
St John's wort, hypericin, and imipramine - Nature
18
Molecular Psychiatry (2001) 6, 547–564 2001 Nature Publishing Group All rights reserved 1359-4184/01 $15.00 www.nature.com/mp ORIGINAL RESEARCH ARTICLE St John’s wort, hypericin, and imipramine: a comparative analysis of mRNA levels in brain areas involved in HPA axis control following short-term and long-term administration in normal and stressed rats V Butterweck 1,2 , H Winterhoff 1 and M Herkenham 2 1 Institute of Pharmacology and Toxicology, Domagkstrasse 12, 48149 Muenster, Germany; 2 Section on Functional Neuroanatomy, National Institute of Mental Health, Bethesda, Maryland 20892, USA Clinical studies demonstrate that the antidepressant efficacy of St John’s wort (Hypericum) is comparable to that of tricyclic antidepressants such as imipramine. Onset of efficacy of these drugs occurs after several weeks of treatment. Therefore, we used in situ hybridization histochemistry to examine in rats the effects of short-term (2 weeks) and long-term (8 weeks) administration of imipramine, Hypericum extract, and hypericin (an active constituent of St John’s wort) on the expression of genes that may be involved in the regulation of the hypotha- lamic-pituitary-adrenal (HPA) axis. Imipramine (15 mg kg −1 ), Hypericum (500 mg kg −1 ), and hypericin (0.2 mg kg −1 ) given daily by gavage for 8 weeks but not for 2 weeks significantly decreased levels of corticotropin-releasing hormone (CRH) mRNA by 16–22% in the hypotha- lamic paraventricular nucleus (PVN) and serotonin 5-HT 1A receptor mRNA by 11–17% in the hippocampus. Only imipramine decreased tyrosine hydroxylase (TH) mRNA levels in the locus coeruleus (by 23%), and only at 8 weeks. The similar delayed effects of the three compounds on gene transcription suggests a shared action on the centers that control HPA axis activity. A second study was performed to assess the effects of long-term imipramine and Hypericum administration on stress-induced changes in gene transcription in stress-responsive circuits. Repeated immobilization stress (2 h daily for 7 days) increased mRNA levels of CRH in the PVN, proopiomelanocortin (POMC) in the anterior pituitary, glutamic acid decarboxylase (GAD 65/67) in the bed nucleus of the stria terminalis (BST), cyclic AMP response element binding protein (CREB) in the hippocampus, and TH in the locus coeruleus. It decreased mRNA levels of 5-HT 1A and brain-derived neurotrophic factor (BDNF) in the hippocampus. Long-term pre- treatment with either imipramine or Hypericum reduced to control levels the stress-induced increases in gene transcription of GAD in the BST, CREB in the hippocampus, and POMC in the pituitary. The stress-induced increases in mRNA levels of CRH in the PVN and TH in the locus coeruleus were reduced by imipramine but not by Hypericum. The stress-induced decreases in BDNF and 5-HT 1A mRNA levels were not prevented by either drug. Taken together, these data show: (1) that Hypericum and hypericin have delayed effects on HPA axis control centers similar to those of imipramine; and (2) that select stress-induced changes in gene transcription in particular brain areas can be prevented by long-term treatment with either the prototypic tricyclic antidepressant imipramine or the herbiceutical St John’s wort. However, imipramine appears to be more effective in blocking stress effects on the HPA axis than the plant extract. Molecular Psychiatry (2001) 6, 547–564. Keywords: antidepressant; hypothalamic-pituitary-adrenal; corticotropin-releasing hormone; para- ventricular hypothalamic nucleus; norepinephrine; locus coeruleus; imipramine; Hypericum; in situ hybridization; gene expression regulation Introduction St John’s wort (Hypericum perforatum L, Hyperi- caceae) is a plant that has been used as a medicinal herb since ancient times. Today, Hypericum is used in Correspondence: M Herkenham, PhD, Section on Functional Neu- roanatomy, NIMH Bldg 36, Rm 2D-15, Bethesda, MD 20892–4070, USA. E-mail: mileshcodon.nih.gov Received 22 December 2000; revised 29 March 2001; accepted 3 April 2001 many countries for the treatment of mild to moderate forms of depression, and its efficacy has been con- firmed in several clinical studies. 1 The therapeutic action of Hypericum is comparable to that of tricyclic antidepressants, whereas its side effects are less pro- nounced, resulting in a better compliance of the patients. 2 Several behavioral studies support the antidepress- ant activity of Hypericum extracts. A methanolic extract significantly reduced immobility time in the
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of St John's wort, hypericin, and imipramine - Nature

Molecular Psychiatry (2001) 6, 547–564 2001 Nature Publishing Group All rights reserved 1359-4184/01 $15.00
www.nature.com/mp
ORIGINAL RESEARCH ARTICLE
St John’s wort, hypericin, and imipramine: acomparative analysis of mRNA levels in brain areasinvolved in HPA axis control following short-term andlong-term administration in normal and stressed ratsV Butterweck1,2, H Winterhoff1 and M Herkenham2
1Institute of Pharmacology and Toxicology, Domagkstrasse 12, 48149 Muenster, Germany; 2Section on FunctionalNeuroanatomy, National Institute of Mental Health, Bethesda, Maryland 20892, USA
Clinical studies demonstrate that the antidepressant efficacy of St John’s wort (Hypericum)is comparable to that of tricyclic antidepressants such as imipramine. Onset of efficacy ofthese drugs occurs after several weeks of treatment. Therefore, we used in situ hybridizationhistochemistry to examine in rats the effects of short-term (2 weeks) and long-term (8 weeks)administration of imipramine, Hypericum extract, and hypericin (an active constituent of StJohn’s wort) on the expression of genes that may be involved in the regulation of the hypotha-lamic-pituitary-adrenal (HPA) axis. Imipramine (15 mg kg−1), Hypericum (500 mg kg−1), andhypericin (0.2 mg kg−1) given daily by gavage for 8 weeks but not for 2 weeks significantlydecreased levels of corticotropin-releasing hormone (CRH) mRNA by 16–22% in the hypotha-lamic paraventricular nucleus (PVN) and serotonin 5-HT1A receptor mRNA by 11–17% in thehippocampus. Only imipramine decreased tyrosine hydroxylase (TH) mRNA levels in the locuscoeruleus (by 23%), and only at 8 weeks. The similar delayed effects of the three compoundson gene transcription suggests a shared action on the centers that control HPA axis activity.A second study was performed to assess the effects of long-term imipramine and Hypericumadministration on stress-induced changes in gene transcription in stress-responsive circuits.Repeated immobilization stress (2 h daily for 7 days) increased mRNA levels of CRH in thePVN, proopiomelanocortin (POMC) in the anterior pituitary, glutamic acid decarboxylase (GAD65/67) in the bed nucleus of the stria terminalis (BST), cyclic AMP response element bindingprotein (CREB) in the hippocampus, and TH in the locus coeruleus. It decreased mRNA levelsof 5-HT1A and brain-derived neurotrophic factor (BDNF) in the hippocampus. Long-term pre-treatment with either imipramine or Hypericum reduced to control levels the stress-inducedincreases in gene transcription of GAD in the BST, CREB in the hippocampus, and POMC inthe pituitary. The stress-induced increases in mRNA levels of CRH in the PVN and TH inthe locus coeruleus were reduced by imipramine but not by Hypericum. The stress-induceddecreases in BDNF and 5-HT1A mRNA levels were not prevented by either drug. Takentogether, these data show: (1) that Hypericum and hypericin have delayed effects on HPA axiscontrol centers similar to those of imipramine; and (2) that select stress-induced changes ingene transcription in particular brain areas can be prevented by long-term treatment witheither the prototypic tricyclic antidepressant imipramine or the herbiceutical St John’s wort.However, imipramine appears to be more effective in blocking stress effects on the HPA axisthan the plant extract. Molecular Psychiatry (2001) 6, 547–564.
Keywords: antidepressant; hypothalamic-pituitary-adrenal; corticotropin-releasing hormone; para-ventricular hypothalamic nucleus; norepinephrine; locus coeruleus; imipramine; Hypericum; in situhybridization; gene expression regulation
IntroductionSt John’s wort (Hypericum perforatum L, Hyperi-caceae) is a plant that has been used as a medicinalherb since ancient times. Today, Hypericum is used in
Correspondence: M Herkenham, PhD, Section on Functional Neu-roanatomy, NIMH Bldg 36, Rm 2D-15, Bethesda, MD 20892–4070,USA. E-mail: milesh�codon.nih.govReceived 22 December 2000; revised 29 March 2001; accepted 3April 2001
many countries for the treatment of mild to moderateforms of depression, and its efficacy has been con-firmed in several clinical studies.1 The therapeuticaction of Hypericum is comparable to that of tricyclicantidepressants, whereas its side effects are less pro-nounced, resulting in a better compliance of thepatients.2
Several behavioral studies support the antidepress-ant activity of Hypericum extracts. A methanolicextract significantly reduced immobility time in the

mRNA changes in the stress axis induced by St John’s wortV Butterweck et al
548
Molecular Psychiatry
forced swimming test.3 After 14 days of daily treatmentin rats, the extract dose dependently reduced prolactinand corticosterone plasma levels.4 In in vitro studies,various extracts have been shown to inhibit synaptoso-mal uptake of norepinephrine, serotonin and dopa-mine5 and to inhibit ligand binding to the GABAA, themu-, delta- and kappa-opioid, and the 5-HT6 and 5-HT7 receptors.6
St John’s wort is a complex mixture of over twodozen constituents.7 The antidepressant activity ofHypericum extracts has been attributed to the phloro-glucinol derivative hyperforin,8,9 to the napthodi-anthrones hypericin and pseudohypericin,10 and toseveral flavonoids.11,12 The activities of the differentHypericum compounds are still a matter of debate, buttaking the previous findings together, it is likely thatthe different individual substances contribute to theantidepressant activity of the crude plant extract in acomplicated manner. In the present study, we studiedhypericin in addition to the full Hypericum extractbecause hypericin displayed high affinity for severalaminergic receptor subtypes in vitro,13 and was activein the Porsolt forced swim test.10,14
Studies that investigate the molecular targets of StJohn’s wort relevant to its antidepressant effects havenot been done. It is not known how St John’s wortaffects key neurochemical systems in the brain in testsin which animals receive therapeutic dose regimens forprotracted periods of time and under normal andstressed conditions. In this study, we employed twosuch tests in rats. The first test is designed to revealneurochemical changes that are selectively associatedwith the delayed onset of therapeutic efficacy inhumans. The second test uses an animal model ofchronic stress that produces defined neurochemicalchanges against which the actions of long-term antide-pressant drug administration can be evaluated. In bothtests, the actions of St John’s wort have been concur-rently compared with those of the prototypic tricyclicantidepressant drug imipramine.The first test is based on a characteristic feature com-
mon to both tricyclic antidepressants and St John’swort—the delay of 2 weeks or more before the thera-peutic effect becomes evident. In an earlier animalstudy designed to examine the association betweenlong-term antidepressant administration and thedelayed alteration in hypothalamic-pituitary-adrenalaxis (HPA) axis activity, corticotropin releasing hor-mone (CRH) mRNA levels in the hypothalamic para-ventricular nucleus (PVN) of rats were found to bedecreased following long-term (8 weeks) but not short-term (2 weeks) treatment with imipramine.15 The sameresults were found with several other antidepressantdrugs selected for their distinctly different primarypharmacological actions.16 Thus in this study, we mea-sured changes in CRH mRNA levels at these two timepoints, and we also measured mRNA expression in anumber of CNS areas involved in and related to HPAaxis control.The second test of the antidepressant efficacy of St
John’s wort employed an animal model of chronic
stress. In humans, prolonged stress is associated withcertain mood disorders.17,18 An important link betweendepression and stress is established by the facts thatthe HPA axis and particular sets of 5-hydroxytrypta-mine (5-HT), norepinephrine, and glutamic acid decar-boxylase (GABA) containing neuronal systems areinvolved both in the pathophysiology of depressionand the neurobiology of stress.19,20 Several clinicalstudies give evidence that depression is associatedwith an activation of the HPA axis.20–22 In the courseof chronic antidepressant drug treatment, the hyperac-tivity of the HPA axis normalizes with clinical recoveryfrom depression.23
Chronic stress in animals results in elevated adreno-corticotropic hormone (ACTH) and glucocorticoid lev-els in plasma and elevated production of CRH in thePVN.24–26 In addition, tyrosine hydroxylase (TH) geneexpression in the locus coeruleus is elevated,24,27 sug-gesting that stress activates the norepinephrine-con-taining neurons that play an important role inbehavioral arousal. Recently, other work has extendedthese findings in animals to include observations ofneurochemical changes in brain structures known toinfluence the HPA axis. These include GABA in thebed nucleus of the stria terminalis (BST), assessed bymeasures of GAD mRNA levels,28 and several neuro-chemical markers in the hippocampus.29
The direction of changes in mRNA levels followingstress is generally opposite to that of the same genes inthe same locations following long-term antidepressanttreatment. However, the relationship between the neur-ochemical effects of chronic stress and those of antide-pressant drug administration has received little study.Thus, in the second study we used an immobilizationstress procedure to: (1) show stress-induced changes ingene expression in selected brain regions: (2) deter-mine whether long-term pre-treatment with eitherimipramine or St John’s wort blocks the stress-inducedchanges; and (3) find differences between the synthetictricyclic antidepressant imipramine and the naturalherbiceutical St John’s wort in their ability to modifythe effects of chronic stress.
Material and methods
Experiment 1—short-term/long-term treatment designMale CD rats (150–180 g, Charles River WIGA, Sulz-feld, Germany) were single housed in a 12 h light/darkcycle, with lights off at 19.00 h, at a constant tempera-ture of 25 ± 1°C and free access to food (Altromin 1324)and tap water. Rats were randomly assigned to the vari-ous experimental groups and weighed daily. Theexperimental procedures used in this work wereofficially approved by the Regierungsprasident, Muns-ter (A 38/93).
Imipramine hydrochloride was obtained from Sigma(Deisenhofen, Germany), and hypericin was obtainedfrom Roth (Karlsruhe, Germany). A lyophilized meth-anolic native Hypericum extract (LI 160, drug/extractratio: 4–7:1) was obtained from Lichtwer Pharma AG(Berlin, Germany). The extract LI 160 was subsequently

