
Copy number variation at the FCGR locus includes FCGR3A, FCGR2C and FCGR3B but not FCGR2A and FCGR2B
11
HUMAN MUTATION MUTATION IN BRIEF HUMAN MUTATION Mutation in Brief #1063, 30:E640-650 (2009) Online OFFICIAL JOURNAL www.hgvs.org Copy Number Variation at the FCGR Locus Includes FCGR3A, FCGR2C and FCGR3B but not FCGR2A and FCGR2B Willemijn B. Breunis 1,2 , Edwin van Mirre 2 , Judy Geissler 2 , Nadja Laddach 3 , Gertjan Wolbink 4 , Ellen van der Schoot 5 , Masja de Haas 5 , Martin de Boer 2 , Dirk Roos 2 and Taco W. Kuijpers 1,2 1 From the Department of Pediatric Hematology, Immunology & Infectious Disease at the Emma Children’s Hospital, Academic Medical Center (AMC), Amsterdam, 1105 AZ, The Netherlands; 2 Department of Blood Cell Research, Sanquin Research and Landsteiner Laboratory, AMC, University of Amsterdam, Amsterdam, 1066 CX, The Netherlands; 3 MRC Holland, Amsterdam, 1057 DN, The Netherlands; 4 Jan van Breemen Institute, Amsterdam, 1056 AB, The Netherlands; 5 Department of Experimental Immunohematology, Sanquin Research and Landsteiner Laboratory, AMC, University of Amsterdam, Amsterdam, 1066 CX, The Netherlands *Correspondence to; T.W. Kuijpers, MD, PhD, Emma Children's Hospital, Academic Medical Center Room G8-205, Meibergdreef 9, 1105 AZ Amsterdam, the Netherlands. Tel: +31-20-5662727, Fax: +31-20-6917735, E-mail: [email protected] Communicated by Pui-Yan Kwok ABSTRACT: Human Fcγ receptors (FcγRs) are glycoproteins that bind the Fc region of IgG. The genes encoding the low-affinity FcγRs are located on chromosome 1q23-24. Beside single nucleotide polymorphisms (SNPs), gene copy number variation (CNV) is now being recognized as an important indicator for inter-individual differences. Recent studies on identifying CNV in the human genome suggest large areas at chromosome 1q23-24 to be involved, and CNV in this region has been associated with manifestations of systemic autoimmune disease. To study both SNPs and CNV of the low-affinity FcγRs in one assay, we have developed a Multiplex Ligation-dependent Probe Amplification (MLPA) assay. A novel CNV for FCGR3A was observed. Similar to FCGR3B and FCGR2C, a gene-dosage effect of FCGR3A was found, that seemed to correlate nicely with the FcγRIIIa expression on NK cells. Next, we delineated the approximate boundaries of CNV at the FCGR locus. Variation in co-segregation of neighboring FCGR genes was limited to four variants, with patterns of Mendelian inheritance. No CNV of the FCGR2A and FCGR2B genes was observed in over 600 individuals. In conclusion, we report a novel CNV of the FCGR3A gene that correlates with FcγRIIIa expression and function on NK cells. Only FCGR3A, FCGR2C and FCGR3B show CNV, in contrast to FCGR2A and FCGR2B. ©2009 Wiley-Liss, Inc. KEY WORDS: Copy number variation, single nucleotide polymorphism, Fcγ receptors INTRODUCTION Human Fcγ receptors (FcγRs) are glycoproteins that bind the Fc region of IgG. Depending on their expression on the many different effector leukocytes, the FcγRs exert different effects (Ravetch, 1994). Based on their affinity © 2009 WILEY-LISS, INC. Received 12 October 2008; accepted revised manuscript 21 January 2009. DOI: 10.1002/humu.20997
Transcript of Copy number variation at the FCGR locus includes FCGR3A, FCGR2C and FCGR3B but not FCGR2A and FCGR2B

HUMAN MUTATION MUTATION IN BRIEF
HUMAN MUTATION Mutation in Brief #1063, 30:E640-650 (2009) Online
OFFICIAL JOURNAL
www.hgvs.org
Copy Number Variation at the FCGR Locus Includes FCGR3A, FCGR2C and FCGR3B but not FCGR2A and FCGR2B Willemijn B. Breunis 1,2, Edwin van Mirre2, Judy Geissler 2, Nadja Laddach 3, Gertjan Wolbink 4, Ellen van der Schoot 5, Masja de Haas5, Martin de Boer 2, Dirk Roos 2 and Taco W. Kuijpers 1,2
1 From the Department of Pediatric Hematology, Immunology & Infectious Disease at the Emma Children’s Hospital, Academic Medical Center (AMC), Amsterdam, 1105 AZ, The Netherlands; 2 Department of Blood Cell Research, Sanquin Research and Landsteiner Laboratory, AMC, University of Amsterdam, Amsterdam, 1066 CX, The Netherlands; 3 MRC Holland, Amsterdam, 1057 DN, The Netherlands; 4 Jan van Breemen Institute, Amsterdam, 1056 AB, The Netherlands; 5 Department of Experimental Immunohematology, Sanquin Research and Landsteiner Laboratory, AMC, University of Amsterdam, Amsterdam, 1066 CX, The Netherlands
*Correspondence to; T.W. Kuijpers, MD, PhD, Emma Children's Hospital, Academic Medical Center Room G8-205, Meibergdreef 9, 1105 AZ Amsterdam, the Netherlands. Tel: +31-20-5662727, Fax: +31-20-6917735, E-mail: [email protected] Communicated by Pui-Yan Kwok
ABSTRACT: Human Fcγ receptors (FcγRs) are glycoproteins that bind the Fc region of IgG. The genes encoding the low-affinity FcγRs are located on chromosome 1q23-24. Beside single nucleotide polymorphisms (SNPs), gene copy number variation (CNV) is now being recognized as an important indicator for inter-individual differences. Recent studies on identifying CNV in the human genome suggest large areas at chromosome 1q23-24 to be involved, and CNV in this region has been associated with manifestations of systemic autoimmune disease. To study both SNPs and CNV of the low-affinity FcγRs in one assay, we have developed a Multiplex Ligation-dependent Probe Amplification (MLPA) assay. A novel CNV for FCGR3A was observed. Similar to FCGR3B and FCGR2C, a gene-dosage effect of FCGR3A was found, that seemed to correlate nicely with the FcγRIIIa expression on NK cells. Next, we delineated the approximate boundaries of CNV at the FCGR locus. Variation in co-segregation of neighboring FCGR genes was limited to four variants, with patterns of Mendelian inheritance. No CNV of the FCGR2A and FCGR2B genes was observed in over 600 individuals. In conclusion, we report a novel CNV of the FCGR3A gene that correlates with FcγRIIIa expression and function on NK cells. Only FCGR3A, FCGR2C and FCGR3B show CNV, in contrast to FCGR2A and FCGR2B. ©2009 Wiley-Liss, Inc.
