
Toll-like receptor 4 ligation enforces tolerogenic properties of oral mucosal Langerhans cells
Upload
independentCategory
view
2download
0
Corneal Langerhans Cell Dynamics After Herpes SimplexVirus Reactivation
Judith Kelvin Miller* Keith A. Laycock,* Melinda M. Nash,* and Jay S. Pepose*"f
Purpose. The authors investigated the progressive changes in the distribution of corneal Lan-gerhans cells (LC) after reactivation of latent herpes simplex virus type I (HSV-1) in mice.
Methods. After corneal inoculation of National Institutes of Health inbred mice with HSV-1and the establishment of latency, viral reactivation was induced by irradiating the ocular sur-face with 250 mj/cm2 of ultraviolet B (UV-B) light.
Results. Subsequent viral replication in the cornea was followed by the migration of the LCtoward the paracentral and central corneal epithelium. These areas are normally devoid of LC.The number of LC in the paracentral and central regions of the eye reached a peak at day 14post-UV-B irradiation. After UV-B irradiation of mice latently infected with HSV-1, the devel-opment of corneal stromal opacification and neovascularization closely followed the migrationof LC toward the central cornea and paralleled the influx of T-cells into the corneal stroma.This pattern was not observed in irradiated uninfected mice.
Conclusions. LC migrate centrally in the corneal epithelium after viral reactivation. There is adirect correlation between the number of LC in the cornea and the degree of persistentstromal opacification. Invest Ophthalmol Vis Sci. 1993;34:2282-2290.
An many tissues, such as the skin and mucous mem-branes, Langerhans cells (LC) appear to be an integralcomponent of the regional immune response.1"5 Theantigen-presenting capabilities6"8 of these migratory,bone marrow-derived,1 la-expressing cells19"12 placethem at a critical outpost to generate local immunityagainst microbial pathogens.
The distribution of LC appears to be compart-mentally localized within specific regions of the ocularsurface. This localization may contribute to some ofthe unique immunologic features of the eye. OcularLC generally reside in the conjunctiva and corneallimbus.13'14 Although the central cornea is normallydevoid of LC, they can be detected in large numbers in
From the Departments of * Ophthalmology and Visual Sciences and f Pathology,Washington University School of Medicine, St. Louis, Missouri.Supported in part fry grants from the Monsanto Corporation, St. Louis, MO (JSP),Fight for Sight C.A90031 Baltimore, MO (JKM), ami Research to Preventlilindness. Inc., New York, New York.Submitted for publication: November 22, 1991; accepted August 25, 1992.Proprietary interest category: N.Reprint requests: Jay S. Pepose, M.D., Ph.D., Department of Ophthalmology andVisual Sciences, Washington University School of Medicine, 660 South Euclid,St. Louis, MO 63110.
all regions of the corneal epithelium after a variety ofinsults, including acute infection with herpes simplexvirus I (HSV-1 ).15"20
The exquisite antigen-presenting properties of LCimplicate them as central in the activation of T-lym-phocytes.121"24 In the eye, which depends on transpar-ent media for visual function, a side effect of the T-cellactivation, proliferation, and cytokine release may bethe opacification of clear corneal tissue.25"30 Damagefrom the inflammatory reactions in the corneal stromainduced by recurrent HSV-1 exposure can lead to irre-versible tissue necrosis, scarring, and neovasculariza-tion.31 The ocular damage observed during HSV-1 re-crudescence appears to arise predominantly from theresponse of the immune system to the presence of thereactivated virus, rather than from viral cytolysis it-se|j-28,29,32
Whereas previous studies have quantified cornealLC after primary ocular infection with HSV-1, we initi-ated experiments to investigate the progressivechanges in the distribution of LC after reactivation oflatent HSV-1. Our interest in reactivation (as opposed
2282:.stig;iiive Ophthalmology & Visual Scienc
Copyright © Association for Research i1993, Vol. 34. No. 7
n and Ophthalmology

Corneal Langerhans Cells After HSV-1 Reactivation 2283
to primary infection) reflects our effort to mimic therelevant clinical situation in which the majority ofsight-threatening ocular disease is the result of viralrecrudescence not subclinical primary infection. Un-derstanding the influence of LC on the manifestationsand severity of recurrent disease will help us exploreways to modulate the immune system to provide a de-fense against HSV-1 while ameliorating the destruc-tive consequences of its responses.
For these studies, we modified a murine ocularmodel of herpetic reactivation3334 recently character-ized in our laboratory,35 in which HSV-1 is isolatedfrom the ocular surface of infected National Institutesof Health inbred mice after ultraviolet B (UV-B) irra-diation.35 In our previous studies, topical corticoste-roids were applied to the ocular surface, starting 2days after irradiation, to increase the yield and detec-tion of infectious virus during the postirradiation pe-riod.35 Steroid treatment was excluded from thecurrent study to avoid any complicating effects thatthe steroids might have on the LC.14 As a conse-quence, the amount of viral shedding at the ocularsurface was reduced and might be below detectablelevels in some animals, as assessed by once-a-day swab-bing. This has been evidenced in our previous work35
by the detection of viral antigens in the basal cornealepithelium, but not in the superficial corneal cells, inselected animals that had negative swab cultures at thetime they were killed. Because of potential limitationsin relying on swab culture to identify all mice withrecurrent keratitis, we present a population study inwhich we compared corneal LC migration in an unin-fected group and a latently HSV-1-infected group ofmice after exposure of both groups to UV-B light.Data from the latter population were substratified,and the results from mice with virus positive eye swabsare also presented separately.
