
Contrasting ecological breadth of co-occurring annual Polygonum species

1988;48:4646-4650. Cancer Res Takashi Sako, Alfred I. Tauber, Arco Y. Jeng, et al. CellsInhibitor, on Human Neutrophils and Primary Mouse Epidermal Contrasting Actions of Staurosporine, a Protein Kinase C
Updated version
http://cancerres.aacrjournals.org/content/48/16/4646
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on August 25, 2014. © 1988 American Association for Cancercancerres.aacrjournals.org Downloaded from
Research. on August 25, 2014. © 1988 American Association for Cancercancerres.aacrjournals.org Downloaded from

(CANCER RESEARCH 48. 4646-4650. August 15. I988|
Contrasting Actions of Staurosporine, a Protein Kinase C Inhibitor, on Human
Neutrophils and Primary Mouse Epidermal CellsTakashi Sako, Alfred I. Tauber,1 Arco Y. Jeng, Stuart H. Yuspa, and Peter M. Blumberg2
Molecular Mechanisms of Tumor Promotion Section [T. S., P. M. B.J and In Vitro Pathogenesis Section fS. H. Y.J, Laboratory of Cellular Carcinogenesis and TumorPromotion, National Cancer Institute, Bethesda, Maryland 20892; William B. Castle Hematology Research Laboratory, Boston City Hospital, and the Departments ofMedicine and Pathology ¡A.I. T.], Boston University School of Medicine, Boston, Massachusetts 021 IS; and Research Department [A. Y. J.¡,Pharmaceutical Division,Cirta-O'eifiy Corporation, Summit, New Jersey 07901
ABSTRACT
Staurosporine, a recently described microbial alkaloid, is uniquelypotent as an inhibitor of protein kinase C in vitro, being active at MMconcentrations rather than the /IM concentrations typical of other inhibitor classes. Like these other inhibitors, however, Staurosporine exhibitsonly limited selectivity among different protein kinases. We report herethat, in intact human neutrophils, IIM concentrations of Staurosporineblocked the action of the phorbol ester tumor promoters. In mouseprimary epidermal cells, on the other hand, Staurosporine failed to blockthe effects of phorbol 12,13-dibutyrate on epidermal growth factor binding and on induction of ornithine decarboxylase and epidermal transglu-taminase. Unexpectedly, Staurosporine induced morphological changesin keratinocytes to a dendritic shape resembling that induced by thephorbol esters. It also induced epidermal transglutaminase and cornifiedenvelope production, markers of the differentiative pathway in the epidermal cells. We conclude that the effectiveness of Staurosporine as aprotein kinase C inhibitor in intact cells may depend markedly on thecell system. Other actions of Staurosporine may predominate, and, inkeratinocytes, its activity is suggestive of a tumor promoter rather thanof an inhibitor of tumor promotion.
INTRODUCTION
Protein kinase C has attracted great research interest ( 1, 2).It is the major receptor of the phorbol ester tumor promoters(3, 4) and mediates one arm of the phosphatidylinositol secondmessenger system. This system is involved in signal transduc-tion for a broad array of hormones and cellular effectors,including both growth factors and several oncogenes (5, 6).
A number of compounds inhibit protein kinase C enzymaticactivity. These include H-73 (7), sphingosine (8), palmitoylcar-
nitine (9), and chlorpromazine (10). These inhibitors unfortunately have proven to be of limited usefulness in biologicalsystems because of a combination of low potency—typicallyMM—andlimited selectivity. H-7, for example, shows similarA'iS for protein kinase C and for the cAMP-dependent and
cGMP-dependent protein kinases, although its Kt for myosinlight chain kinase is 15- to 20-fold greater (7). H-7 is, moreover,competitive with ATP, which has a 7 MMAâ„¢for protein kinaseC ( 11), so effective biological concentrations of H-7 would needto be in appreciable excess over those for ATP in biologicalsystems, which may be HIM.
Recently, a new class of protein kinase C inhibitors have beenreported which are unique in terms of their high potencies.Staurosporine and the related compounds K252a,b inhibit pro-
Received1/21/88;revised5/9/88;acceptedS/13/88.The costs of publication of this article were defrayed in part by the payment
of page charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1Supported by Grants AI20064 and HL33565 from the NIH and by a grantfrom the Fairfax Hospital Association.
2To whom requests for reprints should be addressed.'The abbreviations used are; H-7, l-(5-isoquinolinesulfonyl)-2-methylpipcra-
¿inc;EGF, epidermal growth factor; ODC, ornithine decarboxylase; PDBu.phorbol 12,13-dibutyrate: TGase. transglutaminase; PMA, phorbol 12-myristate13-acetate; FMLP. formylmethionylleucylphenylalanine; 1C».50% inhibitoryconcentration; cAMP, cyclic AMP; cGMP, cyclic GMP.
tein kinase C enzymatic activity in vitro with A^sof 2.7 nM (12)and 20 to 25 nM (13), respectively. Although the details of theirinhibitory mechanisms remain to be established, we have shownthat Staurosporine potently inhibits the proteolytically generated catalytic domain of protein kinase C (14), whereas it hasno effect on phorbol ester binding activity (12, 14). Likewise,inhibition by K252a was reported to be competitive with ATPbut not with the phosphate-accepting substrate or with Ca2+
and phospholipid, modulators of the regulatory domain. LikeH-7, however, Staurosporine and K252a,b show limited selectivity among different protein kinases. Staurosporine inhibitedthe p60v"srctyrosine kinase with an ICso of 6.4 nM (15), and
K252a inhibited the cAMP-dependent and cGMP-dependentprotein kinases with A'iSof 18 and 20 nM, respectively (13).
