
Computational Simulation of Platelet Deposition and Activation: I. Model Development and Properties
10
Computational Simulation of Platelet Deposition and Activation: II. Results for Poiseuille Flow over Collagen ERIK N. SORENSEN, 1 GREG W. BURGREEN, 1,2 WILLIAM R. WAGNER, 1,2 and JAMES F. ANTAKI 1,2 Departments of 1 Bioengineering and 2 Surgery, McGowan Center for Artificial Organ Development, University of Pittsburgh, Pittsburgh, PA (Received 12 August 1998; accepted 3 June 1999) Abstract—We have previously described the development of a two-dimensional computational model of platelet deposition onto biomaterials from flowing blood ~Sorensen et al., Ann. Biomed. Eng. 27:436–448, 1999!. The model requires estima- tion of four parameters to fit it to experimental data: shear- dependent platelet diffusivity and three platelet-deposition- related reaction rate constants. These parameters are estimated for platelet deposition onto a collagen substrate for simple parallel-plate flow of whole blood in both the presence and absence of thrombin. One set of experimental results is used as a benchmark for model-fitting purposes. The ‘‘trained’’ model is then validated by applying it to additional test cases from the literature for parallel-plate Poiseuille flow over collagen at both higher and lower wall shear rates, and in the presence of vari- ous anticoagulants. The predicted values agree very well with the experimental results for the training cases, and good repro- duction of deposition trends and magnitudes is obtained for the heparin, but not the citrate, validation cases. The model is formulated to be easily extended to synthetic biomaterials, as well as to more complex flows. © 1999 Biomedical Engineer- ing Society. @S0090-6964~99!01704-X# Keywords—Thrombosis, Mathematical model, Thrombin, Fluid dynamics, Mass transport, Platelets. INTRODUCTION We have developed a two-dimensional computational model of platelet-mediated thrombogenesis on biomate- rials which was described in Part I. 11 The model, based on continuum assumptions for blood, is comprised of a system of convection-diffusion-reaction equations. It at- tempts to simulate the following events occurring in blood under varying flow conditions: activated and rest- ing platelet-biomaterial adhesion; platelet-platelet aggre- gation at surfaces; generation of platelet-released and platelet-synthesized agonists ~adenosine diphosphate ~ADP! and thromboxane A 2 @ TxA 2 # , respectively!; platelet-phospholipid-dependent thrombin generation; and agonist-induced platelet activation. In Part I, the structure and underlying assumptions of the model were outlined, and its sensitivity to the parameters that would have to be varied to simulate different experimental con- ditions was investigated. In this article, we attempt to validate the model against experimental results, using simple test cases from the literature involving parallel- plate Poiseuille flow of whole blood over a type I col- lagen substrate. One set of training cases is used to estimate the variable model parameters, and then the model is applied to additional validation cases to inves- tigate its robustness under varying fluid shear rates and anticoagulation conditions. METHODS Briefly, the model consists of seven coupled convection-diffusion-reaction equations, one for each simulated species @resting platelets, activated platelets, ADP, TxA 2 , prothrombin, thrombin, and antithrombin III ~ATIII!#. The form of the convection-diffusion- reaction equations employed for each species is ] @ C i # ] t 1div~ u• @ C i #! 5div~ D i • grad@ C i #! 1S i . ~1! Here, div is the divergence operator and grad is the gradient operator. D i refers to the diffusivity of species i in blood ~in cm 2 s 21 !; u is the two-dimensional fluid velocity vector ~in cm s 21 !; @ C i # is the concentration of species i; and S i is a source term for species i. Species- specific events such as agonist generation and platelet activation are implemented in the source terms, while platelet-surface interactions are simulated via surface- flux boundary conditions. The functional forms of all the relevant terms, as well as details of the computational methods employed to solve the discrete equations, can be found in Part I. 11 Additionally, the flow field, neglecting thrombus growth, is obtained by solving the steady-state Navier- Address all correspondence to Erik Sorensen, University of Pitts- burgh, 300 Technology Drive, Room 439 Biotech Center, Pittsburgh, PA 15219. Electronic mail: ensst61@pitt.edu Annals of Biomedical Engineering, Vol. 27, pp. 449–458, 1999 0090-6964/99/27~4!/449/10/$15.00 Printed in the USA. All rights reserved. Copyright © 1999 Biomedical Engineering Society 449
Transcript of Computational Simulation of Platelet Deposition and Activation: I. Model Development and Properties

Annals of Biomedical Engineering,Vol. 27, pp. 449–458, 1999 0090-6964/99/27~4!/449/10/$15.00Printed in the USA. All rights reserved. Copyright © 1999 Biomedical Engineering Society
Computational Simulation of Platelet Deposition and Activation: II.Results for Poiseuille Flow over Collagen
ERIK N. SORENSEN,1 GREG W. BURGREEN,1,2 WILLIAM R. WAGNER,1,2 and JAMES F. ANTAKI 1,2
Departments of1Bioengineering and2Surgery, McGowan Center for Artificial Organ Development, University of Pittsburgh,Pittsburgh, PA
(Received 12 August 1998; accepted 3 June 1999)
f aion
-ar-on-
atplendd aelthethari-ith
prothel is, as-
n,
nalte-
f aat-inst-re-ande
on;he
ereuldn-toingel-l-
tothes-
and
dchs,n-
teletilece-enaln be
ser-
itts-gh,
Abstract—We have previously described the development otwo-dimensional computational model of platelet depositonto biomaterials from flowing blood~Sorensenet al., Ann.Biomed. Eng.27:436–448, 1999!. The model requires estimation of four parameters to fit it to experimental data: shedependent platelet diffusivity and three platelet-depositirelated reaction rate constants. These parameters are estimfor platelet deposition onto a collagen substrate for simparallel-plate flow of whole blood in both the presence aabsence of thrombin. One set of experimental results is usea benchmark for model-fitting purposes. The ‘‘trained’’ modis then validated by applying it to additional test cases fromliterature for parallel-plate Poiseuille flow over collagen at bohigher and lower wall shear rates, and in the presence of vous anticoagulants. The predicted values agree very well wthe experimental results for the training cases, and good reduction of deposition trends and magnitudes is obtained forheparin, but not the citrate, validation cases. The modeformulated to be easily extended to synthetic biomaterialswell as to more complex flows. ©1999 Biomedical Engineering Society.@S0090-6964~99!01704-X#
Keywords—Thrombosis, Mathematical model, ThrombiFluid dynamics, Mass transport, Platelets.