MATERIALS AND METHODS
Experimental DesignMice. Female National Institutes of Health inbred
mice (Harlan Sprague Dawley, Indianapolis, IN) wereinoculated with virus at an average age of 4 weeks. Thefollowing four groups were involved in this study:
1. The test group consisted of 90 mice that werelatently infected with virus and subsequently ex-posed to UV-B light.
2. The UV-B control group consisted of 43 micethat were not infected with virus but were ex-posed to UV-B light.
3. The latent control group consisted of 11 micethat were latently infected with virus but werenot exposed to UV-B light.
4. The untreated control group consisted of 16mice that were neither infected with virus norexposed to UV-B light.
The experiments conformed to the ARVO Resolu-tion on the Use of Animals in Research.
Virus. HSV-1, the McKrae strain, was used for theinoculations. The virus was from a plaque-purifiedstock that had been grown and assayed on Africangreen monkey kidney (Vero) cells in minimum essen-tial medium with Earle's salts and L-glutamine (catalog-no. 320-1095PK, Gibco, Grand Island, NY) contain-ing 5% fetal bovine serum (Biocell, Carson, CA), 100units/ml of penicillin, and 100 Mg/m' of streptomycin.
Anesthesia. The mice were anesthetized with a com-bination of 10 mg/kg of xylazine and 70 mg/kg ofketamine HC1.
Inoculation. After scarification of the cornea of theright eye in a grid pattern with a No. 15 scalpel blade, a5-/xl drop of medium containing 1 X 106 plaque-form-ing units of HSV-1 was placed onto the eye. While theanimal was anesthetized, the uninoculated left eye waskept moist with a 5-̂ tl drop of Hanks balanced saltsolution (catalogno. 21-022-LM, Mediatech, Washing-ton, DC).
Antiserum. To limit the extent of primary HSV-1infection, the mice were inoculated intraperitoneallyat the time of viral inoculation with 1 ml of pooledhuman serum (catalog no. SI, Chemicon, Temecula,CA) containing antibodies to HSV-1 (50% effectivedose, = 1/640).
Establishment of Latency. The mice were housed(five to ten animals per cage) and fed and watered adlibitum for 46 days after the viral inoculation.
UV-B Irradiation. Forty-six days after the viral inoc-ulation, the right eye of each mouse in which HSV-1was to be reactivated and that of each mouse thatserved as an uninfected UV-B-irradiated control wasexposed to 250 mj/cm2 of UV-B light (55 sec of 4.5mW/cm2 at 302 nm on a UVP Chromato-Vue transil-luminator (model TM-20, San Gabriel, CA).
Data Acquisition
Shedding of Virus. Surgical spears (catalog no.8680, Xomed-Treace, Jacksonville, FL) saturated withminimum essential medium with Earle's salts and L-glutamine were used to swab the inoculated eyes of allbut those of the untreated control group on day 3postinoculation and to swab the test mice and UV-Bcontrols once daily on days 1-11 post-UV-B exposure.The swab material was cultured on Vero cells, whichwere examined for virally induced cytopathic effects.Neither topical nor systemic steroids were used in thisreactivation model.
Clinical Observations. The eyes were observedthrough a binocular dissecting microscope and a slit-

2284 Investigative Ophthalmology & Visual Science, June 1993, Vol. 34, No. 7
lamp biomicroscope at prescribed intervals and in amasked fashion. Detailed additional analysis was ac-complished by reviewing the slit-lamp photographs.Stromal opacification was rated on a scale of 0-4,where 0 indicated a clear stroma; 1, a mild stromalopacification; 2, a moderate opacity with discernibleiris features; 3, a dense opacity with loss of defined irisdetail; and 4, a total opacity with no posterior view.Corneal neovascularization, epithelial dendrites andlesions, and associated ocular pathologic conditionswere also noted.
Tissue Preparation. On days 0, 4, 7, 11, 14, 21, and35 post-UV-B irradiation, 6-18 test animals, 3-7 un-infected UV-B controls, and 2-4 uninfected non-UV-B-irradiated controls were killed. On days equivalentto days 35 and 43 post-UV-B irradiation, five or sixnonirradiated latent controls were killed. Their eyeswere enucleated, and the intact globes were placedinto Hanks balanced salt solution. After excision, thecorneas were flattened with the aid of four peripheralrelaxing incisions. The tissue was placed epithelial-sideup into an ethylenediaminetetraacetic acid (EDTA) so-lution (6.83 g of NaCl, 0.2 g of KC1, 1.14 g ofNa2HPO4, 0.2 gof KH2PO4, 0.12 ml of 1% phenol red[optional], 7.6 g of Na4EDTA per 1 1, pH 7.2-7.4),according to published methods.3637 After a 2-hr in-cubation at 37°C in the EDTA solution, the corneaswere transferred to a phosphate-buffered saline solu-tion (PBS, 10 mM sodium phosphate in 0.9% saline)until each was further processed. This step was takento avoid leaving the tissue exposed to EDTA longerthan 2 hr. Extended exposure to EDTA increases thefragility and fragmentation of the epithelium. Eachcornea was placed in a drop of PBS on a glass slide.Jeweler's forceps were used to peel and tease the epi-thelium from the stromal layer. The epithelium wasthen flattened onto the glass slide, the excess PBS wasdrained, and a Pasteur pipette drop of acetone wasplaced onto the epithelium and allowed to dry. Theslides were then immersed in cold acetone for 30 minand then air dried.
