
Restoration of CD44H Expression in Colon Carcinomas Reduces Tumorigenicity

106 American Entomologist bull Summer 2012
Ecologists have conceptualized trophic interactions that deter-mine organism abundance in terms of bottom-up (host plant) and top-down (natural enemies) forces (Hairston et al 1960
White 1978 Power 1992 Walker and Jones 2001) The approach stimulated a wide range of ecological research (Power 1992) A synthesis of the approach remains elusive (Walker and Jones 2001) but a consensus view is that these forces are not mutually exclusive (Lawton and McNeil 1979 Price et al 1980 Shrewsbury and Raupp 2008) It is clear that arthropod populations face a variety of environ-mental risks to which they must adapt that can be described within the bottom-up and top-down heuristic model Research continues to refine and determine the general application of the theory to explain risk handling and the importance of the factors and tradeoffs that are involved Behavior provides an organism a method for responding to environmental stimuli and tracking their changes temporally and spatially (Werner 1992) Our focus is on the potential behaviors that a xylophagous herbivore uses to mitigate the putative risks originating from these environmental forces (Mizell et al 2008)
Hamilton (1971) after Williams (1966) proposed that gregarious behavior of organisms benefited the individual simply by decreasing the probability of predation and in a similar manner nongregarious species also derived a selection benefit from proximity to others
Foster and Treherne (1981) described the phenomenon in fish called the ldquodilution effectrdquo whereby the probability of an individual being a victim is inversely proportional to the number of organisms present in a group Visual response to conspecifics is the implied behavioral mechanism that enables this process Besides the dilution effect derived from the context of gregarious behavior an individualrsquos perception of conspecifics makes it possible to collect a broad range of biological information that may strongly affect other fitness-related behaviors such as foraging success and movement (Lazarus 1979 Foster and Treherne 1981 Lawrence 1985 Stamps 1987 Turchin 1987 Valone 1989)
The glassy-winged sharpshooter Homalodisca vitripennis (Ger-mar) (Hemiptera Cicadellidae) is a primary vector of the bacterium Xylella fastidiosa (Wells et al [1973]) which is the causal agent of Piercersquos disease of grapevine and many other diseases of hardwood trees and herbaceous shrubs (Andersen et al 1989 1992 Redak et al 2004 Mizell et al 2008) As a xylem feeder H vitripennis feeds on a resource depauperate in nutrients and as a result has evolved many specialized physiological and behavioral adaptations such as extreme polyphagy high feeding rate high assimilation efficiency of ingested compounds excretion of ammonia and high vagility in all mobile life stages (Andersen et al 1989 1992 Redak et al 2004
Congener Response Reduces Risks from Bottom-Up and Top-Down Forces Behavioral Parsimony by a Xylophage
Russell F Mizell III Peter C Andersen Brent V Brodbeck and Wayne B Hunter
Abstract Organisms must find food and avoid enemies Homalodisca vitripennis (Germar) is a primary vector of Xylella fastidi-osa the causal agent of Piercersquos disease a deadly disease of grapevines and other plants H vitripennis feeds on xylem tissue depauperate in nutrients and has evolved many specialized physiological and behavioral adaptations Bottom-up mortality risks to H vitripennis include starvation and reduced fecundity through loss of quality hosts Top-down risks include predation egg parasitism by Hymenoptera and fungal pathogens Visual behavior of H vitripennis in response to congeners common heterospecifics and artificial models of conspecifics was investigated Behaviors and the physical properties of the visual cues involved were characterized H vitripennis adults exhibited highly significant positive responses to conspecifics a heterospecific species Oncometopia nigricans and artificial models the size of adult leafhoppers Significant response was not observed to a congeneric Homalodisca insolita with a smaller body size or to models of sizes shapes and colors other than black and red Cadavers of leafhoppers placed on a novel and nutritionally poor host plant stimulated H vitripennis adults to land on branches containing the cadavers H vitripennis also landed preferentially on branches containing conspecific cadavers despite the pres-ence of a cadaver of a major predator Anolis carolinensis lizards or artificial lures resembling spiders This consistent visual response was independent of variable bottom-up (host quality) and top-down (predator presence) risks and it suggests for the first time such parsimonious behavior in insects ie sole use of a live visual cue as a means to reduce mortality risks from bottom-up and top-down forces
ResearcH
107American Entomologist bull Volume 58 Number 2
Tipping et al 2004 Mizell et al 2008) They also contain bacte-rial symbionts thought to be involved in enhancing their nutrition (Wu et al 2006) Two other vector species Oncometopia nigricans (Walker) and H insolita (Walker) commonly occur on host plants with H vitripennis in Florida and Georgia O nigricans feeds mostly on hardwood trees and is often found feeding in heterospecific groups with H vitripennis (Turner and Pollard 1959) H insolita prefers to feed on grasses and although often infesting some of the same plants coincidently with H vitripennis [eg Johnson grass Sorghum halepense (L) Pers and pearl millet Pennisetum glaucum (L) R Br] the two species are not observed routinely feeding closely together or behaving synchronously in multispecies groups as with O nigricans (Turner and Pollard 1959)
H vitripennis and related vectors are strong flyers with keen vi-sion that allows them to detect approaching objects from asymp5 to 10 m distance H vitripennis and other related leafhopper species often feed in a stereotypical head-down posture (Bextine et al 2005) ag-gregated in groups of individual or multiple species on only a few of the many branches available on host plants When disturbed leafhop-pers in such aggregations often respond synchronously by moving en masse away from the stimulus to better conceal their bodies behind the plant structure on which they are feeding Moreover disturbed leafhoppers that fly from the host plant often return immediately and land on the branches near feeding leafhoppers Trap catch of male H vitripennis can be increased by adding cadavers of conspecifics to traps (Mizell et al 2009) In the presence of cadavers the distribution pattern of trapped leafhoppers becomes aggregated near a cadaver Together these behaviors suggest visual response by leafhoppers to conspecifics and perhaps heterospecifics
Bottom-up mortality forces that decrease relative fitness of these xylophagous insects include starvation and reduced fecundity through loss of quality host plants Top-down forces include a high rate of egg parasitism by Hymenoptera (Triapitsyn et al 1998 Lopez et al 2003) fungal pathogens that infect nymphs and adults (Walker and Jones 2001 Boucias et al 2006 Mizell et al 2008) and predation Dragonflies and related visually orienting aerial predators prey on fly-ing adults lizards spiders assassin bugs and damsel bugs threaten feeding and ovipositing adults (Lopez et al 2003 Daane et al 2004 Fournier et al 2008) and the eggs may be consumed by tree crickets Cyrtoxipha spp (Lopez et al 2002) Parasitism by Hymenoptera is also high to eggs (Triapitsyn et al 1998 Lopez et al 2003) and by fungal pathogens that infect nymphs and adults (top-down) (Walker and Jones 2001 Boucias et al 2006 Mizell et al 2008) Mizell et al (2008) developed a behavioral model that provides a conceptual framework to address the critical decisions and cues used by H vitripennis to increase its fitness in response to these putative risks That model does not deal with the potential behaviors that may be associated with the known acoustical vibration communication used by H vitripennis (Percy et al 2008) however methods using cadavers in this study eliminate the confounding effects potentially derived from acoustical communication
Response to conspecifics as a mechanism to increase an organ-ismrsquos survival and fitness has been reported in a diversity of species including fish (Foster and Treherne 1981 Ward et al 2008) birds (Lazarus 1979 Lawrence 1985 Sandlin 2000) lizards (Stamps 1987 1988) and insects (Thompson et al 1987 Turchin 1987 Muller 1998 Dukas 2001 Dukas and Morse 2003) Such ldquosocially acquiredrdquo information is used for foraging habitat assessment opponent assessment and mate choice (Geraldeau et al 2002 and
literature therein) Group foraging is only one of several behavioral situations whereby organisms derive benefit from ldquopublic informa-tionrdquo provided by conspecifics (Valone 1989) An opposing strategy territorial behavior also illustrates the importance of response to conspecifics because gaining defending and maintaining a terri-tory directly affects fitness Thus various responses to conspecifics can mediate bottom-up and top-down factors (Stamps 1987 1988)
The underlying behaviors of insects in response to conspecifics are reported for several taxa Drosophila melanogaster Meigen are attracted to long vertical objects but repelled by small ones (Mainman et al 2008) a repertoire that likely aids the recognition of landing sites and avoidance of predators and other aerial objects Houseflies Fannia canicularis L chase congeners and hoverflies pursue passing objects in the quest for mates (Land and Collett 1974 Collett and Land 1978) The locust Schistocerca gregaria (Forsk) can respond to stripe patterns (Baker 1979) Wood crickets assess danger from spider predators by observing conspecificsrsquo cues and then adopting a stereotypical predatorshyshy-avoidance behavior that is maintained after the conspecifics are gone (Coolen et al 2005) Honey bees Apis mel-lifera L avoid flower locations where they have escaped a predator and change foraging behavior in response to dead bees or spiders (Dukas 2001) To our knowledge however use of conspecifics by insects to mediate bottom-up and top-down factors has never been reported
The visual behavior of H vitripennis in response to conspecif-ics common heterospecifics their potential predators and their responses to artificial models of the insects and their predators were investigated The objectives were to characterize how leafhoppers re-spond to congeners and a heteropecific determine the putative cues associated with recognition and evaluate how these behaviors are applied in mitigating bottom-up and top-down environmental forces
Methods and MaterialsInsects and Plants Cadavers of H vitripennis and other leafhop-
pers species used in the experiments were collected from natural field populations in north Florida found on crape myrtle Lagerstroemia indica Lindl and holly Ilex spp concurrently with the studies In-sects were captured and placed in a freezer and held until needed L indica cultivars are the most frequently used hosts of H vitripennis during the months of JunendashAugust in north Floridandashsouth Georgia Texas and California (Mizell and French 1987 Brodbeck et al 1990 Mizell et al 2005 Northfield et al 2009) By using cadavers rather than live insects in the experiments the potential confounding ef-fects from behavioral communication such as acoustical signaling were eliminated
All experiments were conducted from June to September of 2006ndash2011 at the NFREC (North Florida Research and Education Center)ndashQuincy in north Florida in the field with naturally occurring populations of H vitripennis on various cultivars of crape myrtle L indica or in several 3 times 10 m plantings of sunflower Helianthus an-nus L H vitripennis density on crape myrtle in north Florida usually varies from 1 to 15 individuals per branch at moderate population levels (Brodbeck et al 1990 Mizell et al 2008) In sunflowers a good but less favored host than crape myrtle the number of H vitripennis averaged 35 leafhoppers per plant over the 6 d in that experiment At 1ndash2 h intervals visual counts (the statistical replicates) were made of H vitripennis that landed on treated and untreated branches Counts were recorded from 1000 to 1700 hours when leafhop-per counts are highest (Mizell et al 2008) After counting and to
108 American Entomologist bull Summer 2012
begin the next observation period all responding leafhoppers were induced to leave the plants by shaking the branches These leafhop-pers moved to other branches or to nearby plants Thus the count of the subsequent replicate consisted of new leafhoppers or (much less likely) previous leafhoppers making a new and independent behavioral choice The sex of the responding leafhoppers was noted and recorded when possible
Response to Conspeciics Regular white plastic drinking straws were used in the first experiments to deploy leafhopper cadavers The straws were slit lengthwise on one side to permit them to be fitted snugly over branches H vitripennis cadavers were glued to the straws with hot glue in the lab before field set up a cadaver was placed at each end of the straw and in the middle Several experiments were conducted in this manner by identifying two or more branches po-sitioned ge 50 cm apart on lsquoNatchezrsquo crape myrtle Branches for the experiment were selected for the treatments by assessing where leafhoppers were generally found feeding at the time (Mizell and French 1987) and then placing treatments at those points on other branches In the first experiment the treatments consisted of an untreated control four and eight H vitripennis cadavers per branch A second experiment was conducted using zero three six and nine leafhopper cadavers per branch
Colored Models A third test was conducted using plastic models of the average size of an adult H vitripennis (12 times 3 mm) The models were made from L-shaped pieces cut from the bottom of white plastic yogurt cups The models tested were painted black white or green (Premium Deacutecor decorative enamel General Paint and Manufactur-ing Co Cary IL) and tested along with a fresh H vitripennis cadaver and an untreated control The base of the L-shape model was used to glue the plastic models to the branches such that a silhouette of asymp3 mm was visible to represent the leafhopper bodies Counts were made on the treatment branch and either one or two other branches within ge 50 cm of the main treatment branch
Glued Cadaver Experiments Crape myrtle and sunflower field plantings with populations of leafhoppers were used as host plants Adjacent branches on crape myrtle or main branches on two adjacent sunflowers were chosen and marked in pairs such that they were ge 50 cm apart One branch or plant served as the treatment and the adjacent branch or plant as the untreated control Test branches were maintained in a natural state as much as possible but some leaves were removed similarly on each of the paired branches to facilitate the leafhopper count Treatments were placed directly on parts of the host plants where leafhoppers were typically feeding All wild leafhoppers were removed from the treatment branches to begin the test and after each observational replicate In these later experiments a rapidly drying clear glue (Aleenersquos Fast Grab Tacky Glue Duncan Enterprises Fresno CA) was used to glue the treatments directly on the host branches In all treatments with models and leafhopper cadavers a group of three individuals attached asymp8‒10 cm apart was used The experiments were conducted for several days with treat-ment leafhoppers maintained or replaced to represent naturally occurring leafhoppers as closely as possible
Heterospeciics Tested Adult H insolita were collected using a sweep net or handheld aspirator from grasses and Oncometopia nigricans from Ilex spp near the experimental sites and then placed in a freezer until they were used Heterospecific cadavers were glued onto branches in groups of three and handled as described for H vitripennis
Effects of Acoustical Communication on Response to Cadavers
As described before two branches on crape myrtle separated by ge 50 cm were used for the experiment Both branches received three cadavers asymp8‒10 cm apart located asymp50 cm from the branch tip A sleeve cage asymp30 cm long was added onto the tip of the treated branch above the cadavers Three twist ties were used to form two cages from the sleeve and three live male and three female H vitripennis were added but were segregated by sex into one of the sides of the sleeve to provide potential acoustical communication signals to the treat-ment branch Landed adult H vitripennis were counted at 30‒60 min intervals and compared between the treated and control branches
Colors and Enhanced Models Leafhopper cadavers were sprayed gray (pathogen cadaver model) red or black with acrylic paint (as described before) in the laboratory and allowed to dry They were used in the field as described above Models close to the average size of an adult H vitripennis and larger were cut out of a black foam sheet (3 mm thick) (Darico Foamies Darico Strongville OH) in the following sizes (mm) 3 times 12 (leafhopper dimensions) 10 times 10 10 times 20 and 10 times 30 (Fig 1) A red model in the leafhopper size was also tested Models were glued directly to branches in groups of three as before Rectangular models were tested with the long axis oriented either in the vertical or horizontal plane The leafhopper model was also tested for the effect of contrast by placing it in the middle of a 25 mm square block of white cardboard attached to the branch which served as a background
Bottom-up Baiting of a Poor-Quality Host Plant with Conspecif-ics Lagerstroemia limii Merrill is a species of crape myrtle related to L indica that was imported to the United States from Southeast Asia for cross-breeding In north Florida this species is represented to our knowledge by only four specimens located together at the NFREC‒Quincy Therefore it represents a novel host plant to which H vitripennis would not have been previously exposed except by
Fig 1 Types of models that were evaluated for response by the glassy-winged sharpshooter H vitripennis A Live H vitripennis in natural feeding position and aggregation B H vitripennis cadavers aligned in the experimental positions C Horizontally confi gured leafhopper-sized felt model D Square 10 times 10 mm felt model E Vertically oriented felt model F Horizontally oriented felt model G Felt model on white card for contrast H Vertically oriented model showing leafhopper feeding nearby
109American Entomologist bull Volume 58 Number 2
the local population It also represents a host of ldquopoorrdquo nutritional quality as indicated by the very low number of naturally occurring H vitripennis observed on the plants during the course of the experi-ments (Brodbeck et al 1990) Three open-pollinated seedlings of the same size (lt3 m in height) were available side by side in a row of L indica cultivars and other ornamental plant species The plants had been pruned the spring before the study and each of the L limii plants consisted of about 30 parallel vertical branches asymp2 m long above the bottom stumps of asymp1 m Cadavers of H vitripennis were placed on 10 branches randomly selected from the three L limii plants available as described before Before the treatments were deployed and at each recorded observation all of the available branches were observed for the presence of H vitripennis At no time were there more than four naturally occurring H vitripennis adults observed on the three test plants other than on the 10 branch pairs in the experiment Moreover the leafhoppers occurring naturally on L limii were all single individuals Unlike L indica and sunflowers L limii was fed upon naturally only sparingly if at all by H vitripennis This strongly indicates that this exotic novel host plant also ranked as a very poor to non-host during the time of the experiment (Brodbeck et al 1990 Andersen et al 2005)
Top-down Predators and Predator Models Predaceous lizards the green anoles Anolis carolinensis Voigt were captured locally in north Florida around buildings euthanized using carbon dioxide and held in sealed plastic bags in a freezer before use Individual specimens were only used twice in the field on two successive days for a total of asymp12 h The lizards were attached to treatment branches in their natural hunting positions using fine wire Lizards were tested with and without the addition of leafhopper cadavers as attractants In addition brown artificial bass fishing lures resembling lizards (Su-per Salt Plus 18 cm lizard Chart pumpkin 002-012 Zoom Bait Co Bogart GA) or green or purple 22 cm worms (Classic Manufacturing Co Clermont FL) were used as predator model treatments Artifi-cial fly fishing lures that resemble spiders were purchased (Gander Mountain Ocala FL brookside flies panfish assortment) and used as models The spider lures were of several colors black or yellow bod-ies with black yellow or white legs Lures were mounted singularly on the treatment branches in plain sight by using the attached hook to embed them in a treatment branch in a natural position Cadavers were not combined in the tests with these treatments
Statistics Five to 10 combinations of the treatment‒control pairs were set up in the field and observed repeatedly in all of the tests to provide the statistical replicates After each observation the responding H vitripennis were removed from the treatment branches to reset the counts to zero for the next replicate In all tests H vit-ripennis cadavers were included as the standard treatment The data were analyzed using Proc GLM procedures in SAS 93 (SAS Institute 2009) or the Mann‒Whitney Rank Sum test in SigmaPlot 110 (Systat Software 2009) when the data did not meet the requirements of normality and heterogeneity of variance The probability used for statistical significance was 005
ResultsConspeciic and Heterospeciic Cadavers Adding H vitripen-
nis cadavers to branches significantly increased the response of H vitripennis to the branches containing the cadavers The presence of the straws used only in the early experiments did not appear to influence the visual response (Fig 2 3 and 4 Table 1) The presence of three cadavers however maximized the response because adding
four to nine cadavers did not significantly increase the number of landing H vitripennis Although numerically more were observed on the treatment H vitripennis did not significantly respond to cadavers of H insolita a species about half the size of H vitripennis (Table 1) H vitripennis responded similarly to cadavers of conspecifics and the heterospecific O nigricans a species that is similar in size but differs in color from H vitripennis (Table 1) H vitripennis also responded significantly to H vitripennis cadavers painted red which is much of the body color of newly emerged adults and to cadavers painted gray as models of fungi-infected cadavers (Boucias et al 2006) (Table 1)
Fig 3
Fig 2 Mean number and SE of H vitripennis responding to varying numbers of leafhopper cadavers placed on straws on a L indica branch Letters indicate signifi cant differences P lt 005
Fig 3 Mean number and SE of H vitripennis responding to varying num-bers of leafhopper cadavers placed on straws on a crape myrtle branch and two branches within 50 cm of the main treatment branch Letters indicate signifi cant differences for comparisons within each branch type Where letters are omitted for a branch type there were no signifi cant differences P lt 005
110 American Entomologist bull Summer 2012
Effects of Acoustical Communication on Response to Cadavers Adding live H vitripennis and presumably intraspecific acoustical signals to the treatment branch significantly increased leafhopper response rate relative to the cadaver control by 53 (141 for treat-ment versus 92 on control)
Models H vitripennis response to plastic or felt models was dependent on color size and orientation (Table 1) H vitripennis responded significantly to models of several colors including black (Fig 1 and 3) and red (Table 1) Red would likely appear to these leafhoppers as dark gray or near black (Table 1) Significant response was observed to a black model similar in size to H vitripennis (3 times 12 mm) and to a model of 10 times 10 mm Models gt12 mm oriented verti-cally or horizontally did not elicit a significant response (Table 1)
Bottom-upmdashNovel Poor or Non-host Plant A significant re-sponse was observed to a treatment of three cadavers placed on the branches of the novel and poor-quality host L limii (Table 1) Total counts for all control branches never exceeded four and these were always single leafhoppers
Top-downmdashPredators H vitripennis were not repelled by the presence of any tested predator or predator model (Table 1) Lure treatments presented without cadavers using a frog a yellow or black spider received 25 on the treatments and 4 on the control and were not significantly different (data not shown) Presentations of lizard predators together with H vitripennis cadavers elicited a significant response similar to that of H vitripennis cadavers alone This further indicates a lack of response to predators and positive response to
conspecifics even when in the visual presence of actual predators or predator-like models
DiscussionH vitripennis responded strongly to the visual presence of H
vitripennis cadavers Moreover low numbers of cadavers (three)
Fig 4 Mean number and SE of H vitripennis (GWSS) responding to cadavers and colored models similar in size to its body size Letters indicate significant differences P lt 005 for comparisons between treat-ments by branch
Fig 4
Table 1 Response of the H vitripennis to various visual treatments placed on host plants
TreatmentsMain Branch
No H vitripennis Responding
Adjacent BranchNo H vitripennis Responding
(P value MannndashWhitney U test) [replicatesa]
MannndashWhitney U Statistic
H vitripennis 232 76 (0001) [165] 52395H insolita 63 30 (0998) NS [65] 21125Oncometopia nigricans 197 41 (0001) [129] 25920H vitripennis painted red 69 44 (gt 005) NS [60] 17915H vitripennis painted gray (fungus) 101 42 (0001) [76] 15470
Novel non-host plantL limii 73 36 (0001) [93] 28890
Predator modelsDead anole lizard + 3 H vitripennis cadavers 23 13 (0008) [19] 4130
Felt modelsBlack model = asympH vitripennis 3 times12 mm 108 33 (0001) [87] 15380
Black model = H vitripennis (horizontal) 37 58 (gt007) NS [71] 21145
Black model 10 times10 mm 81 45 (lt0001) [83] 22145Black model 20 times20 mm 28 31 (0241) NS [38] 5840Black model 10 times30 mm vertical 74
66 (0280) NS [69] 21425
Black model 10 times30 mm horizontal 38
40 (0646) NS [55] 29825
Black model 3 times12 mm on white 2 times2 43
36 (0262) NS [50] 10995
Red model 3 times12 mm 127 34 (lt0001) [65] 6045 Acoustical communication Live H vitripennis 141 92 (lt0004) [115] 152675
The numbers are the total leafhoppers that responded to all replicates within each comparison The statistical comparisons were performed on the actual paired numbers of leafhoppers in all of the replicates that responded to the main branch compared with the adjacent branch containing each of the treatments in the individual rows The P values indicate the probability of a larger MannndashWhitney Rank Sum statistic by chance aReplicates = number of pairs observed where count on at least one of the paired treatments was ne 0
111American Entomologist bull Volume 58 Number 2
maximized the behavior The test cadavers were placed asymp8ndash10 cm apart with the total three-cadaver presentation providing a visual cue le20 cm wide The lack of increase by additional cadavers may indicate that leafhopper visual breadth is lt30 cm H vitripennis also responded to cadavers of the similarly sized heterospecific O nigricans that is often found feeding on the same host plants (ie Ilex spp) in bi-species groups The congeneric species H insolita which is much smaller and predominantly a grass feeder however did not elicit a positive response Although H vitripennis do feed on some grass and herbaceous species concurrently with H insolita we have never observed the two species feeding in heterospecific groups or on the same part of a plant Brodbeck et al (1993) reported that the dietary needs for both species were similar although they prefer to feed on different plant tissues (Turner and Pollard 1959) Percy et al (2008) compared the acoustical signals of H vitripennis and the congener H liturata Ball and reported that the signals were different in most parameters measured Some overlap however was observed such that it cannot be ruled out that acoustical signals produced by one Homalodisca sp could be detected by a related species If there is some signal overlap between the two species then it is possible that H vitripennis could derive benefit from detecting signals produced by H insolita acoustically and visually from group defensive movements that are similar to those of H vitripennis
In an attempt to define the physical parameters of the visual cues perceived and used by H vitripennis a group of models of different sizes and orientations on the host plant were tested A black model of 3times12 mm roughly equal to the body dimensions of an adult H vitripennis evoked significant visual attraction as did a 10 times10 mm
black felt model All other larger models in any plane did not evoke significant response when placed either vertically or horizontally on the plant No response was observed to a black model affixed to a contrasting white square card These results along with the lack of response to predators and the smaller congener H insolita suggest that the visual cues used by H vitripennis when in flight are relatively narrowly defined to the size close to its own body dimensions (width le10 times length le12 mm) The heterospecific O nigricans falls within this size range The failure of H vitripennis to respond to the model on the white card along with the orientation limits may indicate that detection of the visual cues from congeners is contextually related in some manner to that provided as background by the branch or its silhouette in space
Brodbeck et al (1990) and Andersen et al (2008) showed that H vitripennis abundance on a plant is highly correlated with leafhopper feeding rate and is indicative of the underlying xylem nutritional qual-ity H vitripennis are normally highly aggregated in the field among and within available host species (especially on higher quality host plants) although there is never a time when 100 of the population is observed on one host species As a rule in north Florida only a low percentage of branches on fair-to-good hosts are infested and they usually contain more than one leafhopper per branch (Mizell and French 1987) Significant H vitripennis response to cadavers when they were placed on the novel and poor host plant L limii provides very strong support for the use of conspecifics as visual cues by H vitripennis in flight to locate potential host plants for bottom-up risk mitigation Furthermore H vitripennis will respond visually to conspecifics independent of host plant quality and familiarity with a host plant species This result supports the idea that H vitripennis do not assess host plant quality before landing and furthermore that this pivotal decision is likely made using gustation at the level of the
xylem tissue (Hunter and Ullman 1994 Hunter et al1996 Backus et al 2005) The role of plant volatiles remains to be fully described but the research of Patt and Setamou (2007 2010) suggests a potential role in H vitripennis behavior
The importance of bottom-up nutrient profiles on fitness of these leafhoppers is well described (Redak et al 2004) Leafhopper ag-gregation however may be of further benefit by inducing chemical changes in host plant physiology that affect fitness such as feeding rate and fecundity (Karban and Baldwin 1997) Andersen et al (2003 2009) addressed this possibility in experiments with five host species They found that moderate-to-large plants of crape myrtle L indica L and plum Prunus salicina Lindl exhibited no increasing effects on nutrient compounds of leafhopper feeding in large numbers nor did leafhopper feeding behavior change when in groups In small soybean plants Glycine max (L) some nutrients were highly reduced (Andersen et al 2003) In citrus H vitripennis feeding increased seven amino acids in protein form but little change in protein amino acids was observed in Vitis vinifera lsquoChardonnayrsquo In citrus and lsquoChardonnayrsquo grape amino acid increased assimilation efficiency as a result of leafhopper feeding These data suggest that H vitripennis feeding does not induce metabolically inhibitory profiles in the xylem (Andersen et al 2009) This along with the induction of proteins and a slight decrease in free amino acids indicates that there may be some induced benefit in nutrition from H vitripennis aggregation (Andersen et al 2009)
Predation is an important mortality risk to H vitripennis in flight and on the host plant during feeding and oviposition (Lopez et al 2003 Daane et al 2005 Suttle and Hoddle 2006 Fournier et al 2008 Mizell et al 2008) Spiders and lizards are the most impor-tant predators of H vitripennis on the plants (Lopez et al 2003 Fournier et al 2008) Top-down risk mitigation by H vitripennis was assessed indirectly by testing visual response by H vitripennis to lizard predators and simulated predator models (fishing lures) that closely resembled living predators The experimental context most closely approximates the risk to H vitripennis during landing on a new branch the time when little or no information on predator presence on the selected branch is available to arriving H vitripennis This is also the time when H vitripennis presents the largest moving visual stimulus to hidden predators Once on the plant landed H vitripennis stop moving and provide a different visual stimulus to would-be predators they also regain the use of positional camouflage and their own visually based longer-range avoidance defense with the protection it provides As described earlier this protection is partially accrued from conspecifics via the ldquodilutionrdquo effect (Foster and Treherne 1981) Similarly heterospecifics present would also contribute to defense
The only other time that H vitripennis move excessively on the host plant is when the males seek mates and the females oviposit (Mizell et al 2008) Although copulation occurs mostly during the scotophase one might hypothesize that the presence of conspecif-ics would also reduce the risk from predation to more vulnerable copulating pairs and ovipositing females This may also be a selec-tion factor that induces H vitripennis females to oviposit on plants upon which they feed but which do not meet the nutritional needs of their nymphs (Brodbeck et al 2007 Mizell et al 2008) The ability to recognize conspecifics as indicators of suitable host plants allows H vitripennis to feed without moving and eliminates search flights for hours or days proportionally reducing in-flight and landing preda-tion risks These behaviors underscore the potential risks involved in
112 American Entomologist bull Summer 2012
finding hosts the tradeoffs faced by female leafhoppers and the value of conspecific recognition in their mitigation An unusual number of mated leafhopper pairs also were observed among the cadaver treatment branches on the mornings of succeeding test days These observations provide further support for the contention concerning top down risk mitigation via conspecific response
Significant response to avoid predators and artificial models was not observed It is unlikely that an arthropod herbivore would be able to develop a search image with such elasticity that it could encompass the wide range of natural enemies potentially encoun-tered The immediate acceptance of the novel and poor host L limii provides further evidence against the occurrence of a learned host image by searching leafhoppers Response to conspecifics would obviate the necessity of developing a predator search image because conspecifics would usually be present only where predators were absent Thus a type of ldquoenemy-free spacerdquo (Jeffries and Lawton 1984) may be represented by conspecific presence Alternatively relative to the risk of landing on a branch without conspecifics even if the conspecific signal was not entirely truthful about the level of risk the dilution effect from conspecific presence would provide increased protection theoretically in direct proportion to the number of con-specifics present Although foragers in many animal groups have an inherent geometry that causes information asymmetries individuals get more information from closer neighbors (Fernandez-Juricic et al 2004) H vitripennis uses keen visual acuity that likely is coupled with acoustical communication as a mechanism to respond rapidly en masse to approaching predators As a result leafhoppers accrue a more equivalent gain from defensive movement coordination (Fernandez-Juricic et al 2004)
On the host plant H vitripennis can detect and respond (and routinely do so in concert perhaps mediated by acoustic commu-nication) to movement of large objects approaching them from off the plant and to smaller objects on the plant Live lizards do elicit a response from feeding H vitripennis as they move over the host plant and approach leafhoppers (Lopez et al 2003) This suggests the occurrence in H vitripennis of a general visual capability (differ-ent from the visual response to conspecifics) to recognize moving objects that would circumvent the necessity of developing an array of species-specific predator search images
H vitripennis responded significantly to cadavers painted gray that served as a model of H vitripennis infected with the parasitic fungal pathogen H homalodiscae (Boucias et al 2006) H vitripennis cadavers infected with this pathogen remain attached for months on the host plant at feeding sites in the feeding position and infect H vitripennis nymphs and adults The infected cadavers are covered by the fungal mycelia that sporulate in cycles and provide a means to infect uninfected feeding H vitripennis (Lietze et al 2010) H homalodiscae-infected H vitripennis attach tenaciously via rhizoids to the plant bark in the summer and are often found attached to this substrate the following spring This positive visual response may increase transmission of the fungus to H vitripennis
When live presumably acoustically active H vitripennis were added to treatment branches the numbers of H vitripennis increased by 53 on the treated branches (Table 1) H vitripennis unlike most insect herbivores has not been shown to use olfaction at any level for courtship or mating In contrast H vitripennis and many Cicadel-lidae and related insect families use acoustical vibrations propagated through the host plant to communicate during courtship and mating and for other functions (Cocroft 2006) This behavioral modality of
H vitripennis however has received only cursory attention (Percy et al 2008) and its potential effect was eliminated in all except this one experiment through the use of cadavers
With respect to host finding the role of olfaction remains un-clear Patt and Setamou (2007) reported that the response to visual cues by H vitripennis nymphs and adults was enhanced by exposure to plant volatiles Patt and Setamou (2010) demonstrated that H vitripennis nymphs could be conditioned to respond to vanilla as a novel host volatile and as a result they argued that associative learning of chemical profiles could enhance leafhopper location and recognition of host plants These authors also speculated on a potential mechanism use of plant volatile profile emissions (Ninemets et al 2002 2004) in host finding and recognition by adult H vitripennis While an interesting hypothesis much research remains to be done to support these ideas A strong response to conspecifics provides a complimentary or alternative mechanism that would eliminate the need for such highly evolved behaviors However one may hypothesize that the presence of plant volatiles could stimulate a more intense leafhopper visual search for conspe-cifics on the host Nevertheless the ready acceptance of the novel and poor host L limii in the presence of conspecifics argues against the need for leafhoppers to learn a visual or olfactory pattern for specific plant species
Trivial movement within and between host plants and host plant patches is germane to understanding the distribution and fitness of H vitripennis (Mizell et al 2008) H vitripennis are strongly visually oriented while feeding on a host plant they can detect movement from 5 to 10 m away Based on the ostensibly narrow sensory ca-pabilities of H vitripennis (lack of pheromones apparently limited use of olfaction) it is not unexpected that H vitripennis would have evolved visual behavior with multiple functions Moreover this simple capability would obviate the need for higher level behaviors such as associative learning at least for adults (Patt and Setamou 2010) Profiles of xylem chemistry can change as much as 10-fold in one day or from day to day in the absence of any overt visual signals to leafhoppers (Andersen et al1992 Brodbeck et al 2010) Because host plant nutrient quality varies unrecognizably to leafhoppers in time and space and between and within a plant species developing a search image to visually identify specific host plants or character-istics related to their quality appears to be extremely difficult and perhaps of little value Moreover host plants usually do not occur in monocultures a situation that would reinforce short-term pattern recognition By contrast delay in finding hosts within a patch can affect information retention (McLinn and Stephens 2006) Relying too heavily on recent experience can be costly especially if the environment and in this case host plant nutrients change rapidly (Mizell and French 1987 Andersen et al 1989 1992 1993 1995ab Brodbeck et al 1990 19951999 McLinn and Stephens 2006) Animals devalue unreliable signals Models predict signal following when the signal reliability exceeds environmental certaintymdashso that information is valuable Signal value however is not based solely on reliability it also depends on the statistical distribution of the environmental states (McLinn and Stephens 2006) Plant quality and predation risks are in constant flux in the case of H vitripennis and extremely variable in time and space H vitripennis handling of bottom-up and top-down forces is tied closely together because the potential payoff and consequences from both accrue on the host plant following a single decision to land H vitripennis appar-ently can make a decision to land based solely on a visual response
113American Entomologist bull Volume 58 Number 2
to conspecifics on host plant branches An arrestment response is then likely reinforced on the plant via acoustical communication as gustation behavior is initiated
Thus presence of conspecifics indicates a ldquosafety zonerdquo for host plant testing and likely provides high-quality (truthfulness of signal) information because all H vitripennis in any one location are under more or less the same dietary constraints Notwithstanding that O nigricans may have somewhat different nutritional requirements it may be important in the mitigation of bottom-up and top-down forces The presence of one or (usually) multiple conspecifics is likely a signal indicating a reasonably high threshold quality of nutrients or else the duration of conspecifics remaining on a host plant would be short This perhaps explains why three conspecifics analogous to the quorum decision of Ward et al (2008) maximize the response Three may be enough to compensate for any variations in individual hunger rate or other uninterpretable behaviors (females not feeding or ovipositing males that feed less) that may be ongoing and could affect the ldquotruth in signalrdquo presented by conspecifics