mRNA changes in the stress axis induced by St John’s wortV Butterweck et al
549characterized by HPLC using the method published byButterweck et al.3 The main constituents were: chlor-ogenic acid 5.5 mg g−1, rutin 20.1 mg g−1, hyperoside20.15 mg g−1, isoquercitrine 10.8 mg g−1, astilbin 2.4 mgg−1, quercitrine 5.0 mg g−1, quercetin 4.6 mg g−1, 13.118-biapigenine 3.8 mg g−1, hyperforin 53.1 mg g−1, andhypericin 2.21 mg g−1.All substances were administered orally using the
gavage technique. The oral administration route had tobe chosen because: (1) both the Hypericum extract andhypericin are barely soluble in aqueous solvents andthus must be administered as a suspension; (2) theHypericum extract suspension is pH 4–5 (for intraperi-toneal injections, solutions should have a 7.0–7.5 pH);(3) for chronic application, intraperitoneal injectionsare contraindicated;30 and (4) gavage—if done prop-erly—is less stressful for animals than intraperitonealinjection.30 For consistency of method, imipraminewas also given by gavage.Drug solutions were prepared fresh daily. Stock com-
pounds were kept in light-tight containers. Control ani-mals received deionized water with an ethanol contentof 160 �l per 10 ml. Imipramine (15 mg) and Hyper-icum extract (500 mg) were first moisturized in 160 �lethanol and then diluted with deionized water to afinal volume of 10 ml. A homogeneous suspension ofthe Hypericum extract was obtained by sonication for5 min. Hypericin is barely soluble in water, therefore5 mg hypericin were nearly dissolved in 2.5 ml ethanoland further diluted with water to a final concentrationof 0.2 mg per 10 ml. The ethanol concentration of thehypericin solution was 1%. The final application vol-ume of each preparation was 10 ml kg−1 b.w. A dosageof 15 mg kg−1 imipramine was chosen because previousbehavioral experiments (V Butterweck, unpublisheddata) indicated that oral dosages below 15 mg wereonly partially active; for chronic treatment, 15 mg kg−1
proved to be active without toxic effects, whereashigher doses administered chronically were toxic. TheHypericum extract (500 mg kg−1) and the hypericin (0.2mg kg−1) dosages were chosen because of their demon-strated efficacy in the forced swimming test.3,10
Animals were killed by decapitation between 9.00and 11.00 h; the last drug administration was the daybefore between 16.00 and 17.00 h. Brains wereremoved, frozen by immersion in 2-methyl butane at−30°C, and stored at −70°C prior to sectioning. Trunkblood was collected on ice-chilled EDTA-coated (10ml) tubes containing 500 KIU aprotinin ml−1, centri-fuged, and plasma was frozen at −70°C.
Experiment 2—stress plus drugs designMale Sprague–Dawley rats (150–180 g. Taconic Farms,Germantown, NY, USA) were housed two per cage ina 12 h light/dark cycle, with lights off at 18.00 h, at aconstant temperature of 25 ± 1°C and free access tofood and tap water. Rats were randomly assigned to thevarious experimental groups and weighed daily. Drugs(imipramine and Hypericum extract) were preparedand administered as in the first experiment. Control
Molecular Psychiatry
animals received deionized water only. Administrationwas performed between 8.00 and 10.00 h every day.The immobilization procedure was approved by the
Animal Care and Use Committee of the National Insti-tute of Mental Health Intramural Research Program.Rats were immobilized for 2 h in the morning duringthe nadir of their circadian rhythm. Animals wereimmobilized by placing the head through stainlesssteel loops and securing each limb with tape to a stain-less steel platform as described previously.31 Theimmobilization schedule used in this study was thestress design from Mamalaki et al.24 A group of 48 ani-mals was subdivided as follows: 16 animals weretreated with imipramine (15 mg kg−1, p.o.), 16 weretreated with Hypericum extract (500 mg kg−1, p.o.), and16 animals received vehicle (deionized water) over aperiod of 7 weeks. In the seventh week, eight animalsof each treatment group were immobilized for 2 h oncedaily for 7 days and killed immediately after the laststress session.An acute paradigm was designed to investigate
whether a single exposure of rats to immobilizationstress could induce changes in mRNA expression inbrain areas involved in HPA axis control, and inaddition, whether acute pre-treatment with imipra-mine could antagonize possible changes in stress-induced gene expression. Thus, a second group of 32animals was divided into an unstressed and a stressedgroup. Unstressed animals received either imipramine(15 mg kg−1 b.w., n = 8) or vehicle (deionized water, n= 8) 2 h prior to decapitation. Stressed animals (n = 8per group) received the same treatment, respectively.Two h after oral administration of test substances, ani-mals were exposed to a single immobilization stress for2 h and were killed immediately after the stress ses-sion. Unstressed animals from the acute and chronicexperiment were habituated by daily handling. Allgroups were killed between 12.00 and 14.00 h.
Measurement of corticosterone andadrenocorticotropic hormone (ACTH)Radioimmunoassay (RIA) of corticosterone was perfor-med using [125I]corticosterone, antiserum, and standardsolution in a kit from ICN Biomedical (Costa Mesa, CA,USA). The assay was adapted to rat serum conditions.Precipitation was done using a second antibody solidphase. ACTH was measured using a DSL kit (Webster,Texas, USA). Plasma samples contained 500 KIU apro-tinin ml−1. Both assays were performed according tomanufacturer’s instruction. The inter- and intraassaycoefficients of variance for ACTH were 10.6% and6.9%, respectively, with a detection limit of 10 pg ml−1.For corticosterone, the inter- and intraassay coef-ficients of variance were 7.2% and 4.4%, with a detec-tion limit of 25 ng ml−1.
In situ hybridization histochemistryGuided by Nissl-stained sections collected during thecutting and by the atlas of Paxinos and Watson,32 cor-onal frozen sections (15 �m-thick) were collected at thelevels of the midportion of the parvocellular region of

mRNA changes in the stress axis induced by St John’s wortV Butterweck et al
550
Molecular Psychiatry
the PVN at the level where the magnocellular nucleusis largest (−1.8 mm), bed nucleus of the stria terminalis(BST) (−0.26 mm), dorsal hippocampus (−3.3 mm),pituitary, and locus coeruleus (−9.7 mm). Sectionswere thaw-mounted onto gelatin-coated slides, dried,and stored at −40°C prior to processing for in situhybridization histochemistry.The in situ hybridization histochemistry procedures
were performed as described previously for ribonucle-otide (cRNA) probes.15 First, tissue sections were pro-cessed by fixation with 4% formaldehyde solution,acetylation with 0.25% acetic anhydride in 0.1 M tri-ethanolamine-HCI, pH 8.0 solution, dehydration withethanol, and delipidation with chloroform. Second, theantisense probes were transcribed from linearized plas-mids using the Riboprobe System (Promega Biotech,Madison, WI, USA) with 35S-UTP (specific activity�1000 Ci mmol−1; New England Nuclear, Boston, MA,USA) and T7, T3, or SP6 RNA polymerase. The cDNAprobes were: a 760-bp fragment of rat CRH (gift of DrJames Herman, University of Kentucky, Lexington, KY,USA), 923 bp of mouse POMC (gift of Dr James Doug-lass, Vollum Institute, Portland, OR, USA), 384 bp ofrat TH (gift of Dr Barry Kosofsky, Harvard MedicalSchool, Boston, MA, USA), 229 bp of rat arginine-vaso-pressin (AVP) (gift of Dr W Scott Young, NIMH,Bethesda, MD, USA), 2.3 kb and 3.2 kb full-lengthsequences of the rat GAD65 and GAD67 (gift of Drs ATobin and N Tillakaratne, UCLA, Los Angeles, CA,USA), rat full-length BDNF33 (gift of Drs J Lauterhornand C Gall, University of California Irvine, CA, USA),rat full-length CREB (gift of Dr Stephen Hyman, NIMH,Bethesda, MD, USA) and a 900-bp BAL I/PVU II frag-ment of the rat serotonin 5-HT1A receptor gene (gift ofDr Paul Albert, University of Ottawa, Ontario, Canada).The radiolabeled probes were diluted in a riboprobe
hybridization buffer and applied to brain sections(approximately 500 000 CPM per section). After over-night incubation at 55°C in a humidified chamber,slides containing brain sections were washed first in20 �g ml−1 RNase solution and then 1 h each in 2× SSC(50°C) and 0.2× SSC (55°C and 60°C) solutions toreduce non-specific binding of the probe. The slideswere then dehydrated with ethanol and air-dried forautoradiography.Slides and 14C plastic standards containing known
amounts of radioactivity (American Radiochemicals, StLouis, MO, USA) were placed in x-ray cassettes,apposed to film (BioMax MR, Kodak, Rochester, NY,USA) for periods ranging from 1 to 72 h, and developedin an automatic processor (X-OMAT, Kodak). To deter-mine anatomical localization of hybridized AVP cRNAat the cellular level, sections were dipped in nucleartrack emulsion (NTB-2, Kodak), exposed for 18 h,developed (D-19, Kodak), and lightly counterstainedwith cresyl violet.
Data analysis and presentationAutoradiographic images were digitized with a solid-state camera (CCD-72, Dage-MTI) and a Macintoshcomputer using NIH Image software (Wayne Rasband,
NIMH). Transmittance measurements were convertedto dpm mg−1 plastic using the calibration curve(Rodbard equation) generated from the standards.Brain structures were identified according to the atlasof Paxinos and Watson.32 Light transmittance throughthe film at PVN (CRH and AVP in the parvocellulardivision), BST (GAD65/67), hippocampus (5-HT1A,BDNF, CREB), anterior and intermediate lobes of pitu-itary (POMC), and locus coeruleus (TH) was measuredby outlining the structure with the mouse cursor.Mouse cursor control was used to outline the selectedstructure. The average value for each animal in experi-mental or control groups (based on four measurementsper animal) was used to calculate group means (n = 5–8 per group).
For analysis of induction of AVP mRNA in cellswithin the parvocellular portion of the PVN that pro-jects to the median eminence (the CRH-containing zoneof the PVNpc), emulsion-coated slides were analyzedwith a Leica DMR microscope. The silver grains in theemulsion layer were visualized with epi-illuminationpassing through a POL cube filter (which exclusivelyselects for polarized light reflected off the silver grains)and captured as a ‘darkfield’ image with the Dage-MTIcamera using NIH Image. The captured POL image wasinverted and background flattened using the 2D rollingball function. Density slice was then used to select onlylabeled cells, and the median eminence projecting zone(determined on the basis of the corresponding CRHmRNA image from the same animal) was outlinedusing the mouse cursor. A calibrated area measurementwas then made; this number was directly proportionalto the number of labeled cells in the PVNpc.
Two-way ANOVAs (time and drug treatment orstress and drug treatment) were used to compare spe-cific mRNA levels in control vs treated groups. TheStudent–Newman–Keuls test was used for post-hoccomparisons of mRNA levels in each region. A cri-terion level of P � 0.05 was used to determine signifi-cance.
Results
Experiment 1—short-term/long-term treatment studyExamples of the expression patterns of each of themRNA probes are shown in Figure 1. Densitometryshowed that CRH mRNA levels in the PVN were notsignificantly changed at 2 weeks (short-termadministration) but were significantly decreased at 8weeks (long-term administration) in all three druggroups examined (Table 1). CRH mRNA levels werereduced 20–22% by imipramine and hypericin (P �0.001) and 16% by Hypericum extract (P � 0.01) at 8weeks. No significant changes were observed formRNA expression of AVP, measured by both densito-metry (data not shown) and cell counting (Table 1), inthe parvocellular portion of the PVN at either timepoint.
After 2 weeks of oral treatment, imipramine andhypericin but not Hypericum extract significantlydecreased POMC mRNA expression in the anterior lobe