Immunostaining. PBS was used for all washing andsoaking. All incubations were performed in a coveredslide tray (catalog no. M6304, Baxter, McGaw Park,IL) with 50-100 /il of a given solution.
After encircling the tissue with a water-imperme-able substance (PAP pen, Research Products Interna-tional, Mt. Prospect, IL), the epithelium was coveredwith the primary antibody, rat anti-mouse la (a 1:100dilution in PBS of catalog no. 1199293, BoehringerMannheim, Indianapolis, IN), and the slides were in-cubated at 4CC overnight (16 hr).
After washing off the primary antibody, soakingthe slide for 4 min, and draining off the excess PBS,the secondary antibody, biotinylated rabbit anti-rat (a1:100 dilution in PBS of catalog no. BA4000, Vector,
Burlingame, CA) was applied. The slides were thenincubated for 2 hr at room temp. The secondaryantibody was then washed off, and the slides weresoaked 4 min.
The slides were then processed with the avidin-biotin complex procedure of the Vectastain Peroxi-dase Standard PK4000 kit (Vector). This incubationwas performed at room temperature for 45 min and,after washing, was followed by a 13-min room tempera-ture incubation in 3-amino-9-ethylcarbazole solution(17.8 mmol/1 sodium acetate, pH 5.1, 0.02% H2O2,0.85 mol/1 3-amino-9-ethylcarbazole [catalog no.A5754, Sigma, St. Louis, MO], made as 7.94 mol/1 indimethyl sulfoxide (catalog no. D5879, Sigma). Afterremoval of the 3-amino-9-ethylcarbazole, Mayer's he-matoxylin stain (catalog no. MHS-32, Sigma) was ap-plied for 1 min at room temperature. The counterstainwas washed off, and the slides were allowed to dry. Adrop of Gelvatol (20 g of polyvinyl alcohol [catalog no.P8136, Sigma] in 80 ml of PBS and 40 ml of glycerol)was placed on top of the epithelium, and a cover slipwas placed over the Gelvatol.
Detection and Quantitation of LC. The slides wereviewed through a Zeiss (Thornwood, NY) binocularmicroscope. Cells identified morphologically as LCwere counted with the aid of a 100-square grid thatencompassed 0.16 mm2 at 250X power. The corneawas partitioned into three concentric circles by divid-ing the overall radius of the corneal flat mount intothirds. The number of LC per 100-square grid werecounted in the entire center region and at four pointsaround the paracentral and peripheral regions. Thedata were then converted to the average number ofcells per millimeter squared for a given region of theepithelium.
Immunostaining for Macrophages, T-Cells, and HSV-1 Antigens. Cryostat-cut 8-fim sections of corneal tissuewere fixed in chilled acetone and stained with mono-clonal antibodies against macrophage (a 1:30 dilutionin PBS of Anti-Mac-1, catalog no. 1118 129, Boeh-ringer Mannheim) or T-cells (a 1:30 dilution in PBS ofAnti-Thy 1.2, catalog no. 1331, Becton Dickinson, SanJose, CA). Formaldehyde-fixed, paraffin-embeddedtissues were cut to an 8-jum thickness and analyzedwith rabbit polyclonal antibodies against HSV anti-gens (a 1:50 dilution, catalog no. AXL-237, AccurateChemical and Scientific, Westbury, NY). The avidin-biotin complex kit described earlier was used for theseanalyses.
Photography. Photographs were taken through aZeiss Axioskop with an MCI00 attachment. ASA 400,35-mm black-and-white film (XP1, ILFORD, Chicago,IL) or ASA 200, 35-mm color film (Kodacolor Gold,Eastman Kodak, Rochester, NY) was used for the origi-nal LC photographs. Technical Pan black-and-whitefilm (catalog no. 129 7563, Eastman Kodak), shot at

Corneal Langerhans Cells After HSV-1 Reactivation 2285
V
FIGURE I. Photomicrographs illustrating the morphologic characteristics of LC detected inthe corneal epithelium with antibodies to la antigen. (A) This corneal epithelium is from theleft eye of a latent control mouse (HSV-1 infected by the right cornea, non-UV-B irradiated)and was taken at a time in our experiment equivalent to 43 days post-UV-B-irradiation(X250). (B) This corneal epithelium is from a test mouse (HSV-I infected, UV-B irradiated)and was taken ] I days posl-UV-B irradiation (X250).