INTRODUCTION
We have developed a two-dimensional computatiomodel of platelet-mediated thrombogenesis on biomarials which was described in Part I.11 The model, basedon continuum assumptions for blood, is comprised osystem of convection-diffusion-reaction equations. Ittempts to simulate the following events occurringblood under varying flow conditions: activated and reing platelet-biomaterial adhesion; platelet-platelet agggation at surfaces; generation of platelet-releasedplatelet-synthesized agonists~adenosine diphosphat~ADP! and thromboxane A2 @TxA2#, respectively!;platelet-phospholipid-dependent thrombin generatiand agonist-induced platelet activation. In Part I, t
Address all correspondence to Erik Sorensen, University of Pburgh, 300 Technology Drive, Room 439 Biotech Center, PittsburPA 15219. Electronic mail: [email protected]
449
ed
s
-
structure and underlying assumptions of the model woutlined, and its sensitivity to the parameters that wohave to be varied to simulate different experimental coditions was investigated. In this article, we attemptvalidate the model against experimental results, ussimple test cases from the literature involving parallplate Poiseuille flow of whole blood over a type I colagen substrate. One set of training cases is usedestimate the variable model parameters, and thenmodel is applied to additional validation cases to invetigate its robustness under varying fluid shear ratesanticoagulation conditions.
METHODS
Briefly, the model consists of seven coupleconvection-diffusion-reaction equations, one for easimulated species@resting platelets, activated plateletADP, TxA2, prothrombin, thrombin, and antithrombiIII ~ATIII !#. The form of the convection-diffusionreaction equations employed for each species is
]@Ci #
]t1div~u•@Ci # !5div~Di•grad@Ci # !1Si . ~1!
Here, div is the divergence operator andgrad is thegradient operator.Di refers to the diffusivity of speciesiin blood ~in cm2 s21!; u is the two-dimensional fluidvelocity vector~in cm s21!; @Ci # is the concentration ofspeciesi; and Si is a source term for speciesi. Species-specific events such as agonist generation and plaactivation are implemented in the source terms, whplatelet-surface interactions are simulated via surfaflux boundary conditions. The functional forms of all threlevant terms, as well as details of the computatiomethods employed to solve the discrete equations, cafound in Part I.11
Additionally, the flow field, neglecting thrombugrowth, is obtained by solving the steady-state Navi

ible
lveesn,
ll
t
g-oflelel-telects
g
therein
-er
nto
n-pa-
f
ts,tiveof
tede.
2
m-pon
-e
I
in-portes-
ndac-see
a-
450 SORENSEN et al.
Stokes equations, assuming blood is an incompressNewtonian fluid:
r~u–grad„u…!52div~p!1mDu, ~2!
div„u…50. ~3!
Here, D is the Laplacian operator;r is fluid density~ing cm23!; p is pressure~in dynes cm22!; and m is theasymptotic ~high-shear! viscosity of blood ~ing cm21 s21!. Since all the cases considered herein invoparallel-plate flow, the incompressible Navier-Stokequations are exactly satisfied by the Poiseuille solutio2
which is independent of blood density and viscosity:
ux5UmaxS 12~H2y!2
H2 D ,
~4!
uy50.
Here,y is measured from the bottom of the channel,H isthe channel half height~in cm!, and Umax is the maxi-mum velocity~at the centerline! in cm s21. Note that fora Newtonian fluid,
gw5~]ux /]y!wall5~2•Umax•~H2y!/H2!y50
52•Umax/H, ~5!
and henceux can be written as a function of the washear rategw .
Model Training Cases
The variable model parameters@resting platelet-surface adhesion ratekrs ~cm s21!, activated platelet-surface adhesion ratekas ~cm s21!, activated platelet-platelet adhesion ratekaa ~cm s21!, and shear-dependenplatelet ~PLT! diffusivity DPLT ~cm s22!# were estimatedby fitting the model to the experimental results of Waner and Hubbell,13 who studied the axial dependenceplatelet deposition from flowing, heparinized whoblood onto a type I collagen surface using a paralplate flow chamber. Since these studies measured pladeposition as a function of time and examined the effeof removing thrombin @via the active-site inhibitorFPRCH2Cl ~PPACK!#, the results are ideal for calibratinour model.
Model parameters specific to these cases andequations from Part I to which they are relevant alisted in Table 1; all other parameters are as givenTables 2 and 3 of Part I.11 The resting platelet concentration was estimated by comparing Fig. 6 of Wagn
t
and Hubbell,13 which shows maximal platelet depositiofor heparinized blood as a function of platelet count,the maximum value in Fig. 1 of that report, which cotains the results we are attempting to predict. TherameterDs is the Keller correction factor7 to the Brown-ian diffusivity of the ‘‘large’’ species ~prothrombin,thrombin, and ATIII! based on the wall shear rate o1000 s21 used by Wagner and Hubbell,13 while the valuefor the rate of thrombin generation by activated platelefat , has been increased at the collagen surface relato its bulk value to account for higher observed ratesthrombin generation from activated platelets stimulaby both collagen and thrombin versus thrombin alon9
The heparin concentration listed corresponds to theU ml21 used in the experiments.~U is National Institutesof Health Units.! Finally, the value of 1.0 assumed foruimplies that all resting platelets deposited on the throbogenic type I collagen surface become activated ucontact.
The experiments of Wagner and Hubbell13 employeda 200mm high320 cm long channel, with the inlet portion ~3 cm in length! coated with relatively nonadhesivalbumin and the remaining 17 cm coated with typecollagen. Using the reported wall shear rate of 1000 s21,a Poiseuille solution for the flow field@Eqs. ~4! and ~5!#can be computed, and the resulting velocity termsserted into the transport equations. Then, the transmodel, consisting of the seven coupled speciconservation equations@Eq. ~1!#, can be solved oncesuitable boundary and initial conditions are imposed avalues for the platelet-surface and platelet-platelet retivities and platelet diffusivity are assumed. In the cawhere PPACK was used to inhibit thrombin, ththrombin-related species~prothrombin, thrombin, andATIII ! were eliminated from the model. This is equiv
TABLE 1. Specific parameter values for training cases.