mRNA changes in the stress axis induced by St John’s wortV Butterweck et al
551
Figure 1 Examples of autoradiographic images, obtained in control animals, for the ribonucleotide probes hybridized in experi-ment 1. A darkfield microphotograph of an emulsion-coated section hybridized for arginine vasopressin (AVP) mRNA illustratesthe discrete cellular labeling in the magnocellular (condensed cluster of labeled cells) and parvocellular (scattered labeled cells)divisions of the hypothalamic paraventricular nucleus (PVN). A photograph of the film exposed to a corticotropin-releasinghormone (CRH) mRNA-hybridized section is shown at the same magnification. The zone of CRH mRNA labeling, in the parvocel-lular PVN, was used to determine the portion of the PVN that was outlined for AVP mRNA quantification. The remaining filmimages, captured at lower magnification, are examples of proopiomelanocortin (POMC) mRNA hybridization in the pituitary(long exposure is shown, used for measurement of the expression in the anterior lobe; a shorter exposure was used for measure-ment of the darkly labeled intermediate lobe); tyrosine hydroxylase (TH) mRNA in the locus coeruleus, and serotonin receptor1A(5-HT1A) mRNA in the hippocampus. Bars measure 0.5 mm.
Table 1 Expression of mRNAs in rat brain after short-term and long-term antidepressant treatment
Brain region/mRNA Control Imipramine Hypericum extract Hypericin
2 weeks 8 weeks 2 weeks 8 weeks 2 weeks 8 weeks 2 weeks 8 weeks
PVNCRH 100 ± 3 100 ± 3 98 ± 2 78 ± 3** 96 ± 2 84 ± 3** 96 ± 3 80 ± 3**AVP 100 ± 13 100 ± 7 87 ± 6 87 ± 10 73 ± 11 97 ± 5 87 ± 12 102 ± 10
PituitaryAnterior lobe, POMC 100 ± 5 100 ± 6 70 ± 8** 82 ± 9* 88 ± 11 75 ± 12* 68 ± 6** 78 ± 10*Intermediate lobe, 100 ± 18 100 ± 12 91 ± 7 84 ± 9 77 ± 9 78 ± 5 89 ± 14 91 ± 9POMC
Hippocampus5-HT1A
CA1 100 ± 3 100 ± 2 99 ± 2 83 ± 4** 98 ± 3 89 ± 3* 98 ± 3 89 ± 2**CA3 100 ± 4 100 ± 5 100 ± 5 81 ± 7 97 ± 2 89 ± 8 92 ± 4 87 ± 5DG 100 ± 3 100 ± 4 102 ± 3 88 ± 5 97 ± 2 90 ± 3 96 ± 4 96 ± 3
Locus coeruleusTH 100 ± 4 100 ± 4 86 ± 4 77 ± 2** 93 ± 6 89 ± 2 103 ± 7 107 ± 5
Values are mean ± SEM based on the average of four sections for each brain region per animal (n = 6–8 per group) and areexpressed as a percentage of the respective 2-week or 8-week control. Values significantly different from the respective 2-weekor 8-week control animals are expressed as *P � 0.05 and **P � 0.01, Student–Newman–Keuls post hoc test.For reference, raw values (DPM mg−1) for each of the control means are: CRH in PVN 2 wk 1734; 8 wk 1754.POMC in anterior lobe 2 wk 2212; 8 wk 2132; intermediate lobe 2 wk 23358; 8 wk 22572.5-HT1A in CA1 2 wk 943; 8 wk 927; CA3 2 wk 1064; 8 wk 1074; DG 2 wk 1321; 8 wk 1327.TH in LC 2 wk 4544; 8 wk 4397. AVP in PVNpc (sq �m) 2 wk 3835; 8 wk 4089.
Molecular Psychiatry

mRNA changes in the stress axis induced by St John’s wortV Butterweck et al
552
Molecular Psychiatry
of the pituitary (30–32%; P � 0.001) (Table 1). After 8weeks of chronic drug treatment, POMC mRNA levelswere reduced 18% by imipramine (P � 0.05), 25% byHypericum extract (P � 0.05), and 22% by hypericin(P � 0.05). No treatment effects were observed in theintermediate lobe of the pituitary.After 2 weeks of daily treatment, no significant
changes in 5-HT1A receptor mRNA levels were foundfor any of the drugs (Table 1). Long-term treatment (8weeks) with imipramine, Hypericum extract, andhypericin significantly decreased 5-HT1A receptormRNA expression by 11–17% (P � 0.001; P � 0.01)in hippocampal field CA1 relative to control (Table 1).Similar decreases were observed in CA3 and DG butwere not significant in these areas.No changes in TH mRNA levels were evident after 2
weeks for any treatment (Table 1). After 8 weeks ofdaily imipramine administration, TH gene expressionlevels were decreased by 23% (P � 0.001). Hypericumextract produced a slight (11%) but not significantdecrease in TH gene expression after 8 weeks, andhypericin did not alter TH mRNA levels.The decrease in POMC mRNA levels in the anterior
pituitary of animals treated with imipramine, Hyper-icum extract, and hypericin for 2 weeks was associatedwith a significant reduction of plasma ACTH (25–39%,P � 0.001) and corticosterone (51–57%, P � 0.001) lev-els (Table 2). No changes in hormone levels wereobserved after 8 weeks.Adrenal gland weights were not significantly altered
by 2 weeks or 8 weeks of chronic antidepressant treat-ment. Body weight was significantly decreased byimipramine (13%, P � 0.05) and Hypericum extract(12%, P � 0.05) after 2 weeks of daily treatment butwas not affected after 8 weeks. Hypericin slightlyincreased body weight (8%, P � 0.05) after 8 weeks(Table 2).
Experiment 2—stress plus antidepressant drugadministration
CRH and AVP mRNA in the PVNpc Neither acuteimipramine pre-treatment, acute immobilization stress,nor acute imipramine plus stress had a significanteffect on gene expression of CRH and AVP in the par-
Table 2 Plasma hormone levels and adrenal and body weights in short-term and long-term treatment groups
Control Imipramine Hypericum extract Hypericin
2 weeks 8 weeks 2 weeks 8 weeks 2 weeks 8 weeks 2 weeks 8 weeks
ACTH (pg ml−1) 224 ± 26 203 ± 14 138 ± 13** 182 ± 13 170 ± 9** 205 ± 13 155 ± 19** 182 ± 13Corticosterone (ng ml−1) 131 ± 26 148 ± 19 65 ± 11** 159 ± 18 62 ± 16** 201 ± 20 57 ± 7** 148 ± 14Adrenal gland weight 42.3 ± 2.7 49.7 ± 2.7 35.7 ± 1.7 43.8 ± 4.1 44.5 ± 1.8 52.4 ± 3.8 37 ± 3.1 44.8 ± 2.9(mg)Body weight 330 ± 11 390 ± 15 289 ± 9* 369 ± 7 291 ± 9* 372 ± 12 311 ± 8 419 ± 11*
Values expressed as mean ± SEM (n = 6–8). Initial body weights for the control animals were 160–200 g.Values significantly different from the respective 2-week or 8-week control animals are expressed as *P � 0.05 and **P � 0.01.
vocellular PVN (Table 3). A 28% non-significantincrease in AVP mRNA level was seen by densitometryof the PVNpc in the acute stress condition relative tocontrol (data not shown). In contrast, long-term admin-istration (7 weeks) of imipramine (15 mg kg−1 p.o.) orHypericum extract (500 mg kg−1 p.o.) significantlydecreased CRH mRNA levels in the PVN (Figures 2 and3), replicating the results of the first experiment. CRHmRNA levels were reduced 15–17% (P � 0.05) byimipramine and by Hypericum extract (P � 0.05). Inuntreated animals, chronic immobilization stress (2 hper day for 7 days) significantly increased CRH mRNAexpression (26%, P � 0.01) in the PVN (Figures 2b, 3),replicating earlier work.24 The stress-induced elevationof CRH mRNA expression was blocked by chronicimipramine administration (16%, P � 0.01) but not bythe extract (Figure 3). No changes were observed foreither expression density or numbers of cells express-ing AVP mRNA in the parvocellular PVN in any of thechronic conditions.
POMC mRNA levels in the pituitary Neither acuteimipramine pre-treatment, acute immobilization stress,nor acute imipramine plus stress had a significanteffect on gene expression of POMC in the anterior lobeof the pituitary (Table 3). In the intermediate lobe ofthe pituitary, acute imipramine treatment alone had noeffect on the POMC gene expression, whereas acutestress produced a slight but not significant increase inPOMC mRNA levels (Table 3).
Seven weeks of chronic administration of imipra-mine or Hypericum extract significantly reducedPOMC mRNA levels in the anterior lobe of the pituitaryof unstressed groups (Figure 4). POMC mRNA levelswere reduced 27% (P � 0.05) by imipramine and 20%(P � 0.05) by Hypericum extract. Repeated immobiliz-ation significantly increased POMCmRNA levels in theanterior lobe (50%, P � 0.01) of control animals (Figure4). The stress-induced elevated POMC mRNAexpression in the anterior lobe was blocked by treat-ment with imipramine (22%, P � 0.05) and Hypericumextract (26%, P � 0.05) (Figure 4).
Although less pronounced, similar effects wereobserved in the intermediate lobe of the pituitary.Chronic stress increased POMC mRNA expression to