In intact platelets, Staurosporine blocked the phorbol ester-induced rise in intracellular calcium (ICso = 27 nM) and phorbolester-induced alkalinization (at a Staurosporine concentrationof 100 nM) (16). Effects of Staurosporine on the response tophorbol esters in other systems have not as yet been reported.Other biological effects of Staurosporine include induction ofdifferentiation in human NB-1 neuroblastoma cells at 20 nM(17) and inhibition of platelet aggregation in response to collagen or ADP at MMconcentrations (18). Cytotoxic concentrations ranged from 4 x 10~12M for HeLa S3 cells upon a 72-hincubation (12) to 2.1 X IO"7 M for the NB-1 cells (17). The
related rebeccamycin displayed an ICso of 6 ng/m\ for a 1-hexposure of human lung adenocarcinoma cells (19). The stability of Staurosporine under culture conditions has not beendetermined.
We analyze here the effect of Staurosporine on phorbol esteraction in two biological systems—human neutrophils andmouse primary epidermal cells. Neutrophils have been extensively studied as a model system for stimulus-response coupling(20). The NADPH-oxidase of neutrophils can be activated boththrough protein kinase C-dependent and -independent pathways. Mouse primary epidermal cells are of great relevance forexploring the tumor-promoting activity of the phorbol esters.Subpopulations of the epidermal cells display both proliferativeand differentiative responses; the induction of the differentiativepathway is thought to be essential for the promotion response(21,22).
MATERIALS AND METHODS
PDBU (Sigma, St. Louis, MO) was dissolved in dimethyl sulfoxideat a concentration of 50 fig/ml, stored at -20'C, and diluted with
medium containing serum for treatment of cells. The final concentration of dimethyl sulfoxide did not exceed 0.1%. Staurosporine, thegenerous gift of Kyowa Hakko Kogyo Co., Ltd., was dissolved indimethyl sulfoxide at a concentration of 2 mM, stored at -20°C in
small aliquots, and diluted with medium containing serum to the desiredconcentration immediately before treatment of cells.
Neutrophils were isolated from normal human donors as described(23). Neutrophil respiratory burst activation was assessed by measuringO2~ generation by a modification of the method of Cohen and Chov-
4646
Research. on August 25, 2014. © 1988 American Association for Cancercancerres.aacrjournals.org Downloaded from
CONTRASTING ACTIONS OF STAUROSPOR1NE
aniec (24). Briefly, neutrophils (IO6 cells) were incubated for 3 min at37°Cin 140 mM NaCl-3 mM KC1-1 HIMK2HPO4-1 mM CaCl2-l mM
MgCU-5 mM glucose, pH 7.4, 0.9-ml final volume, in the presence orabsence of the indicated concentrations of staurosporine. An additional0.10 ml of the above solution containing cytochrome <•(final concen
tration in 1 ml, 30 MM),PMA (final concentration in 1 ml, 100 ng/ml),and, for the reference cuvette, Superoxide dismutase (30 /ig) was addedto yield a final volume of 1.0 ml. The sample was incubated for 5 minat 37°Cwith continuous monitoring of absorbance at 550 nm. For
measurement of staurosporine inhibition of opsonized zymosan orarachidonate-induced respiratory burst activity, PMA was replaced by2-mg/ml particles or 82 nM (final concentration) arachidonic acid,respectively; for stimulation by FMLP, 5 ^M FMLP was used in placeof PMA, and 5 Mg/ml of cytochalasin B were included in the 3-minpreincubation period.
Cell Culture. Epidermal cells were prepared from newborn BALB/cmice by the trypsin flotation technique and cultured in low calcium(0.05 mM) Eagle's minimal essential medium (MA Bioproducts, Walk-
ersville, MD) prepared with reduced calcium and supplemented with8% Chelex-treated fetal calf serum (Reheis Chemical Co., Kankakee,IL) as described previously (25). All media contained 1% antibiotic-antimycotic solution (Gibco, Grand Island, NY). Cells were plated atan initial density of 0.24 x 106/cm2 in Costar 6-well tissue cultureplates for assay of EGF binding and in 60-mm Falcon tissue culturedishes for determination of ODC and TGase induction and cornifiedenvelope formation. Media were changed daily for the first 3 days andevery 2 to 3 days thereafter. All experiments were done on confluentcells at 3 to 7 days after plating. Cultures were routinely incubated at37°Cin a humidified atmosphere of 5% CO2 in air.
Enzyme Assays. For assay of ODC, TGase, and protein, cells weretreated as described in the figure legends. They were then washed 3times with Dulbecco's phosphate-buffered saline (0.2 g of KC1-0.2 g of
KH2PO4-8 g of NaCl-2.16 g of Na2HPO„-7H2O/liter), frozen on dryice, and stored at -20°Cuntil assay. The cells were then scraped from
the culture dish with 300 //I of lysis buffer containing 0.02 M sodiumphosphate (pH 7.5), 0.5 mM EDTA, 10 mM dithiothreitol, and 50 Mg/ml of phenylmethanesulfonyl fluoride. Aliquots were analyzed for ODCor for TGase and for protein.
