
COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE FOR MANAGEMENT OF...
50
i COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE FOR MANAGEMENT OF DIABETES. PARTIAL FULFILMENT OF THE DEGREE PROGRAM IN PHARMACEUTICAL SCIENCES RESEARCH PROJECT REPORT WRITTEN BY: OMOLO CALVIN ANDEVE REGISTRATION NUMBER: 2009/2346 COURSE NAME: BACHELORS DEGREE IN PHARMACEUTICAL SCIENCES DEPARTMENT: PHARMACEUTICAL RESEARCH PROJECT SUPERVISOR: DR. H.MAURICE DATE OF SUBMISSION: 4 th AUGUST 2013 ST JOHN’S UNIVERSITY OF TANZANIA SCHOOL OF PHARMACY
Transcript of COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE FOR MANAGEMENT OF...

i
COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE FOR MANAGEMENT OF
DIABETES.
PARTIAL FULFILMENT OF THE DEGREE PROGRAM IN PHARMACEUTICAL
SCIENCES
RESEARCH PROJECT REPORT
WRITTEN BY: OMOLO CALVIN ANDEVE
REGISTRATION NUMBER: 2009/2346
COURSE NAME: BACHELORS DEGREE IN PHARMACEUTICAL
SCIENCES
DEPARTMENT: PHARMACEUTICAL RESEARCH PROJECT
SUPERVISOR: DR. H.MAURICE
DATE OF SUBMISSION: 4th AUGUST 2013
ST JOHN’S UNIVERSITY OF TANZANIA
SCHOOL OF PHARMACY

i
DECLARATION
I do declare that this dissertation work entitled “Computational methods in designing
novel inhibitors of deoxyhypusine synthase for management of Diabetes’’ which is sub-
mitted by me in partial fulfillment of the requirement for the completion of bachelor’s de-
gree in pharmaceutical sciences comprises only my original work and due acknowledge-
ment has been made in the text to all other material used.
OMOL CALVIN ANDEVE
2009/2346

ii
CERTIFICATION
The undersigned certifies that he has read and hereby recommend for acceptance by St.
John’s University of Tanzania, a dissertation entitled Computational methods in design-
ing novel inhibitors of deoxyhypusine synthase for management of Diabetes, in partial
fulfillment of the requirement for the award of of bachelor’s degree in pharmaceutical sci-
ences
Supervisor: Dr. H. Maurice
Signature:
Date:

iii
ACKNOWLEDGEMENT
The most notable source of guidance, inspiration and motivation was my advisor,
Dr. Maurice Bishop, whom I owe a great deal of thanks for taking me under his wing and
allowing me to take up some of his knowledge, insight and wisdom. He has not only made
us to work hard but guided us towards orienting ourselves research and learn invaluable
skills towards drug discovery and medicinal chemistry.
I am highly grateful to the honorable, Dr. Joseph Mhando, for his ever helping attitude
and encouraging us to excel in studies. Besides, he has been a source of inspiration during
my entire period of four years in St. Johns University
I am also thankful to rest of the classmates for their cooperation during the whole of this
four years we have been working together and cooperating with each other.
This acknowledgement will not complete until I pay my respectful homage to my family
whose enthusiasm to see this work complete was as infectious as their inspiration. Lastly
but not least I give my thanks to the Almighty God for the gift of life, wisdom, persever-
ance, health and humility as I travelled this journey.
Omolo Calvin Andeve

iv
ABSTRACT
Diabetes mellitus is one of the most common non-communicable diseases, and if uncon-
trolled, targets multi-organ systems with serious debilitating and life-threatening sequel.
Despite having multifaceted aetiologies the most common of its manifestation is hypergly-
cemia by the patients. At the molecular level, pancreatic cell loss by apoptosis appears to
play an important role in the development of insulin deficiency and the onset and/or pro-
gression of the disease. Beta cells population are dynamic and undergoes compensatory
changes to maintain euglycemia. The structural parameters that allow mass changes (rep-
lication, neogenesis, cell volume changes, and cell death) are stimulated by many precur-
sors. Hypusine has been found to play an important role in the turnover of beta cells through
initiation of inflammation cascade. Inhibition of hypusine synthesis will go a long way in
increasing resistance of beta cells to apoptosis. This paper will illustrate computational
methods to design novel of inhibitors of deoxyhypusine synthase, an enzyme that plays an
important role in the synthesis of hypusine. The process will include finding computational
model of deoxyhypusine synthase, designing its ligand, and then using different libraries
to scan for various chemicals that will have lower binding energy to deoxyhypusine syn-
thase energy than spamidine. The results obtained will pave way for designing and synthe-
sising chemicals that can be potent deoxyhypusine synthase inhibitors.

v
List of Abbreviations
TID 1: diabetes type 1
TID 2: Diabetes type 2
β - cell: pancreatic βeta cells
Bcl2: B cell lymphoma 2
BAX: Bcl-2–associated X protein
BAK: Bcl-2 homologous antagonist/killer
cAMP: Cyclic adenosine monophosphate
CADD: Computer Aided Drug Designing
CAMD: Computer Aided Molecular Designing
EIF5A: Eukaryotic translation initiation factor 5A
IL-1beta: interleukin-1beta
IFNgamma: interferon-gamma
TNFalpha: tumor necrosis factor-alpha
DC: dentritic cells
HDACs: histone deacetylases
TSA: trichostatin A
SAHA: suberoylanilide hydroxamic acid
IAPs: Inhibitor of apoptosis proteins
NFκB: nuclear factor kappa-light-chain-enhancer of activated B cells JNKs:
MRNAs: messenner RNA designing
HIV: Human Immuno defincency Virus
iNOS: Nitric Acid synthase Inhibitor
NK: Natural Killer
DHS: deoxyhypusine Synthase
PDB: Protein Data Bank
QSAR: Quantitative Structure Activity Relationship
CNI-1493: uanylhydrazone
SBDD: Structure-Based Drug Design
TNF: Tumor necrosis factor
VS: Virtual screenin

vi
List of Figure
Figure 1.1 to 1.5 agents that inhibit transcription of NFKb………………………………...7
Figure 2.1 guanylhrazone…………………………………………………….…………….13
Figure 2.2 – 2. Exixting inhobitors of DHS…………………………………………...…...14 .
Figure 3.0 Docking task flow chart………………………………………………………...17
Figure 4.1.1 DHS 3D imageof DHS……………….………………………………..… …21
Figure 4.1.2 3-D Structure of DHS -2……..………………………………………….…...22
Figure 4.1.3 Ramachandran for Deoxyhypusine synthaseProtein generated by Discovery stu-
dio 3.5………………………………………………………………………..….……...37
Figure 4.3.1 to 4.3.10 2D ligand 1 – 10 ………………………………………..….….22 - 27
Figure 4.4.1 2D Pharmacophore Model of Top Ligands………………………………28 - 32
Figure 4.6.1 ligand 10 binding scope……………………………………………………...33
Figure 4.6.2 pocket surface Hbonding…………..…………………………….……...….35
Figure 4.6. 3 Pocket binding pattern…………………………………………………..…..37 ….
Figure 4.6.4 aromaticizity of the pocket surface………………………………...……….39
Figure 4.6.5 Sas ……………………………………………………………………………40
Figure 4.6.6 hydrophobicity of the pocket surface………………………………………38