mRNA changes in the stress axis induced by St John’s wortV Butterweck et al
553Table 3 Expression of mRNA in acute conditions
Brain region/mRNA Control Imipramine
1 2 3 4no stress acute stress no stress no stress
PVNCRH 100 ± 2 99 ± 3 110 ± 4 111 ± 2AVP 100 ± 14 109 ± 9 118 ± 15 101 ± 12
PituitaryAnterior lobe, POMC 100 ± 5 100 ± 6 94 ± 8 103 ± 9Intermediate lobe, POMC 100 ± 7 130 ± 5 100 ± 8 121 ± 7
BSTGAD 65anteromedial 100 ± 7 106 ± 2 100 ± 6 103 ± 6anterodorsal 100 ± 6 104 ± 4 96 ± 5 100 ± 4ventral 100 ± 8 122 ± 5 110 ± 7 114 ± 5GAD 67anteromedial 100 ± 4 108 ± 7 92 ± 5 102 ± 6anterodorsal 100 ± 3 105 ± 6 97 ± 7 99 ± 6ventral 100 ± 2 117 ± 5 96 ± 7 104 ± 6
1:2 P � 20.05Hippocampus
5-HT1A
CA1 100 ± 3 103 ± 3 102 ± 4 93 ± 5CA3 100 ± 7 120 ± 8 104 ± 8 97 ± 9
2:4 P � 0.05DG 100 ± 5 101 ± 4 95 ± 5 86 ± 4
2:4 P � 20.05BDNFCA1 100 ± 3 93 ± 2 92 ± 23 91 ± 3CA3 100 ± 2 98 ± 3 98 ± 2 93 ± 3DG 100 ± 5 68 ± 5 83 ± 5 58 ± 2
1:2 P � 0.01 1:3 P � 0.01 2:4 P � 0.053:4 P � 0.01
CREBCA1 100 ± 5 106 ± 4 98 ± 3 97 ± 5CA3 100 ± 4 102 ± 3 100 ± 4 97 ± 5DG 100 ± 5 107 ± 3 97 ± 4 96 ± 5
Locus coeruleusTH 100 ± 3 125 ± 1 102 ± 4 104 ± 7
1:2 P � 0.05 2:4 P � 0.05
Values are mean ± SEM based on the average of four sections for each brain region per animal (n = 5–8 per group) and areexpressed as a percentage of the no stress control (group 1).Values significantly different from the respective comparison group, Student–Newman–Keuls post hoc test, are shown in thecells. For reference, mean values (DPM mg−1) for the no stress control group are: CRH in PVN 1449; POMC in Ant. Lobe 782,Intermed. Lobe 26855; GAD65 in AM 2982, AD 3612, V 3868; GAD67 in AM 2801, AD 3447, V 3943; 5-HT1A in CA1 503, CA3440, DG 764; BDNF in CA1 635, CA3 1298, DG 1481; CREB in CA1 562, CA3 590, DG 759; TH in LC 2983. Value for AVP inPVNpc (sq �m) 3229.
124% of control (P � 0.05). This increase was pre-vented by long-term treatment with imipramine or theextract (Figure 4).
ACTH and corticosterone plasma levels Acute immo-bilization stress produced a significant elevation (P �0.05) of plasma ACTH and corticosterone levels com-pared to the non-stressed rats. Acute pre-treatmentwith imipramine had no effect on the stress-inducedincreases in plasma hormone levels. The ACTH values(pg ml−1) for the different treatment groups in the acute
Molecular Psychiatry
stress experiment were: vehicle no stress = 203 ± 38,imipramine no stress = 142 ± 14, vehicle + acute stress= 666 ± 63, imipramine + acute stress = 529 ± 15.Corticosterone values (ng ml−1) were: vehicle no stress= 83 ± 32, imipramine no stress = 94 ± 25, vehicle +stress = 459 ± 62, imipramine + stress = 573 ± 52.The chronic stress paradigm caused a significant
elevation of basal plasma hormone levels relative to thenon-stressed control group. The stress-inducedincreases in corticosterone plasma levels were signifi-cantly reduced in animals that were pre-treated with
mRNA changes in the stress axis induced by St John’s wortV Butterweck et al
554
Molecular Psychiatry
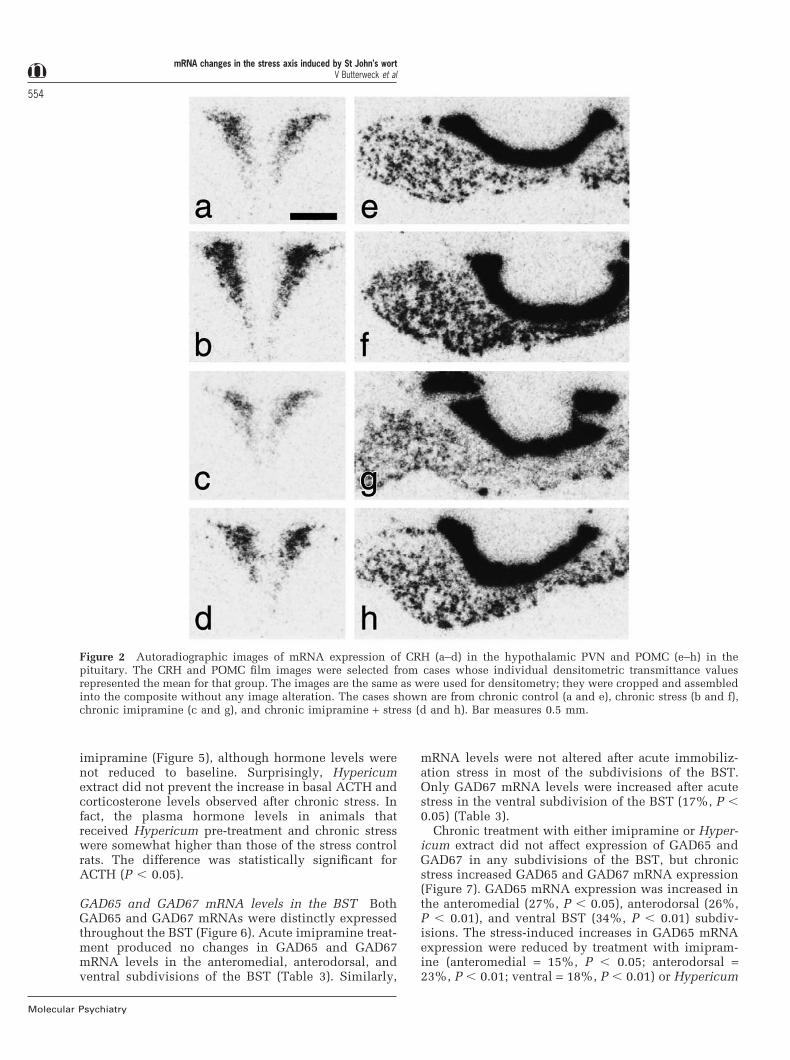
Figure 2 Autoradiographic images of mRNA expression of CRH (a–d) in the hypothalamic PVN and POMC (e–h) in thepituitary. The CRH and POMC film images were selected from cases whose individual densitometric transmittance valuesrepresented the mean for that group. The images are the same as were used for densitometry; they were cropped and assembledinto the composite without any image alteration. The cases shown are from chronic control (a and e), chronic stress (b and f),chronic imipramine (c and g), and chronic imipramine + stress (d and h). Bar measures 0.5 mm.
imipramine (Figure 5), although hormone levels werenot reduced to baseline. Surprisingly, Hypericumextract did not prevent the increase in basal ACTH andcorticosterone levels observed after chronic stress. Infact, the plasma hormone levels in animals thatreceived Hypericum pre-treatment and chronic stresswere somewhat higher than those of the stress controlrats. The difference was statistically significant forACTH (P � 0.05).
GAD65 and GAD67 mRNA levels in the BST BothGAD65 and GAD67 mRNAs were distinctly expressedthroughout the BST (Figure 6). Acute imipramine treat-ment produced no changes in GAD65 and GAD67mRNA levels in the anteromedial, anterodorsal, andventral subdivisions of the BST (Table 3). Similarly,
mRNA levels were not altered after acute immobiliz-ation stress in most of the subdivisions of the BST.Only GAD67 mRNA levels were increased after acutestress in the ventral subdivision of the BST (17%, P �0.05) (Table 3).
Chronic treatment with either imipramine or Hyper-icum extract did not affect expression of GAD65 andGAD67 in any subdivisions of the BST, but chronicstress increased GAD65 and GAD67 mRNA expression(Figure 7). GAD65 mRNA expression was increased inthe anteromedial (27%, P � 0.05), anterodorsal (26%,P � 0.01), and ventral BST (34%, P � 0.01) subdiv-isions. The stress-induced increases in GAD65 mRNAexpression were reduced by treatment with imipram-ine (anteromedial = 15%, P � 0.05; anterodorsal =23%, P � 0.01; ventral = 18%, P � 0.01) or Hypericum

mRNA changes in the stress axis induced by St John’s wortV Butterweck et al
555
Figure 3 CRH and AVP mRNA expression in the parvocellu-lar paraventricular nucleus (PVN) in the chronic conditions.Groups are: 1, chronic no stress; 2, chronic stress; 3, chronicimipramine; 4, chronic imipramine + stress; 5, chronic Hyper-icum extract; and 6, chronic Hypericum extract + stress. *P� 0.05; †P � 0.01.
Figure 4 POMC mRNA expression in the anterior and inter-mediate lobes of the pituitary in the chronic conditions.Groups are: 1, chronic no stress; 2, chronic stress; 3, chronicimipramine; 4, chronic imipramine + stress, 5, chronic Hyper-icum extract; and 6, chronic Hypericum extract + stress. *P� 0.05; †P � 0.01.
extract (anteromedial = 25% P � 0.01; anterodorsal =18%, P � 0.01; ventral = 22%, P � 0.01), respectively(Figure 7). Similarly, chronic stress-induced increasesin GAD67 mRNA expression levels were observed inthe anteromedial (24%, P � 0.05), anterodorsal (19%,P � 0.05) and ventral (26%, P � 0.01) subdivisions ofthe BST. After chronic treatment with imipramine orHypericum extract, stress-induced elevations inGAD67 mRNA levels were no longer apparent, and thestress-blocking effect was significant in the anterodor-sal and ventral subdivisions (Figure 7).
Molecular Psychiatry
Figure 5 Adrenocorticotropin (ACTH) and corticosteronelevels in plasma in the chronic conditions. Groups are: 1,chronic no stress; 2, chronic stress; 3, chronic imipramine; 4,chronic imipramine ± stress; 5, chronic Hypericum extract;and 6, chronic Hypericum extract + stress. *P � 0.05; †P �0.01.
5-HT1A mRNA levels in the hippocampus All cell lay-ers of the hippocampus expressed moderate levels of5-HT1A mRNA (Figure 8). Neither acute stress nor acuteimipramine had effects on 5-HT1A mRNA levels(Table 3).Long-term treatment with either imipramine or Hyp-
ericum extract caused a significant decrease in 5-HT1A
mRNA levels across all subfields of the hippocampusrelative to control (Table 4). The decreases were 17–32% in the unstressed imipramine group and 19–28%in the Hypericum group. Animals receiving chronicimmobilization stress also showed a decrease in 5-HT1A mRNA levels across all hippocampal subfields(Figure 8b). The decreases were 18–37% (P � 0.01) inthe stressed control group relative to unstressed con-trol. The stressed groups treated with imipramine orHypericum extract for 7 weeks had 5-HT1A mRNA lev-els that were not different from those of the untreatedstressed control group (Table 4).