ODC was measured by the release of 14CO2from [l-MC]ornithine
(57 mCi/mmol; Amersham, Arlington Heights, IL) as described (26).TGase activity was assayed by transfer of [2,3-3H]putrescine (New
England Nuclear, Boston, MA; 20 to 40 Ci/mmol) to casein (27).Protein was determined by the method of Lowry et al. (28) afterdestruction of dithiothreitol with chloramine T (29).
EGF binding was assayed with '"I-EGF (150 to 200 /iCi/Mg; New
England Nuclear, Boston, MA), with or without unlabeled EGF, l ¿ig/ml (receptor grade; Collaborative Research, Lexington, MA), as previously described (30, 31). Treatment of cells with staurosporine and/orPDBu was carried out in freshly changed low calcium medium containing 8% chelex-treated serum for l h at 37°Cunless otherwise indicated.
Following treatment, cells were washed 2 times with binding assaybuffer (Eagle's minimal essential medium containing 1 mg/ml of bovine
serum albumin), PDBu was readded, and the cells were incubated with'"1-labeled EGF at 0°Cfor 5 h. The plates were washed 3 times with
binding assay buffer, and cell-associated radioactivity was determinedby liquid scintillation counting. Data were expressed as cpm/well.
The number of cornified cells produced in confluent dishes of mouseepidermal cells was measured as described by Rice and Green (32). Inthis assay, cornified cells were quantified by their resistance to lysis bydetergent in the presence of reducing agent. Attached cells from three60-mm dishes were trypsinized, pooled, and extracted with 2% sodiumdodecyl sulfate containing 20 HIMdithiothreitol for 10 min at 90°C.
Normal cells are solubili/ed, but the envelopes of cornified cells remainintact. Data were expressed as the percentage of cornified cells relativeto the total number of cells per dish.
RESULTS
Inhibition of NADPH-Oxidase Activation. Staurosporine wasa potent inhibitor of Superoxide generation by human neutro
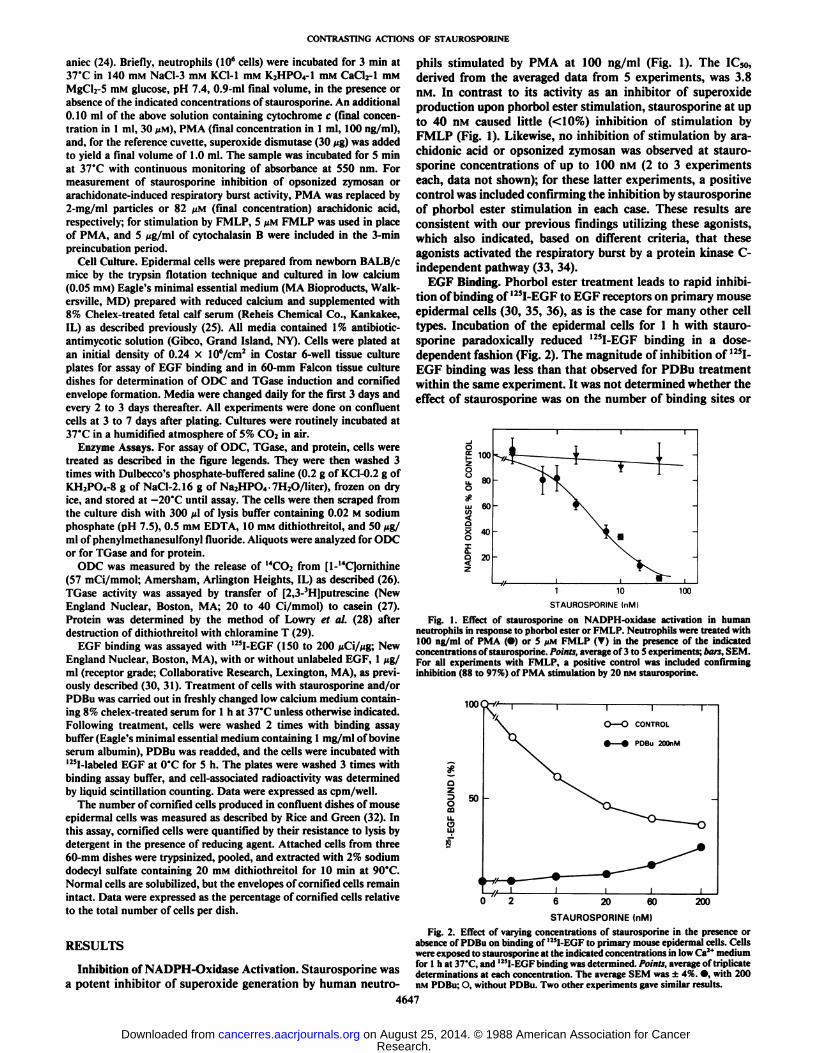
phils stimulated by PMA at 100 ng/ml (Fig. 1). The IC50,derived from the averaged data from 5 experiments, was 3.8nM. In contrast to its activity as an inhibitor of Superoxideproduction upon phorbol ester stimulation, staurosporine at upto 40 nM caused little (<10%) inhibition of stimulation byFMLP (Fig. 1). Likewise, no inhibition of stimulation by arachidonic acid or opsonized zymosan was observed at staurosporine concentrations of up to 100 nM (2 to 3 experimentseach, data not shown); for these latter experiments, a positivecontrol was included confirming the inhibition by staurosporineof phorbol ester stimulation in each case. These results areconsistent with our previous findings utilizing these agonists,which also indicated, based on different criteria, that theseagonists activated the respiratory burst by a protein kinase C-independent pathway (33, 34).