KEY WORDS: Copy number variation, single nucleotide polymorphism, Fcγ receptors
INTRODUCTION
Human Fcγ receptors (FcγRs) are glycoproteins that bind the Fc region of IgG. Depending on their expression on the many different effector leukocytes, the FcγRs exert different effects (Ravetch, 1994). Based on their affinity
© 2009 WILEY-LISS, INC.
Received 12 October 2008; accepted revised manuscript 21 January 2009.
DOI: 10.1002/humu.20997

E641 CNV at the FCGR Locus
for monomeric IgG, three types of FcγRs are discriminated. Type I is a high-affinity receptor, whereas type II and type III are low-affinity receptors (Galon, et al., 1997; Maenaka, et al., 2001).
Genetic variation within the low-affinity receptors has been associated with different diseases. FcγRII (CD32) is encoded by three very homologous genes, FCGR2A (MIM# 146790), FCGR2B (MIM# 604590) and FCGR2C (MIM# 612169). In contrast, FcγRIII (CD16) is encoded by two genes, FCGR3A (MIM# 146740) and FCGR3B (MIM# 610665), which also show a very high degree of sequence homology. FcγRIIIa is expressed on NK cells, monocytes and macrophages, whereas FcγRIIIb is expressed solely on neutrophils. Both genes contain single nucleotide polymorphisms (SNPs) that influence receptor function. Koene et al. have shown that FcγRIIIa-V158 has a higher affinity for IgG1 and IgG3 than does FcγIIIa-F158 (Koene, et al., 1997). On FcγRIIIb the Human Neutrophil Antigen system is located. FcγRIIIb-HNA1a internalizes IgG1- or IgG3-opsonized particles more efficiently than does FcγRIIIb-HNA1b (Huizinga, et al., 1989).
Besides SNPs, gene Copy Number Variation (CNV) (Redon, et al., 2006), is now recognized as an important indicator for inter-individual differences. From a functional perspective, gene copy number differences may contribute to variation in gene expression, at the transcript and/or protein level. In turn, altered FcγR levels can alter the functional capacity of a given cell in response to activation by IgG complexes. In the human gene cluster for IgG receptors, deletions and duplications are known for FCGR3B and the adjacent FCGR2C gene (De Haas, et al., 1995; Koene, et al., 1996). Recent large-scale studies on identifying CNV in the human genome suggest large areas at chromosome 1q23-24 to be involved, encompassing the afore-mentioned FCGR genes and including FCGR2A, FCGR2B and FCGR3A (Database of Genomic, ; de Smith, et al., 2007; Iafrate, et al., 2004; Redon, et al., 2006; Wong, et al., 2007). Variation in the copy number of FCGR3B has recently been associated with glomerulonephritis in the autoimmune disease systemic lupus erythematosus (SLE) (Aitman, et al., 2006) and with susceptibility to systemic autoimmunity (Fanciulli, et al., 2007).
To study the genetic variation of this complex gene cluster we developed a Multiplex Ligation-dependent Probe Amplification (MLPA) assay that enabled us to study the genetic variation, including SNPs and CNV, in the genes encoding the low-affinity FcγRs (FCGR2A, FCGR2B, FCGR2C, FCGR3A and FCGR3B) in one assay (Breunis, et al., 2008). Here, we narrowed down the area of CNV within this locus, because we did not observe any CNV for the FCGR2A and the FCGR2B gene in more than 600 individuals. However, we report a novel CNV in FCGR3A that correlates with the expression and function of FcγRIIIa on NK cells.
