
Complex Salt Bridges in Proteins: Statistical Analysis of Structure and Function
10
J. Mol. Biol. (1995) 254, 761–770 Complex Salt Bridges in Proteins: Statistical Analysis of Structure and Function Boaz Musafia, Virginia Buchner and Dorit Arad* We developed an algorithm to analyze the distribution and geometry of Department of Molecular simple and complex salt bridges in 94 proteins selected from the Protein Biology and Biotechnology Data Bank. In this study, the term ‘‘salt bridging’’ denotes both non-bonded The George S. Wise Faculty of Life Sciences, Tel-Aviv and hydrogen-bonded paired electrostatic interactions between acidic carboxyl groups and basic amino groups in single or adjacent protein University, Ramat Aviv 39040 Tel Aviv, Israel chains. We defined complex salt bridges as those joining more than two charged residues, including Asp, Glu, Lys and Arg, and excluding His. The survey related the following special features of complex salt bridges. (1) The abundance of complex salt bridges is high; one-third of all residues participating in salt-bridge formation were part of complex salt bridges. (2) The geometry of the interaction between acidic and basic residues is very similar in simple and complex salt bridges. Adding one residue to a simple interaction represents a minor change in the geometry but provides the molecule with a more complex interaction, a phenomenon that may explain the cooperative effect of salt bridges in proteins. Such moderate changes in salt-bridge networks can be generated stepwise and reversibly without trapping the protein in a local energetic minimum. (3) One important role of complex salt bridges is connecting protein subunits or joining two secondary structures to form quaternary structures, where they can connect as many as five secondary structure units. (4) Arginine serves as a key connector and/or a branching unit because its geometry allows three possible directions of interactions. The information gained from this study of complex salt bridges should enhance the understanding of protein structure. 7 1995 Academic Press Limited Keywords: ion-pairing; protein–protein interactions; protein structure; *Corresponding author intersubunit interactions; arginine Introduction Salt-bridge interactions are among the more thoroughly investigated functional groups in pro- tein research. Salt bridging plays an important role in the structure and function of proteins (Perutz, 1978, 1994; Hendsch & Tibor, 1994). One group of salt bridges, which we call ‘‘complex’’, has not been thoroughly studied as a separate entity. To distinguish between the terms ‘‘simple’’ and ‘‘complex’’ salt bridges as used in this study, we defined a simple salt bridge as a non-bonded or hydrogen-bonded ion-paired interaction that joins a single pair of charged amino acid residues, whereas a complex salt bridge joins more than two residues (such as the triad Asp-Arg-Asp) in single or adjacent protein chains. We believe that complex salt bridges deserve special treatment because our recent experience in modeling a breast cancer cell surface antigen, the muc1 gene product H23Ag, suggested that a multiple formation of complex salt bridges might be involved in intermolecular interactions (Arad et al ., 1992). Complex salt bridges could be important because of the cooperativity involved (Horovitz et al ., 1990; Horovitz, 1995; Marqusee & Sauer, 1994). Korn & Burnett (1991) found that some subunits in allosteric proteins bind pre- dominantly through ion pairs and hydrogen bonds. New information on complex salt bridges should contribute to the understanding of protein structure. The goal of this study was to analyze and compare the distribution, geometry, and function of complex versus simple salt bridges in proteins Abbreviations used: HMG-3, hemagglutinin-3; MDH, malate dehydrogenase; PDB, Protein Data Bank; MW, Mann-Whitney Wilcoxon test. 0022–2836/95/490761–10 $12.00/0 7 1995 Academic Press Limited
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Complex Salt Bridges in Proteins: Statistical Analysis of Structure and Function

J. Mol. Biol. (1995) 254, 761–770
Complex Salt Bridges in Proteins: Statistical Analysisof Structure and Function
Boaz Musafia, Virginia Buchner and Dorit Arad*
We developed an algorithm to analyze the distribution and geometry ofDepartment of Molecularsimple and complex salt bridges in 94 proteins selected from the ProteinBiology and BiotechnologyData Bank. In this study, the term ‘‘salt bridging’’ denotes both non-bondedThe George S. Wise Faculty
of Life Sciences, Tel-Aviv and hydrogen-bonded paired electrostatic interactions between acidiccarboxyl groups and basic amino groups in single or adjacent proteinUniversity, Ramat Aviv
39040 Tel Aviv, Israel chains. We defined complex salt bridges as those joining more than twocharged residues, including Asp, Glu, Lys and Arg, and excluding His. Thesurvey related the following special features of complex salt bridges. (1)The abundance of complex salt bridges is high; one-third of all residuesparticipating in salt-bridge formation were part of complex salt bridges.(2) The geometry of the interaction between acidic and basic residues isvery similar in simple and complex salt bridges. Adding one residue to asimple interaction represents a minor change in the geometry but providesthe molecule with a more complex interaction, a phenomenon that mayexplain the cooperative effect of salt bridges in proteins. Such moderatechanges in salt-bridge networks can be generated stepwise and reversiblywithout trapping the protein in a local energetic minimum. (3) Oneimportant role of complex salt bridges is connecting protein subunits orjoining two secondary structures to form quaternary structures, where theycan connect as many as five secondary structure units. (4) Arginine servesas a key connector and/or a branching unit because its geometry allowsthree possible directions of interactions. The information gained from thisstudy of complex salt bridges should enhance the understanding of proteinstructure.