Arthropod populations face a variety of environmental risks to which they must adapt that can be described within the bottom-up and top-down heuristic model however ldquoNo creature can do all thingsrdquo and genetic and other constraints are many Thus tradeoffs are required for organisms to adapt and sustain over evolutionary time (Rolf 1992) Activity or movement is highly related to resource acquisition but it is also related to mortality risks a cost and as a result mediates one of the most general tradeoffs faced by organ-isms (Werner 1992) Information gathering also comes at a cost because of the loss of time potentially spent on other activities (Eliassen et al 2007) Moreover where resources vary temporally or for example predation risk is highly unpredictable the value of information decreases with time such that a forager should place greater value on the newest available information (Eliassen et al 2007) An individualrsquos response to conspecifics makes it possible to collect a broad range of real-time biological data that may reduce the tradeoffs and costs imposed by required activity Foraging suc-cess and predator avoidance behaviors directly related to fitness are strongly affected as a result (Lazarus 1979 Foster and Treherne 1981 Lawrence 1985 Stamps 1987 Turchin 1987) The environment of xylophagous leafhoppers is highly stochastic in time and space and their extreme polyphagy requires exposure to a wide array of plant structures changing chemical profiles and associated predators Response to conspecifics offers a simple parsimonious mechanism to eliminate the need to develop sophisticated physiological or behavioral mechanisms to handle this array of visual and chemical cues that accompany the extreme uncertainty presented by resource finding and predator avoidance
In conclusion H vitripennis exhibits at least two major types of visually based behaviors While in the normal feeding position (Bex-tine et al 2005) they can detect movement from asymp10 m distance and move behind the branch to hide themselves from the approaching object This response is often done synchronously by the leafhoppers present More importantly when searching for and landing on a host plant where the risk of predation may be high they can respond to cues presented by conspecifics These consist of a narrowly defined stimulus roughly equal in size to the H vitripennis body but not gt12 mm To our knowledge this is the first report of an arthropod exhibit-ing behavioral parsimony in which one signal a visual cue (response to conspecifics) is used to mitigate immediate risks from bottom-up and top-down forces In the present case leafhoppers exhibit a ldquofixedrdquo
response response to conspecifics to bottom-up and top-down forces independent of their value in the environmental context Avail-ability and spatial distribution of host plants and their nutritional quality are critical to the survival and fitness of H vitripennis The availability of xylem fluid within and between host plants changes continuously and rapidly over time and space requiring much trivial movement and emigration by leafhoppers These natural fluctuations in plant xylem chemistry are the primary basis of the bottom-up and top-down risks to xylophagous leafhoppers because they determine the movement and seasonal use of host plants by H vitripennis (be-havior and performance) (Mizell and French 1987 Andersen et al 1989 1992 1995ab 2005 2008 Brodbeck et al 1990 19951999) Previously several novel physiological and behavioral adaptations critical to leafhopper survival were documented (Mizell et al 2008) Response to conspecifics during host plant searching movements is another novel but fundamentally important behavior exhibited by these interesting disease vectors that allow them to survive and endure as xylophages
AcknowledgmentsThe research was partially funded by a grant to R F Mizell III and P
C Andersen from the California Department of Food and Agriculture Piercersquos DiseasemdashGlassy-winged Sharpshooter Research Program We thank Gary W Knox for use of the experimental plot containing the Lagerstroemia limii We thank Tobin Northfield Joseph Patt and Arash Rashad for helpful comments on a previous draft of the manuscript
References CitedAndersen P C B V Brodbeck and R M Mizell III 1989 Metabolism
of amino acids organic acids and sugars extracted from the xylem fluid of four host plants by Homalodisca coagulata Entomol Exp Appl 50 149ndash159
Andersen P C B V Brodbeck and R M Mizell III 1992 Feeding by the leafhopper Homalodisca coagulata in relation to xylem fluid chemistry and tension J Insect Physiol 38 611ndash622
Andersen P C B V Brodbeck and R M Mizell III 1993 Diurnal variations in the chemistry of xylem fluid from Lagerstroemia indica an endogenous circadian rhythm Physiol Plant 89 783-790
Andersen P C B V Brodbeck and R M Mizell III 1995a Diurnal variation in tension osmolarity and the composition of nitrogen and carbon assimilates in xylem fluid of Prunus persica Vitis hybrid and Prunus communis J Am Hortic Sci 120 600ndash604
Andersen P C B V Brodbeck and R M Mizell III 1995b Water stress- and nutrient-mediated changes in the water relations and the amino acids organic acids and sugars in xylem fluid of Prunus salicina and Lagerstroemia indica J Am Soc Hortic Sci 120 36ndash42
Andersen P C B V Brodbeck and R M Mizell III 2003 Plant and insect characteristics in response to increasing density of Homalodisca coagulata on three host species a quantification of assimilate extraction Entomol Exp Appl 107 57ndash68
Andersen P C B V Brodbeck R M Mizell III and S Oden 2005 The abundance and feeding of Homalodisca coagulata (Homoptera Auche-norrhyncha Cicadellidae) on Vitis genotypes in north Florida Environ Entomol 34 466ndash478
Andersen PC R M Mizell III B V Brodbeck T Beckman and G Krewer 2008 Abundance and consumption rates of glassy-winged sharpshooter (Hemiptera Cicadellidae) on peaches and plums J En-tomol Sci 43 394ndash407
Andersen P C B V Brodbeck and R M Mizell III 2009 Assimilation efficiency of free and protein amino acids by the glassy-winged sharp-shooter Homalodisca vitripennis (Germar) (Hemiptera Cicadellidae Cicadellinae) feeding on Citrus sinensis and Vitis vinifera Fla Entomol 92 116ndash122
Backus E J Habibi F Yan and M Ellersieck 2005 Stylet penetration by adult Homalodisca coagulata on grape electrical penetration graph
114 American Entomologist bull Summer 2012
waveform characterization tissue correlation and possible implica-tions for transmission of Xylella fastidiosa Ann Entomol Soc Am 98 787ndash813
Baker P S 1979 Flying locust visual responses in a radial wind tunnel J Comp Physiol 131 39ndash47
Bextine B R B C Jackson D B Harshman and Miller 2005 Homalo-disca coagulata (Hemiptera Cicadellidae) feeding posture Ann Ento-mol Soc Am 98 814ndash819
Boucias D G D W Scharf S E Breaux S D Purcell and R M Mizell III 2006 Studies on the fungi associated with the glassy-winged sharpshooter Homalodisca coagulata with emphasis on a new species Hirsutella homalodiscae nom prov BioControl 52 231ndash258
Brodbeck BV R M Mizell III and P C Andersen 1990 Amino acids as determinants of host preference for the xylemfeeding leafhopper Homalodisca coagulata Oecologia 83 338ndash345
Brodbeck B V P C Andersen and R M Mizell III 1993 Physiological and behavioral adaptations of three species of leafhoppers in response to the dilute nutrient content of xylem fluid J Insect Physiol 39 73ndash81
Brodbeck B V P C Andersen and R M Mizell III 1995 Differential utilization of nutrients during development by the xylophagous leafhop-per Homalodisca coagulata Entomol Exp Appl 75 279ndash289
Brodbeck BV P C Andersen and R M Mizell III 1999 The effects of total dietary nitrogen and amino acid balance on the development of xylophagous leafhoppers Arch Insect Biochem Physiol 42 37ndash50
Brodbeck B V P C Andersen and R M Mizell III 2007 Preference-performance linkage of the xylem feeding leafhopper Homalodisca vit-ripennis (Hemiptera Cicadellidae) Environ Entomol 36 1512ndash1522
Brodbeck B V P C Andersen and R M Mizell III 2011 Nutrient media-tion of behavioral plasticity and resource allocation in a xylem-feeding leafhopper Oecol 165 111-122
Cocroft R B H J Shugart K T Konrad and K Tibbs 2006 Variation in plant substrates and its consequences for insect vibrational com-munication Ethology 112 779ndash789
Collett T S and M F Land 1978 How hoverflies compute interception courses J Comp Physiol 125 191ndash204
Coolen I O Dangles and J Casas 2005 Social learning in non-colonial insects Curr Biol 15 1931-1935
Daane K M Johnson T Ruiz and J M Hashim 2005 Host plant influence on glassy-winged sharpshooter and its natural enemies pp 100ndash104 In Proceedings 2005 Piercersquos Disease Research Symposium California Department of Food and Agriculture Sacramento
Dukas R 2001 Effects of perceived danger on flower choice by bees Ecol Lett 4 327ndash333
Dukas R and D H Morse 2003 Crab spiders affect flower visitation by bees Oikos 101 157ndash163
Eliassen S E C Jargensen M Mangel and J Garl 2007 Exploration or exploitation life expectancy changes the value of learning in foraging strategies Oikos 116 513ndash523
Fernandez-Juricic E B Kerr P Bednekoff and D Stephens 2004 When are two heads better than one Visual perception and informa-tion transfer affect vigilance coordination in foraging groups Behav Ecol 6 898ndash906
Foster W A and J E Treherne 1981 Evidence for the dilution effect in the selfish herd from fish predation on a marine insect Nature 293 466ndash467
Fournier V J Hagler K Daane J de Leoacuten and R Groves 2008 Iden-tifying the predator complex of Homalodisca vitripennis (Hemiptera Cicadellidae) a comparative study of the efficacy of an ELISA and PCR gut content bioassay Oecologia 157 629ndash640
Giraldeau L T J Valone and J J Templeton 2002 Potential disadvan-tages of using socially acquired information Phil Trans R Soc Lond B 357 1559ndash1566
Hamiliton WD 1971 Geometry for the selfish herd J Theror Biol 31295-311
Hairston N G J E Smith and L B Slobodkin 1960 Community struc-ture population control and competition Am Midl Nat 106 249ndash257
Hunter W B and D E Ullman 1994 Precibarial and cibarial chemosen-sillae in the western flower thrips Frankliniella occidentalis (Pergande)(Thysanoptera Thripidae) Int J Insect Morphol Embryol 23 69ndash83
Hunter W B E Hiebert S Webb J E Polston and J H Tsai 1996 Precibarial and cibarial chemosensilla in the whitefly Bemisia tabaci
(Gennadius) (Homoptera Aleyrodidae) Int J Insect Morphol Embryol 25 295ndash304
Jeffries M J and J H Lawton 1984 Enemy-free space and the structure of ecological communities Biol J Linn Soc 23 269-286
Karban R and L T Baldwin 1997 Induced response to herbivory University of Chicago Press Chicago
Land M F and T S Collett 1974 Chasing behavior of houseflies (Fannia canicularis) J Comp Physiol 80 331ndash357
Lawrence E S 1985 Vigilance during ldquoeasyrdquo and ldquodifficultrdquo foraging tasks Anim Behav 33 1373ndash1375
Lawton J H and S McNeil1979 Between the devil and the deep blue sea on the problem of being a herbivore pp 223ndash244 In R M Anderson B D Turner and L R Taylor [Eds] Population dynamics Blackwell Oxford England Blackwell Sci Publ 434 pp
Lazarus J 1979 Flock size and behavior in captive red-billed weaverbirds (Quelea quelea) implications for social facilitation and the functions of flocking Behavior 71 1ndash2
Lietze V R M Mizell III and D G Boucias 2011 Transmission of the mycopathogen Hirsutella spp to nymphs and adults of the glassy-winged sharpshooters Homalodisca vitripennis (=coagulata) in the greenhouse Fla Entomol 94 106-108
Lopez R F R Mizell III P C Andersen and B V Brodbeck 2003 Impact of natural enemies on populations of the glassy-winged sharpshooter Homalodisca coagulata (Say) in north Florida In R VanDriesche [Ed] First international symposium on biological control of arthropods US Dep Agric For Ser Pub FHTET-03-05 484ndash486
Mainman G A D Straw and M H Dickson 2008 A simple vision-based algorithm for decision making in flying Drosophila Curr Biol 18 464ndash470
McLinn C and D W Stephens 2006 What makes information valu-able signal reliability and environmental uncertainty Anim Behav 71 1119ndash1129
Mizell R F III and W J French 1987 Leafhopper vectors of phony peach disease feeding site preference and survival on infected and uninfected peach and seasonal response to selected host plants J Entomol Sci 22 1122
Mizell R F III P C Andersen P C Phillips and P Oevering 2005 Population dynamics and interactions between the glassy-winged sharpshooter and the host plants in response to California phenology pp 117ndash121 In Proceedings 2005 Piercersquos Disease Research Symposium California Department of Food and Agriculture Sacramento
Mizell R F III C Tipping P C Andersen B V Brodbeck T Northfield and W Hunter W 2008 Behavioral model for the glassy-winged sharp-shooter Homalodisca vitripennis (Hemiptera Cicadellidae) optimiza-tion of host plant utilization and management implications Environ Entomol 37 1049ndash1062
Mizell R F III PC Andersen and B V Brodbeck 2009 Improved detec-tion monitoring and management of the glassy-winged sharpshooter pp 40ndash43 In Proceedings 2009 Piercersquos Disease Research Symposium California Department of Food and Agriculture Sacramento
Muller KL 1998 The role of conspecifics in habitat settlement in a ter-ritorial grasshopper Anim Behav 56 479ndash485
Ninemets Uuml M Riechstein M Staudt G Siefert and J Tenhunen 2002 Stomatal constraints may affect emission of oxygenated monoterpenoids from foliage of Pinus pinea Plant Physiol 130 1371ndash1385
Ninemets Uuml F Loreto and M Riechstein 2004 Physiological and physiochemical controls on foliar volatile organic compound emissions Trends Plant Sci 9 180ndash186
Northfield T R F Mizell III P C Andersen B V Brodbeck T C Riddle and W B Hunter 2009 Dispersal patch leaving and aggregation of the glassy-winged sharpshooter Homalodisca vitripennis (Hemiptera Cicadellidae) Environ Entomol 38 183ndash191
Patt J and M Setamou 2007 Olfactory and visual stimuli affecting host plant detection in Homalodisca coagulata (Hemiptera Cicadellidae) Environ Entomol 36 142ndash150
Patt J and M Setamou 2010 Recognition of novel volatile cues by the nymph of the glassy-winged sharpshooter Homalodisca vitripennis (Cicadellidae) J Insect Behav DOI 101007s10905-010-9214-z 23 290-302
Percy D M E A Boyd and M S Hoddle 2008 Observations of acoustical signaling in three sharpshooters Homalodisca vitripennis Homalodisca
115American Entomologist bull Volume 58 Number 2
liturata and Graphocephala atropunctata (Hemiptera Cicadellidae) Ann Entomol Soc 101 253ndash259
Power M E 1992 Top-down and bottom-up forces in food webs do plants have primacy Ecologia 73 733ndash746
Price P W C E Bouton P Gross B A McPheron J N Thompson and A E Wise 1980 Interactions among three trophic levels influence of plants on interactions between insect herbivores and natural enemies Annu Rev Ecol Syst 11 41ndash66
Redak R M J Blua K L Lopes A H Purcell R F Mizell III and P C Andersen 2004 The biology and control of xylem sap-feeding insect vectors of Xylella fastidiosa and their relation to disease epidemiology Annu Rev Entomol 49 243ndash270
Rolf D A 1992 The evolution of life history traits Chapman amp Hall NYSandlin E A 2000 Cue use affects resource subdivision among three
coexisting hummingbird species Behav Ecol 11 550-559SAS Institute 2009 Proc GLM SAS 93 Statistical Analysis System Cary NCShrewsbury P and M Raupp 2008 Do top-down or bottom-up forces
determine Stephanitis pyroides abundance in urban landscapes Ecol Appl 16 262ndash273
Systat Software 2009 SigmaPlot for Windows 110 Systat Software Bangalore India
Stamps J A 1987 Conspecifics as cues to territory quality a preference of juvenile lizards (Anolis aeneus) for previously used territories Am Nat 129 629ndash642
Stamps J A 1988 Conspecific attraction and aggregation in territorial species Am Nat 131 330ndash347
Thompson J D S C Peterson and L D Harder 1987 Response to traplining bumble bees to competition experiments shifts in feeding location and efficiency Oecologia 71 295ndash300
Tipping C R F Mizell III and P C Andersen 2004 Adaptations impor-tant for the dispersal of immature stages of three species of leafhopper (Homoptera Cicadellidae) Fla Entomol 87 372ndash379
Triapitsyn S V R F Mizell III J L Bossert and J E Carlton 1998 Egg parasitoids of glassy-winged sharpshooter Homalodisca coagulata (Homoptera Cicadellidae) Fla Entomol 81 243ndash251
Turner W F and H N Pollard 1959 Life histories and behavior of five insect vectors of phony peach disease Tech Bull US Dep Agric 1188
Turchin P 1987 The role of aggregation in the response of Mexican bean beetles to host-plant density Oecologia 71 577ndash582
Valone T J 1989 Group foraging public information and patch simula-tion Oikos 56 357ndash363
Walker M and T H Jones 2001 Relative roles of top-down and bottom-up forces in terrestrial tritrophic plant-insect herbivore-natural enemy systems Oikos 93 177ndash187
Ward A J W D J T Sumpter I D Couzin P J B Hart and J Krause 2008 Quorum decision-making facilitates information transfer in fish shoals PNAS 105 6948ndash6953
Wells J M B C Raju H Y Hung W G Weisburg L Mandelco-Paul and D J Brenner 1987 Xylella fastidiosa gen nov sp nov gram-negative xylem-limited fastidious plant bacteria related to Xanthomonas spp Int J Syst Bacteriol 37 136ndash143
Werner E E 1992 Individual behavior and higher-order species interac-tions Am Nat (suppl) 140 S5ndashS32
White T R C 1978 The importance of relative shortage of food in animal ecology Oecologia 33 71ndash86
Williams G C 1966 Adaptation and natural selection Princeton University Press Princeton NJ
Wu D S C Daugherty S E Van Aken G H Pai K L Watkins H Khouri L J Tallon J M Zaborsky H E Dunbar P L Tran N A Moran J A Eisen 2006 Metabolic complementarity and genomics of the dual symbiosis of sharpshooters PloS-Biology 4 e188
Russell F Mizell III Is a Professor of Entomology (rfmizellufledu) Peter C Andersen is a Professor of Horticulture (peaufledu) and Brent V Brod-beck (bvbufledu) is a Biological Scientist all located at the University of Floridarsquos North Florida Research and Education Center-Quincy 155 Research Rd Quincy Fl 32351 Wayne B Hunter (whunterushrlarsusdagov) is a research entomologist with the USDA Agricultural Research Service at the US Horticultural Research Lab 2001 South Rock Rd Ft Pierce FL 34945 For over 20 years they have collaborated in research on the biology ecology and behavior of the leafhopper vectors of Xylella fastidiosa
With our high qualityLepidoptera Eggs and LarvaeBeet Army Worm bull Cabbage LooperCorn Earworm bull Diamondback MothFall Armyworm bull Tobacco Budworm
Save Money Save Time Get Results
Lepidoptera Eggs and LarvaeInsect Diets and Ingredients
Rearing Supplies and Equipment
Entomology Division
800-996-9908 US and Canada908-996-2155 Internationalsalesinsectrearingcom
wwwinsectrearingcom

107American Entomologist bull Volume 58 Number 2
Tipping et al 2004 Mizell et al 2008) They also contain bacte-rial symbionts thought to be involved in enhancing their nutrition (Wu et al 2006) Two other vector species Oncometopia nigricans (Walker) and H insolita (Walker) commonly occur on host plants with H vitripennis in Florida and Georgia O nigricans feeds mostly on hardwood trees and is often found feeding in heterospecific groups with H vitripennis (Turner and Pollard 1959) H insolita prefers to feed on grasses and although often infesting some of the same plants coincidently with H vitripennis [eg Johnson grass Sorghum halepense (L) Pers and pearl millet Pennisetum glaucum (L) R Br] the two species are not observed routinely feeding closely together or behaving synchronously in multispecies groups as with O nigricans (Turner and Pollard 1959)
H vitripennis and related vectors are strong flyers with keen vi-sion that allows them to detect approaching objects from asymp5 to 10 m distance H vitripennis and other related leafhopper species often feed in a stereotypical head-down posture (Bextine et al 2005) ag-gregated in groups of individual or multiple species on only a few of the many branches available on host plants When disturbed leafhop-pers in such aggregations often respond synchronously by moving en masse away from the stimulus to better conceal their bodies behind the plant structure on which they are feeding Moreover disturbed leafhoppers that fly from the host plant often return immediately and land on the branches near feeding leafhoppers Trap catch of male H vitripennis can be increased by adding cadavers of conspecifics to traps (Mizell et al 2009) In the presence of cadavers the distribution pattern of trapped leafhoppers becomes aggregated near a cadaver Together these behaviors suggest visual response by leafhoppers to conspecifics and perhaps heterospecifics
Bottom-up mortality forces that decrease relative fitness of these xylophagous insects include starvation and reduced fecundity through loss of quality host plants Top-down forces include a high rate of egg parasitism by Hymenoptera (Triapitsyn et al 1998 Lopez et al 2003) fungal pathogens that infect nymphs and adults (Walker and Jones 2001 Boucias et al 2006 Mizell et al 2008) and predation Dragonflies and related visually orienting aerial predators prey on fly-ing adults lizards spiders assassin bugs and damsel bugs threaten feeding and ovipositing adults (Lopez et al 2003 Daane et al 2004 Fournier et al 2008) and the eggs may be consumed by tree crickets Cyrtoxipha spp (Lopez et al 2002) Parasitism by Hymenoptera is also high to eggs (Triapitsyn et al 1998 Lopez et al 2003) and by fungal pathogens that infect nymphs and adults (top-down) (Walker and Jones 2001 Boucias et al 2006 Mizell et al 2008) Mizell et al (2008) developed a behavioral model that provides a conceptual framework to address the critical decisions and cues used by H vitripennis to increase its fitness in response to these putative risks That model does not deal with the potential behaviors that may be associated with the known acoustical vibration communication used by H vitripennis (Percy et al 2008) however methods using cadavers in this study eliminate the confounding effects potentially derived from acoustical communication
Response to conspecifics as a mechanism to increase an organ-ismrsquos survival and fitness has been reported in a diversity of species including fish (Foster and Treherne 1981 Ward et al 2008) birds (Lazarus 1979 Lawrence 1985 Sandlin 2000) lizards (Stamps 1987 1988) and insects (Thompson et al 1987 Turchin 1987 Muller 1998 Dukas 2001 Dukas and Morse 2003) Such ldquosocially acquiredrdquo information is used for foraging habitat assessment opponent assessment and mate choice (Geraldeau et al 2002 and
literature therein) Group foraging is only one of several behavioral situations whereby organisms derive benefit from ldquopublic informa-tionrdquo provided by conspecifics (Valone 1989) An opposing strategy territorial behavior also illustrates the importance of response to conspecifics because gaining defending and maintaining a terri-tory directly affects fitness Thus various responses to conspecifics can mediate bottom-up and top-down factors (Stamps 1987 1988)
The underlying behaviors of insects in response to conspecifics are reported for several taxa Drosophila melanogaster Meigen are attracted to long vertical objects but repelled by small ones (Mainman et al 2008) a repertoire that likely aids the recognition of landing sites and avoidance of predators and other aerial objects Houseflies Fannia canicularis L chase congeners and hoverflies pursue passing objects in the quest for mates (Land and Collett 1974 Collett and Land 1978) The locust Schistocerca gregaria (Forsk) can respond to stripe patterns (Baker 1979) Wood crickets assess danger from spider predators by observing conspecificsrsquo cues and then adopting a stereotypical predatorshyshy-avoidance behavior that is maintained after the conspecifics are gone (Coolen et al 2005) Honey bees Apis mel-lifera L avoid flower locations where they have escaped a predator and change foraging behavior in response to dead bees or spiders (Dukas 2001) To our knowledge however use of conspecifics by insects to mediate bottom-up and top-down factors has never been reported
The visual behavior of H vitripennis in response to conspecif-ics common heterospecifics their potential predators and their responses to artificial models of the insects and their predators were investigated The objectives were to characterize how leafhoppers re-spond to congeners and a heteropecific determine the putative cues associated with recognition and evaluate how these behaviors are applied in mitigating bottom-up and top-down environmental forces
Methods and MaterialsInsects and Plants Cadavers of H vitripennis and other leafhop-
pers species used in the experiments were collected from natural field populations in north Florida found on crape myrtle Lagerstroemia indica Lindl and holly Ilex spp concurrently with the studies In-sects were captured and placed in a freezer and held until needed L indica cultivars are the most frequently used hosts of H vitripennis during the months of JunendashAugust in north Floridandashsouth Georgia Texas and California (Mizell and French 1987 Brodbeck et al 1990 Mizell et al 2005 Northfield et al 2009) By using cadavers rather than live insects in the experiments the potential confounding ef-fects from behavioral communication such as acoustical signaling were eliminated
All experiments were conducted from June to September of 2006ndash2011 at the NFREC (North Florida Research and Education Center)ndashQuincy in north Florida in the field with naturally occurring populations of H vitripennis on various cultivars of crape myrtle L indica or in several 3 times 10 m plantings of sunflower Helianthus an-nus L H vitripennis density on crape myrtle in north Florida usually varies from 1 to 15 individuals per branch at moderate population levels (Brodbeck et al 1990 Mizell et al 2008) In sunflowers a good but less favored host than crape myrtle the number of H vitripennis averaged 35 leafhoppers per plant over the 6 d in that experiment At 1ndash2 h intervals visual counts (the statistical replicates) were made of H vitripennis that landed on treated and untreated branches Counts were recorded from 1000 to 1700 hours when leafhop-per counts are highest (Mizell et al 2008) After counting and to
108 American Entomologist bull Summer 2012
begin the next observation period all responding leafhoppers were induced to leave the plants by shaking the branches These leafhop-pers moved to other branches or to nearby plants Thus the count of the subsequent replicate consisted of new leafhoppers or (much less likely) previous leafhoppers making a new and independent behavioral choice The sex of the responding leafhoppers was noted and recorded when possible
Response to Conspeciics Regular white plastic drinking straws were used in the first experiments to deploy leafhopper cadavers The straws were slit lengthwise on one side to permit them to be fitted snugly over branches H vitripennis cadavers were glued to the straws with hot glue in the lab before field set up a cadaver was placed at each end of the straw and in the middle Several experiments were conducted in this manner by identifying two or more branches po-sitioned ge 50 cm apart on lsquoNatchezrsquo crape myrtle Branches for the experiment were selected for the treatments by assessing where leafhoppers were generally found feeding at the time (Mizell and French 1987) and then placing treatments at those points on other branches In the first experiment the treatments consisted of an untreated control four and eight H vitripennis cadavers per branch A second experiment was conducted using zero three six and nine leafhopper cadavers per branch
Colored Models A third test was conducted using plastic models of the average size of an adult H vitripennis (12 times 3 mm) The models were made from L-shaped pieces cut from the bottom of white plastic yogurt cups The models tested were painted black white or green (Premium Deacutecor decorative enamel General Paint and Manufactur-ing Co Cary IL) and tested along with a fresh H vitripennis cadaver and an untreated control The base of the L-shape model was used to glue the plastic models to the branches such that a silhouette of asymp3 mm was visible to represent the leafhopper bodies Counts were made on the treatment branch and either one or two other branches within ge 50 cm of the main treatment branch
Glued Cadaver Experiments Crape myrtle and sunflower field plantings with populations of leafhoppers were used as host plants Adjacent branches on crape myrtle or main branches on two adjacent sunflowers were chosen and marked in pairs such that they were ge 50 cm apart One branch or plant served as the treatment and the adjacent branch or plant as the untreated control Test branches were maintained in a natural state as much as possible but some leaves were removed similarly on each of the paired branches to facilitate the leafhopper count Treatments were placed directly on parts of the host plants where leafhoppers were typically feeding All wild leafhoppers were removed from the treatment branches to begin the test and after each observational replicate In these later experiments a rapidly drying clear glue (Aleenersquos Fast Grab Tacky Glue Duncan Enterprises Fresno CA) was used to glue the treatments directly on the host branches In all treatments with models and leafhopper cadavers a group of three individuals attached asymp8‒10 cm apart was used The experiments were conducted for several days with treat-ment leafhoppers maintained or replaced to represent naturally occurring leafhoppers as closely as possible
Heterospeciics Tested Adult H insolita were collected using a sweep net or handheld aspirator from grasses and Oncometopia nigricans from Ilex spp near the experimental sites and then placed in a freezer until they were used Heterospecific cadavers were glued onto branches in groups of three and handled as described for H vitripennis
Effects of Acoustical Communication on Response to Cadavers
As described before two branches on crape myrtle separated by ge 50 cm were used for the experiment Both branches received three cadavers asymp8‒10 cm apart located asymp50 cm from the branch tip A sleeve cage asymp30 cm long was added onto the tip of the treated branch above the cadavers Three twist ties were used to form two cages from the sleeve and three live male and three female H vitripennis were added but were segregated by sex into one of the sides of the sleeve to provide potential acoustical communication signals to the treat-ment branch Landed adult H vitripennis were counted at 30‒60 min intervals and compared between the treated and control branches
Colors and Enhanced Models Leafhopper cadavers were sprayed gray (pathogen cadaver model) red or black with acrylic paint (as described before) in the laboratory and allowed to dry They were used in the field as described above Models close to the average size of an adult H vitripennis and larger were cut out of a black foam sheet (3 mm thick) (Darico Foamies Darico Strongville OH) in the following sizes (mm) 3 times 12 (leafhopper dimensions) 10 times 10 10 times 20 and 10 times 30 (Fig 1) A red model in the leafhopper size was also tested Models were glued directly to branches in groups of three as before Rectangular models were tested with the long axis oriented either in the vertical or horizontal plane The leafhopper model was also tested for the effect of contrast by placing it in the middle of a 25 mm square block of white cardboard attached to the branch which served as a background
Bottom-up Baiting of a Poor-Quality Host Plant with Conspecif-ics Lagerstroemia limii Merrill is a species of crape myrtle related to L indica that was imported to the United States from Southeast Asia for cross-breeding In north Florida this species is represented to our knowledge by only four specimens located together at the NFREC‒Quincy Therefore it represents a novel host plant to which H vitripennis would not have been previously exposed except by
Fig 1 Types of models that were evaluated for response by the glassy-winged sharpshooter H vitripennis A Live H vitripennis in natural feeding position and aggregation B H vitripennis cadavers aligned in the experimental positions C Horizontally confi gured leafhopper-sized felt model D Square 10 times 10 mm felt model E Vertically oriented felt model F Horizontally oriented felt model G Felt model on white card for contrast H Vertically oriented model showing leafhopper feeding nearby
109American Entomologist bull Volume 58 Number 2
the local population It also represents a host of ldquopoorrdquo nutritional quality as indicated by the very low number of naturally occurring H vitripennis observed on the plants during the course of the experi-ments (Brodbeck et al 1990) Three open-pollinated seedlings of the same size (lt3 m in height) were available side by side in a row of L indica cultivars and other ornamental plant species The plants had been pruned the spring before the study and each of the L limii plants consisted of about 30 parallel vertical branches asymp2 m long above the bottom stumps of asymp1 m Cadavers of H vitripennis were placed on 10 branches randomly selected from the three L limii plants available as described before Before the treatments were deployed and at each recorded observation all of the available branches were observed for the presence of H vitripennis At no time were there more than four naturally occurring H vitripennis adults observed on the three test plants other than on the 10 branch pairs in the experiment Moreover the leafhoppers occurring naturally on L limii were all single individuals Unlike L indica and sunflowers L limii was fed upon naturally only sparingly if at all by H vitripennis This strongly indicates that this exotic novel host plant also ranked as a very poor to non-host during the time of the experiment (Brodbeck et al 1990 Andersen et al 2005)
Top-down Predators and Predator Models Predaceous lizards the green anoles Anolis carolinensis Voigt were captured locally in north Florida around buildings euthanized using carbon dioxide and held in sealed plastic bags in a freezer before use Individual specimens were only used twice in the field on two successive days for a total of asymp12 h The lizards were attached to treatment branches in their natural hunting positions using fine wire Lizards were tested with and without the addition of leafhopper cadavers as attractants In addition brown artificial bass fishing lures resembling lizards (Su-per Salt Plus 18 cm lizard Chart pumpkin 002-012 Zoom Bait Co Bogart GA) or green or purple 22 cm worms (Classic Manufacturing Co Clermont FL) were used as predator model treatments Artifi-cial fly fishing lures that resemble spiders were purchased (Gander Mountain Ocala FL brookside flies panfish assortment) and used as models The spider lures were of several colors black or yellow bod-ies with black yellow or white legs Lures were mounted singularly on the treatment branches in plain sight by using the attached hook to embed them in a treatment branch in a natural position Cadavers were not combined in the tests with these treatments
Statistics Five to 10 combinations of the treatment‒control pairs were set up in the field and observed repeatedly in all of the tests to provide the statistical replicates After each observation the responding H vitripennis were removed from the treatment branches to reset the counts to zero for the next replicate In all tests H vit-ripennis cadavers were included as the standard treatment The data were analyzed using Proc GLM procedures in SAS 93 (SAS Institute 2009) or the Mann‒Whitney Rank Sum test in SigmaPlot 110 (Systat Software 2009) when the data did not meet the requirements of normality and heterogeneity of variance The probability used for statistical significance was 005
ResultsConspeciic and Heterospeciic Cadavers Adding H vitripen-
nis cadavers to branches significantly increased the response of H vitripennis to the branches containing the cadavers The presence of the straws used only in the early experiments did not appear to influence the visual response (Fig 2 3 and 4 Table 1) The presence of three cadavers however maximized the response because adding
four to nine cadavers did not significantly increase the number of landing H vitripennis Although numerically more were observed on the treatment H vitripennis did not significantly respond to cadavers of H insolita a species about half the size of H vitripennis (Table 1) H vitripennis responded similarly to cadavers of conspecifics and the heterospecific O nigricans a species that is similar in size but differs in color from H vitripennis (Table 1) H vitripennis also responded significantly to H vitripennis cadavers painted red which is much of the body color of newly emerged adults and to cadavers painted gray as models of fungi-infected cadavers (Boucias et al 2006) (Table 1)
Fig 3
Fig 2 Mean number and SE of H vitripennis responding to varying numbers of leafhopper cadavers placed on straws on a L indica branch Letters indicate signifi cant differences P lt 005
Fig 3 Mean number and SE of H vitripennis responding to varying num-bers of leafhopper cadavers placed on straws on a crape myrtle branch and two branches within 50 cm of the main treatment branch Letters indicate signifi cant differences for comparisons within each branch type Where letters are omitted for a branch type there were no signifi cant differences P lt 005
110 American Entomologist bull Summer 2012
Effects of Acoustical Communication on Response to Cadavers Adding live H vitripennis and presumably intraspecific acoustical signals to the treatment branch significantly increased leafhopper response rate relative to the cadaver control by 53 (141 for treat-ment versus 92 on control)
Models H vitripennis response to plastic or felt models was dependent on color size and orientation (Table 1) H vitripennis responded significantly to models of several colors including black (Fig 1 and 3) and red (Table 1) Red would likely appear to these leafhoppers as dark gray or near black (Table 1) Significant response was observed to a black model similar in size to H vitripennis (3 times 12 mm) and to a model of 10 times 10 mm Models gt12 mm oriented verti-cally or horizontally did not elicit a significant response (Table 1)
Bottom-upmdashNovel Poor or Non-host Plant A significant re-sponse was observed to a treatment of three cadavers placed on the branches of the novel and poor-quality host L limii (Table 1) Total counts for all control branches never exceeded four and these were always single leafhoppers
Top-downmdashPredators H vitripennis were not repelled by the presence of any tested predator or predator model (Table 1) Lure treatments presented without cadavers using a frog a yellow or black spider received 25 on the treatments and 4 on the control and were not significantly different (data not shown) Presentations of lizard predators together with H vitripennis cadavers elicited a significant response similar to that of H vitripennis cadavers alone This further indicates a lack of response to predators and positive response to
conspecifics even when in the visual presence of actual predators or predator-like models
DiscussionH vitripennis responded strongly to the visual presence of H
vitripennis cadavers Moreover low numbers of cadavers (three)
Fig 4 Mean number and SE of H vitripennis (GWSS) responding to cadavers and colored models similar in size to its body size Letters indicate significant differences P lt 005 for comparisons between treat-ments by branch
Fig 4
Table 1 Response of the H vitripennis to various visual treatments placed on host plants
TreatmentsMain Branch
No H vitripennis Responding
Adjacent BranchNo H vitripennis Responding
(P value MannndashWhitney U test) [replicatesa]
MannndashWhitney U Statistic
H vitripennis 232 76 (0001) [165] 52395H insolita 63 30 (0998) NS [65] 21125Oncometopia nigricans 197 41 (0001) [129] 25920H vitripennis painted red 69 44 (gt 005) NS [60] 17915H vitripennis painted gray (fungus) 101 42 (0001) [76] 15470
Novel non-host plantL limii 73 36 (0001) [93] 28890
Predator modelsDead anole lizard + 3 H vitripennis cadavers 23 13 (0008) [19] 4130
Felt modelsBlack model = asympH vitripennis 3 times12 mm 108 33 (0001) [87] 15380
Black model = H vitripennis (horizontal) 37 58 (gt007) NS [71] 21145
Black model 10 times10 mm 81 45 (lt0001) [83] 22145Black model 20 times20 mm 28 31 (0241) NS [38] 5840Black model 10 times30 mm vertical 74
66 (0280) NS [69] 21425
Black model 10 times30 mm horizontal 38
40 (0646) NS [55] 29825
Black model 3 times12 mm on white 2 times2 43
36 (0262) NS [50] 10995
Red model 3 times12 mm 127 34 (lt0001) [65] 6045 Acoustical communication Live H vitripennis 141 92 (lt0004) [115] 152675
The numbers are the total leafhoppers that responded to all replicates within each comparison The statistical comparisons were performed on the actual paired numbers of leafhoppers in all of the replicates that responded to the main branch compared with the adjacent branch containing each of the treatments in the individual rows The P values indicate the probability of a larger MannndashWhitney Rank Sum statistic by chance aReplicates = number of pairs observed where count on at least one of the paired treatments was ne 0
111American Entomologist bull Volume 58 Number 2
maximized the behavior The test cadavers were placed asymp8ndash10 cm apart with the total three-cadaver presentation providing a visual cue le20 cm wide The lack of increase by additional cadavers may indicate that leafhopper visual breadth is lt30 cm H vitripennis also responded to cadavers of the similarly sized heterospecific O nigricans that is often found feeding on the same host plants (ie Ilex spp) in bi-species groups The congeneric species H insolita which is much smaller and predominantly a grass feeder however did not elicit a positive response Although H vitripennis do feed on some grass and herbaceous species concurrently with H insolita we have never observed the two species feeding in heterospecific groups or on the same part of a plant Brodbeck et al (1993) reported that the dietary needs for both species were similar although they prefer to feed on different plant tissues (Turner and Pollard 1959) Percy et al (2008) compared the acoustical signals of H vitripennis and the congener H liturata Ball and reported that the signals were different in most parameters measured Some overlap however was observed such that it cannot be ruled out that acoustical signals produced by one Homalodisca sp could be detected by a related species If there is some signal overlap between the two species then it is possible that H vitripennis could derive benefit from detecting signals produced by H insolita acoustically and visually from group defensive movements that are similar to those of H vitripennis
In an attempt to define the physical parameters of the visual cues perceived and used by H vitripennis a group of models of different sizes and orientations on the host plant were tested A black model of 3times12 mm roughly equal to the body dimensions of an adult H vitripennis evoked significant visual attraction as did a 10 times10 mm
black felt model All other larger models in any plane did not evoke significant response when placed either vertically or horizontally on the plant No response was observed to a black model affixed to a contrasting white square card These results along with the lack of response to predators and the smaller congener H insolita suggest that the visual cues used by H vitripennis when in flight are relatively narrowly defined to the size close to its own body dimensions (width le10 times length le12 mm) The heterospecific O nigricans falls within this size range The failure of H vitripennis to respond to the model on the white card along with the orientation limits may indicate that detection of the visual cues from congeners is contextually related in some manner to that provided as background by the branch or its silhouette in space
Brodbeck et al (1990) and Andersen et al (2008) showed that H vitripennis abundance on a plant is highly correlated with leafhopper feeding rate and is indicative of the underlying xylem nutritional qual-ity H vitripennis are normally highly aggregated in the field among and within available host species (especially on higher quality host plants) although there is never a time when 100 of the population is observed on one host species As a rule in north Florida only a low percentage of branches on fair-to-good hosts are infested and they usually contain more than one leafhopper per branch (Mizell and French 1987) Significant H vitripennis response to cadavers when they were placed on the novel and poor host plant L limii provides very strong support for the use of conspecifics as visual cues by H vitripennis in flight to locate potential host plants for bottom-up risk mitigation Furthermore H vitripennis will respond visually to conspecifics independent of host plant quality and familiarity with a host plant species This result supports the idea that H vitripennis do not assess host plant quality before landing and furthermore that this pivotal decision is likely made using gustation at the level of the
