphytane andthe lack of the isoprenoid crocetane (cf. Blumenberg et al., 2004;Niemann and Elvert, 2008). In accordance with the dominance ofmicrite and the scarcity of early diagenetic fibrous cement in theAwanuiII limestone, this indicates that seepage was rather diffusive than advec-tive at this ancient seep site (cf. Peckmann et al., 2009). Based on thelowest δ13Ccarbonate value of the Awanui II limestone of −12.1‰, itwould not have been possible to confirm that the seepage fluidscontained methane, as it is revealed by the biomarker data. The onlymoderately low δ13Ccarbonate values corroborate that a substantial degreeof admixture of marine carbonate with methane-derived carbonatemust have contributed to carbonate formation to explain such compara-bly high values, again pointing to diffusive rather than advective seepage(cf. Peckmann et al., 2009). The source of ββ-C32 hopanoic acid in theAwanui II limestone cannot be assessed with certainty, but its δ13Cvalue of −42‰, quite similar to the value of anteiso-C15 fatty acidassigned to sulfate reducers, agrees best with a derivation fromsulfate-reducing bacteria involved in AOM. When found in seep lime-stones, this compound has also been interpreted to derive from aerobic
deposits from New Zealand and their fauna, Palaeogeography,eo.2012.10.033

Fig. 12. Fauna of the seep deposit Awanui I, Raukumara Peninsula, New Zealand. A, Nucinella? sp. (TM8823). B, Ezolucina? “tall” (TM8824). C, Nipponothracia? sp. (TM8825). D,carditiid (TM8826). E, unidentified heterodont bivalve (TM8827). F, small, fusiform gastropod (Atresius? Columbellid?) (TM8828).
13S. Kiel et al. / Palaeogeography, Palaeoclimatology, Palaeoecology xxx (2012) xxx–xxx
methanotrophs in case of an even stronger 13C-depletion (Birgel et al.,2011) or unknown marine bacteria not involved in AOM in case of less13C-depletion (Kuechler et al., 2012). Abundant framboidal pyrite inthin sections from Awanui II supports the presence of sulfate reductionduring carbonate formation.
The Waipiro III limestone yielded similar, only moderately lowδ13Ccarbonate values like the Awanui II limestone. Unfortunately, thislimestone proved to be too biodegraded to evaluate past biogeochemi-cal processes and the composition of seepage fluids with certainty. Thepresence of PMI agrees with the seep interpretation and AOM. With noδ13C value of this compound available owing to biodegradation, itcannot yet be excluded that PMI derives from non-methanotrophicarchaea. Nonetheless, the paragenetic sequence of the Waipiro III lime-stone, containing microbial carbonate fabrics such as clotted micriteand the sum of other observations including carbon stable isotopesof carbonate, biomarkers, and fauna (see below), agree best with aninterpretation of the limestone as a hydrocarbon-seep deposit.
Waipiro I limestones exhibit a paragenetic sequence with phasesthat are typical of seep deposits, including different varieties ofauthigenic micrite (cf. Campbell, 2006), fibrous, banded and botryoidalcement (cf. Teichert et al., 2005), and early formed silica replacingfibrous cement (cf. Kuechler et al., 2012). Spheroidal textures like thecoated grains and the smaller spheroids of the Waipiro I limestone arenot particularly common in seep deposits, but similar textures havebeen recognized in some instances (Jenkins et al., 2008; De Boeveret al., 2011). The lowest δ13C value of coated grains of −25.4‰ agreeswith hydrocarbon oxidation as a major source of carbonate. Oxidationof methane seems likely, but on the basis of δ13Ccarbonate values alone,oxidation of crude oil cannot be excluded, although it should be stressedthat−25.4‰ is rather close to the negative limit of values expected toresult from oil degradation (cf. Peckmann and Thiel, 2004).
Similar to the Awanui II limestone, the Awanui I limestone is domi-nated by micrite that cements detrital background sediment. Thislithology also contains minor phases including different varieties ofnon-detrital micrite, banded and botryoidal cement, and early formedsilica replacing fibrous cement, all in accord with a seep interpretation.
Please cite this article as: Kiel, S., et al., Cretaceous methane-seepPalaeoclimatology, Palaeoecology (2012), http://dx.doi.org/10.1016/j.pala
Such an inference is confirmed by the lowest δ13C value of −29.2‰found for micrite. Values close to −30‰ of carbonate phases formingat marine seeps are unlikely to reflect biodegradation of crude oil andare best explained by oxidation ofmethane. The presence of 13C-deplet-ed authigenic micrite (−13.9‰) and banded and botryoidal cement(−13.3‰) in the Owhena limestone agrees with carbonate forma-tion at a seep, but does not allow us to assess the composition ofhydrocarbons in the seepage fluids. It is unusual for seep deposits(cf. Campbell, 2006) that the late diagenetic equant spar cementsfrom Owhena and Waipiro III (one sample analyzed from each;Fig. 10) suggest a trend to lower δ
13C values. But based on the limited
number of isotope measurements, contemplation of the significance isconsidered premature. For the same reason, any extrapolation ofoxygen isotope patterns seems inappropriate given the small data set.Unlike carbon isotopes, oxygen isotopes aremuchmore prone to diage-netic alteration. The low δ18O values of all samples, but particularly theextremely low values of Owhena samples reflect alteration of primaryisotopic compositions in accordance with the moderate to poor pre-servation of lipid biomarkers, both pointing to substantial thermalmaturation.