vii
List of Tables
Table 4.2 Drug like top ten ligands properties………………………………….27
Table 4.6.1 receptor surface H-bonds ………………………………………….35
Table 4.6.2 Aromaticity of the receptor surface………………………………..36
Table 4.6.3 Binding pattern………………………………………………...…..38
Table 4.6.4. sas………………………………………………………………....39
Table 4.6.5 hydrophobicity……………………………………………………..40

viii
Table of Contents ABSTRACT ............................................................................................................................................. iv
List of Abbreviations .............................................................................................................................. v
List of Figure ......................................................................................................................................... vi
List of Tables......................................................................................................................................... vii
1.0 Introduction ..................................................................................................................................... 1
1.1 Diabetes and inflammation .......................................................................................................... 1
1.6 CONTRIBUTION OF THE PROJECT TO DRUG DEVELOPEMENT .................................................... 4
2.0 Literature Review............................................................................................................................. 5
2.1 Current Novel inhibitors of deoxy Hypusine Synthase ................................................................ 8
3.0 Material and Methods ................................................................................................................... 11
3.1 MATERIALS USED ....................................................................................................................... 11
3.2 METHODOLOGY ......................................................................................................................... 12
4.0 Results and Discussion ................................................................................................................... 15
4.1 3-D Structure of deoxy hypusine synthase ................................................................................ 15
4.3 Drug like Top 10 ligands ............................................................................................................. 17
4.4 Deoxyhypusine synthase with top ligands ................................................................................. 22
4.5 Generated Pharmacophore Model ............................................................................................ 28
4.6 Receptor Binding Site analysis ................................................................................................... 30
4.8 Discussion .................................................................................................................................. 37
4.9 Limitation of the Study. ............................................................................................................. 38
5.0 Conclusion ..................................................................................................................................... 39
6.0 Future Plan of Work ....................................................................................................................... 40
References ............................................................................................................................................... 41

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
1
1.0 Introduction
1.1 Diabetes and inflammation
Diabetes mellitus describes a group of chronic diseases where absolute or relative lack of in-
sulin leads to aberrancies in glucose, proteins and fat metabolism, causing acute and long-
term complications. Contrary to what is known hyperglycemia is the hallmark for clinical
diagnosis, however metabolic disturbances in diabetes mellitus are not limited to glucose but
incorporate most processes of the intermediary metabolism of nutrients, including proteins
and lipids. Diabetes mellitus is associated with insulin resistance, but that is not the whole
story because rare mutations in the insulin signaling cascade occur. As it is known the dis-
ease does not occur if insulin release from the pancreatic cells in the islets of Langerhans is
intact.1 Preserving a functional cell mass is therefore the primary target of novel treatments
aimed at curing and preventing diabetes mellitus.
With respect to pathogenesis, the strict contrast between T1D and T2D is highly generalised.
There is increasing recognition that T1D and T2D may represent extremes of a continuous
spectrum with a dominating β-cell defect at one end and dominating insulin resistance at the
other2. However, when not putting into perspective diabetes caused by rare mutations in in-
sulin signaling, insulin resistance is neither necessary nor sufficient to cause diabetes,
whereas defects in the pancreatic beta cells is both a necessary and sufficient to cause the
disease. This concept is held by studies indicating progressive decrease in beta cell function
and mass in T2D Autoimmune islet inflammation and pancreatic β-cell obliteration are long
documented causes of T1D, although it is contested if the molecular effector mechanisms
involve principally classical cytotoxic T-cell–mediated or predominantly inflammatory cyto-
kine-mediated β-cell killing or both.
Recently, a unifying hypothesis was proposed by the observation that all these stimuli lead
to the initiation of inflammatory mediators in the pancreatic islets that cause β-cell oblitera-
tion by triggering cascades in cells similar to those in T1D. Thus, in spite of their dissimilar
genetic origins, the immune and metabolic progression of both T1D and T2D, seem to con

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
2
gregate on mutual extracellular inflammatory stressors in the islets and intracellular gesticu-
lating induced by these stressors3
Extra cellular inflammation markers are linked to signalling of inflammatory process is
probably consequence of hypusinated protein eif5a. This protein is hypusinated by the en-
zymes deoxyhypusine synthase and deoxyhypusine hydroxylase. The hypusinated protein
plays an important role of stimulating cytokines. Pancreatic beta cell damage caused by pro-
inflammatory cytokines interleukin-1beta (IL-1beta), interferon-gamma (IFNgamma) and
tumor necrosis factor-alpha (TNFalpha). Hypusinated owes their stimulation to hypusinated
elF5A, which attains this through gesticulation via transmembrane potential translocation of
Bax to the mitochondria, consequently release of cytochrome c, caspase activation, and up-
regulation of p53. EIF5A expression is significantly upregulates DC (dentritic cells) and
maturation which are in turn responsible for the production of caspaces which are responsi-
ble in causing apoptosis. Inhibition of hypusination goes a long way in shielding pancreatic
beta cells to apoptosis
Figure 1.1Adopted from Shao Chin Lee and Shazib Pervaiz 20064
Erstwhile studies have refer to molecules with protective effects on the beta cells from cyto-
kines, because the act as antioxidants or anti-inflammatory agents. For example, resveratrol
even in the presence of cytokines results in restoration of sustainability of the pancreatic
beta cells, possibly by NAD-dependent deacetylase sirtuin-15 activation. Furthermore

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
3
O O
NHOH
NHtrichostatin
Larsen et al. established that molecules that inhibits histone deacetylases (HDACs) with tri-
chostatin A (TSA) or suberoylanilide hydroxamic acid (SAHA) averts cytokine prompted
beta cell death, most probably by diminishing NFκB transactivation6 Hence, numerous
agents like Alsterpaullone, pyrazole derivatives, dexamethasone, have shown to score highly
in the tanimoto score index with significant results even though acting with different mecha-
nisms as protectants beta cells from cytokine induced apoptosis.
OHOH
F
H
OHOH
O
Dexamethasone Figure 1.2
OH
OH
OH
resveratrol
Figure 1.3
Figure 1.4

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
4
NHNH
OH
O
O
suberoylanilide hydroxamic acid
Figure 1.5
1.6 CONTRIBUTION OF THE PROJECT TO DRUG DEVELOPEMENT
The number of new drugs and drugs in pipeline compared to drugs which are becoming ab-
solute is lower. Faster methods of drug discovery have to be found so that new agents are
available for combating diseases which many of the current drugs in the market are becom-
ing are failing to treat. Computational chemistry is one such method which speeds up drug
discoveries and one of the most economical methods.
Diabetes is one of the debilitating disease, with prevalence rates increasing each and every
day. Most causes of diabetes have been so far been targeted however high turnover of pan-
creatic beta cells is one of the cause of diabetes type 2. Most drugs discovery research ha-
ven’t involved the effect of inflammation as the cause of apoptosis of the pancreatic beta
cells. This paper will pay way for further research in the discovery of drugs which will pro-
vide invaluable insight in the development of drugs that will help in controlling and prevent-
ing diabetes type two.