xylem tissue (Hunter and Ullman 1994 Hunter et al1996 Backus et al 2005) The role of plant volatiles remains to be fully described but the research of Patt and Setamou (2007 2010) suggests a potential role in H vitripennis behavior
The importance of bottom-up nutrient profiles on fitness of these leafhoppers is well described (Redak et al 2004) Leafhopper ag-gregation however may be of further benefit by inducing chemical changes in host plant physiology that affect fitness such as feeding rate and fecundity (Karban and Baldwin 1997) Andersen et al (2003 2009) addressed this possibility in experiments with five host species They found that moderate-to-large plants of crape myrtle L indica L and plum Prunus salicina Lindl exhibited no increasing effects on nutrient compounds of leafhopper feeding in large numbers nor did leafhopper feeding behavior change when in groups In small soybean plants Glycine max (L) some nutrients were highly reduced (Andersen et al 2003) In citrus H vitripennis feeding increased seven amino acids in protein form but little change in protein amino acids was observed in Vitis vinifera lsquoChardonnayrsquo In citrus and lsquoChardonnayrsquo grape amino acid increased assimilation efficiency as a result of leafhopper feeding These data suggest that H vitripennis feeding does not induce metabolically inhibitory profiles in the xylem (Andersen et al 2009) This along with the induction of proteins and a slight decrease in free amino acids indicates that there may be some induced benefit in nutrition from H vitripennis aggregation (Andersen et al 2009)
Predation is an important mortality risk to H vitripennis in flight and on the host plant during feeding and oviposition (Lopez et al 2003 Daane et al 2005 Suttle and Hoddle 2006 Fournier et al 2008 Mizell et al 2008) Spiders and lizards are the most impor-tant predators of H vitripennis on the plants (Lopez et al 2003 Fournier et al 2008) Top-down risk mitigation by H vitripennis was assessed indirectly by testing visual response by H vitripennis to lizard predators and simulated predator models (fishing lures) that closely resembled living predators The experimental context most closely approximates the risk to H vitripennis during landing on a new branch the time when little or no information on predator presence on the selected branch is available to arriving H vitripennis This is also the time when H vitripennis presents the largest moving visual stimulus to hidden predators Once on the plant landed H vitripennis stop moving and provide a different visual stimulus to would-be predators they also regain the use of positional camouflage and their own visually based longer-range avoidance defense with the protection it provides As described earlier this protection is partially accrued from conspecifics via the ldquodilutionrdquo effect (Foster and Treherne 1981) Similarly heterospecifics present would also contribute to defense
The only other time that H vitripennis move excessively on the host plant is when the males seek mates and the females oviposit (Mizell et al 2008) Although copulation occurs mostly during the scotophase one might hypothesize that the presence of conspecif-ics would also reduce the risk from predation to more vulnerable copulating pairs and ovipositing females This may also be a selec-tion factor that induces H vitripennis females to oviposit on plants upon which they feed but which do not meet the nutritional needs of their nymphs (Brodbeck et al 2007 Mizell et al 2008) The ability to recognize conspecifics as indicators of suitable host plants allows H vitripennis to feed without moving and eliminates search flights for hours or days proportionally reducing in-flight and landing preda-tion risks These behaviors underscore the potential risks involved in
112 American Entomologist bull Summer 2012
finding hosts the tradeoffs faced by female leafhoppers and the value of conspecific recognition in their mitigation An unusual number of mated leafhopper pairs also were observed among the cadaver treatment branches on the mornings of succeeding test days These observations provide further support for the contention concerning top down risk mitigation via conspecific response
Significant response to avoid predators and artificial models was not observed It is unlikely that an arthropod herbivore would be able to develop a search image with such elasticity that it could encompass the wide range of natural enemies potentially encoun-tered The immediate acceptance of the novel and poor host L limii provides further evidence against the occurrence of a learned host image by searching leafhoppers Response to conspecifics would obviate the necessity of developing a predator search image because conspecifics would usually be present only where predators were absent Thus a type of ldquoenemy-free spacerdquo (Jeffries and Lawton 1984) may be represented by conspecific presence Alternatively relative to the risk of landing on a branch without conspecifics even if the conspecific signal was not entirely truthful about the level of risk the dilution effect from conspecific presence would provide increased protection theoretically in direct proportion to the number of con-specifics present Although foragers in many animal groups have an inherent geometry that causes information asymmetries individuals get more information from closer neighbors (Fernandez-Juricic et al 2004) H vitripennis uses keen visual acuity that likely is coupled with acoustical communication as a mechanism to respond rapidly en masse to approaching predators As a result leafhoppers accrue a more equivalent gain from defensive movement coordination (Fernandez-Juricic et al 2004)
On the host plant H vitripennis can detect and respond (and routinely do so in concert perhaps mediated by acoustic commu-nication) to movement of large objects approaching them from off the plant and to smaller objects on the plant Live lizards do elicit a response from feeding H vitripennis as they move over the host plant and approach leafhoppers (Lopez et al 2003) This suggests the occurrence in H vitripennis of a general visual capability (differ-ent from the visual response to conspecifics) to recognize moving objects that would circumvent the necessity of developing an array of species-specific predator search images
H vitripennis responded significantly to cadavers painted gray that served as a model of H vitripennis infected with the parasitic fungal pathogen H homalodiscae (Boucias et al 2006) H vitripennis cadavers infected with this pathogen remain attached for months on the host plant at feeding sites in the feeding position and infect H vitripennis nymphs and adults The infected cadavers are covered by the fungal mycelia that sporulate in cycles and provide a means to infect uninfected feeding H vitripennis (Lietze et al 2010) H homalodiscae-infected H vitripennis attach tenaciously via rhizoids to the plant bark in the summer and are often found attached to this substrate the following spring This positive visual response may increase transmission of the fungus to H vitripennis
When live presumably acoustically active H vitripennis were added to treatment branches the numbers of H vitripennis increased by 53 on the treated branches (Table 1) H vitripennis unlike most insect herbivores has not been shown to use olfaction at any level for courtship or mating In contrast H vitripennis and many Cicadel-lidae and related insect families use acoustical vibrations propagated through the host plant to communicate during courtship and mating and for other functions (Cocroft 2006) This behavioral modality of
H vitripennis however has received only cursory attention (Percy et al 2008) and its potential effect was eliminated in all except this one experiment through the use of cadavers
With respect to host finding the role of olfaction remains un-clear Patt and Setamou (2007) reported that the response to visual cues by H vitripennis nymphs and adults was enhanced by exposure to plant volatiles Patt and Setamou (2010) demonstrated that H vitripennis nymphs could be conditioned to respond to vanilla as a novel host volatile and as a result they argued that associative learning of chemical profiles could enhance leafhopper location and recognition of host plants These authors also speculated on a potential mechanism use of plant volatile profile emissions (Ninemets et al 2002 2004) in host finding and recognition by adult H vitripennis While an interesting hypothesis much research remains to be done to support these ideas A strong response to conspecifics provides a complimentary or alternative mechanism that would eliminate the need for such highly evolved behaviors However one may hypothesize that the presence of plant volatiles could stimulate a more intense leafhopper visual search for conspe-cifics on the host Nevertheless the ready acceptance of the novel and poor host L limii in the presence of conspecifics argues against the need for leafhoppers to learn a visual or olfactory pattern for specific plant species
Trivial movement within and between host plants and host plant patches is germane to understanding the distribution and fitness of H vitripennis (Mizell et al 2008) H vitripennis are strongly visually oriented while feeding on a host plant they can detect movement from 5 to 10 m away Based on the ostensibly narrow sensory ca-pabilities of H vitripennis (lack of pheromones apparently limited use of olfaction) it is not unexpected that H vitripennis would have evolved visual behavior with multiple functions Moreover this simple capability would obviate the need for higher level behaviors such as associative learning at least for adults (Patt and Setamou 2010) Profiles of xylem chemistry can change as much as 10-fold in one day or from day to day in the absence of any overt visual signals to leafhoppers (Andersen et al1992 Brodbeck et al 2010) Because host plant nutrient quality varies unrecognizably to leafhoppers in time and space and between and within a plant species developing a search image to visually identify specific host plants or character-istics related to their quality appears to be extremely difficult and perhaps of little value Moreover host plants usually do not occur in monocultures a situation that would reinforce short-term pattern recognition By contrast delay in finding hosts within a patch can affect information retention (McLinn and Stephens 2006) Relying too heavily on recent experience can be costly especially if the environment and in this case host plant nutrients change rapidly (Mizell and French 1987 Andersen et al 1989 1992 1993 1995ab Brodbeck et al 1990 19951999 McLinn and Stephens 2006) Animals devalue unreliable signals Models predict signal following when the signal reliability exceeds environmental certaintymdashso that information is valuable Signal value however is not based solely on reliability it also depends on the statistical distribution of the environmental states (McLinn and Stephens 2006) Plant quality and predation risks are in constant flux in the case of H vitripennis and extremely variable in time and space H vitripennis handling of bottom-up and top-down forces is tied closely together because the potential payoff and consequences from both accrue on the host plant following a single decision to land H vitripennis appar-ently can make a decision to land based solely on a visual response
113American Entomologist bull Volume 58 Number 2
to conspecifics on host plant branches An arrestment response is then likely reinforced on the plant via acoustical communication as gustation behavior is initiated
Thus presence of conspecifics indicates a ldquosafety zonerdquo for host plant testing and likely provides high-quality (truthfulness of signal) information because all H vitripennis in any one location are under more or less the same dietary constraints Notwithstanding that O nigricans may have somewhat different nutritional requirements it may be important in the mitigation of bottom-up and top-down forces The presence of one or (usually) multiple conspecifics is likely a signal indicating a reasonably high threshold quality of nutrients or else the duration of conspecifics remaining on a host plant would be short This perhaps explains why three conspecifics analogous to the quorum decision of Ward et al (2008) maximize the response Three may be enough to compensate for any variations in individual hunger rate or other uninterpretable behaviors (females not feeding or ovipositing males that feed less) that may be ongoing and could affect the ldquotruth in signalrdquo presented by conspecifics
Arthropod populations face a variety of environmental risks to which they must adapt that can be described within the bottom-up and top-down heuristic model however ldquoNo creature can do all thingsrdquo and genetic and other constraints are many Thus tradeoffs are required for organisms to adapt and sustain over evolutionary time (Rolf 1992) Activity or movement is highly related to resource acquisition but it is also related to mortality risks a cost and as a result mediates one of the most general tradeoffs faced by organ-isms (Werner 1992) Information gathering also comes at a cost because of the loss of time potentially spent on other activities (Eliassen et al 2007) Moreover where resources vary temporally or for example predation risk is highly unpredictable the value of information decreases with time such that a forager should place greater value on the newest available information (Eliassen et al 2007) An individualrsquos response to conspecifics makes it possible to collect a broad range of real-time biological data that may reduce the tradeoffs and costs imposed by required activity Foraging suc-cess and predator avoidance behaviors directly related to fitness are strongly affected as a result (Lazarus 1979 Foster and Treherne 1981 Lawrence 1985 Stamps 1987 Turchin 1987) The environment of xylophagous leafhoppers is highly stochastic in time and space and their extreme polyphagy requires exposure to a wide array of plant structures changing chemical profiles and associated predators Response to conspecifics offers a simple parsimonious mechanism to eliminate the need to develop sophisticated physiological or behavioral mechanisms to handle this array of visual and chemical cues that accompany the extreme uncertainty presented by resource finding and predator avoidance
In conclusion H vitripennis exhibits at least two major types of visually based behaviors While in the normal feeding position (Bex-tine et al 2005) they can detect movement from asymp10 m distance and move behind the branch to hide themselves from the approaching object This response is often done synchronously by the leafhoppers present More importantly when searching for and landing on a host plant where the risk of predation may be high they can respond to cues presented by conspecifics These consist of a narrowly defined stimulus roughly equal in size to the H vitripennis body but not gt12 mm To our knowledge this is the first report of an arthropod exhibit-ing behavioral parsimony in which one signal a visual cue (response to conspecifics) is used to mitigate immediate risks from bottom-up and top-down forces In the present case leafhoppers exhibit a ldquofixedrdquo
response response to conspecifics to bottom-up and top-down forces independent of their value in the environmental context Avail-ability and spatial distribution of host plants and their nutritional quality are critical to the survival and fitness of H vitripennis The availability of xylem fluid within and between host plants changes continuously and rapidly over time and space requiring much trivial movement and emigration by leafhoppers These natural fluctuations in plant xylem chemistry are the primary basis of the bottom-up and top-down risks to xylophagous leafhoppers because they determine the movement and seasonal use of host plants by H vitripennis (be-havior and performance) (Mizell and French 1987 Andersen et al 1989 1992 1995ab 2005 2008 Brodbeck et al 1990 19951999) Previously several novel physiological and behavioral adaptations critical to leafhopper survival were documented (Mizell et al 2008) Response to conspecifics during host plant searching movements is another novel but fundamentally important behavior exhibited by these interesting disease vectors that allow them to survive and endure as xylophages
AcknowledgmentsThe research was partially funded by a grant to R F Mizell III and P
C Andersen from the California Department of Food and Agriculture Piercersquos DiseasemdashGlassy-winged Sharpshooter Research Program We thank Gary W Knox for use of the experimental plot containing the Lagerstroemia limii We thank Tobin Northfield Joseph Patt and Arash Rashad for helpful comments on a previous draft of the manuscript
References CitedAndersen P C B V Brodbeck and R M Mizell III 1989 Metabolism
of amino acids organic acids and sugars extracted from the xylem fluid of four host plants by Homalodisca coagulata Entomol Exp Appl 50 149ndash159
Andersen P C B V Brodbeck and R M Mizell III 1992 Feeding by the leafhopper Homalodisca coagulata in relation to xylem fluid chemistry and tension J Insect Physiol 38 611ndash622
Andersen P C B V Brodbeck and R M Mizell III 1993 Diurnal variations in the chemistry of xylem fluid from Lagerstroemia indica an endogenous circadian rhythm Physiol Plant 89 783-790
Andersen P C B V Brodbeck and R M Mizell III 1995a Diurnal variation in tension osmolarity and the composition of nitrogen and carbon assimilates in xylem fluid of Prunus persica Vitis hybrid and Prunus communis J Am Hortic Sci 120 600ndash604
Andersen P C B V Brodbeck and R M Mizell III 1995b Water stress- and nutrient-mediated changes in the water relations and the amino acids organic acids and sugars in xylem fluid of Prunus salicina and Lagerstroemia indica J Am Soc Hortic Sci 120 36ndash42
Andersen P C B V Brodbeck and R M Mizell III 2003 Plant and insect characteristics in response to increasing density of Homalodisca coagulata on three host species a quantification of assimilate extraction Entomol Exp Appl 107 57ndash68
Andersen P C B V Brodbeck R M Mizell III and S Oden 2005 The abundance and feeding of Homalodisca coagulata (Homoptera Auche-norrhyncha Cicadellidae) on Vitis genotypes in north Florida Environ Entomol 34 466ndash478
Andersen PC R M Mizell III B V Brodbeck T Beckman and G Krewer 2008 Abundance and consumption rates of glassy-winged sharpshooter (Hemiptera Cicadellidae) on peaches and plums J En-tomol Sci 43 394ndash407
Andersen P C B V Brodbeck and R M Mizell III 2009 Assimilation efficiency of free and protein amino acids by the glassy-winged sharp-shooter Homalodisca vitripennis (Germar) (Hemiptera Cicadellidae Cicadellinae) feeding on Citrus sinensis and Vitis vinifera Fla Entomol 92 116ndash122
Backus E J Habibi F Yan and M Ellersieck 2005 Stylet penetration by adult Homalodisca coagulata on grape electrical penetration graph
114 American Entomologist bull Summer 2012
waveform characterization tissue correlation and possible implica-tions for transmission of Xylella fastidiosa Ann Entomol Soc Am 98 787ndash813
Baker P S 1979 Flying locust visual responses in a radial wind tunnel J Comp Physiol 131 39ndash47
Bextine B R B C Jackson D B Harshman and Miller 2005 Homalo-disca coagulata (Hemiptera Cicadellidae) feeding posture Ann Ento-mol Soc Am 98 814ndash819
Boucias D G D W Scharf S E Breaux S D Purcell and R M Mizell III 2006 Studies on the fungi associated with the glassy-winged sharpshooter Homalodisca coagulata with emphasis on a new species Hirsutella homalodiscae nom prov BioControl 52 231ndash258
Brodbeck BV R M Mizell III and P C Andersen 1990 Amino acids as determinants of host preference for the xylemfeeding leafhopper Homalodisca coagulata Oecologia 83 338ndash345
Brodbeck B V P C Andersen and R M Mizell III 1993 Physiological and behavioral adaptations of three species of leafhoppers in response to the dilute nutrient content of xylem fluid J Insect Physiol 39 73ndash81
Brodbeck B V P C Andersen and R M Mizell III 1995 Differential utilization of nutrients during development by the xylophagous leafhop-per Homalodisca coagulata Entomol Exp Appl 75 279ndash289
Brodbeck BV P C Andersen and R M Mizell III 1999 The effects of total dietary nitrogen and amino acid balance on the development of xylophagous leafhoppers Arch Insect Biochem Physiol 42 37ndash50
Brodbeck B V P C Andersen and R M Mizell III 2007 Preference-performance linkage of the xylem feeding leafhopper Homalodisca vit-ripennis (Hemiptera Cicadellidae) Environ Entomol 36 1512ndash1522
Brodbeck B V P C Andersen and R M Mizell III 2011 Nutrient media-tion of behavioral plasticity and resource allocation in a xylem-feeding leafhopper Oecol 165 111-122
Cocroft R B H J Shugart K T Konrad and K Tibbs 2006 Variation in plant substrates and its consequences for insect vibrational com-munication Ethology 112 779ndash789
Collett T S and M F Land 1978 How hoverflies compute interception courses J Comp Physiol 125 191ndash204
Coolen I O Dangles and J Casas 2005 Social learning in non-colonial insects Curr Biol 15 1931-1935
Daane K M Johnson T Ruiz and J M Hashim 2005 Host plant influence on glassy-winged sharpshooter and its natural enemies pp 100ndash104 In Proceedings 2005 Piercersquos Disease Research Symposium California Department of Food and Agriculture Sacramento
Dukas R 2001 Effects of perceived danger on flower choice by bees Ecol Lett 4 327ndash333
Dukas R and D H Morse 2003 Crab spiders affect flower visitation by bees Oikos 101 157ndash163
Eliassen S E C Jargensen M Mangel and J Garl 2007 Exploration or exploitation life expectancy changes the value of learning in foraging strategies Oikos 116 513ndash523
Fernandez-Juricic E B Kerr P Bednekoff and D Stephens 2004 When are two heads better than one Visual perception and informa-tion transfer affect vigilance coordination in foraging groups Behav Ecol 6 898ndash906
Foster W A and J E Treherne 1981 Evidence for the dilution effect in the selfish herd from fish predation on a marine insect Nature 293 466ndash467
Fournier V J Hagler K Daane J de Leoacuten and R Groves 2008 Iden-tifying the predator complex of Homalodisca vitripennis (Hemiptera Cicadellidae) a comparative study of the efficacy of an ELISA and PCR gut content bioassay Oecologia 157 629ndash640
Giraldeau L T J Valone and J J Templeton 2002 Potential disadvan-tages of using socially acquired information Phil Trans R Soc Lond B 357 1559ndash1566
Hamiliton WD 1971 Geometry for the selfish herd J Theror Biol 31295-311
Hairston N G J E Smith and L B Slobodkin 1960 Community struc-ture population control and competition Am Midl Nat 106 249ndash257
Hunter W B and D E Ullman 1994 Precibarial and cibarial chemosen-sillae in the western flower thrips Frankliniella occidentalis (Pergande)(Thysanoptera Thripidae) Int J Insect Morphol Embryol 23 69ndash83
Hunter W B E Hiebert S Webb J E Polston and J H Tsai 1996 Precibarial and cibarial chemosensilla in the whitefly Bemisia tabaci
(Gennadius) (Homoptera Aleyrodidae) Int J Insect Morphol Embryol 25 295ndash304
Jeffries M J and J H Lawton 1984 Enemy-free space and the structure of ecological communities Biol J Linn Soc 23 269-286
Karban R and L T Baldwin 1997 Induced response to herbivory University of Chicago Press Chicago
Land M F and T S Collett 1974 Chasing behavior of houseflies (Fannia canicularis) J Comp Physiol 80 331ndash357
Lawrence E S 1985 Vigilance during ldquoeasyrdquo and ldquodifficultrdquo foraging tasks Anim Behav 33 1373ndash1375
Lawton J H and S McNeil1979 Between the devil and the deep blue sea on the problem of being a herbivore pp 223ndash244 In R M Anderson B D Turner and L R Taylor [Eds] Population dynamics Blackwell Oxford England Blackwell Sci Publ 434 pp
Lazarus J 1979 Flock size and behavior in captive red-billed weaverbirds (Quelea quelea) implications for social facilitation and the functions of flocking Behavior 71 1ndash2
Lietze V R M Mizell III and D G Boucias 2011 Transmission of the mycopathogen Hirsutella spp to nymphs and adults of the glassy-winged sharpshooters Homalodisca vitripennis (=coagulata) in the greenhouse Fla Entomol 94 106-108
Lopez R F R Mizell III P C Andersen and B V Brodbeck 2003 Impact of natural enemies on populations of the glassy-winged sharpshooter Homalodisca coagulata (Say) in north Florida In R VanDriesche [Ed] First international symposium on biological control of arthropods US Dep Agric For Ser Pub FHTET-03-05 484ndash486
Mainman G A D Straw and M H Dickson 2008 A simple vision-based algorithm for decision making in flying Drosophila Curr Biol 18 464ndash470
McLinn C and D W Stephens 2006 What makes information valu-able signal reliability and environmental uncertainty Anim Behav 71 1119ndash1129
Mizell R F III and W J French 1987 Leafhopper vectors of phony peach disease feeding site preference and survival on infected and uninfected peach and seasonal response to selected host plants J Entomol Sci 22 1122
Mizell R F III P C Andersen P C Phillips and P Oevering 2005 Population dynamics and interactions between the glassy-winged sharpshooter and the host plants in response to California phenology pp 117ndash121 In Proceedings 2005 Piercersquos Disease Research Symposium California Department of Food and Agriculture Sacramento
Mizell R F III C Tipping P C Andersen B V Brodbeck T Northfield and W Hunter W 2008 Behavioral model for the glassy-winged sharp-shooter Homalodisca vitripennis (Hemiptera Cicadellidae) optimiza-tion of host plant utilization and management implications Environ Entomol 37 1049ndash1062
Mizell R F III PC Andersen and B V Brodbeck 2009 Improved detec-tion monitoring and management of the glassy-winged sharpshooter pp 40ndash43 In Proceedings 2009 Piercersquos Disease Research Symposium California Department of Food and Agriculture Sacramento
Muller KL 1998 The role of conspecifics in habitat settlement in a ter-ritorial grasshopper Anim Behav 56 479ndash485
Ninemets Uuml M Riechstein M Staudt G Siefert and J Tenhunen 2002 Stomatal constraints may affect emission of oxygenated monoterpenoids from foliage of Pinus pinea Plant Physiol 130 1371ndash1385
Ninemets Uuml F Loreto and M Riechstein 2004 Physiological and physiochemical controls on foliar volatile organic compound emissions Trends Plant Sci 9 180ndash186
Northfield T R F Mizell III P C Andersen B V Brodbeck T C Riddle and W B Hunter 2009 Dispersal patch leaving and aggregation of the glassy-winged sharpshooter Homalodisca vitripennis (Hemiptera Cicadellidae) Environ Entomol 38 183ndash191
Patt J and M Setamou 2007 Olfactory and visual stimuli affecting host plant detection in Homalodisca coagulata (Hemiptera Cicadellidae) Environ Entomol 36 142ndash150
Patt J and M Setamou 2010 Recognition of novel volatile cues by the nymph of the glassy-winged sharpshooter Homalodisca vitripennis (Cicadellidae) J Insect Behav DOI 101007s10905-010-9214-z 23 290-302
Percy D M E A Boyd and M S Hoddle 2008 Observations of acoustical signaling in three sharpshooters Homalodisca vitripennis Homalodisca
115American Entomologist bull Volume 58 Number 2
liturata and Graphocephala atropunctata (Hemiptera Cicadellidae) Ann Entomol Soc 101 253ndash259
Power M E 1992 Top-down and bottom-up forces in food webs do plants have primacy Ecologia 73 733ndash746
Price P W C E Bouton P Gross B A McPheron J N Thompson and A E Wise 1980 Interactions among three trophic levels influence of plants on interactions between insect herbivores and natural enemies Annu Rev Ecol Syst 11 41ndash66
Redak R M J Blua K L Lopes A H Purcell R F Mizell III and P C Andersen 2004 The biology and control of xylem sap-feeding insect vectors of Xylella fastidiosa and their relation to disease epidemiology Annu Rev Entomol 49 243ndash270
Rolf D A 1992 The evolution of life history traits Chapman amp Hall NYSandlin E A 2000 Cue use affects resource subdivision among three
coexisting hummingbird species Behav Ecol 11 550-559SAS Institute 2009 Proc GLM SAS 93 Statistical Analysis System Cary NCShrewsbury P and M Raupp 2008 Do top-down or bottom-up forces
determine Stephanitis pyroides abundance in urban landscapes Ecol Appl 16 262ndash273
Systat Software 2009 SigmaPlot for Windows 110 Systat Software Bangalore India
Stamps J A 1987 Conspecifics as cues to territory quality a preference of juvenile lizards (Anolis aeneus) for previously used territories Am Nat 129 629ndash642
Stamps J A 1988 Conspecific attraction and aggregation in territorial species Am Nat 131 330ndash347
Thompson J D S C Peterson and L D Harder 1987 Response to traplining bumble bees to competition experiments shifts in feeding location and efficiency Oecologia 71 295ndash300
Tipping C R F Mizell III and P C Andersen 2004 Adaptations impor-tant for the dispersal of immature stages of three species of leafhopper (Homoptera Cicadellidae) Fla Entomol 87 372ndash379
Triapitsyn S V R F Mizell III J L Bossert and J E Carlton 1998 Egg parasitoids of glassy-winged sharpshooter Homalodisca coagulata (Homoptera Cicadellidae) Fla Entomol 81 243ndash251
Turner W F and H N Pollard 1959 Life histories and behavior of five insect vectors of phony peach disease Tech Bull US Dep Agric 1188
Turchin P 1987 The role of aggregation in the response of Mexican bean beetles to host-plant density Oecologia 71 577ndash582
Valone T J 1989 Group foraging public information and patch simula-tion Oikos 56 357ndash363
Walker M and T H Jones 2001 Relative roles of top-down and bottom-up forces in terrestrial tritrophic plant-insect herbivore-natural enemy systems Oikos 93 177ndash187
Ward A J W D J T Sumpter I D Couzin P J B Hart and J Krause 2008 Quorum decision-making facilitates information transfer in fish shoals PNAS 105 6948ndash6953
Wells J M B C Raju H Y Hung W G Weisburg L Mandelco-Paul and D J Brenner 1987 Xylella fastidiosa gen nov sp nov gram-negative xylem-limited fastidious plant bacteria related to Xanthomonas spp Int J Syst Bacteriol 37 136ndash143
Werner E E 1992 Individual behavior and higher-order species interac-tions Am Nat (suppl) 140 S5ndashS32
White T R C 1978 The importance of relative shortage of food in animal ecology Oecologia 33 71ndash86
Williams G C 1966 Adaptation and natural selection Princeton University Press Princeton NJ
Wu D S C Daugherty S E Van Aken G H Pai K L Watkins H Khouri L J Tallon J M Zaborsky H E Dunbar P L Tran N A Moran J A Eisen 2006 Metabolic complementarity and genomics of the dual symbiosis of sharpshooters PloS-Biology 4 e188
Russell F Mizell III Is a Professor of Entomology (rfmizellufledu) Peter C Andersen is a Professor of Horticulture (peaufledu) and Brent V Brod-beck (bvbufledu) is a Biological Scientist all located at the University of Floridarsquos North Florida Research and Education Center-Quincy 155 Research Rd Quincy Fl 32351 Wayne B Hunter (whunterushrlarsusdagov) is a research entomologist with the USDA Agricultural Research Service at the US Horticultural Research Lab 2001 South Rock Rd Ft Pierce FL 34945 For over 20 years they have collaborated in research on the biology ecology and behavior of the leafhopper vectors of Xylella fastidiosa
With our high qualityLepidoptera Eggs and LarvaeBeet Army Worm bull Cabbage LooperCorn Earworm bull Diamondback MothFall Armyworm bull Tobacco Budworm
Save Money Save Time Get Results
Lepidoptera Eggs and LarvaeInsect Diets and Ingredients
Rearing Supplies and Equipment
Entomology Division
800-996-9908 US and Canada908-996-2155 Internationalsalesinsectrearingcom
wwwinsectrearingcom

108 American Entomologist bull Summer 2012
begin the next observation period all responding leafhoppers were induced to leave the plants by shaking the branches These leafhop-pers moved to other branches or to nearby plants Thus the count of the subsequent replicate consisted of new leafhoppers or (much less likely) previous leafhoppers making a new and independent behavioral choice The sex of the responding leafhoppers was noted and recorded when possible
Response to Conspeciics Regular white plastic drinking straws were used in the first experiments to deploy leafhopper cadavers The straws were slit lengthwise on one side to permit them to be fitted snugly over branches H vitripennis cadavers were glued to the straws with hot glue in the lab before field set up a cadaver was placed at each end of the straw and in the middle Several experiments were conducted in this manner by identifying two or more branches po-sitioned ge 50 cm apart on lsquoNatchezrsquo crape myrtle Branches for the experiment were selected for the treatments by assessing where leafhoppers were generally found feeding at the time (Mizell and French 1987) and then placing treatments at those points on other branches In the first experiment the treatments consisted of an untreated control four and eight H vitripennis cadavers per branch A second experiment was conducted using zero three six and nine leafhopper cadavers per branch
Colored Models A third test was conducted using plastic models of the average size of an adult H vitripennis (12 times 3 mm) The models were made from L-shaped pieces cut from the bottom of white plastic yogurt cups The models tested were painted black white or green (Premium Deacutecor decorative enamel General Paint and Manufactur-ing Co Cary IL) and tested along with a fresh H vitripennis cadaver and an untreated control The base of the L-shape model was used to glue the plastic models to the branches such that a silhouette of asymp3 mm was visible to represent the leafhopper bodies Counts were made on the treatment branch and either one or two other branches within ge 50 cm of the main treatment branch
Glued Cadaver Experiments Crape myrtle and sunflower field plantings with populations of leafhoppers were used as host plants Adjacent branches on crape myrtle or main branches on two adjacent sunflowers were chosen and marked in pairs such that they were ge 50 cm apart One branch or plant served as the treatment and the adjacent branch or plant as the untreated control Test branches were maintained in a natural state as much as possible but some leaves were removed similarly on each of the paired branches to facilitate the leafhopper count Treatments were placed directly on parts of the host plants where leafhoppers were typically feeding All wild leafhoppers were removed from the treatment branches to begin the test and after each observational replicate In these later experiments a rapidly drying clear glue (Aleenersquos Fast Grab Tacky Glue Duncan Enterprises Fresno CA) was used to glue the treatments directly on the host branches In all treatments with models and leafhopper cadavers a group of three individuals attached asymp8‒10 cm apart was used The experiments were conducted for several days with treat-ment leafhoppers maintained or replaced to represent naturally occurring leafhoppers as closely as possible
Heterospeciics Tested Adult H insolita were collected using a sweep net or handheld aspirator from grasses and Oncometopia nigricans from Ilex spp near the experimental sites and then placed in a freezer until they were used Heterospecific cadavers were glued onto branches in groups of three and handled as described for H vitripennis
Effects of Acoustical Communication on Response to Cadavers
As described before two branches on crape myrtle separated by ge 50 cm were used for the experiment Both branches received three cadavers asymp8‒10 cm apart located asymp50 cm from the branch tip A sleeve cage asymp30 cm long was added onto the tip of the treated branch above the cadavers Three twist ties were used to form two cages from the sleeve and three live male and three female H vitripennis were added but were segregated by sex into one of the sides of the sleeve to provide potential acoustical communication signals to the treat-ment branch Landed adult H vitripennis were counted at 30‒60 min intervals and compared between the treated and control branches
Colors and Enhanced Models Leafhopper cadavers were sprayed gray (pathogen cadaver model) red or black with acrylic paint (as described before) in the laboratory and allowed to dry They were used in the field as described above Models close to the average size of an adult H vitripennis and larger were cut out of a black foam sheet (3 mm thick) (Darico Foamies Darico Strongville OH) in the following sizes (mm) 3 times 12 (leafhopper dimensions) 10 times 10 10 times 20 and 10 times 30 (Fig 1) A red model in the leafhopper size was also tested Models were glued directly to branches in groups of three as before Rectangular models were tested with the long axis oriented either in the vertical or horizontal plane The leafhopper model was also tested for the effect of contrast by placing it in the middle of a 25 mm square block of white cardboard attached to the branch which served as a background
Bottom-up Baiting of a Poor-Quality Host Plant with Conspecif-ics Lagerstroemia limii Merrill is a species of crape myrtle related to L indica that was imported to the United States from Southeast Asia for cross-breeding In north Florida this species is represented to our knowledge by only four specimens located together at the NFREC‒Quincy Therefore it represents a novel host plant to which H vitripennis would not have been previously exposed except by
Fig 1 Types of models that were evaluated for response by the glassy-winged sharpshooter H vitripennis A Live H vitripennis in natural feeding position and aggregation B H vitripennis cadavers aligned in the experimental positions C Horizontally confi gured leafhopper-sized felt model D Square 10 times 10 mm felt model E Vertically oriented felt model F Horizontally oriented felt model G Felt model on white card for contrast H Vertically oriented model showing leafhopper feeding nearby
109American Entomologist bull Volume 58 Number 2
the local population It also represents a host of ldquopoorrdquo nutritional quality as indicated by the very low number of naturally occurring H vitripennis observed on the plants during the course of the experi-ments (Brodbeck et al 1990) Three open-pollinated seedlings of the same size (lt3 m in height) were available side by side in a row of L indica cultivars and other ornamental plant species The plants had been pruned the spring before the study and each of the L limii plants consisted of about 30 parallel vertical branches asymp2 m long above the bottom stumps of asymp1 m Cadavers of H vitripennis were placed on 10 branches randomly selected from the three L limii plants available as described before Before the treatments were deployed and at each recorded observation all of the available branches were observed for the presence of H vitripennis At no time were there more than four naturally occurring H vitripennis adults observed on the three test plants other than on the 10 branch pairs in the experiment Moreover the leafhoppers occurring naturally on L limii were all single individuals Unlike L indica and sunflowers L limii was fed upon naturally only sparingly if at all by H vitripennis This strongly indicates that this exotic novel host plant also ranked as a very poor to non-host during the time of the experiment (Brodbeck et al 1990 Andersen et al 2005)
Top-down Predators and Predator Models Predaceous lizards the green anoles Anolis carolinensis Voigt were captured locally in north Florida around buildings euthanized using carbon dioxide and held in sealed plastic bags in a freezer before use Individual specimens were only used twice in the field on two successive days for a total of asymp12 h The lizards were attached to treatment branches in their natural hunting positions using fine wire Lizards were tested with and without the addition of leafhopper cadavers as attractants In addition brown artificial bass fishing lures resembling lizards (Su-per Salt Plus 18 cm lizard Chart pumpkin 002-012 Zoom Bait Co Bogart GA) or green or purple 22 cm worms (Classic Manufacturing Co Clermont FL) were used as predator model treatments Artifi-cial fly fishing lures that resemble spiders were purchased (Gander Mountain Ocala FL brookside flies panfish assortment) and used as models The spider lures were of several colors black or yellow bod-ies with black yellow or white legs Lures were mounted singularly on the treatment branches in plain sight by using the attached hook to embed them in a treatment branch in a natural position Cadavers were not combined in the tests with these treatments
Statistics Five to 10 combinations of the treatment‒control pairs were set up in the field and observed repeatedly in all of the tests to provide the statistical replicates After each observation the responding H vitripennis were removed from the treatment branches to reset the counts to zero for the next replicate In all tests H vit-ripennis cadavers were included as the standard treatment The data were analyzed using Proc GLM procedures in SAS 93 (SAS Institute 2009) or the Mann‒Whitney Rank Sum test in SigmaPlot 110 (Systat Software 2009) when the data did not meet the requirements of normality and heterogeneity of variance The probability used for statistical significance was 005
ResultsConspeciic and Heterospeciic Cadavers Adding H vitripen-
nis cadavers to branches significantly increased the response of H vitripennis to the branches containing the cadavers The presence of the straws used only in the early experiments did not appear to influence the visual response (Fig 2 3 and 4 Table 1) The presence of three cadavers however maximized the response because adding
four to nine cadavers did not significantly increase the number of landing H vitripennis Although numerically more were observed on the treatment H vitripennis did not significantly respond to cadavers of H insolita a species about half the size of H vitripennis (Table 1) H vitripennis responded similarly to cadavers of conspecifics and the heterospecific O nigricans a species that is similar in size but differs in color from H vitripennis (Table 1) H vitripennis also responded significantly to H vitripennis cadavers painted red which is much of the body color of newly emerged adults and to cadavers painted gray as models of fungi-infected cadavers (Boucias et al 2006) (Table 1)
Fig 3
Fig 2 Mean number and SE of H vitripennis responding to varying numbers of leafhopper cadavers placed on straws on a L indica branch Letters indicate signifi cant differences P lt 005
Fig 3 Mean number and SE of H vitripennis responding to varying num-bers of leafhopper cadavers placed on straws on a crape myrtle branch and two branches within 50 cm of the main treatment branch Letters indicate signifi cant differences for comparisons within each branch type Where letters are omitted for a branch type there were no signifi cant differences P lt 005
110 American Entomologist bull Summer 2012
Effects of Acoustical Communication on Response to Cadavers Adding live H vitripennis and presumably intraspecific acoustical signals to the treatment branch significantly increased leafhopper response rate relative to the cadaver control by 53 (141 for treat-ment versus 92 on control)
Models H vitripennis response to plastic or felt models was dependent on color size and orientation (Table 1) H vitripennis responded significantly to models of several colors including black (Fig 1 and 3) and red (Table 1) Red would likely appear to these leafhoppers as dark gray or near black (Table 1) Significant response was observed to a black model similar in size to H vitripennis (3 times 12 mm) and to a model of 10 times 10 mm Models gt12 mm oriented verti-cally or horizontally did not elicit a significant response (Table 1)
Bottom-upmdashNovel Poor or Non-host Plant A significant re-sponse was observed to a treatment of three cadavers placed on the branches of the novel and poor-quality host L limii (Table 1) Total counts for all control branches never exceeded four and these were always single leafhoppers