5.2. Biogeographic relationships
At family and genus level, the faunas of the seep deposits atWaipiroBay and Port Awanui resemble those of other Cretaceous seep deposits.This supports the concept of the ‘Late Mesozoic Seep Fauna’: a faunathat is largely restricted to deep-water methane-seep deposits of lateMesozoic age, with a world-wide distribution (cf. Kiel and Peckmann,2008; Kiel, 2010). In comparing similarities of the species of the Creta-ceous New Zealand seeps to those of other Late Cretaceous seeps,close biogeographic relations to northern Japan, California and, to alesser extent, northern Spain, emerge (Table 1). This applies to boththe Late Albian to mid-Cenomanian faunas at Port Awanui and themid-Campanian faunas at Waipiro Bay. There is little similarity at thespecies or genus level with the Early Cretaceous Tethyan seep faunasin southern France and the Ukraine (Thieuloy, 1972; Kiel and
deposits from New Zealand and their fauna, Palaeogeography,eo.2012.10.033
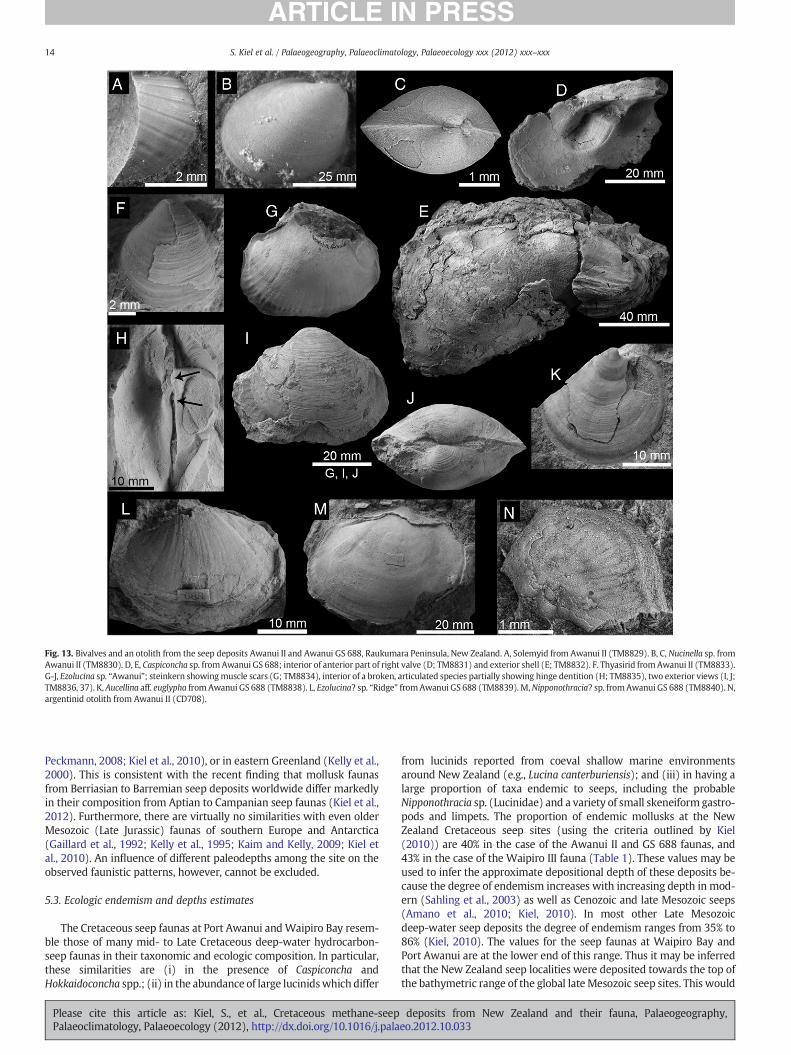
Fig. 13. Bivalves and an otolith from the seep deposits Awanui II and Awanui GS 688, Raukumara Peninsula, New Zealand. A, Solemyid from Awanui II (TM8829). B, C, Nucinella sp. fromAwanui II (TM8830). D, E, Caspiconcha sp. fromAwanui GS 688; interior of anterior part of right valve (D; TM8831) and exterior shell (E; TM8832). F. Thyasirid fromAwanui II (TM8833).G-J, Ezolucina sp. “Awanui”; steinkern showingmuscle scars (G; TM8834), interior of a broken, articulated species partially showing hinge dentition (H; TM8835), two exterior views (I, J;TM8836, 37). K, Aucellina aff. euglypha fromAwanui GS 688 (TM8838). L, Ezolucina? sp. “Ridge” fromAwanui GS 688 (TM8839). M,Nipponothracia? sp. fromAwanui GS 688 (TM8840). N,argentinid otolith from Awanui II (CD708).
14 S. Kiel et al. / Palaeogeography, Palaeoclimatology, Palaeoecology xxx (2012) xxx–xxx
Peckmann, 2008; Kiel et al., 2010), or in eastern Greenland (Kelly et al.,2000). This is consistent with the recent finding that mollusk faunasfrom Berriasian to Barremian seep deposits worldwide differ markedlyin their composition from Aptian to Campanian seep faunas (Kiel et al.,2012). Furthermore, there are virtually no similarities with even olderMesozoic (Late Jurassic) faunas of southern Europe and Antarctica(Gaillard et al., 1992; Kelly et al., 1995; Kaim and Kelly, 2009; Kiel etal., 2010). An influence of different paleodepths among the site on theobserved faunistic patterns, however, cannot be excluded.
5.3. Ecologic endemism and depths estimates
The Cretaceous seep faunas at Port Awanui andWaipiro Bay resem-ble those of many mid- to Late Cretaceous deep-water hydrocarbon-seep faunas in their taxonomic and ecologic composition. In particular,these similarities are (i) in the presence of Caspiconcha andHokkaidoconcha spp.; (ii) in the abundance of large lucinidswhich differ
Please cite this article as: Kiel, S., et al., Cretaceous methane-seepPalaeoclimatology, Palaeoecology (2012), http://dx.doi.org/10.1016/j.pala
from lucinids reported from coeval shallow marine environmentsaround New Zealand (e.g., Lucina canterburiensis); and (iii) in having alarge proportion of taxa endemic to seeps, including the probableNipponothracia sp. (Lucinidae) and a variety of small skeneiform gastro-pods and limpets. The proportion of endemic mollusks at the NewZealand Cretaceous seep sites (using the criteria outlined by Kiel(2010)) are 40% in the case of the Awanui II and GS 688 faunas, and43% in the case of the Waipiro III fauna (Table 1). These values may beused to infer the approximate depositional depth of these deposits be-cause the degree of endemism increases with increasing depth in mod-ern (Sahling et al., 2003) as well as Cenozoic and late Mesozoic seeps(Amano et al., 2010; Kiel, 2010). In most other Late Mesozoicdeep-water seep deposits the degree of endemism ranges from 35% to86% (Kiel, 2010). The values for the seep faunas at Waipiro Bay andPort Awanui are at the lower end of this range. Thus it may be inferredthat the New Zealand seep localities were deposited towards the top ofthe bathymetric range of the global lateMesozoic seep sites. This would
deposits from New Zealand and their fauna, Palaeogeography,eo.2012.10.033

Fig. 14. Gastropods from the seep deposits Awanui II and Awanui GS 688, RaukumaraPeninsula, New Zealand. A. Retiskenea? sp. from Awanui GS 688 (TM8841). B. Planispiralgastropod from Awanui II (TM8842). C. Hokkaidoconcha sp. 1 from Awanui GS 688(TM8843). D. Hokkaidoconcha sp. 2 from Awanui II (TM8844). E. Aporrhaid gastropodfrom Awanui GS 688 (TM8845).