EGF Binding. Phorbol ester treatment leads to rapid inhibition of binding of '"I-EGF to EGF receptors on primary mouse
epidermal cells (30, 35, 36), as is the case for many other celltypes. Incubation of the epidermal cells for l h with staurosporine paradoxically reduced I25I-EGF binding in a dose-dependent fashion (Fig. 2). The magnitude of inhibition of 125I-
EGF binding was less than that observed for PDBu treatmentwithin the same experiment. It was not determined whether theeffect of staurosporine was on the number of binding sites or
100
UL 80
o#uj 60
O
Eo 20
10
STAUROSPORINE InMI
100
Fig. 1. Effect of staurosporine on NADPH-oxidase activation in humanneutrophils in response to phorbol ester or FMLP. Neutrophils were treated with100 ng/ml of PMA (•)or 5 JIM FMLP (T) in the presence of the indicatedconcentrations of staurosporine. Points, average of 3 to 5 experiments; bars, SEM.For all experiments with FMLP, a positive control was included confirminginhibition (88 to 97%) of PMA stimulation by 20 nM staurosporine.
100>j
DOm
50
20 60 200
STAUROSPORINE (nM)
Fig. 2. Effect of varying concentrations of staurosporine in the presence orabsence of PDBu on binding of '"I-EGF to primary mouse epidermal cells. Cellswere exposed to staurosporine at the indicated concentrations in low ( a: mediumfor I h at 37*C, and '"I-EGF binding was determined. Points, average of triplicate
determinations at each concentration. The average SEM was ±4%. •,with 200nM PDBu; O, without PDBu. Two other experiments gave similar results.
4647
Research. on August 25, 2014. © 1988 American Association for Cancercancerres.aacrjournals.org Downloaded from
CONTRASTING ACTIONS OF STAUROSPORINE
on binding affinity. The inhibition of EGF binding by PDBuwas a little decreased by staurosporine at high concentrations,although complete blockage was not observed.
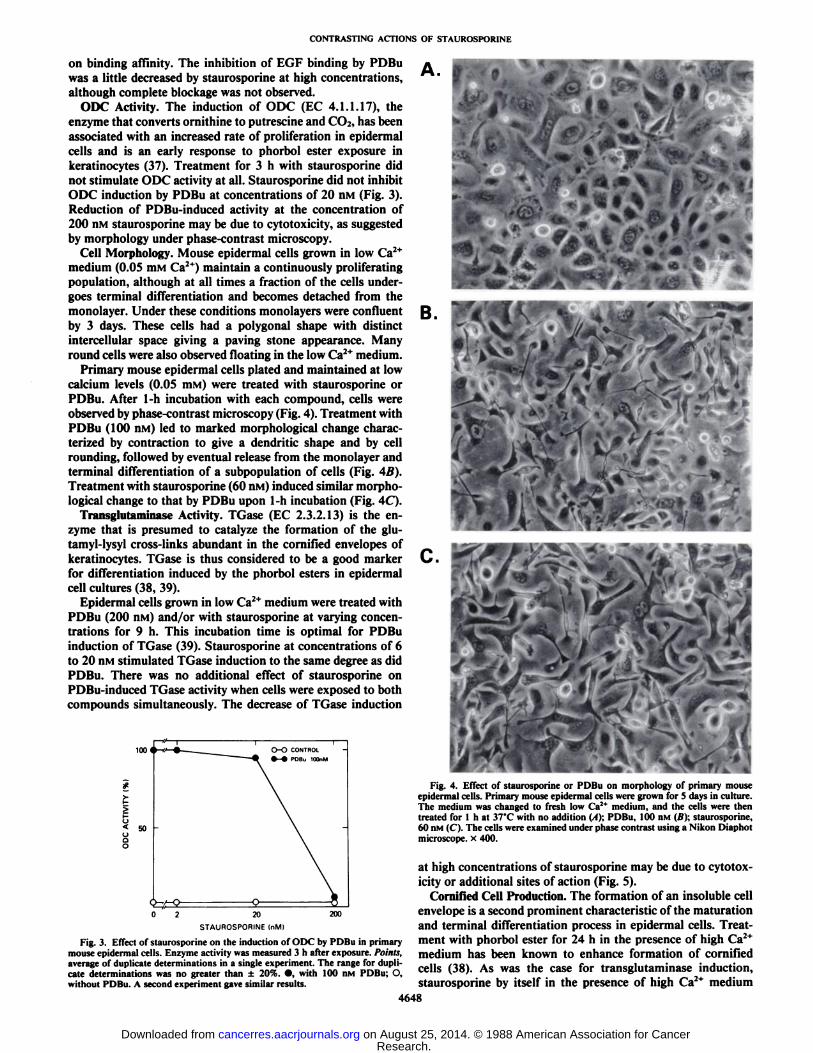
ODC Activity. The induction of ODC (EC 4.1.1.17), theenzyme that converts ornithine to putrescine and CO2, has beenassociated with an increased rate of proliferation in epidermalcells and is an early response to phorbol ester exposure inkeratinocytes (37). Treatment for 3 h with staurosporine didnot stimulate ODC activity at all. Staurosporine did not inhibitODC induction by PDBu at concentrations of 20 nM (Fig. 3).Reduction of PDBu-induced activity at the concentration of200 nM staurosporine may be due to cytotoxicity, as suggestedby morphology under phase-contrast microscopy.