Parameter Value Eq.a Ref.
[RP] 23108 PLT ml21 (1), (5), (7) 13b
C0 (for [RP]) 1.23108 PLT ml21 (19) b
Ds 1.3631025 cm2 s21 (18) 7c
[H] 0.417 mM (4) 13d
fat surface 3.2231028 U PLT21 s21 (9), (10) 9e
(mM PT)21
u 1.00 (7), (13)–(15) ¯
aRelevant equations from Part I (Ref. 11).bValues for [AP] are 5% of these values (to simulate 5% back-ground PLT activation).
cUsed to correct diffusivities of ‘‘large’’ species (thrombin, pro-thrombin, ATIII) for the effect of red blood cells in shear flow.
dFor 2 U ml21 heparin as used in Ref. 13, assuming specificactivity of 300 U mg21 and molecular weight of 16 kDa.
eSee Table 1 of Ref. 9: value for PLTs stimulated bythrombin1collagen.

tsingse
theeto
th,tes
inggo-
hetelyndofofer
is-sedionndthell’s-the
,00eteereware
ria-eeanted
re,
letdif-oth
petletap
n-oi-
ng
5
0,u-e
aridh asgthrid
.2 s
terlledasn-
wer,000ainime
omonsca-forhisuesote,
451Simulation of Platelet Deposition: II. Results
lent to assuming that PPACK completely inhibiplatelet-mediated thrombin generation. In the remainfour equations, all parameters were identical to thoused for the heparin-only case.
Our transport model was solved on a 100mm311 mmrectangular domain designed to simulate a portion oflower half of Wagner and Hubbell’s flow chamber. Thfirst 1 mm of the computational domain was designedreplicate a portion of the 3 cm albumin entrance lengwhere Wagner and Hubbell13 observed negligible plateledeposition. Accordingly, platelet-surface reactivitithere were arbitrarily set to 1028 of the values for the 10mm collagen region, and the parameteru was set to zero.This had the effect of preventing the adherent restplatelets in this region from activating and releasing anists. The results of Wagner and Hubbell13 indicate thatthe experimental platelet deposition curves have reacan essentially constant, asymptotic value approxima10 mm into the collagen portion of the chamber, athose authors report data for only the first 15 mmcollagen. Therefore, we chose 10 mm for the lengththe collagen portion of our computational domain, raththan the full 17 cm used in the experiments.
Initial and boundary conditions were imposed as dcussed in Part I with constant inlet concentrations ufor all species, a wall-skewed quadratic concentratprofile employed for resting and activated platelets, a5% background platelet activation assumed. Sinceupper wall was collagen-coated in Wagner and Hubbeexperiments, the 100mm upper plane of our computational domain was treated as a symmetry plane, andboundary condition (]@Ci #/]y)y5100mm50 was imposedon all species.
The mesh used for these cases was the same 10node unstructured triangular mesh used for the paramsensitivity studies in Part I; mesh refinement studies wconducted as described therein. Since time accuracyan important consideration for these cases, time-stepfinement studies were also conducted. Negligible vation in root-mean-square species concentrations betwsimulations was found for physical time steps less th2.5 s for a 75 s simulation. For the results presenherein, a time step of 0.5 s was used.
Validation Cases
Two sets of experimental results from the literatuthose of Hubbell and McIntire6 and of Alevriadouet al.,1
were selected to validate the model after the platereaction rate constants for collagen and the plateletfusivity were estimated using the training cases. In bstudies, parallel-plate flow chambers with 200mm slitwidths were used, human blood was perfused, and tycollagen was the substrate. Thus, the same inlet, ousymmetry-plane, and surface boundary conditions as
d
0r
s-
n
I,-
plied for the training cases were used, with inlet concetration magnitudes altered appropriately, and the Pseuille flow field was recomputed using the prevailiwall shear rate for the study being simulated.
Hubbell and McIntire6 measured deposition over 2mm of collagen after 120 s of flow of heparinized~10U ml21! whole human blood at wall shear rates of 10500, and 1500 s21; the two lower shear rates were simlated here. The 100 s21 wall shear rate is very near thlower limit for Newtonian blood behavior;8 however, asa first approximation, we have continued to employPoiseuille flow field for this case. For both cases, a gwith the same mesh density and 1 mm entrance lengtthat used in the previous cases, but with the total lenextended to 26 mm, was constructed. This larger gcontained 24,000 mesh points, and a time step of 1was used for the model solution. Alevriadouet al.1 mea-sured platelet deposition along 10 mm of collagen af60 s of flow of citrated whole human blood at a washear rate of 1500 s21. By the same reasoning as for thheparin1PPACK training case, the thrombin-relateequations were not solved for this case, since it wassumed that citrate completely inhibited thrombin geeration. Additionally, since the geometry of the flochamber is basically identical to that of the chambused in the training cases, the same unstructured 10node mesh was employed, with the first 1 mm agassumed to be a nonadhesive entrance length. A tstep of 0.6 s was used in the simulations.
Table 2 lists the parameters which were altered frthe training cases to match the experimental conditifor the validation cases. Neither study gave any indition of average platelet counts in the perfusateswhich platelet deposition results were reported. For treason, the concentrations given in Table 2 are valthat gave the best fits to the experimental results. N
TABLE 2. Parameters altered for validation cases. „N/A5notapplicable for this case. …
ParameterAlevriadou et al.
(Ref. 1)Hubbell and McIntire
(Ref. 6)
[RP]a 2.53108 PLT ml21 2.03108 PLT ml21
C0 (for [RP])a 1.53108 PLT ml21 1.23108 PLT ml21
DsN/Ab 1.3631026 cm2 s21
(gw5100 s21);6.8031026 cm2 s21
(gw5500 s21)[H] 0.00 mM 2.09 mMc
aValues for [AP] are 5% of these values (to simulate 5% back-ground PLT activation).
b‘‘Large’’ species (thrombin, prothrombin, ATIII) eliminated fromthe model for this case due to assumption of 100% thrombininhibition by citrate.
cFor 10 U ml21 heparin as used in Ref. 6, assuming specificactivity of 300 U mg21 and molecular weight of 16 kDa.