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
5
2.0 Literature Review
Diabetes, a syndrome of glucose homeostasis, which has escalated to near pandemic propor-
tions and may be the single most central risk factor for heart, renal, and ocular diseases. Dys-
function and damage of islet beta cells, which is caused in part by the systemic or local release
of pro-inflammatory cytokines, bring about all forms of diabetes. A most important effort in
diabetes findings in recent years has been to ascertain new factors or pathways that can be
therapeutically pinpointed to reduce cytokine action on the beta cell.
Regulation of cellular activities is done through posttranslational alterations of proteins, which
may comprise phosphorylation, acetylation and methylation. These alterations are typically
attached to numerous types of proteins by several enzymes, and thereby instantaneously reg-
ulate a wide assortment of protein functions. Consequently, most signaling pathways are eas-
ily terminated, enabling maintenance of cellular integrity even if the modification of a single
signaling molecule is disrupted, signaling can still be achieved. An outstanding exclusion is
hypusine. This essential PTM is limited to a single protein: the eukaryotic initiation factor 5A
(eIF-5A) 7
Recent studies have suggested that an ancient and poorly understood protein, eif5a, may be
important to cytokine release and signaling. Interestingly, eif5a is the only protein which con-
tains a unique amino acid hypusine, which is derived from sparmidine that originates from
alteration of lysine amino acid residue. This modification is catalyzed by the sequential ac-
tions of enzymes deoxyhypusine synthase and deoxyhypusine hydroxylase which can be in-
hibited. Because the hypusine modification is solely dependent on eif5a action in cytokine
signaling the inhibition of modification could be an invaluable target for anti-diabetic agents8.
Hypusine is uncommon amino acid that can be traced in all eukaryotes and in some Achaea,
but not in bacteria. The only known protein containing hypusine is eukaryotic translation ini-
tiation factor 5a (eif5a), nature has committed two enzymes. Deoxyhypusine synthase and
deoxyhypusine hydroxylase, are the enzymes used to modify one unique lysine residue on a

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
6
single Polypeptide, eif-5a, into hypusine.9 The commitment of the process to this two enzymes
underlines the importance of targeting these enzymes in creating novel anti diabet agents.
EIF5A is the only known protein to contain the atypical amino acid hypusine (Nε-[4-amino-
2-hydroxybutyl]-lysine). in formation of eif5a the paramount step, deoxyhypusine synthase
(DHS) transfers an aminobutyl moiety from the polyamine spermidine to the ε-amino group
of lysine 50 in eIF5A to form deoxyhypusine; subsequently, deoxyhypusine hydroxylase
(DOHH) hydroxylates deoxyhypusine to the final hypusine residue (in figure1 below ). Both
deoxyhypusine synthatse and deoxyhypusine hydroxylase only act on eIF5A, and in human
pancreatic islets there is only a single protein (eIF5A) which is observed to form 3H-spermi-
dine when radioactive hydrogen (3H) is incorporated. The hypusinated form of eIF5A is con-
sidered as the functional form, and until this moment most known functions of eIF5A are
owed to its hypusinated form. Thus, for processes dependent upon hypusinated eIF5A, target-
ing of hypusination serves as a specific means to impede eIF5A action.
Recently, studies have associated hypusinated eIF5A in specific disease progressions. Re-
markably, findings of Hauber and colleagues have connected hypusinated eIF5A in the nucleo
cytoplasmic transport of mRNAs involved in HIV progression in T cells (Hauber et al., 2005).
These findings Indicates a fascinating conversion whereby viruses have evolved to “hijack”
specific functionalities of hypusinated elF5A for the benefit of self-propagation and spread-
ing. Consequently, inhibition of hypusination of elF5A may have a therapeutic value in dis-
ease states such as diabetes and HIV. In this respect, recent studies have implicated a permis-
sive role for eIF5A in inflammation. Depletion of eIF5A in vivo (using liposomal siRNA)
was amazingly shown to safeguard mice from fatal doses of lipopolysaccharides of in a mu-
rine model of endotoxin prompted sepsis (Moore et al., 2008). In this model, systemic inten-
sities of pro-inflammatory cytokine and chemokines were reduced, indicative of a pro-inflam-
matory activity of eIF5A10

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
7
Figure 2 . 0 . Chemistry o f hypusine formation. The figure depicts formation of hypusine in
a reaction involving the Lys50 (lysine at position 50) residue of eIF5A, spermidine, and the
enzymes deoxyhypusine synthase and deoxyhypusi ne hydroxylase.
In both type 1 and type 2 diabetes, pancreatic islet dysfunction domino effect is due to cyto-
kine facilitated inflammation. The pervasive eukaryotic translation commencement factor 5A
(eIF5A), which is the only protein to contain the amino acid hypusine, contributes to the pro-
duction of pro-inflammatory cytokines. The pathogenesis of islet beta cell dysfunction in type
1 and type 2 diabetes includes inflammatory signaling induced by cytokines, leading acutely
to production of inducible nitric oxide synthase (iNOS) and nitric oxide11. This is mediated
through the increased coding of iNOS. Inhibition of translation inhibition factor in mice re-
sulted to reduced levels of iNOS in rodent beta cells and human islets and safeguarded the
mice against the development of glucose intolerance the low dose streptozotocin model of
diabetes. Further investigation exposed that hypusine is vital in part for nuclear export of
iNOS-encoding mRNA, a process that entangled the export protein exportin1.12 Pro-inflam-
matory cytokines acutely trigger NF-κB–mediated transcription of the Nos2 gene encoding
iNOS 13. Production of nitric oxide by iNOS plays an important role to the early pathogenesis
of β cell dysfunction in reaction to cytokines, as nitric oxide impedes proteins associated in
aerobic glycolysis and the electron transport chain, this act diminishes ATP production.14.
This impairment in ATP production limits the coupling of glycolysis to insulin release in the
β cell. In the longer term, both the iNOS-dependent and -independent effects of cytokine sig-
naling lead to eventual islet death15.

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
8
Thus to preserve islet function and reduce the high turnover rate due to apoptosis, it is imper-
ative to identify and counter the mechanisms that mediate islet responsiveness to pro-inflam-
matory cytokines. This can be achieved by inhibiting deoxy hypusine synthase, enzyme in-
volved in the synthesis of hypusinated elF5A.
2.1 Current Novel inhibitors of deoxy Hypusine Synthase
In search for novel inhibitors of purified plasmodium DHS, the guanylhydrazone (CNI-1493),
a novel selective pro-inflammatory cytokine inhibitor used in clinical phase II for the treat-
ment of Crohn’s disease, inhibited the enzyme of the parasite. In vitro it decreases serum TNF
levels of the mice. It was proposed that the new mechanism of CNI-1493 was instigated by a
decrease in modified eIF-5A biosynthesis with a downstream outcome on the TNF synthesis
of the host.16 From the current data, CNI-1493 is considered to be a promising drug for anti-
malarial therapy, anti-inflammatory, and anti-cytokine, anti-neoplastic and anti-viral for HIV
especially for patients with multi-drug resistant viral strains. This was because of its joint
action, i.e., the decrease in eIF-5A biosynthesis in both parasite and host and in the host’s cell
TNF biosynthesis.
N
N+H H
NNN
+
N
H
H
NNN
+
H
NHN
O
N
N
NN N
+
H
H
O
H
HH
H
H
H
H
H
H
H
guanylhydrazone
Figure 2.1
It has been recently reported that down-regulation of eukaryotic initiation factor 5A (eIF5A)
in Imatinib treated K562 cells. Additionally, the inhibition of eIF5A by siRNA in combination
with Imatinib has been shown to exert synergistic cytotoxic effects on BCR-ABL positive cell
lines. Based on the structure of known deoxyhypusine synthase (DHS) inhibitors such as CNI-
1493, a drug design approach was applied to develop potential compounds targeting DHS. A