MATERIALS AND METHODS
Subjects DNA was available for genotyping from 129 healthy adult white volunteers, 116 patients with idiopathic
thrombocytopenia (ITP; MIM# 188030), 159 patients diagnosed with Kawasaki disease (KD; MIM# 611775), 112 patients with rheumatoid arthritis (RA; MIM# 180300) and 106 women who became sensitized against RhD despite adequate anti-D immunoglobulin prophylaxis in the majority of cases. All cases are of Northern-European ancestry. Informed consent was obtained from all parents of patients <12 years old and from all patients >12 years of age. The study was approved by the Medical Ethics Committee of the Academic Medical Center (AMC) in Amsterdam and was performed in accordance with the Declaration of Helsinki.
Multiplex Ligation-dependent Probe Amplification MLPA was performed as described extensively (Breunis, et al., 2008). In short, MLPA probes were designed
specific for the FCGR2A, FCGR2B, FCGR2C, FCGR3A and FCGR3B genes. In addition two synthetic probes specific for the HSPA6 (MIM# 140555) and HSPA7 (MIM# 140556) gene were added.
HSPA6: CGAGAGGCTCTCAACTGGGCGGGAAGGTGCGGGAAGGTGC- GGAAAGGTTCGCGAAAGTTCGCGGCGGCGGGGGTCGGG HSPA7: CGGGCGGGAGGCTCTCGACTGGGCGGGAAGGTGCGGGAAG- GTTCGCGGCGGCGGGGTCGGGGAGGTGCAAAAG
The MLPA assay was performed according to the first description by Schouten et al (Schouten, et al., 2002).

Breunis et al. E642
Sequencing Since a low probe signal can be caused by either a deletion or a SNP that interferes with accurate probe binding,
the area encompassing the ligation point of the probe 3A-D01 and 3A-D05 were sequenced. For the 3A-D01 an FCGR3A-specific product of 470 bp was amplified with the sense primer 5'-AGATGCTCAGCCACAGACCT-3' and the antisense primer 5'-CCCATCTTGGCTTGTCCTA-3'. For 3A-D05 an FCGR3A-specific product of 393 bp was amplified with the sense primer 5'-CAGTGTCAACCATCTCATCATTATT-3' and the antisense primer 5'-CAGCTACTCACTGGGGCTTC-3'. The PCR conditions were 94○C for 5 min, 35 cycles of 94○C for 1 min, 60○C for 1 min and 72○C for 1 min, followed by 10 min at 72○C. The PCR product was purified and sequenced as described before (van Mirre, et al., 2006).
Quantitative RT- PCR To confirm our observation of CNV for FCGR3A, we set up a relative quantification assay by means of the
LightCycler instrument. The same specific primer pairs were used as for sequencing. The reaction was performed with Lightcycler Faststart DNA MasterPLUS SYBR green 1 mix (Roche). The annealing temperature for all primer pairs was 60○C.
A serial dilution of DNA of a reference sample (2 copies of FCGR3A) was used to construct a standard curve. To correct for DNA input the gene CYBB (MIM# 300481) was used as a reference.
Monoclonal antibodies and reagents The following monoclonal antibodies (mAb) against FcγRs were used: CD16 (anti-FcγRIII, IgG1 isotype, clone
3G8; Sanquin, Amsterdam, The Netherlands); CD16-APC-Cy7 (anti-FcγRIII, IgG1 isotype, clone 3G8, BD Pharmingen, Alphen aan de Rijn, The Netherlands). For the detection of NK cells CD56-APC (isotype IgG1, clone B159, BD Pharmingen) was used. Relevant isotype controls were IgG1-APC (clone MOPC-21, BD Pharmingen) and IgG1-APC-Cy7 (clone X40, BD Pharmingen). Expression of these markers was analyzed by tri-color flowcytometry on a FACS LSRII machine (BD Biosciences) with BD FACS Diva Software (version 5.0.1, BD Biosciences, San Jose, CA, USA).
Evaluation of FcγRIIIa expression levels Whole blood was diluted 1:10 and incubated with CD56-APC and CD16-APC-Cy7. Expression of FcγRIIIa
was determined on NK cells by flow cytometry. NK cells were identified within the lymphocyte population as CD56+ cells. Measurements were performed in triplicate.
Redirected Antibody-dependent Cytotoxicity Assay The cytotoxicity of NK cells was assessed by incubating NK cells in the presence of FcγR-bearing P815 cells as
targets, as described previously (Breunis, et al., 2008). In short, PBL were mixed with target cells at E:T ratios of 50:1, 25:1, 12:1 and 6:1, in the presence or absence of 5 μg/ml anti-FcγRIII (clone 3G8). After 4 h cell death was determined by flow cytometry. The percentage of specific cell death was calculated by subtracting the percentage of spontaneous cell death from experimental samples.
Statistics Genotyping data were analyzed by Fisher's exact tests, a p-value of <0.05 was considered statistically
significant. Results of the in vitro data are expressed as mean ± SEM. Where applicable, Student’s t test or one-way ANOVA was used.

E643 CNV at the FCGR Locus
RESULTS
Distribution of FCGR3A CNV Genetic variation in the FCGR gene cluster was studied by means of an MLPA assay. Previously, we reported about the distribution of FCGR gene CNV in 100 white volunteers and 116 white ITP
patients (Breunis, et al., 2008). The FCGR2C, FCGR3A and the FCGR3B genes showed variation in gene copy number.