15S. Kiel et al. / Palaeogeography, Palaeoclimatology, Palaeoecology xxx (2012) xxx–xxx
place theNew Zealand Cretaceous seeps in an outer shelf to upper slopesetting. The argentinid otolith found at Awanui II (Fig. 13N)might serveas an independent control for this estimate: extant argentinid fish aregenerally found on the outer shelf to upper slope (Paxton and Cohen,1999). Around New Zealand these fish are found mainly below 300 mdepth (Jacob et al., 1998), supporting an outer shelf to upper slope set-ting for the fossils. The fauna at Awanui I has a very low proportion ofendemics, which partlymay be explained by poor preservation and cor-responding difficulties in taxonomic identifications.
5.4. New Zealand seep faunas through time
The marine mollusk fauna of New Zealand is characterized by itshigh degree of endemism in both the Recent and Cenozoic (Beu andMaxwell, 1990; Spencer et al., 2009). It is therefore noteworthy, asoutlined below, that the Cretaceous to present-day seep faunas ofNewZealand did not develop a strong local endemismbut consist large-ly of taxa known from coeval vents and seeps around theworld.Moderntaxonomic accounts of the broader Cretaceous mollusk fauna of NewZealand are few (Spencer et al., 2009), but a screening of the available
Please cite this article as: Kiel, S., et al., Cretaceous methane-seepPalaeoclimatology, Palaeoecology (2012), http://dx.doi.org/10.1016/j.pala
literature suggests that many species belong to cosmopolitan genera,as well as some to genera that are restricted to the Southern Hemi-sphere (i.e., Conchothyra and Struthioptera (Stilwell, 2003) and refer-ences therein). As described herein, both the Albian-Cenomanian andthe Campanian seep faunas in New Zealand consist largely of generaknown from coeval seeps worldwide.
More than a dozen Cenozoic fossil seep deposits in New Zealandoccur in lower to upper Miocene strata of the East Coast Basin, NorthIsland (Campbell et al., 2008a). The fauna of these deposits is diverseat the generic level and consists of typical seep-inhabiting taxa such asthe chemosymbiotic bivalves Bathymodiolus, Gigantidas, Lucinoma,Thyasira and Calyptogena (sensu lato), and the bacteria-grazing gastro-pods Provanna, Homalopoma and possibly Serradonta (Saether et al.,2010a, 2010b, 2012; Saether, 2011). In addition, there is a suite ofsponges, corals, crustaceans, brachiopods and other mollusks thatwere probably inhabitants of the surrounding seafloor and which tookadvantage of the abundance of food at the seep sites, rather thanbeing seep specialists (Saether, 2011). Most of the typical Mioceneseep taxa from New Zealand are known also from other Miocene seepdeposits around the world (Taviani, 1994; Gill et al., 2005; Majimaet al., 2005; Amano et al., 2010; Kiel, 2010). This is an interestingcontrast to the general Miocene mollusk fauna of New Zealand: at thegenus level, about 20% of the shelf mollusks are endemic and about34% of the mollusks occupied deeper waters beyond the shelf (AlanBeu, pers. comm. 2012, based on taxonomic data held in the SynopticDataset described in Crampton et al. (2006a)).
The majority of the extant vent and seep fauna around NewZealand is composed of taxa known from other vents and seepsaround the world, including the bivalves Calyptogena (sensu lato),Bathymodiolus, Gigantidas, Vulcanidas, Acharax and Lucinoma, the gas-tropods Provanna, Xylodiscula and Lurifax, and the siboglinid tubeworm Lamellibrachia (Lewis and Marshall, 1996; Cosel and Marshall,2003, 2010; Baco et al., 2010). In contrast, the overall, extant marinemollusk fauna of the New Zealand region consists of about 82%endemic species and about 10% endemic genera (calculated fromunpublished data compilations supplied by D. Gordon (NIWA,Wellington), based on the taxonomic lists of Spencer et al. (2009)).
In summary, since Cretaceous times, seeps in New Zealand havebeen colonized repeatedly or perhaps even continuously by membersof the global seep fauna, whereas the general fauna of the NewZealand region shows an often considerable degree of endemism, atleast in relation to mollusks. The historical and ecological reasonsfor these remarkably different degrees of endemism of New Zealand'sdeep-water fauna are still unclear. Further research will likely provideinsights into the roles of geographic isolation, climate change andevolutionary rates in shaping the shallow and deep-water faunas ofNew Zealand.