ASA 200, was used for the macrophage and T-cellphotographs. Filters, times, and F-stops were varied tooptimize the contrast in each sample.
Statistics. The averages, standard errors of themeans, two-tailed t-tests, and regression analyses werecalculated on an Apple Macintosh (Cupertino, CA)with Microsoft Excel (Bellevue, WA) and StatView IIand plotted with Cricket Graph 1.2 and Apple Mac-Draw II (Malvern, PA).
Data Presentation. The data in Figures 2, 3, and 5are presented as the means ± the standard errors ofthe means.
RESULTS
Identification of LCLC were easily identified by the reddish-brown reac-tion product formed after incubation with the lamonoclonal antibody and immunoperoxidase develop-ment. The cells contained scanty amounts of cyto-plasm and had many delicate dendritic processes thatwere seen interdigitating between neighboring epithe-lial cells (Fig. 1).
Control Animals: Uninfected, UV-B IrradiatedThe changes observed in the distribution of cornealLC after exposure to the UV-B used to initiate HSV-1reactivation in our murine model were transient (Fig.2). The initial trend in the uninfected mice was a slightdecrease in the number of LC in the periphery of thecorneas. LC were virtually absent in the paracentraland central regions of the cornea during this same
period. After the early decrease in the number of LCin the peripheral cornea, the number of LC increasedbetween days 7-21 in the periphery, paracentral re-gion, and center of the corneas of these uninfectedUV-B-irradiated animals, but to a lesser degree than inthe infected animals (Figs. 2, 3). The number of LC inthe central and paracentral regions decreased afterday 21, approaching baseline levels. The number ofLC in the periphery on day 35 was greater than that onday 0.
300UNINFECTED MICE
10 20 30DAY POST-UV IRRADIATION
FIGURE 2. Uninfected, UV-B-irradiated mice. These data il-lustrate the progressive changes in the number of LC in theperipheral, paracentral, and central regions of the cornealepithelium as the time post-UV-B irradiation increases. Theyare presented as the means accompanied by the standarderrors of the means.
2286 Investigative Ophthalmology & Visual Science, June 1993, Vol. 34, No. 7
LATENTLY INFECTED MICE
300
LANGERHANSCELLS,
PER mm2
10 20 30
DAY POST-UV IRRADIATION
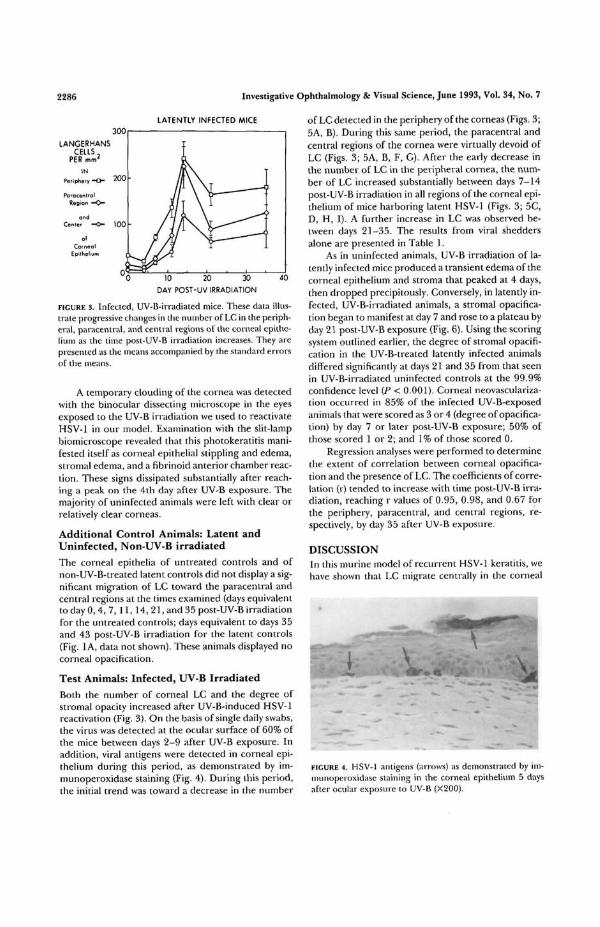
FIGURE 3. Infected, UV-B-irradiated mice. These data illus-trate progressive changes in the number of LC in the periph-eral, paracentral, and central regions of the corneal epithe-lium as the time post-UV-B irradiation increases. They arepresented as the means accompanied by the standard errorsof the means.
A temporary clouding of the cornea was detectedwith the binocular dissecting microscope in the eyesexposed to the UV-B irradiation we used to reactivateHSV-1 in our model. Examination with the slit-lampbiomicroscope revealed that this photokeratitis mani-fested itself as corneal epithelial stippling and edema,stromal edema, and a fibrinoid anterior chamber reac-tion. These signs dissipated substantially after reach-ing a peak on the 4th day after UV-B exposure. Themajority of uninfected animals were left with clear orrelatively clear corneas.
Additional Control Animals: Latent andUninfected, Non-UV-B irradiatedThe corneal epithelia of untreated controls and ofnon-UV-B-treated latent controls did not display a sig-nificant migration of LC toward the paracentral andcentral regions at the times examined (days equivalenttoday 0,4, 7, 11, 14, 21, and 35 post-UV-B irradiationfor the untreated controls; days equivalent to days 35and 43 post-UV-B irradiation for the latent controls(Fig. 1A, data not shown). These animals displayed nocorneal opacification.