Top-downmdashPredators H vitripennis were not repelled by the presence of any tested predator or predator model (Table 1) Lure treatments presented without cadavers using a frog a yellow or black spider received 25 on the treatments and 4 on the control and were not significantly different (data not shown) Presentations of lizard predators together with H vitripennis cadavers elicited a significant response similar to that of H vitripennis cadavers alone This further indicates a lack of response to predators and positive response to
conspecifics even when in the visual presence of actual predators or predator-like models
DiscussionH vitripennis responded strongly to the visual presence of H
vitripennis cadavers Moreover low numbers of cadavers (three)
Fig 4 Mean number and SE of H vitripennis (GWSS) responding to cadavers and colored models similar in size to its body size Letters indicate significant differences P lt 005 for comparisons between treat-ments by branch
Fig 4
Table 1 Response of the H vitripennis to various visual treatments placed on host plants
TreatmentsMain Branch
No H vitripennis Responding
Adjacent BranchNo H vitripennis Responding
(P value MannndashWhitney U test) [replicatesa]
MannndashWhitney U Statistic
H vitripennis 232 76 (0001) [165] 52395H insolita 63 30 (0998) NS [65] 21125Oncometopia nigricans 197 41 (0001) [129] 25920H vitripennis painted red 69 44 (gt 005) NS [60] 17915H vitripennis painted gray (fungus) 101 42 (0001) [76] 15470
Novel non-host plantL limii 73 36 (0001) [93] 28890
Predator modelsDead anole lizard + 3 H vitripennis cadavers 23 13 (0008) [19] 4130
Felt modelsBlack model = asympH vitripennis 3 times12 mm 108 33 (0001) [87] 15380
Black model = H vitripennis (horizontal) 37 58 (gt007) NS [71] 21145
Black model 10 times10 mm 81 45 (lt0001) [83] 22145Black model 20 times20 mm 28 31 (0241) NS [38] 5840Black model 10 times30 mm vertical 74
66 (0280) NS [69] 21425
Black model 10 times30 mm horizontal 38
40 (0646) NS [55] 29825
Black model 3 times12 mm on white 2 times2 43
36 (0262) NS [50] 10995
Red model 3 times12 mm 127 34 (lt0001) [65] 6045 Acoustical communication Live H vitripennis 141 92 (lt0004) [115] 152675
The numbers are the total leafhoppers that responded to all replicates within each comparison The statistical comparisons were performed on the actual paired numbers of leafhoppers in all of the replicates that responded to the main branch compared with the adjacent branch containing each of the treatments in the individual rows The P values indicate the probability of a larger MannndashWhitney Rank Sum statistic by chance aReplicates = number of pairs observed where count on at least one of the paired treatments was ne 0
111American Entomologist bull Volume 58 Number 2
maximized the behavior The test cadavers were placed asymp8ndash10 cm apart with the total three-cadaver presentation providing a visual cue le20 cm wide The lack of increase by additional cadavers may indicate that leafhopper visual breadth is lt30 cm H vitripennis also responded to cadavers of the similarly sized heterospecific O nigricans that is often found feeding on the same host plants (ie Ilex spp) in bi-species groups The congeneric species H insolita which is much smaller and predominantly a grass feeder however did not elicit a positive response Although H vitripennis do feed on some grass and herbaceous species concurrently with H insolita we have never observed the two species feeding in heterospecific groups or on the same part of a plant Brodbeck et al (1993) reported that the dietary needs for both species were similar although they prefer to feed on different plant tissues (Turner and Pollard 1959) Percy et al (2008) compared the acoustical signals of H vitripennis and the congener H liturata Ball and reported that the signals were different in most parameters measured Some overlap however was observed such that it cannot be ruled out that acoustical signals produced by one Homalodisca sp could be detected by a related species If there is some signal overlap between the two species then it is possible that H vitripennis could derive benefit from detecting signals produced by H insolita acoustically and visually from group defensive movements that are similar to those of H vitripennis
In an attempt to define the physical parameters of the visual cues perceived and used by H vitripennis a group of models of different sizes and orientations on the host plant were tested A black model of 3times12 mm roughly equal to the body dimensions of an adult H vitripennis evoked significant visual attraction as did a 10 times10 mm
black felt model All other larger models in any plane did not evoke significant response when placed either vertically or horizontally on the plant No response was observed to a black model affixed to a contrasting white square card These results along with the lack of response to predators and the smaller congener H insolita suggest that the visual cues used by H vitripennis when in flight are relatively narrowly defined to the size close to its own body dimensions (width le10 times length le12 mm) The heterospecific O nigricans falls within this size range The failure of H vitripennis to respond to the model on the white card along with the orientation limits may indicate that detection of the visual cues from congeners is contextually related in some manner to that provided as background by the branch or its silhouette in space
Brodbeck et al (1990) and Andersen et al (2008) showed that H vitripennis abundance on a plant is highly correlated with leafhopper feeding rate and is indicative of the underlying xylem nutritional qual-ity H vitripennis are normally highly aggregated in the field among and within available host species (especially on higher quality host plants) although there is never a time when 100 of the population is observed on one host species As a rule in north Florida only a low percentage of branches on fair-to-good hosts are infested and they usually contain more than one leafhopper per branch (Mizell and French 1987) Significant H vitripennis response to cadavers when they were placed on the novel and poor host plant L limii provides very strong support for the use of conspecifics as visual cues by H vitripennis in flight to locate potential host plants for bottom-up risk mitigation Furthermore H vitripennis will respond visually to conspecifics independent of host plant quality and familiarity with a host plant species This result supports the idea that H vitripennis do not assess host plant quality before landing and furthermore that this pivotal decision is likely made using gustation at the level of the
xylem tissue (Hunter and Ullman 1994 Hunter et al1996 Backus et al 2005) The role of plant volatiles remains to be fully described but the research of Patt and Setamou (2007 2010) suggests a potential role in H vitripennis behavior
The importance of bottom-up nutrient profiles on fitness of these leafhoppers is well described (Redak et al 2004) Leafhopper ag-gregation however may be of further benefit by inducing chemical changes in host plant physiology that affect fitness such as feeding rate and fecundity (Karban and Baldwin 1997) Andersen et al (2003 2009) addressed this possibility in experiments with five host species They found that moderate-to-large plants of crape myrtle L indica L and plum Prunus salicina Lindl exhibited no increasing effects on nutrient compounds of leafhopper feeding in large numbers nor did leafhopper feeding behavior change when in groups In small soybean plants Glycine max (L) some nutrients were highly reduced (Andersen et al 2003) In citrus H vitripennis feeding increased seven amino acids in protein form but little change in protein amino acids was observed in Vitis vinifera lsquoChardonnayrsquo In citrus and lsquoChardonnayrsquo grape amino acid increased assimilation efficiency as a result of leafhopper feeding These data suggest that H vitripennis feeding does not induce metabolically inhibitory profiles in the xylem (Andersen et al 2009) This along with the induction of proteins and a slight decrease in free amino acids indicates that there may be some induced benefit in nutrition from H vitripennis aggregation (Andersen et al 2009)
Predation is an important mortality risk to H vitripennis in flight and on the host plant during feeding and oviposition (Lopez et al 2003 Daane et al 2005 Suttle and Hoddle 2006 Fournier et al 2008 Mizell et al 2008) Spiders and lizards are the most impor-tant predators of H vitripennis on the plants (Lopez et al 2003 Fournier et al 2008) Top-down risk mitigation by H vitripennis was assessed indirectly by testing visual response by H vitripennis to lizard predators and simulated predator models (fishing lures) that closely resembled living predators The experimental context most closely approximates the risk to H vitripennis during landing on a new branch the time when little or no information on predator presence on the selected branch is available to arriving H vitripennis This is also the time when H vitripennis presents the largest moving visual stimulus to hidden predators Once on the plant landed H vitripennis stop moving and provide a different visual stimulus to would-be predators they also regain the use of positional camouflage and their own visually based longer-range avoidance defense with the protection it provides As described earlier this protection is partially accrued from conspecifics via the ldquodilutionrdquo effect (Foster and Treherne 1981) Similarly heterospecifics present would also contribute to defense
The only other time that H vitripennis move excessively on the host plant is when the males seek mates and the females oviposit (Mizell et al 2008) Although copulation occurs mostly during the scotophase one might hypothesize that the presence of conspecif-ics would also reduce the risk from predation to more vulnerable copulating pairs and ovipositing females This may also be a selec-tion factor that induces H vitripennis females to oviposit on plants upon which they feed but which do not meet the nutritional needs of their nymphs (Brodbeck et al 2007 Mizell et al 2008) The ability to recognize conspecifics as indicators of suitable host plants allows H vitripennis to feed without moving and eliminates search flights for hours or days proportionally reducing in-flight and landing preda-tion risks These behaviors underscore the potential risks involved in
112 American Entomologist bull Summer 2012
finding hosts the tradeoffs faced by female leafhoppers and the value of conspecific recognition in their mitigation An unusual number of mated leafhopper pairs also were observed among the cadaver treatment branches on the mornings of succeeding test days These observations provide further support for the contention concerning top down risk mitigation via conspecific response
Significant response to avoid predators and artificial models was not observed It is unlikely that an arthropod herbivore would be able to develop a search image with such elasticity that it could encompass the wide range of natural enemies potentially encoun-tered The immediate acceptance of the novel and poor host L limii provides further evidence against the occurrence of a learned host image by searching leafhoppers Response to conspecifics would obviate the necessity of developing a predator search image because conspecifics would usually be present only where predators were absent Thus a type of ldquoenemy-free spacerdquo (Jeffries and Lawton 1984) may be represented by conspecific presence Alternatively relative to the risk of landing on a branch without conspecifics even if the conspecific signal was not entirely truthful about the level of risk the dilution effect from conspecific presence would provide increased protection theoretically in direct proportion to the number of con-specifics present Although foragers in many animal groups have an inherent geometry that causes information asymmetries individuals get more information from closer neighbors (Fernandez-Juricic et al 2004) H vitripennis uses keen visual acuity that likely is coupled with acoustical communication as a mechanism to respond rapidly en masse to approaching predators As a result leafhoppers accrue a more equivalent gain from defensive movement coordination (Fernandez-Juricic et al 2004)
On the host plant H vitripennis can detect and respond (and routinely do so in concert perhaps mediated by acoustic commu-nication) to movement of large objects approaching them from off the plant and to smaller objects on the plant Live lizards do elicit a response from feeding H vitripennis as they move over the host plant and approach leafhoppers (Lopez et al 2003) This suggests the occurrence in H vitripennis of a general visual capability (differ-ent from the visual response to conspecifics) to recognize moving objects that would circumvent the necessity of developing an array of species-specific predator search images
H vitripennis responded significantly to cadavers painted gray that served as a model of H vitripennis infected with the parasitic fungal pathogen H homalodiscae (Boucias et al 2006) H vitripennis cadavers infected with this pathogen remain attached for months on the host plant at feeding sites in the feeding position and infect H vitripennis nymphs and adults The infected cadavers are covered by the fungal mycelia that sporulate in cycles and provide a means to infect uninfected feeding H vitripennis (Lietze et al 2010) H homalodiscae-infected H vitripennis attach tenaciously via rhizoids to the plant bark in the summer and are often found attached to this substrate the following spring This positive visual response may increase transmission of the fungus to H vitripennis
When live presumably acoustically active H vitripennis were added to treatment branches the numbers of H vitripennis increased by 53 on the treated branches (Table 1) H vitripennis unlike most insect herbivores has not been shown to use olfaction at any level for courtship or mating In contrast H vitripennis and many Cicadel-lidae and related insect families use acoustical vibrations propagated through the host plant to communicate during courtship and mating and for other functions (Cocroft 2006) This behavioral modality of
H vitripennis however has received only cursory attention (Percy et al 2008) and its potential effect was eliminated in all except this one experiment through the use of cadavers
With respect to host finding the role of olfaction remains un-clear Patt and Setamou (2007) reported that the response to visual cues by H vitripennis nymphs and adults was enhanced by exposure to plant volatiles Patt and Setamou (2010) demonstrated that H vitripennis nymphs could be conditioned to respond to vanilla as a novel host volatile and as a result they argued that associative learning of chemical profiles could enhance leafhopper location and recognition of host plants These authors also speculated on a potential mechanism use of plant volatile profile emissions (Ninemets et al 2002 2004) in host finding and recognition by adult H vitripennis While an interesting hypothesis much research remains to be done to support these ideas A strong response to conspecifics provides a complimentary or alternative mechanism that would eliminate the need for such highly evolved behaviors However one may hypothesize that the presence of plant volatiles could stimulate a more intense leafhopper visual search for conspe-cifics on the host Nevertheless the ready acceptance of the novel and poor host L limii in the presence of conspecifics argues against the need for leafhoppers to learn a visual or olfactory pattern for specific plant species
Trivial movement within and between host plants and host plant patches is germane to understanding the distribution and fitness of H vitripennis (Mizell et al 2008) H vitripennis are strongly visually oriented while feeding on a host plant they can detect movement from 5 to 10 m away Based on the ostensibly narrow sensory ca-pabilities of H vitripennis (lack of pheromones apparently limited use of olfaction) it is not unexpected that H vitripennis would have evolved visual behavior with multiple functions Moreover this simple capability would obviate the need for higher level behaviors such as associative learning at least for adults (Patt and Setamou 2010) Profiles of xylem chemistry can change as much as 10-fold in one day or from day to day in the absence of any overt visual signals to leafhoppers (Andersen et al1992 Brodbeck et al 2010) Because host plant nutrient quality varies unrecognizably to leafhoppers in time and space and between and within a plant species developing a search image to visually identify specific host plants or character-istics related to their quality appears to be extremely difficult and perhaps of little value Moreover host plants usually do not occur in monocultures a situation that would reinforce short-term pattern recognition By contrast delay in finding hosts within a patch can affect information retention (McLinn and Stephens 2006) Relying too heavily on recent experience can be costly especially if the environment and in this case host plant nutrients change rapidly (Mizell and French 1987 Andersen et al 1989 1992 1993 1995ab Brodbeck et al 1990 19951999 McLinn and Stephens 2006) Animals devalue unreliable signals Models predict signal following when the signal reliability exceeds environmental certaintymdashso that information is valuable Signal value however is not based solely on reliability it also depends on the statistical distribution of the environmental states (McLinn and Stephens 2006) Plant quality and predation risks are in constant flux in the case of H vitripennis and extremely variable in time and space H vitripennis handling of bottom-up and top-down forces is tied closely together because the potential payoff and consequences from both accrue on the host plant following a single decision to land H vitripennis appar-ently can make a decision to land based solely on a visual response
113American Entomologist bull Volume 58 Number 2
to conspecifics on host plant branches An arrestment response is then likely reinforced on the plant via acoustical communication as gustation behavior is initiated
Thus presence of conspecifics indicates a ldquosafety zonerdquo for host plant testing and likely provides high-quality (truthfulness of signal) information because all H vitripennis in any one location are under more or less the same dietary constraints Notwithstanding that O nigricans may have somewhat different nutritional requirements it may be important in the mitigation of bottom-up and top-down forces The presence of one or (usually) multiple conspecifics is likely a signal indicating a reasonably high threshold quality of nutrients or else the duration of conspecifics remaining on a host plant would be short This perhaps explains why three conspecifics analogous to the quorum decision of Ward et al (2008) maximize the response Three may be enough to compensate for any variations in individual hunger rate or other uninterpretable behaviors (females not feeding or ovipositing males that feed less) that may be ongoing and could affect the ldquotruth in signalrdquo presented by conspecifics
Arthropod populations face a variety of environmental risks to which they must adapt that can be described within the bottom-up and top-down heuristic model however ldquoNo creature can do all thingsrdquo and genetic and other constraints are many Thus tradeoffs are required for organisms to adapt and sustain over evolutionary time (Rolf 1992) Activity or movement is highly related to resource acquisition but it is also related to mortality risks a cost and as a result mediates one of the most general tradeoffs faced by organ-isms (Werner 1992) Information gathering also comes at a cost because of the loss of time potentially spent on other activities (Eliassen et al 2007) Moreover where resources vary temporally or for example predation risk is highly unpredictable the value of information decreases with time such that a forager should place greater value on the newest available information (Eliassen et al 2007) An individualrsquos response to conspecifics makes it possible to collect a broad range of real-time biological data that may reduce the tradeoffs and costs imposed by required activity Foraging suc-cess and predator avoidance behaviors directly related to fitness are strongly affected as a result (Lazarus 1979 Foster and Treherne 1981 Lawrence 1985 Stamps 1987 Turchin 1987) The environment of xylophagous leafhoppers is highly stochastic in time and space and their extreme polyphagy requires exposure to a wide array of plant structures changing chemical profiles and associated predators Response to conspecifics offers a simple parsimonious mechanism to eliminate the need to develop sophisticated physiological or behavioral mechanisms to handle this array of visual and chemical cues that accompany the extreme uncertainty presented by resource finding and predator avoidance
In conclusion H vitripennis exhibits at least two major types of visually based behaviors While in the normal feeding position (Bex-tine et al 2005) they can detect movement from asymp10 m distance and move behind the branch to hide themselves from the approaching object This response is often done synchronously by the leafhoppers present More importantly when searching for and landing on a host plant where the risk of predation may be high they can respond to cues presented by conspecifics These consist of a narrowly defined stimulus roughly equal in size to the H vitripennis body but not gt12 mm To our knowledge this is the first report of an arthropod exhibit-ing behavioral parsimony in which one signal a visual cue (response to conspecifics) is used to mitigate immediate risks from bottom-up and top-down forces In the present case leafhoppers exhibit a ldquofixedrdquo
response response to conspecifics to bottom-up and top-down forces independent of their value in the environmental context Avail-ability and spatial distribution of host plants and their nutritional quality are critical to the survival and fitness of H vitripennis The availability of xylem fluid within and between host plants changes continuously and rapidly over time and space requiring much trivial movement and emigration by leafhoppers These natural fluctuations in plant xylem chemistry are the primary basis of the bottom-up and top-down risks to xylophagous leafhoppers because they determine the movement and seasonal use of host plants by H vitripennis (be-havior and performance) (Mizell and French 1987 Andersen et al 1989 1992 1995ab 2005 2008 Brodbeck et al 1990 19951999) Previously several novel physiological and behavioral adaptations critical to leafhopper survival were documented (Mizell et al 2008) Response to conspecifics during host plant searching movements is another novel but fundamentally important behavior exhibited by these interesting disease vectors that allow them to survive and endure as xylophages
AcknowledgmentsThe research was partially funded by a grant to R F Mizell III and P
C Andersen from the California Department of Food and Agriculture Piercersquos DiseasemdashGlassy-winged Sharpshooter Research Program We thank Gary W Knox for use of the experimental plot containing the Lagerstroemia limii We thank Tobin Northfield Joseph Patt and Arash Rashad for helpful comments on a previous draft of the manuscript
References CitedAndersen P C B V Brodbeck and R M Mizell III 1989 Metabolism
of amino acids organic acids and sugars extracted from the xylem fluid of four host plants by Homalodisca coagulata Entomol Exp Appl 50 149ndash159
Andersen P C B V Brodbeck and R M Mizell III 1992 Feeding by the leafhopper Homalodisca coagulata in relation to xylem fluid chemistry and tension J Insect Physiol 38 611ndash622
Andersen P C B V Brodbeck and R M Mizell III 1993 Diurnal variations in the chemistry of xylem fluid from Lagerstroemia indica an endogenous circadian rhythm Physiol Plant 89 783-790
Andersen P C B V Brodbeck and R M Mizell III 1995a Diurnal variation in tension osmolarity and the composition of nitrogen and carbon assimilates in xylem fluid of Prunus persica Vitis hybrid and Prunus communis J Am Hortic Sci 120 600ndash604
Andersen P C B V Brodbeck and R M Mizell III 1995b Water stress- and nutrient-mediated changes in the water relations and the amino acids organic acids and sugars in xylem fluid of Prunus salicina and Lagerstroemia indica J Am Soc Hortic Sci 120 36ndash42
Andersen P C B V Brodbeck and R M Mizell III 2003 Plant and insect characteristics in response to increasing density of Homalodisca coagulata on three host species a quantification of assimilate extraction Entomol Exp Appl 107 57ndash68
Andersen P C B V Brodbeck R M Mizell III and S Oden 2005 The abundance and feeding of Homalodisca coagulata (Homoptera Auche-norrhyncha Cicadellidae) on Vitis genotypes in north Florida Environ Entomol 34 466ndash478
Andersen PC R M Mizell III B V Brodbeck T Beckman and G Krewer 2008 Abundance and consumption rates of glassy-winged sharpshooter (Hemiptera Cicadellidae) on peaches and plums J En-tomol Sci 43 394ndash407
Andersen P C B V Brodbeck and R M Mizell III 2009 Assimilation efficiency of free and protein amino acids by the glassy-winged sharp-shooter Homalodisca vitripennis (Germar) (Hemiptera Cicadellidae Cicadellinae) feeding on Citrus sinensis and Vitis vinifera Fla Entomol 92 116ndash122
Backus E J Habibi F Yan and M Ellersieck 2005 Stylet penetration by adult Homalodisca coagulata on grape electrical penetration graph
114 American Entomologist bull Summer 2012
waveform characterization tissue correlation and possible implica-tions for transmission of Xylella fastidiosa Ann Entomol Soc Am 98 787ndash813
Baker P S 1979 Flying locust visual responses in a radial wind tunnel J Comp Physiol 131 39ndash47
Bextine B R B C Jackson D B Harshman and Miller 2005 Homalo-disca coagulata (Hemiptera Cicadellidae) feeding posture Ann Ento-mol Soc Am 98 814ndash819
Boucias D G D W Scharf S E Breaux S D Purcell and R M Mizell III 2006 Studies on the fungi associated with the glassy-winged sharpshooter Homalodisca coagulata with emphasis on a new species Hirsutella homalodiscae nom prov BioControl 52 231ndash258
Brodbeck BV R M Mizell III and P C Andersen 1990 Amino acids as determinants of host preference for the xylemfeeding leafhopper Homalodisca coagulata Oecologia 83 338ndash345
Brodbeck B V P C Andersen and R M Mizell III 1993 Physiological and behavioral adaptations of three species of leafhoppers in response to the dilute nutrient content of xylem fluid J Insect Physiol 39 73ndash81
Brodbeck B V P C Andersen and R M Mizell III 1995 Differential utilization of nutrients during development by the xylophagous leafhop-per Homalodisca coagulata Entomol Exp Appl 75 279ndash289
Brodbeck BV P C Andersen and R M Mizell III 1999 The effects of total dietary nitrogen and amino acid balance on the development of xylophagous leafhoppers Arch Insect Biochem Physiol 42 37ndash50
Brodbeck B V P C Andersen and R M Mizell III 2007 Preference-performance linkage of the xylem feeding leafhopper Homalodisca vit-ripennis (Hemiptera Cicadellidae) Environ Entomol 36 1512ndash1522
Brodbeck B V P C Andersen and R M Mizell III 2011 Nutrient media-tion of behavioral plasticity and resource allocation in a xylem-feeding leafhopper Oecol 165 111-122
Cocroft R B H J Shugart K T Konrad and K Tibbs 2006 Variation in plant substrates and its consequences for insect vibrational com-munication Ethology 112 779ndash789
Collett T S and M F Land 1978 How hoverflies compute interception courses J Comp Physiol 125 191ndash204
Coolen I O Dangles and J Casas 2005 Social learning in non-colonial insects Curr Biol 15 1931-1935
Daane K M Johnson T Ruiz and J M Hashim 2005 Host plant influence on glassy-winged sharpshooter and its natural enemies pp 100ndash104 In Proceedings 2005 Piercersquos Disease Research Symposium California Department of Food and Agriculture Sacramento
Dukas R 2001 Effects of perceived danger on flower choice by bees Ecol Lett 4 327ndash333
Dukas R and D H Morse 2003 Crab spiders affect flower visitation by bees Oikos 101 157ndash163
Eliassen S E C Jargensen M Mangel and J Garl 2007 Exploration or exploitation life expectancy changes the value of learning in foraging strategies Oikos 116 513ndash523
Fernandez-Juricic E B Kerr P Bednekoff and D Stephens 2004 When are two heads better than one Visual perception and informa-tion transfer affect vigilance coordination in foraging groups Behav Ecol 6 898ndash906
Foster W A and J E Treherne 1981 Evidence for the dilution effect in the selfish herd from fish predation on a marine insect Nature 293 466ndash467
Fournier V J Hagler K Daane J de Leoacuten and R Groves 2008 Iden-tifying the predator complex of Homalodisca vitripennis (Hemiptera Cicadellidae) a comparative study of the efficacy of an ELISA and PCR gut content bioassay Oecologia 157 629ndash640
Giraldeau L T J Valone and J J Templeton 2002 Potential disadvan-tages of using socially acquired information Phil Trans R Soc Lond B 357 1559ndash1566
Hamiliton WD 1971 Geometry for the selfish herd J Theror Biol 31295-311
Hairston N G J E Smith and L B Slobodkin 1960 Community struc-ture population control and competition Am Midl Nat 106 249ndash257
Hunter W B and D E Ullman 1994 Precibarial and cibarial chemosen-sillae in the western flower thrips Frankliniella occidentalis (Pergande)(Thysanoptera Thripidae) Int J Insect Morphol Embryol 23 69ndash83
Hunter W B E Hiebert S Webb J E Polston and J H Tsai 1996 Precibarial and cibarial chemosensilla in the whitefly Bemisia tabaci
(Gennadius) (Homoptera Aleyrodidae) Int J Insect Morphol Embryol 25 295ndash304
Jeffries M J and J H Lawton 1984 Enemy-free space and the structure of ecological communities Biol J Linn Soc 23 269-286
Karban R and L T Baldwin 1997 Induced response to herbivory University of Chicago Press Chicago
Land M F and T S Collett 1974 Chasing behavior of houseflies (Fannia canicularis) J Comp Physiol 80 331ndash357
Lawrence E S 1985 Vigilance during ldquoeasyrdquo and ldquodifficultrdquo foraging tasks Anim Behav 33 1373ndash1375
Lawton J H and S McNeil1979 Between the devil and the deep blue sea on the problem of being a herbivore pp 223ndash244 In R M Anderson B D Turner and L R Taylor [Eds] Population dynamics Blackwell Oxford England Blackwell Sci Publ 434 pp
Lazarus J 1979 Flock size and behavior in captive red-billed weaverbirds (Quelea quelea) implications for social facilitation and the functions of flocking Behavior 71 1ndash2
Lietze V R M Mizell III and D G Boucias 2011 Transmission of the mycopathogen Hirsutella spp to nymphs and adults of the glassy-winged sharpshooters Homalodisca vitripennis (=coagulata) in the greenhouse Fla Entomol 94 106-108
Lopez R F R Mizell III P C Andersen and B V Brodbeck 2003 Impact of natural enemies on populations of the glassy-winged sharpshooter Homalodisca coagulata (Say) in north Florida In R VanDriesche [Ed] First international symposium on biological control of arthropods US Dep Agric For Ser Pub FHTET-03-05 484ndash486
Mainman G A D Straw and M H Dickson 2008 A simple vision-based algorithm for decision making in flying Drosophila Curr Biol 18 464ndash470
McLinn C and D W Stephens 2006 What makes information valu-able signal reliability and environmental uncertainty Anim Behav 71 1119ndash1129
Mizell R F III and W J French 1987 Leafhopper vectors of phony peach disease feeding site preference and survival on infected and uninfected peach and seasonal response to selected host plants J Entomol Sci 22 1122
Mizell R F III P C Andersen P C Phillips and P Oevering 2005 Population dynamics and interactions between the glassy-winged sharpshooter and the host plants in response to California phenology pp 117ndash121 In Proceedings 2005 Piercersquos Disease Research Symposium California Department of Food and Agriculture Sacramento
Mizell R F III C Tipping P C Andersen B V Brodbeck T Northfield and W Hunter W 2008 Behavioral model for the glassy-winged sharp-shooter Homalodisca vitripennis (Hemiptera Cicadellidae) optimiza-tion of host plant utilization and management implications Environ Entomol 37 1049ndash1062
Mizell R F III PC Andersen and B V Brodbeck 2009 Improved detec-tion monitoring and management of the glassy-winged sharpshooter pp 40ndash43 In Proceedings 2009 Piercersquos Disease Research Symposium California Department of Food and Agriculture Sacramento
Muller KL 1998 The role of conspecifics in habitat settlement in a ter-ritorial grasshopper Anim Behav 56 479ndash485
Ninemets Uuml M Riechstein M Staudt G Siefert and J Tenhunen 2002 Stomatal constraints may affect emission of oxygenated monoterpenoids from foliage of Pinus pinea Plant Physiol 130 1371ndash1385
Ninemets Uuml F Loreto and M Riechstein 2004 Physiological and physiochemical controls on foliar volatile organic compound emissions Trends Plant Sci 9 180ndash186
Northfield T R F Mizell III P C Andersen B V Brodbeck T C Riddle and W B Hunter 2009 Dispersal patch leaving and aggregation of the glassy-winged sharpshooter Homalodisca vitripennis (Hemiptera Cicadellidae) Environ Entomol 38 183ndash191
Patt J and M Setamou 2007 Olfactory and visual stimuli affecting host plant detection in Homalodisca coagulata (Hemiptera Cicadellidae) Environ Entomol 36 142ndash150
Patt J and M Setamou 2010 Recognition of novel volatile cues by the nymph of the glassy-winged sharpshooter Homalodisca vitripennis (Cicadellidae) J Insect Behav DOI 101007s10905-010-9214-z 23 290-302
Percy D M E A Boyd and M S Hoddle 2008 Observations of acoustical signaling in three sharpshooters Homalodisca vitripennis Homalodisca
115American Entomologist bull Volume 58 Number 2
liturata and Graphocephala atropunctata (Hemiptera Cicadellidae) Ann Entomol Soc 101 253ndash259
Power M E 1992 Top-down and bottom-up forces in food webs do plants have primacy Ecologia 73 733ndash746
Price P W C E Bouton P Gross B A McPheron J N Thompson and A E Wise 1980 Interactions among three trophic levels influence of plants on interactions between insect herbivores and natural enemies Annu Rev Ecol Syst 11 41ndash66
Redak R M J Blua K L Lopes A H Purcell R F Mizell III and P C Andersen 2004 The biology and control of xylem sap-feeding insect vectors of Xylella fastidiosa and their relation to disease epidemiology Annu Rev Entomol 49 243ndash270
Rolf D A 1992 The evolution of life history traits Chapman amp Hall NYSandlin E A 2000 Cue use affects resource subdivision among three
coexisting hummingbird species Behav Ecol 11 550-559SAS Institute 2009 Proc GLM SAS 93 Statistical Analysis System Cary NCShrewsbury P and M Raupp 2008 Do top-down or bottom-up forces
determine Stephanitis pyroides abundance in urban landscapes Ecol Appl 16 262ndash273
Systat Software 2009 SigmaPlot for Windows 110 Systat Software Bangalore India
Stamps J A 1987 Conspecifics as cues to territory quality a preference of juvenile lizards (Anolis aeneus) for previously used territories Am Nat 129 629ndash642
Stamps J A 1988 Conspecific attraction and aggregation in territorial species Am Nat 131 330ndash347
Thompson J D S C Peterson and L D Harder 1987 Response to traplining bumble bees to competition experiments shifts in feeding location and efficiency Oecologia 71 295ndash300
Tipping C R F Mizell III and P C Andersen 2004 Adaptations impor-tant for the dispersal of immature stages of three species of leafhopper (Homoptera Cicadellidae) Fla Entomol 87 372ndash379
Triapitsyn S V R F Mizell III J L Bossert and J E Carlton 1998 Egg parasitoids of glassy-winged sharpshooter Homalodisca coagulata (Homoptera Cicadellidae) Fla Entomol 81 243ndash251
Turner W F and H N Pollard 1959 Life histories and behavior of five insect vectors of phony peach disease Tech Bull US Dep Agric 1188
Turchin P 1987 The role of aggregation in the response of Mexican bean beetles to host-plant density Oecologia 71 577ndash582
Valone T J 1989 Group foraging public information and patch simula-tion Oikos 56 357ndash363
Walker M and T H Jones 2001 Relative roles of top-down and bottom-up forces in terrestrial tritrophic plant-insect herbivore-natural enemy systems Oikos 93 177ndash187
Ward A J W D J T Sumpter I D Couzin P J B Hart and J Krause 2008 Quorum decision-making facilitates information transfer in fish shoals PNAS 105 6948ndash6953
Wells J M B C Raju H Y Hung W G Weisburg L Mandelco-Paul and D J Brenner 1987 Xylella fastidiosa gen nov sp nov gram-negative xylem-limited fastidious plant bacteria related to Xanthomonas spp Int J Syst Bacteriol 37 136ndash143
Werner E E 1992 Individual behavior and higher-order species interac-tions Am Nat (suppl) 140 S5ndashS32
White T R C 1978 The importance of relative shortage of food in animal ecology Oecologia 33 71ndash86
Williams G C 1966 Adaptation and natural selection Princeton University Press Princeton NJ
Wu D S C Daugherty S E Van Aken G H Pai K L Watkins H Khouri L J Tallon J M Zaborsky H E Dunbar P L Tran N A Moran J A Eisen 2006 Metabolic complementarity and genomics of the dual symbiosis of sharpshooters PloS-Biology 4 e188
Russell F Mizell III Is a Professor of Entomology (rfmizellufledu) Peter C Andersen is a Professor of Horticulture (peaufledu) and Brent V Brod-beck (bvbufledu) is a Biological Scientist all located at the University of Floridarsquos North Florida Research and Education Center-Quincy 155 Research Rd Quincy Fl 32351 Wayne B Hunter (whunterushrlarsusdagov) is a research entomologist with the USDA Agricultural Research Service at the US Horticultural Research Lab 2001 South Rock Rd Ft Pierce FL 34945 For over 20 years they have collaborated in research on the biology ecology and behavior of the leafhopper vectors of Xylella fastidiosa
With our high qualityLepidoptera Eggs and LarvaeBeet Army Worm bull Cabbage LooperCorn Earworm bull Diamondback MothFall Armyworm bull Tobacco Budworm
Save Money Save Time Get Results
Lepidoptera Eggs and LarvaeInsect Diets and Ingredients
Rearing Supplies and Equipment
Entomology Division
800-996-9908 US and Canada908-996-2155 Internationalsalesinsectrearingcom
wwwinsectrearingcom
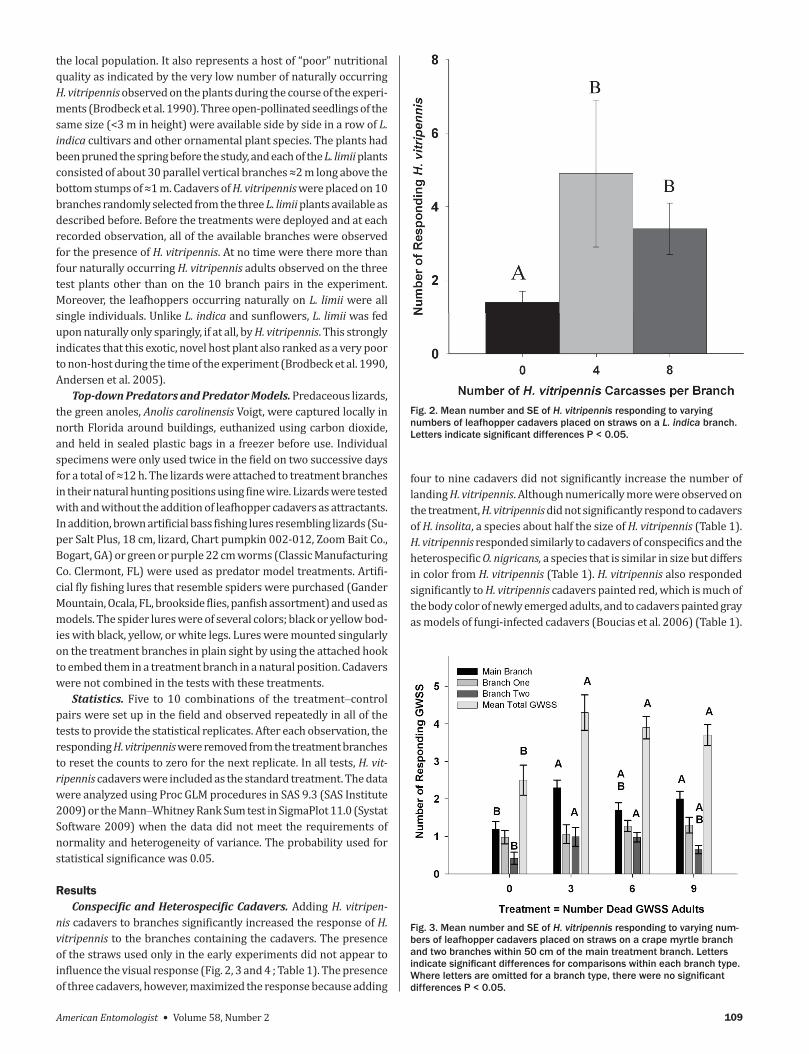
109American Entomologist bull Volume 58 Number 2
the local population It also represents a host of ldquopoorrdquo nutritional quality as indicated by the very low number of naturally occurring H vitripennis observed on the plants during the course of the experi-ments (Brodbeck et al 1990) Three open-pollinated seedlings of the same size (lt3 m in height) were available side by side in a row of L indica cultivars and other ornamental plant species The plants had been pruned the spring before the study and each of the L limii plants consisted of about 30 parallel vertical branches asymp2 m long above the bottom stumps of asymp1 m Cadavers of H vitripennis were placed on 10 branches randomly selected from the three L limii plants available as described before Before the treatments were deployed and at each recorded observation all of the available branches were observed for the presence of H vitripennis At no time were there more than four naturally occurring H vitripennis adults observed on the three test plants other than on the 10 branch pairs in the experiment Moreover the leafhoppers occurring naturally on L limii were all single individuals Unlike L indica and sunflowers L limii was fed upon naturally only sparingly if at all by H vitripennis This strongly indicates that this exotic novel host plant also ranked as a very poor to non-host during the time of the experiment (Brodbeck et al 1990 Andersen et al 2005)
Top-down Predators and Predator Models Predaceous lizards the green anoles Anolis carolinensis Voigt were captured locally in north Florida around buildings euthanized using carbon dioxide and held in sealed plastic bags in a freezer before use Individual specimens were only used twice in the field on two successive days for a total of asymp12 h The lizards were attached to treatment branches in their natural hunting positions using fine wire Lizards were tested with and without the addition of leafhopper cadavers as attractants In addition brown artificial bass fishing lures resembling lizards (Su-per Salt Plus 18 cm lizard Chart pumpkin 002-012 Zoom Bait Co Bogart GA) or green or purple 22 cm worms (Classic Manufacturing Co Clermont FL) were used as predator model treatments Artifi-cial fly fishing lures that resemble spiders were purchased (Gander Mountain Ocala FL brookside flies panfish assortment) and used as models The spider lures were of several colors black or yellow bod-ies with black yellow or white legs Lures were mounted singularly on the treatment branches in plain sight by using the attached hook to embed them in a treatment branch in a natural position Cadavers were not combined in the tests with these treatments
Statistics Five to 10 combinations of the treatment‒control pairs were set up in the field and observed repeatedly in all of the tests to provide the statistical replicates After each observation the responding H vitripennis were removed from the treatment branches to reset the counts to zero for the next replicate In all tests H vit-ripennis cadavers were included as the standard treatment The data were analyzed using Proc GLM procedures in SAS 93 (SAS Institute 2009) or the Mann‒Whitney Rank Sum test in SigmaPlot 110 (Systat Software 2009) when the data did not meet the requirements of normality and heterogeneity of variance The probability used for statistical significance was 005
ResultsConspeciic and Heterospeciic Cadavers Adding H vitripen-
nis cadavers to branches significantly increased the response of H vitripennis to the branches containing the cadavers The presence of the straws used only in the early experiments did not appear to influence the visual response (Fig 2 3 and 4 Table 1) The presence of three cadavers however maximized the response because adding
four to nine cadavers did not significantly increase the number of landing H vitripennis Although numerically more were observed on the treatment H vitripennis did not significantly respond to cadavers of H insolita a species about half the size of H vitripennis (Table 1) H vitripennis responded similarly to cadavers of conspecifics and the heterospecific O nigricans a species that is similar in size but differs in color from H vitripennis (Table 1) H vitripennis also responded significantly to H vitripennis cadavers painted red which is much of the body color of newly emerged adults and to cadavers painted gray as models of fungi-infected cadavers (Boucias et al 2006) (Table 1)
Fig 3
Fig 2 Mean number and SE of H vitripennis responding to varying numbers of leafhopper cadavers placed on straws on a L indica branch Letters indicate signifi cant differences P lt 005
Fig 3 Mean number and SE of H vitripennis responding to varying num-bers of leafhopper cadavers placed on straws on a crape myrtle branch and two branches within 50 cm of the main treatment branch Letters indicate signifi cant differences for comparisons within each branch type Where letters are omitted for a branch type there were no signifi cant differences P lt 005
110 American Entomologist bull Summer 2012
Effects of Acoustical Communication on Response to Cadavers Adding live H vitripennis and presumably intraspecific acoustical signals to the treatment branch significantly increased leafhopper response rate relative to the cadaver control by 53 (141 for treat-ment versus 92 on control)
Models H vitripennis response to plastic or felt models was dependent on color size and orientation (Table 1) H vitripennis responded significantly to models of several colors including black (Fig 1 and 3) and red (Table 1) Red would likely appear to these leafhoppers as dark gray or near black (Table 1) Significant response was observed to a black model similar in size to H vitripennis (3 times 12 mm) and to a model of 10 times 10 mm Models gt12 mm oriented verti-cally or horizontally did not elicit a significant response (Table 1)
Bottom-upmdashNovel Poor or Non-host Plant A significant re-sponse was observed to a treatment of three cadavers placed on the branches of the novel and poor-quality host L limii (Table 1) Total counts for all control branches never exceeded four and these were always single leafhoppers