mRNA changes in the stress axis induced by St John’s wortV Butterweck et al
556
Molecular Psychiatry
Figure 6 Autoradiographic image of GAD65 mRNAexpression in the bed nucleus of the stria terminalis (BST).A similar pattern and density of expression was seen forGAD67. The parcellation of BST was based on Bowers et al28
and the Paxinos and Watson atlas32 plates 19 and 20 as fol-lows: anteromedial BST (AM, encompassing atlas divisionsBSTMA); anterodorsal BST (AD, encompassing atlasdivisions BSTLD, BSTLJ, and BSTI), and ventral (V, BSTV).Bar measures 1 mm.
BDNF mRNA levels in the hippocampus All changesin BDNF mRNA expression in the hippocampus wereconfined to the dentate gyrus granule cell layer (Tables3 and 4). Both acute imipramine treatment and acuteimmobilization stress produced decreases in BDNFmRNA levels in the dentate gyrus relative to theunstressed control group (17%, P � 0.01 and 32%, P� 0.01, respectively) (Table 3). A single imipramineadministration prior to acute stress further down-regu-lated BDNF mRNA expression relative to the stressedcontrol (15%, P � 0.05).Similarly, both chronic antidepressant treatment as
well as chronic stress decreased BDNF mRNAexpression in the dentate gyrus. Chronic antidepress-ant treatment alone decreased BDNF mRNA levels by15–20% (P � 0.01) (Table 4). Chronic stress also mark-edly reduced mRNA expression, by 44% (P � 0.01)(Figure 8c, d). Chronic administration of imipramine orHypericum extract did not prevent the stress-induceddecrease in BDNF mRNA. In fact, BDNF levels werecomparable to those of the stressed control group(Table 4).
CREB mRNA levels in the hippocampus No signifi-cant change in CREB mRNA expression was seen in thehippocampus with the acute stress paradigm (Table 3).After long-term treatment with imipramine and Hyper-icum extract, CREB mRNA levels remained unchangedin all subdivisions of the hippocampus (Table 4). CREBmRNA expression was increased after chronic stress inthe CA1 field (16%, P � 0.05) (Figure 8e, f) and in thedentate gyrus (13%, P � 0.05). Chronic administrationof imipramine and Hypericum extract resulted in CREBmRNA levels in stressed animals that were close to thelevels of the unstressed control group (Table 4).
Figure 7 GAD65 and 67 mRNA expression in the bednucleus of the stria terminalis (BST) in the chronic con-ditions. BST subdivisions are anteromedial (AM), anterodor-sal (AD), and ventral (V). Groups are: 1, chronic no stress; 2,chronic stress; 3, chronic imipramine; 4, chronic imipramine+ stress; 5, chronic Hypericum extract; and 6, chronic Hyper-icum extract + stress. *P � 0.05; †P � 0.01.
TH mRNA levels in the locus coeruleus Acuteexposure to immobilization stress increased (25%, P �0.05) TH mRNA levels in the locus coeruleus (Table3). Acute treatment with imipramine alone had noeffect on TH gene expression. The stress-induced acuteincrease of TH mRNA levels was normalized by acutepre-treatment with imipramine (17%, P � 0.05) (Table3). Chronic exposure to immobilization stress signifi-cantly increased TH mRNA levels in the locus coeru-leus (19%, P � 0.05). Pre-treatment with imipraminebut not the plant extract normalized the stress-inducedelevation of TH mRNA (P � 0.01) (Figures 9 and 10).
Discussion
Experiment 1—chronic administration effectsThe major finding of the first study is that St John’swort (Hypericum extract) altered gene expression lev-els in brain areas involved in HPA axis control after

mRNA changes in the stress axis induced by St John’s wortV Butterweck et al
557
Figure 8 Autoradiographic film images of mRNA expression of 5-HT1A (a and b), BDNF (c and d), and CREB (e and f) in thehippocampus. Notice the differences in density of labeling between the chronic control (no stress) at the top with chronicstress at the bottom. Images were selected as described in Figure 1 caption. Hippocampal fields labeled are Cornu AmmonisCA1 and CA3 and the dentate gyrus (DG). Bar measures 1 mm.
Table 4 Expression of mRNA in the hippocampus in chronic conditions
Brain region/ Control Imipramine Hypericum extractmRNA
1 2 3 4 5 6no stress chronic stress no stress chronic stress no stress chronic stress
5-HT1A
CA1 100 ± 3 82 ± 4 83 ± 2 79 ± 5 79 ± 1 81 ± 31:2 P � 0.01 1:3 P � 0.05 1:5 P � 0.01
CA3 100 ± 4 63 ± 8 68 ± 4 70 ± 9 72 ± 9 78 ± 71:2 P � 0.01 1:3 P � 0.01 1:5 P � 0.05
DG 100 ± 5 81 ± 4 80 ± 4 69 ± 4 81 ± 4 77 ± 51:2 P � 0.01 1:3 P � 0.01 1:5 P � 0.01
BDNFCA1 100 ± 3 92 ± 4 91 ± 4 93 ± 3 91 ± 3 89 ± 5CA3 100 ± 3 98 ± 2 103 ± 2 98 ± 3 95 ± 4 88 ± 4DG 100 ± 2 56 ± 3 80 ± 5 55 ± 3 85 ± 4 53 ± 4
1:2 P � 0.01 1:3 P � 0.01 1:5 P � 0.01CREBCA1 100 ± 3 116 ± 3 100 ± 5 114 ± 4 104 ± 2 106 ± 6
1:2 P � 0.05CA3 100 ± 4 105 ± 3 90 ± 4 101 ± 3 94 ± 2 94 ± 6DG 100 ± 5 113 ± 3 94 ± 5 107 ± 4 99 ± 3 100 ± 6
1:2 P � 0.05
Values are mean ± SEM based on the average of four sections for each brain region per animal (n = 5–8 per group) and areexpressed as a percentage of the no stress control (group 1). Values significantly different (Student–Newman–Keuls post hoctest) from the respective comparison group are designated in the cells.
long-term administration, similar to the changes elic-ited by the prototypic synthetic antidepressant imipra-mine—a finding that has not been previously reportedin an in vivo model. The neurochemical actions of Hyp-ericum extract could also be demonstrated with purehypericin. It appears, therefore, that this naphthodi-anthrone is a major active principle of St John’s wortthat may contribute to its therapeutic effect. Hypericinis a stable compound that is convenient and inexpen-
Molecular Psychiatry
sive to bioextract, making it particularly attractive forfurther exploitation and characterization for potentialuse in CNS mood disorders.The drug administration paradigm and the selection
of molecular targets was as described by Brady et al.15
All three drugs—Hypericum extract, hypericin, andimipramine—decreased CRH mRNA expression in thehypothalamus after long-term (8 weeks) but not short-term (2 weeks) administration. The imipramine-

mRNA changes in the stress axis induced by St John’s wortV Butterweck et al
558
Molecular Psychiatry
Figure 9 Autoradiographic film images of TH mRNA expression in the locus coeruleus (LC) on the right side of the brain.The three conditions shown are chronic control (a), chronic stress (b), and chronic imipramine + stress (c). Images were selectedas described in Figure 1 caption. Insets show the same image after blurring in NIH Image and coding with a five-shades-of-gray scale in order to highlight the density differences. All images were manipulated identically. Bar measures 0.2 mm.
Figure 10 TH mRNA expression in the locus coeruleus inthe chronic conditions. Groups are: 1, chronic no stress; 2,chronic stress; 3, chronic imipramine; 4, chronic imipramine+ stress; 5, chronic Hypericum extract; and 6, chronic Hyper-icum extract + stress. *P � 0.05; †P � 0.01.
induced delayed decrease in CRH mRNA levels in thePVN reported previously15 was with 5 mg kg−1 givenintraperitoneally, whereas in the present study 15 mgkg−1 were given orally by gavage. The time-dependentdecrease in CRH mRNA levels was also induced byfluoxetine, idazoxan and phenelzine in the same para-digm.16 Thus, Hypericum, hypericin, and several syn-thetic antidepressants act in a similar delayed fashionto affect expression levels of genes controlling activityof the HPA axis.Imipramine and hypericin significantly reduced
POMCmRNA levels in the anterior lobe of the pituitaryat both 2 and 8 weeks. Inconsistent effects have beenreported in previous studies measuring POMC mRNAlevels after administration with imipramine, fluoxet-
ine, idazoxan, fluoxetine, and citalopram.15,16,34 It issurprising that at 2 weeks, POMC mRNA and plasmaACTH and corticosterone levels were decreased whileCRH and AVP mRNA expression levels wereunchanged. At 8 weeks, anterior pituitary POMCmRNA levels and hypothalamic CRH mRNA levelswere reduced, and plasma ACTH and corticosteronelevels were back to baseline. The functional conse-quences of these findings are not clear. The relativelyhigh baseline levels of ACTH might be a reason forthese dissociative effects after 2 and 8 weeks. It can bespeculated that different adrenal responsiveness afterboth time points might play an important role inmediating the ACTH effects.
5-HT has long been implicated in the biological basisof depression as well as in the mechanism of actionof antidepressant drugs.35 Animal studies suggest thatincreases in neurotransmission at postsynaptic 5-HT1A
receptors may mediate the therapeutic effects of someantidepressant drugs.36 The 5-HT1A receptor has beenidentified as an inhibitory somatodendritic autorecep-tor in the raphe serotonergic cells and as a postsynapticreceptor in serotonergic terminal fields.37 The highestdensity of 5-HT1A receptor binding sites occurs in thehippocampus and other limbic regions.38 A role forhippocampal 5-HT1A in the pathophysiology of mooddisorders has been explored, but data from postmortemhuman brain studies have been inconsistent, showingunchanged,39 increased,40 or reduced 5-HT1A receptorbinding in the hippocampus of patients with majordepressive disorders who died by suicide.29
We found that 2-week imipramine administrationhad no effect on 5-HT1A receptor mRNA expression inthe hippocampus, consistent with Lopez et al.29 How-ever, long-term treatment with all three agents signifi-cantly decreased 5-HT1A receptor mRNA expression inCA1 of the hippocampus. Other studies have shownsmall decreases41,42 in 5-HT1A receptor number after