Test Animals: Infected, UV-B Irradiated
Both the number of corneal LC and the degree ofstromal opacity increased after UV-B-induced HSV-1reactivation (Fig. 3). On the basis of single daily swabs,the virus was detected at the ocular surface of 60% ofthe mice between days 2-9 after UV-B exposure. Inaddition, viral antigens were detected in corneal epi-thelium during this period, as demonstrated by im-munoperoxidase staining (Fig. 4). During this period,the initial trend was toward a decrease in the number
of LC detected in the periphery of the corneas (Figs. 3;5A, B). During this same peiiod, the paracentral andcentral regions of the cornea were virtually devoid ofLC (Figs. 3; 5A, B, F, G). After the early decrease inthe number of LC in the peripheral cornea, the num-ber of LC increased substantially between days 7-14post-UV-B irradiation in all regions of the corneal epi-thelium of mice harboring latent HSV-1 (Figs. 3; 5C,D, H, I). A further increase in LC was observed be-tween days 21-35. The results from viral sheddersalone are presented in Table 1.
As in uninfected animals, UV-B irradiation of la-tently infected mice produced a transient edema of thecorneal epithelium and stroma that peaked at 4 days,then dropped precipitously. Conversely, in latently in-fected, UV-B-irradiated animals, a stromal opacifica-tion began to manifest at day 7 and rose to a plateau byday 21 post-UV-B exposure (Fig. 6). Using the scoringsystem outlined earlier, the degree of stromal opacifi-cation in the UV-B-treated latently infected animalsdiffered significantly at days 21 and 35 from that seenin UV-B-irradiated uninfected controls at the 99.9%confidence level (P < 0.001). Corneal neovasculariza-tion occurred in 85% of the infected UV-B-exposedanimals that were scored as 3 or 4 (degree of opacifica-tion) by day 7 or later post-UV-B exposure; 50% ofthose scored 1 or 2; and 1% of those scored 0.
Regression analyses were performed to determinethe extent of correlation between corneal opacifica-tion and the presence of LC. The coefficients of corre-lation (r) tended to increase with time post-UV-B irra-diation, reaching r values of 0.95, 0.98, and 0.67 forthe periphery, paracentral, and central regions, re-spectively, by day 35 after UV-B exposure.
DISCUSSIONIn this inurine model of recurrent HSV-1 keratitis, wehave shown that LC migrate centrally in the corneal
FIGURE 4. HSV-1 antigens (arrows) as demonstrated by im-munoperoxidase staining in the corneal epithelium 5 daysafter ocular exposure to UV-B (X200).

Corneal Langerhans Cells After HSV-1 Reactivation
r»AV n
2287
DAYd DAY 11 DAY 14B C D
DAY 3S
FIGURE 5. Photomicrographs illustrating the progressive changes in the number of LC in thecorneal epithelium after the UV-B irradiation of the corneas of mice harboring latent HSV-1(X125). The magnification is lower than that in Figure 1 (125X versus 250X) to allow for awider field of view. A-E show the peripheral and paracentral regions of the corneal epithe-lium on days 0, 4, 11, 14, and 35 post-UV-B irradiation, respectively. F-J show the centralregion of the corneal epithelium on days 0, 4, 11, 14, and 35 post-UV-B-irradiation, respec-tively.
epithelium after viral reactivation. The virally inducedmigration of LC temporally precedes the developmentof herpetic stromal disease (Figs. 3, 6). These findingsare consistent with those of previous studies of pri-mary infection15'26 that demonstrated that an increasein the number of LC in the more central regions of thecornea during the time of primary HSV-1 infectioncan promote stromal disease.
The intensity of the UV-B used in our model maybe expected to damage functionally the LC it en-counters38 and also may underlie the initial drop inperipheral LC in both the uninfected and latently in-fected groups. However, the migratory LC that infil-trate the cornea in substantial numbers by day 7 post-UV-B-induced reactivation are unlikely to be the samecells that were exposed to the UV-B. Those LC we
observe at this later time are, therefore, potentiallycapable of presenting HSV-1 antigens to incoming T-cells.8
The central migration of LC demonstrated afterHSV-1 reactivation shares some common featureswith the changes in corneal LC distribution observedafter primary HSV-1 infection.15-17182026 However, ina recurrent infection, the migration of LC toward thecentral cornea is delayed in comparison with that ob-served during primary infection. The later time pointfor the peak in the number of LC observed in thisstudy of reactivated virus (day 14) compared with thatobserved in earlier experiments of primary infection(days 7-11) may represent the lag time between thereactivation in the trigeminal ganglia and the appear-ance of the replicating virus at the ocular surface. Al-
TABLE l. Langerhans Cells in the Cornea of Animalswith Virus-Positive Eye Swabs
Day Post-UVReactivation
047
11142135
Peripheral
33.6 ± 3.428.9 ± 7.069.7+ 8.0
137.6 ± 26.0232.2 ±59.1152.2 ±41.8208.7 ± 64.0
Region of Cornea
Paracentral
15.6 ± 4.17.8 ± 3.1
17.7 ± 2.971.7 ±20.0
179.5 ±67.990.6 ±31.3
156.2 ± 69.0
Central
5.7 d0.0 d
14.6 d47.2 d64.6 d79.7-
110.5 :
t 2.7b 0.0b 5.5b 18.5b30.0b70.3b49.4
Dala are expressed as LC per square millimeter ± SEM.