To investigate the frequency of the CNV of FCGR3A, we genotyped additional healthy white volunteers (n=129 in total). Apart from ITP, two other autoinflammatory diseases were studied, in which a role for FcγRs has been suggested, i.e. KD, as the most common pediatric vasculitis of the heart (n=159) and RA, as the most common rheumatic disease (n=112). Also 106 RhD-negative women, who became sensitized despite preventive anti-RhD immunoglobulin prophylaxis, were genotyped. CNV for the FCGR3A gene was observed in 36 individuals, with 24 (3.9%) individuals having 3 gene copies and 12 (1.9%) carrying only a single FCGR3A gene copy (Table 1). We did not observe any individual with a complete absence of the FCGR3A gene. CNV in FCGR3A was confirmed by quantitative PCR (data not shown). To validate the MLPA results, areas encompassing the ligation points of the probes were sequenced to exclude that genetic variation caused by SNPs at the annealing site for the probes could influence the probe signal (data not shown). The cDNA of individuals carrying only one allele of FCGR3A were sequenced to exclude any heterozygous variation within the sequence (data not shown).
Table 1. FCGR3A CNV frequencies FCGR3A Controls ITP KD RA RhD-
0 0 (0%) 0 (0%) 0 (0%) 0 (0%) 0 (0%) 1 2 (1.6%) 4 (3.4%) 3 (1.9%) 1 (0.9%) 2 (1.9%) 2 122 (94.6%) 108 (93.1%) 152 (95.6%) 107 (95.5%) 97 (91.5%) 3 5 (3.9%) 4 (3.4%) 4 (2.5%) 4 (3.6%) 7 (6.6%)
Individuals with one, two or three copies of the FCGR3A gene were observed. No significant differences were observed when the frequencies of the control group were compared to any of the patient groups. Controls = healthy white, ITP = idiopathic thrombocytopenia, KD = Kawasaki Disease, RA= rheumatoid arthritis, RhD-= rhesus-D negative women who became sensitized against RhD. Data as number and (percentage).
CNV of FCGR3A correlates with FcγRIIIa surface expression and function of NK cells To evaluate whether the CNV in FCGR3A correlates with FcγRIIIa expression levels, we obtained blood from
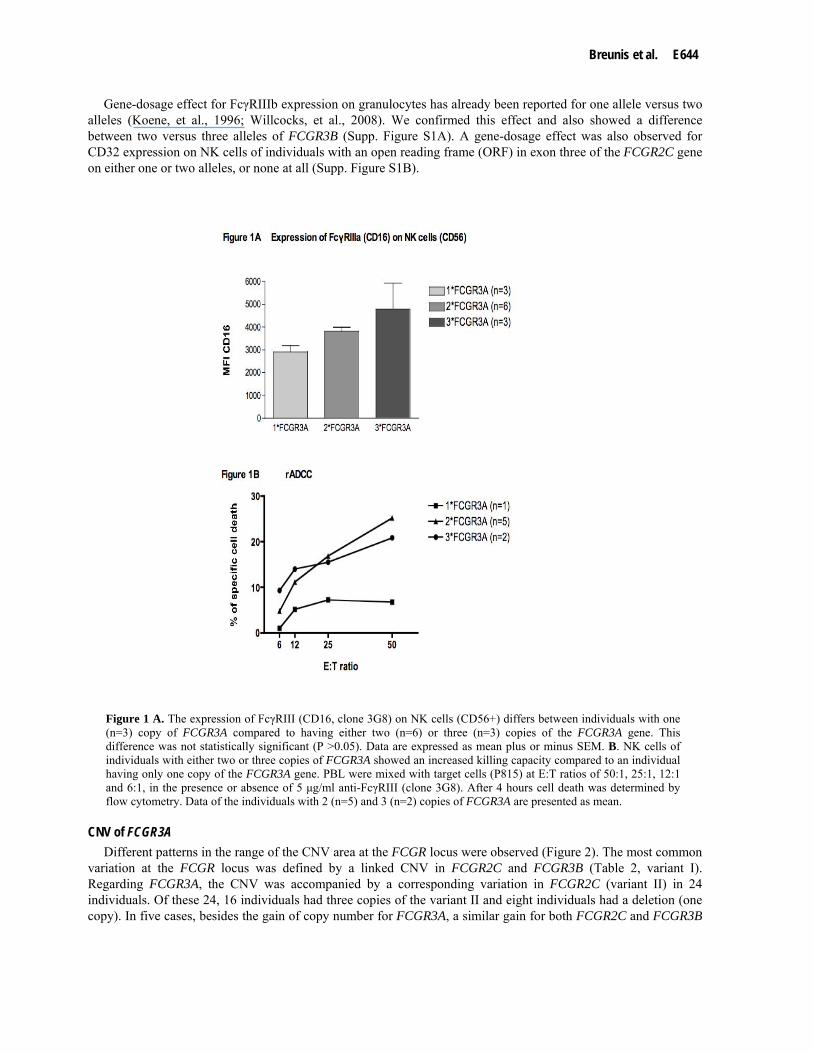
individuals having either one (n=3), two (n=6) or three (n=3) FCGR3A genes and determined expression levels by flow cytometry. Testing the NK cells (CD56+), a gene-dosage effect was observed for FcγRIIIa (CD16) expression, i.e. individuals having one FCGR3A copy had a lower expression than individuals having either two or three copies of the FCGR3A gene (Figure 1A). However, these differences in expression of CD16 were not significant, probably due to the low number of individuals tested. Due to the low frequency of the CNV for FCGR3A more samples were not available to test the expression.