Cell Morphology. Mouse epidermal cells grown in low Ca2+medium (0.05 mvi Ca2+) maintain a continuously proliferating
population, although at all times a fraction of the cells undergoes terminal differentiation and becomes detached from themonolayer. Under these conditions monolayers were confluentby 3 days. These cells had a polygonal shape with distinctintercellular space giving a paving stone appearance. Manyround cells were also observed floating in the low Ca2+ medium.
Primary mouse epidermal cells plated and maintained at lowcalcium levels (0.05 mM) were treated with staurosporine orPDBu. After 1-h incubation with each compound, cells wereobserved by phase-contrast microscopy (Fig. 4). Treatment withPDBu (100 nM) led to marked morphological change characterized by contraction to give a dendritic shape and by cellrounding, followed by eventual release from the monolayer andterminal differentiation of a subpopulation of cells (Fig. 4B).Treatment with staurosporine (60 nM) induced similar morphological change to that by PDBu upon 1-h incubation (Fig. 4C).
Transglutaminase Activity. TGase (EC 2.3.2.13) is the enzyme that is presumed to catalyze the formation of the glu-tamyl-lysyl cross-links abundant in the cornified envelopes ofkeratinocytes. TGase is thus considered to be a good markerfor differentiation induced by the phorbol esters in epidermalcell cultures (38, 39).
Epidermal cells grown in low Ca2+ medium were treated with
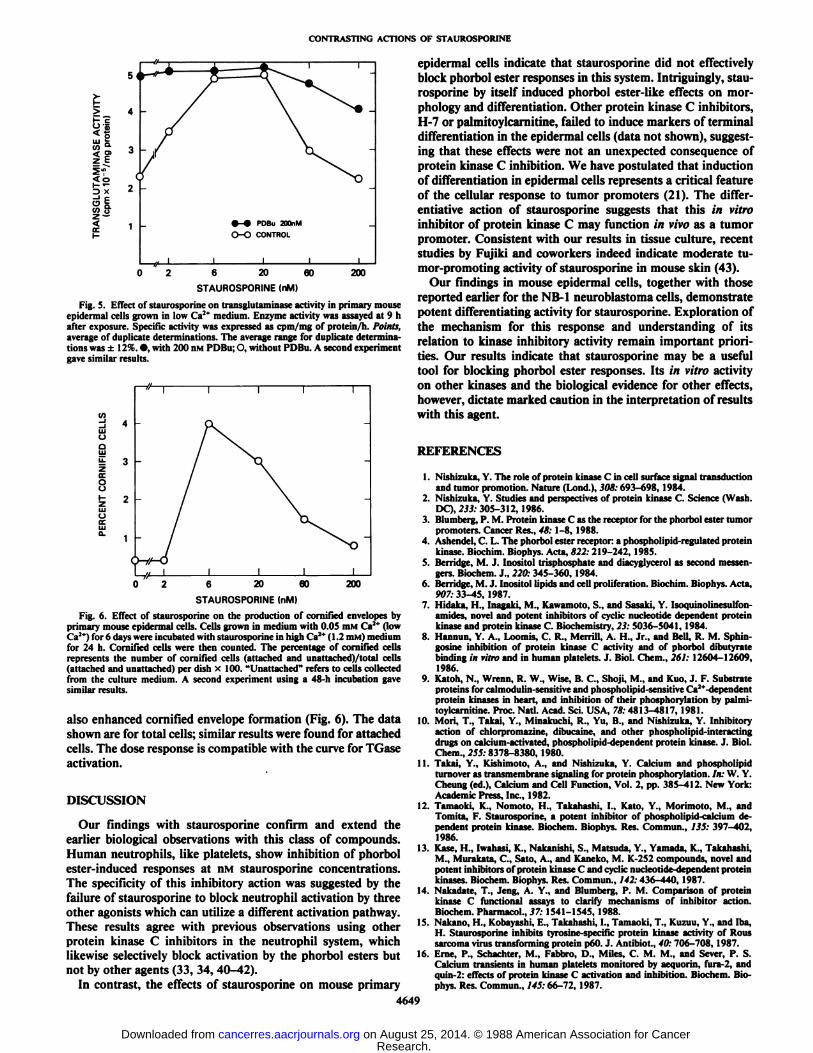
PDBu (200 nM) and/or with staurosporine at varying concentrations for 9 h. This incubation time is optimal for PDBuinduction of TGase (39). Staurosporine at concentrations of 6to 20 nM stimulated TGase induction to the same degree as didPDBu. There was no additional effect of staurosporine onPDBu-induced TGase activity when cells were exposed to bothcompounds simultaneously. The decrease of TGase induction
A.