7 1995 Academic Press Limited
Keywords: ion-pairing; protein–protein interactions; protein structure;*Corresponding author intersubunit interactions; arginine
Introduction
Salt-bridge interactions are among the morethoroughly investigated functional groups in pro-tein research. Salt bridging plays an important rolein the structure and function of proteins (Perutz,1978, 1994; Hendsch & Tibor, 1994). One group ofsalt bridges, which we call ‘‘complex’’, has not beenthoroughly studied as a separate entity. Todistinguish between the terms ‘‘simple’’ and‘‘complex’’ salt bridges as used in this study, wedefined a simple salt bridge as a non-bonded orhydrogen-bonded ion-paired interaction that joins asingle pair of charged amino acid residues, whereasa complex salt bridge joins more than two residues
(such as the triad Asp-Arg-Asp) in single oradjacent protein chains.
We believe that complex salt bridges deservespecial treatment because our recent experiencein modeling a breast cancer cell surface antigen,the muc1 gene product H23Ag, suggested thata multiple formation of complex salt bridgesmight be involved in intermolecular interactions(Arad et al., 1992). Complex salt bridges could beimportant because of the cooperativity involved(Horovitz et al., 1990; Horovitz, 1995; Marqusee &Sauer, 1994). Korn & Burnett (1991) found thatsome subunits in allosteric proteins bind pre-dominantly through ion pairs and hydrogen bonds.New information on complex salt bridges shouldcontribute to the understanding of protein structure.
The goal of this study was to analyze andcompare the distribution, geometry, and function ofcomplex versus simple salt bridges in proteins
Abbreviations used: HMG-3, hemagglutinin-3; MDH,malate dehydrogenase; PDB, Protein Data Bank; MW,Mann-Whitney Wilcoxon test.
0022–2836/95/490761–10 $12.00/0 7 1995 Academic Press Limited
Complex Salt Bridging in Proteins762
selected from a database of protein structures, usingan algorithm that we developed for this study. Theresults indicate that the main role of complex saltbridges is to connect two or more protein subunitsor two protein structures. Complex salt bridges thusenable reversible and stepwise change in proteininteractions.
Results and Discussion
General observations
A total of 1105 salt bridges were found in the 94proteins analyzed (see Table 1). The results of theanalysis are arranged as follows: (1) salt bridges atthe primary, secondary, and quaternary-structurelevels; (2) H-bonded salt bridges; (3) spatialgeometries and functions of different amino acidresidues; (4) salt bridges at the atomic level. In allcases, both simple and complex salt bridges weredetected.
Statistical analysis of the protein structuresrevealed that 60% of the proteins contained complexsalt bridges. In 22% of the total salt bridges, theacid–base interaction had a C–O–N angle <90°,which did not allow hydrogen bonding. The resultspresented in Table 2 show that about one-third of thecharged amino acids participating in salt-bridgeformation were located in complex salt bridges(three or more amino acids). Salt bridges formed bya triad of charged residues were the most abundant.The abundance of salt bridges decreased as thenumber of residues in the bridge increased; saltbridges of six or more residues occurred only once.The influenza virus hemagglutinin-3 (HMG-3),which was unique in this group, contained thelargest complex salt bridge, a complex network of 15charged amino acid residues interacting amongthemselves to connect three enzyme subunits.