2288 Investigative Ophthalmology & Visual Science, June 1993, Vol. 34, No. 7
DEGREE OFSTROMAL
OPACIFICATION
0= CLEAR STROMAi> TOTAL OPACITY
10 20 30
DAYS POST-UV IRRADIATION
FIGURE 6. Corneal stromal opacification after UV-B irradia-tion of mice latently infected with HSV-1. Stromal opacifica-tion was rated on a scale of 0-4, in which 0 indicated a clearstroma; 1, a mild stromal opacification; 2, a moderate opac-ity with discernible iris features; 3, a dense opacity with lossof defined iris detail; and 4, total opacity with no posteriorview. At days 21 and 35, the two groups differed signifi-cantly at the 99.9% confidence level (P < 0.001).
ternatively, the later peak in LC migration observedafter reactivation (as opposed to primary infection)may reflect transient depletion of LC from the ocularsurface in the period immediately after UV-B, requir-ing additional time to repopulate the conjunctiva andlimbus before subsequent central migration canoccur.
In preliminary experiments in this laboratory withHSV-1 reactivation in mice, we have shown the infil-tration into the cornea of different classes of T-cells topeak at day 10 or later after viral reactivation (Fig. 7)and a delayed-type hypersensitivity response to HSV-1antigens to rise sharply until day 10 and then to con-tinue to increase more slowly (data not shown). Thesepilot studies have also demonstrated that a large num-ber of macrophages are present in the stroma by day 3postreactivation (Fig. 8), but few macrophages everenter the epithelium. These findings, taken in con-junction with the data we have presented here, pro-duce a sequence of observations in which neuronalreactivation of virus, retrograde transport, and subse-quent corneal replication of HSV-1 is followed, first,by the migration of macrophages into the stroma (day3) and, then, by the movement of LC toward the cen-tral regions of the corneal epithelium (beginning atday 7 and peaking at day 14, Fig. 3). These events areaccompanied and followed, first, by infiltration of T-lymphocytes into the cornea (beginning at day 7 andpeaking at day 10 or later) and, then, by a persistentcorneal opacification (Fig. 6) and neovascularization(day 21 and beyond).
It is our working hypothesis that reactivated virusreplicating in the cornea (Fig. 4 shows the detection ofHSV antigens) induces macrophages39 and corneal epi-
FIGURE 7. Immuncytochemical staining of corneal T-cell in-filtrates detected 10 days after UV-B irradiation of mice la-tently infected with HSV-1 (Thy 1.2 monoclonal antibody,X250).
thelium40 to release cytokines (eg, interleukin-1 andtumor necrosis factor-a), which in turn, attract LC tothe epithelium, either directly or through other down-stream cytokines such as interleukin-8 or monocytechemotactic activation factor.41"43 Antigen-presentingcorneal LC and stromal macrophages, perhapsthrough the release of their own products30 or in con-junction with interleukin-8,43 induce T-cells (and, pos-sibly, additional LC) to infiltrate the cornea. The infil-tration of macrophages and T-cells and/or the releaseof their products results in corneal opacification. Arepetition of this sequence as a result of multiple viralrecurrences further increases stromal opacification.
FIGURE 8. Immuncylochemical staining of corneal macro-phage infiltrates detected 3 days after UV-B irradiation ofmice latently infected with HSV-1 (Mac-1 monoclonal anti-body, X 250).

Corneal Langerhans Cells After HSV-1 Reactivation 2289
In addition, both interleukin-1 and -2 have been impli-cated in the production of corneal neovasculariza-tion,44'45 along with other components of the inflam-matory response.4647 Ongoing experiments designedto test the various components of this hypothesis maylead to therapeutic modalities to modulate the damag-ing ocular immune response to HSV-1 and to a betterunderstanding of existing treatment regimens.
In summary, we found a correlation between thenumber of LC present in the cornea and the degree ofpersistent stromal opacification in our murine modelof herpetic reactivation. To our knowledge, this is thefirst reported study of corneal LC dynamics afterHSV-1 reactivation. Viral reactivation, as opposed toprimary infection, has considerable clinical signifi-cance. In humans, it is, generally, reactivation of HSV-1, rather than primary infection, that eventually leadsto visually significant corneal stromal disease.48 Wehave demonstrated that our murine reactivationmodel mimics this most significant component of thehuman ocular disease.
Key Words
Langerhans cells, herpes simplex virus type 1, viral reactiva-tion, mouse model of reactivation, corneal epithelium
Acknowledgments
The authors thank Stephen S. Yoon and Danna Tauber fortechnical assistance, Carla Terrell for assistance in prepara-tion of the manuscript, Drs. Thomas S. Kupper and ThomasA. Ferguson for helpful discussion, and Drs. Donald S. Levyand Stephanie A. Skolik for technical advice.