We subsequently examined whether FcγRIIIa expression levels correlated to NK cell function by assessing rADCC capacity of these cells from individuals having different copy numbers of FCGR3A. For this purpose we obtained blood from five healthy controls with two copies of FCGR3A and two healthy controls with three copies, as well as one control with one copy of FCGR3A. The antibody-dependent killing capacity of NK cells with either two or three FCGR3A copies did not differ significantly. However, NK cells from the individual with one FCGR3A displayed a marked decrease in antibody-dependent killing capacity (Figure 1B).
Breunis et al. E644
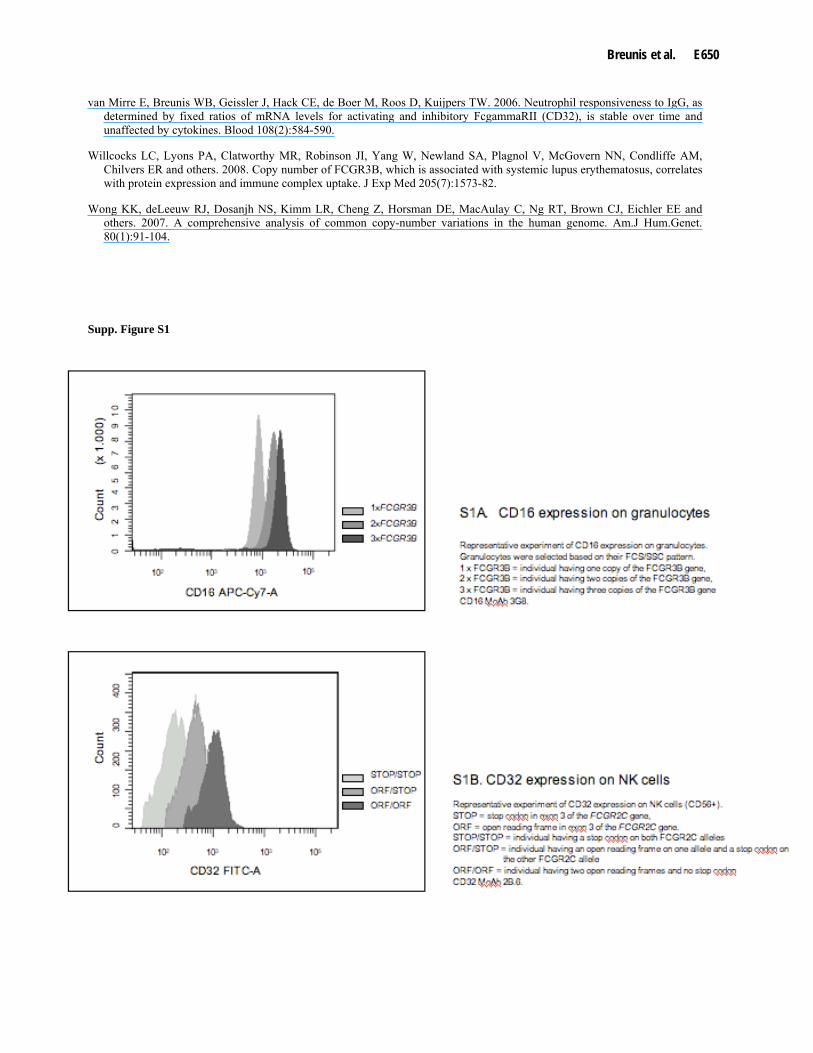
Gene-dosage effect for FcγRIIIb expression on granulocytes has already been reported for one allele versus two alleles (Koene, et al., 1996; Willcocks, et al., 2008). We confirmed this effect and also showed a difference between two versus three alleles of FCGR3B (Supp. Figure S1A). A gene-dosage effect was also observed for CD32 expression on NK cells of individuals with an open reading frame (ORF) in exon three of the FCGR2C gene on either one or two alleles, or none at all (Supp. Figure S1B).
Figure 1 A. The expression of FcγRIII (CD16, clone 3G8) on NK cells (CD56+) differs between individuals with one (n=3) copy of FCGR3A compared to having either two (n=6) or three (n=3) copies of the FCGR3A gene. This difference was not statistically significant (P >0.05). Data are expressed as mean plus or minus SEM. B. NK cells of individuals with either two or three copies of FCGR3A showed an increased killing capacity compared to an individual having only one copy of the FCGR3A gene. PBL were mixed with target cells (P815) at E:T ratios of 50:1, 25:1, 12:1 and 6:1, in the presence or absence of 5 μg/ml anti-FcγRIII (clone 3G8). After 4 hours cell death was determined by flow cytometry. Data of the individuals with 2 (n=5) and 3 (n=2) copies of FCGR3A are presented as mean.
CNV of FCGR3A Different patterns in the range of the CNV area at the FCGR locus were observed (Figure 2). The most common
variation at the FCGR locus was defined by a linked CNV in FCGR2C and FCGR3B (Table 2, variant I). Regarding FCGR3A, the CNV was accompanied by a corresponding variation in FCGR2C (variant II) in 24 individuals. Of these 24, 16 individuals had three copies of the variant II and eight individuals had a deletion (one copy). In five cases, besides the gain of copy number for FCGR3A, a similar gain for both FCGR2C and FCGR3B

E645 CNV at the FCGR Locus
was observed (variant III). An isolated gain in CNV for FCGR3A (variant IV) was observed in three cases. An isolated loss (one copy) of FCGR3A was observed in four cases.