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
9
report of the biological evaluation of selected novel
(DHSI-15) as compared to established (CNI-1493, deox-
yspergualin) DHS inhibitors, showed that upon the com-
pounds tested, DHSI-15 and deoxyspergualin exerted
strongest anti-proliferative effects on BCR-ABL cells including Imatinib resistant mutants.
Both compounds were able to induce apoptosis/necrosis during long term incubation of BCR-
ABL positive BA/F3 derivates. Finally it showed that deoxyspergualin was able to inhibit
proliferation of CD34+ progenitor cells from CML patients. Which lead to conclusion that
inhibition of deoxyhypusine synthase (DHS) can be supportive for the anti-proliferative treat-
ment of leukemia.17
Inhibition of deoxyhypusine synthase by 1-guanidino-7-aminoheptane (GC7) suppressed tu-
mor cell growth. Using GC7 as a prototype three classes of diamine analogs, namely, guani-
dino-, pyrimidino-, and hydroxamate derivatives were synthesized and tested, as potential in-
hibitors for deoxyhypusine synthase. In the study showed that (i) among all the compounds
tested, GC7 remained to be the most potent inhibitor for deoxyhypusine synthase; (ii) N,N′-
bispyrimidino-1,9-diaminononane, although a poor inhibitor of deoxyhypusine synthase,
were a potent growth inhibitor; and (iii) one of the hydroxamate derivatives, 6-aminohexanoic
hydroxamate (HC6), prominently induced the differentiation of mouse neuroblastoma cells at
sub-mill molar concentrations18
NH NH2
NH
NH2
1-guanidino-7-aminoheptane
Figure 2.2
N
N N NH2
H
N,N'-bispyrimidino-1,9-diaminononane
Figure 2.3
NH2N
H
OH
O
6-aminohexanoic hydroxamate

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
10
Figure 2.4
The above findings merits for further investigation for inhibitors of this enzyme, which can
result to data and structures of invaluable importance to finding drug targets for diabetes and
even other diseases which effective cures haven’t been found yet. This project will provide a
paramount insight towards achieving that

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
11
3.0 Material and Methods
3.1 MATERIALS USED
The materials used will included
3.11 Hardware
i. Laptop
ii. Flash drive
iii. Wires internet modem
iv. Printer
3.12 Software
Argus lab
It’s a Molecular modelling, graphics, and drug design program, Contains both the GADock and Ar-
gusDock docking engines and the AScore scoring function with a preliminary set of parameters that
helps will help docking and scoring of ligands on the binding sites.
Ligand scout
LigandScout is a software tool that will allow to rapidly and transparently derive 3D pharmacophores
from structural data of macromolecule/ligand complexes in a fully automated and convenient way.
ACD Chemsketch ver 12.01
ACD/ChemSketch Freeware is a drawing package that allows you to draw chemical structures includ-
ing organics, organometallics, polymers, and Markush structures. It also includes features such as cal-
culation of molecular properties (e.g., molecular weight, density, molar refractivity etc.), 2D and 3D
structure cleaning and viewing, functionality for naming structures, and prediction of logP.
Accerlys Discovery Studio 3.5 visualizer
DS Visualizer is a free, feature-rich molecular modeling environment, for both small mole-
cule and macromolecule applications. Built from, and fully compatible with the latest release
of our enterprise-ready expert modeling product, Discovery Studio 3.5, experts and col-
leagues can seamlessly and efficiently exchange results, without loss of either time or scien-
tific information.

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
12
The project undertaking involved:
i. Binding site and ligand identification
ii. Virtual screening
iii. Docking
iv. Active sites analysis
3.2 METHODOLOGY
Figure 3.0
3.2.1Binding site and Ligand identification
Protein target coordicates extracted from the PDB entry corresponded to the Crystal structure
of the Deoxyhypusine Enzyme (PDB code: 1ROZ). All water molecules were removed from
the active site. File modification were necessary for the docking programs
3.2.2 Virtual screening.
The screening stage involved the following step
i. The first stage involved searching and downloading several drug screening libraries
ii. The crystal structure containing the enzyme with ligand bound to it was opened using
ligand scout software.

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
13
iii. The using the software the pharmacophore of the ligand was created.
iv. Then the libraries was attached, then the software was set to align and calculate
pharmacophore score of the ligand comparing with the agents in the libraries.
v. After screening the score was displayed, using the software the scores were arranged in
the descending order.
vi. The agents with highest score i.e. ascore of fifty from various libraries was drawn using
Argus lab software, then they were docked on the binding site and their docking ener-
gies calculated and obtained.
vii. The compound with lowest binding energy was selected as the lead hit target.
3.2.3 Binding pocket analysis
The binding pocket was analyzed using Ligand scout 3.1 and acceryls discovery studio 3.5.the
software’s had the ability to simulate and map out the ligand biding site interactions, neighbor-
ing residues, amino acids making the pocket and the atoms in the pocket.
After docking the docking energy was recorded and the ligands showing lower docking energy
than natural ligand to the enzyme were selected and the binding complex between the ligand
and the target protein was saved in .pbd file format. The ligand complex file was then trans-
ferred to the accerlys Discovery studio 3.5 where the binding pattern, the binding pocket, hy-
drophobicity and the Hydrogen bonding of the binding site were analyzed. After analysis top
ten drug like ligands were chosen. Then chosen ligands were further analyzed to find the strik-
ing features for their pocket binding and the residues in the pocket they bind to.
3.2.4 Collection Data
Data results were be obtained from the simulation of the computer software’s which was used
as follows in the project. Data collection parameter and consideration for accurate data were
illustrated in the following text. The X-ray structures of protein ligand complexes were selected
basing on High resolution and well-defined atomic positions of the ligand. The X-ray pose
was considered as the bioactive one and further used as the reference to evaluate computed
conformations of total of the ligands used to compose a data set. The chemical diversity
of ligands was assessed to note the molecule attachments that are responsible for activity of the
ligand. Correlation was observed between ligand’s various attached and decrease in the docking
energies. Both the size of the protein and the conformational flexibility of the ligand were fur-

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
14
ther used to classify binding sites and discuss docking performance of the ligand. Since dock-
ing was based on not only shape complementary but also favorable ligand receptor interactions,
docking results were examined with regard to the polar surface, hydrophobicity, hydrogen
bonding and ionizability area of the ligand. Special attention was paid to hydrophobic
ligands, as well as very polar ligands.