Table 2. Salt bridges and residues participating in simpleand complex salt bridgesSalt-bridge type(number of residues) Total salt bridges Total residues
Simple (2) 845 (76%) 1690 (65%)Complex (3) 166 498Complex (4) 74 296Complex (5) 15 75Complex (6) 2 12Complex (7) 1 7Complex (8) 1 8Complex (15) 1 15Total 1105 2601
Primary structure level
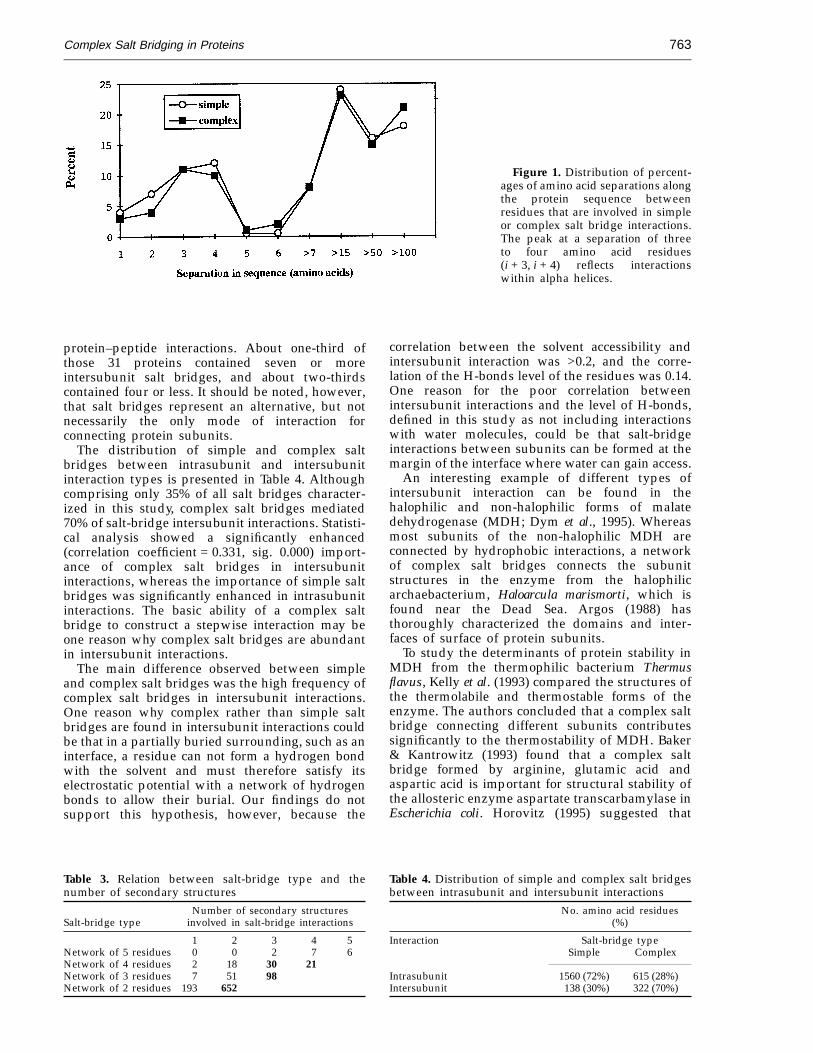
Figure 1 shows that in both simple and complexsalt bridges, most charged residues participating insalt-bridge formation were separated by chains ofseven or more amino acids. The results are similarto those of an earlier study on ion pairing inproteins by Barlow & Thornton (1983). We observeda second peak at separations of three to four aminoacids (i + 3, i + 4). Fifty percent of the i + 3, i + 4separations were within alpha helices, whichreflects the interaction of oppositely chargedresidues within an alpha helix (correlation coef-ficient = 0.4530, sig. = 0.000). Marqusee & Baldwin(1987) and Pingchiang et al. (1992) reported thatspecific salt-bridge interactions stabilize the alpha-helical structure in model short peptides.
Secondary structure level
We investigated the types of structural units thatare connected by complex salt bridges. Table 3shows that the number of secondary structuresconnected by complex salt bridges increases indirect relation to the increase in the number ofamino acid residues in the network of the saltbridge. In general, three-amino-acid salt bridgesconnect three secondary structure units, four-amino-acid salt bridges connect four secondarystructure units, and so on, which raises the questionof quantity versus quality. Although only one-thirdof the residues participating in salt-bridge for-mation are found in complex bridges, a complexsalt-bridge interaction has the unique ability tofocus on a single zone that connects as many as fivesecondary structure units. It can be concluded thatthis special character of converging multiplesecondary structure units may be an important toolthat the protein uses for forming tertiary andquaternary structures.