To obtain more insight in the CNV at the FCGR locus, synthetic probes for the genes HSPA6 and HSPA7 located within this locus (Figure 2) were designed and added to the MLPA assay. HSPA6 is a stress-induced heat-shock gene encoding a 70-kD protein; it shares about 95% homology with the HSPA7 gene (Leung, et al., 1992). HSPA6 is located on chromosome 1 between FCGR2A and FCGR3A, HSPA7 is located at the 3’ UTR site of FCGR2C (Figure 2). Individuals with no variation in gene copy number at the FCGR region also did not show CNV for either HSPA6 or HSPA7. Variant I was always accompanied with CNV for the HSPA7 gene, as can be expected by the location of the gene. The individual deficient for FCGR3B and FCGR2C also did not have the HSPA7 gene. In Table 3, we show the detailed CNV and SNP data in these genes in the white healthy controls carrying a variant copy number of the FCGR3A gene. In most cases the CNV for FCGR3A was accompanied by CNV for HSPA6, although exceptions do exist (samples C4 and C7, Table 3). Within the control group, CNV of FCGR3A occurred on a background of different SNPs. No absolute linkage between SNPs and CNV was observed (Table 3). For instance, both variants were seen in the individuals having only one copy of the gene (158V and 158F) for the SNP at position 158 (V>F) in the FCGR3A gene.
The dotted lines in Figure 2 indicate that the exact breakpoints of the CNV are not known, because no probes were located in the areas between the annotated genes.
The frequency of CNV of the FCGR3A gene in the observed study populations is low, and no significant difference between the control group and the subgroups included in the study were observed. The transmission of different CNV variants was tested in pedigrees (Figure 3). Transmission of the FCGR3A CNV could be tested in a pedigree of one of the patients with only one copy of FCGR3A. The mother of the patient also showed the loss of one of the FCGR3A gene copies (Figure 3B). All pedigrees tested showed inheritance of the CNV according to expected Mendelian inheritance patterns.
Table 2. Frequencies of the observed variants within the subgroups
Four different patterns of CNV (I-IV) were observed. An individual can either have two copies of the variant, three copies (duplication) of the variant or one copy (deletion). The numbers of individuals with either one or three copies are mentioned in the table. I = CNV of FCGR2C + FCGR3B, II = CNV of FCGR3A + FCGR2C, III = CNV of FCGR3A + FCGR2C + FCGR3B, IV = CNV of FCGR3A. ITP= idiopathic thrombocytopenia, KD= Kawasaki Disease, RA= Rheumatoid Arthritis, RhD = rhesus-D negative women. N = total number of individuals.
Copy Number I II III IV Controls (n=129) 3 13 3 1 1 1 9 1 0 1 ITP (n = 116) 3 7 3 0 1 1 6 4 0 0 KD (n = 159) 3 15 3 0 1 1 10 1 0 2 RA (n = 112) 3 5 3 1 0 1 10 1 0 0 RhD- (n= 106) 3 11 4 3 0 1 5 1 0 1

Breunis et al. E646
Table 3. FCGR2 and FCGR3 CNV and SNP data of control samples with FCGR3A CNV
Complete genotyping data of the 7 controls (C1-C7) showing CNV for the FCGR3A. C= Dutch control. 2A = FCGR2A, 131H/R = histidine to arginine amino acid substitution at position 131 in FCGR2A, HSPA6 = Heat-shock protein A6, 3A = FCGR3A, 158V/F = valine to phenylalanine amino acid substitution at position 158 in FCGR3A, 2C = FCGR2C, HSPA7 = heat shock protein A7, 3B = FCGR3B, HNA1a/HNA1b = human neutrophil antigen1a and 1b (formerly know as NA1 and NA2), HNA1c = human neutrophil antigen 1c (formerly known as SH), 2B = FCGR2B, 232I/T = isoleucine to threonine amino acid substitution at position 232 in FCGR2B.
Figure 2. Black boxes refer to the genes located in this area on chromosome 1.The arrows underneath the black boxes refer to the direction of the genes. Four different patterns (I-IV) in the range of CNV were observed, as indicated with the solid gray lines. All variants can either be a gain or loss of the area. The dotted gray lines on both sides of the CNV areas indicate that the exact borders of the CNV are not known.
sample 2A 131H/R
HSPA6 3A 158V/F
2C HSPA7 3B HNA1a/HNA1b HNA1c
2B 232I/T
C1 2 HR 3 3 FFF 3 3 2 HNA1aHNA1b - 2 II C2 2 HR 3 3 FFF 3 3 2 HNA1aHNA1a - 2 II C3 2 RR 3 3 FFF 3 3 3 HNA1aHNA1aHNA1b - 2 II C4 2 HR 2 3 VVF 3 2 2 HNA1aHNA1a - 2 II C5 2 HH 3 3 VFF 3 2 2 HNA1bHNA1b - 2 IT C6 2 RR 1 1 F 1 2 2 HNA1aHNA1b - 2 II C7 2 HR 2 1 V 2 2 2 HNA1bHNA1b - 2 IT

E647 CNV at the FCGR Locus
Figure 3. CNV transmission in pedigrees. A. Pedigree with no variation in CNV for the low affinity FCGR genes FCGR2A, FCGR2B, FCGR2C, FCGR3A and FCGR3B. All tested members within the family have a stop codon in exon 3 of FCGR2C. B. Pedigree with a loss of one copy of FCGR3A (variant IV) C. Pedigree with one copy of FCGR2C and FCGR3B (variant I) D. Pedigree with three copies of FCGR2C and FCGR3B (variant I). St = stop codon in exon 3 of FCGR2C, Orf = open reading frame in exon 3 of FCGR2C. All tested SNPs transmitted according to Mendel's law of inheritance.