forde
hali-for
en
andofionby
1en-n-ofusoru-gedi-andof-t.
ehe
ndsnh.si-i-sti-in
n-the
on-in,xi-
thee-e
452 SORENSEN et al.
however, that this was the only parameter fitting donethese cases; rather than estimating the shear-depenplatelet diffusivity, we assumed it to vary linearly witthe wall shear rate, so that the values used for the vdation cases were interpolated from the best-fit valuethe 1000 s21 wall shear rate of the training cases, givbelow.
RESULTS
Training Cases
The experimental results presented by WagnerHubbell13 and our model solutions correspond to 75 sflow. The platelet-surface and platelet-platelet reactrate constants and platelet diffusivity were adjusted
FIGURE 1. Comparison of axial platelet deposition on col-lagen as predicted by the model vs the experimental resultsof Wagner and Hubbell „Ref. 13… for whole blood with 2U ml21 heparin alone „heparin only … and with the thrombinactive-site inhibitor PPACK „Heparin 1PPACK … after 75 s offlow.
nt
trial and error to yield the best fit to the data. Figurecompares our computational results with the experimtal data;x50 corresponds to the start of the collagecoated region. Figure 1 shows the axial distributiontotal platelet deposition, i.e., platelet-surface plplatelet-platelet adhesion, for heparin onlyheparin1PPACK as the anticoagulant. In Fig. 1, the nmerically predicted deposition values have been averaover 200mm intervals. This was done to give approxmately the same number of data points as in WagnerHubbell’s results and to mimic the averaging effectthe 43104 mm2 field of view of the fluorescent microscope used to track platelet deposition in that repor13
Values of kaa5kas54.631023 cm s21, krs53.731023 cm s21, and DPLT51050•Db,PLT51.6631026 cm2 s21 for resting and activated platelets gavthe best agreement with both experimental curves. Tcomputational results reproduce the experimental trein total deposition very well, with the no-thrombi(heparin1PPACK) results an especially close matcHowever, the heparin-only model underpredicts depotion far downstream of the inlet while slightly overestmating values near the inlet. Since no uncertainty emates were given for the platelet deposition valuesWagner and Hubbell’s report,13 it is unclear whether ourresults fall within the range of experimental error.
Figure 2 shows the time-varying, platelet-activatiothreshold-normalized concentration of each agonist atrepresentative pointx52.5 mm, y50 mm. The thrombinconcentration continues to increase with time due to ctinued platelet deposition. For the case with thrombthe ADP concentration reaches a plateau by appromately 5 s and remains there throughout the rest ofsimulation, likely due to its release by activating platlets. While the TxA2 concentration continues to increas
FIGURE 2. Predicted normalized agonist concentrations for the training cases vs time at a point on the wall „y 50… 2.5 mmdownstream of the beginning of the collagen surface. Normalized concentration 5agonist concentration/agonist’s platelet acti-vation threshold concentration „see Table 2 of Part I for the thresholds used …. „A… Model with thrombin. „B… Model withoutthrombin.

453Simulation of Platelet Deposition: II. Results
FIGURE 3. Predicted axial distribution of normalized agonist concentrations for the training case with thrombin at 75 s.Normalized concentration 5agonist concentration/agonist’s platelet activation threshold concentration „see Table 2 of Part I forthe thresholds used …. „A… Concentrations at the wall. Inset: Magnified detail of ADP and TxA 2 trends. „B… Concentrations 25 mmfrom the wall.
tsheibituede
d thg aofy
M,let
s-wsxial
inan
i-xi-P
ter
are
rsetothealllk,
m.-
e,
t aen-ulkanandelsrinn-
u-
4.dlelelyodee
,
ea-
n-nlo-thenmi-n-ti-
ely
ation,
due to platelet activation, its level is well below iactivation threshold throughout the simulation. For tcase where PPACK was used to completely inhthrombin, the ADP concentration increases initially, dto surface-induced agonist release, then begins tocrease rapidly as surface saturation is approached anbulk-released agonists begin to wash away, reachinrelatively negligible value after approximately 30 sflow. The normalized TxA2 concentration remains verlow throughout. The maximum concentration of TxA2
observed in all simulations was approximately 320 nslightly more than half its assumed threshold for plateactivation.
The effects of flow and inhibitors on the spatial ditribution of agonists are shown in Fig. 3. Figure 3 shonormalized agonist concentrations as a function of aposition at two lateral stations, the wall (y50) and y525mm, after 75 s of flow. Results for the no-thrombcase have been omitted because the normalized ADPTxA2 concentrations are negligible at this time@see Fig.2~B!#. At the wall, it is clear that thrombin is the domnant agonist, with concentrations ranging from appromately 40- to 600-fold greater than threshold. The ADconcentrations range from about five- to eightfold greathan threshold, while the levels of TxA2, when ex-pressed in terms of its platelet activation potential,about half the threshold level. At a distance of 25mmfrom the wall, the shape of the ADP curve is the reveof that at the wall: it is increasing axially comparedthe steady decrease at the wall. This is likely due toincreased convection at this location relative to the wsince ADP released from platelets activated in the buat this location will be more rapidly washed downstreaThe levels of TxA2 are negligible. Relative to their respective values at the wall, the level of thrombin aty525mm drops approximately two orders of magnitud
-e
d
whereas the level of ADP decreases by only aboufactor of 4. Some of the decrease in thrombin conctration is because the rate of thrombin generation on bactivated platelets is an order of magnitude lower ththat selected for platelets activated by both agonistscollagen at the wall. However, given the disparate levof the two agonists at the wall, it is apparent that hepahas a very noticeable effect on the bulk thrombin cocentration. This inhibitory effect was confirmed by nmerical experiments conducted without heparin~data notshown!. In these simulations, the ADP and TxA2 con-centrations were similar to the values shown in Fig.However, without heparin, the maximum normalizethrombin concentration at the wall was roughly doubthe maximum value in the simulation with heparin, whiat y525mm, the maximum normalized value was neartwo orders of magnitude greater than the value for blowith 2 U ml21 heparin. We note that these results agrqualitatively with the simulations of Folie and McIntire5
who considered four agonists~thrombin, ADP, TxA2,and the von Willebrand factor! and heparin inhibition.