6. Conclusions
Evidence from petrography, negative δ13C values of early diageneticmicrites (as low as−29.2‰) and 13C-depleted molecular fossils clearlyshow that the investigated limestones from the eastern Raukumarapeninsula, North Island, New Zealand, are ancient methane seepdeposits. Dinoflagellate dating indicates a Late Albian to mid-Cenomanian age for the deposits from Port Awanui, a mid-Campanianage for the deposits from Waipiro Bay, and also a mid-Campanian agefor the deposit from Owhena Stream. The associated mollusk fauna issimilar to Late Mesozoic seep faunas worldwide and shows close bio-geographic relationships to Cretaceous North Pacific seep faunas.When traced through geologic time, seeps in New Zealand have repeat-edly been colonized by mollusks of the global deep-water seep faunasince the Cretaceous, while the general marine mollusk fauna of NewZealand, including the deep-water fauna, often shows a considerabledegree of endemism.
deposits from New Zealand and their fauna, Palaeogeography,eo.2012.10.033

Fig. 15. Fossils from the seep deposits at Waipiro Bay, Raukumara Peninsula, New Zealand. A, B. Ezolucina sp. “Waipiro” from Waipiro I (TM8846). C-E. Serradonta sp. from WaipiroIII (TM8847). F Naticid? gastropod from Waipiro III (TM8848). G. Callianassa claw from Waipiro III (AR2508). H. Hokkaidoconcha sp. 2 (?) from Waipiro III (TM8849). I-L.Abyssochrysoid gastropod from Waipiro III (TM8850–53).
16 S. Kiel et al. / Palaeogeography, Palaeoclimatology, Palaeoecology xxx (2012) xxx–xxx
Acknowledgments
Thanks to Alan Beu (GNS Science, Lower Hutt) for information onthe Cenozoic mollusks of New Zealand; D. Gordon (NIWA, Wellington)for supplying unpublished data on the living fauna of NewZealand; DirkNolf (Brussels) for his comment on the otolith; Gerhard Hundertmarkand Tanja Stegemann (University of Göttingen) for photography; LouiseCotterall (University of Auckland) for assistance with drafting; NevilleHudson (University of Auckland) for help with fossil collections; KrisSaether and Ivana Alfaro (Auckland) for aiding rock sample prepara-tion; Birgit Wild and Andreas Richter (University of Vienna) for aidmeasuring compound-specific stable carbon isotopes; Beatrix Bethke(University of Vienna) for assistance setting up the lipid biomarker laband sample extraction; and Axel Hackmann (University of Göttingen)and Andrés Arcila (University of Auckland) for thin section preparation.Alan Beu and an anonymous reviewer are thanked for their constructivecriticism. JSC and PS acknowledge support from the Ministry of Science
Please cite this article as: Kiel, S., et al., Cretaceous methane-seepPalaeoclimatology, Palaeoecology (2012), http://dx.doi.org/10.1016/j.pala
and Innovation programmes C050X905, Petroleum Source Rocks andfluids and GNS Science's Direct Crown Funded programme GlobalChange Though Time, Biostratigraphy and Time Scales (GCT8), PS fur-ther acknowledges support from the Carlsberg Foundation (Copenha-gen) during fieldwork in 1999; KAC acknowledges support from theRoyal Society of NewZealandMarsden fund; and SK acknowledges sup-port from the Deutsche Forschungsgemeinschaft through grant Ki802/6-1. We also acknowledge use of information contained within theNew Zealand Fossil Record File (http://www.fred.org.nz/).
References
Agirrezabala, L.M., Kiel, S., Blumenberg, M., Schäfer, N., Reitner, J., this volume. Outcropanalogues of pockmarks and associated methane-seep carbonates: a case studyfrom Lower Cretaceous (Albian) of the Basque–Cantabrian Basin, western Pyrenees.Palaeogeography, Palaeoclimatology, Palaeoecology.
Amano, K., Kiel, S., 2007. Fossil vesicomyid bivalves from the North Pacific region. Veliger49, 270–293.
deposits from New Zealand and their fauna, Palaeogeography,eo.2012.10.033

17S. Kiel et al. / Palaeogeography, Palaeoclimatology, Palaeoecology xxx (2012) xxx–xxx
Amano, K., Jenkins, R.G., Hikida, Y., 2007. A new gigantic Nucinella (Bivalvia:Solemyoida) from the Cretaceous cold-seep deposit in Hokkaido, northern Japan.Veliger 49, 84–90.
Amano, K., Jenkins, R.G., Kurihara, Y., Kiel, S., 2008. A new genus for Vesicomya inflataKanie and Nishida, a lucinid shell convergent with that of vesicomyids, from Creta-ceous strata of Hokkaido, Japan. Veliger 50, 255–262.
Amano, K., Jenkins, R.G., Aikawa, M., Nobuhara, T., 2010. A Miocene chemosyntheticcommunity from the Ogaya Formation in Joetsu: evidence for depth-related eco-logic control among fossil seep communities in the Japan Sea back-arc basin.Palaeogeography, Palaeoclimatology, Palaeoecology 286, 164–170.
Backhouse, J., 2006. Albian (Lower Cretaceous) dinoflagellate cysts biostratigraphy ofthe Lower Gearle Siltstone, southern Carnarvon Basin, Western Australia. Palynology30, 43–68.
Baco, A.R., Rowden, A.A., Levin, L.A., Smith, C.R., Bowden,D.A., 2010. Initial characterizationof cold seep faunal communities on the New Zealand Hikurangi margin. MarineGeology 272, 251–259.
Batten, D.J., 1999. Small palynomorphs. In: Jones, T.P., Rowe, N.P. (Eds.), Fossil Plantsand Spores: Modern Techniques. Geological Society, London, pp. 15–19.
Beu, A.G., Maxwell, P.A., 1990. Cenozoic Mollusca of New Zealand. New ZealandGeological Survey Paleontological Bulletin 58, 1–518.
Birgel, D., Thiel, V., Hinrichs, K.-U., Elvert, M., Campbell, K.A., Reitner, J., Farmer, J.D.,Peckmann, J., 2006. Lipid biomarker patterns of methane-seep microbialites fromtheMesozoic convergent margin of California. Organic Geochemistry 37, 1289–1302.
Birgel, D., Elvert, M., Han, X., Peckmann, J., 2008a. 13C-depleted biphytanic diacids astracers of past anaerobic oxidation of methane. Organic Geochemistry 39, 152–156.
Birgel, D., Himmler, T., Freiwald, A., Peckmann, J., 2008b. A new constraint on the antiq-uity of anaerobic oxidation of methane: Late Pennsylvanian seep limestones fromsouthern Namibia. Geology 36, 543–546.