mRNA changes in the stress axis induced by St John’s wortV Butterweck et al
559long-term antidepressant administration. The mech-anisms responsible for the delayed decrease in 5-HT1A
receptor mRNA expression after daily treatment withHypericum extract, and hypericin, and imipramine areunknown. It can be speculated that alterations in cen-tral catecholaminergic, serotonergic and/or cholinergicsystems are involved. Chronic antidepressants increaseserotonergic neurotransmission in the hippocampus43
and elevate extracellular 5-HT concentrations in thenerve endings,44 possibly causing a postsynaptic down-regulation of the 5-HT1A receptors.Long-term treatment with imipramine significantly
decreased TH mRNA levels in the locus coeruleus,whereas Hypericum extract and hypericin had no effecton TH message. Previous studies also showed thatchronic imipramine treatment decreased THexpression,15,45 whereas chronic fluoxetine, idazoxanand phenelzine increased TH mRNA levels.16 Brady etal16 suggested that the therapeutic efficacy of activatingdrugs such as fluoxetine and phenelzine in the treat-ment of atypical depression may result from theirability to increase mRNA levels of TH in the locuscoeruleus. Because Hypericum extract and hypericinhad no effect on TH mRNA levels, it can be speculatedthat the natural substances have an action profile thatis different from that of classical antidepressants.
Experiment 2—actions in a stress paradigmAnalysis of relative hybridization densities demon-strated that chronic stress more so than acute stressinduced pronounced changes in levels of geneexpression of stress-related molecules and that chronicoral pre-treatment with imipramine or St John’s wort(Hypericum extract) countered a number of thesechanges in an anatomically specific manner. The stressaxis, comprised of the CNS nuclei and pathways thatcontrol glucocorticoid secretion elicited by psychologi-cal stressors such as immobilization, is principallycentered on the hypothalamic parvocellular PVN andits neurotransmitters CRH and AVP. Neural circuitsthat control PVN CRH activity have been demonstratedby tract-tracing, lesion, and electrophysiology studiesand by studies showing gene or protein regulation inthe afferent pathways (summarized in Herman andCullinan46). Acute and/or repeated stress elevates lev-els of transmitters, enzymes, and/or immediate-earlygenes in the PVN and its major afferent control centerssuch as the brainstem catecholamine cell groups, theamygdala, and the BST. Inputs from hippocampus andprefrontal cortex may inhibit the stress response. Wefocused our efforts on well-studied stress-responsivebrain areas involved in HPA axis control, but it is likelythat changes in gene expression levels occurred inother neurotransmitter systems and locations.
CRH and AVP mRNA expression In this study,chronic immobilization stress increased CRH geneexpression in the PVN. The stress-induced increase inCRH mRNA levels was blocked by long-term (7 weeks)pre-treatment with imipramine but not Hypericumextract. AVP mRNA density and numbers of expressing
Molecular Psychiatry
cells in the PVNpc were not affected by either stress orby antidepressant treatments. It is well established thatthe levels of CRH mRNA in the PVNpc increase inresponse to stress.24,47–50 However, the situation is lessclear for AVP mRNA, with some studies showing noresponse and others showing a small increase either inmessage levels or in numbers of cells expressing mess-age.25,48,50–54 The different transcript responses of CRHand AVP to stress imply different regulatory mech-anisms for these genes.55,56 Evidence for progressivechanges in AVP and CRH mRNA levels after repeatedstress has been reported.57
Our present data suggest that imipramine but notHypericum extract exerts its antidepressant activity byblocking the stress-induced changes in CRH geneexpression, which might be relevant to its therapeuticactions. This interpretation is tempered by the factsthat we do not know whether changes in CRH mRNAexpression levels are reflected in changes in theamount of CRH released from nerve terminals, orwhether these changes are affecting the hormonal out-put of the pituitary and adrenal glands. We also cannotexclude the possibility that isolated substances of Hyp-ericum might have stress-blocking properties that aremasked in the whole preparation.
POMC mRNA levels in the pituitary and plasma hor-mone levels The elevation in CRH mRNA levels afterrepeated immobilization stress in the PVN wasaccompanied by significant elevation of pituitaryPOMC mRNA levels in the anterior pituitary, asreported previously.24 Chronic stress-related changesin POMC mRNA levels in the anterior pituitary wereaccompanied by significant elevations of plasma ACTHand corticosterone levels. When imipramine and Hyp-ericum extract were administered concomitantly withchronic immobilization stress, up-regulation of POMCmRNA levels was prevented. These observations are ingood correlation with our previous findings.Although both antidepressants alone reduced POMC
gene expression in the anterior pituitary, plasma ACTHand corticosterone levels were not different from con-trol. The same pattern of data occurred in experiment1. The functional consequences of these findings arenot clear. The relatively high basal levels of ACTHmight be a reason for the dissociative effects afterchronic treatment. It can be also speculated that differ-ent adrenal responsiveness after chronic treatmentmight feed back to affect ACTH output.Immobilization stress induced a strong ACTH and
corticosterone response. Long-term administration ofimipramine reduced stress-induced increases incorticosterone plasma levels, a finding which wasreported recently.29 However, Hypericum extract wasunable to prevent the stress-induced elevation inplasma hormone levels. A similar result was observedfor zimelidine and fluoxetine, two specific serotoninreuptake inhibitors (SSRIs).29 Hypericum might havean influence on the serotonergic system comparable tothat of the SSRIs. There is evidence from the literaturethat different compounds from Hypericum differen-

mRNA changes in the stress axis induced by St John’s wortV Butterweck et al
560
Molecular Psychiatry
tially affect neurotransmitter systems, warrantingfurther study of individual extracts.
GAD65/67 mRNA expression There is an increasingbody of evidence suggesting that GABA plays animportant role in the therapeutic effects ofantidepressant/antipanic drugs. In clinical studies, thelevels of GABA in plasma have been found to be sig-nificantly lower in depressed patients than in con-trols.17,18 GABA agonists have been demonstrated topossess antidepressant and antipanic effects in animaland clinical studies.19,20 Phenelzine, a non-selectivemonoamine oxidase inhibitor, is efficacious in thetreatment of depression and panic disorder, and it hasbeen shown to elevate brain GABA levels.58,59 Simi-larly, imipramine has been reported to acutely enhanceGABA release in the brain.60
We found that neither of the long-term treatmentsgave rise to significant changes in levels of mRNAencoding GAD65 and GAD67 in the BST, one of themajor sources of afferent inputs to the PVN.61 Ourresults are in good correlation with the results of Laiet al62 who observed no changes in levels of GAD65and GAD67 mRNAs in cortex after short-term and long-term phenelzine and imipramine treatment. Therefore,it appears that antidepressants elevate GABA levelswithout altering mRNA levels of GAD65 and GAD67in the places measured.GABA is known to inhibit the release of ACTH and
corticosterone in vivo and reduce CRH release fromhypothalamic explants.63 It has been noted that PVNprojecting GABAergic BST cell populations are part ofa feedback circuit that serves to inhibit the stressresponses, suggesting that these cell groups should beactivated by stressful stimuli.28 Stress-induced GAD65and GAD67 mRNA increases in the BST subdivisionscomport with the results of Bowers et al.28
Interestingly, the stress-induced increases in GAD65and GAD67 gene expression were reduced by long-term pre-treatment with either imipramine or Hyper-icum extract. Whereas a decrease in the inhibition ofthe PVN by the BST seems paradoxical given that theCRH mRNA levels in the PVN are also decreased byantidepressants, a overall return to homeostasis may bethe more important aspect of the consequence ofchronic administration. The data suggest that inhi-bition of the HPA axis by a circuit originating in thehippocampus and relayed through the BST is not cru-cial for restraining the stress response during chronicantidepressant administration. Alternatively, changesin activity of local GABAergic interneurons in thehypothalamus28 may ‘reverse’ the sign of the BST inputto the PVN.
5-HT1A mRNA expression Confirming experiment 1,long-term treatment with imipramine and Hypericumextract significantly decreased 5-HT1A receptor mRNAexpression in the hippocampus. Chronic but not acuteimmobilization stress also reduced levels of 5-HT1A
mRNA. The decrease in 5-HT1A gene expression is con-sistent across various stress experiments.29,64 Imipra-
mine and Hypericum extract were unable to preventthe stress-induced decrease in 5-HT1A mRNA levels inthe hippocampus. Our results are different from thoseof Lopez et al29 who showed that imipramine adminis-tered concomitantly with chronic unpredictable stressprevented the reduction in 5-HT1A mRNA expression.The authors suggest that this effect might be steroid-mediated because elevated levels of glucocorticoidshave been shown to downregulate 5-HT1A mRNAexpression and binding in the hippocampus,65 andimipramine was able to decrease corticosterone levelsin chronically stressed rats. In our study, imipraminewas also able to reduce the stress-induced increase incorticosterone levels, casting doubt on this expla-nation. It is possible that differences in the nature ofthe stressor may underlie the different outcomes.
Imaging studies of primary mood disorder haveshown a decrease in 5-HT1A binding capacity in thehuman mesiotemporal cortex.66,67 In the study by Sarg-ent et al,67 antidepressants did not restore thereduction.
CREB and BDNF mRNA expression Recent work hasraised the possibility that affective disorders may havean organic component that involves cell survival.68
Among the many long-term targets of antidepressanttreatments, neurotrophins may play an important role.Neurotrophins promote the growth and developmentof immature neurons and enhance the survival andfunction of adult neurons.69 Therefore it can be specu-lated that antidepressants mediate their actions par-tially via BDNF, the most abundant neurotrophin in thebrain. Stress-induced atrophy of hippocampal neuronsmay contribute to the loss of hippocampal control ofthe HPA axis and hypercortisolism that often occurs indepression.70,71 It has been proposed that BDNFexpression in the hippocampus may oppose the dam-age and may be part of a critical link between stress-induced hypercortisolism and antidepressant drugactions.72 Finally, BDNF mRNA expression is regulatedby CREB,73 whose activity could thereby underlie someof the long-term effects of antidepressant treatment.74
In this study, acute immobilization stress decreasedlevels of BDNF mRNA selectively in the dentate gyrus,confirming the results of Smith et al.75 Chronic stressalso reduced BDNF gene expression selectively in thedentate gyrus; no effects were observed in the CA fieldsof the hippocampus. Chronic but not acute stressincreased CREB mRNA levels in CA1 and the dentategyrus. Therefore, the two markers were not linked.
Surprisingly, antidepressant treatment alone (acutelyand chronically, with imipramine and Hypericum) alsodecreased BDNF mRNA levels selectively in the den-tate gyrus. In contrast, Nibuya et al72 showed that theantidepressants imipramine, desipramine, sertraline,and mianserin significantly increased BDNF geneexpression in the hippocampus after a treatment periodof 21 days, whereas Kuroda et al76 found no effect onBDNF mRNA levels after chronic tianeptine treatment.One reason for those discrepancies might be the useof different treatment paradigms. In our present study,