20
STAUROSPORINE InM)
200
Fig. 3. Effect of slaurosporine on the induction of ODC by PDBu in primarymouse epidermal cells. Enzyme activity was measured 3 h after exposure. Points,average of duplicate determinations in a single experiment. The range for duplicate determinations was no greater than ±20%. •,with 100 nM PDBu; O,without PDBu. A second experiment gave similar results.
%Jy*- 'l *J 0 P
fröRT«?'*-**?*\•J--* _ ^_ *¿f/: *
»r,T'* aT^^^ifr*•^^•''•:B.
C.<f"V^fiiT^.\?*^i
r3M
Fig. 4. Effect of staurosporine or PDBu on morphology of primary mouseepidermal cells. Primary mouse epidermal cells were grown for 5 days in culture.The medium was changed to fresh low <V medium, and the cells were thentreated for l h at 37'C with no addition (A); PDBu, 100 nM (B); staurosporine,
60 nM (C). The cells were examined under phase contrast using a Nikon Diaphotmicroscope, x 400.
at high concentrations of staurosporine may be due to cytotoxicity or additional sites of action (Fig. 5).
Cornified Cell Production. The formation of an insoluble cellenvelope is a second prominent characteristic of the maturationand terminal differentiation process in epidermal cells. Treatment with phorbol ester for 24 h in the presence of high Ca2+
medium has been known to enhance formation of cornifiedcells (38). As was the case for transglutaminase induction,staurosporine by itself in the presence of high Ca2+ medium
4648
Research. on August 25, 2014. © 1988 American Association for Cancercancerres.aacrjournals.org Downloaded from
CONTRASTING ACTIONS OF STAUROSPORINE
6 20
STAUROSPORINE (nM)
60 200
Fig. S. Effect of staurosporine on transglulaminase activity in primary mouseepidermal cells grown in low Ca2* medium. Enzyme activity was assayed at 9 hafter exposure. Specific activity was expressed as cpm/mg of protein 'h. Points,
average of duplicate determinations. The average range for duplicate determinations was ±12%. •.with 200 nM PDBu; O, without PDBu. A second experimentgave similar results.
60 200
STAUROSPORINE (nM)
Fig. 6. Effect of staurosporine on the production of cornified envelopes byprimary mouse epidermal cells. Cells grown in medium with 0.05 IHMl V ' (lowCa2*) for 6 days were incubated with staurosporine in high ( V ' (1.2 rn\i) medium
for 24 h. CorniPied cells were then counted. The percentage of cornified cellsrepresents the number of cornified cells (attached and unattached)/total cells(attached and unattached) per dish x 100. "Unattached" refers to cells collectedfrom the culture medium. A second experiment using a 48-h incubation gavesimilar results.
also enhanced cornified envelope formation (Fig. 6). The datashown are for total cells; similar results were found for attachedcells. The dose response is compatible with the curve for TGaseactivation.
DISCUSSION
Our findings with staurosporine confirm and extend theearlier biological observations with this class of compounds.Human neutrophils, like platelets, show inhibition of phorbolester-induced responses at nM staurosporine concentrations.The specificity of this inhibitory action was suggested by thefailure of staurosporine to block neutrophil activation by threeother agonists which can utilize a different activation pathway.These results agree with previous observations using otherprotein kinase C inhibitors in the neutrophil system, whichlikewise selectively block activation by the phorbol esters butnot by other agents (33, 34, 40-42).
In contrast, the effects of staurosporine on mouse primary
epidermal cells indicate that staurosporine did not effectivelyblock phorbol ester responses in this system. Intriguingly, staurosporine by itself induced phorbol ester-like effects on morphology and differentiation. Other protein kinase C inhibitors,H-7 or palmitoylcarnitine, failed to induce markers of terminaldifferentiation in the epidermal cells (data not shown), suggesting that these effects were not an unexpected consequence ofprotein kinase C inhibition. We have postulated that inductionof differentiation in epidermal cells represents a critical featureof the cellular response to tumor promoters (21). The differ-entiative action of staurosporine suggests that this //; vitroinhibitor of protein kinase C may function in vivo as a tumorpromoter. Consistent with our results in tissue culture, recentstudies by Fujiki and coworkers indeed indicate moderate tumor-promoting activity of staurosporine in mouse skin (43).
Our findings in mouse epidermal cells, together with thosereported earlier for the NB-1 neuroblastoma cells, demonstratepotent differentiating activity for staurosporine. Exploration ofthe mechanism for this response and understanding of itsrelation to kinase inhibitory activity remain important priorities. Our results indicate that staurosporine may be a usefultool for blocking phorbol ester responses. Its in vitro activityon other kinases and the biological evidence for other effects,however, dictate marked caution in the interpretation of resultswith this agent.
REFERENCES
1. Nishizuka, Y. The role of protein kinase C in cell surface signal transductionand tumor promotion. Nature (Lond.), 308:693-698, 1984.
2. Nishizuka. Y. Studies and perspectives of protein kinase C. Science (Wash.DC), 233: 305-312, 1986.
3. Blumberg, P. M. Protein kinase C as the receptor for the phorbol ester tumorpromoters. Cancer Res., 48: 1-8, 1988.
4. Ashendel, C. 1 . The phorbol ester receptor: a phospholipid-regulated proteinkinase. Biochim. Biophys. Acta, 822: 219-242, 1985.