Birgel, D., Feng, D., Roberts, H.H., Peckmann, J., 2011. Changing redox conditions at coldseeps as revealed by authigenic carbonates from Alaminos Canyon, northern Gulfof Mexico. Chemical Geology 285, 82–96.
Blumenberg, M., Seifert, R., Reitner, J., Pape, T., Michaelis, W., 2004. Membrane lipidpatterns typify distinct anaerobic methanotrophic consortia. Proceedings of theNational Academy of Sciences of the United States of America 101, 11111–11116.
Campbell, K.A., 2006. Hydrocarbon seep and hydrothermal vent paleoenvironments andpaleontology: past developments and future research directions. Palaeogeography,Palaeoclimatology, Palaeoecology 232, 362–407.
Campbell, K.A., Bottjer, D.J., 1995a. Brachiopods and chemosymbiotic bivalves in Phan-erozoic hydrothermal vent and cold seep environments. Geology 23, 321–324.
Campbell, K.A., Bottjer, D.J., 1995b. Peregrinella: an Early Cretaceous cold-seep-restricted brachiopod. Paleobiology 24, 461–478.
Campbell, K.A., Francis, D.A., Collins, M., Gregory, M.R., Nelson, C.S., Greinert, J., Aharon,P., 2008a. Hydrocarbon seep-carbonates of a Miocene forearc (East Coast Basin),North Island, New Zealand. Sedimentary Geology 204, 83–105.
Campbell, K.A., Peterson, D., Alfaro, A.C., 2008b. Two new species of Retiskenea?(Gastropoda: Neomphalidae) from Lower Cretaceous hydrocarbon seep-carbonatesof northern California. Journal of Paleontology 82, 140–153.
Cookson, I.C., Eisenack, A., 1970. Cretaceousmicroplankton from the Eucla Basin,WesternAustralia. Proceedings of the Royal Society of Victoria 83, 137–157.
Cooper, R.A.E., 2004. The New Zealand geological timescale. Institute of Geological andNuclear Sciences Monograph 22, 1–284.
Cosel, R.v, Marshall, B.A., 2003. Two new species of large mussels (Bivalvia: Mytilidae)from active submarine volcanoes and a cold seep off the eastern North Island ofNew Zealand, with description of a new genus. Nautilus 117, 31–46.
Cosel, R.v, Marshall, B.A., 2010. A new genus and species of large mussel (Bivalvia:Mytilidae) from the Kermadec Ridge. Tuhinga: Records of the Museum of NewZealand Te Papa Tongarewa 21, 59–73.
Crampton, J.S., Foote, M., Beu, A.G., Cooper, R.A., Matcham, I., Jones, C.M., Maxwell, P.A.,Marshall, B.A., 2006a. Second-order sequence stratigraphic controls on the qualityof the fossil record at an active margin: New Zealand Eocene to Recent shelfmolluscs. Palaios 21, 86–105.
Crampton, J.S., Schiøler, P., Roncaglia, L., 2006b. Detection of Late Cretaceous eustaticsignatures using quantitative biostratigraphy. Geological Society of America Bulletin118, 975–990.
Dando, P.R., Southward, A.J., 1986. Chemoautotrophy in bivalve molluscs of the genusThyasira. Journal of the marine biological Association of the United Kingdom 66,915–929.
Dando, P.R., Southward, A.J., Southward, E.C., 1986. Chemoautotrophic symbionts inthe gills of the bivalve mollusc Lucinoma borealis and the sediment chemistry ofits habitat. Proceedings of the Royal Society of London B 227, 227–247.
De Boever, E., Birgel, D., Muchez, P., Peckmann, J., Dimitrov, L., Swennen, R., 2011. Fab-ric and formation of grapestone concretions within an unusual ancient methaneseep system (Eocene, Bulgaria). Terra Nova 23, 56–61.
Dubilier, N., Bergin, C., Lott, C., 2008. Symbiotic diversity in marine animals: the art ofharnessing chemosynthesis. Nature Reviews Microbiology 6, 725–740.
Dufour, S.C., 2005. Gill anatomy and the evolution of symbiosis in the bivalve familyThyasiridae. The Biological Bulletin 208, 200–212.
Felbeck, H., 1981. Chemoautotrophic potential of the hydrothermal vent tube worm,Riftia pachyptila Jones (Vestimentifera). Science 213, 336–338.
Fensome, R.A., MacRae, R.A., Williams, G.L., 2008. DINOFLAJ2, Version 1. Data Series no. 1.American Association of Stratigraphic Palynologists. (http://dinoflaj.smu.ca/wiki/main_page).
Field, B.D., Uruski, C., Beu, A.G., Browne, G., Crampton, J.S., Funnell, R., Killops, S., Laird,M.G., Mazengarb, C., Morgans, H., Rait, G., Smale, D., Strong, P., 1997. Cretaceous–Cenozoic geology and petroleum systems of the East Coast region, New Zealand.Institute of Geological and Nuclear Sciences Monograph 13, 1–244 (+ enclosures).
Please cite this article as: Kiel, S., et al., Cretaceous methane-seepPalaeoclimatology, Palaeoecology (2012), http://dx.doi.org/10.1016/j.pala
Fisher, C.R., Childress, J.J., 1986. Translocation of fixed carbon from symbiotic bacteriato host tissues in the gutless bivalve Solemya reidi. Marine Biology 93, 59–68.
Gaillard, C., Rio, M., Rolin, Y., 1992. Fossil chemosynthetic communities related to vents orseeps in sedimentary basins: the pseudobioherms of southeastern France comparedto other world examples. Palaios 7, 451–465.
Gill, F.L., Harding, I.C., Little, C.T.S., Todd, J.A., 2005. Palaeogene and Neogene cold seepcommunities in Barbados, Trinidad and Venezuela: an overview. Palaeogeography,Palaeoclimatology, Palaeoecology 227, 191–209.
Hammer, Ø., Nakrem, H.A., Little, C.T.S., Hryniewicz, K., Sandy, M.R., Hurum, J.H.,Druckenmiller, P., Knutsen, E.M., Høyberget, M., 2011. Hydrocarbon seeps close tothe Jurassic–Cretaceous boundary, Svalbard. Palaeogeography, Palaeoclimatology,Palaeoecology 306, 15–26.