References
1. Stingl G, Tamaki K, Katz SI. Origin and function ofepidermal Langerhans cells. Immunol Rev. 1980;53:149-174.
2. Sprecher E, Becker Y. Langerhans cell density andactivity in mouse skin and lymph nodes affect herpessimplex type 1 (HSV-1) pathogenicity. Arch Virol.1989;107:191-205.
3. Hayashi Y, Aurelian L. Immunity to herpes simplexvirus type 2: Viral antigen-presenting capacity of epi-dermal cells and its impairment by ultraviolet irradia-tion./ Immunol. 1986; 136:1087-1092.
4. Braathen LR, Berle E, Mobech-Hanssen U, ThorsbyE. Studies on human epidermal Langerhans cells: II.Activation of human T lymphocytes to herpes simplexvirus. Ada Dermatovener (Stockh). J 980; 60:381-387.
5. Silberberg I, Baer RL, Rosenthal SA, Thorbecke GJ,Berezowsky V. Langerhans cells in skin, lymphatic-likevessels and draining lymph nodes in allergic contactsensitivity (abstract). Clin Res. 1975;23:231 A.
6. Braathen LR, Thorsby E. Studies on human epider-mal Langerhans cells: Allo-activating and antigen-pre-senting capacity. ScaiulJ Immunol. 1980; 11:401-408.
7. Rasanen L, Lehio M, Reunala T, Jansen C, LehtinenM, Leinikki P. Langerhans cell- and T-lympbocyte
functions in patients with atopic dermatitis with dis-seminated cutaneous herpes simplex virus infection. JInvest Dermatol. 1987;89:15-18.
8. Williams NA, Hill TJ, Hooper DC. Murine epidermalantigen-presenting cells in primary and secondary T-cell proliferative responses to herpes simplex virus invitro. Immunology. 1991 ;72:34-39.
9. Klareskog L, Forsuin U, Tjernlund UM, Rask L, Pe-terson PA. Expression of la antigen-like molecules oncells in the corneal epithelium. Invest Ophlhalmol VisSci. 1979;18:310-313.
10. Klareskog L, Tjernlund UM, Forsum U, Peterson PA.Epidermal Langerhans cells express la antigens. Na-ture. 1977; 268:248-250.
11. Rowden G, Lewis MG, Sullivan AK. la antigen expres-sion on human epidermal Langerhans cells. Nature.1977;268:247-248.
12. Rowden G. Expression of la antigens on Langerhanscells in mice, guinea pigs, and man. / Invest Dermatol.1980;75:22-31.
13. Rodrigues MM, Rowden G, Hackett J, Bakos 1. Lan-gerhans cells in the normal conjunctiva and periph-eral cornea of selected species. Invest Ophlhalmol VisSri. 1981;21:759-765.
14. Gillette TE, Chandler JW, Greiner JV. Langerhanscells of the ocular surface. Ophthalmology. 1982;89:700-710.
15. Jager MJ, Atherton S, Bradley D, Streilein JW. Herpe-tic stromal keratitis in mice: Less reversibility in thepresence of Langerhans cells in the central cornea.Curr Eye Res. 1991; 10(suppl):69-73.
16. Roussel TJ, Osato MS, Wilhelmus KR. Corneal Lan-gerhans' cell migration following ocular con tact hyper-sensitivity. Cornea. 1983;2:27-30.
17. Asbell PA, Kamenar T. The response of Langerhanscells in the cornea to herpetic keratitis. CUIT Eye Res.1987;6:179-182.
18. Lewkowicz-Moss SJ, Shimeld C, Lipworth K, Hill TJ,Blyth WA, Easty DL. Quantitative studies on Langer-hans cells in mouse corneal epithelium following in-fection with herpes simplex virus. Exp Eye Res.1987;45:127-140.
19. Streilein JW, Toews GB, Bergstresser PR. Corneal al-lografts fail to express la antigens. Nature.1979;282:326-327.
20. Pepose JS. The relationship of corneal Langerhanscells to herpes simplex antigens during dendritic kera-titis. Curr Eye Res. 1989;8:851-858.
21. Aiba S, Katz SI. Phenotypic and functional character-istics of in vivo-activated Langerhans cells. J Immunol.1990; 145:2791-2796.
22. Yasumoto S, Okabe N, Mori R. Role of epidermalLangerhans cells in resistance to herpes simplex virusinfection. Arch Virol. 1986;90:261-271.
23. Stingl G, Katz SI, Green 1, Shevach EM. The func-tional role of Langerhans cells. J Invest Dermatol.1980;74:315-318.
24. Silberberg-Sinakin I, Thorbecke GJ, Baer RL, Ro-senthal SA, Berezowsky V. Antigen-bearing Langer-hans cells in skin, dermal lymphatics and in lymphnodes. Cell Immunol. 1976; 25:137-151.

2290 Investigative Ophthalmology & Visual Science, June 1993, Vol. 34, No. 7
25. Rouse BT. Immunopathology of herpesvirus infec-tions. In: Roizman B, Lopez C, eds. The Herpesviruses.vol. 4. New York: Plenum Press; 1985:103-119.