mRNA changes in the stress axis induced by St John’s wortV Butterweck et al
561antidepressants were administered over a period of 7weeks, whereas Nibuya et al72 treated rats for 3 weeks.Alternatively, dose differences might underlie theopposite outcomes. Nibuya et al72 administered imi-pramine at a dose of 15 mg kg−1 i.p., whereas weadministered 15 mg kg−1 by gavage. The comparativelyhigh i.p. dose could have side effects that induce BDNFmRNA expression. Further studies are necessary toclarify the role of antidepressants on BDNF geneexpression and the relationship between mRNA levelsand neurotrophin function vis-a-vis cell proliferation,maintenance, and survival.Long-term antidepressant pre-treatment had no effect
on the stress-induced decrease in BDNF geneexpression in the dentate gyrus. Nibuya et al72 showedthat long-term pre-treatment (21 days) with severalclasses of antidepressants was able to antagonize thesignificant decrease in BDNF mRNA levels after acute(45 min) restraint stress. Together with the data fromour study, it can be speculated that chronic antide-pressant pre-treatment can prevent the decrease ofBDNF gene expression after acute but not chronicstress.It has been shown that administration of cortico-
sterone (10 mg per rat) reduced the level of BDNFmRNA in the dentate gyrus.75 In our present study, glu-cocorticoids were significantly elevated after acute andchronic stress. Although chronic treatment with imi-pramine was able to reduce the increase in plasmacorticosterone, baseline levels were not reached. Pre-treatment with Hypericum extract even increasedplasma hormone levels. Therefore our data suggest thatthe decreased induction of BDNF mRNA in the dentategyrus after chronic stress is mediated via elevated glu-cocorticoid levels.In the present study, none of the antidepressants
alone had a significant effect on CREB mRNA in thehippocampus, whereas chronic but not acute stressslightly increased its expression. The increasedexpression was not maintained in the stressed animalsreceiving chronic imipramine or plant extract. Thefunctional significance of this small effect is not clear,though it supports the hypothesis that hippocampalactivity, as reflected by CREB mRNA expression, ispositively linked to the activity of the HPA axis.Nibuya et al74 showed that chronic (21 days) admin-
istration of different types of antidepressant drugs sig-nificantly increased levels of CREB mRNA in the rathippocampus. Moreover, increased expression of CREBmRNA was observed after 10 days, but not 3 days, ofantidepressant treatment, suggesting that upregulationof CREB is dependent on repeated antidepressant treat-ment. However, we did not see any drug-induced upre-gulation at 7 weeks. Therefore, it can be speculated thatincreases in CREB mRNA levels are related to shorter-term or high-dose effects of antidepressants and thatthey are not of important relevance for the long-termtreatment with therapeutic doses.
TH mRNA expression The norepinephrine-locuscoeruleus and CRH-PVN systems often respond in
Molecular Psychiatry
similar directions in response to acute and chronicstress and to chronic antidepressant drug treatment(reviewed in Brady77). The results of the present studyconfirm previous reports that chronic stress and antide-pressant treatments regulate the norepinephrine sys-tem by increasing or decreasing, respectively, theexpression of TH in the locus coeruleus.24,45,78 More-over, the data demonstrate that the increased THmRNA levels in response to chronic stress could benormalized by pre-treatment with imipramine for 7weeks. Interestingly, chronic pre-treatment with Hyp-ericum extract again had no effect on TH geneexpression alone (as in experiment 1) or in combi-nation with chronic stress, suggesting that the naturalantidepressant might have a unique mode of action anda unique therapeutic indication.Norepinephrine is a potent stimulus of hypothalamic
CRH release.79 Because tricyclic antidepressantsdecrease the firing rate of locus coeruleus neurons,77
the imipramine-induced decrease in TH geneexpression may be involved in mediating the decreasein CRH gene expression in the PVN. However, the PVNand the locus coeruleus are only weakly linked ana-tomically, and a dissociation between gene expressionactivity in the locus coeruleus (TH) and the PVN (CRH)has been shown in the chronic drug administrationparadigm. The ability of antidepressant treatments tomaintain control levels of TH in the presence of stressmay be relevant to their therapeutic action.
Summary
Long-term but not short-term daily administration ofall three drugs decreased mRNA expression of CRHmRNA in the hypothalamic PVN. The delayed actionof the drugs on the key central determinant of HPAaxis activity replicates the earlier work15,16 and vali-dates this model as a test of antidepressant drugactions relevant to clinical efficacy. Long-termadministration of the three drugs also reduced POMCmRNA levels in the anterior lobe of the pituitary,though plasma hormone levels of ACTH and cortico-sterone returned to control levels. These findings areconsistent with the concept of a resetting of basalactivity in the HPA axis by chronic antidepressantdrugs. Long-term administration of all three drugsdecreased mRNA levels of the serotonin 5-HT1A
receptor in the hippocampus, suggesting commonactions on aspects of serotonin transmission. Thetested drugs had different effects on TH mRNAexpression in the locus coeruleus. The individualneurochemical profiles of imipramine, Hypericum(St John’s wort), and hypericin may be relevant totheir particular therapeutic efficacies in mood dis-orders.Repeated immobilization stress produced a persist-
ent alteration of mRNA expression of CRH in thePVN, POMC in the anterior pituitary, TH in the locuscoeruleus, GAD65/67 in the BST, and CREB, BDNFand 5-HT1A in the hippocampus; and it increasedplasma hormone levels of ACTH and corticosterone.

mRNA changes in the stress axis induced by St John’s wortV Butterweck et al
562
Molecular Psychiatry
Long-term pre-treatment with either imipramine or StJohn’s wort reduce to control levels the stress-induced increases in gene expression of GAD65 andGAD67 in the BST and POMC in the anterior pitu-itary. The stress-induced decreases in BDNF and 5-HT1A mRNA levels in hippocampus were not pre-vented by either imipramine or St John’s wort. Thestress-induced increases in mRNA levels of CRH inthe PVN and TH in the locus coeruleus were reducedby imipramine but not by the plant extract, indicatingthat St John’s wort only partially affects the stress cir-cuits. Our results, therefore, show that there are dif-ferences between the tricyclic antidepressant imipra-mine and the ‘natural’ herbiceutical St John’s wort intheir ability to modify the effects of chronic stress.Imipramine appears to be more effective in blockingstress effects on the HPA axis, but both drugs countergene expression changes induced in select nuclei ofthe brain’s stress circuitry. Imipramine, therefore,may be a more effective antidepressant drug fordepressive disorders that share features with chronicstress and activation of the HPA axis.
Acknowledgements
Funding was supplied by the NIMH IntramuralResearch Program and by Lichtwer Pharma (for VB).
References
1 Wheatly D. Hypericum extract—potential in the treatment ofdepression. CNS Drugs 1998; 9: 431–440.
2 Philipp M, Kohnen R, Hiller KO. Hypericum extract versus imipra-mine or placebo in patients with moderate depression: randomisedmulticentre study of treatment for eight weeks. Br Med J 1999; 319:1534–1538.
3 Butterweck V, Wall A, Lieflander-Wulf U, Winterhoff H, NahrstedtA. Effects of the total extract and fractions of Hypericum perfor-atum in animal assays for antidepressant activity. Pharmacopsych-iatry 1997; 30 Suppl 2: 117–124.
4 Winterhoff H, Butterweck V, Nahrstedt A, Gumbinger H, Schulz V,Erping S et al Pharmakologische. Untersuchungen zur antidepress-iven Wirkung von Hypericum perforatum L. In: Loew D, RietbrockN (eds). Phytopharmaka in Forschung und klinischer Anwendung.Steinkopff Verlag: Darmstadt, 1995, pp 39–56.
5 Muller WE, Rolli M, Schafer C, Hafner U. Effects of hypericumextract (LI 160) in biochemical models of antidepressant activity.Pharmacopsychiatry 1997; 30 Suppl 2: 102–107.
6 Simmen U, Burkard W, Berger K, Schaffner W, Lundstrom K.Extracts and constituents of Hypericum perforatum inhibit thebinding of various ligands to recombinant receptors expressed withthe Semliki Forest virus system. J Recept Signal Transduct Res1999; 19: 59–74.
7 Nahrstedt A, Butterweck V. Biologically active and other chemicalconstituents of the herb of Hypericum perforatum L. Pharmacopsy-chiatry 1997; 30 Suppl 2: 129–134.
8 Chatterjee SS, Bhattacharya SK, Wonnemann M, Singer A, MullerWE. Hyperforin as a possible antidepressant component of hyper-icum extracts. Life Sci 1998; 63: 499–510.
9 Singer A, Wonnemann M, Muller WE. Hyperforin, a major antide-pressant constituent of St John’s Wort, inhibits serotonin uptakeby elevating free intracellular Na+. J Pharmacol Exp Ther 1999; 290:1363–1368.
10 Butterweck V, Petereit F, Winterhoff H, Nahrstedt A. Solubilizedhypericin and pseudohypericin from Hypericum perforatum exertantidepressant activity in the forced swimming test. Planta Medica1998; 64: 291–294.
11 Butterweck V, Jurgenliemk G, Nahrstedt A, Winterhoff H. Flavono-ids from Hypericum perforatum show antidepressant activity in theforced swimming test. Planta Med 2000; 66: 3–6.
12 Calapai G, Crupi A, Firenzuoli F, Costantino G, Inferrera G, CampoGM et al. Effects of Hypericum perforatum on levels of 5-hydroxy-tryptamine, noradrenaline and dopamine in the cortex, diencepha-lon and brainstem of the rat. J Pharm Pharmacol 1999; 51: 723–728.
13 Butterweck V, Nahrstedt A, Evans J, Rauser L, Savafe J, Popadak Bet al. In vitro receptor screening of pure constituents of St John’swort reveals novel interactions with a number of GPCR’s. Soc Neu-rosci Abs 2001; 27: (in press).
14 Butterweck V, Korte B, Winterhoff H. Pharmacological and endo-crine effects of Hypericum perforatum and hypericin after repeatedtreatment. Pharmacospsychiatry 2001; (in press).
15 Brady LS, Whitfield H, Jr, Fox RJ, Gold PW, Herkenham M. Long-term antidepressant administration alters corticotropin-releasinghormone, tyrosine hydroxylase, and mineralocorticoid receptorgene expression in rat brain. Therapeutic implications. J Clin Invest1991; 87: 831–837.
16 Brady LS, Gold PW, Herkenham M, Lynn AB, Whitfield H Jr. Theantidepressants fluoxetine, idazoxan and phenelzine alter cortico-tropin-releasing hormone and tyrosine hydroxylase mRNA levelsin rat brain: therapeutic implications. Brain Res 1992; 572: 117–125.
17 Petty F. Plasma concentrations of gamma-aminobutyric acid(GABA) and mood disorders: a blood test for manic depressive dis-ease? Clin Chem 1994; 40: 296–302.
18 Plaznik A, Palejko W, Stefanski R, Kostowski W. Open fieldbehavior of rats reared in different social conditions: the effects ofstress and imipramine. Pol J Pharmacol 1993; 45: 243–252.
19 Lloyd KG, Zivkovic B, Scatton B, Morselli PL, Bartholini G. Thegabaergic hypothesis of depression. Prog NeuropsychopharmacolBiol Psychiatry 1989; 13: 341–351.
20 Breslow MF, Fankhauser MP, Potter RL, Meredith KE, Misiaszek J,Hope DG Jr. Role of gamma-aminobutyric acid in antipanic drugefficacy. Am J Psychiatry 1989; 146: 353–356.
21 Holsboer F, Barden N. Antidepressants and hypothalamic-pitu-itary-adrenocortical regulation. Endocr Rev 1996; 17: 187–205.
22 Barden N, Reul JM, Holsboer F. Do antidepressants stabilize moodthrough actions on the hypothalamic-pituitary-adrenocortical sys-tem? Trends Neurosci 1995; 18: 6–11.
23 Heuser I, Bissette G, Dettling M, Schweiger U, Gotthardt U,Schmider J et al. Cerebrospinal fluid concentrations of corticotro-pin-releasing hormone, vasopressin, and somatostatin in depressedpatients and healthy controls: response to amitriptyline treatment.Depress Anxiety 1998; 8: 71–79.
24 Mamalaki E, Kvetnansky R, Brady LS, Gold PW, Herkenham M.Repeated immobilization stress alters tyrosine hydroxylase, corti-cotropin-releasing hormone, and corticosteroid receptor mRNAlevels in rat brain. J Neuroendocrinol 1992; 4: 689–699.
25 Herman JP, Adams D, Prewitt C. Regulatory changes in neuroen-docrine stress—integrative circuitry produced by a variable stressparadigm. Neuroendocrinology 1995; 61: 180–190.
26 Sawchenko PE, Arias CA, Mortrud MT. Local tetrodotoxin blockschronic stress effects on corticotropin-releasing factor and vaso-pressin messenger ribonucleic acids in hypophysiotropic neurons.J Neuroendocrinol 1993; 5: 341–348.
27 Watanabe Y, McKittrick CR, Blanchard DC, Blanchard RJ, McEwenBS, Sakai RR. Effects of chronic social stress on tyrosinehydroxylase mRNA and protein levels. Brain Res Mol Brain Res1995; 32: 176–180.
28 Bowers G, Cullinan WE, Herman JP. Region-specific regulation ofglutamic acid decarboxylase (GAD) mRNA expression in centralstress circuits. J Neurosci 1998; 18: 5938–5947.
29 Lopez JF, Chalmers DT, Little KY, Watson SJ. Regulation of sero-tonin1A, glucocorticoid, and mineralocorticoid receptor in rat andhuman hippocampus: implications for the neurobiology ofdepression. Biol Psychiatry 1998; 43: 547–573.
30 Wolfensohn S, Lloyd M. Handbook of Laboratory Animal Manage-ment and Welfare. Oxford University Press: Oxford, 1994.
31 Kvetnansky R, Sun CL, Lake CR, Thoa N, Torda T, Kopin IJ. Effectof handing and forced immobilization on rat plasma levels of epi-nephrine, norepinephrine, and dopamine-�-hydroxylase. Endo-crinology 1978; 103: 1868–1874.