Helby, R., Morgan, R., Partridge, A.D., 1987. A palynological zonation of the AustralianMesozoic. Memoir of the Association of Australasian Palaeontologists 4, 1–94.
Henderson, R.A., 1970. Ammonoidea from the Mata Series (Santonian-Maastrichtian)of New Zealand. Special Papers in Palaeontology 6, 1–82.
Henderson, R.A., 1973. Clarene and Raukumara Series (Albian-'Santonian) Ammonoideafrom New Zealand. Journal of the Royal Society of New Zealand 3, 71–123.
Hikida, Y., Suzuki, S., Togo, Y., Ijiri, A., 2003. An exceptionallywell-preserved seep communityfrom the Cretaceous Yezo forearc basin in Hokkaido, northern Japan. PaleontologicalResearch 7, 329–342.
Hollis, C.J., Beu, A.G., Crampton, J.S., Jones, A.F., 2010. Calibration of the New ZealandCretaceous–Cenozoic timescale to GTS2004. GNS Science Report 2010/43, pp. 1–20.
Jacob, W., McClatchie, S., Probert, P.K., Hurst, R.J., 1998. Demersal fish assemblages offsouthern New Zealand in relation to depth and temperature. Deep-Sea ResearchI 45, 2119–2155.
Jenkins, R.G., Kaim, A., Hikida, Y., 2007. Antiquity of the substrate choice amongacmaeid limpets from the Late Cretaceous chemosynthesis-based communities.Acta Palaeontologica Polonica 52, 369–373.
Jenkins, R.G., Hikida, Y., Chikaraishi, Y., Ohkouchi, N., Tanabe, K., 2008. Microbially in-duced formation of ooid-like coated grains in the Late Cretaceous methane-seepdeposits of the Nakagawa area, Hokkaido, northern Japan. Island Arc 17, 261–269.
Jenkins, R.G., Kaim, A., Little, C.T.S., Iba, Y., Tanabe, K., Campbell, K.A., in press. Worldwidedistribution of modiomorphid bivalve genus Caspiconcha in late Mesozoic hydrocar-bon seeps. Acta Palaeontologica Polonica. http://dx.doi.org/10.4202/app.2011.0026.
Johnson, S.B., Warén, A., Lee, R.W., Kano, Y., Kaim, A., Davis, A., Strong, E.E., Vrijenhoek,R.C., 2010. Rubyspira, new genus and two new species of bone-eating deep-seasnails with ancient habits. The Biological Bulletin 219.
Kaim, A., Kelly, S.R.A., 2009. Mass occurrence of hokkaidoconchid gastropods in theUpper Jurassic methane seep carbonate from Alexander Island, Antarctica. AntarcticScience 21, 279–284.
Kaim, A., Jenkins, R.G., Warén, A., 2008. Provannid and provannid-like gastropods fromLate Cretaceous cold seeps of Hokkaido (Japan) and the fossil record of theProvannidae (Gastropoda: Abyssochrysoidea). Zoological Journal of the LinneanSociety 154, 421–436.
Kaim, A., Jenkins, R.G., Hikida, Y., 2009. Gastropods from Late Cretaceous hydrocarbonseep deposits in Omagari and Yasukawa, Nakagawa area, Hokkaido, Japan. ActaPalaeontologica Polonica 54, 463–690.
Kanie, Y., Sakai, T., 1997. Chemsynthetic thraciid bivalve Nipponothracia, gen. nov. fromthe Lower Cretaceous andMiddleMiocenemudstones in Japan. Venus 56, 205–220.
Kase, T., Kurihara, Y., Hagino, K., 2007. Middle Miocene chemosynthetic thraciidNipponothracia gigantea (Shikama, 1968) from central Japan is a large lucinidbivalve (Lucinoidea; Mollusca). Veliger 49, 294–302.
Kelly, S.R.A., Ditchfield, P.W., Doubleday, P.A., Marshall, J.D., 1995. An Upper Jurassicmethane-seep limestone from the Fossil Bluff Group forearc basin of AlexanderIsland, Antarctica. Journal of Sedimentary Research A65, 274–282.
Kelly, S.R.A., Blanc, E., Price, S.P., Withham, A.G., 2000. Early Cretaceous giant bivalvesfrom seep-related limestone mounds, Wollaston Forland, Northeast Greenland.In: Harper, E.M., Taylor, J.D., Crame, J.A. (Eds.), The evolutionary biology of theBivalvia. Geological Society of London, Special Publication, London, pp. 227–246.
Kiel, S., 2010. On the potential generality of depth-related ecologic structure in cold-seepcommunities: Cenozoic andMesozoic examples. Palaeogeography, Palaeoclimatology,Palaeoecology 295, 245–257.
Kiel, S., Little, C.T.S., 2006. Cold seep mollusks are older than the general marinemollusk fauna. Science 313, 1429–1431.
Kiel, S., Peckmann, J., 2008. Paleoecology and evolutionary significance of an EarlyCretaceous Peregrinella-dominated hydrocarbon-seep deposit on the CrimeanPeninsula. Palaios 23, 751–759.
Kiel, S., Amano, K., Jenkins, R.G., 2008a. Bivalves from Cretaceous cold-seep deposits onHokkaido, Japan. Acta Palaeontologica Polonica 53, 525–537.
Kiel, S., Campbell, K.A., Elder,W.P., Little, C.T.S., 2008b. Jurassic and Cretaceous gastropodsfrom hydrocarbon-seeps in forearc basin and accretionary prism settings, California.Acta Palaeontologica Polonica 53, 679–703.
Kiel, S., Campbell, K.A., Gaillard, C., 2010. New and little known mollusks from ancientchemosynthetic environments. Zootaxa 2390, 26–48.
Kiel, S., Wiese, F., Titus, A.L., 2012. Shallow-water methane-seep faunas in theCenomanian Western Interior Seaway: No evidence for onshore–offshore adapta-tions to deep-sea vents. Geology 40, 839–842.
Knittel, K., Boetius, A., 2009. Anaerobic oxidation of methane: progress with anunknown process. Annual Reviews of Microbiology 63, 311–334.