Top-downmdashPredators H vitripennis were not repelled by the presence of any tested predator or predator model (Table 1) Lure treatments presented without cadavers using a frog a yellow or black spider received 25 on the treatments and 4 on the control and were not significantly different (data not shown) Presentations of lizard predators together with H vitripennis cadavers elicited a significant response similar to that of H vitripennis cadavers alone This further indicates a lack of response to predators and positive response to
conspecifics even when in the visual presence of actual predators or predator-like models
DiscussionH vitripennis responded strongly to the visual presence of H
vitripennis cadavers Moreover low numbers of cadavers (three)
Fig 4 Mean number and SE of H vitripennis (GWSS) responding to cadavers and colored models similar in size to its body size Letters indicate significant differences P lt 005 for comparisons between treat-ments by branch
Fig 4
Table 1 Response of the H vitripennis to various visual treatments placed on host plants
TreatmentsMain Branch
No H vitripennis Responding
Adjacent BranchNo H vitripennis Responding
(P value MannndashWhitney U test) [replicatesa]
MannndashWhitney U Statistic
H vitripennis 232 76 (0001) [165] 52395H insolita 63 30 (0998) NS [65] 21125Oncometopia nigricans 197 41 (0001) [129] 25920H vitripennis painted red 69 44 (gt 005) NS [60] 17915H vitripennis painted gray (fungus) 101 42 (0001) [76] 15470
Novel non-host plantL limii 73 36 (0001) [93] 28890
Predator modelsDead anole lizard + 3 H vitripennis cadavers 23 13 (0008) [19] 4130
Felt modelsBlack model = asympH vitripennis 3 times12 mm 108 33 (0001) [87] 15380
Black model = H vitripennis (horizontal) 37 58 (gt007) NS [71] 21145
Black model 10 times10 mm 81 45 (lt0001) [83] 22145Black model 20 times20 mm 28 31 (0241) NS [38] 5840Black model 10 times30 mm vertical 74
66 (0280) NS [69] 21425
Black model 10 times30 mm horizontal 38
40 (0646) NS [55] 29825
Black model 3 times12 mm on white 2 times2 43
36 (0262) NS [50] 10995
Red model 3 times12 mm 127 34 (lt0001) [65] 6045 Acoustical communication Live H vitripennis 141 92 (lt0004) [115] 152675
The numbers are the total leafhoppers that responded to all replicates within each comparison The statistical comparisons were performed on the actual paired numbers of leafhoppers in all of the replicates that responded to the main branch compared with the adjacent branch containing each of the treatments in the individual rows The P values indicate the probability of a larger MannndashWhitney Rank Sum statistic by chance aReplicates = number of pairs observed where count on at least one of the paired treatments was ne 0
111American Entomologist bull Volume 58 Number 2
maximized the behavior The test cadavers were placed asymp8ndash10 cm apart with the total three-cadaver presentation providing a visual cue le20 cm wide The lack of increase by additional cadavers may indicate that leafhopper visual breadth is lt30 cm H vitripennis also responded to cadavers of the similarly sized heterospecific O nigricans that is often found feeding on the same host plants (ie Ilex spp) in bi-species groups The congeneric species H insolita which is much smaller and predominantly a grass feeder however did not elicit a positive response Although H vitripennis do feed on some grass and herbaceous species concurrently with H insolita we have never observed the two species feeding in heterospecific groups or on the same part of a plant Brodbeck et al (1993) reported that the dietary needs for both species were similar although they prefer to feed on different plant tissues (Turner and Pollard 1959) Percy et al (2008) compared the acoustical signals of H vitripennis and the congener H liturata Ball and reported that the signals were different in most parameters measured Some overlap however was observed such that it cannot be ruled out that acoustical signals produced by one Homalodisca sp could be detected by a related species If there is some signal overlap between the two species then it is possible that H vitripennis could derive benefit from detecting signals produced by H insolita acoustically and visually from group defensive movements that are similar to those of H vitripennis
In an attempt to define the physical parameters of the visual cues perceived and used by H vitripennis a group of models of different sizes and orientations on the host plant were tested A black model of 3times12 mm roughly equal to the body dimensions of an adult H vitripennis evoked significant visual attraction as did a 10 times10 mm
black felt model All other larger models in any plane did not evoke significant response when placed either vertically or horizontally on the plant No response was observed to a black model affixed to a contrasting white square card These results along with the lack of response to predators and the smaller congener H insolita suggest that the visual cues used by H vitripennis when in flight are relatively narrowly defined to the size close to its own body dimensions (width le10 times length le12 mm) The heterospecific O nigricans falls within this size range The failure of H vitripennis to respond to the model on the white card along with the orientation limits may indicate that detection of the visual cues from congeners is contextually related in some manner to that provided as background by the branch or its silhouette in space
Brodbeck et al (1990) and Andersen et al (2008) showed that H vitripennis abundance on a plant is highly correlated with leafhopper feeding rate and is indicative of the underlying xylem nutritional qual-ity H vitripennis are normally highly aggregated in the field among and within available host species (especially on higher quality host plants) although there is never a time when 100 of the population is observed on one host species As a rule in north Florida only a low percentage of branches on fair-to-good hosts are infested and they usually contain more than one leafhopper per branch (Mizell and French 1987) Significant H vitripennis response to cadavers when they were placed on the novel and poor host plant L limii provides very strong support for the use of conspecifics as visual cues by H vitripennis in flight to locate potential host plants for bottom-up risk mitigation Furthermore H vitripennis will respond visually to conspecifics independent of host plant quality and familiarity with a host plant species This result supports the idea that H vitripennis do not assess host plant quality before landing and furthermore that this pivotal decision is likely made using gustation at the level of the
xylem tissue (Hunter and Ullman 1994 Hunter et al1996 Backus et al 2005) The role of plant volatiles remains to be fully described but the research of Patt and Setamou (2007 2010) suggests a potential role in H vitripennis behavior
The importance of bottom-up nutrient profiles on fitness of these leafhoppers is well described (Redak et al 2004) Leafhopper ag-gregation however may be of further benefit by inducing chemical changes in host plant physiology that affect fitness such as feeding rate and fecundity (Karban and Baldwin 1997) Andersen et al (2003 2009) addressed this possibility in experiments with five host species They found that moderate-to-large plants of crape myrtle L indica L and plum Prunus salicina Lindl exhibited no increasing effects on nutrient compounds of leafhopper feeding in large numbers nor did leafhopper feeding behavior change when in groups In small soybean plants Glycine max (L) some nutrients were highly reduced (Andersen et al 2003) In citrus H vitripennis feeding increased seven amino acids in protein form but little change in protein amino acids was observed in Vitis vinifera lsquoChardonnayrsquo In citrus and lsquoChardonnayrsquo grape amino acid increased assimilation efficiency as a result of leafhopper feeding These data suggest that H vitripennis feeding does not induce metabolically inhibitory profiles in the xylem (Andersen et al 2009) This along with the induction of proteins and a slight decrease in free amino acids indicates that there may be some induced benefit in nutrition from H vitripennis aggregation (Andersen et al 2009)
Predation is an important mortality risk to H vitripennis in flight and on the host plant during feeding and oviposition (Lopez et al 2003 Daane et al 2005 Suttle and Hoddle 2006 Fournier et al 2008 Mizell et al 2008) Spiders and lizards are the most impor-tant predators of H vitripennis on the plants (Lopez et al 2003 Fournier et al 2008) Top-down risk mitigation by H vitripennis was assessed indirectly by testing visual response by H vitripennis to lizard predators and simulated predator models (fishing lures) that closely resembled living predators The experimental context most closely approximates the risk to H vitripennis during landing on a new branch the time when little or no information on predator presence on the selected branch is available to arriving H vitripennis This is also the time when H vitripennis presents the largest moving visual stimulus to hidden predators Once on the plant landed H vitripennis stop moving and provide a different visual stimulus to would-be predators they also regain the use of positional camouflage and their own visually based longer-range avoidance defense with the protection it provides As described earlier this protection is partially accrued from conspecifics via the ldquodilutionrdquo effect (Foster and Treherne 1981) Similarly heterospecifics present would also contribute to defense
The only other time that H vitripennis move excessively on the host plant is when the males seek mates and the females oviposit (Mizell et al 2008) Although copulation occurs mostly during the scotophase one might hypothesize that the presence of conspecif-ics would also reduce the risk from predation to more vulnerable copulating pairs and ovipositing females This may also be a selec-tion factor that induces H vitripennis females to oviposit on plants upon which they feed but which do not meet the nutritional needs of their nymphs (Brodbeck et al 2007 Mizell et al 2008) The ability to recognize conspecifics as indicators of suitable host plants allows H vitripennis to feed without moving and eliminates search flights for hours or days proportionally reducing in-flight and landing preda-tion risks These behaviors underscore the potential risks involved in
112 American Entomologist bull Summer 2012
finding hosts the tradeoffs faced by female leafhoppers and the value of conspecific recognition in their mitigation An unusual number of mated leafhopper pairs also were observed among the cadaver treatment branches on the mornings of succeeding test days These observations provide further support for the contention concerning top down risk mitigation via conspecific response
Significant response to avoid predators and artificial models was not observed It is unlikely that an arthropod herbivore would be able to develop a search image with such elasticity that it could encompass the wide range of natural enemies potentially encoun-tered The immediate acceptance of the novel and poor host L limii provides further evidence against the occurrence of a learned host image by searching leafhoppers Response to conspecifics would obviate the necessity of developing a predator search image because conspecifics would usually be present only where predators were absent Thus a type of ldquoenemy-free spacerdquo (Jeffries and Lawton 1984) may be represented by conspecific presence Alternatively relative to the risk of landing on a branch without conspecifics even if the conspecific signal was not entirely truthful about the level of risk the dilution effect from conspecific presence would provide increased protection theoretically in direct proportion to the number of con-specifics present Although foragers in many animal groups have an inherent geometry that causes information asymmetries individuals get more information from closer neighbors (Fernandez-Juricic et al 2004) H vitripennis uses keen visual acuity that likely is coupled with acoustical communication as a mechanism to respond rapidly en masse to approaching predators As a result leafhoppers accrue a more equivalent gain from defensive movement coordination (Fernandez-Juricic et al 2004)
On the host plant H vitripennis can detect and respond (and routinely do so in concert perhaps mediated by acoustic commu-nication) to movement of large objects approaching them from off the plant and to smaller objects on the plant Live lizards do elicit a response from feeding H vitripennis as they move over the host plant and approach leafhoppers (Lopez et al 2003) This suggests the occurrence in H vitripennis of a general visual capability (differ-ent from the visual response to conspecifics) to recognize moving objects that would circumvent the necessity of developing an array of species-specific predator search images
H vitripennis responded significantly to cadavers painted gray that served as a model of H vitripennis infected with the parasitic fungal pathogen H homalodiscae (Boucias et al 2006) H vitripennis cadavers infected with this pathogen remain attached for months on the host plant at feeding sites in the feeding position and infect H vitripennis nymphs and adults The infected cadavers are covered by the fungal mycelia that sporulate in cycles and provide a means to infect uninfected feeding H vitripennis (Lietze et al 2010) H homalodiscae-infected H vitripennis attach tenaciously via rhizoids to the plant bark in the summer and are often found attached to this substrate the following spring This positive visual response may increase transmission of the fungus to H vitripennis
When live presumably acoustically active H vitripennis were added to treatment branches the numbers of H vitripennis increased by 53 on the treated branches (Table 1) H vitripennis unlike most insect herbivores has not been shown to use olfaction at any level for courtship or mating In contrast H vitripennis and many Cicadel-lidae and related insect families use acoustical vibrations propagated through the host plant to communicate during courtship and mating and for other functions (Cocroft 2006) This behavioral modality of
H vitripennis however has received only cursory attention (Percy et al 2008) and its potential effect was eliminated in all except this one experiment through the use of cadavers
With respect to host finding the role of olfaction remains un-clear Patt and Setamou (2007) reported that the response to visual cues by H vitripennis nymphs and adults was enhanced by exposure to plant volatiles Patt and Setamou (2010) demonstrated that H vitripennis nymphs could be conditioned to respond to vanilla as a novel host volatile and as a result they argued that associative learning of chemical profiles could enhance leafhopper location and recognition of host plants These authors also speculated on a potential mechanism use of plant volatile profile emissions (Ninemets et al 2002 2004) in host finding and recognition by adult H vitripennis While an interesting hypothesis much research remains to be done to support these ideas A strong response to conspecifics provides a complimentary or alternative mechanism that would eliminate the need for such highly evolved behaviors However one may hypothesize that the presence of plant volatiles could stimulate a more intense leafhopper visual search for conspe-cifics on the host Nevertheless the ready acceptance of the novel and poor host L limii in the presence of conspecifics argues against the need for leafhoppers to learn a visual or olfactory pattern for specific plant species
Trivial movement within and between host plants and host plant patches is germane to understanding the distribution and fitness of H vitripennis (Mizell et al 2008) H vitripennis are strongly visually oriented while feeding on a host plant they can detect movement from 5 to 10 m away Based on the ostensibly narrow sensory ca-pabilities of H vitripennis (lack of pheromones apparently limited use of olfaction) it is not unexpected that H vitripennis would have evolved visual behavior with multiple functions Moreover this simple capability would obviate the need for higher level behaviors such as associative learning at least for adults (Patt and Setamou 2010) Profiles of xylem chemistry can change as much as 10-fold in one day or from day to day in the absence of any overt visual signals to leafhoppers (Andersen et al1992 Brodbeck et al 2010) Because host plant nutrient quality varies unrecognizably to leafhoppers in time and space and between and within a plant species developing a search image to visually identify specific host plants or character-istics related to their quality appears to be extremely difficult and perhaps of little value Moreover host plants usually do not occur in monocultures a situation that would reinforce short-term pattern recognition By contrast delay in finding hosts within a patch can affect information retention (McLinn and Stephens 2006) Relying too heavily on recent experience can be costly especially if the environment and in this case host plant nutrients change rapidly (Mizell and French 1987 Andersen et al 1989 1992 1993 1995ab Brodbeck et al 1990 19951999 McLinn and Stephens 2006) Animals devalue unreliable signals Models predict signal following when the signal reliability exceeds environmental certaintymdashso that information is valuable Signal value however is not based solely on reliability it also depends on the statistical distribution of the environmental states (McLinn and Stephens 2006) Plant quality and predation risks are in constant flux in the case of H vitripennis and extremely variable in time and space H vitripennis handling of bottom-up and top-down forces is tied closely together because the potential payoff and consequences from both accrue on the host plant following a single decision to land H vitripennis appar-ently can make a decision to land based solely on a visual response
113American Entomologist bull Volume 58 Number 2
to conspecifics on host plant branches An arrestment response is then likely reinforced on the plant via acoustical communication as gustation behavior is initiated
Thus presence of conspecifics indicates a ldquosafety zonerdquo for host plant testing and likely provides high-quality (truthfulness of signal) information because all H vitripennis in any one location are under more or less the same dietary constraints Notwithstanding that O nigricans may have somewhat different nutritional requirements it may be important in the mitigation of bottom-up and top-down forces The presence of one or (usually) multiple conspecifics is likely a signal indicating a reasonably high threshold quality of nutrients or else the duration of conspecifics remaining on a host plant would be short This perhaps explains why three conspecifics analogous to the quorum decision of Ward et al (2008) maximize the response Three may be enough to compensate for any variations in individual hunger rate or other uninterpretable behaviors (females not feeding or ovipositing males that feed less) that may be ongoing and could affect the ldquotruth in signalrdquo presented by conspecifics
Arthropod populations face a variety of environmental risks to which they must adapt that can be described within the bottom-up and top-down heuristic model however ldquoNo creature can do all thingsrdquo and genetic and other constraints are many Thus tradeoffs are required for organisms to adapt and sustain over evolutionary time (Rolf 1992) Activity or movement is highly related to resource acquisition but it is also related to mortality risks a cost and as a result mediates one of the most general tradeoffs faced by organ-isms (Werner 1992) Information gathering also comes at a cost because of the loss of time potentially spent on other activities (Eliassen et al 2007) Moreover where resources vary temporally or for example predation risk is highly unpredictable the value of information decreases with time such that a forager should place greater value on the newest available information (Eliassen et al 2007) An individualrsquos response to conspecifics makes it possible to collect a broad range of real-time biological data that may reduce the tradeoffs and costs imposed by required activity Foraging suc-cess and predator avoidance behaviors directly related to fitness are strongly affected as a result (Lazarus 1979 Foster and Treherne 1981 Lawrence 1985 Stamps 1987 Turchin 1987) The environment of xylophagous leafhoppers is highly stochastic in time and space and their extreme polyphagy requires exposure to a wide array of plant structures changing chemical profiles and associated predators Response to conspecifics offers a simple parsimonious mechanism to eliminate the need to develop sophisticated physiological or behavioral mechanisms to handle this array of visual and chemical cues that accompany the extreme uncertainty presented by resource finding and predator avoidance
In conclusion H vitripennis exhibits at least two major types of visually based behaviors While in the normal feeding position (Bex-tine et al 2005) they can detect movement from asymp10 m distance and move behind the branch to hide themselves from the approaching object This response is often done synchronously by the leafhoppers present More importantly when searching for and landing on a host plant where the risk of predation may be high they can respond to cues presented by conspecifics These consist of a narrowly defined stimulus roughly equal in size to the H vitripennis body but not gt12 mm To our knowledge this is the first report of an arthropod exhibit-ing behavioral parsimony in which one signal a visual cue (response to conspecifics) is used to mitigate immediate risks from bottom-up and top-down forces In the present case leafhoppers exhibit a ldquofixedrdquo
response response to conspecifics to bottom-up and top-down forces independent of their value in the environmental context Avail-ability and spatial distribution of host plants and their nutritional quality are critical to the survival and fitness of H vitripennis The availability of xylem fluid within and between host plants changes continuously and rapidly over time and space requiring much trivial movement and emigration by leafhoppers These natural fluctuations in plant xylem chemistry are the primary basis of the bottom-up and top-down risks to xylophagous leafhoppers because they determine the movement and seasonal use of host plants by H vitripennis (be-havior and performance) (Mizell and French 1987 Andersen et al 1989 1992 1995ab 2005 2008 Brodbeck et al 1990 19951999) Previously several novel physiological and behavioral adaptations critical to leafhopper survival were documented (Mizell et al 2008) Response to conspecifics during host plant searching movements is another novel but fundamentally important behavior exhibited by these interesting disease vectors that allow them to survive and endure as xylophages
AcknowledgmentsThe research was partially funded by a grant to R F Mizell III and P
C Andersen from the California Department of Food and Agriculture Piercersquos DiseasemdashGlassy-winged Sharpshooter Research Program We thank Gary W Knox for use of the experimental plot containing the Lagerstroemia limii We thank Tobin Northfield Joseph Patt and Arash Rashad for helpful comments on a previous draft of the manuscript
References CitedAndersen P C B V Brodbeck and R M Mizell III 1989 Metabolism
of amino acids organic acids and sugars extracted from the xylem fluid of four host plants by Homalodisca coagulata Entomol Exp Appl 50 149ndash159
Andersen P C B V Brodbeck and R M Mizell III 1992 Feeding by the leafhopper Homalodisca coagulata in relation to xylem fluid chemistry and tension J Insect Physiol 38 611ndash622
Andersen P C B V Brodbeck and R M Mizell III 1993 Diurnal variations in the chemistry of xylem fluid from Lagerstroemia indica an endogenous circadian rhythm Physiol Plant 89 783-790
Andersen P C B V Brodbeck and R M Mizell III 1995a Diurnal variation in tension osmolarity and the composition of nitrogen and carbon assimilates in xylem fluid of Prunus persica Vitis hybrid and Prunus communis J Am Hortic Sci 120 600ndash604
Andersen P C B V Brodbeck and R M Mizell III 1995b Water stress- and nutrient-mediated changes in the water relations and the amino acids organic acids and sugars in xylem fluid of Prunus salicina and Lagerstroemia indica J Am Soc Hortic Sci 120 36ndash42
Andersen P C B V Brodbeck and R M Mizell III 2003 Plant and insect characteristics in response to increasing density of Homalodisca coagulata on three host species a quantification of assimilate extraction Entomol Exp Appl 107 57ndash68
Andersen P C B V Brodbeck R M Mizell III and S Oden 2005 The abundance and feeding of Homalodisca coagulata (Homoptera Auche-norrhyncha Cicadellidae) on Vitis genotypes in north Florida Environ Entomol 34 466ndash478
Andersen PC R M Mizell III B V Brodbeck T Beckman and G Krewer 2008 Abundance and consumption rates of glassy-winged sharpshooter (Hemiptera Cicadellidae) on peaches and plums J En-tomol Sci 43 394ndash407
Andersen P C B V Brodbeck and R M Mizell III 2009 Assimilation efficiency of free and protein amino acids by the glassy-winged sharp-shooter Homalodisca vitripennis (Germar) (Hemiptera Cicadellidae Cicadellinae) feeding on Citrus sinensis and Vitis vinifera Fla Entomol 92 116ndash122
Backus E J Habibi F Yan and M Ellersieck 2005 Stylet penetration by adult Homalodisca coagulata on grape electrical penetration graph
114 American Entomologist bull Summer 2012
waveform characterization tissue correlation and possible implica-tions for transmission of Xylella fastidiosa Ann Entomol Soc Am 98 787ndash813
Baker P S 1979 Flying locust visual responses in a radial wind tunnel J Comp Physiol 131 39ndash47
Bextine B R B C Jackson D B Harshman and Miller 2005 Homalo-disca coagulata (Hemiptera Cicadellidae) feeding posture Ann Ento-mol Soc Am 98 814ndash819
Boucias D G D W Scharf S E Breaux S D Purcell and R M Mizell III 2006 Studies on the fungi associated with the glassy-winged sharpshooter Homalodisca coagulata with emphasis on a new species Hirsutella homalodiscae nom prov BioControl 52 231ndash258
Brodbeck BV R M Mizell III and P C Andersen 1990 Amino acids as determinants of host preference for the xylemfeeding leafhopper Homalodisca coagulata Oecologia 83 338ndash345
Brodbeck B V P C Andersen and R M Mizell III 1993 Physiological and behavioral adaptations of three species of leafhoppers in response to the dilute nutrient content of xylem fluid J Insect Physiol 39 73ndash81
Brodbeck B V P C Andersen and R M Mizell III 1995 Differential utilization of nutrients during development by the xylophagous leafhop-per Homalodisca coagulata Entomol Exp Appl 75 279ndash289
Brodbeck BV P C Andersen and R M Mizell III 1999 The effects of total dietary nitrogen and amino acid balance on the development of xylophagous leafhoppers Arch Insect Biochem Physiol 42 37ndash50
Brodbeck B V P C Andersen and R M Mizell III 2007 Preference-performance linkage of the xylem feeding leafhopper Homalodisca vit-ripennis (Hemiptera Cicadellidae) Environ Entomol 36 1512ndash1522
Brodbeck B V P C Andersen and R M Mizell III 2011 Nutrient media-tion of behavioral plasticity and resource allocation in a xylem-feeding leafhopper Oecol 165 111-122
Cocroft R B H J Shugart K T Konrad and K Tibbs 2006 Variation in plant substrates and its consequences for insect vibrational com-munication Ethology 112 779ndash789
Collett T S and M F Land 1978 How hoverflies compute interception courses J Comp Physiol 125 191ndash204
Coolen I O Dangles and J Casas 2005 Social learning in non-colonial insects Curr Biol 15 1931-1935
Daane K M Johnson T Ruiz and J M Hashim 2005 Host plant influence on glassy-winged sharpshooter and its natural enemies pp 100ndash104 In Proceedings 2005 Piercersquos Disease Research Symposium California Department of Food and Agriculture Sacramento
Dukas R 2001 Effects of perceived danger on flower choice by bees Ecol Lett 4 327ndash333
Dukas R and D H Morse 2003 Crab spiders affect flower visitation by bees Oikos 101 157ndash163
Eliassen S E C Jargensen M Mangel and J Garl 2007 Exploration or exploitation life expectancy changes the value of learning in foraging strategies Oikos 116 513ndash523
Fernandez-Juricic E B Kerr P Bednekoff and D Stephens 2004 When are two heads better than one Visual perception and informa-tion transfer affect vigilance coordination in foraging groups Behav Ecol 6 898ndash906
Foster W A and J E Treherne 1981 Evidence for the dilution effect in the selfish herd from fish predation on a marine insect Nature 293 466ndash467
Fournier V J Hagler K Daane J de Leoacuten and R Groves 2008 Iden-tifying the predator complex of Homalodisca vitripennis (Hemiptera Cicadellidae) a comparative study of the efficacy of an ELISA and PCR gut content bioassay Oecologia 157 629ndash640
Giraldeau L T J Valone and J J Templeton 2002 Potential disadvan-tages of using socially acquired information Phil Trans R Soc Lond B 357 1559ndash1566
Hamiliton WD 1971 Geometry for the selfish herd J Theror Biol 31295-311
Hairston N G J E Smith and L B Slobodkin 1960 Community struc-ture population control and competition Am Midl Nat 106 249ndash257
Hunter W B and D E Ullman 1994 Precibarial and cibarial chemosen-sillae in the western flower thrips Frankliniella occidentalis (Pergande)(Thysanoptera Thripidae) Int J Insect Morphol Embryol 23 69ndash83
Hunter W B E Hiebert S Webb J E Polston and J H Tsai 1996 Precibarial and cibarial chemosensilla in the whitefly Bemisia tabaci
(Gennadius) (Homoptera Aleyrodidae) Int J Insect Morphol Embryol 25 295ndash304
Jeffries M J and J H Lawton 1984 Enemy-free space and the structure of ecological communities Biol J Linn Soc 23 269-286
Karban R and L T Baldwin 1997 Induced response to herbivory University of Chicago Press Chicago
Land M F and T S Collett 1974 Chasing behavior of houseflies (Fannia canicularis) J Comp Physiol 80 331ndash357
Lawrence E S 1985 Vigilance during ldquoeasyrdquo and ldquodifficultrdquo foraging tasks Anim Behav 33 1373ndash1375
Lawton J H and S McNeil1979 Between the devil and the deep blue sea on the problem of being a herbivore pp 223ndash244 In R M Anderson B D Turner and L R Taylor [Eds] Population dynamics Blackwell Oxford England Blackwell Sci Publ 434 pp
Lazarus J 1979 Flock size and behavior in captive red-billed weaverbirds (Quelea quelea) implications for social facilitation and the functions of flocking Behavior 71 1ndash2
Lietze V R M Mizell III and D G Boucias 2011 Transmission of the mycopathogen Hirsutella spp to nymphs and adults of the glassy-winged sharpshooters Homalodisca vitripennis (=coagulata) in the greenhouse Fla Entomol 94 106-108
Lopez R F R Mizell III P C Andersen and B V Brodbeck 2003 Impact of natural enemies on populations of the glassy-winged sharpshooter Homalodisca coagulata (Say) in north Florida In R VanDriesche [Ed] First international symposium on biological control of arthropods US Dep Agric For Ser Pub FHTET-03-05 484ndash486
Mainman G A D Straw and M H Dickson 2008 A simple vision-based algorithm for decision making in flying Drosophila Curr Biol 18 464ndash470
McLinn C and D W Stephens 2006 What makes information valu-able signal reliability and environmental uncertainty Anim Behav 71 1119ndash1129
Mizell R F III and W J French 1987 Leafhopper vectors of phony peach disease feeding site preference and survival on infected and uninfected peach and seasonal response to selected host plants J Entomol Sci 22 1122
Mizell R F III P C Andersen P C Phillips and P Oevering 2005 Population dynamics and interactions between the glassy-winged sharpshooter and the host plants in response to California phenology pp 117ndash121 In Proceedings 2005 Piercersquos Disease Research Symposium California Department of Food and Agriculture Sacramento
Mizell R F III C Tipping P C Andersen B V Brodbeck T Northfield and W Hunter W 2008 Behavioral model for the glassy-winged sharp-shooter Homalodisca vitripennis (Hemiptera Cicadellidae) optimiza-tion of host plant utilization and management implications Environ Entomol 37 1049ndash1062
Mizell R F III PC Andersen and B V Brodbeck 2009 Improved detec-tion monitoring and management of the glassy-winged sharpshooter pp 40ndash43 In Proceedings 2009 Piercersquos Disease Research Symposium California Department of Food and Agriculture Sacramento
Muller KL 1998 The role of conspecifics in habitat settlement in a ter-ritorial grasshopper Anim Behav 56 479ndash485
Ninemets Uuml M Riechstein M Staudt G Siefert and J Tenhunen 2002 Stomatal constraints may affect emission of oxygenated monoterpenoids from foliage of Pinus pinea Plant Physiol 130 1371ndash1385
Ninemets Uuml F Loreto and M Riechstein 2004 Physiological and physiochemical controls on foliar volatile organic compound emissions Trends Plant Sci 9 180ndash186
Northfield T R F Mizell III P C Andersen B V Brodbeck T C Riddle and W B Hunter 2009 Dispersal patch leaving and aggregation of the glassy-winged sharpshooter Homalodisca vitripennis (Hemiptera Cicadellidae) Environ Entomol 38 183ndash191
Patt J and M Setamou 2007 Olfactory and visual stimuli affecting host plant detection in Homalodisca coagulata (Hemiptera Cicadellidae) Environ Entomol 36 142ndash150
Patt J and M Setamou 2010 Recognition of novel volatile cues by the nymph of the glassy-winged sharpshooter Homalodisca vitripennis (Cicadellidae) J Insect Behav DOI 101007s10905-010-9214-z 23 290-302
Percy D M E A Boyd and M S Hoddle 2008 Observations of acoustical signaling in three sharpshooters Homalodisca vitripennis Homalodisca
115American Entomologist bull Volume 58 Number 2
liturata and Graphocephala atropunctata (Hemiptera Cicadellidae) Ann Entomol Soc 101 253ndash259
Power M E 1992 Top-down and bottom-up forces in food webs do plants have primacy Ecologia 73 733ndash746
Price P W C E Bouton P Gross B A McPheron J N Thompson and A E Wise 1980 Interactions among three trophic levels influence of plants on interactions between insect herbivores and natural enemies Annu Rev Ecol Syst 11 41ndash66
Redak R M J Blua K L Lopes A H Purcell R F Mizell III and P C Andersen 2004 The biology and control of xylem sap-feeding insect vectors of Xylella fastidiosa and their relation to disease epidemiology Annu Rev Entomol 49 243ndash270
Rolf D A 1992 The evolution of life history traits Chapman amp Hall NYSandlin E A 2000 Cue use affects resource subdivision among three
coexisting hummingbird species Behav Ecol 11 550-559SAS Institute 2009 Proc GLM SAS 93 Statistical Analysis System Cary NCShrewsbury P and M Raupp 2008 Do top-down or bottom-up forces
determine Stephanitis pyroides abundance in urban landscapes Ecol Appl 16 262ndash273
Systat Software 2009 SigmaPlot for Windows 110 Systat Software Bangalore India
Stamps J A 1987 Conspecifics as cues to territory quality a preference of juvenile lizards (Anolis aeneus) for previously used territories Am Nat 129 629ndash642
Stamps J A 1988 Conspecific attraction and aggregation in territorial species Am Nat 131 330ndash347
Thompson J D S C Peterson and L D Harder 1987 Response to traplining bumble bees to competition experiments shifts in feeding location and efficiency Oecologia 71 295ndash300
Tipping C R F Mizell III and P C Andersen 2004 Adaptations impor-tant for the dispersal of immature stages of three species of leafhopper (Homoptera Cicadellidae) Fla Entomol 87 372ndash379
Triapitsyn S V R F Mizell III J L Bossert and J E Carlton 1998 Egg parasitoids of glassy-winged sharpshooter Homalodisca coagulata (Homoptera Cicadellidae) Fla Entomol 81 243ndash251
Turner W F and H N Pollard 1959 Life histories and behavior of five insect vectors of phony peach disease Tech Bull US Dep Agric 1188
Turchin P 1987 The role of aggregation in the response of Mexican bean beetles to host-plant density Oecologia 71 577ndash582
Valone T J 1989 Group foraging public information and patch simula-tion Oikos 56 357ndash363
Walker M and T H Jones 2001 Relative roles of top-down and bottom-up forces in terrestrial tritrophic plant-insect herbivore-natural enemy systems Oikos 93 177ndash187
Ward A J W D J T Sumpter I D Couzin P J B Hart and J Krause 2008 Quorum decision-making facilitates information transfer in fish shoals PNAS 105 6948ndash6953
Wells J M B C Raju H Y Hung W G Weisburg L Mandelco-Paul and D J Brenner 1987 Xylella fastidiosa gen nov sp nov gram-negative xylem-limited fastidious plant bacteria related to Xanthomonas spp Int J Syst Bacteriol 37 136ndash143
Werner E E 1992 Individual behavior and higher-order species interac-tions Am Nat (suppl) 140 S5ndashS32
White T R C 1978 The importance of relative shortage of food in animal ecology Oecologia 33 71ndash86
Williams G C 1966 Adaptation and natural selection Princeton University Press Princeton NJ
Wu D S C Daugherty S E Van Aken G H Pai K L Watkins H Khouri L J Tallon J M Zaborsky H E Dunbar P L Tran N A Moran J A Eisen 2006 Metabolic complementarity and genomics of the dual symbiosis of sharpshooters PloS-Biology 4 e188
Russell F Mizell III Is a Professor of Entomology (rfmizellufledu) Peter C Andersen is a Professor of Horticulture (peaufledu) and Brent V Brod-beck (bvbufledu) is a Biological Scientist all located at the University of Floridarsquos North Florida Research and Education Center-Quincy 155 Research Rd Quincy Fl 32351 Wayne B Hunter (whunterushrlarsusdagov) is a research entomologist with the USDA Agricultural Research Service at the US Horticultural Research Lab 2001 South Rock Rd Ft Pierce FL 34945 For over 20 years they have collaborated in research on the biology ecology and behavior of the leafhopper vectors of Xylella fastidiosa
With our high qualityLepidoptera Eggs and LarvaeBeet Army Worm bull Cabbage LooperCorn Earworm bull Diamondback MothFall Armyworm bull Tobacco Budworm
Save Money Save Time Get Results
Lepidoptera Eggs and LarvaeInsect Diets and Ingredients
Rearing Supplies and Equipment
Entomology Division
800-996-9908 US and Canada908-996-2155 Internationalsalesinsectrearingcom
wwwinsectrearingcom
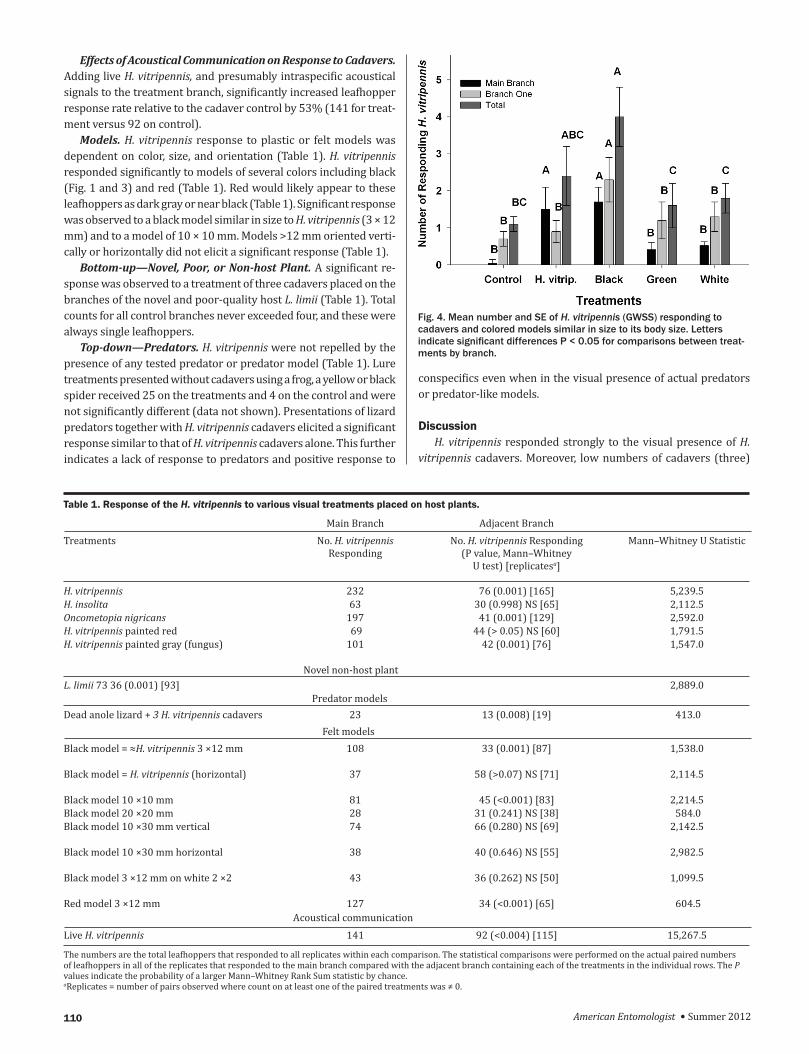
110 American Entomologist bull Summer 2012
Effects of Acoustical Communication on Response to Cadavers Adding live H vitripennis and presumably intraspecific acoustical signals to the treatment branch significantly increased leafhopper response rate relative to the cadaver control by 53 (141 for treat-ment versus 92 on control)
Models H vitripennis response to plastic or felt models was dependent on color size and orientation (Table 1) H vitripennis responded significantly to models of several colors including black (Fig 1 and 3) and red (Table 1) Red would likely appear to these leafhoppers as dark gray or near black (Table 1) Significant response was observed to a black model similar in size to H vitripennis (3 times 12 mm) and to a model of 10 times 10 mm Models gt12 mm oriented verti-cally or horizontally did not elicit a significant response (Table 1)
Bottom-upmdashNovel Poor or Non-host Plant A significant re-sponse was observed to a treatment of three cadavers placed on the branches of the novel and poor-quality host L limii (Table 1) Total counts for all control branches never exceeded four and these were always single leafhoppers
Top-downmdashPredators H vitripennis were not repelled by the presence of any tested predator or predator model (Table 1) Lure treatments presented without cadavers using a frog a yellow or black spider received 25 on the treatments and 4 on the control and were not significantly different (data not shown) Presentations of lizard predators together with H vitripennis cadavers elicited a significant response similar to that of H vitripennis cadavers alone This further indicates a lack of response to predators and positive response to
conspecifics even when in the visual presence of actual predators or predator-like models
DiscussionH vitripennis responded strongly to the visual presence of H
vitripennis cadavers Moreover low numbers of cadavers (three)
Fig 4 Mean number and SE of H vitripennis (GWSS) responding to cadavers and colored models similar in size to its body size Letters indicate significant differences P lt 005 for comparisons between treat-ments by branch
Fig 4
Table 1 Response of the H vitripennis to various visual treatments placed on host plants
TreatmentsMain Branch
No H vitripennis Responding
Adjacent BranchNo H vitripennis Responding
(P value MannndashWhitney U test) [replicatesa]
MannndashWhitney U Statistic
H vitripennis 232 76 (0001) [165] 52395H insolita 63 30 (0998) NS [65] 21125Oncometopia nigricans 197 41 (0001) [129] 25920H vitripennis painted red 69 44 (gt 005) NS [60] 17915H vitripennis painted gray (fungus) 101 42 (0001) [76] 15470
Novel non-host plantL limii 73 36 (0001) [93] 28890
Predator modelsDead anole lizard + 3 H vitripennis cadavers 23 13 (0008) [19] 4130
Felt modelsBlack model = asympH vitripennis 3 times12 mm 108 33 (0001) [87] 15380
Black model = H vitripennis (horizontal) 37 58 (gt007) NS [71] 21145
Black model 10 times10 mm 81 45 (lt0001) [83] 22145Black model 20 times20 mm 28 31 (0241) NS [38] 5840Black model 10 times30 mm vertical 74
66 (0280) NS [69] 21425
Black model 10 times30 mm horizontal 38
40 (0646) NS [55] 29825
Black model 3 times12 mm on white 2 times2 43
36 (0262) NS [50] 10995
Red model 3 times12 mm 127 34 (lt0001) [65] 6045 Acoustical communication Live H vitripennis 141 92 (lt0004) [115] 152675
The numbers are the total leafhoppers that responded to all replicates within each comparison The statistical comparisons were performed on the actual paired numbers of leafhoppers in all of the replicates that responded to the main branch compared with the adjacent branch containing each of the treatments in the individual rows The P values indicate the probability of a larger MannndashWhitney Rank Sum statistic by chance aReplicates = number of pairs observed where count on at least one of the paired treatments was ne 0
111American Entomologist bull Volume 58 Number 2
maximized the behavior The test cadavers were placed asymp8ndash10 cm apart with the total three-cadaver presentation providing a visual cue le20 cm wide The lack of increase by additional cadavers may indicate that leafhopper visual breadth is lt30 cm H vitripennis also responded to cadavers of the similarly sized heterospecific O nigricans that is often found feeding on the same host plants (ie Ilex spp) in bi-species groups The congeneric species H insolita which is much smaller and predominantly a grass feeder however did not elicit a positive response Although H vitripennis do feed on some grass and herbaceous species concurrently with H insolita we have never observed the two species feeding in heterospecific groups or on the same part of a plant Brodbeck et al (1993) reported that the dietary needs for both species were similar although they prefer to feed on different plant tissues (Turner and Pollard 1959) Percy et al (2008) compared the acoustical signals of H vitripennis and the congener H liturata Ball and reported that the signals were different in most parameters measured Some overlap however was observed such that it cannot be ruled out that acoustical signals produced by one Homalodisca sp could be detected by a related species If there is some signal overlap between the two species then it is possible that H vitripennis could derive benefit from detecting signals produced by H insolita acoustically and visually from group defensive movements that are similar to those of H vitripennis
In an attempt to define the physical parameters of the visual cues perceived and used by H vitripennis a group of models of different sizes and orientations on the host plant were tested A black model of 3times12 mm roughly equal to the body dimensions of an adult H vitripennis evoked significant visual attraction as did a 10 times10 mm
black felt model All other larger models in any plane did not evoke significant response when placed either vertically or horizontally on the plant No response was observed to a black model affixed to a contrasting white square card These results along with the lack of response to predators and the smaller congener H insolita suggest that the visual cues used by H vitripennis when in flight are relatively narrowly defined to the size close to its own body dimensions (width le10 times length le12 mm) The heterospecific O nigricans falls within this size range The failure of H vitripennis to respond to the model on the white card along with the orientation limits may indicate that detection of the visual cues from congeners is contextually related in some manner to that provided as background by the branch or its silhouette in space