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
15
4.0 Results and Discussion
4.1 3-D Structure of deoxy hypusine synthase
Generated Deoxyhypusine synthase protein 3D structures from Ligand scout 3.1 software is
illustrated below:
Figure 4.1.1 3-D Structure of of DHS (side view)
Figure 4.1.2 3D Structure DHS-2 (from acceryl discovery studio)

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
16
RAMACHADRAN PLOT:
Figure 4.1.3 Ramachandran for DHS Protein generated by discovery stu-
dio 3.5.
The Ramachandran above showed the phi-psi torsion angles for all residues in the structure
(except those at the chain termini). The white areas corresponded to which conformations where
atoms in the polypeptide come closer than the sum of their van der Waals radii. These regions
were statically disallowed for all amino acids except glycine which is unique in that it lacks a
side chain. The green regions corresponded to conformations where there are no steric clashes,
i.e. these are the allowed regions namely the sheet conformations. The red areas
show the allowed regions if slightly shorter van der Waals radii were used in the calculation,
i.e. the atoms are allowed to come a little closer together and it brought out an additional region
which corresponded to the left-handed helix.

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
17
4.3 Drug like Top 10 ligands
Figure 4.3.1 to 4.3.10 chemical structures of top ten drug like ligands
Ligand 1 (C19H20N2OS3)
Figure 4.3.1 ligand 1
Ligand 2 (C26H20ClF6NO2)
Figure 4.3.2
COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
18
Ligand 3(C21H21Cl4F3N2O4S)
Figure 4.3.3
Ligand 4 (C24H15NO5)
Figure 4.3.4

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
19
Ligand 5 (C30H28Cl4F5N3O6S)
Figure 4.3.5
Ligand 6(C37H39Cl2F3N8O4)
Figure 4.3.6

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
20
Ligand 7(C26H22Cl3F5N2O4)
Figure 4.3.7
Ligand 8 (C38H37Cl2F3N8O6)
Fiugre 4.3

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
21
Ligand 9(C25H16ClF3N2O4)
Figure 4.3.9
Ligand 10 (C30H26Cl3F6N3O6S)
Figure 4.3.10 C30H26Cl3F6N3O6S

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
22
Table 4.3 properties of the top ten ligands ligand
Pharmacoscore Docking Energy Log P
Molecular
weight
1 46.76 - 12.1208 Kcal/mol 5.33 388.57
2 44.34 -12.3954 Kcal/mol 8.53 527.89
3 56.77 -12.4551 Kcal/mol 4.22 596.27
4 44.23 - 13.0358 Kcal/mol 5.52 397.38
5 66.78 -13.1674 Kcal/mol 6.48 795.43
6 71.68 -13.5186 Kcal/mol 4.04 787.66
7 56.77 -13.6983 Kcal/mol 6.44 627.81
8 76.33 -13.9683 Kcal/mol 6.03 829.65
9 48.98 -13.9782 Kcal/mol 2.26 500.85
10
55.33 - 14.8698 Kcal/mol 6.05
776.96
4.4 Deoxyhypusine synthase with top ligands
Deoxyhypusine synthase residues interactions with top ligands as shown in Figures 4.4.1 to
4.4.10 below, the green colour represents Vander Waals bonds, pink represents electrostatic
bonds, purple colour represents covalent bonding, and the grey colour shows metal interactions.
Figure 4.4.1 ligand 1 residues interactions

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
23
Figure 4.4.2 ligand 2 residues interactions
Figure 4.4.1 ligand 2 binding to the residues
Figure 4.4.3 Deoxyhypusine synthase and ligand 3

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
24
Figure 4.4.4 Ligand 4 interaction with residues
Figure 4.4.5 3D Deoxyhypusine synthase residue atoms and Ligand 5

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
25
Figure 4.4.6 Deoxyhypusine synthase and Ligand 6
Figure 4.4.7 Deoxyhypusine synthase and Ligand 7

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
26
Figure 4.4.8 Deoxyhypusine synthase and Ligand 8
Figure 4.4.9 ligand 9 residues interactions
Figure 4.5.9 2D ligand 9 interaction

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
27
Figure 4.5 2D ligand 10 interaction with hypusine Deoxyhypusine synthase
(Ligand with the lowest binding energy)

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
28
4.5 Generated Pharmacophore Model
After finding the top drug like molecules, we have done structural alignment of these
molecules and generated a pharmacophore of these molecules [Structures of top ligands
obtained from virtual screening were analyzed and it was found that there were two
types of ligands which had lower binding energy. The includeded;
Type one had the following features
- 2H-chromene structure
5
6
10
7
9
8
3
2
4
O1
Figure 4.5.1
- On the chromic structure; ligands with methyl at position 7 had lower docking energies.
O NH2
OH
3-aminopropane-1,2-diol
Figure 4.5.2
- 3-aminopropane-1,2-diol bridge between the chrome and benzyl moiety has been noted in the
ligand which have shown to have lower binding energy.
- Ligands with Heavily substituted benzyl moiety attached to the amino of the 3-aminopropane -
1,2-diol had ligands with lower docking energies.
F
F
F
S
O
ONH2
Cl
Cl
2-(dichloromethyl)-6-(trifluoromethyl)benzenesulfonamide
Figure 4.5.3