Kojima, S., Fujikura, K., Okutani, T., 2004. Multiple trans-Pacific migrations of deep-seavent/seep-endemic bivalves in the family Vesicomyidae. Molecular Phylogeneticsand Evolution 32, 396–406.
Kuechler, R.R., Birgel, D., Kiel, S., Freiwald, A., Goedert, J.L., Thiel, V., Peckmann, J., 2012.Miocene methane-derived carbonates from southwestern Washington, USA, and amodel for silicification at seeps. Lethaia 45, 259–273.
deposits from New Zealand and their fauna, Palaeogeography,eo.2012.10.033

18 S. Kiel et al. / Palaeogeography, Palaeoclimatology, Palaeoecology xxx (2012) xxx–xxx
Lewis, K.B., Marshall, B.A., 1996. Seep faunas and other indicators of methane-richdewatering on New Zealand convergent margins. New Zealand Journal of Geologyand Geophysics 39, 181–200.
Majima, R., Nobuhara, T., Kitazaki, T., 2005. Review of fossil chemosynthetic assem-blages in Japan. Palaeogeography, Palaeoclimatology, Palaeoecology 227, 86–123.
Mazengarb, C., Speden, I.G., 2000. Geology of the Raukumara area. Institute of Geologicaland Nuclear Sciences 1:250000 geological map 6, 1–60 + map.
Mazengarb, C., Francis, D.A., Moore, P.R., 1991. Sheet Y16, Tauwhareparae. Geologicalmap of New Zealand 1:50000. DSIR, Wellington.
McKay, A., 1887. On the geology of East Auckland and the northerndistrict ofHawke's Bay.New Zealand Geological Survey reports of geological explorations 18, 182–219.
Moore, P.R., 1988. Stratigraphy, composition and environment of deposition of theWhangai Formation and associated Late Cretaceous–Paleocene rocks, easternNorth Island, New Zealand. New Zealand Geological Survey Bulletin 100, 1–82.
Morgan, R., 1980. Palynostratigraphy of the Australian Early and Middle Cretaceous.Memoirs of the Geological Survey of New South Wales. Palaeontology 18, 1–153.
Niemann, H., Elvert, M., 2008. Diagnostic lipid biomarker and stable carbon isotope sig-natures of microbial communities mediating the anaerobic oxidation of methanewith sulphate. Organic Geochemistry 39, 1668–1677.
Nobuhara, T., Onda, D., Kikuchi, N., Kondo, Y., Matsubara, K., Amano, K., Jenkins, R.G.,Hikida, Y., Majima, R., 2008. Lithofacies and fossil assemblages of the Upper Creta-ceous Sada Limestone, Shimanto City, Kochi Prefecture, Shikoku, Japan. Fossils, ThePalaeontological Society of Japan 84, 47–60.
Oliver, P.G., Taylor, J.D., 2012. Bacterial symbiosis in the Nucinellidae (Bivalvia:Solemyida) with description of two new species. Journal of Molluscan Studies 78,81–91.
Olu, K., Cordes, E.E., Fisher, C.R., Brooks, J.M., Sibuet, M., Desbruyères, D., 2010. Biogeogra-phy and potential exchanges among the Atlantic equatorial belt cold-seep faunas.PLoS One 5, e11967.
Ongley, M., Macpherson, E.O., 1928. The geology of the Waiapu Subdivision,Raukumara Division. New Zealand Geological Survey Bulletin 30, 1–79 (+ maps).
Partridge, A.D., 2006. Jurassic - Early Cretaceous spore-pollen and dinocyst zonationsfor Australia. In: Monteil. E. (coord.), Australian Mesozoic and Cenozoic palynologyzonations-updated to the 2004 Geological Time Scale. Geoscience Australia Record2006/23 (wall chart), ISBN 1 921 236 05 1 (available at https://www.ga.gov.au/products/servlet/controller?event=GEOCAT_DETAILS&catno=64962).
Paull, C.K., Jull, A.J.T., Toolin, L.J., Linick, T., 1985. Stable isotope evidence for chemosyn-thesis in an abyssal seep community. Nature 317, 709–711.
Paxton, J.R., Cohen, D.M., 1999. Families Argentinidae, Microstomatidae, Bathylagidae,and Opisthoproctidae. In: Carpenter, K.E., Niem, V.H. (Eds.), FAO species identifica-tion guide for fishery purposes, vol. 3. FAO, Rome, pp. 1886–1887.
Peckmann, J., Thiel, V., 2004. Carbon cycling at ancient methane-seeps. Chemical Geology205, 443–467.
Peckmann, J., Campbell, K.A., Walliser, O.H., Reitner, J., 2007. A Late Devonianhydrocarbon-seep deposit dominated by dimerelloid brachiopods, Morocco. Palaios22, 114–122.
Peckmann, J., Birgel, D., Kiel, S., 2009. Molecular fossils reveal fluid composition andflow intensity at a Cretaceous seep. Geology 37, 847–850.
Peckmann, J., Kiel, S., Sandy, M.R., Taylor, D.G., Goedert, J.L., 2011. Mass occurrences ofthe brachiopod Halorella in Late Triassic methane-seep deposits, Eastern Oregon.Journal of Geology 119, 207–220.
Phillips, C.J., 1985. Upper Cretaceous and Tertiary geology of the upper Waitahaia River,Raukumara Peninsula, New Zealand. New Zealand Journal of Geology and Geophysics28, 595–607.
Pick, M.C., 1962. The stratigraphy, structure, and economic geology of the Cretaceous-Tertiary rocks of the Waiapu district, New Zealand. Todd Oil Exploration Co. Ltd.unpublished PhD thesis, University of Bristol, and unpublished open-file petroleumreport 608, Ministry of Economic Development, Wellington, New Zealand Thesis,Bristol, 532 pp.
Riding, J.B., Crame, J.A., 2002. Aptian to Coniacian (Early-Late Cretaceous) palynostratigraphyof the Gustav Group, James Ross Basin, Antarctica. Cretaceous Research 23, 739–760.