5. Berridge, M. J. Inositol trisphosphate and diacyglycerol as second messengers. Biochem. J., 220: 345-360, 1984.
6. Berridge, M. J. Inositol lipids and cell proliferation. Biochim. Biophys. Acta,907; 33-45, 1987.
7. Hidaka, H., Inagaki, M., Kawamoto, S., and Sasaki, Y. Isoquinolinesulfon-amides, novel and potent inhibitors of cyclic nucleotide dependent proteinkinase and protein kinase C. Biochemistry, 23: 5036-5041, 1984.
8. Hannun, Y. A., Loomis, C. R., Merrill, A. H., Jr., and Bell, R. M. Sphin-gosine inhibition of protein kinase C activity and of phorbol dibutyratebinding in vitro and in human platelets. J. Biol. Chem., 261: 12604-12609,1986.
9. Katoh, N., Wrenn, R. W., Wise, B. C., Shoji, M., and Kuo, J. F. Substrateproteins for calmodulin-sensitive and phospholipid-sensitive Ca2*-dependent
protein kinases in heart, and inhibition of their phosphorylation by palmitoylcarnitine. Proc. Nati. Acad. Sci. USA, 78:4813-4817, 1981.
10. Mori, T., Takai, Y., Minakuchi, R., Yu, B., and Nishizuka, Y. Inhibitoryaction of chlorpromazine, dibucaine, and other phospholipid-interactingdrugs on calcium-activated, phospholipid-dependent protein kinase. J. Biol.Chem., 255: 8378-8380, 1980.
11. Takai, Y., Kishimoto, A., and Nishizuka, Y. Calcium and phospholipidturnover as transmembrane signaling for protein phosphorylation. In: W. Y.Cheung (ed.). Calcium and Cell Function, Vol. 2, pp. 385-412. New York:Academic Press, Inc., 1982.
12. Tamaoki, K . Nomoto, H., Takahashi, I., Kalo, Y., Morimmo. M., andTornita, F. Staurosporine, a potent inhibitor of phospholipid-calcium dependent protein kinase. Biochem. Biophys. Res. Commun., 135: 397-402,1986.
13. Käse,H., Iwahasi, K., Nakanishi, S., Matsuda, Y., Yamada, K., Takahashi,M., Murakata, C., Sato, A., and Kaneko, M. K-252 compounds, novel andpotent inhibitors of protein kinase C and cyclic nucleotide-dependent proteinkinases. Biochem. Biophys. Res. Commun., 142:436-440, 1987.
14. Nakadate, T., Jeng, A. Y., and Blumberg, P. M. Comparison of proteinkinase C functional assays to clarify mechanisms of inhibitor action.Biochem. Pharmacol., 37: 1541-1545, 1988.
15. Nakano, H., Kobayashi, E., Takahashi, I., Tamaoki, T., Kuzuu, Y., and Iba,H. Staurosporine inhibits tyrosine specific protein kinase activity of Roussarcoma virus transforming protein p60. J. Antibiot., 40: 706-708, 1987.
16. Erne, P., Schachter, M., Fabbro, D., Miles, C. M. M., and Sever, P. S.Calcium transients in human platelets monitored by aequorin, fura-2, andquin-2: effects of protein kinase C activation and inhibition. Biochem. Biophys. Res. Commun., 145:66-12, 1987.
4649
Research. on August 25, 2014. © 1988 American Association for Cancercancerres.aacrjournals.org Downloaded from

CONTRASTING ACTIONS OF STAUROSPOR1NE
17. Morioka, II.. Ishihara, M., Shibai, II.. and Suzuki, T. Staurosporine-induceddifferentiation in a human neuroblastoma cell line MM. Agrie. Biol. Chem.,49: 1959-1964. 1985.
18. Oka, S., Kodama. M.. Takeda, H., Tomizuka, N., and Suzuki, T. Stauro-sporine, a potent platelet aggregation inhibitor from a Streptomyces species.Agrie. Biol. Chem., 50: 2723-2728, 1986.
19. Bush. J. A.. Long, B. H., Catino. J. }.. and Bradner. W. T. Production andbiological activity of rebeccamycin, a novel antitumor agent. J. Anulimi,. 40:668-678. 1987. "
20. Tauber. A. I. Protein kinase C and the activation of the human neutrophilNADPH-oxidase. Blood, 69: 711 -720, 1987.
21. Yuspa. S. H., Hennings, H., and Lichti, U. Initiator and promoter inducedspecific changes in epidermal function and biological potential. J. Cell.Biochem., 17: 245-257, 1981.
22. Parkinson, E. K. Defective responses of transformed keratinocytes to terminal differentiation stimuli. Their role in epidermal tumor promotion bypin u Imi esters and by deep skin wounding. Br. J. Cancer, 52:479-493,1985.
23. Borregaard. N.. and Tauber, A. I. Subcellular localization of the humanneutrophil NADPH-oxidase: ft cytochrome and associated flavoprotein. J.Biol Chem.. 259: 47-52, 1984.