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
29
When the ligand had 2-(dichloromethyl-6-(trifluoromethyl)benzylsulfonamide resulted to ligand
that had docking energy of -14.698Kcal/mol. When the sulfonamide substituent is replaced with
Hydroxyl moiety the docking energy rose to -13.156Kcal/mol. Ligands without the sulfonamide
moiety and and when the ligand had Chloro instead of dichloromethyl moiety the docking energy
rose to -136903Kcal/mol.
The second set of ligands consisted of two acridin-9-amine moieties linked with 2-((2-4-butyla-
mino-3-oxobutyl)amino)ethyl)amine-N-((methylamino)methyl. The substitutions on the moieties
had various impact on the docking energies of the ligands
Figure 4.5.4
Figure 4.5.5
COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
30
4.6 Receptor Binding Site analysis
Figure 4.3 Pharmacophore Model of Top Ligands
4.6.1 Binding pocket analysis by POCKET program
By running the POCKET program, the binding pocket from 3D structure of Deoxyhypu-
sine synthase(represented in PDB format) was analyzed and the key interaction sites
within the binding pocket were also derived.
Figure 4.6.1 Ligand 10 binding site interactions
4.6.1Binding pocket hydrophobicity
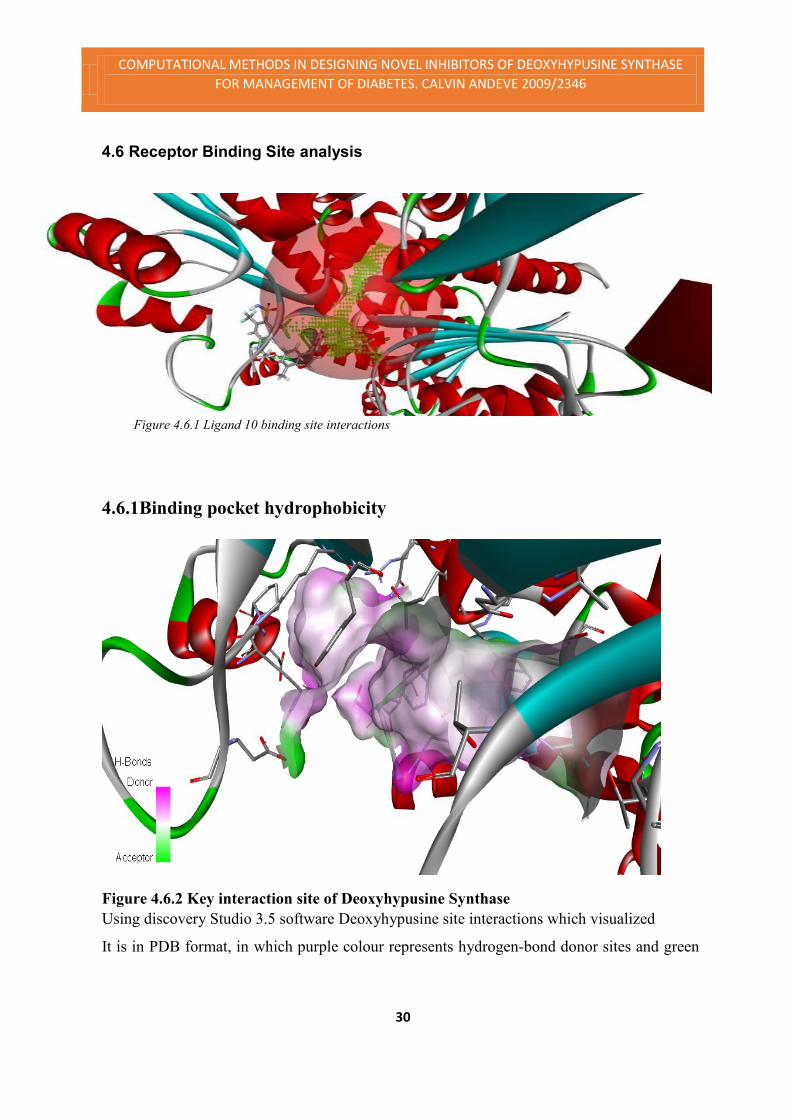
Figure 4.6.2 Key interaction site of Deoxyhypusine Synthase
Using discovery Studio 3.5 software Deoxyhypusine site interactions which visualized
It is in PDB format, in which purple colour represents hydrogen-bond donor sites and green

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
31
colour represent hydrogen-bond acceptor sites; and carbon atoms (grey) represent hydropho-
bic sites [see Figure.4.6.1]. Table 4.6.1 shows the number of receptor atoms with the ligands.
It illustrates the exposed groups, Pi-Pi interactions. During the ligand receptor interactions.
The exposed protein groups were 284 and buried protein groups were 264. And a total of 81
ligand atoms interfaced with the receptor and total of 513 pocket atoms had interface with the
ligand. The table also shows 8 H-Bond donors and I Hbond acceptor in the pocket, that’s why
the pink colour dominates above in figure 4.6.2
Table 4.6.1 interface of receptor atoms with ligand 10
Name Parent Size
Buried Protein Groups 262
Exposed Protein Groups 284
Main-Chain HB Donors Protein Groups 2
Side-Chain HB Donors Protein Groups 6
Side-Chain HB Acceptors Protein Groups 1
Main-Chain Polar Contacts Protein Groups 7
Side-Chain Polar Contacts Protein Groups 12
Main-Chain Nonpolar Contacts Protein Groups 3
Side-Chain Nonpolar Contacts Protein Groups 11
Main-Chain Mixed Contacts Protein Groups 13
Side-Chain Mixed Contacts Protein Groups 21
Pi-Pi Interactions Protein Groups 1
Pi-Cation Interactions (charge) Protein Groups 1
Interface Ligand Atoms with DHS Interface Groups 81
Interface Pocket Atoms with DHS Interface Groups 513
Current Ligand <TreeRoot> 1

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
32
Figure 4.6.2 shows the aromatic edge of the binding pocket
The table above shows the aromatic interactions at the binding site. The interaction involving
the edge of the benzyl moiety are colored in blue and those involving the face are colored in
orange.
Table 4.6.2 receptor aromatic face interactions
Name
Col-
our Parent X Y Y
Dis-
tance End1
End
2
Lambda
angle
Theta
an-
gle1
Theta
angle2
Lig10 -
B:PHE47
2
Pi-Pi
Interac 52.937 4.251 31.169 4.70576 Lig10
B:P
HE4
72 12.912
43.94
8 56.77
Lig10 -
B:ARG48
6:NH1
Pi-Cat-
ion In-
terac 53.11
-
0.031 33.124 5.49343 Lig10
B:A
RG4
86:N
H1
Lig10 -
B:ARG48
6:NH2
Pi-Cat-
ion In-
terac 53.532
-
0.894 32.493 6.28949 Lig10
B:A
RG4
86:N
H2
The table above shows the a sample of of aromatic interactions involving the face of the
benyl moieties in ligand 10 and those of phenyl groups of phenylalanine 427 which resulted
to the Pi-pi interaction, arginine 486 which resulted to the Pi-cation interactions.