The axial distributions of activated platelets for thmodel with and without thrombin at the same two loctions as above~wall, y50, andy525mm! are shown inFig. 4. At the wall, trends in activated platelet concetration for the simulation with thrombin mirror trends iagonist concentration, with more activated plateletscated where agonist concentrations are higher. Inbulk flow (y525mm), activated platelet concentratioincreases with downstream distance due to the concotant rise in bulk agonists. Despite the much higher cocentrations of agonists at the wall, the maximum acvated platelet concentration there is only approximatthree times the maximum concentration aty525mm.This is due to rapid consumption of activated plateletsthe wall by platelet-surface and platelet-platelet adhes

454 SORENSEN et al.
FIGURE 4. Predicted axial distribution of activated platelets for training cases at 75 s. Closed squares, concentration at the wall;open diamonds, concentration 25 mm from the wall. „A… Model with thrombin. „B… Model without thrombin.
off
cee.am
the
letnd
n
6
xi-en-ell,esn-
ed,ed,ith
letia-
00
as modeled by thekaa and kas terms. For the no-thrombin case, the concentration at the wall fallsrapidly due to deposition, while aty525mm the con-centration varies only slightly with axial distance sinlittle or no bulk activation is taking place in this casThe slight decrease in concentration with downstredistance aty525mm is likely due to diffusion of plate-lets toward the wall to replace those consumed bysurface.
Validation Cases
Figure 5 shows model predictions of total platedeposition versus the experimental data of Hubbell aMcIntire6 for 120 s of flow of heparinized whole humablood at wall shear rates of 100 and 500 s21, for whichthe linearly interpolated platelet diffusivities were 1.6
31027 and 8.3331027 cm2 s21, respectively. Again, themodel-predicted data were averaged to give appromately the same number of points as for the experimtal data. Forgw5100 s21, the model predicts the shapof the deposition curve and its magnitudes fairly wealthough, similar to the heparin-only training case, valunear the inlet are slightly overestimated, while dowstream values are underestimated. Forgw5500 s21, onthe other hand, the shape of the curve is well predictbut the amount of deposition is generally overpredictwith the error highest near the inlet and decreasing wdownstream distance.
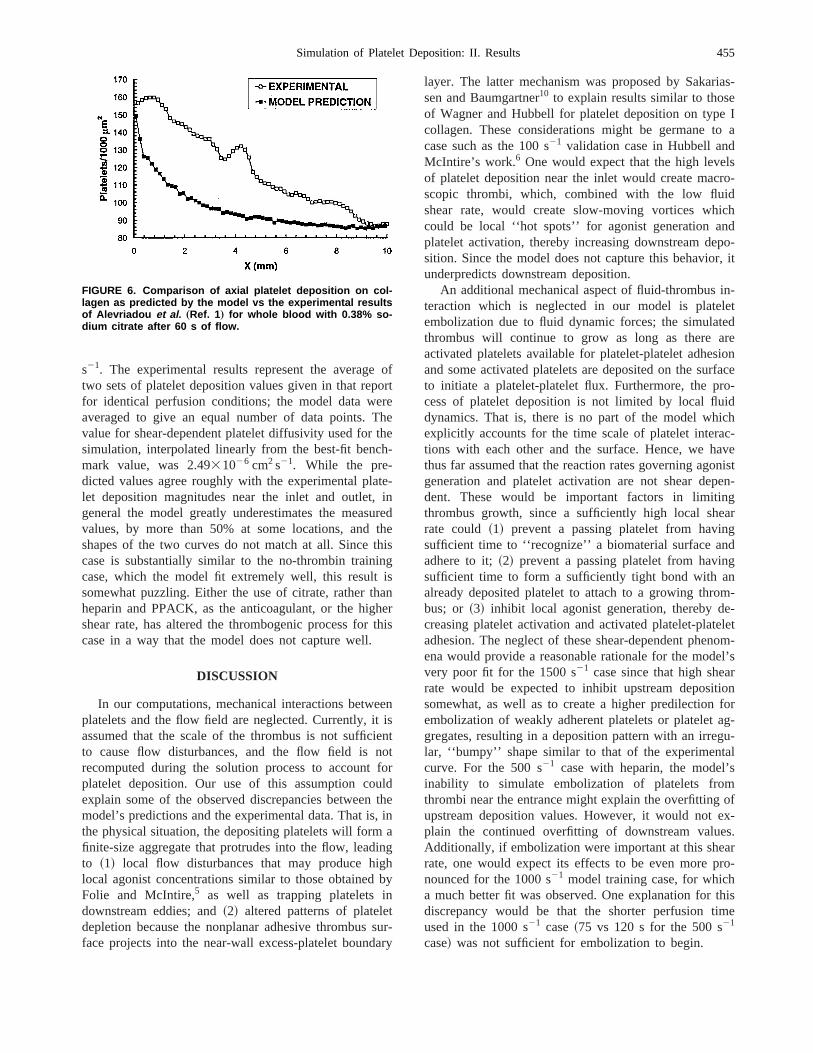
Figure 6 compares the model’s predicted total platedeposition values to the experimental results of Alevrdou et al.1 for 60 s of flow of whole human blood with0.38% wt/vol sodium citrate at a wall shear rate of 15
FIGURE 5. Comparison of axial platelet deposition on collagen as predicted by the model vs the experimental results of Hubbelland McIntire „Ref. 6… for whole blood with 10 U ml 21 heparin after 120 s of flow. „A… Wall shear rate 5100 s21. „B… Wall shearrate5500 s21.
oortrehe
theh-
te-in
urethethisngisanherthi
enis
ienotfor
uldth
, ina
ingghby
ntsuary
ias-e
Io a
lsro-dchdpo-r, it
n-letedreionfaceo-idchc-avenisten-gargdgnm-e-eletom-
el’sr
ionforg-
gu-l
s
ofx-s.
arro-
hisme
455Simulation of Platelet Deposition: II. Results
s21. The experimental results represent the averagetwo sets of platelet deposition values given in that repfor identical perfusion conditions; the model data weaveraged to give an equal number of data points. Tvalue for shear-dependent platelet diffusivity used forsimulation, interpolated linearly from the best-fit bencmark value, was 2.4931026 cm2 s21. While the pre-dicted values agree roughly with the experimental plalet deposition magnitudes near the inlet and outlet,general the model greatly underestimates the measvalues, by more than 50% at some locations, andshapes of the two curves do not match at all. Sincecase is substantially similar to the no-thrombin trainicase, which the model fit extremely well, this resultsomewhat puzzling. Either the use of citrate, rather thheparin and PPACK, as the anticoagulant, or the higshear rate, has altered the thrombogenic process forcase in a way that the model does not capture well.