Roncaglia, L., Field, B.D., Raine, J.I., Schiøler, P., Wilson, G.J., 1999. Dinoflagellate biostra-tigraphy of Piripauan–Haumurian (Upper Cretaceous) sections from northeastSouth Island, New Zealand. Cretaceous Research 20, 271–314.
Saether, K.P., 2011. A taxonomic and palaeobiogeographic study of the fossil fauna ofMiocene hydrocarbon seep deposits, North Island, New Zealand. PhD Thesis,University of Auckland, Auckland, 537 pp.
Please cite this article as: Kiel, S., et al., Cretaceous methane-seepPalaeoclimatology, Palaeoecology (2012), http://dx.doi.org/10.1016/j.pala
Saether, K.P., Little, C.T.S., Campbell, K.A., 2010a. A new fossil provannid gastropodfrom Miocene hydrocarbon seep deposits, East Coast Basin, North Island, NewZealand. Acta Palaeontologica Polonica 55, 507–517.
Saether, K.P., Little, C.T.S., Campbell, K.A., Marshall, B.A., Collins, M., Alfaro, A.C., 2010b.New fossil mussels (Bivalvia: Mytilidae) from Miocene hydrocarbon seep deposits,North Island, New Zealand, with general remarks on vent and seep mussels.Zootaxa 2577, 1–45.
Saether, K.P., Little, C.T.S., Marshall, B.A., Campbell, K.A., 2012. New fossil limpet(Patellogastropoda: Pectinodontidae) from Miocene hydrocarbon seep deposits,East Coast Basin, North Island, New Zealand: systematics and palaeoecology.Molluscan Research 32, 1–15.
Sahling, H., Galkin, S.V., Salyuk, A., Greinert, J., Foerstel, H., Piepenburg, D., Suess, E., 2003.Depth-related structure and ecological significance of cold-seep communities—a casestudy from the Sea of Okhotsk. Deep-sea Research I 50, 1391–1409.
Sandy, M.R., 1995. A review of some Palaeozoic and Mesozoic brachiopods as membersof cold seep chemosynthetic communities: “unusual” palaeoecology and anoma-lous palaeobiogeographic pattern explained. Földtani Közlöny 125, 241–258.
Saul, L.R., Squires, R.L., Goedert, J.L., 1996. A new genus of cryptic lucinid? Bivalve fromEocene cold seeps and turbidite-influenced mudstone, western Washington. Jour-nal of Paleontology 70, 788–794.
Schiøler, P., Crampton, J.S., King, P., 2000. Palynostratigraphic analysis of a measuredsection through the Karekare, Owhena and Whangai formations (Upper Cretaceous)at Waitahaia River-“Owhena Stream” (Raukumara Peninsula). Institute of Geological& Nuclear Sciences Science Report 31, 1–24.
Sibuet, M., Olu, K., 1998. Biogeography, biodiversity and fluid dependence of deep-seacold-seep communities at active and passive margins. Deep-Sea Research II 45,517–567.
Spencer, H.G., Marshall, B.A., Maxwell, P.A., Grant-Mackie, J.A., Stilwell, J.D., Willan, R.C.,Campbell, H.J., Crampton, J.S., Henderson, R.A., Bradshaw, M.A., Waterhouse, J.B.,Pojeta, J., 2009. Phylum Mollusca: chitons, clams, tusk shells, squids and kin. In:Gordon, D.P. (Ed.), New Zealand inventory of biodiversity. 1. Kingdom Animalia:Radiata, Lophotrochozoa, Deuterostomia. Canterbury University Press, Christchurch,pp. 161–254.
Stanton, T.W., 1895. Contributions to the Cretaceous paleontology of the Pacific coast:the fauna of the Knoxville beds. United States Geological Survey Bulletin 133,1–132.
Stilwell, J.D., 2003. Diversity, distribution, and evolutionary history of the Austral, LateCretaceous to Eocene, gastropod Struthioptera (Stromboidea: Aporrhaidae). Journal ofthe Royal Society of New Zealand 33, 315–340.
Taviani, M., 1994. The “calcari a Lucina” macrofauna reconsidered: deep-sea faunaloases from Miocene-age cold vents in the Romagna Apennine, Italy. Geo-MarineLetters 14, 185–191.
Taylor, J.D., Glover, E.A., 2006. Lucinidae (Bivalvia)—the most diverse group ofchemosymbiotic molluscs. Zoological Journal of the Linnean Society 148, 421–438.
Taylor, J.D., Glover, E.A., 2010. Chemosymbiotic bivalves. In: Kiel, S. (Ed.), The Vent andSeep Biota. : Topics in Geobiology. Springer, Heidelberg, pp. 107–136.
Teichert, B.M.A., Bohrmann, G., Suess, E., 2005. Chemoherms on Hydrate Ridge—uniquemicrobially-mediated carbonate build-ups growing into the water column.Palaeogeography, Palaeoclimatology, Palaeoecology 227, 67–85.
Thiel, V., Peckmann, J., Richnow, H.H., Luth, U., Reitner, J., Michaelis, W., 2001a. Molec-ular signals for anaerobic methane oxidation in Black Sea seep carbonates and amicrobial mat. Marine Chemistry 73, 97–112.
Thiel, V., Peckmann, J., Schmale, O., Reitner, J., Michaelis, W., 2001b. A new straight-chain hydrocarbon biomarker associated with anaerobic methane cycling. OrganicGeochemistry 32, 1019–1023.
Thieuloy, J.-P., 1972. Biostratigraphie des lentilles a peregrinelles (Brachiopodes) del'Hauterivien de Rottier (Drome, France). Geobios 5, 5–53.
Van Dover, C.L., German, C.R., Speer, K.G., Parson, L.M., Vrijenhoek, R.C., 2002. Evolutionand biogeography of deep-sea vent and seep invertebrates. Science 295,1253–1257.
Wilson, G.J., 1984. New Zealand Late Jurassic to Eocene dinoflagellate biostratigraphy—a summary. Newsletters on Stratigraphy 13, 104–117.
deposits from New Zealand and their fauna, Palaeogeography,eo.2012.10.033




