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
33
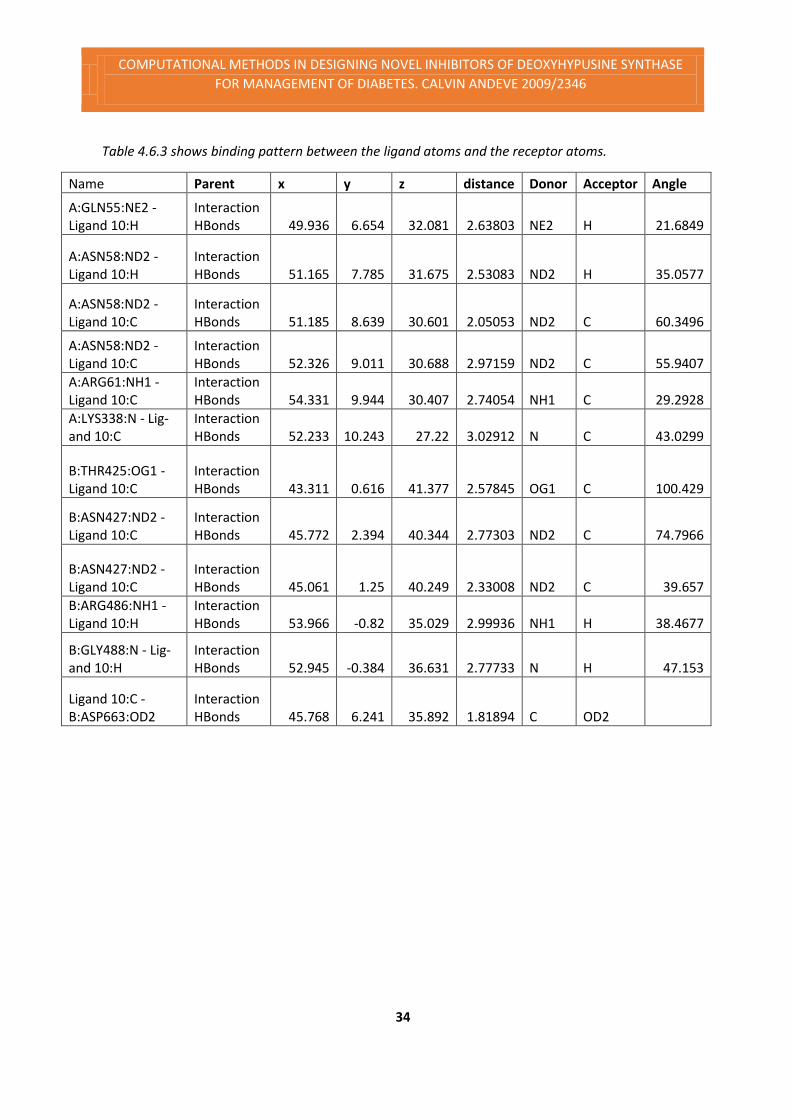
Figure 4.6.3 Ligand 10 pocket binding pattern
Figure 4.6.3 Ligand 10 pocket binding pattern
The figure above shows the ligand pocket binding pattern. The pattern is visualized by the
purple like threads showing the bond interactions. The below (table 4.6.3) sums up the interac-
tions of the bonds and the ligand. The table sums up the ligand atom type of interaction. The
bond distance ant the type of interaction i.e. whether acceptor or donor and the angle bond
interactions form.
COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
34
Table 4.6.3 shows binding pattern between the ligand atoms and the receptor atoms.
Name Parent x y z distance Donor Acceptor Angle
A:GLN55:NE2 - Ligand 10:H
Interaction HBonds 49.936 6.654 32.081 2.63803 NE2 H 21.6849
A:ASN58:ND2 - Ligand 10:H
Interaction HBonds 51.165 7.785 31.675 2.53083 ND2 H 35.0577
A:ASN58:ND2 - Ligand 10:C
Interaction HBonds 51.185 8.639 30.601 2.05053 ND2 C 60.3496
A:ASN58:ND2 - Ligand 10:C
Interaction HBonds 52.326 9.011 30.688 2.97159 ND2 C 55.9407
A:ARG61:NH1 - Ligand 10:C
Interaction HBonds 54.331 9.944 30.407 2.74054 NH1 C 29.2928
A:LYS338:N - Lig-and 10:C
Interaction HBonds 52.233 10.243 27.22 3.02912 N C 43.0299
B:THR425:OG1 - Ligand 10:C
Interaction HBonds 43.311 0.616 41.377 2.57845 OG1 C 100.429
B:ASN427:ND2 - Ligand 10:C
Interaction HBonds 45.772 2.394 40.344 2.77303 ND2 C 74.7966
B:ASN427:ND2 - Ligand 10:C
Interaction HBonds 45.061 1.25 40.249 2.33008 ND2 C 39.657
B:ARG486:NH1 - Ligand 10:H
Interaction HBonds 53.966 -0.82 35.029 2.99936 NH1 H 38.4677
B:GLY488:N - Lig-and 10:H
Interaction HBonds 52.945 -0.384 36.631 2.77733 N H 47.153
Ligand 10:C - B:ASP663:OD2
Interaction HBonds 45.768 6.241 35.892 1.81894 C OD2

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
35
Figure 4.6.4 SAS analysis was used to evaluate how deep a given residue is buried The SAS
of each structure was calculated by Solvent Accessibility Calculation of Discovery Studio 3.5
software with probe radius of 1.4 Å. The differences in SAS among residues in the models
were determined. Two residue groups were created: 1) exposed group, which contained resi-
dues with maximum SAS values greater than 25%, and 2) buried group, which contained resi-
dues with maximum SAS less than 10%. These SAS data were used in the subsequent epitope
screening. From the table the exposed groups were 284 and buried groups were 262.
Table 4.6.4 shows the exposed and buried proteins due to receptor ligand interaction.
Name Visible Parent Size
Buried Yes Protein Groups 262
Exposed Yes Protein Groups 284
Interface Ligand Atoms (DHS with ligand 10) Yes Interface Groups 81
Interface Pocket Atoms (DHS with ligand 10) Yes Interface Groups 513
Current Ligand Yes <TreeRoot> 1

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
36
Figure 4.6.5 hydrophobicity of the pocket surface due to receptor ligand interaction
The figure above show the hydrophobicity and hydrophilic interfaces due to the receptor ligand
interface. The blue colour indicates the hydrophilic interfaces and moving towards the brown
colour indicates the increase in hydrophobicity. The table below goes to indicate the number of
hydrophobic and hydrophilic groups
Table 4.6.5. Hydrophobicity of the pocket surface
Name Visible Parent size
Backbone Yes Protein Groups 2,628
Sidechain Yes Protein Groups 2,516
Hydrophobic Yes Protein Groups 264
Hydrophilic Yes Protein Groups 214
Acidic Yes Protein Groups 74
Basic Yes Protein Groups 71
Ligand 9 Yes Ligand Groups 1
Interface Receptor Atoms (DHS and ligand 10) Yes Interface Groups 10,250
Interface Ligand Atoms (DHS with ligand10) Yes Interface Groups 81
Interface Pocket Atoms (DHS with ligand 10) Yes Interface Groups 513
Current Ligand Yes <TreeRoot> 1

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
37
4.8 Discussion
The available drugs for diabetes target, Insulin Secretagogues, euglycemics: (Medicines that
bring the blood glucose into the normal range), alpha-glucosidase inhibitors: starch blockers
(delay digestion and absorption of carbohydrates), thiazolidinediones (Glitazones or tzds),
decrease insulin resistance in the body (muscle and fat tissues), glp-1 analogs: increase insu-
lin secretion, reduce glucose release from liver after meals, delay food emptying from stom-
ach and promote satiety19
The project aimed at looking at managing diabetes differently, in designing DHS inhibitors
which will go a long way in inhibiting the destruction of the pancreatic beta cells through in-
hibiting apoptosis of pancreatic beta cells.
The 3D structure of Deoxyhypusine synthase was downloaded from the protein database-
website, and it was used as the binding site for the docking in the search for drug like ligands
for designing deoxyhypusine synthase inhibitors. We carried out screening of the possible
ligands from ZINC, ASINEX, AMBINTER and HANTINTON databases. Top ten ligands
which were obtained from virtual screening using Argus lab program had energy scores
ranging from – 14.8698 to - 12.1208 Kcal/mol [see Table 4.3]. Three dimensional structures
of top ten ligand molecules were visualized and it was found that these ligand or drugs like
molecules were dock in the same to the protein. Energy score which is the sum of van der
Waals and electrostatic components. More negative energy score (kcal/mol) corresponds to
the more binding affinity.
Further analysis of the pocket was done ligand 10, which was the ligand with the least free
docking energy. Using the ligand several values of hydrophobicity, Hbonding, aromaticity,
solvent accessibility surface of the pocket surface were obtained using the discovery studio
3.5. It was clear that the above parameters played a paramount role in the binding affinity of
the pocket to the ligand. And the ligand to bind had to have several functional groups that
directly affected the above named parameters of the receptor binding surface.
This data signifies that after further optimization process these probable leads can generate a
potent inhibitor for the Deoxyhypusine synthase. After visualizing the Deoxyhypusine syn-
thase with top ten ligand complexes, it was found that above ligands were best fitted in the