DISCUSSION
In our computations, mechanical interactions betweplatelets and the flow field are neglected. Currently, itassumed that the scale of the thrombus is not sufficto cause flow disturbances, and the flow field is nrecomputed during the solution process to accountplatelet deposition. Our use of this assumption coexplain some of the observed discrepancies betweenmodel’s predictions and the experimental data. That isthe physical situation, the depositing platelets will formfinite-size aggregate that protrudes into the flow, leadto ~1! local flow disturbances that may produce hilocal agonist concentrations similar to those obtainedFolie and McIntire,5 as well as trapping platelets idownstream eddies; and~2! altered patterns of plateledepletion because the nonplanar adhesive thrombusface projects into the near-wall excess-platelet bound
FIGURE 6. Comparison of axial platelet deposition on col-lagen as predicted by the model vs the experimental resultsof Alevriadou et al. „Ref. 1… for whole blood with 0.38% so-dium citrate after 60 s of flow.
f
d
s
t
e
r-
layer. The latter mechanism was proposed by Sakarsen and Baumgartner10 to explain results similar to thosof Wagner and Hubbell for platelet deposition on typecollagen. These considerations might be germane tcase such as the 100 s21 validation case in Hubbell andMcIntire’s work.6 One would expect that the high leveof platelet deposition near the inlet would create macscopic thrombi, which, combined with the low fluishear rate, would create slow-moving vortices whicould be local ‘‘hot spots’’ for agonist generation anplatelet activation, thereby increasing downstream desition. Since the model does not capture this behaviounderpredicts downstream deposition.
An additional mechanical aspect of fluid-thrombus iteraction which is neglected in our model is plateembolization due to fluid dynamic forces; the simulatthrombus will continue to grow as long as there aactivated platelets available for platelet-platelet adhesand some activated platelets are deposited on the surto initiate a platelet-platelet flux. Furthermore, the prcess of platelet deposition is not limited by local fludynamics. That is, there is no part of the model whiexplicitly accounts for the time scale of platelet interations with each other and the surface. Hence, we hthus far assumed that the reaction rates governing agogeneration and platelet activation are not shear depdent. These would be important factors in limitinthrombus growth, since a sufficiently high local sherate could ~1! prevent a passing platelet from havinsufficient time to ‘‘recognize’’ a biomaterial surface anadhere to it;~2! prevent a passing platelet from havinsufficient time to form a sufficiently tight bond with aalready deposited platelet to attach to a growing throbus; or ~3! inhibit local agonist generation, thereby dcreasing platelet activation and activated platelet-platadhesion. The neglect of these shear-dependent phenena would provide a reasonable rationale for the modvery poor fit for the 1500 s21 case since that high shearate would be expected to inhibit upstream depositsomewhat, as well as to create a higher predilectionembolization of weakly adherent platelets or platelet agregates, resulting in a deposition pattern with an irrelar, ‘‘bumpy’’ shape similar to that of the experimentacurve. For the 500 s21 case with heparin, the model’inability to simulate embolization of platelets fromthrombi near the entrance might explain the overfittingupstream deposition values. However, it would not eplain the continued overfitting of downstream valueAdditionally, if embolization were important at this sherate, one would expect its effects to be even more pnounced for the 1000 s21 model training case, for whicha much better fit was observed. One explanation for tdiscrepancy would be that the shorter perfusion tiused in the 1000 s21 case~75 vs 120 s for the 500 s21
case! was not sufficient for embolization to begin.

anstiretheeas ok.
u-ua-w-orttrobustosr-iont-ite-
eingre.g
theirecsignelylde.ch
ghctein
on.ceion,no-ofaset-aceady
ur-iononnel isx-ityllsions as,
eell-mtionra-
tybe
earsula-nglyto
ear-asn-is
o-r-
forf-
let-
stsueheththision
d inv-
edu-
Itsgh-
ti-ciesurre-h
hadgb-an
ver,r-of
456 SORENSEN et al.
The issues of shear-dependent platelet adhesionactivation reaction rates could be addressed by invegating functional kinetic relationships which are mocomplex than the simple first-order ones included inpresent model and which take into account the shdependence of the reactions. Addressing the problemfluid-thrombus interaction is a more challenging tasFirst, simulating thrombus growth would require copling the mass transport equations to the fluid flow eqtions, necessitating a computationally expensive flofield solution at every iteration of the mass transpequations. Second, a reasonably accurate way of inducing the mechanical interactions between the thromand the flow field would have to be incorporated inboth sets of equations. Fogelson4 has proposed doing thiby incorporating a fictitious body force into the NavieStokes equations which would account for deformatof the flow field by thrombus growth. However, simulaing embolization would be more challenging, sincewould require depicting the shape of the thrombus somhow so that the fluid dynamic forces on it could baccurately computed, as well as knowing somethabout the fracture mechanics of the thrombus structu
Despite the computational complexity of simulatinthese phenomena, the possibility of their inclusion inmodel needs to be considered since they have a dbearing on the usefulness of the simulations as deaids. For example, in attempting to design a relativ‘‘thrombosis-proof’’ device for long-term use, one wouneed to know when or if thrombus growth will stabilizThe present model would only be able to simulate sua phenomenon in a particular way: for a sufficiently hishear rate, generated agonists might be convequickly away and would not be able to accumulateconcentrations sufficient to cause platelet activatiCombined with a sufficiently low rate of platelet-surfaadhesion, this would lead to a cessation of depositbut not until the surface concentration reached a molayer level, which would clearly not be representativeall biomaterials. The time at which a monolayer wreached could be controlled by varying the platelsurface and platelet-platelet reaction rates, but surfdeposition would never reach a submonolayer stestate.