Brodbeck et al (1990) and Andersen et al (2008) showed that H vitripennis abundance on a plant is highly correlated with leafhopper feeding rate and is indicative of the underlying xylem nutritional qual-ity H vitripennis are normally highly aggregated in the field among and within available host species (especially on higher quality host plants) although there is never a time when 100 of the population is observed on one host species As a rule in north Florida only a low percentage of branches on fair-to-good hosts are infested and they usually contain more than one leafhopper per branch (Mizell and French 1987) Significant H vitripennis response to cadavers when they were placed on the novel and poor host plant L limii provides very strong support for the use of conspecifics as visual cues by H vitripennis in flight to locate potential host plants for bottom-up risk mitigation Furthermore H vitripennis will respond visually to conspecifics independent of host plant quality and familiarity with a host plant species This result supports the idea that H vitripennis do not assess host plant quality before landing and furthermore that this pivotal decision is likely made using gustation at the level of the
xylem tissue (Hunter and Ullman 1994 Hunter et al1996 Backus et al 2005) The role of plant volatiles remains to be fully described but the research of Patt and Setamou (2007 2010) suggests a potential role in H vitripennis behavior
The importance of bottom-up nutrient profiles on fitness of these leafhoppers is well described (Redak et al 2004) Leafhopper ag-gregation however may be of further benefit by inducing chemical changes in host plant physiology that affect fitness such as feeding rate and fecundity (Karban and Baldwin 1997) Andersen et al (2003 2009) addressed this possibility in experiments with five host species They found that moderate-to-large plants of crape myrtle L indica L and plum Prunus salicina Lindl exhibited no increasing effects on nutrient compounds of leafhopper feeding in large numbers nor did leafhopper feeding behavior change when in groups In small soybean plants Glycine max (L) some nutrients were highly reduced (Andersen et al 2003) In citrus H vitripennis feeding increased seven amino acids in protein form but little change in protein amino acids was observed in Vitis vinifera lsquoChardonnayrsquo In citrus and lsquoChardonnayrsquo grape amino acid increased assimilation efficiency as a result of leafhopper feeding These data suggest that H vitripennis feeding does not induce metabolically inhibitory profiles in the xylem (Andersen et al 2009) This along with the induction of proteins and a slight decrease in free amino acids indicates that there may be some induced benefit in nutrition from H vitripennis aggregation (Andersen et al 2009)
Predation is an important mortality risk to H vitripennis in flight and on the host plant during feeding and oviposition (Lopez et al 2003 Daane et al 2005 Suttle and Hoddle 2006 Fournier et al 2008 Mizell et al 2008) Spiders and lizards are the most impor-tant predators of H vitripennis on the plants (Lopez et al 2003 Fournier et al 2008) Top-down risk mitigation by H vitripennis was assessed indirectly by testing visual response by H vitripennis to lizard predators and simulated predator models (fishing lures) that closely resembled living predators The experimental context most closely approximates the risk to H vitripennis during landing on a new branch the time when little or no information on predator presence on the selected branch is available to arriving H vitripennis This is also the time when H vitripennis presents the largest moving visual stimulus to hidden predators Once on the plant landed H vitripennis stop moving and provide a different visual stimulus to would-be predators they also regain the use of positional camouflage and their own visually based longer-range avoidance defense with the protection it provides As described earlier this protection is partially accrued from conspecifics via the ldquodilutionrdquo effect (Foster and Treherne 1981) Similarly heterospecifics present would also contribute to defense
The only other time that H vitripennis move excessively on the host plant is when the males seek mates and the females oviposit (Mizell et al 2008) Although copulation occurs mostly during the scotophase one might hypothesize that the presence of conspecif-ics would also reduce the risk from predation to more vulnerable copulating pairs and ovipositing females This may also be a selec-tion factor that induces H vitripennis females to oviposit on plants upon which they feed but which do not meet the nutritional needs of their nymphs (Brodbeck et al 2007 Mizell et al 2008) The ability to recognize conspecifics as indicators of suitable host plants allows H vitripennis to feed without moving and eliminates search flights for hours or days proportionally reducing in-flight and landing preda-tion risks These behaviors underscore the potential risks involved in
112 American Entomologist bull Summer 2012
finding hosts the tradeoffs faced by female leafhoppers and the value of conspecific recognition in their mitigation An unusual number of mated leafhopper pairs also were observed among the cadaver treatment branches on the mornings of succeeding test days These observations provide further support for the contention concerning top down risk mitigation via conspecific response
Significant response to avoid predators and artificial models was not observed It is unlikely that an arthropod herbivore would be able to develop a search image with such elasticity that it could encompass the wide range of natural enemies potentially encoun-tered The immediate acceptance of the novel and poor host L limii provides further evidence against the occurrence of a learned host image by searching leafhoppers Response to conspecifics would obviate the necessity of developing a predator search image because conspecifics would usually be present only where predators were absent Thus a type of ldquoenemy-free spacerdquo (Jeffries and Lawton 1984) may be represented by conspecific presence Alternatively relative to the risk of landing on a branch without conspecifics even if the conspecific signal was not entirely truthful about the level of risk the dilution effect from conspecific presence would provide increased protection theoretically in direct proportion to the number of con-specifics present Although foragers in many animal groups have an inherent geometry that causes information asymmetries individuals get more information from closer neighbors (Fernandez-Juricic et al 2004) H vitripennis uses keen visual acuity that likely is coupled with acoustical communication as a mechanism to respond rapidly en masse to approaching predators As a result leafhoppers accrue a more equivalent gain from defensive movement coordination (Fernandez-Juricic et al 2004)
On the host plant H vitripennis can detect and respond (and routinely do so in concert perhaps mediated by acoustic commu-nication) to movement of large objects approaching them from off the plant and to smaller objects on the plant Live lizards do elicit a response from feeding H vitripennis as they move over the host plant and approach leafhoppers (Lopez et al 2003) This suggests the occurrence in H vitripennis of a general visual capability (differ-ent from the visual response to conspecifics) to recognize moving objects that would circumvent the necessity of developing an array of species-specific predator search images
H vitripennis responded significantly to cadavers painted gray that served as a model of H vitripennis infected with the parasitic fungal pathogen H homalodiscae (Boucias et al 2006) H vitripennis cadavers infected with this pathogen remain attached for months on the host plant at feeding sites in the feeding position and infect H vitripennis nymphs and adults The infected cadavers are covered by the fungal mycelia that sporulate in cycles and provide a means to infect uninfected feeding H vitripennis (Lietze et al 2010) H homalodiscae-infected H vitripennis attach tenaciously via rhizoids to the plant bark in the summer and are often found attached to this substrate the following spring This positive visual response may increase transmission of the fungus to H vitripennis
When live presumably acoustically active H vitripennis were added to treatment branches the numbers of H vitripennis increased by 53 on the treated branches (Table 1) H vitripennis unlike most insect herbivores has not been shown to use olfaction at any level for courtship or mating In contrast H vitripennis and many Cicadel-lidae and related insect families use acoustical vibrations propagated through the host plant to communicate during courtship and mating and for other functions (Cocroft 2006) This behavioral modality of
H vitripennis however has received only cursory attention (Percy et al 2008) and its potential effect was eliminated in all except this one experiment through the use of cadavers
With respect to host finding the role of olfaction remains un-clear Patt and Setamou (2007) reported that the response to visual cues by H vitripennis nymphs and adults was enhanced by exposure to plant volatiles Patt and Setamou (2010) demonstrated that H vitripennis nymphs could be conditioned to respond to vanilla as a novel host volatile and as a result they argued that associative learning of chemical profiles could enhance leafhopper location and recognition of host plants These authors also speculated on a potential mechanism use of plant volatile profile emissions (Ninemets et al 2002 2004) in host finding and recognition by adult H vitripennis While an interesting hypothesis much research remains to be done to support these ideas A strong response to conspecifics provides a complimentary or alternative mechanism that would eliminate the need for such highly evolved behaviors However one may hypothesize that the presence of plant volatiles could stimulate a more intense leafhopper visual search for conspe-cifics on the host Nevertheless the ready acceptance of the novel and poor host L limii in the presence of conspecifics argues against the need for leafhoppers to learn a visual or olfactory pattern for specific plant species
Trivial movement within and between host plants and host plant patches is germane to understanding the distribution and fitness of H vitripennis (Mizell et al 2008) H vitripennis are strongly visually oriented while feeding on a host plant they can detect movement from 5 to 10 m away Based on the ostensibly narrow sensory ca-pabilities of H vitripennis (lack of pheromones apparently limited use of olfaction) it is not unexpected that H vitripennis would have evolved visual behavior with multiple functions Moreover this simple capability would obviate the need for higher level behaviors such as associative learning at least for adults (Patt and Setamou 2010) Profiles of xylem chemistry can change as much as 10-fold in one day or from day to day in the absence of any overt visual signals to leafhoppers (Andersen et al1992 Brodbeck et al 2010) Because host plant nutrient quality varies unrecognizably to leafhoppers in time and space and between and within a plant species developing a search image to visually identify specific host plants or character-istics related to their quality appears to be extremely difficult and perhaps of little value Moreover host plants usually do not occur in monocultures a situation that would reinforce short-term pattern recognition By contrast delay in finding hosts within a patch can affect information retention (McLinn and Stephens 2006) Relying too heavily on recent experience can be costly especially if the environment and in this case host plant nutrients change rapidly (Mizell and French 1987 Andersen et al 1989 1992 1993 1995ab Brodbeck et al 1990 19951999 McLinn and Stephens 2006) Animals devalue unreliable signals Models predict signal following when the signal reliability exceeds environmental certaintymdashso that information is valuable Signal value however is not based solely on reliability it also depends on the statistical distribution of the environmental states (McLinn and Stephens 2006) Plant quality and predation risks are in constant flux in the case of H vitripennis and extremely variable in time and space H vitripennis handling of bottom-up and top-down forces is tied closely together because the potential payoff and consequences from both accrue on the host plant following a single decision to land H vitripennis appar-ently can make a decision to land based solely on a visual response
113American Entomologist bull Volume 58 Number 2
to conspecifics on host plant branches An arrestment response is then likely reinforced on the plant via acoustical communication as gustation behavior is initiated
Thus presence of conspecifics indicates a ldquosafety zonerdquo for host plant testing and likely provides high-quality (truthfulness of signal) information because all H vitripennis in any one location are under more or less the same dietary constraints Notwithstanding that O nigricans may have somewhat different nutritional requirements it may be important in the mitigation of bottom-up and top-down forces The presence of one or (usually) multiple conspecifics is likely a signal indicating a reasonably high threshold quality of nutrients or else the duration of conspecifics remaining on a host plant would be short This perhaps explains why three conspecifics analogous to the quorum decision of Ward et al (2008) maximize the response Three may be enough to compensate for any variations in individual hunger rate or other uninterpretable behaviors (females not feeding or ovipositing males that feed less) that may be ongoing and could affect the ldquotruth in signalrdquo presented by conspecifics
Arthropod populations face a variety of environmental risks to which they must adapt that can be described within the bottom-up and top-down heuristic model however ldquoNo creature can do all thingsrdquo and genetic and other constraints are many Thus tradeoffs are required for organisms to adapt and sustain over evolutionary time (Rolf 1992) Activity or movement is highly related to resource acquisition but it is also related to mortality risks a cost and as a result mediates one of the most general tradeoffs faced by organ-isms (Werner 1992) Information gathering also comes at a cost because of the loss of time potentially spent on other activities (Eliassen et al 2007) Moreover where resources vary temporally or for example predation risk is highly unpredictable the value of information decreases with time such that a forager should place greater value on the newest available information (Eliassen et al 2007) An individualrsquos response to conspecifics makes it possible to collect a broad range of real-time biological data that may reduce the tradeoffs and costs imposed by required activity Foraging suc-cess and predator avoidance behaviors directly related to fitness are strongly affected as a result (Lazarus 1979 Foster and Treherne 1981 Lawrence 1985 Stamps 1987 Turchin 1987) The environment of xylophagous leafhoppers is highly stochastic in time and space and their extreme polyphagy requires exposure to a wide array of plant structures changing chemical profiles and associated predators Response to conspecifics offers a simple parsimonious mechanism to eliminate the need to develop sophisticated physiological or behavioral mechanisms to handle this array of visual and chemical cues that accompany the extreme uncertainty presented by resource finding and predator avoidance
In conclusion H vitripennis exhibits at least two major types of visually based behaviors While in the normal feeding position (Bex-tine et al 2005) they can detect movement from asymp10 m distance and move behind the branch to hide themselves from the approaching object This response is often done synchronously by the leafhoppers present More importantly when searching for and landing on a host plant where the risk of predation may be high they can respond to cues presented by conspecifics These consist of a narrowly defined stimulus roughly equal in size to the H vitripennis body but not gt12 mm To our knowledge this is the first report of an arthropod exhibit-ing behavioral parsimony in which one signal a visual cue (response to conspecifics) is used to mitigate immediate risks from bottom-up and top-down forces In the present case leafhoppers exhibit a ldquofixedrdquo
response response to conspecifics to bottom-up and top-down forces independent of their value in the environmental context Avail-ability and spatial distribution of host plants and their nutritional quality are critical to the survival and fitness of H vitripennis The availability of xylem fluid within and between host plants changes continuously and rapidly over time and space requiring much trivial movement and emigration by leafhoppers These natural fluctuations in plant xylem chemistry are the primary basis of the bottom-up and top-down risks to xylophagous leafhoppers because they determine the movement and seasonal use of host plants by H vitripennis (be-havior and performance) (Mizell and French 1987 Andersen et al 1989 1992 1995ab 2005 2008 Brodbeck et al 1990 19951999) Previously several novel physiological and behavioral adaptations critical to leafhopper survival were documented (Mizell et al 2008) Response to conspecifics during host plant searching movements is another novel but fundamentally important behavior exhibited by these interesting disease vectors that allow them to survive and endure as xylophages
AcknowledgmentsThe research was partially funded by a grant to R F Mizell III and P
C Andersen from the California Department of Food and Agriculture Piercersquos DiseasemdashGlassy-winged Sharpshooter Research Program We thank Gary W Knox for use of the experimental plot containing the Lagerstroemia limii We thank Tobin Northfield Joseph Patt and Arash Rashad for helpful comments on a previous draft of the manuscript
References CitedAndersen P C B V Brodbeck and R M Mizell III 1989 Metabolism
of amino acids organic acids and sugars extracted from the xylem fluid of four host plants by Homalodisca coagulata Entomol Exp Appl 50 149ndash159
Andersen P C B V Brodbeck and R M Mizell III 1992 Feeding by the leafhopper Homalodisca coagulata in relation to xylem fluid chemistry and tension J Insect Physiol 38 611ndash622
Andersen P C B V Brodbeck and R M Mizell III 1993 Diurnal variations in the chemistry of xylem fluid from Lagerstroemia indica an endogenous circadian rhythm Physiol Plant 89 783-790
Andersen P C B V Brodbeck and R M Mizell III 1995a Diurnal variation in tension osmolarity and the composition of nitrogen and carbon assimilates in xylem fluid of Prunus persica Vitis hybrid and Prunus communis J Am Hortic Sci 120 600ndash604
Andersen P C B V Brodbeck and R M Mizell III 1995b Water stress- and nutrient-mediated changes in the water relations and the amino acids organic acids and sugars in xylem fluid of Prunus salicina and Lagerstroemia indica J Am Soc Hortic Sci 120 36ndash42
Andersen P C B V Brodbeck and R M Mizell III 2003 Plant and insect characteristics in response to increasing density of Homalodisca coagulata on three host species a quantification of assimilate extraction Entomol Exp Appl 107 57ndash68
Andersen P C B V Brodbeck R M Mizell III and S Oden 2005 The abundance and feeding of Homalodisca coagulata (Homoptera Auche-norrhyncha Cicadellidae) on Vitis genotypes in north Florida Environ Entomol 34 466ndash478
Andersen PC R M Mizell III B V Brodbeck T Beckman and G Krewer 2008 Abundance and consumption rates of glassy-winged sharpshooter (Hemiptera Cicadellidae) on peaches and plums J En-tomol Sci 43 394ndash407
Andersen P C B V Brodbeck and R M Mizell III 2009 Assimilation efficiency of free and protein amino acids by the glassy-winged sharp-shooter Homalodisca vitripennis (Germar) (Hemiptera Cicadellidae Cicadellinae) feeding on Citrus sinensis and Vitis vinifera Fla Entomol 92 116ndash122
Backus E J Habibi F Yan and M Ellersieck 2005 Stylet penetration by adult Homalodisca coagulata on grape electrical penetration graph
114 American Entomologist bull Summer 2012
waveform characterization tissue correlation and possible implica-tions for transmission of Xylella fastidiosa Ann Entomol Soc Am 98 787ndash813
Baker P S 1979 Flying locust visual responses in a radial wind tunnel J Comp Physiol 131 39ndash47
Bextine B R B C Jackson D B Harshman and Miller 2005 Homalo-disca coagulata (Hemiptera Cicadellidae) feeding posture Ann Ento-mol Soc Am 98 814ndash819
Boucias D G D W Scharf S E Breaux S D Purcell and R M Mizell III 2006 Studies on the fungi associated with the glassy-winged sharpshooter Homalodisca coagulata with emphasis on a new species Hirsutella homalodiscae nom prov BioControl 52 231ndash258
Brodbeck BV R M Mizell III and P C Andersen 1990 Amino acids as determinants of host preference for the xylemfeeding leafhopper Homalodisca coagulata Oecologia 83 338ndash345
Brodbeck B V P C Andersen and R M Mizell III 1993 Physiological and behavioral adaptations of three species of leafhoppers in response to the dilute nutrient content of xylem fluid J Insect Physiol 39 73ndash81
Brodbeck B V P C Andersen and R M Mizell III 1995 Differential utilization of nutrients during development by the xylophagous leafhop-per Homalodisca coagulata Entomol Exp Appl 75 279ndash289
Brodbeck BV P C Andersen and R M Mizell III 1999 The effects of total dietary nitrogen and amino acid balance on the development of xylophagous leafhoppers Arch Insect Biochem Physiol 42 37ndash50
Brodbeck B V P C Andersen and R M Mizell III 2007 Preference-performance linkage of the xylem feeding leafhopper Homalodisca vit-ripennis (Hemiptera Cicadellidae) Environ Entomol 36 1512ndash1522
Brodbeck B V P C Andersen and R M Mizell III 2011 Nutrient media-tion of behavioral plasticity and resource allocation in a xylem-feeding leafhopper Oecol 165 111-122
Cocroft R B H J Shugart K T Konrad and K Tibbs 2006 Variation in plant substrates and its consequences for insect vibrational com-munication Ethology 112 779ndash789
Collett T S and M F Land 1978 How hoverflies compute interception courses J Comp Physiol 125 191ndash204
Coolen I O Dangles and J Casas 2005 Social learning in non-colonial insects Curr Biol 15 1931-1935
Daane K M Johnson T Ruiz and J M Hashim 2005 Host plant influence on glassy-winged sharpshooter and its natural enemies pp 100ndash104 In Proceedings 2005 Piercersquos Disease Research Symposium California Department of Food and Agriculture Sacramento
Dukas R 2001 Effects of perceived danger on flower choice by bees Ecol Lett 4 327ndash333
Dukas R and D H Morse 2003 Crab spiders affect flower visitation by bees Oikos 101 157ndash163
Eliassen S E C Jargensen M Mangel and J Garl 2007 Exploration or exploitation life expectancy changes the value of learning in foraging strategies Oikos 116 513ndash523
Fernandez-Juricic E B Kerr P Bednekoff and D Stephens 2004 When are two heads better than one Visual perception and informa-tion transfer affect vigilance coordination in foraging groups Behav Ecol 6 898ndash906
Foster W A and J E Treherne 1981 Evidence for the dilution effect in the selfish herd from fish predation on a marine insect Nature 293 466ndash467
Fournier V J Hagler K Daane J de Leoacuten and R Groves 2008 Iden-tifying the predator complex of Homalodisca vitripennis (Hemiptera Cicadellidae) a comparative study of the efficacy of an ELISA and PCR gut content bioassay Oecologia 157 629ndash640
Giraldeau L T J Valone and J J Templeton 2002 Potential disadvan-tages of using socially acquired information Phil Trans R Soc Lond B 357 1559ndash1566
Hamiliton WD 1971 Geometry for the selfish herd J Theror Biol 31295-311
Hairston N G J E Smith and L B Slobodkin 1960 Community struc-ture population control and competition Am Midl Nat 106 249ndash257
Hunter W B and D E Ullman 1994 Precibarial and cibarial chemosen-sillae in the western flower thrips Frankliniella occidentalis (Pergande)(Thysanoptera Thripidae) Int J Insect Morphol Embryol 23 69ndash83
Hunter W B E Hiebert S Webb J E Polston and J H Tsai 1996 Precibarial and cibarial chemosensilla in the whitefly Bemisia tabaci
(Gennadius) (Homoptera Aleyrodidae) Int J Insect Morphol Embryol 25 295ndash304
Jeffries M J and J H Lawton 1984 Enemy-free space and the structure of ecological communities Biol J Linn Soc 23 269-286
Karban R and L T Baldwin 1997 Induced response to herbivory University of Chicago Press Chicago
Land M F and T S Collett 1974 Chasing behavior of houseflies (Fannia canicularis) J Comp Physiol 80 331ndash357
Lawrence E S 1985 Vigilance during ldquoeasyrdquo and ldquodifficultrdquo foraging tasks Anim Behav 33 1373ndash1375
Lawton J H and S McNeil1979 Between the devil and the deep blue sea on the problem of being a herbivore pp 223ndash244 In R M Anderson B D Turner and L R Taylor [Eds] Population dynamics Blackwell Oxford England Blackwell Sci Publ 434 pp
Lazarus J 1979 Flock size and behavior in captive red-billed weaverbirds (Quelea quelea) implications for social facilitation and the functions of flocking Behavior 71 1ndash2
Lietze V R M Mizell III and D G Boucias 2011 Transmission of the mycopathogen Hirsutella spp to nymphs and adults of the glassy-winged sharpshooters Homalodisca vitripennis (=coagulata) in the greenhouse Fla Entomol 94 106-108
Lopez R F R Mizell III P C Andersen and B V Brodbeck 2003 Impact of natural enemies on populations of the glassy-winged sharpshooter Homalodisca coagulata (Say) in north Florida In R VanDriesche [Ed] First international symposium on biological control of arthropods US Dep Agric For Ser Pub FHTET-03-05 484ndash486
Mainman G A D Straw and M H Dickson 2008 A simple vision-based algorithm for decision making in flying Drosophila Curr Biol 18 464ndash470
McLinn C and D W Stephens 2006 What makes information valu-able signal reliability and environmental uncertainty Anim Behav 71 1119ndash1129
Mizell R F III and W J French 1987 Leafhopper vectors of phony peach disease feeding site preference and survival on infected and uninfected peach and seasonal response to selected host plants J Entomol Sci 22 1122
Mizell R F III P C Andersen P C Phillips and P Oevering 2005 Population dynamics and interactions between the glassy-winged sharpshooter and the host plants in response to California phenology pp 117ndash121 In Proceedings 2005 Piercersquos Disease Research Symposium California Department of Food and Agriculture Sacramento
Mizell R F III C Tipping P C Andersen B V Brodbeck T Northfield and W Hunter W 2008 Behavioral model for the glassy-winged sharp-shooter Homalodisca vitripennis (Hemiptera Cicadellidae) optimiza-tion of host plant utilization and management implications Environ Entomol 37 1049ndash1062
Mizell R F III PC Andersen and B V Brodbeck 2009 Improved detec-tion monitoring and management of the glassy-winged sharpshooter pp 40ndash43 In Proceedings 2009 Piercersquos Disease Research Symposium California Department of Food and Agriculture Sacramento
Muller KL 1998 The role of conspecifics in habitat settlement in a ter-ritorial grasshopper Anim Behav 56 479ndash485
Ninemets Uuml M Riechstein M Staudt G Siefert and J Tenhunen 2002 Stomatal constraints may affect emission of oxygenated monoterpenoids from foliage of Pinus pinea Plant Physiol 130 1371ndash1385
Ninemets Uuml F Loreto and M Riechstein 2004 Physiological and physiochemical controls on foliar volatile organic compound emissions Trends Plant Sci 9 180ndash186
Northfield T R F Mizell III P C Andersen B V Brodbeck T C Riddle and W B Hunter 2009 Dispersal patch leaving and aggregation of the glassy-winged sharpshooter Homalodisca vitripennis (Hemiptera Cicadellidae) Environ Entomol 38 183ndash191
Patt J and M Setamou 2007 Olfactory and visual stimuli affecting host plant detection in Homalodisca coagulata (Hemiptera Cicadellidae) Environ Entomol 36 142ndash150
Patt J and M Setamou 2010 Recognition of novel volatile cues by the nymph of the glassy-winged sharpshooter Homalodisca vitripennis (Cicadellidae) J Insect Behav DOI 101007s10905-010-9214-z 23 290-302
Percy D M E A Boyd and M S Hoddle 2008 Observations of acoustical signaling in three sharpshooters Homalodisca vitripennis Homalodisca
115American Entomologist bull Volume 58 Number 2
liturata and Graphocephala atropunctata (Hemiptera Cicadellidae) Ann Entomol Soc 101 253ndash259
Power M E 1992 Top-down and bottom-up forces in food webs do plants have primacy Ecologia 73 733ndash746
Price P W C E Bouton P Gross B A McPheron J N Thompson and A E Wise 1980 Interactions among three trophic levels influence of plants on interactions between insect herbivores and natural enemies Annu Rev Ecol Syst 11 41ndash66
Redak R M J Blua K L Lopes A H Purcell R F Mizell III and P C Andersen 2004 The biology and control of xylem sap-feeding insect vectors of Xylella fastidiosa and their relation to disease epidemiology Annu Rev Entomol 49 243ndash270
Rolf D A 1992 The evolution of life history traits Chapman amp Hall NYSandlin E A 2000 Cue use affects resource subdivision among three
coexisting hummingbird species Behav Ecol 11 550-559SAS Institute 2009 Proc GLM SAS 93 Statistical Analysis System Cary NCShrewsbury P and M Raupp 2008 Do top-down or bottom-up forces
determine Stephanitis pyroides abundance in urban landscapes Ecol Appl 16 262ndash273
Systat Software 2009 SigmaPlot for Windows 110 Systat Software Bangalore India
Stamps J A 1987 Conspecifics as cues to territory quality a preference of juvenile lizards (Anolis aeneus) for previously used territories Am Nat 129 629ndash642
Stamps J A 1988 Conspecific attraction and aggregation in territorial species Am Nat 131 330ndash347
Thompson J D S C Peterson and L D Harder 1987 Response to traplining bumble bees to competition experiments shifts in feeding location and efficiency Oecologia 71 295ndash300
Tipping C R F Mizell III and P C Andersen 2004 Adaptations impor-tant for the dispersal of immature stages of three species of leafhopper (Homoptera Cicadellidae) Fla Entomol 87 372ndash379
Triapitsyn S V R F Mizell III J L Bossert and J E Carlton 1998 Egg parasitoids of glassy-winged sharpshooter Homalodisca coagulata (Homoptera Cicadellidae) Fla Entomol 81 243ndash251
Turner W F and H N Pollard 1959 Life histories and behavior of five insect vectors of phony peach disease Tech Bull US Dep Agric 1188
Turchin P 1987 The role of aggregation in the response of Mexican bean beetles to host-plant density Oecologia 71 577ndash582
Valone T J 1989 Group foraging public information and patch simula-tion Oikos 56 357ndash363
Walker M and T H Jones 2001 Relative roles of top-down and bottom-up forces in terrestrial tritrophic plant-insect herbivore-natural enemy systems Oikos 93 177ndash187
Ward A J W D J T Sumpter I D Couzin P J B Hart and J Krause 2008 Quorum decision-making facilitates information transfer in fish shoals PNAS 105 6948ndash6953
Wells J M B C Raju H Y Hung W G Weisburg L Mandelco-Paul and D J Brenner 1987 Xylella fastidiosa gen nov sp nov gram-negative xylem-limited fastidious plant bacteria related to Xanthomonas spp Int J Syst Bacteriol 37 136ndash143
Werner E E 1992 Individual behavior and higher-order species interac-tions Am Nat (suppl) 140 S5ndashS32
White T R C 1978 The importance of relative shortage of food in animal ecology Oecologia 33 71ndash86
Williams G C 1966 Adaptation and natural selection Princeton University Press Princeton NJ
Wu D S C Daugherty S E Van Aken G H Pai K L Watkins H Khouri L J Tallon J M Zaborsky H E Dunbar P L Tran N A Moran J A Eisen 2006 Metabolic complementarity and genomics of the dual symbiosis of sharpshooters PloS-Biology 4 e188
Russell F Mizell III Is a Professor of Entomology (rfmizellufledu) Peter C Andersen is a Professor of Horticulture (peaufledu) and Brent V Brod-beck (bvbufledu) is a Biological Scientist all located at the University of Floridarsquos North Florida Research and Education Center-Quincy 155 Research Rd Quincy Fl 32351 Wayne B Hunter (whunterushrlarsusdagov) is a research entomologist with the USDA Agricultural Research Service at the US Horticultural Research Lab 2001 South Rock Rd Ft Pierce FL 34945 For over 20 years they have collaborated in research on the biology ecology and behavior of the leafhopper vectors of Xylella fastidiosa
With our high qualityLepidoptera Eggs and LarvaeBeet Army Worm bull Cabbage LooperCorn Earworm bull Diamondback MothFall Armyworm bull Tobacco Budworm
Save Money Save Time Get Results
Lepidoptera Eggs and LarvaeInsect Diets and Ingredients
Rearing Supplies and Equipment
Entomology Division
800-996-9908 US and Canada908-996-2155 Internationalsalesinsectrearingcom
wwwinsectrearingcom

111American Entomologist bull Volume 58 Number 2
maximized the behavior The test cadavers were placed asymp8ndash10 cm apart with the total three-cadaver presentation providing a visual cue le20 cm wide The lack of increase by additional cadavers may indicate that leafhopper visual breadth is lt30 cm H vitripennis also responded to cadavers of the similarly sized heterospecific O nigricans that is often found feeding on the same host plants (ie Ilex spp) in bi-species groups The congeneric species H insolita which is much smaller and predominantly a grass feeder however did not elicit a positive response Although H vitripennis do feed on some grass and herbaceous species concurrently with H insolita we have never observed the two species feeding in heterospecific groups or on the same part of a plant Brodbeck et al (1993) reported that the dietary needs for both species were similar although they prefer to feed on different plant tissues (Turner and Pollard 1959) Percy et al (2008) compared the acoustical signals of H vitripennis and the congener H liturata Ball and reported that the signals were different in most parameters measured Some overlap however was observed such that it cannot be ruled out that acoustical signals produced by one Homalodisca sp could be detected by a related species If there is some signal overlap between the two species then it is possible that H vitripennis could derive benefit from detecting signals produced by H insolita acoustically and visually from group defensive movements that are similar to those of H vitripennis
In an attempt to define the physical parameters of the visual cues perceived and used by H vitripennis a group of models of different sizes and orientations on the host plant were tested A black model of 3times12 mm roughly equal to the body dimensions of an adult H vitripennis evoked significant visual attraction as did a 10 times10 mm
black felt model All other larger models in any plane did not evoke significant response when placed either vertically or horizontally on the plant No response was observed to a black model affixed to a contrasting white square card These results along with the lack of response to predators and the smaller congener H insolita suggest that the visual cues used by H vitripennis when in flight are relatively narrowly defined to the size close to its own body dimensions (width le10 times length le12 mm) The heterospecific O nigricans falls within this size range The failure of H vitripennis to respond to the model on the white card along with the orientation limits may indicate that detection of the visual cues from congeners is contextually related in some manner to that provided as background by the branch or its silhouette in space
Brodbeck et al (1990) and Andersen et al (2008) showed that H vitripennis abundance on a plant is highly correlated with leafhopper feeding rate and is indicative of the underlying xylem nutritional qual-ity H vitripennis are normally highly aggregated in the field among and within available host species (especially on higher quality host plants) although there is never a time when 100 of the population is observed on one host species As a rule in north Florida only a low percentage of branches on fair-to-good hosts are infested and they usually contain more than one leafhopper per branch (Mizell and French 1987) Significant H vitripennis response to cadavers when they were placed on the novel and poor host plant L limii provides very strong support for the use of conspecifics as visual cues by H vitripennis in flight to locate potential host plants for bottom-up risk mitigation Furthermore H vitripennis will respond visually to conspecifics independent of host plant quality and familiarity with a host plant species This result supports the idea that H vitripennis do not assess host plant quality before landing and furthermore that this pivotal decision is likely made using gustation at the level of the
xylem tissue (Hunter and Ullman 1994 Hunter et al1996 Backus et al 2005) The role of plant volatiles remains to be fully described but the research of Patt and Setamou (2007 2010) suggests a potential role in H vitripennis behavior
The importance of bottom-up nutrient profiles on fitness of these leafhoppers is well described (Redak et al 2004) Leafhopper ag-gregation however may be of further benefit by inducing chemical changes in host plant physiology that affect fitness such as feeding rate and fecundity (Karban and Baldwin 1997) Andersen et al (2003 2009) addressed this possibility in experiments with five host species They found that moderate-to-large plants of crape myrtle L indica L and plum Prunus salicina Lindl exhibited no increasing effects on nutrient compounds of leafhopper feeding in large numbers nor did leafhopper feeding behavior change when in groups In small soybean plants Glycine max (L) some nutrients were highly reduced (Andersen et al 2003) In citrus H vitripennis feeding increased seven amino acids in protein form but little change in protein amino acids was observed in Vitis vinifera lsquoChardonnayrsquo In citrus and lsquoChardonnayrsquo grape amino acid increased assimilation efficiency as a result of leafhopper feeding These data suggest that H vitripennis feeding does not induce metabolically inhibitory profiles in the xylem (Andersen et al 2009) This along with the induction of proteins and a slight decrease in free amino acids indicates that there may be some induced benefit in nutrition from H vitripennis aggregation (Andersen et al 2009)
Predation is an important mortality risk to H vitripennis in flight and on the host plant during feeding and oviposition (Lopez et al 2003 Daane et al 2005 Suttle and Hoddle 2006 Fournier et al 2008 Mizell et al 2008) Spiders and lizards are the most impor-tant predators of H vitripennis on the plants (Lopez et al 2003 Fournier et al 2008) Top-down risk mitigation by H vitripennis was assessed indirectly by testing visual response by H vitripennis to lizard predators and simulated predator models (fishing lures) that closely resembled living predators The experimental context most closely approximates the risk to H vitripennis during landing on a new branch the time when little or no information on predator presence on the selected branch is available to arriving H vitripennis This is also the time when H vitripennis presents the largest moving visual stimulus to hidden predators Once on the plant landed H vitripennis stop moving and provide a different visual stimulus to would-be predators they also regain the use of positional camouflage and their own visually based longer-range avoidance defense with the protection it provides As described earlier this protection is partially accrued from conspecifics via the ldquodilutionrdquo effect (Foster and Treherne 1981) Similarly heterospecifics present would also contribute to defense
The only other time that H vitripennis move excessively on the host plant is when the males seek mates and the females oviposit (Mizell et al 2008) Although copulation occurs mostly during the scotophase one might hypothesize that the presence of conspecif-ics would also reduce the risk from predation to more vulnerable copulating pairs and ovipositing females This may also be a selec-tion factor that induces H vitripennis females to oviposit on plants upon which they feed but which do not meet the nutritional needs of their nymphs (Brodbeck et al 2007 Mizell et al 2008) The ability to recognize conspecifics as indicators of suitable host plants allows H vitripennis to feed without moving and eliminates search flights for hours or days proportionally reducing in-flight and landing preda-tion risks These behaviors underscore the potential risks involved in
112 American Entomologist bull Summer 2012
finding hosts the tradeoffs faced by female leafhoppers and the value of conspecific recognition in their mitigation An unusual number of mated leafhopper pairs also were observed among the cadaver treatment branches on the mornings of succeeding test days These observations provide further support for the contention concerning top down risk mitigation via conspecific response
Significant response to avoid predators and artificial models was not observed It is unlikely that an arthropod herbivore would be able to develop a search image with such elasticity that it could encompass the wide range of natural enemies potentially encoun-tered The immediate acceptance of the novel and poor host L limii provides further evidence against the occurrence of a learned host image by searching leafhoppers Response to conspecifics would obviate the necessity of developing a predator search image because conspecifics would usually be present only where predators were absent Thus a type of ldquoenemy-free spacerdquo (Jeffries and Lawton 1984) may be represented by conspecific presence Alternatively relative to the risk of landing on a branch without conspecifics even if the conspecific signal was not entirely truthful about the level of risk the dilution effect from conspecific presence would provide increased protection theoretically in direct proportion to the number of con-specifics present Although foragers in many animal groups have an inherent geometry that causes information asymmetries individuals get more information from closer neighbors (Fernandez-Juricic et al 2004) H vitripennis uses keen visual acuity that likely is coupled with acoustical communication as a mechanism to respond rapidly en masse to approaching predators As a result leafhoppers accrue a more equivalent gain from defensive movement coordination (Fernandez-Juricic et al 2004)
On the host plant H vitripennis can detect and respond (and routinely do so in concert perhaps mediated by acoustic commu-nication) to movement of large objects approaching them from off the plant and to smaller objects on the plant Live lizards do elicit a response from feeding H vitripennis as they move over the host plant and approach leafhoppers (Lopez et al 2003) This suggests the occurrence in H vitripennis of a general visual capability (differ-ent from the visual response to conspecifics) to recognize moving objects that would circumvent the necessity of developing an array of species-specific predator search images
H vitripennis responded significantly to cadavers painted gray that served as a model of H vitripennis infected with the parasitic fungal pathogen H homalodiscae (Boucias et al 2006) H vitripennis cadavers infected with this pathogen remain attached for months on the host plant at feeding sites in the feeding position and infect H vitripennis nymphs and adults The infected cadavers are covered by the fungal mycelia that sporulate in cycles and provide a means to infect uninfected feeding H vitripennis (Lietze et al 2010) H homalodiscae-infected H vitripennis attach tenaciously via rhizoids to the plant bark in the summer and are often found attached to this substrate the following spring This positive visual response may increase transmission of the fungus to H vitripennis
When live presumably acoustically active H vitripennis were added to treatment branches the numbers of H vitripennis increased by 53 on the treated branches (Table 1) H vitripennis unlike most insect herbivores has not been shown to use olfaction at any level for courtship or mating In contrast H vitripennis and many Cicadel-lidae and related insect families use acoustical vibrations propagated through the host plant to communicate during courtship and mating and for other functions (Cocroft 2006) This behavioral modality of