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
38
cavity of receptor and visualization with Discovery Studio 3.5 found that these ligands are
interact in same active site. There were notable structures in the top ligands obtained from
virtual screening, the structures can be used as an indicators to choose the ligand that could
be used as top Targets to be used as modal for hit modification to so that to have lead struc-
tures for synthesis. Analyzing further moieties of the structures it was found that, there were
two sets of ligands interacting in the pocket. The one that contained the 2H-chromene and 3-
aminopropane-1,2-diol bridge between the chrome and benzyl as the pharmacophoric struc-
ture. And the second set of ligands consisted of acridin-9-amine moieties linked with 2-((2-4-
butylamino-3-oxobutyl)amino)ethyl)amine-N-((methylamino)methyl[see Figure4.5.4 and
figure 4.5.5].
4.9 Limitation of the Study.
The study can considered a success, however there were several factors that were stumbling
blocks towards the study, which will be enlighten below.
The first issue was most of the software’s we used were the free versions of the software, this
was an issue because most of the free versions have limited capabilities compared to the pro-
fessional editions. This made it difficult to analyse and visualisation of some features of the
Ligand –receptor interaction. Even it reached a time interligand suspended the application for
academic licences from St. Johns’ university.
The time allocation was really limited that’s why only few top ligands were found, due to that
we couldn’t conclusively term the top ligands as potent inhibitors of DHS
The money allocated wasn’t enough to carry out the projects. Costs for data mining which in-
cluded internet cost were really high.
Lack of powerful machines to carry out the Docking and visualisation algorithms was a limit-
ing factor. It took days to dock a ligand on the receptor site.

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
39
5.0 Conclusion
The present project was aimed at finding novel drug like molecules for the selective
competitive inhibition of Deoxyhypusine synthase. Deoxyhypusine synthase are
playing crucial role in initiation of pancreatic beta cells apoptosis. The 3D structure of
this enzyme was obtained from the Protein Data bank site(PDB Code: 1ROZ) was
considered as template and generate 3D structure of Deoxyhypusine synthase for hu-
mans. These proteins were used for virtual screening for finding novel ligands
having better energy scores using the program Argus Lab and database of commer-
cially available chemical moieties. We used anticancer drug eflornithine as the ligand
for screening. Through Argus lab software we found better energy score ligands.
We have found a novel seed molecules containing 2H-chromene and 3-aminopro-
pane-1,2-diol bridge and another set of ligands containing acridin-9-amine moieties
linked with 2-((2-4-butylamino-3-oxobutyl)amino)ethyl)amine-N-((methylamino)me-
thyl, basing these new targets future lead targets can be crafted from them and ligands
containing 2H-chromene and 3-aminopropane-1,2-diol bridge and another set of lig-
ands containing acridin-9-amine moieties linked with 2-((2-4-butylamino-3-oxobu-
tyl)amino)ethyl)amine-N-((methylamino)methyl may act as a potent and specific in-
hibitor for Deoxyhypusine synthase; though their efficacy, toxicity and pharmacoki-
netic properties need to be studied experimentally.

COMPUTATIONAL METHODS IN DESIGNING NOVEL INHIBITORS OF DEOXYHYPUSINE SYNTHASE
FOR MANAGEMENT OF DIABETES. CALVIN ANDEVE 2009/2346
40
6.0 Future Plan of Work
These structures with similar to structures those obtained by virtual screening i.e. 2H-
chromene and 3-aminopropane-1,2-diol bridge and another set of ligands containing acridin-
9-amine moieties linked with 2-((2-4-butylamino-3-oxobutyl)amino)ethyl)amine-N-((me-
thylamino)methyl might be potent and specific inhibitors for DHS though more study is
required to confirm the physiochemical properties and safety of the drugs.
Molecular dynamics study of these Deoxyhypusine synthase inhibitors complexes with top
scored ligands reported herein, would throw more light on the thermodynamic stability
aspect of the ligand- receptor complexes. Substructure-pharmacophore studies of these
top scored ligands might help in building more specific and high affinity inhibitors. Moreo-
ver, during the filtering process more substantive ligands might have got missed, therefore
alternative parameters may be applied and new structures worked out and the seed structure
assessed. Such results may help to corroborate the present findings. However, wet experi-
ments involving chemical synthesis and testing the designed molecules in vivo using specific
cell lines would be necessary to arrive at definite conclusions

References
1 Raj SM, et al. No association of multipletype 2 diabetes loci with type 1 diabetes. Diabetologia. 2009
2 Barrett JC, et al. Genome-wide association study and meta-analysis find that over 40 loci affect risk of type 1 diabetes. Nat.Genet 2009
3 McCarthy MI, Lindgren CM, Type 2 diabetes: new genes, new understanding, TrendsGenet. 2008
4 Shao Chin Lee and Shazib Pervaiz , Apoptosis in the pathophysiology of diabetes mellitus, IJBCB
2006
5 Pacholec M, et al, resveratrol are not direct activators of SIRT1. J Biol Chem, pub med
6 Larsen L et al, Inhibition of histone deacetylases prevents cytokine-induced toxicity in beta cells Dia-
betologia 2007, Pubmed
7 Henning S. Et al, Protein-Protein-Interaction Network Organization of the Hypusine Modification System 2012, MCP press 8 Abbruzzese A, The role of eukaryotic initiation factor 5A in the control of cell proliferation and apoptosis
2001.
9 Kuang Yu Chen and David L. Jao, Chemistry of Hypusine Formation on Eukaryotic Initiation Factor 5A in Bio-logical Systems 1999
10 Abbruzzese A., The role of eukaryotic initiation factor 5A in the control of cell proliferation and apop-
tosis, 2001
11 Yurika N. et al, Regulation of deoxyhypusine synthase by p38 MAPK links cytokine signaling to the transla-
tion of iNOS in pancreatic islets, Indiana University School of Medicine, Indianapolis 2011
12 Bernhard M. et al, The unique hypusine modification of eIF5A promotes islet β cell inflammation and
dysfunction in mice, JCI 2011
13 Eizirik DL, Mandrup-Poulsen T. A, choice of death the signal-transduction of immune-mediated beta-
cell apoptosis. Diabetologia. 2001;
14 Leist M, et al. Inhibition of mitochondrial ATP generation by nitric oxide switches apoptosis to necro-
sis. Exp Cell Res. 1999.
15 Chambers KT, Unverferth JA, Weber SM, Wek RC, Urano F, Corbett JA. The role of nitric oxide and
the unfolded protein response in cytokine-induced beta-cell death.Diabetes. 2008.
16 Sommer M-N, et al. Screening assay for the identification of deoxyhypusine synthase inhibitors. J,
Biomol. Screen. 2004, Pubmed
17 Patrick. Z et al, Investigational New Drugs: Evaluation of deoxyhypusine synthase inhibitors targeting
BCR-ABL positive leukemias, Volume 30, Issue 6 December 2012, Springer US
18 Jiebo L et al, Aminohexanoic hydroxamate is a potent inducer of the differentiation of mouse neuro-
blastoma cells, Cancer Letters, Volume 160, Issue 1, 10 November 2000
19http://dtc.ucsf.edu/types-of-diabetes/type2/treatment-of-type-2-diabetes/medications-and-thera-
pies/type-2-non-insulin-therapies/table-of-medications/ visited on 7th August 2013.




