Since the concentration of platelets at a reactive sface directly determines the amount of platelet depositin our simulations, solution accuracy depends greatlyhow platelet transport to the wall is modeled. Thus, oof the most important deficiencies in the present modeour relatively crude method of inducing a near-wall ecess. With a true near-wall platelet excess, the diffusivof platelets would be low near the wall, where red ceare excluded, and much higher near the core regwhere most of the red cells reside. Our model usesingle value for diffusivity at all transverse location
d-
rf
-
t
d
,
which could help explain its predictive error, since thhigher diffusivity near the wall gradually destroys thskewed concentration profile. Furthermore, red-ceinduced platelet motion should be directed away frothe channel center regardless of the platelet concentragradient, whereas an enhanced diffusivity requires a gdient to produce platelet motion. Also, platelet diffusiviis currently an unknown parameter and must thereforeestimated for each case to be fit. In this article, a linvariation of platelet diffusivity with wall shear rate waassumed, so that a great deal of the error in our simtions could be attributed to a linear fit being the wrofunctional form for this parameter. To more accuratereproduce this phenomenon, it would be preferablehave a more accurate mathematical description of nwall excess. One model of this phenomenon which hshown the ability to reproduce experimental platelet cocentration profiles for flow in small-diameter tubesEckstein’s concept of red-cell-induced drift.3,15
Thrombin is clearly the most important platelet agnist in our simulations, with its concentration at the suface and in the bulk much higher than its thresholdplatelet activation. This occurs despite the inhibitory efects of heparin and despite the fact that our plateactivation rate constant@see Part I, Eq.~3!# weightsthreshold-normalized concentrations of all agoniequally. It is possible that thrombin’s dominance is dto the generation model we employed, which allows trate of thrombin production to increase linearly wiplatelet deposition and imposes no upper bound onrate. Although a linear increase in thrombin generatwith platelet concentration was shown by Rosinget al.,9
the studies were conducted using platelets suspendebuffer medium, rather than whole blood, thereby remoing the inhibitory effects of various plasma proteins~e.g.,C and S!. We intend to investigate more sophisticatthrombin generation functions for use in future reformlations of the model.
As modeled, ADP is also an important agonist.concentration was also generally suprathreshold throuout the domain, with normalized levels 25mm from thewall comparable to those of thrombin. The TxA2 con-centrations were generally well below the platelet acvation threshold we proposed. Consequently, this spedid not play a significant role in platelet activation in osimulations. This is consistent with the experimentalsults of Turneret al.,12 who observed that aspirin, whicinhibits platelets’ ability to synthesize TxA2, failed todecrease platelet deposition on type I collagen andvery little effect on platelet aggregation under flowinconditions. It is worth noting that our results were otained despite the fact that the model gave plateletsinexhaustible capacity to generate this agonist. Howeit is possible that our simplistic model of agonist inteaction may not correctly capture the contribution

erac
is-onc-odu-
ourf-ticis
tedveronn-en
ledithnwith
al-
, ager
ahiser
ehistele
be-
on,, ital-
emf-c-d-
hasics
ts,b-ib-
nistoutacets
yn-oningt toition
lythe
thee
st-ex-t2
h-in,rce
, isheotenxAalkac-it
ues,form-
s,be-Ais
edw-
no-ive-itionssf-
elet
lypt-I.ri-
457Simulation of Platelet Deposition: II. Results
TxA2, or that this agonist might participate in othimportant thrombogenic processes aside from platelettivation. Wagner and Hubbell14 postulated a role forTxA2 in thrombus stabilization, observing that admintration of aspirin reduced platelet-platelet adhesiontype I collagen. However, statistically significant redutions in deposition for aspirinized versus control blowere only obtained at longer flow times than those simlated here. We intend to investigate improvements toactivation function that better mimic the synergistic efects of various agonists, although quantitative kineinformation on such interactions in the literaturesparse.
CONCLUSIONS
We have presented a model of platelet-mediathrombogenesis which has been calibrated against sesets of experimental results for platelet depositiontype I collagen under simple parallel-plate flow. In geeral, the predicted results demonstrate good agreemwith the experimental trends in spite of many unmodeeffects, such as fluid-thrombus interactions. Starting wa case with a 1000 s21 wall shear rate and only heparias the anticoagulant, the model was able to fit cases~1! PPACK added as the anticoagulant at the same wshear rate; and~2! a fivefold higher heparin concentration, a wall shear rate lowered by a factor of 2 or 10domain 2.5 times as long, and an almost twofold lonperfusion time. We believe this indicates, at least inpreliminary fashion, the robustness of the model for trange of conditions. However, for the case with a highwall shear rate~1500 s21! and sodium citrate as thanticoagulant, a much poorer fit was observed. Tcould be due to a shear-dependent decrease in plaadhesion or agonist generation kinetics which onlycomes apparent at shear rates greater than 1000 s21, or toa fluid-dynamic mechanical effect such as embolizatiall of which are neglected in the present model. Orcould be the case that citrate’s chelating effect on ccium significantly alters the behavior of the entire systrelative to that of normal whole blood, i.e., that its efects go well beyond merely inhibiting thrombin prodution, which is all that was simulated in the present stuies. Either way, it is apparent that the citrate caserevealed that the model did not capture all of the physof the system being simulated.
As for the model’s behavior with regard to agonisthe distinctly different platelet deposition patterns oserved when thrombin is present versus when it is inhited can be explained by considering the ways agogeneration and platelet activation are simulated. Withthrombin, the rate of agonist generation at the surfwill depend on ~1! the rate at which resting plateleadhere, activate, and degranulate, releasing ADP; and~2!