mRNA changes in the stress axis induced by St John’s wortV Butterweck et al
56332 Paxinos G, Watson C. The Rat Brain in Stereotaxic Coordinates,4th edn. Academic Press: San Diego, 1998.
33 Isackson PJ, Huntsman MM, Murray KD, Gall CM. BDNF mRNAexpression is increased in adult rat forebrain after limbic seizures:temporal patterns of induction distinct from NGF. Neuron 1991; 6:937–948.
34 Jensen JB, Jessop DS, Harbuz MS, Mork A, Sanchez C, MikkelsenJD. Acute and long-term treatments with the selective serotoninreputake inhibitor citalopram modulate the HPA axis activity atdifferent levels in male rats. J Neuroendocrinol 1999; 11: 465–471.
35 Meltzer HY. Role of serotonin in depression. Ann N Y Acad Sci1990; 600: 486–499.
36 Welner SA, De Montigny C, Desroches J, Desjardins P, Suranyi-Cadotte BE. Autoradiographic quantification of serotonin1A recep-tors in rat brain following antidepressant drug treatment. Synapse1989; 4: 347–352.
37 Hall MD, el Mestikawy S, Emerit MB, Pichat L, Hamon M, GozlanH. [3H]8-hydroxy-2-(di-n -propylamino)tetralin binding to pre- andpostsynaptic 5-hydroxytryptamine sites in various regions of therat brain. J Neurochem 1985; 44: 1685–1696.
38 Marcinkiewicz M, Verge D, Gozlan H, Pichat L, Hamon M. Auto-radiographic evidence for the heterogeneity of 5-HT1 sites in therat brain. Brain Res 1984; 291: 159–163.
39 Stockmeier CA, Dilley GE, Shapiro LA, Overholser JC, ThompsonPA, Meltzer HY. Serotonin receptors in suicide victims with majordepression. Neuropsychopharmacology 1997; 16: 162–173.
40 Arango V, Underwood MD, Gubbi AV, Mann JJ. Localized alter-ations in pre- and postsynaptic serotonin binding sites in the ven-trolateral prefrontal cortex of suicide victims. Brain Res 1995; 688:121–133.
41 Mizuta T, Segawa T. Chronic effects of imipramine and lithiumon 5-HT receptor subtypes in rat frontal cortex, hippocampus andchoroid plexus: quantitative receptor autoradiographic analysis.Jpn J Pharmacol 1989; 50: 315–326.
42 Subhash MN, Srinivas BN, Vinod KY, Jagadeesh S. Modulation of5-HT1A receptor mediated response by fluoxetine in rat brain. JNeural Transm 2000; 107: 377–387.
43 Newman ME, Gur E, Dremencov E, Garcia F, Lerer B, Van de KarLD. Chronic clomipramine alters presynaptic 5-HT(1B) and post-synaptic 5-HT(1A) receptor sensitivity in rat hypothalamus andhippocampus, respectively. Neuropharmacology 2000; 39: 2309–2317.
44 Yoshioka M, Matsumoto M, Numazawa R, Togashi H, Smith CB,Saito H. Changes in the regulation of 5-hydroxytryptamine releaseby alpha2-adrenoceptors in the rat hippocampus after long-termdesipramine treatment. Eur J Pharmacol 1995; 294: 565–570.
45 Nestler EJ, McMahon A, Sabban EL, Tallman JF, Duman RS.Chronic antidepressant administration decreases the expression oftyrosine hydroxylase in the rat locus coeruleus. Proc Natl Acad SciUSA 1990; 87: 7522–7526.
46 Herman JP, Cullinan WE. Neurocircuitry of stress: central controlof the hypothalamo-pituitary-adrenocortical axis. Trends Neurosci1997; 20: 78–84.
47 Harbuz MS, Lightman SL. Responses of hypothalamic and pituitarymRNA to physical and psychological stress in the rat. J Endocrinol1989; 122: 705–711.
48 Harbuz MS, Jessop DS, Lightman SL, Chowdrey HS. The effectsof restraint or hypertonic saline stress on corticotrophin-releasingfactor, arginine vasopressin, and proenkephalin A mRNAs in theCFY, Sprague–Dawley and Wistar strains of rat. Brain Res 1994;667: 6–12.
49 Kalin NH, Takahashi LK, Chen FL. Restraint stress increases corti-cotropin-releasing hormone mRNA content in the amygdala andparaventricular nucleus. Brain Res 1994; 656: 182–186.
50 Makino S, Smith MA, Gold PW. Increased expression of corticotro-pin-releasing hormone and vasopressin messenger ribonucleic acid(mRNA) in the hypothalamic paraventricular nucleus duringrepeated stress: association with reduction in glucocorticoid recep-tor mRNA levels. Endocrinology 1995; 136: 3299–3309.
51 Bartanusz V, Aubry JM, Jezova D, Baffi J, Kiss JZ. Up-regulation ofvasopressin mRNA in paraventricular hypophysiotrophic neuronsafter acute immobilization stress. Neuroendocrinology 1993; 58:625–629.
52 Herman JP. In situ hybridization analysis of vasopressin gene tran-
Molecular Psychiatry
scription in the paraventricular and supraoptic nuclei of the rat:regulation by stress and glucocorticoids. J Comp Neurol 1995; 363:15–27.
53 Lightman SL, Young WSI. Corticotrophin-releasing factor, vaso-pressin and proopiomelanocortin mRNA responses to stress andopiates in the rat. J Physiol (Lond) 1988; 403: 511–523.
54 Darlington DN, Barraclough CA, Gann DS. Hypotensive hemo-rrhage elevates corticotropin-releasing hormone messenger ribo-nucleic acid (mRNA) but not vasopressin mRNA in the rat hypo-thalamus. Endocrinology 1992; 130: 1281–1288.
55 Herman JP, Wiegand SJ, Watson SJ. Regulation of basal corticotro-pin-releasing hormone and arginine vasopressin messenger ribo-nucleic acid expression in the paraventricular nucleus: effects ofselective hypothalamic deafferentations. Endocrinology 1990; 127:2408–2417.
56 Schmidt ED, Janszen AW, Binnekade R, Tilders FJ. Transient sup-pression of resting corticosterone levels induces sustained increaseof AVP stores in hypothalamic CRH-neurons of rats. J Neuroendo-crinol 1997; 9: 69–77.
57 Ma XM, Lightman SL. The arginine vasopressin and corticotro-phin-releasing hormone gene transcription responses to varied fre-quencies of repeated stress in rats. J Physiol (Lond) 1998; 510:605–614.
58 Johnson MR, Lydiard RB, Ballenger JC. Panic disorder. Pathophysi-ology and drug treatment. Drugs 1995; 49: 328–344.
59 Stewart JW, McGrath PJ, Quitkin FM, Rabkin JG, Harrison W,Wager S et al. Chronic depression: response to placebo, imipra-mine, and phenelzine. J Clin Psychopharmacol 1993; 13: 391–396.
60 Korf J, Venema K. Desmethylimipramine enhances the release ofendogenous GABA and other neurotransmitter amino acids fromthe rat thalamus. J Neurochem 1983; 40: 946–950.
61 Sawchenko PE, Swanson LW. The organization of forebrain affer-ents to the paraventricular and supraoptic nuclei of the rat. J CompNeurol 1983; 218: 121–144.
62 Lai CT, Tanay VA, Charrois GJ, Baker GB, Bateson AN. Effects ofphenelzine and imipramine on the steady-state levels of mRNAsthat encode glutamic acid decarboxylase (GAD67 and GAD65), theGABA transporter GAT-1 and GABA transaminase in rat cortex.Naunyn Schmiedebergs Arch Pharmacol 1998; 357: 32–38.
63 Delbende C, Delarue C, Lefebvre H, Bunel DT, Szafarczyk A,Mocaer E et al. Glucocorticoids, transmitters and stress. Br J Psy-chiatry 1992; Suppl 15: 24–35.
64 Watanabe Y, Sakai RR, McEwen BS, Mendelson S. Stress and anti-depressant effects on hippocampal and cortical 5-HT1A and 5-HT2receptors and transport sites for serotonin. Brain Res 1993; 615:87–94.
65 Meijer OC, Van Oosten RV, De Kloet ER. Elevated basal troughlevels of corticosterone suppress hippocampal 5-hydroxy-tryptamine(1A) receptor expression in adrenally intact rats:implication for the pathogenesis of depression. Neuroscience 1997;80: 419–426.
66 Drevets WC, Frank E, Price JC, Kupfer DJ, Holt D, Greer PJ et al.PET imaging of serotonin 1A receptor binding in depression. BiolPsychiatry 1999; 46: 1375–1387.
67 Sargent PA, Kjaer KH, Bench CJ, Rabiner EA, Messa C, Meyer J et al.Brain serotonin1A receptor binding measured by position emissiontomography with [11C]WAY-100635: effects of depression and anti-depressant treatment. Arch Gen Psychiatry 2000; 57: 174–180.
68 Jacobs BL, Praag H, Gage FH. Adult brain neurogenesis and psy-chiatry: a novel theory of depression. Mol Psychiatry 2000; 5:262–269.
69 Lindsay RM, Wiegand SJ, Altar CA, DiStefano PS. Neurotrophicfactors: from molecule to man. Trends Neurosci 1994; 17: 182–190.
70 Herman JP, Schafer MK, Young EA, Thompson R, Douglass J, AkilH et al. Evidence for hippocampal regulation of neuroendocrineneurons of the hypothalamo-pituitary-adrenocortical axis. J Neuro-sci 1989; 9: 3072–3082.
71 Young EA, Haskett RF, Murphy-Weinberg V, Watson SJ, Akil H.Loss of glucocorticoid fast feedback in depression. Arch Gen Psy-chiatry 1991; 48: 693–699.
72 Nibuya M, Morinobu S, Duman RS. Regulation of BDNF and trkBmRNA in rat brain by chronic electroconvulsive seizure and antide-pressant drug treatments. J Neurosci 1995; 15: 7539–7547.
73 Tao X, Finkbeiner S, Arnold DB, Shaywitz AJ, Greenberg ME. Ca2+

mRNA changes in the stress axis induced by St John’s wortV Butterweck et al
564
Molecular Psychiatry
influx regulates BDNF transcription by a CREB family transcriptionfactor-dependent mechanism. Neuron 1998; 20: 709–726.
74 Nibuya M, Nestler EJ, Duman RS. Chronic antidepressant adminis-tration increases the expression of cAMP response element bindingprotein (CREB) in rat hippocampus. J Neurosci 1996; 16: 2365–2372.
75 Smith MA, Makino S, Kvetnansky R, Post RM. Stress and glucocort-icoids affect the expression of brain-derived neurotrophic factorand neurotrophin-3 mRNAs in the hippocampus. J Neurosci 1995;15: 1768–1777.
76 Kuroda Y, McEwen BS. Effect of chronic restraint stress and tianep-
This article is a United States Government Work paper as defined by the US Copyright Act.
tine on growth factors, growth-associated protein-43 and microtu-bule-associated protein 2 mRNA expression in the rat hippocam-pus. Brain Res Mol Brain Res 1998; 59: 35–39.
77 Brady LS. Stress, antidepressant drugs, and the locus coeruleus.Brain Res Bull 1994; 35: 545–556.
78 Melia KR, Nestler EJ, Duman RS. Chronic imipramine treatmentnormalizes levels of tyrosine hydroxylase in the locus coeruleus ofchronically stressed rats. Psychopharmacology 1992; 108: 23–26.
79 Calogero AE, Gallucci WT, Chrousos GP, Gold PW. Catecholamineeffects upon rat hypothalamic corticotropin-releasing hormonesecretion in vitro. J Clin Invest 1988; 82: 839–846.




