26. McLeish W, Rubsamen P, Atherton SS, Streilein JW.Immunobiology of Langerhans cells on the ocularsurface: II. Role of central corneal Langerhans cells instromal keratitis following experimental HSV-1 infec-tion in mice. Reg Immunol. 1989; 2:236-243.
27. Russell RC, Naisisse MP, Larsen HS, Rouse BT. Roleof T-lymphocytes in the pathogenesis of herpetic stro-mal keratitis. Invest Ophthalmol Vis Sci. 1984; 25:938-944.
28. Metcalf JF, Kaufman HE. Herpetic stromal keratitis:Evidence for cell-mediated immunopathogenesis. AmJ Ophthalmol. 1976;82:827-834.
29. Newell CK, Martin S, Sendele D, Mercadal CM, RouseBT. Herpes simplex virus-induced stromal keratitis:Role of T-lymphocyte subsets in immunopathology. JVirol. 1989;63:769-775.
30. Youinou P, Colin J, Miossec P. Levels of T lympho-cytes, but not specific antibodies, reflect clinical pre-sentation in human herpes simplex keratitis. hit ArchAllergy Appl Immunol. 1982;69:400-401.
31. Foster CS, Opremcak EM, Rice B, et al. Clinical,pathologic, and immunopathologic characteristics ofexperimental murine herpes simplex virus stromalkeratitis and uveitis is controlled by gene productsfrom the Igh-1 locus on chromosome 12. Trans AmOphthalmol Soc. 1987;85:293-311.
32. Pepose JS. Herpes simplex keratitis: Role of viral in-fection versus immune response. Surv Ophthalmol.1991; 35:345-352.
33. Shimeld C, Hill T, Blyth B, Easty D. An improvedmodel of recurrent herpetic eye disease in mice. CurrEye Res. 1989;8:1193-1205.
34. Shimeld C, Hill TJ, Blyth WA, Easty DL. Reactivationof latent infection and induction of recurrent herpeticeye disease in mice./ Gen Virol. 1990;71:397-404.
35. Laycock KA, Lee SF, Brady RH, Pepose JS. Character-ization of a murine model of recurrent herpes simplexviral keratitis induced by ultraviolet-B radiation. InvestOphthalmol Vis Sci. 199 J; 32:2741-2746.
36. Mackenzie 1C, Squier CA. Cytochemical identificationof ATPase-positive Langerhans cells in EDTA-sepa-
rated sheets of mouse epidermis. Br J Dermatol.1975;92:523-533.
37. Gillette TE, Chandler JW. Immunofluorescence andhistochemistry of corneal epithelial flat mounts: Useof EDTA. CurrEyeRes. 1981; 1:249-253.
38. Cooper KD, Fox P, Neises G, Katz SI. Effects of ultra-violet radiation on human epidermal cell alloantigenpresentation: Initial depression of Langerhans cell-dependent function is followed by the appearance ofT6~ Dr+ cells that enhance epidermal alloantigen pre-sentation./ Immunol. 1985; 134:129-137.
39. Rook G. Cell-mediated immune responses. In: Roitt I,Brostoff J, Male D, eds. Immunology. St. Louis: CVMosby; 1989:9.1-9.14.
40. Niederkorn JY, Peeler JS, Mellon J. Phagocytosis ofpaniculate antigens by corneal epithelial cells stimu-lates interleukin-1 secretion and migration of Langer-hans cells into the central cornea. Reg Immunol.1989;2:83-90.
41. Kupper TS. Immune and inflammatory processes incutaneous tissues: Mechanisms and speculations. /Clin Invest. 1990;86:1783-1789.
42. Sica A, Wang JM, Colotta F, et al. Monocyte chemo-tactic and activating factor gene expression induced inendothelial cells by IL-1 and tumor necrosis factor. /Immunol. 1990; 144:3034-3038.
43. Oppenheim JJ, Zachariae COC, Mukaida N, Matsu-shima K. Properties of the novel proinflammatory su-pergene "intercrine" cytokine family. Annu Rev Im-munol. 1991;9:617-648.
44. Lutty GA, Liu SH, Prendergast RA. Angiogenic lym-phokines of activated T-cell origin. Invest OphthalmolVis Sci. 1983;24:1595-1601.
45. BenEzra D, Hemo I, Maftzir G. In vivo angiogenicactivity of interleukins. Arch Ophthalmol. 1990; 108:573-576.
46. Klintworth GK, Burger PC. Neovascularization of thecornea: Current concepts of its pathogenesis. hit Oph-thalmol Clin. 1983;23:27-39.
47. Hendricks RL, Epstein RJ, Tumpey T. The effect ofcellular immune tolerance to HSV-1 antigens on theimmunopathology of HSV-1 keratitis. Invest Ophthal-mol Vis Sci. 1989;30:105-115.
48. LiesegangTJ. Epidemiology of ocular herpes simplex.Arch Ophthalmol. 1989; 107:1160-1165.
Copyright © 2022 FDOKUMEN




