-
al
t
l
t
the rate at which activated, surface-bound platelets sthesize TxA2. This surface-generated agonist productiwill induce platelet activation and agonist generationthe bulk fluid. However, recall that, in our model, restinplatelets can only adhere to the exposed surface, noother deposited platelets, and that the surface deposrate goes to zero at the surface saturation limit@see PartI, Eqs.~11! and~12!#. Thus, after surface saturation, onactivated platelet-platelet adhesion can occur, andonly sources of agonist generation will be TxA2 synthe-sis by activated platelets~at the surface and in the bulk!and ADP release by activating platelets~in the bulkonly!. At a high-shear rate such as that used inmodel-training simulations, the bulk agonists will bquickly convected away by the flow. Despite the worcase assumption that activated platelets have an inhaustible capacity for TxA2 synthesis, Fig. 2 shows thalevels of this agonist remain approximately a factor ofbelow the level required for platelet activation througout our simulations. Thus, in the absence of thrombvery little platelet activation, and therefore little furtheactivated platelet-platelet adhesion, will take place onthe surface-saturation limit is reached.
The rate of thrombin generation, on the other handproportional to local platelet concentration, either on tsurface or in the bulk. Therefore, when thrombin is ncompletely inhibited, platelets will adhere to the collagsurface and become activated, generating ADP and T2
and providing a phospholipid surface for additionthrombin production. This will lead to a higher bulconcentration of agonists, causing increased platelettivation and deposition. After the surface saturation limis reached, activated platelet-platelet adhesion continwhich provides even more phospholipid surfacethrombin generation, thereby leading to increased throbin production and further bulk activation. Thuthrombin-mediated platelet activation and depositionhave as a positive-feedback system. Theoretically, Tx2
synthesis should have a similar effect, since its ratealso proportional to the total concentration of activatplatelets, both at the surface and in the bulk flow. Hoever, the subthreshold levels of TxA2 found in our simu-lations prevent it from making a significant contributioto the rate of platelet activation. Conversely, in the nthrombin case surface saturation leads to a negatfeedback cascade, in which increased platelet deposresults in more rapid saturation, which results in leADP generation, which, combined with the washing efect of the high-shear flow and the low rate of TxA2
synthesis, eventually leads to a cessation of new platdeposition.
While the results presented in this article pertain onto a type I collagen substrate, the model is readily adaable to other biomaterials, as was discussed in Part11
We are currently attempting to fit the model to expe

toleti-
ngletn kseayde-
byt,
du-theyhis
i,,
entn
-
ort
n:
flet-
isny-
in
d
nf
ncem
. F.nd
S.l-
ow-l
sis
in
in
ed
458 SORENSEN et al.
mental results for synthetic biomaterials as well assituations involving more complex flows than the simpPoiseuille fields investigated herein. Also under invesgation are ways of refining the model by incorporatifluid-thrombus interaction, red-cell-induced platetransport enhancement, and shear-dependent reactionetics. It is anticipated that the culmination of theefforts will be a thrombosis model which may one daid in the design of thrombus-resistant blood-wettedvices.
ACKNOWLEDGMENTS
The work presented in this article was supportedthe McGowan Center for Artificial Organ Developmenthe Keck Foundation, and a Whitaker Foundation graate student research fellowship. The authors expressgratitude to Dr. W. K. Anderson of the NASA LangleResearch Center for generously making availablefluid dynamics analysis code,FUN2D, from which wederived our transport equation solver.
REFERENCES
1Alevriadou, B. R., J. L. Moake, N. A. Turner, Z. M. RuggerB. J. Folie, M. D. Phillips, A. B. Schreiber, M. E. Hrindaand L. V. McIntire. Real-time analysis of shear-dependthrombus formation and its blockade by inhibitors of voWillebrand factor binding to platelets.Blood 81:1263–1276,1993.
2Batchelor, G. K. An Introduction to Fluid Dynamics. Cambridge: Cambridge University Press, 1967.
3Eckstein, E. C., and F. Belgacem. Model of platelet transpin flowing blood with drift and diffusion terms.Biophys. J.60:53–69, 1991.
4Fogelson, A. L. Continuum models of platelet aggregatioformulation and mechanical properties.SIAM (Soc. Ind. Appl.Math.) J. Appl. Math.52:1089–1110, 1992.
i-
ir
5Folie, B. J., and L. V. McIntire. Mathematical analysis omural thrombogenesis. Concentration profiles of plateactivating agents and effects of viscous shear flow.Biophys.J. 56:1121–1141, 1989.
6Hubbell, J. A., and L. V. McIntire. Visualization and analysof mural thrombogenesis on collagen, polyurethane andlon. Biomaterials7:354–363, 1986.
7Keller, K. H. Effect of fluid shear on mass transportflowing blood. Fed. Proc.30:1591–1599, 1971.
8Munter, W. A., and P. D. Stein. Newtonian behavior of blooat high rates of shear.Biorheology10:501–508, 1973.
9Rosing, J., J. L. M. L. van Rijn, E. M. Bevers, G. vaDieijen, P. Comfurius, and R. F. A. Zwaal. The role oactivated human platelets in prothrombin and factorX acti-vation. Blood 65:319–332, 1985.
10Sakariassen, K. S., and H. R. Baumgartner. Axial dependeof platelet-collagen interactions in flowing blood. Upstreathrombus growth impairs downstream platelet adhesion.Ar-teriosclerosis (Dallas)9:33–42, 1989.
11Sorensen, E. N., G. W. Burgreen, W. R. Wagner, and JAntaki. Computational simulation of platelet deposition aactivation: I. Model development and properties.Ann.Biomed. Eng.27:436–448, 1999.
12Turner, N. A., J. L. Moake, S. G. Kamat, A. I. Schafer, N.Kleiman, R. Jordan, and L. V. McIntire. Comparative reatime effects on platelet adhesion and aggregation under fling conditions of in vivo aspirin, heparin, and monoclonaantibody fragment against glycoprotein IIb/IIIa.Circulation91:1354–1362, 1995.
13Wagner, W. R., and J. A. Hubbell. Local thrombin syntheand fibrin formation in anin vitro thrombosis model result inplatelet recruitment and thrombus stabilization on collagenheparinized blood.J. Lab. Clin. Med.116:636–650, 1989.
14Wagner, W. R., and J. A. Hubbell. Evidence for a rolethrombus stabilization for thromboxane A2 in human plateletdeposition on collagen.J. Lab. Clin. Med. 119:690–697,1992.
15Yeh, C., A. C. Calvez, and E. C. Eckstein. An estimatshape function for drift in a platelet-transport model.Biophys.J. 67:1252–1259, 1994.




