H vitripennis however has received only cursory attention (Percy et al 2008) and its potential effect was eliminated in all except this one experiment through the use of cadavers
With respect to host finding the role of olfaction remains un-clear Patt and Setamou (2007) reported that the response to visual cues by H vitripennis nymphs and adults was enhanced by exposure to plant volatiles Patt and Setamou (2010) demonstrated that H vitripennis nymphs could be conditioned to respond to vanilla as a novel host volatile and as a result they argued that associative learning of chemical profiles could enhance leafhopper location and recognition of host plants These authors also speculated on a potential mechanism use of plant volatile profile emissions (Ninemets et al 2002 2004) in host finding and recognition by adult H vitripennis While an interesting hypothesis much research remains to be done to support these ideas A strong response to conspecifics provides a complimentary or alternative mechanism that would eliminate the need for such highly evolved behaviors However one may hypothesize that the presence of plant volatiles could stimulate a more intense leafhopper visual search for conspe-cifics on the host Nevertheless the ready acceptance of the novel and poor host L limii in the presence of conspecifics argues against the need for leafhoppers to learn a visual or olfactory pattern for specific plant species
Trivial movement within and between host plants and host plant patches is germane to understanding the distribution and fitness of H vitripennis (Mizell et al 2008) H vitripennis are strongly visually oriented while feeding on a host plant they can detect movement from 5 to 10 m away Based on the ostensibly narrow sensory ca-pabilities of H vitripennis (lack of pheromones apparently limited use of olfaction) it is not unexpected that H vitripennis would have evolved visual behavior with multiple functions Moreover this simple capability would obviate the need for higher level behaviors such as associative learning at least for adults (Patt and Setamou 2010) Profiles of xylem chemistry can change as much as 10-fold in one day or from day to day in the absence of any overt visual signals to leafhoppers (Andersen et al1992 Brodbeck et al 2010) Because host plant nutrient quality varies unrecognizably to leafhoppers in time and space and between and within a plant species developing a search image to visually identify specific host plants or character-istics related to their quality appears to be extremely difficult and perhaps of little value Moreover host plants usually do not occur in monocultures a situation that would reinforce short-term pattern recognition By contrast delay in finding hosts within a patch can affect information retention (McLinn and Stephens 2006) Relying too heavily on recent experience can be costly especially if the environment and in this case host plant nutrients change rapidly (Mizell and French 1987 Andersen et al 1989 1992 1993 1995ab Brodbeck et al 1990 19951999 McLinn and Stephens 2006) Animals devalue unreliable signals Models predict signal following when the signal reliability exceeds environmental certaintymdashso that information is valuable Signal value however is not based solely on reliability it also depends on the statistical distribution of the environmental states (McLinn and Stephens 2006) Plant quality and predation risks are in constant flux in the case of H vitripennis and extremely variable in time and space H vitripennis handling of bottom-up and top-down forces is tied closely together because the potential payoff and consequences from both accrue on the host plant following a single decision to land H vitripennis appar-ently can make a decision to land based solely on a visual response
113American Entomologist bull Volume 58 Number 2
to conspecifics on host plant branches An arrestment response is then likely reinforced on the plant via acoustical communication as gustation behavior is initiated
Thus presence of conspecifics indicates a ldquosafety zonerdquo for host plant testing and likely provides high-quality (truthfulness of signal) information because all H vitripennis in any one location are under more or less the same dietary constraints Notwithstanding that O nigricans may have somewhat different nutritional requirements it may be important in the mitigation of bottom-up and top-down forces The presence of one or (usually) multiple conspecifics is likely a signal indicating a reasonably high threshold quality of nutrients or else the duration of conspecifics remaining on a host plant would be short This perhaps explains why three conspecifics analogous to the quorum decision of Ward et al (2008) maximize the response Three may be enough to compensate for any variations in individual hunger rate or other uninterpretable behaviors (females not feeding or ovipositing males that feed less) that may be ongoing and could affect the ldquotruth in signalrdquo presented by conspecifics
Arthropod populations face a variety of environmental risks to which they must adapt that can be described within the bottom-up and top-down heuristic model however ldquoNo creature can do all thingsrdquo and genetic and other constraints are many Thus tradeoffs are required for organisms to adapt and sustain over evolutionary time (Rolf 1992) Activity or movement is highly related to resource acquisition but it is also related to mortality risks a cost and as a result mediates one of the most general tradeoffs faced by organ-isms (Werner 1992) Information gathering also comes at a cost because of the loss of time potentially spent on other activities (Eliassen et al 2007) Moreover where resources vary temporally or for example predation risk is highly unpredictable the value of information decreases with time such that a forager should place greater value on the newest available information (Eliassen et al 2007) An individualrsquos response to conspecifics makes it possible to collect a broad range of real-time biological data that may reduce the tradeoffs and costs imposed by required activity Foraging suc-cess and predator avoidance behaviors directly related to fitness are strongly affected as a result (Lazarus 1979 Foster and Treherne 1981 Lawrence 1985 Stamps 1987 Turchin 1987) The environment of xylophagous leafhoppers is highly stochastic in time and space and their extreme polyphagy requires exposure to a wide array of plant structures changing chemical profiles and associated predators Response to conspecifics offers a simple parsimonious mechanism to eliminate the need to develop sophisticated physiological or behavioral mechanisms to handle this array of visual and chemical cues that accompany the extreme uncertainty presented by resource finding and predator avoidance
In conclusion H vitripennis exhibits at least two major types of visually based behaviors While in the normal feeding position (Bex-tine et al 2005) they can detect movement from asymp10 m distance and move behind the branch to hide themselves from the approaching object This response is often done synchronously by the leafhoppers present More importantly when searching for and landing on a host plant where the risk of predation may be high they can respond to cues presented by conspecifics These consist of a narrowly defined stimulus roughly equal in size to the H vitripennis body but not gt12 mm To our knowledge this is the first report of an arthropod exhibit-ing behavioral parsimony in which one signal a visual cue (response to conspecifics) is used to mitigate immediate risks from bottom-up and top-down forces In the present case leafhoppers exhibit a ldquofixedrdquo
response response to conspecifics to bottom-up and top-down forces independent of their value in the environmental context Avail-ability and spatial distribution of host plants and their nutritional quality are critical to the survival and fitness of H vitripennis The availability of xylem fluid within and between host plants changes continuously and rapidly over time and space requiring much trivial movement and emigration by leafhoppers These natural fluctuations in plant xylem chemistry are the primary basis of the bottom-up and top-down risks to xylophagous leafhoppers because they determine the movement and seasonal use of host plants by H vitripennis (be-havior and performance) (Mizell and French 1987 Andersen et al 1989 1992 1995ab 2005 2008 Brodbeck et al 1990 19951999) Previously several novel physiological and behavioral adaptations critical to leafhopper survival were documented (Mizell et al 2008) Response to conspecifics during host plant searching movements is another novel but fundamentally important behavior exhibited by these interesting disease vectors that allow them to survive and endure as xylophages
AcknowledgmentsThe research was partially funded by a grant to R F Mizell III and P
C Andersen from the California Department of Food and Agriculture Piercersquos DiseasemdashGlassy-winged Sharpshooter Research Program We thank Gary W Knox for use of the experimental plot containing the Lagerstroemia limii We thank Tobin Northfield Joseph Patt and Arash Rashad for helpful comments on a previous draft of the manuscript
References CitedAndersen P C B V Brodbeck and R M Mizell III 1989 Metabolism
of amino acids organic acids and sugars extracted from the xylem fluid of four host plants by Homalodisca coagulata Entomol Exp Appl 50 149ndash159
Andersen P C B V Brodbeck and R M Mizell III 1992 Feeding by the leafhopper Homalodisca coagulata in relation to xylem fluid chemistry and tension J Insect Physiol 38 611ndash622
Andersen P C B V Brodbeck and R M Mizell III 1993 Diurnal variations in the chemistry of xylem fluid from Lagerstroemia indica an endogenous circadian rhythm Physiol Plant 89 783-790
Andersen P C B V Brodbeck and R M Mizell III 1995a Diurnal variation in tension osmolarity and the composition of nitrogen and carbon assimilates in xylem fluid of Prunus persica Vitis hybrid and Prunus communis J Am Hortic Sci 120 600ndash604
Andersen P C B V Brodbeck and R M Mizell III 1995b Water stress- and nutrient-mediated changes in the water relations and the amino acids organic acids and sugars in xylem fluid of Prunus salicina and Lagerstroemia indica J Am Soc Hortic Sci 120 36ndash42
Andersen P C B V Brodbeck and R M Mizell III 2003 Plant and insect characteristics in response to increasing density of Homalodisca coagulata on three host species a quantification of assimilate extraction Entomol Exp Appl 107 57ndash68
Andersen P C B V Brodbeck R M Mizell III and S Oden 2005 The abundance and feeding of Homalodisca coagulata (Homoptera Auche-norrhyncha Cicadellidae) on Vitis genotypes in north Florida Environ Entomol 34 466ndash478
Andersen PC R M Mizell III B V Brodbeck T Beckman and G Krewer 2008 Abundance and consumption rates of glassy-winged sharpshooter (Hemiptera Cicadellidae) on peaches and plums J En-tomol Sci 43 394ndash407
Andersen P C B V Brodbeck and R M Mizell III 2009 Assimilation efficiency of free and protein amino acids by the glassy-winged sharp-shooter Homalodisca vitripennis (Germar) (Hemiptera Cicadellidae Cicadellinae) feeding on Citrus sinensis and Vitis vinifera Fla Entomol 92 116ndash122
Backus E J Habibi F Yan and M Ellersieck 2005 Stylet penetration by adult Homalodisca coagulata on grape electrical penetration graph
114 American Entomologist bull Summer 2012
waveform characterization tissue correlation and possible implica-tions for transmission of Xylella fastidiosa Ann Entomol Soc Am 98 787ndash813
Baker P S 1979 Flying locust visual responses in a radial wind tunnel J Comp Physiol 131 39ndash47
Bextine B R B C Jackson D B Harshman and Miller 2005 Homalo-disca coagulata (Hemiptera Cicadellidae) feeding posture Ann Ento-mol Soc Am 98 814ndash819
Boucias D G D W Scharf S E Breaux S D Purcell and R M Mizell III 2006 Studies on the fungi associated with the glassy-winged sharpshooter Homalodisca coagulata with emphasis on a new species Hirsutella homalodiscae nom prov BioControl 52 231ndash258
Brodbeck BV R M Mizell III and P C Andersen 1990 Amino acids as determinants of host preference for the xylemfeeding leafhopper Homalodisca coagulata Oecologia 83 338ndash345
Brodbeck B V P C Andersen and R M Mizell III 1993 Physiological and behavioral adaptations of three species of leafhoppers in response to the dilute nutrient content of xylem fluid J Insect Physiol 39 73ndash81
Brodbeck B V P C Andersen and R M Mizell III 1995 Differential utilization of nutrients during development by the xylophagous leafhop-per Homalodisca coagulata Entomol Exp Appl 75 279ndash289
Brodbeck BV P C Andersen and R M Mizell III 1999 The effects of total dietary nitrogen and amino acid balance on the development of xylophagous leafhoppers Arch Insect Biochem Physiol 42 37ndash50
Brodbeck B V P C Andersen and R M Mizell III 2007 Preference-performance linkage of the xylem feeding leafhopper Homalodisca vit-ripennis (Hemiptera Cicadellidae) Environ Entomol 36 1512ndash1522
Brodbeck B V P C Andersen and R M Mizell III 2011 Nutrient media-tion of behavioral plasticity and resource allocation in a xylem-feeding leafhopper Oecol 165 111-122
Cocroft R B H J Shugart K T Konrad and K Tibbs 2006 Variation in plant substrates and its consequences for insect vibrational com-munication Ethology 112 779ndash789
Collett T S and M F Land 1978 How hoverflies compute interception courses J Comp Physiol 125 191ndash204
Coolen I O Dangles and J Casas 2005 Social learning in non-colonial insects Curr Biol 15 1931-1935
Daane K M Johnson T Ruiz and J M Hashim 2005 Host plant influence on glassy-winged sharpshooter and its natural enemies pp 100ndash104 In Proceedings 2005 Piercersquos Disease Research Symposium California Department of Food and Agriculture Sacramento
Dukas R 2001 Effects of perceived danger on flower choice by bees Ecol Lett 4 327ndash333
Dukas R and D H Morse 2003 Crab spiders affect flower visitation by bees Oikos 101 157ndash163
Eliassen S E C Jargensen M Mangel and J Garl 2007 Exploration or exploitation life expectancy changes the value of learning in foraging strategies Oikos 116 513ndash523
Fernandez-Juricic E B Kerr P Bednekoff and D Stephens 2004 When are two heads better than one Visual perception and informa-tion transfer affect vigilance coordination in foraging groups Behav Ecol 6 898ndash906
Foster W A and J E Treherne 1981 Evidence for the dilution effect in the selfish herd from fish predation on a marine insect Nature 293 466ndash467
Fournier V J Hagler K Daane J de Leoacuten and R Groves 2008 Iden-tifying the predator complex of Homalodisca vitripennis (Hemiptera Cicadellidae) a comparative study of the efficacy of an ELISA and PCR gut content bioassay Oecologia 157 629ndash640
Giraldeau L T J Valone and J J Templeton 2002 Potential disadvan-tages of using socially acquired information Phil Trans R Soc Lond B 357 1559ndash1566
Hamiliton WD 1971 Geometry for the selfish herd J Theror Biol 31295-311
Hairston N G J E Smith and L B Slobodkin 1960 Community struc-ture population control and competition Am Midl Nat 106 249ndash257
Hunter W B and D E Ullman 1994 Precibarial and cibarial chemosen-sillae in the western flower thrips Frankliniella occidentalis (Pergande)(Thysanoptera Thripidae) Int J Insect Morphol Embryol 23 69ndash83
Hunter W B E Hiebert S Webb J E Polston and J H Tsai 1996 Precibarial and cibarial chemosensilla in the whitefly Bemisia tabaci
(Gennadius) (Homoptera Aleyrodidae) Int J Insect Morphol Embryol 25 295ndash304
Jeffries M J and J H Lawton 1984 Enemy-free space and the structure of ecological communities Biol J Linn Soc 23 269-286
Karban R and L T Baldwin 1997 Induced response to herbivory University of Chicago Press Chicago
Land M F and T S Collett 1974 Chasing behavior of houseflies (Fannia canicularis) J Comp Physiol 80 331ndash357
Lawrence E S 1985 Vigilance during ldquoeasyrdquo and ldquodifficultrdquo foraging tasks Anim Behav 33 1373ndash1375
Lawton J H and S McNeil1979 Between the devil and the deep blue sea on the problem of being a herbivore pp 223ndash244 In R M Anderson B D Turner and L R Taylor [Eds] Population dynamics Blackwell Oxford England Blackwell Sci Publ 434 pp
Lazarus J 1979 Flock size and behavior in captive red-billed weaverbirds (Quelea quelea) implications for social facilitation and the functions of flocking Behavior 71 1ndash2
Lietze V R M Mizell III and D G Boucias 2011 Transmission of the mycopathogen Hirsutella spp to nymphs and adults of the glassy-winged sharpshooters Homalodisca vitripennis (=coagulata) in the greenhouse Fla Entomol 94 106-108
Lopez R F R Mizell III P C Andersen and B V Brodbeck 2003 Impact of natural enemies on populations of the glassy-winged sharpshooter Homalodisca coagulata (Say) in north Florida In R VanDriesche [Ed] First international symposium on biological control of arthropods US Dep Agric For Ser Pub FHTET-03-05 484ndash486
Mainman G A D Straw and M H Dickson 2008 A simple vision-based algorithm for decision making in flying Drosophila Curr Biol 18 464ndash470
McLinn C and D W Stephens 2006 What makes information valu-able signal reliability and environmental uncertainty Anim Behav 71 1119ndash1129
Mizell R F III and W J French 1987 Leafhopper vectors of phony peach disease feeding site preference and survival on infected and uninfected peach and seasonal response to selected host plants J Entomol Sci 22 1122
Mizell R F III P C Andersen P C Phillips and P Oevering 2005 Population dynamics and interactions between the glassy-winged sharpshooter and the host plants in response to California phenology pp 117ndash121 In Proceedings 2005 Piercersquos Disease Research Symposium California Department of Food and Agriculture Sacramento
Mizell R F III C Tipping P C Andersen B V Brodbeck T Northfield and W Hunter W 2008 Behavioral model for the glassy-winged sharp-shooter Homalodisca vitripennis (Hemiptera Cicadellidae) optimiza-tion of host plant utilization and management implications Environ Entomol 37 1049ndash1062
Mizell R F III PC Andersen and B V Brodbeck 2009 Improved detec-tion monitoring and management of the glassy-winged sharpshooter pp 40ndash43 In Proceedings 2009 Piercersquos Disease Research Symposium California Department of Food and Agriculture Sacramento
Muller KL 1998 The role of conspecifics in habitat settlement in a ter-ritorial grasshopper Anim Behav 56 479ndash485
Ninemets Uuml M Riechstein M Staudt G Siefert and J Tenhunen 2002 Stomatal constraints may affect emission of oxygenated monoterpenoids from foliage of Pinus pinea Plant Physiol 130 1371ndash1385
Ninemets Uuml F Loreto and M Riechstein 2004 Physiological and physiochemical controls on foliar volatile organic compound emissions Trends Plant Sci 9 180ndash186
Northfield T R F Mizell III P C Andersen B V Brodbeck T C Riddle and W B Hunter 2009 Dispersal patch leaving and aggregation of the glassy-winged sharpshooter Homalodisca vitripennis (Hemiptera Cicadellidae) Environ Entomol 38 183ndash191
Patt J and M Setamou 2007 Olfactory and visual stimuli affecting host plant detection in Homalodisca coagulata (Hemiptera Cicadellidae) Environ Entomol 36 142ndash150
Patt J and M Setamou 2010 Recognition of novel volatile cues by the nymph of the glassy-winged sharpshooter Homalodisca vitripennis (Cicadellidae) J Insect Behav DOI 101007s10905-010-9214-z 23 290-302
Percy D M E A Boyd and M S Hoddle 2008 Observations of acoustical signaling in three sharpshooters Homalodisca vitripennis Homalodisca
115American Entomologist bull Volume 58 Number 2
liturata and Graphocephala atropunctata (Hemiptera Cicadellidae) Ann Entomol Soc 101 253ndash259
Power M E 1992 Top-down and bottom-up forces in food webs do plants have primacy Ecologia 73 733ndash746
Price P W C E Bouton P Gross B A McPheron J N Thompson and A E Wise 1980 Interactions among three trophic levels influence of plants on interactions between insect herbivores and natural enemies Annu Rev Ecol Syst 11 41ndash66
Redak R M J Blua K L Lopes A H Purcell R F Mizell III and P C Andersen 2004 The biology and control of xylem sap-feeding insect vectors of Xylella fastidiosa and their relation to disease epidemiology Annu Rev Entomol 49 243ndash270
Rolf D A 1992 The evolution of life history traits Chapman amp Hall NYSandlin E A 2000 Cue use affects resource subdivision among three
coexisting hummingbird species Behav Ecol 11 550-559SAS Institute 2009 Proc GLM SAS 93 Statistical Analysis System Cary NCShrewsbury P and M Raupp 2008 Do top-down or bottom-up forces
determine Stephanitis pyroides abundance in urban landscapes Ecol Appl 16 262ndash273
Systat Software 2009 SigmaPlot for Windows 110 Systat Software Bangalore India
Stamps J A 1987 Conspecifics as cues to territory quality a preference of juvenile lizards (Anolis aeneus) for previously used territories Am Nat 129 629ndash642
Stamps J A 1988 Conspecific attraction and aggregation in territorial species Am Nat 131 330ndash347
Thompson J D S C Peterson and L D Harder 1987 Response to traplining bumble bees to competition experiments shifts in feeding location and efficiency Oecologia 71 295ndash300
Tipping C R F Mizell III and P C Andersen 2004 Adaptations impor-tant for the dispersal of immature stages of three species of leafhopper (Homoptera Cicadellidae) Fla Entomol 87 372ndash379
Triapitsyn S V R F Mizell III J L Bossert and J E Carlton 1998 Egg parasitoids of glassy-winged sharpshooter Homalodisca coagulata (Homoptera Cicadellidae) Fla Entomol 81 243ndash251
Turner W F and H N Pollard 1959 Life histories and behavior of five insect vectors of phony peach disease Tech Bull US Dep Agric 1188
Turchin P 1987 The role of aggregation in the response of Mexican bean beetles to host-plant density Oecologia 71 577ndash582
Valone T J 1989 Group foraging public information and patch simula-tion Oikos 56 357ndash363
Walker M and T H Jones 2001 Relative roles of top-down and bottom-up forces in terrestrial tritrophic plant-insect herbivore-natural enemy systems Oikos 93 177ndash187
Ward A J W D J T Sumpter I D Couzin P J B Hart and J Krause 2008 Quorum decision-making facilitates information transfer in fish shoals PNAS 105 6948ndash6953
Wells J M B C Raju H Y Hung W G Weisburg L Mandelco-Paul and D J Brenner 1987 Xylella fastidiosa gen nov sp nov gram-negative xylem-limited fastidious plant bacteria related to Xanthomonas spp Int J Syst Bacteriol 37 136ndash143
Werner E E 1992 Individual behavior and higher-order species interac-tions Am Nat (suppl) 140 S5ndashS32
White T R C 1978 The importance of relative shortage of food in animal ecology Oecologia 33 71ndash86
Williams G C 1966 Adaptation and natural selection Princeton University Press Princeton NJ
Wu D S C Daugherty S E Van Aken G H Pai K L Watkins H Khouri L J Tallon J M Zaborsky H E Dunbar P L Tran N A Moran J A Eisen 2006 Metabolic complementarity and genomics of the dual symbiosis of sharpshooters PloS-Biology 4 e188
Russell F Mizell III Is a Professor of Entomology (rfmizellufledu) Peter C Andersen is a Professor of Horticulture (peaufledu) and Brent V Brod-beck (bvbufledu) is a Biological Scientist all located at the University of Floridarsquos North Florida Research and Education Center-Quincy 155 Research Rd Quincy Fl 32351 Wayne B Hunter (whunterushrlarsusdagov) is a research entomologist with the USDA Agricultural Research Service at the US Horticultural Research Lab 2001 South Rock Rd Ft Pierce FL 34945 For over 20 years they have collaborated in research on the biology ecology and behavior of the leafhopper vectors of Xylella fastidiosa
With our high qualityLepidoptera Eggs and LarvaeBeet Army Worm bull Cabbage LooperCorn Earworm bull Diamondback MothFall Armyworm bull Tobacco Budworm
Save Money Save Time Get Results
Lepidoptera Eggs and LarvaeInsect Diets and Ingredients
Rearing Supplies and Equipment
Entomology Division
800-996-9908 US and Canada908-996-2155 Internationalsalesinsectrearingcom
wwwinsectrearingcom

112 American Entomologist bull Summer 2012
finding hosts the tradeoffs faced by female leafhoppers and the value of conspecific recognition in their mitigation An unusual number of mated leafhopper pairs also were observed among the cadaver treatment branches on the mornings of succeeding test days These observations provide further support for the contention concerning top down risk mitigation via conspecific response
Significant response to avoid predators and artificial models was not observed It is unlikely that an arthropod herbivore would be able to develop a search image with such elasticity that it could encompass the wide range of natural enemies potentially encoun-tered The immediate acceptance of the novel and poor host L limii provides further evidence against the occurrence of a learned host image by searching leafhoppers Response to conspecifics would obviate the necessity of developing a predator search image because conspecifics would usually be present only where predators were absent Thus a type of ldquoenemy-free spacerdquo (Jeffries and Lawton 1984) may be represented by conspecific presence Alternatively relative to the risk of landing on a branch without conspecifics even if the conspecific signal was not entirely truthful about the level of risk the dilution effect from conspecific presence would provide increased protection theoretically in direct proportion to the number of con-specifics present Although foragers in many animal groups have an inherent geometry that causes information asymmetries individuals get more information from closer neighbors (Fernandez-Juricic et al 2004) H vitripennis uses keen visual acuity that likely is coupled with acoustical communication as a mechanism to respond rapidly en masse to approaching predators As a result leafhoppers accrue a more equivalent gain from defensive movement coordination (Fernandez-Juricic et al 2004)
On the host plant H vitripennis can detect and respond (and routinely do so in concert perhaps mediated by acoustic commu-nication) to movement of large objects approaching them from off the plant and to smaller objects on the plant Live lizards do elicit a response from feeding H vitripennis as they move over the host plant and approach leafhoppers (Lopez et al 2003) This suggests the occurrence in H vitripennis of a general visual capability (differ-ent from the visual response to conspecifics) to recognize moving objects that would circumvent the necessity of developing an array of species-specific predator search images
H vitripennis responded significantly to cadavers painted gray that served as a model of H vitripennis infected with the parasitic fungal pathogen H homalodiscae (Boucias et al 2006) H vitripennis cadavers infected with this pathogen remain attached for months on the host plant at feeding sites in the feeding position and infect H vitripennis nymphs and adults The infected cadavers are covered by the fungal mycelia that sporulate in cycles and provide a means to infect uninfected feeding H vitripennis (Lietze et al 2010) H homalodiscae-infected H vitripennis attach tenaciously via rhizoids to the plant bark in the summer and are often found attached to this substrate the following spring This positive visual response may increase transmission of the fungus to H vitripennis
When live presumably acoustically active H vitripennis were added to treatment branches the numbers of H vitripennis increased by 53 on the treated branches (Table 1) H vitripennis unlike most insect herbivores has not been shown to use olfaction at any level for courtship or mating In contrast H vitripennis and many Cicadel-lidae and related insect families use acoustical vibrations propagated through the host plant to communicate during courtship and mating and for other functions (Cocroft 2006) This behavioral modality of
H vitripennis however has received only cursory attention (Percy et al 2008) and its potential effect was eliminated in all except this one experiment through the use of cadavers
With respect to host finding the role of olfaction remains un-clear Patt and Setamou (2007) reported that the response to visual cues by H vitripennis nymphs and adults was enhanced by exposure to plant volatiles Patt and Setamou (2010) demonstrated that H vitripennis nymphs could be conditioned to respond to vanilla as a novel host volatile and as a result they argued that associative learning of chemical profiles could enhance leafhopper location and recognition of host plants These authors also speculated on a potential mechanism use of plant volatile profile emissions (Ninemets et al 2002 2004) in host finding and recognition by adult H vitripennis While an interesting hypothesis much research remains to be done to support these ideas A strong response to conspecifics provides a complimentary or alternative mechanism that would eliminate the need for such highly evolved behaviors However one may hypothesize that the presence of plant volatiles could stimulate a more intense leafhopper visual search for conspe-cifics on the host Nevertheless the ready acceptance of the novel and poor host L limii in the presence of conspecifics argues against the need for leafhoppers to learn a visual or olfactory pattern for specific plant species
Trivial movement within and between host plants and host plant patches is germane to understanding the distribution and fitness of H vitripennis (Mizell et al 2008) H vitripennis are strongly visually oriented while feeding on a host plant they can detect movement from 5 to 10 m away Based on the ostensibly narrow sensory ca-pabilities of H vitripennis (lack of pheromones apparently limited use of olfaction) it is not unexpected that H vitripennis would have evolved visual behavior with multiple functions Moreover this simple capability would obviate the need for higher level behaviors such as associative learning at least for adults (Patt and Setamou 2010) Profiles of xylem chemistry can change as much as 10-fold in one day or from day to day in the absence of any overt visual signals to leafhoppers (Andersen et al1992 Brodbeck et al 2010) Because host plant nutrient quality varies unrecognizably to leafhoppers in time and space and between and within a plant species developing a search image to visually identify specific host plants or character-istics related to their quality appears to be extremely difficult and perhaps of little value Moreover host plants usually do not occur in monocultures a situation that would reinforce short-term pattern recognition By contrast delay in finding hosts within a patch can affect information retention (McLinn and Stephens 2006) Relying too heavily on recent experience can be costly especially if the environment and in this case host plant nutrients change rapidly (Mizell and French 1987 Andersen et al 1989 1992 1993 1995ab Brodbeck et al 1990 19951999 McLinn and Stephens 2006) Animals devalue unreliable signals Models predict signal following when the signal reliability exceeds environmental certaintymdashso that information is valuable Signal value however is not based solely on reliability it also depends on the statistical distribution of the environmental states (McLinn and Stephens 2006) Plant quality and predation risks are in constant flux in the case of H vitripennis and extremely variable in time and space H vitripennis handling of bottom-up and top-down forces is tied closely together because the potential payoff and consequences from both accrue on the host plant following a single decision to land H vitripennis appar-ently can make a decision to land based solely on a visual response
113American Entomologist bull Volume 58 Number 2
to conspecifics on host plant branches An arrestment response is then likely reinforced on the plant via acoustical communication as gustation behavior is initiated
Thus presence of conspecifics indicates a ldquosafety zonerdquo for host plant testing and likely provides high-quality (truthfulness of signal) information because all H vitripennis in any one location are under more or less the same dietary constraints Notwithstanding that O nigricans may have somewhat different nutritional requirements it may be important in the mitigation of bottom-up and top-down forces The presence of one or (usually) multiple conspecifics is likely a signal indicating a reasonably high threshold quality of nutrients or else the duration of conspecifics remaining on a host plant would be short This perhaps explains why three conspecifics analogous to the quorum decision of Ward et al (2008) maximize the response Three may be enough to compensate for any variations in individual hunger rate or other uninterpretable behaviors (females not feeding or ovipositing males that feed less) that may be ongoing and could affect the ldquotruth in signalrdquo presented by conspecifics
Arthropod populations face a variety of environmental risks to which they must adapt that can be described within the bottom-up and top-down heuristic model however ldquoNo creature can do all thingsrdquo and genetic and other constraints are many Thus tradeoffs are required for organisms to adapt and sustain over evolutionary time (Rolf 1992) Activity or movement is highly related to resource acquisition but it is also related to mortality risks a cost and as a result mediates one of the most general tradeoffs faced by organ-isms (Werner 1992) Information gathering also comes at a cost because of the loss of time potentially spent on other activities (Eliassen et al 2007) Moreover where resources vary temporally or for example predation risk is highly unpredictable the value of information decreases with time such that a forager should place greater value on the newest available information (Eliassen et al 2007) An individualrsquos response to conspecifics makes it possible to collect a broad range of real-time biological data that may reduce the tradeoffs and costs imposed by required activity Foraging suc-cess and predator avoidance behaviors directly related to fitness are strongly affected as a result (Lazarus 1979 Foster and Treherne 1981 Lawrence 1985 Stamps 1987 Turchin 1987) The environment of xylophagous leafhoppers is highly stochastic in time and space and their extreme polyphagy requires exposure to a wide array of plant structures changing chemical profiles and associated predators Response to conspecifics offers a simple parsimonious mechanism to eliminate the need to develop sophisticated physiological or behavioral mechanisms to handle this array of visual and chemical cues that accompany the extreme uncertainty presented by resource finding and predator avoidance
In conclusion H vitripennis exhibits at least two major types of visually based behaviors While in the normal feeding position (Bex-tine et al 2005) they can detect movement from asymp10 m distance and move behind the branch to hide themselves from the approaching object This response is often done synchronously by the leafhoppers present More importantly when searching for and landing on a host plant where the risk of predation may be high they can respond to cues presented by conspecifics These consist of a narrowly defined stimulus roughly equal in size to the H vitripennis body but not gt12 mm To our knowledge this is the first report of an arthropod exhibit-ing behavioral parsimony in which one signal a visual cue (response to conspecifics) is used to mitigate immediate risks from bottom-up and top-down forces In the present case leafhoppers exhibit a ldquofixedrdquo
response response to conspecifics to bottom-up and top-down forces independent of their value in the environmental context Avail-ability and spatial distribution of host plants and their nutritional quality are critical to the survival and fitness of H vitripennis The availability of xylem fluid within and between host plants changes continuously and rapidly over time and space requiring much trivial movement and emigration by leafhoppers These natural fluctuations in plant xylem chemistry are the primary basis of the bottom-up and top-down risks to xylophagous leafhoppers because they determine the movement and seasonal use of host plants by H vitripennis (be-havior and performance) (Mizell and French 1987 Andersen et al 1989 1992 1995ab 2005 2008 Brodbeck et al 1990 19951999) Previously several novel physiological and behavioral adaptations critical to leafhopper survival were documented (Mizell et al 2008) Response to conspecifics during host plant searching movements is another novel but fundamentally important behavior exhibited by these interesting disease vectors that allow them to survive and endure as xylophages
AcknowledgmentsThe research was partially funded by a grant to R F Mizell III and P
C Andersen from the California Department of Food and Agriculture Piercersquos DiseasemdashGlassy-winged Sharpshooter Research Program We thank Gary W Knox for use of the experimental plot containing the Lagerstroemia limii We thank Tobin Northfield Joseph Patt and Arash Rashad for helpful comments on a previous draft of the manuscript
References CitedAndersen P C B V Brodbeck and R M Mizell III 1989 Metabolism
of amino acids organic acids and sugars extracted from the xylem fluid of four host plants by Homalodisca coagulata Entomol Exp Appl 50 149ndash159
Andersen P C B V Brodbeck and R M Mizell III 1992 Feeding by the leafhopper Homalodisca coagulata in relation to xylem fluid chemistry and tension J Insect Physiol 38 611ndash622
Andersen P C B V Brodbeck and R M Mizell III 1993 Diurnal variations in the chemistry of xylem fluid from Lagerstroemia indica an endogenous circadian rhythm Physiol Plant 89 783-790
Andersen P C B V Brodbeck and R M Mizell III 1995a Diurnal variation in tension osmolarity and the composition of nitrogen and carbon assimilates in xylem fluid of Prunus persica Vitis hybrid and Prunus communis J Am Hortic Sci 120 600ndash604
Andersen P C B V Brodbeck and R M Mizell III 1995b Water stress- and nutrient-mediated changes in the water relations and the amino acids organic acids and sugars in xylem fluid of Prunus salicina and Lagerstroemia indica J Am Soc Hortic Sci 120 36ndash42
Andersen P C B V Brodbeck and R M Mizell III 2003 Plant and insect characteristics in response to increasing density of Homalodisca coagulata on three host species a quantification of assimilate extraction Entomol Exp Appl 107 57ndash68
Andersen P C B V Brodbeck R M Mizell III and S Oden 2005 The abundance and feeding of Homalodisca coagulata (Homoptera Auche-norrhyncha Cicadellidae) on Vitis genotypes in north Florida Environ Entomol 34 466ndash478
Andersen PC R M Mizell III B V Brodbeck T Beckman and G Krewer 2008 Abundance and consumption rates of glassy-winged sharpshooter (Hemiptera Cicadellidae) on peaches and plums J En-tomol Sci 43 394ndash407
Andersen P C B V Brodbeck and R M Mizell III 2009 Assimilation efficiency of free and protein amino acids by the glassy-winged sharp-shooter Homalodisca vitripennis (Germar) (Hemiptera Cicadellidae Cicadellinae) feeding on Citrus sinensis and Vitis vinifera Fla Entomol 92 116ndash122
Backus E J Habibi F Yan and M Ellersieck 2005 Stylet penetration by adult Homalodisca coagulata on grape electrical penetration graph
114 American Entomologist bull Summer 2012
waveform characterization tissue correlation and possible implica-tions for transmission of Xylella fastidiosa Ann Entomol Soc Am 98 787ndash813
Baker P S 1979 Flying locust visual responses in a radial wind tunnel J Comp Physiol 131 39ndash47
Bextine B R B C Jackson D B Harshman and Miller 2005 Homalo-disca coagulata (Hemiptera Cicadellidae) feeding posture Ann Ento-mol Soc Am 98 814ndash819
Boucias D G D W Scharf S E Breaux S D Purcell and R M Mizell III 2006 Studies on the fungi associated with the glassy-winged sharpshooter Homalodisca coagulata with emphasis on a new species Hirsutella homalodiscae nom prov BioControl 52 231ndash258
Brodbeck BV R M Mizell III and P C Andersen 1990 Amino acids as determinants of host preference for the xylemfeeding leafhopper Homalodisca coagulata Oecologia 83 338ndash345
Brodbeck B V P C Andersen and R M Mizell III 1993 Physiological and behavioral adaptations of three species of leafhoppers in response to the dilute nutrient content of xylem fluid J Insect Physiol 39 73ndash81
Brodbeck B V P C Andersen and R M Mizell III 1995 Differential utilization of nutrients during development by the xylophagous leafhop-per Homalodisca coagulata Entomol Exp Appl 75 279ndash289
Brodbeck BV P C Andersen and R M Mizell III 1999 The effects of total dietary nitrogen and amino acid balance on the development of xylophagous leafhoppers Arch Insect Biochem Physiol 42 37ndash50
Brodbeck B V P C Andersen and R M Mizell III 2007 Preference-performance linkage of the xylem feeding leafhopper Homalodisca vit-ripennis (Hemiptera Cicadellidae) Environ Entomol 36 1512ndash1522
Brodbeck B V P C Andersen and R M Mizell III 2011 Nutrient media-tion of behavioral plasticity and resource allocation in a xylem-feeding leafhopper Oecol 165 111-122
Cocroft R B H J Shugart K T Konrad and K Tibbs 2006 Variation in plant substrates and its consequences for insect vibrational com-munication Ethology 112 779ndash789
Collett T S and M F Land 1978 How hoverflies compute interception courses J Comp Physiol 125 191ndash204
Coolen I O Dangles and J Casas 2005 Social learning in non-colonial insects Curr Biol 15 1931-1935
Daane K M Johnson T Ruiz and J M Hashim 2005 Host plant influence on glassy-winged sharpshooter and its natural enemies pp 100ndash104 In Proceedings 2005 Piercersquos Disease Research Symposium California Department of Food and Agriculture Sacramento
Dukas R 2001 Effects of perceived danger on flower choice by bees Ecol Lett 4 327ndash333
Dukas R and D H Morse 2003 Crab spiders affect flower visitation by bees Oikos 101 157ndash163
Eliassen S E C Jargensen M Mangel and J Garl 2007 Exploration or exploitation life expectancy changes the value of learning in foraging strategies Oikos 116 513ndash523
Fernandez-Juricic E B Kerr P Bednekoff and D Stephens 2004 When are two heads better than one Visual perception and informa-tion transfer affect vigilance coordination in foraging groups Behav Ecol 6 898ndash906
Foster W A and J E Treherne 1981 Evidence for the dilution effect in the selfish herd from fish predation on a marine insect Nature 293 466ndash467
Fournier V J Hagler K Daane J de Leoacuten and R Groves 2008 Iden-tifying the predator complex of Homalodisca vitripennis (Hemiptera Cicadellidae) a comparative study of the efficacy of an ELISA and PCR gut content bioassay Oecologia 157 629ndash640
Giraldeau L T J Valone and J J Templeton 2002 Potential disadvan-tages of using socially acquired information Phil Trans R Soc Lond B 357 1559ndash1566
Hamiliton WD 1971 Geometry for the selfish herd J Theror Biol 31295-311
Hairston N G J E Smith and L B Slobodkin 1960 Community struc-ture population control and competition Am Midl Nat 106 249ndash257
Hunter W B and D E Ullman 1994 Precibarial and cibarial chemosen-sillae in the western flower thrips Frankliniella occidentalis (Pergande)(Thysanoptera Thripidae) Int J Insect Morphol Embryol 23 69ndash83
Hunter W B E Hiebert S Webb J E Polston and J H Tsai 1996 Precibarial and cibarial chemosensilla in the whitefly Bemisia tabaci
(Gennadius) (Homoptera Aleyrodidae) Int J Insect Morphol Embryol 25 295ndash304
Jeffries M J and J H Lawton 1984 Enemy-free space and the structure of ecological communities Biol J Linn Soc 23 269-286
Karban R and L T Baldwin 1997 Induced response to herbivory University of Chicago Press Chicago
Land M F and T S Collett 1974 Chasing behavior of houseflies (Fannia canicularis) J Comp Physiol 80 331ndash357
Lawrence E S 1985 Vigilance during ldquoeasyrdquo and ldquodifficultrdquo foraging tasks Anim Behav 33 1373ndash1375
Lawton J H and S McNeil1979 Between the devil and the deep blue sea on the problem of being a herbivore pp 223ndash244 In R M Anderson B D Turner and L R Taylor [Eds] Population dynamics Blackwell Oxford England Blackwell Sci Publ 434 pp
Lazarus J 1979 Flock size and behavior in captive red-billed weaverbirds (Quelea quelea) implications for social facilitation and the functions of flocking Behavior 71 1ndash2
Lietze V R M Mizell III and D G Boucias 2011 Transmission of the mycopathogen Hirsutella spp to nymphs and adults of the glassy-winged sharpshooters Homalodisca vitripennis (=coagulata) in the greenhouse Fla Entomol 94 106-108