Breunis et al. E648
DISCUSSION
CNV has been recognized as an important indicator for inter-individual differences. As CNVs, when located in annotated gene regions, can have functional consequences, it is important when performing association studies to study both SNPs and CNV. The complexity of the observed variations in the human genome is a major hurdle to detect unequivocally the range and impact of CNV and poses great challenges to understand and translate the genetic information into clinical science. Up till now a few associations studies have already demonstrated an association between CNV and disease-susceptibility (Ionita-Laza, et al., 2009). By means of an FCGR-specific MLPA assay we were able to study both functional SNPs and CNV in one test system. Large-scale studies on identifying CNV in the human genome have repeatedly suggested CNV at the FCGR locus, including the human FCGR2A and FCGR2B genes (Database of Genomic, ; de Smith, et al., 2007; Iafrate, et al., 2004; Redon, et al., 2006; Wong, et al., 2007). This could be of great functional and clinical importance, because FCGR2B encodes the only inhibitory IgG receptor, FcγRIIb, whereas the other low-affinity FcγRs are all activating receptors. The third FCGR2 gene, FCGR2C, is likely to be the result of an unequal cross-over event between the 5' part of FCGR2B and the 3' part of the FCGR2A gene. There is very high homology between the FCGR genes, and great care ought to be taken when interpreting the results from such large-scale studies, because the probes used are not in all cases specific enough to discriminate between the individual FCGR2A, FCGR2B and FCGR2C genes.
Since the extent of the region of genetic variation has not been well defined to date, we used our MLPA assay to delineate the variation in gene copy number at the FCGR locus more precisely and were able to show CNV for the FCGR3A, FCGR3B and FCGR2C genes. In the 622 individuals investigated we did not observe anyone with CNV for the FCGR2A or FCGR2B genes.
Because the frequency of CNV for FCGR3A is low, we were limited in our possibilities to study the functional consequences of this CNV. However, our expression data and rADCC results suggest a relevant gene-dosage effect. A more pronounced effect was observed when the rADCC of NK cells from individuals with one versus two copies of FCGR3A was compared than when we looked at individuals with two versus three FCGR3A. This can be explained by the relative difference of 100% versus 50% in additional FcγRIIIa surface expression comparing one with two alleles or two with three alleles.
CNV of FCGR3A might play a role in the severity of certain pathologic conditions, as has been reported for the CNV of FCGR3B and the risk of glomerulonephritis in patients suffering from SLE (Aitman, et al., 2006). The observed frequency of the CNV for FCGR3A in our study was between 0.9 and 6.6% in all subgroups, and no significant association with susceptibility to the studied hematologic and immunologic conditions was observed. So far, we did not observe an individual with a complete absence of the FCGR3A gene. From our data one would estimate the frequency for a complete FCGR3A deficiency to be 1 : 10,000 in white individuals. FcγRIIIb-deficiency has been reported in completely healthy individuals (De Haas, et al., 1995), in contrast to Fanciulli et al. (Fanciulli, et al., 2007), who suggested that FCGR3B deficiency confers a very high risk of auto-immunity.
In conclusion, we have extended the current insight in CNV at the FCGR locus by delineating its variation and boundaries. In contrast to FCGR3A, FCGR2C and FCGR3B, no CNV for the FCGR2A and FCGR2B was observed in more than 600 Caucasian individuals. Not only does CNV in FCGR3A seem to correlate with FcγRIIIa expression levels as determined on the NK cells in peripheral blood, but also with FcγRIIIa-dependent cellular activity, as shown by the capacity to perform rADCC. The FCGR3A gene shows CNV at a frequency of approximately 1 : 20 (either gain or loss of FCGR3A alleles), either alone or together with FCGR2C or with both FCGR2C and FCGR3B combined. More research on CNV within the FcγR gene cluster might reveal the influence of this locus on the outcome of both infectious and autoimmune diseases to a better extent.
ACKNOWLEDGMENTS
We thank J. Schouten for his assistance in the development of the FCGR-specific MLPA. We thank Dr. M. Bruin, Dr. H.R. Koene and Dr. M. Peters for their contribution in the collection of the patient materials. We thank Dr. G. Vidarsson for critically reading the manuscript.
Funding: This work was supported by grants from the Netherlands Genomic Initiative (NROG, 050-71-315). W.B.B. was supported by ZonMw (920-03-391).

E649 CNV at the FCGR Locus
Authorship: W.B.B. and E.v.M. designed and performed research, analyzed data and wrote the paper. J.G. performed research. M.d.B. provided vital analytical tools, N.L. helped to develop the MLPA assay, G.W. and E.vd.S. provided patient materials and provided intellectual input, M.d.H. provided intellectual input, D.R. and T.W.K. designed the overall study, supervised the research and helped write the paper.
Conflict-of-interest disclosure: The authors declare no competing financial interest
REFERENCES Aitman TJ, Dong R, Vyse TJ, Norsworthy PJ, Johnson MD, Smith J, Mangion J, Roberton-Lowe C, Marshall AJ, Petretto E
and others. 2006. Copy number polymorphism in Fcgr3 predisposes to glomerulonephritis in rats and humans. Nature 439(7078):851-855.