24. Cohen, H. J., and Chovaniec, M. E. Superoxide generation by digitonin-stimulated guinea pig granulocytes: a basis for a continuous assay for monitoring Superoxide production and for the study of the activation of thegenerating system. J. Clin. Invest., 61: 1081-1087, 1978.
25. Hennings, H., Michael, D., Cheng, C, Steinert, P., Holbrook, K., and Yuspa,S. H. Calcium regulation of growth and differentiation of mouse epidermalcells in culture. Cell, 19: 245-254, 1980.
26. Lichti, U., and Gottesman, M. M. Genetic evidence that a phorbol estertumor promoter stimulates ornithine decarboxylase activity by a pathwaythat is independent of cyclic AMP-dependent protein kinases in CHO cells.J. Cell. Physiol., 7/3:433-439, 1982.
27. Sako. T., Yuspa, S. H., Herald, C. H., Pettit, G. R., and Blumberg, P. M.Partial parallelism and partial blockade by bryostatin 1 of effects of phorbolester tumor promoters on primary mouse epidermal cells. Cancer Res., 47:5445-5450, 1987.
28. Lowry, O. H., Rosebrough, N. J., Farr, A. L., and Randall, R. J. Proteinmeasurement with the Polin phenol reagent. J. Biol. Chem., 193: 265-275,1951.
29. Higuchi, M., and Yoshida. F. Lowry determination of protein in the presenceof sulfhydryl compounds or other reducing agents. Anal. Biochem., 77:542-547, 1977.
30. Strickland, J. E., Jetten, A. M., Kawamura, H., and Yuspa, S. H. Interactionof epidermal growth factor with basal and differentiating epidermal cells of
mice resistant and sensitive to carcinogenesis. Carcinogenesis (I.muÃ.).5:735-740, 1984.
31. Jetten, A. M. Retinoids specifically enhance the number of epidermal growthfactor receptors. Nature (Lond.), 284: 626-629, 1980.
32. Rice, R. H., and Green, H. The cornified envelope of terminally differentiatedhuman epidermal keratinocytes consists of cross-linked protein. Cell. II:417-422, 1977.
33. Maridonneau-Parini, I., Tringale, S. M., and Tauber, A. I. Identification ofdistinct activation pathways of the human neutrophil NADPH-oxidase. J.Immunol., 137: 2925-2929, 1986.
34. Maridonneau-Parini, I., and Tauber, A. I. Activation of NADPH-oxidase byarachidonic acid involves phospholipase A_-in intact human neutrophils butnot in the cell-free system. Biochem. Biophys. Res. Commun., 138: 1099-1105. 1986.
35. Murray, A. W., and Fusenig, N. E. Binding of epidermal growth factor toprimary and permanent cultures of mouse epidermal cells: inhibition bytumor-promoting phorbol esters. Cancer Lett., 7:71-77, 1979.
36. Lockyer, J. M., Bowden, G. T., Matrisian, L. M., and Magun, B. E. Tumorpromoter-induced inhibition of epidermal growth factor binding to culturedmouse primary epidermal cells. Cancer Res., 41: 2308-2314, 1981.
37. Lichti, U., Patterson, E., Hennings, H., and Yuspa, S. H. The tumor promoter12-O-tetradecanoylphorbol-13-acetate induces ornithine decarboxylase inproliferating basal cells but not in differentiating cells from mouse epidermis.J. Cell. Physiol., 107: 261-270, 1981.
38. Yuspa, S. H., Ben, T., and Hennings, H. The induction of epidermal trans-glutaminase and terminal differentiation by tumor promoters in culturedepidermal cells. Carcinogenesis (Lond.), 4: 1413-1418, 1983.
39. Yuspa, S. H., Ben, T., Hennings, H., and Lichti, U. Phorbol ester tumorpromoters induce epidermal transglutaminase. Biochem. Biophys. Res. Commun., 97: 700-708, 1980.
40. Naccache, P. H., Molski, M. M., and Sha'afi, R. J. Polymixin B inhibitsphorbol 12-myristrate 13-acetate, but not chemotactic factor, induced effectsin rabbit neutrophils. FEBS Lett., 193: 227-230, 1985.
41. Gerard, C., McPhail, L. C., Marfat, A., Slimier-Gerard, N. P., Bass, D. A.,and McCall, C. E. Role of protein kinases in stimulation of human polymor-phonuclear leukocyte oxidative metabolism by various agonists. J. Clin.Invest., 77: 61-65, 1986.
42. Sha'afi, R. I., Molski, T. F. P., Huang, C. K., and Naccache, P. I. The
inhibition of neutrophil responsiveness caused by phorbol esters is blockedby the protein kinase C inhibitor H-7. Biochem. Biophys. Res. Commun.,137: 50-60, 1986.
43. Fujiki, H., Suganuma, M., Suguri, H., Yoshizawa, S., Hirota, M., Takagi,K., and Sugimura, T. Diversity in chemical nature of and mechanism ofresponse to tumor promoters. In: T. J. Slaga (éd.).Dermal Carcinogenesis,Research Directions for Human Relevance. New York: Raven Press, in press,1988.
4650
Research. on August 25, 2014. © 1988 American Association for Cancercancerres.aacrjournals.org Downloaded from
Copyright © 2022 FDOKUMEN




