Lopez R F R Mizell III P C Andersen and B V Brodbeck 2003 Impact of natural enemies on populations of the glassy-winged sharpshooter Homalodisca coagulata (Say) in north Florida In R VanDriesche [Ed] First international symposium on biological control of arthropods US Dep Agric For Ser Pub FHTET-03-05 484ndash486
Mainman G A D Straw and M H Dickson 2008 A simple vision-based algorithm for decision making in flying Drosophila Curr Biol 18 464ndash470
McLinn C and D W Stephens 2006 What makes information valu-able signal reliability and environmental uncertainty Anim Behav 71 1119ndash1129
Mizell R F III and W J French 1987 Leafhopper vectors of phony peach disease feeding site preference and survival on infected and uninfected peach and seasonal response to selected host plants J Entomol Sci 22 1122
Mizell R F III P C Andersen P C Phillips and P Oevering 2005 Population dynamics and interactions between the glassy-winged sharpshooter and the host plants in response to California phenology pp 117ndash121 In Proceedings 2005 Piercersquos Disease Research Symposium California Department of Food and Agriculture Sacramento
Mizell R F III C Tipping P C Andersen B V Brodbeck T Northfield and W Hunter W 2008 Behavioral model for the glassy-winged sharp-shooter Homalodisca vitripennis (Hemiptera Cicadellidae) optimiza-tion of host plant utilization and management implications Environ Entomol 37 1049ndash1062
Mizell R F III PC Andersen and B V Brodbeck 2009 Improved detec-tion monitoring and management of the glassy-winged sharpshooter pp 40ndash43 In Proceedings 2009 Piercersquos Disease Research Symposium California Department of Food and Agriculture Sacramento
Muller KL 1998 The role of conspecifics in habitat settlement in a ter-ritorial grasshopper Anim Behav 56 479ndash485
Ninemets Uuml M Riechstein M Staudt G Siefert and J Tenhunen 2002 Stomatal constraints may affect emission of oxygenated monoterpenoids from foliage of Pinus pinea Plant Physiol 130 1371ndash1385
Ninemets Uuml F Loreto and M Riechstein 2004 Physiological and physiochemical controls on foliar volatile organic compound emissions Trends Plant Sci 9 180ndash186
Northfield T R F Mizell III P C Andersen B V Brodbeck T C Riddle and W B Hunter 2009 Dispersal patch leaving and aggregation of the glassy-winged sharpshooter Homalodisca vitripennis (Hemiptera Cicadellidae) Environ Entomol 38 183ndash191
Patt J and M Setamou 2007 Olfactory and visual stimuli affecting host plant detection in Homalodisca coagulata (Hemiptera Cicadellidae) Environ Entomol 36 142ndash150
Patt J and M Setamou 2010 Recognition of novel volatile cues by the nymph of the glassy-winged sharpshooter Homalodisca vitripennis (Cicadellidae) J Insect Behav DOI 101007s10905-010-9214-z 23 290-302
Percy D M E A Boyd and M S Hoddle 2008 Observations of acoustical signaling in three sharpshooters Homalodisca vitripennis Homalodisca
115American Entomologist bull Volume 58 Number 2
liturata and Graphocephala atropunctata (Hemiptera Cicadellidae) Ann Entomol Soc 101 253ndash259
Power M E 1992 Top-down and bottom-up forces in food webs do plants have primacy Ecologia 73 733ndash746
Price P W C E Bouton P Gross B A McPheron J N Thompson and A E Wise 1980 Interactions among three trophic levels influence of plants on interactions between insect herbivores and natural enemies Annu Rev Ecol Syst 11 41ndash66
Redak R M J Blua K L Lopes A H Purcell R F Mizell III and P C Andersen 2004 The biology and control of xylem sap-feeding insect vectors of Xylella fastidiosa and their relation to disease epidemiology Annu Rev Entomol 49 243ndash270
Rolf D A 1992 The evolution of life history traits Chapman amp Hall NYSandlin E A 2000 Cue use affects resource subdivision among three
coexisting hummingbird species Behav Ecol 11 550-559SAS Institute 2009 Proc GLM SAS 93 Statistical Analysis System Cary NCShrewsbury P and M Raupp 2008 Do top-down or bottom-up forces
determine Stephanitis pyroides abundance in urban landscapes Ecol Appl 16 262ndash273
Systat Software 2009 SigmaPlot for Windows 110 Systat Software Bangalore India
Stamps J A 1987 Conspecifics as cues to territory quality a preference of juvenile lizards (Anolis aeneus) for previously used territories Am Nat 129 629ndash642
Stamps J A 1988 Conspecific attraction and aggregation in territorial species Am Nat 131 330ndash347
Thompson J D S C Peterson and L D Harder 1987 Response to traplining bumble bees to competition experiments shifts in feeding location and efficiency Oecologia 71 295ndash300
Tipping C R F Mizell III and P C Andersen 2004 Adaptations impor-tant for the dispersal of immature stages of three species of leafhopper (Homoptera Cicadellidae) Fla Entomol 87 372ndash379
Triapitsyn S V R F Mizell III J L Bossert and J E Carlton 1998 Egg parasitoids of glassy-winged sharpshooter Homalodisca coagulata (Homoptera Cicadellidae) Fla Entomol 81 243ndash251
Turner W F and H N Pollard 1959 Life histories and behavior of five insect vectors of phony peach disease Tech Bull US Dep Agric 1188
Turchin P 1987 The role of aggregation in the response of Mexican bean beetles to host-plant density Oecologia 71 577ndash582
Valone T J 1989 Group foraging public information and patch simula-tion Oikos 56 357ndash363
Walker M and T H Jones 2001 Relative roles of top-down and bottom-up forces in terrestrial tritrophic plant-insect herbivore-natural enemy systems Oikos 93 177ndash187
Ward A J W D J T Sumpter I D Couzin P J B Hart and J Krause 2008 Quorum decision-making facilitates information transfer in fish shoals PNAS 105 6948ndash6953
Wells J M B C Raju H Y Hung W G Weisburg L Mandelco-Paul and D J Brenner 1987 Xylella fastidiosa gen nov sp nov gram-negative xylem-limited fastidious plant bacteria related to Xanthomonas spp Int J Syst Bacteriol 37 136ndash143
Werner E E 1992 Individual behavior and higher-order species interac-tions Am Nat (suppl) 140 S5ndashS32
White T R C 1978 The importance of relative shortage of food in animal ecology Oecologia 33 71ndash86
Williams G C 1966 Adaptation and natural selection Princeton University Press Princeton NJ
Wu D S C Daugherty S E Van Aken G H Pai K L Watkins H Khouri L J Tallon J M Zaborsky H E Dunbar P L Tran N A Moran J A Eisen 2006 Metabolic complementarity and genomics of the dual symbiosis of sharpshooters PloS-Biology 4 e188
Russell F Mizell III Is a Professor of Entomology (rfmizellufledu) Peter C Andersen is a Professor of Horticulture (peaufledu) and Brent V Brod-beck (bvbufledu) is a Biological Scientist all located at the University of Floridarsquos North Florida Research and Education Center-Quincy 155 Research Rd Quincy Fl 32351 Wayne B Hunter (whunterushrlarsusdagov) is a research entomologist with the USDA Agricultural Research Service at the US Horticultural Research Lab 2001 South Rock Rd Ft Pierce FL 34945 For over 20 years they have collaborated in research on the biology ecology and behavior of the leafhopper vectors of Xylella fastidiosa
With our high qualityLepidoptera Eggs and LarvaeBeet Army Worm bull Cabbage LooperCorn Earworm bull Diamondback MothFall Armyworm bull Tobacco Budworm
Save Money Save Time Get Results
Lepidoptera Eggs and LarvaeInsect Diets and Ingredients
Rearing Supplies and Equipment
Entomology Division
800-996-9908 US and Canada908-996-2155 Internationalsalesinsectrearingcom
wwwinsectrearingcom

113American Entomologist bull Volume 58 Number 2
to conspecifics on host plant branches An arrestment response is then likely reinforced on the plant via acoustical communication as gustation behavior is initiated
Thus presence of conspecifics indicates a ldquosafety zonerdquo for host plant testing and likely provides high-quality (truthfulness of signal) information because all H vitripennis in any one location are under more or less the same dietary constraints Notwithstanding that O nigricans may have somewhat different nutritional requirements it may be important in the mitigation of bottom-up and top-down forces The presence of one or (usually) multiple conspecifics is likely a signal indicating a reasonably high threshold quality of nutrients or else the duration of conspecifics remaining on a host plant would be short This perhaps explains why three conspecifics analogous to the quorum decision of Ward et al (2008) maximize the response Three may be enough to compensate for any variations in individual hunger rate or other uninterpretable behaviors (females not feeding or ovipositing males that feed less) that may be ongoing and could affect the ldquotruth in signalrdquo presented by conspecifics
Arthropod populations face a variety of environmental risks to which they must adapt that can be described within the bottom-up and top-down heuristic model however ldquoNo creature can do all thingsrdquo and genetic and other constraints are many Thus tradeoffs are required for organisms to adapt and sustain over evolutionary time (Rolf 1992) Activity or movement is highly related to resource acquisition but it is also related to mortality risks a cost and as a result mediates one of the most general tradeoffs faced by organ-isms (Werner 1992) Information gathering also comes at a cost because of the loss of time potentially spent on other activities (Eliassen et al 2007) Moreover where resources vary temporally or for example predation risk is highly unpredictable the value of information decreases with time such that a forager should place greater value on the newest available information (Eliassen et al 2007) An individualrsquos response to conspecifics makes it possible to collect a broad range of real-time biological data that may reduce the tradeoffs and costs imposed by required activity Foraging suc-cess and predator avoidance behaviors directly related to fitness are strongly affected as a result (Lazarus 1979 Foster and Treherne 1981 Lawrence 1985 Stamps 1987 Turchin 1987) The environment of xylophagous leafhoppers is highly stochastic in time and space and their extreme polyphagy requires exposure to a wide array of plant structures changing chemical profiles and associated predators Response to conspecifics offers a simple parsimonious mechanism to eliminate the need to develop sophisticated physiological or behavioral mechanisms to handle this array of visual and chemical cues that accompany the extreme uncertainty presented by resource finding and predator avoidance
In conclusion H vitripennis exhibits at least two major types of visually based behaviors While in the normal feeding position (Bex-tine et al 2005) they can detect movement from asymp10 m distance and move behind the branch to hide themselves from the approaching object This response is often done synchronously by the leafhoppers present More importantly when searching for and landing on a host plant where the risk of predation may be high they can respond to cues presented by conspecifics These consist of a narrowly defined stimulus roughly equal in size to the H vitripennis body but not gt12 mm To our knowledge this is the first report of an arthropod exhibit-ing behavioral parsimony in which one signal a visual cue (response to conspecifics) is used to mitigate immediate risks from bottom-up and top-down forces In the present case leafhoppers exhibit a ldquofixedrdquo
response response to conspecifics to bottom-up and top-down forces independent of their value in the environmental context Avail-ability and spatial distribution of host plants and their nutritional quality are critical to the survival and fitness of H vitripennis The availability of xylem fluid within and between host plants changes continuously and rapidly over time and space requiring much trivial movement and emigration by leafhoppers These natural fluctuations in plant xylem chemistry are the primary basis of the bottom-up and top-down risks to xylophagous leafhoppers because they determine the movement and seasonal use of host plants by H vitripennis (be-havior and performance) (Mizell and French 1987 Andersen et al 1989 1992 1995ab 2005 2008 Brodbeck et al 1990 19951999) Previously several novel physiological and behavioral adaptations critical to leafhopper survival were documented (Mizell et al 2008) Response to conspecifics during host plant searching movements is another novel but fundamentally important behavior exhibited by these interesting disease vectors that allow them to survive and endure as xylophages
AcknowledgmentsThe research was partially funded by a grant to R F Mizell III and P
C Andersen from the California Department of Food and Agriculture Piercersquos DiseasemdashGlassy-winged Sharpshooter Research Program We thank Gary W Knox for use of the experimental plot containing the Lagerstroemia limii We thank Tobin Northfield Joseph Patt and Arash Rashad for helpful comments on a previous draft of the manuscript
References CitedAndersen P C B V Brodbeck and R M Mizell III 1989 Metabolism
of amino acids organic acids and sugars extracted from the xylem fluid of four host plants by Homalodisca coagulata Entomol Exp Appl 50 149ndash159
Andersen P C B V Brodbeck and R M Mizell III 1992 Feeding by the leafhopper Homalodisca coagulata in relation to xylem fluid chemistry and tension J Insect Physiol 38 611ndash622
Andersen P C B V Brodbeck and R M Mizell III 1993 Diurnal variations in the chemistry of xylem fluid from Lagerstroemia indica an endogenous circadian rhythm Physiol Plant 89 783-790
Andersen P C B V Brodbeck and R M Mizell III 1995a Diurnal variation in tension osmolarity and the composition of nitrogen and carbon assimilates in xylem fluid of Prunus persica Vitis hybrid and Prunus communis J Am Hortic Sci 120 600ndash604
Andersen P C B V Brodbeck and R M Mizell III 1995b Water stress- and nutrient-mediated changes in the water relations and the amino acids organic acids and sugars in xylem fluid of Prunus salicina and Lagerstroemia indica J Am Soc Hortic Sci 120 36ndash42
Andersen P C B V Brodbeck and R M Mizell III 2003 Plant and insect characteristics in response to increasing density of Homalodisca coagulata on three host species a quantification of assimilate extraction Entomol Exp Appl 107 57ndash68
Andersen P C B V Brodbeck R M Mizell III and S Oden 2005 The abundance and feeding of Homalodisca coagulata (Homoptera Auche-norrhyncha Cicadellidae) on Vitis genotypes in north Florida Environ Entomol 34 466ndash478
Andersen PC R M Mizell III B V Brodbeck T Beckman and G Krewer 2008 Abundance and consumption rates of glassy-winged sharpshooter (Hemiptera Cicadellidae) on peaches and plums J En-tomol Sci 43 394ndash407
Andersen P C B V Brodbeck and R M Mizell III 2009 Assimilation efficiency of free and protein amino acids by the glassy-winged sharp-shooter Homalodisca vitripennis (Germar) (Hemiptera Cicadellidae Cicadellinae) feeding on Citrus sinensis and Vitis vinifera Fla Entomol 92 116ndash122
Backus E J Habibi F Yan and M Ellersieck 2005 Stylet penetration by adult Homalodisca coagulata on grape electrical penetration graph
114 American Entomologist bull Summer 2012
waveform characterization tissue correlation and possible implica-tions for transmission of Xylella fastidiosa Ann Entomol Soc Am 98 787ndash813
Baker P S 1979 Flying locust visual responses in a radial wind tunnel J Comp Physiol 131 39ndash47
Bextine B R B C Jackson D B Harshman and Miller 2005 Homalo-disca coagulata (Hemiptera Cicadellidae) feeding posture Ann Ento-mol Soc Am 98 814ndash819
Boucias D G D W Scharf S E Breaux S D Purcell and R M Mizell III 2006 Studies on the fungi associated with the glassy-winged sharpshooter Homalodisca coagulata with emphasis on a new species Hirsutella homalodiscae nom prov BioControl 52 231ndash258
Brodbeck BV R M Mizell III and P C Andersen 1990 Amino acids as determinants of host preference for the xylemfeeding leafhopper Homalodisca coagulata Oecologia 83 338ndash345
Brodbeck B V P C Andersen and R M Mizell III 1993 Physiological and behavioral adaptations of three species of leafhoppers in response to the dilute nutrient content of xylem fluid J Insect Physiol 39 73ndash81
Brodbeck B V P C Andersen and R M Mizell III 1995 Differential utilization of nutrients during development by the xylophagous leafhop-per Homalodisca coagulata Entomol Exp Appl 75 279ndash289
Brodbeck BV P C Andersen and R M Mizell III 1999 The effects of total dietary nitrogen and amino acid balance on the development of xylophagous leafhoppers Arch Insect Biochem Physiol 42 37ndash50
Brodbeck B V P C Andersen and R M Mizell III 2007 Preference-performance linkage of the xylem feeding leafhopper Homalodisca vit-ripennis (Hemiptera Cicadellidae) Environ Entomol 36 1512ndash1522
Brodbeck B V P C Andersen and R M Mizell III 2011 Nutrient media-tion of behavioral plasticity and resource allocation in a xylem-feeding leafhopper Oecol 165 111-122
Cocroft R B H J Shugart K T Konrad and K Tibbs 2006 Variation in plant substrates and its consequences for insect vibrational com-munication Ethology 112 779ndash789
Collett T S and M F Land 1978 How hoverflies compute interception courses J Comp Physiol 125 191ndash204
Coolen I O Dangles and J Casas 2005 Social learning in non-colonial insects Curr Biol 15 1931-1935
Daane K M Johnson T Ruiz and J M Hashim 2005 Host plant influence on glassy-winged sharpshooter and its natural enemies pp 100ndash104 In Proceedings 2005 Piercersquos Disease Research Symposium California Department of Food and Agriculture Sacramento
Dukas R 2001 Effects of perceived danger on flower choice by bees Ecol Lett 4 327ndash333
Dukas R and D H Morse 2003 Crab spiders affect flower visitation by bees Oikos 101 157ndash163
Eliassen S E C Jargensen M Mangel and J Garl 2007 Exploration or exploitation life expectancy changes the value of learning in foraging strategies Oikos 116 513ndash523
Fernandez-Juricic E B Kerr P Bednekoff and D Stephens 2004 When are two heads better than one Visual perception and informa-tion transfer affect vigilance coordination in foraging groups Behav Ecol 6 898ndash906
Foster W A and J E Treherne 1981 Evidence for the dilution effect in the selfish herd from fish predation on a marine insect Nature 293 466ndash467
Fournier V J Hagler K Daane J de Leoacuten and R Groves 2008 Iden-tifying the predator complex of Homalodisca vitripennis (Hemiptera Cicadellidae) a comparative study of the efficacy of an ELISA and PCR gut content bioassay Oecologia 157 629ndash640
Giraldeau L T J Valone and J J Templeton 2002 Potential disadvan-tages of using socially acquired information Phil Trans R Soc Lond B 357 1559ndash1566
Hamiliton WD 1971 Geometry for the selfish herd J Theror Biol 31295-311
Hairston N G J E Smith and L B Slobodkin 1960 Community struc-ture population control and competition Am Midl Nat 106 249ndash257
Hunter W B and D E Ullman 1994 Precibarial and cibarial chemosen-sillae in the western flower thrips Frankliniella occidentalis (Pergande)(Thysanoptera Thripidae) Int J Insect Morphol Embryol 23 69ndash83
Hunter W B E Hiebert S Webb J E Polston and J H Tsai 1996 Precibarial and cibarial chemosensilla in the whitefly Bemisia tabaci
(Gennadius) (Homoptera Aleyrodidae) Int J Insect Morphol Embryol 25 295ndash304
Jeffries M J and J H Lawton 1984 Enemy-free space and the structure of ecological communities Biol J Linn Soc 23 269-286
Karban R and L T Baldwin 1997 Induced response to herbivory University of Chicago Press Chicago
Land M F and T S Collett 1974 Chasing behavior of houseflies (Fannia canicularis) J Comp Physiol 80 331ndash357
Lawrence E S 1985 Vigilance during ldquoeasyrdquo and ldquodifficultrdquo foraging tasks Anim Behav 33 1373ndash1375
Lawton J H and S McNeil1979 Between the devil and the deep blue sea on the problem of being a herbivore pp 223ndash244 In R M Anderson B D Turner and L R Taylor [Eds] Population dynamics Blackwell Oxford England Blackwell Sci Publ 434 pp
Lazarus J 1979 Flock size and behavior in captive red-billed weaverbirds (Quelea quelea) implications for social facilitation and the functions of flocking Behavior 71 1ndash2
Lietze V R M Mizell III and D G Boucias 2011 Transmission of the mycopathogen Hirsutella spp to nymphs and adults of the glassy-winged sharpshooters Homalodisca vitripennis (=coagulata) in the greenhouse Fla Entomol 94 106-108
Lopez R F R Mizell III P C Andersen and B V Brodbeck 2003 Impact of natural enemies on populations of the glassy-winged sharpshooter Homalodisca coagulata (Say) in north Florida In R VanDriesche [Ed] First international symposium on biological control of arthropods US Dep Agric For Ser Pub FHTET-03-05 484ndash486
Mainman G A D Straw and M H Dickson 2008 A simple vision-based algorithm for decision making in flying Drosophila Curr Biol 18 464ndash470
McLinn C and D W Stephens 2006 What makes information valu-able signal reliability and environmental uncertainty Anim Behav 71 1119ndash1129
Mizell R F III and W J French 1987 Leafhopper vectors of phony peach disease feeding site preference and survival on infected and uninfected peach and seasonal response to selected host plants J Entomol Sci 22 1122
Mizell R F III P C Andersen P C Phillips and P Oevering 2005 Population dynamics and interactions between the glassy-winged sharpshooter and the host plants in response to California phenology pp 117ndash121 In Proceedings 2005 Piercersquos Disease Research Symposium California Department of Food and Agriculture Sacramento
Mizell R F III C Tipping P C Andersen B V Brodbeck T Northfield and W Hunter W 2008 Behavioral model for the glassy-winged sharp-shooter Homalodisca vitripennis (Hemiptera Cicadellidae) optimiza-tion of host plant utilization and management implications Environ Entomol 37 1049ndash1062
Mizell R F III PC Andersen and B V Brodbeck 2009 Improved detec-tion monitoring and management of the glassy-winged sharpshooter pp 40ndash43 In Proceedings 2009 Piercersquos Disease Research Symposium California Department of Food and Agriculture Sacramento
Muller KL 1998 The role of conspecifics in habitat settlement in a ter-ritorial grasshopper Anim Behav 56 479ndash485
Ninemets Uuml M Riechstein M Staudt G Siefert and J Tenhunen 2002 Stomatal constraints may affect emission of oxygenated monoterpenoids from foliage of Pinus pinea Plant Physiol 130 1371ndash1385
Ninemets Uuml F Loreto and M Riechstein 2004 Physiological and physiochemical controls on foliar volatile organic compound emissions Trends Plant Sci 9 180ndash186
Northfield T R F Mizell III P C Andersen B V Brodbeck T C Riddle and W B Hunter 2009 Dispersal patch leaving and aggregation of the glassy-winged sharpshooter Homalodisca vitripennis (Hemiptera Cicadellidae) Environ Entomol 38 183ndash191
Patt J and M Setamou 2007 Olfactory and visual stimuli affecting host plant detection in Homalodisca coagulata (Hemiptera Cicadellidae) Environ Entomol 36 142ndash150
Patt J and M Setamou 2010 Recognition of novel volatile cues by the nymph of the glassy-winged sharpshooter Homalodisca vitripennis (Cicadellidae) J Insect Behav DOI 101007s10905-010-9214-z 23 290-302
Percy D M E A Boyd and M S Hoddle 2008 Observations of acoustical signaling in three sharpshooters Homalodisca vitripennis Homalodisca
115American Entomologist bull Volume 58 Number 2
liturata and Graphocephala atropunctata (Hemiptera Cicadellidae) Ann Entomol Soc 101 253ndash259
Power M E 1992 Top-down and bottom-up forces in food webs do plants have primacy Ecologia 73 733ndash746
Price P W C E Bouton P Gross B A McPheron J N Thompson and A E Wise 1980 Interactions among three trophic levels influence of plants on interactions between insect herbivores and natural enemies Annu Rev Ecol Syst 11 41ndash66
Redak R M J Blua K L Lopes A H Purcell R F Mizell III and P C Andersen 2004 The biology and control of xylem sap-feeding insect vectors of Xylella fastidiosa and their relation to disease epidemiology Annu Rev Entomol 49 243ndash270
Rolf D A 1992 The evolution of life history traits Chapman amp Hall NYSandlin E A 2000 Cue use affects resource subdivision among three
coexisting hummingbird species Behav Ecol 11 550-559SAS Institute 2009 Proc GLM SAS 93 Statistical Analysis System Cary NCShrewsbury P and M Raupp 2008 Do top-down or bottom-up forces
determine Stephanitis pyroides abundance in urban landscapes Ecol Appl 16 262ndash273
Systat Software 2009 SigmaPlot for Windows 110 Systat Software Bangalore India
Stamps J A 1987 Conspecifics as cues to territory quality a preference of juvenile lizards (Anolis aeneus) for previously used territories Am Nat 129 629ndash642
Stamps J A 1988 Conspecific attraction and aggregation in territorial species Am Nat 131 330ndash347
Thompson J D S C Peterson and L D Harder 1987 Response to traplining bumble bees to competition experiments shifts in feeding location and efficiency Oecologia 71 295ndash300
Tipping C R F Mizell III and P C Andersen 2004 Adaptations impor-tant for the dispersal of immature stages of three species of leafhopper (Homoptera Cicadellidae) Fla Entomol 87 372ndash379
Triapitsyn S V R F Mizell III J L Bossert and J E Carlton 1998 Egg parasitoids of glassy-winged sharpshooter Homalodisca coagulata (Homoptera Cicadellidae) Fla Entomol 81 243ndash251
Turner W F and H N Pollard 1959 Life histories and behavior of five insect vectors of phony peach disease Tech Bull US Dep Agric 1188
Turchin P 1987 The role of aggregation in the response of Mexican bean beetles to host-plant density Oecologia 71 577ndash582
Valone T J 1989 Group foraging public information and patch simula-tion Oikos 56 357ndash363
Walker M and T H Jones 2001 Relative roles of top-down and bottom-up forces in terrestrial tritrophic plant-insect herbivore-natural enemy systems Oikos 93 177ndash187
Ward A J W D J T Sumpter I D Couzin P J B Hart and J Krause 2008 Quorum decision-making facilitates information transfer in fish shoals PNAS 105 6948ndash6953
Wells J M B C Raju H Y Hung W G Weisburg L Mandelco-Paul and D J Brenner 1987 Xylella fastidiosa gen nov sp nov gram-negative xylem-limited fastidious plant bacteria related to Xanthomonas spp Int J Syst Bacteriol 37 136ndash143
Werner E E 1992 Individual behavior and higher-order species interac-tions Am Nat (suppl) 140 S5ndashS32
White T R C 1978 The importance of relative shortage of food in animal ecology Oecologia 33 71ndash86
Williams G C 1966 Adaptation and natural selection Princeton University Press Princeton NJ
Wu D S C Daugherty S E Van Aken G H Pai K L Watkins H Khouri L J Tallon J M Zaborsky H E Dunbar P L Tran N A Moran J A Eisen 2006 Metabolic complementarity and genomics of the dual symbiosis of sharpshooters PloS-Biology 4 e188
Russell F Mizell III Is a Professor of Entomology (rfmizellufledu) Peter C Andersen is a Professor of Horticulture (peaufledu) and Brent V Brod-beck (bvbufledu) is a Biological Scientist all located at the University of Floridarsquos North Florida Research and Education Center-Quincy 155 Research Rd Quincy Fl 32351 Wayne B Hunter (whunterushrlarsusdagov) is a research entomologist with the USDA Agricultural Research Service at the US Horticultural Research Lab 2001 South Rock Rd Ft Pierce FL 34945 For over 20 years they have collaborated in research on the biology ecology and behavior of the leafhopper vectors of Xylella fastidiosa
With our high qualityLepidoptera Eggs and LarvaeBeet Army Worm bull Cabbage LooperCorn Earworm bull Diamondback MothFall Armyworm bull Tobacco Budworm
Save Money Save Time Get Results
Lepidoptera Eggs and LarvaeInsect Diets and Ingredients
Rearing Supplies and Equipment
Entomology Division
800-996-9908 US and Canada908-996-2155 Internationalsalesinsectrearingcom
wwwinsectrearingcom

114 American Entomologist bull Summer 2012
waveform characterization tissue correlation and possible implica-tions for transmission of Xylella fastidiosa Ann Entomol Soc Am 98 787ndash813
Baker P S 1979 Flying locust visual responses in a radial wind tunnel J Comp Physiol 131 39ndash47
Bextine B R B C Jackson D B Harshman and Miller 2005 Homalo-disca coagulata (Hemiptera Cicadellidae) feeding posture Ann Ento-mol Soc Am 98 814ndash819
Boucias D G D W Scharf S E Breaux S D Purcell and R M Mizell III 2006 Studies on the fungi associated with the glassy-winged sharpshooter Homalodisca coagulata with emphasis on a new species Hirsutella homalodiscae nom prov BioControl 52 231ndash258
Brodbeck BV R M Mizell III and P C Andersen 1990 Amino acids as determinants of host preference for the xylemfeeding leafhopper Homalodisca coagulata Oecologia 83 338ndash345
Brodbeck B V P C Andersen and R M Mizell III 1993 Physiological and behavioral adaptations of three species of leafhoppers in response to the dilute nutrient content of xylem fluid J Insect Physiol 39 73ndash81
Brodbeck B V P C Andersen and R M Mizell III 1995 Differential utilization of nutrients during development by the xylophagous leafhop-per Homalodisca coagulata Entomol Exp Appl 75 279ndash289
Brodbeck BV P C Andersen and R M Mizell III 1999 The effects of total dietary nitrogen and amino acid balance on the development of xylophagous leafhoppers Arch Insect Biochem Physiol 42 37ndash50
Brodbeck B V P C Andersen and R M Mizell III 2007 Preference-performance linkage of the xylem feeding leafhopper Homalodisca vit-ripennis (Hemiptera Cicadellidae) Environ Entomol 36 1512ndash1522
Brodbeck B V P C Andersen and R M Mizell III 2011 Nutrient media-tion of behavioral plasticity and resource allocation in a xylem-feeding leafhopper Oecol 165 111-122
Cocroft R B H J Shugart K T Konrad and K Tibbs 2006 Variation in plant substrates and its consequences for insect vibrational com-munication Ethology 112 779ndash789
Collett T S and M F Land 1978 How hoverflies compute interception courses J Comp Physiol 125 191ndash204
Coolen I O Dangles and J Casas 2005 Social learning in non-colonial insects Curr Biol 15 1931-1935
Daane K M Johnson T Ruiz and J M Hashim 2005 Host plant influence on glassy-winged sharpshooter and its natural enemies pp 100ndash104 In Proceedings 2005 Piercersquos Disease Research Symposium California Department of Food and Agriculture Sacramento
Dukas R 2001 Effects of perceived danger on flower choice by bees Ecol Lett 4 327ndash333
Dukas R and D H Morse 2003 Crab spiders affect flower visitation by bees Oikos 101 157ndash163
Eliassen S E C Jargensen M Mangel and J Garl 2007 Exploration or exploitation life expectancy changes the value of learning in foraging strategies Oikos 116 513ndash523
Fernandez-Juricic E B Kerr P Bednekoff and D Stephens 2004 When are two heads better than one Visual perception and informa-tion transfer affect vigilance coordination in foraging groups Behav Ecol 6 898ndash906
Foster W A and J E Treherne 1981 Evidence for the dilution effect in the selfish herd from fish predation on a marine insect Nature 293 466ndash467
Fournier V J Hagler K Daane J de Leoacuten and R Groves 2008 Iden-tifying the predator complex of Homalodisca vitripennis (Hemiptera Cicadellidae) a comparative study of the efficacy of an ELISA and PCR gut content bioassay Oecologia 157 629ndash640
Giraldeau L T J Valone and J J Templeton 2002 Potential disadvan-tages of using socially acquired information Phil Trans R Soc Lond B 357 1559ndash1566
Hamiliton WD 1971 Geometry for the selfish herd J Theror Biol 31295-311
Hairston N G J E Smith and L B Slobodkin 1960 Community struc-ture population control and competition Am Midl Nat 106 249ndash257
Hunter W B and D E Ullman 1994 Precibarial and cibarial chemosen-sillae in the western flower thrips Frankliniella occidentalis (Pergande)(Thysanoptera Thripidae) Int J Insect Morphol Embryol 23 69ndash83
Hunter W B E Hiebert S Webb J E Polston and J H Tsai 1996 Precibarial and cibarial chemosensilla in the whitefly Bemisia tabaci
(Gennadius) (Homoptera Aleyrodidae) Int J Insect Morphol Embryol 25 295ndash304
Jeffries M J and J H Lawton 1984 Enemy-free space and the structure of ecological communities Biol J Linn Soc 23 269-286
Karban R and L T Baldwin 1997 Induced response to herbivory University of Chicago Press Chicago
Land M F and T S Collett 1974 Chasing behavior of houseflies (Fannia canicularis) J Comp Physiol 80 331ndash357
Lawrence E S 1985 Vigilance during ldquoeasyrdquo and ldquodifficultrdquo foraging tasks Anim Behav 33 1373ndash1375
Lawton J H and S McNeil1979 Between the devil and the deep blue sea on the problem of being a herbivore pp 223ndash244 In R M Anderson B D Turner and L R Taylor [Eds] Population dynamics Blackwell Oxford England Blackwell Sci Publ 434 pp
Lazarus J 1979 Flock size and behavior in captive red-billed weaverbirds (Quelea quelea) implications for social facilitation and the functions of flocking Behavior 71 1ndash2
Lietze V R M Mizell III and D G Boucias 2011 Transmission of the mycopathogen Hirsutella spp to nymphs and adults of the glassy-winged sharpshooters Homalodisca vitripennis (=coagulata) in the greenhouse Fla Entomol 94 106-108
Lopez R F R Mizell III P C Andersen and B V Brodbeck 2003 Impact of natural enemies on populations of the glassy-winged sharpshooter Homalodisca coagulata (Say) in north Florida In R VanDriesche [Ed] First international symposium on biological control of arthropods US Dep Agric For Ser Pub FHTET-03-05 484ndash486
Mainman G A D Straw and M H Dickson 2008 A simple vision-based algorithm for decision making in flying Drosophila Curr Biol 18 464ndash470
McLinn C and D W Stephens 2006 What makes information valu-able signal reliability and environmental uncertainty Anim Behav 71 1119ndash1129
Mizell R F III and W J French 1987 Leafhopper vectors of phony peach disease feeding site preference and survival on infected and uninfected peach and seasonal response to selected host plants J Entomol Sci 22 1122
Mizell R F III P C Andersen P C Phillips and P Oevering 2005 Population dynamics and interactions between the glassy-winged sharpshooter and the host plants in response to California phenology pp 117ndash121 In Proceedings 2005 Piercersquos Disease Research Symposium California Department of Food and Agriculture Sacramento
Mizell R F III C Tipping P C Andersen B V Brodbeck T Northfield and W Hunter W 2008 Behavioral model for the glassy-winged sharp-shooter Homalodisca vitripennis (Hemiptera Cicadellidae) optimiza-tion of host plant utilization and management implications Environ Entomol 37 1049ndash1062
Mizell R F III PC Andersen and B V Brodbeck 2009 Improved detec-tion monitoring and management of the glassy-winged sharpshooter pp 40ndash43 In Proceedings 2009 Piercersquos Disease Research Symposium California Department of Food and Agriculture Sacramento
Muller KL 1998 The role of conspecifics in habitat settlement in a ter-ritorial grasshopper Anim Behav 56 479ndash485
Ninemets Uuml M Riechstein M Staudt G Siefert and J Tenhunen 2002 Stomatal constraints may affect emission of oxygenated monoterpenoids from foliage of Pinus pinea Plant Physiol 130 1371ndash1385
Ninemets Uuml F Loreto and M Riechstein 2004 Physiological and physiochemical controls on foliar volatile organic compound emissions Trends Plant Sci 9 180ndash186
Northfield T R F Mizell III P C Andersen B V Brodbeck T C Riddle and W B Hunter 2009 Dispersal patch leaving and aggregation of the glassy-winged sharpshooter Homalodisca vitripennis (Hemiptera Cicadellidae) Environ Entomol 38 183ndash191
Patt J and M Setamou 2007 Olfactory and visual stimuli affecting host plant detection in Homalodisca coagulata (Hemiptera Cicadellidae) Environ Entomol 36 142ndash150
Patt J and M Setamou 2010 Recognition of novel volatile cues by the nymph of the glassy-winged sharpshooter Homalodisca vitripennis (Cicadellidae) J Insect Behav DOI 101007s10905-010-9214-z 23 290-302
Percy D M E A Boyd and M S Hoddle 2008 Observations of acoustical signaling in three sharpshooters Homalodisca vitripennis Homalodisca
115American Entomologist bull Volume 58 Number 2
liturata and Graphocephala atropunctata (Hemiptera Cicadellidae) Ann Entomol Soc 101 253ndash259
Power M E 1992 Top-down and bottom-up forces in food webs do plants have primacy Ecologia 73 733ndash746
Price P W C E Bouton P Gross B A McPheron J N Thompson and A E Wise 1980 Interactions among three trophic levels influence of plants on interactions between insect herbivores and natural enemies Annu Rev Ecol Syst 11 41ndash66
Redak R M J Blua K L Lopes A H Purcell R F Mizell III and P C Andersen 2004 The biology and control of xylem sap-feeding insect vectors of Xylella fastidiosa and their relation to disease epidemiology Annu Rev Entomol 49 243ndash270
Rolf D A 1992 The evolution of life history traits Chapman amp Hall NYSandlin E A 2000 Cue use affects resource subdivision among three
coexisting hummingbird species Behav Ecol 11 550-559SAS Institute 2009 Proc GLM SAS 93 Statistical Analysis System Cary NCShrewsbury P and M Raupp 2008 Do top-down or bottom-up forces
determine Stephanitis pyroides abundance in urban landscapes Ecol Appl 16 262ndash273
Systat Software 2009 SigmaPlot for Windows 110 Systat Software Bangalore India
Stamps J A 1987 Conspecifics as cues to territory quality a preference of juvenile lizards (Anolis aeneus) for previously used territories Am Nat 129 629ndash642
Stamps J A 1988 Conspecific attraction and aggregation in territorial species Am Nat 131 330ndash347
Thompson J D S C Peterson and L D Harder 1987 Response to traplining bumble bees to competition experiments shifts in feeding location and efficiency Oecologia 71 295ndash300
Tipping C R F Mizell III and P C Andersen 2004 Adaptations impor-tant for the dispersal of immature stages of three species of leafhopper (Homoptera Cicadellidae) Fla Entomol 87 372ndash379
Triapitsyn S V R F Mizell III J L Bossert and J E Carlton 1998 Egg parasitoids of glassy-winged sharpshooter Homalodisca coagulata (Homoptera Cicadellidae) Fla Entomol 81 243ndash251
Turner W F and H N Pollard 1959 Life histories and behavior of five insect vectors of phony peach disease Tech Bull US Dep Agric 1188
Turchin P 1987 The role of aggregation in the response of Mexican bean beetles to host-plant density Oecologia 71 577ndash582
Valone T J 1989 Group foraging public information and patch simula-tion Oikos 56 357ndash363
Walker M and T H Jones 2001 Relative roles of top-down and bottom-up forces in terrestrial tritrophic plant-insect herbivore-natural enemy systems Oikos 93 177ndash187
Ward A J W D J T Sumpter I D Couzin P J B Hart and J Krause 2008 Quorum decision-making facilitates information transfer in fish shoals PNAS 105 6948ndash6953
Wells J M B C Raju H Y Hung W G Weisburg L Mandelco-Paul and D J Brenner 1987 Xylella fastidiosa gen nov sp nov gram-negative xylem-limited fastidious plant bacteria related to Xanthomonas spp Int J Syst Bacteriol 37 136ndash143
Werner E E 1992 Individual behavior and higher-order species interac-tions Am Nat (suppl) 140 S5ndashS32
White T R C 1978 The importance of relative shortage of food in animal ecology Oecologia 33 71ndash86
Williams G C 1966 Adaptation and natural selection Princeton University Press Princeton NJ
Wu D S C Daugherty S E Van Aken G H Pai K L Watkins H Khouri L J Tallon J M Zaborsky H E Dunbar P L Tran N A Moran J A Eisen 2006 Metabolic complementarity and genomics of the dual symbiosis of sharpshooters PloS-Biology 4 e188
Russell F Mizell III Is a Professor of Entomology (rfmizellufledu) Peter C Andersen is a Professor of Horticulture (peaufledu) and Brent V Brod-beck (bvbufledu) is a Biological Scientist all located at the University of Floridarsquos North Florida Research and Education Center-Quincy 155 Research Rd Quincy Fl 32351 Wayne B Hunter (whunterushrlarsusdagov) is a research entomologist with the USDA Agricultural Research Service at the US Horticultural Research Lab 2001 South Rock Rd Ft Pierce FL 34945 For over 20 years they have collaborated in research on the biology ecology and behavior of the leafhopper vectors of Xylella fastidiosa
With our high qualityLepidoptera Eggs and LarvaeBeet Army Worm bull Cabbage LooperCorn Earworm bull Diamondback MothFall Armyworm bull Tobacco Budworm
Save Money Save Time Get Results
Lepidoptera Eggs and LarvaeInsect Diets and Ingredients
Rearing Supplies and Equipment
Entomology Division
800-996-9908 US and Canada908-996-2155 Internationalsalesinsectrearingcom
wwwinsectrearingcom

115American Entomologist bull Volume 58 Number 2
liturata and Graphocephala atropunctata (Hemiptera Cicadellidae) Ann Entomol Soc 101 253ndash259
Power M E 1992 Top-down and bottom-up forces in food webs do plants have primacy Ecologia 73 733ndash746
Price P W C E Bouton P Gross B A McPheron J N Thompson and A E Wise 1980 Interactions among three trophic levels influence of plants on interactions between insect herbivores and natural enemies Annu Rev Ecol Syst 11 41ndash66
Redak R M J Blua K L Lopes A H Purcell R F Mizell III and P C Andersen 2004 The biology and control of xylem sap-feeding insect vectors of Xylella fastidiosa and their relation to disease epidemiology Annu Rev Entomol 49 243ndash270
Rolf D A 1992 The evolution of life history traits Chapman amp Hall NYSandlin E A 2000 Cue use affects resource subdivision among three
coexisting hummingbird species Behav Ecol 11 550-559SAS Institute 2009 Proc GLM SAS 93 Statistical Analysis System Cary NCShrewsbury P and M Raupp 2008 Do top-down or bottom-up forces
determine Stephanitis pyroides abundance in urban landscapes Ecol Appl 16 262ndash273
Systat Software 2009 SigmaPlot for Windows 110 Systat Software Bangalore India
Stamps J A 1987 Conspecifics as cues to territory quality a preference of juvenile lizards (Anolis aeneus) for previously used territories Am Nat 129 629ndash642
Stamps J A 1988 Conspecific attraction and aggregation in territorial species Am Nat 131 330ndash347
Thompson J D S C Peterson and L D Harder 1987 Response to traplining bumble bees to competition experiments shifts in feeding location and efficiency Oecologia 71 295ndash300
Tipping C R F Mizell III and P C Andersen 2004 Adaptations impor-tant for the dispersal of immature stages of three species of leafhopper (Homoptera Cicadellidae) Fla Entomol 87 372ndash379
Triapitsyn S V R F Mizell III J L Bossert and J E Carlton 1998 Egg parasitoids of glassy-winged sharpshooter Homalodisca coagulata (Homoptera Cicadellidae) Fla Entomol 81 243ndash251
Turner W F and H N Pollard 1959 Life histories and behavior of five insect vectors of phony peach disease Tech Bull US Dep Agric 1188
Turchin P 1987 The role of aggregation in the response of Mexican bean beetles to host-plant density Oecologia 71 577ndash582
Valone T J 1989 Group foraging public information and patch simula-tion Oikos 56 357ndash363
Walker M and T H Jones 2001 Relative roles of top-down and bottom-up forces in terrestrial tritrophic plant-insect herbivore-natural enemy systems Oikos 93 177ndash187
Ward A J W D J T Sumpter I D Couzin P J B Hart and J Krause 2008 Quorum decision-making facilitates information transfer in fish shoals PNAS 105 6948ndash6953
Wells J M B C Raju H Y Hung W G Weisburg L Mandelco-Paul and D J Brenner 1987 Xylella fastidiosa gen nov sp nov gram-negative xylem-limited fastidious plant bacteria related to Xanthomonas spp Int J Syst Bacteriol 37 136ndash143
Werner E E 1992 Individual behavior and higher-order species interac-tions Am Nat (suppl) 140 S5ndashS32
White T R C 1978 The importance of relative shortage of food in animal ecology Oecologia 33 71ndash86
Williams G C 1966 Adaptation and natural selection Princeton University Press Princeton NJ
Wu D S C Daugherty S E Van Aken G H Pai K L Watkins H Khouri L J Tallon J M Zaborsky H E Dunbar P L Tran N A Moran J A Eisen 2006 Metabolic complementarity and genomics of the dual symbiosis of sharpshooters PloS-Biology 4 e188
Russell F Mizell III Is a Professor of Entomology (rfmizellufledu) Peter C Andersen is a Professor of Horticulture (peaufledu) and Brent V Brod-beck (bvbufledu) is a Biological Scientist all located at the University of Floridarsquos North Florida Research and Education Center-Quincy 155 Research Rd Quincy Fl 32351 Wayne B Hunter (whunterushrlarsusdagov) is a research entomologist with the USDA Agricultural Research Service at the US Horticultural Research Lab 2001 South Rock Rd Ft Pierce FL 34945 For over 20 years they have collaborated in research on the biology ecology and behavior of the leafhopper vectors of Xylella fastidiosa
With our high qualityLepidoptera Eggs and LarvaeBeet Army Worm bull Cabbage LooperCorn Earworm bull Diamondback MothFall Armyworm bull Tobacco Budworm
Save Money Save Time Get Results
Lepidoptera Eggs and LarvaeInsect Diets and Ingredients
Rearing Supplies and Equipment
Entomology Division
800-996-9908 US and Canada908-996-2155 Internationalsalesinsectrearingcom
wwwinsectrearingcom
Copyright © 2022 FDOKUMEN




