Breunis WB, van Mirre E, Bruin M, Geissler J, de Boer M, Peters M, Roos D, De Haas M, Koene HR, Kuijpers TW. 2008. Copy number variation of the activating FCGR2C gene predisposes to idiopathic thrombocytopenic purpura. Blood 111(3):1029-1038.
Database of Genomic Variants. http://projects.tcag.ca/variation.
De Haas M, Kleijer M, van Zwieten R, Roos D, dem Borne AE. 1995. Neutrophil Fc gamma RIIIb deficiency, nature, and clinical consequences: a study of 21 individuals from 14 families. Blood 86(6):2403-2413.
de Smith AJ, Tsalenko A, Sampas N, Scheffer A, Yamada NA, Tsang P, Ben Dor A, Yakhini Z, Ellis RJ, Bruhn L and others. 2007. Array CGH analysis of copy number variation identifies 1284 new genes variant in healthy white males: implications for association studies of complex diseases. Hum.Mol.Genet. 16(23):2783-2794.
Fanciulli M, Norsworthy PJ, Petretto E, Dong R, Harper L, Kamesh L, Heward JM, Gough SC, de Smith A, Blakemore AI and others. 2007. FCGR3B copy number variation is associated with susceptibility to systemic, but not organ-specific, autoimmunity. Nat.Genet. 39(6):721-723.
Galon J, Robertson MW, Galinha A, Mazieres N, Spagnoli R, Fridman WH, Sautes C. 1997. Affinity of the interaction between Fc gamma receptor type III (Fc gammaRIII) and monomeric human IgG subclasses. Role of Fc gammaRIII glycosylation. Eur.J Immunol. 27(8):1928-1932.
Huizinga TW, Kerst M, Nuyens JH, Vlug A, dem Borne AE, Roos D, Tetteroo PA. 1989. Binding characteristics of dimeric IgG subclass complexes to human neutrophils. J Immunol. 142(7):2359-2364.
Iafrate AJ, Feuk L, Rivera MN, Listewnik ML, Donahoe PK, Qi Y, Scherer SW, Lee C. 2004. Detection of large-scale variation in the human genome. Nat.Genet. 36(9):949-951.
Ionita-Laza I, Rogers AJ, Lange C, Raby BA, Lee C. 2009. Genetic association analysis of copy-number variation (CNV) in human disease pathogenesis. Genomics 93(1):22-6.
Koene HR, De Haas M, Kleijer M, Roos D, dem Borne AE. 1996. NA-phenotype-dependent differences in neutrophil Fc gamma RIIIb expression cause differences in plasma levels of soluble Fc gamma RIII. Br.J.Haematol. 93(1):235-241.
Koene HR, Kleijer M, Algra J, Roos D, dem Borne AE, De Haas M. 1997. Fc gammaRIIIa-158V/F polymorphism influences the binding of IgG by natural killer cell Fc gammaRIIIa, independently of the Fc gammaRIIIa-48L/R/H phenotype. Blood 90(3):1109-1114.
Leung TK, Hall C, Rajendran M, Spurr NK, Lim L. 1992. The human heat-shock genes HSPA6 and HSPA7 are both expressed and localize to chromosome 1. Genomics 12(1):74-79.
Maenaka K, van der Merwe PA, Stuart DI, Jones EY, Sondermann P. 2001. The human low affinity Fcgamma receptors IIa, IIb, and III bind IgG with fast kinetics and distinct thermodynamic properties. J Biol.Chem. 276(48):44898-44904.
Ravetch JV. 1994. Fc receptors: rubor redux. Cell 78(4):553-560.
Redon R, Ishikawa S, Fitch KR, Feuk L, Perry GH, Andrews TD, Fiegler H, Shapero MH, Carson AR, Chen W and others. 2006. Global variation in copy number in the human genome. Nature 444(7118):444-454.
Schouten JP, McElgunn CJ, Waaijer R, Zwijnenburg D, Diepvens F, Pals G. 2002. Relative quantification of 40 nucleic acid sequences by multiplex ligation-dependent probe amplification. Nucleic Acids Res. 30(12):e57.
Breunis et al. E650
van Mirre E, Breunis WB, Geissler J, Hack CE, de Boer M, Roos D, Kuijpers TW. 2006. Neutrophil responsiveness to IgG, as determined by fixed ratios of mRNA levels for activating and inhibitory FcgammaRII (CD32), is stable over time and unaffected by cytokines. Blood 108(2):584-590.
Willcocks LC, Lyons PA, Clatworthy MR, Robinson JI, Yang W, Newland SA, Plagnol V, McGovern NN, Condliffe AM, Chilvers ER and others. 2008. Copy number of FCGR3B, which is associated with systemic lupus erythematosus, correlates with protein expression and immune complex uptake. J Exp Med 205(7):1573-82.
Wong KK, deLeeuw RJ, Dosanjh NS, Kimm LR, Cheng Z, Horsman DE, MacAulay C, Ng RT, Brown CJ, Eichler EE and others. 2007. A comprehensive analysis of common copy-number variations in the human genome. Am.J Hum.Genet. 80(1):91-104.
Supp. Figure S1




