Quaternary structure level
Quaternary protein structure analysis revealedthat complex salt bridges are especially importantfor connecting different parts of intact proteinchains. Salt bridges mediated intersubunit inter-actions in 26 of 48 (54%) protein structures that wereinvolved either in intersubunit interactions or in
Table 1. List of proteins chosen for salt-bridge analysis(PDB entry)1 1acx 25 1ppt 49 2kai 73 3icb2 1bds 26 1prc 50 2liv 74 3pgk3 1bmv 27 1rbb 51 2ltn 75 3pgm4 1brd 28 1rhd 52 2mba 76 3ts15 1cc5 29 1rmu 53 2mev 77 451c6 1cd4 30 1rnt 54 2pab 78 4ait7 1cho 31 1sgt 55 2pka 79 4dfr8 1cms 32 1sn3 56 2stv 80 4i1b9 1coh 33 1tgs 57 2taa 81 4mdh
10 1crn 34 1thi 58 2tbv 82 4sbv11 1cse 35 1tnf 59 2tmv 83 4sgb12 1cy3 36 1wrp 60 2utg 84 4tln13 1ecd 37 1wsy 61 3adk 85 5acn14 1etu 38 1ypi 62 3b5c 86 5cpv15 1fc2 39 256b 63 3blm 87 5tnc16 1fx1 40 2act 64 3cpa 88 5xia17 1gcr 41 2aza 65 3csc 89 6pcy18 1gd1 42 2bp2 66 3dfr 90 7at119 1lh3 43 2ca2 67 3ebx 91 8adh20 1mbn 44 2ccy 68 3er3 92 8cat21 1p04 45 2cla 69 3gap 93 8pti22 1paz 46 2gbp 70 3grs 94 9api23 1pfk 47 2gn5 71 3hmg24 1phh 48 2hla 72 3hvp
Complex Salt Bridging in Proteins 763
Figure 1. Distribution of percent-ages of amino acid separations alongthe protein sequence betweenresidues that are involved in simpleor complex salt bridge interactions.The peak at a separation of threeto four amino acid residues(i + 3, i + 4) reflects interactionswithin alpha helices.
correlation between the solvent accessibility andintersubunit interaction was >0.2, and the corre-lation of the H-bonds level of the residues was 0.14.One reason for the poor correlation betweenintersubunit interactions and the level of H-bonds,defined in this study as not including interactionswith water molecules, could be that salt-bridgeinteractions between subunits can be formed at themargin of the interface where water can gain access.
An interesting example of different types ofintersubunit interaction can be found in thehalophilic and non-halophilic forms of malatedehydrogenase (MDH; Dym et al., 1995). Whereasmost subunits of the non-halophilic MDH areconnected by hydrophobic interactions, a networkof complex salt bridges connects the subunitstructures in the enzyme from the halophilicarchaebacterium, Haloarcula marismorti, which isfound near the Dead Sea. Argos (1988) hasthoroughly characterized the domains and inter-faces of surface of protein subunits.
To study the determinants of protein stability inMDH from the thermophilic bacterium Thermusflavus, Kelly et al. (1993) compared the structures ofthe thermolabile and thermostable forms of theenzyme. The authors concluded that a complex saltbridge connecting different subunits contributessignificantly to the thermostability of MDH. Baker& Kantrowitz (1993) found that a complex saltbridge formed by arginine, glutamic acid andaspartic acid is important for structural stability ofthe allosteric enzyme aspartate transcarbamylase inEscherichia coli. Horovitz (1995) suggested that
protein–peptide interactions. About one-third ofthose 31 proteins contained seven or moreintersubunit salt bridges, and about two-thirdscontained four or less. It should be noted, however,that salt bridges represent an alternative, but notnecessarily the only mode of interaction forconnecting protein subunits.
The distribution of simple and complex saltbridges between intrasubunit and intersubunitinteraction types is presented in Table 4. Althoughcomprising only 35% of all salt bridges character-ized in this study, complex salt bridges mediated70% of salt-bridge intersubunit interactions. Statisti-cal analysis showed a significantly enhanced(correlation coefficient = 0.331, sig. 0.000) import-ance of complex salt bridges in intersubunitinteractions, whereas the importance of simple saltbridges was significantly enhanced in intrasubunitinteractions. The basic ability of a complex saltbridge to construct a stepwise interaction may beone reason why complex salt bridges are abundantin intersubunit interactions.
The main difference observed between simpleand complex salt bridges was the high frequency ofcomplex salt bridges in intersubunit interactions.One reason why complex rather than simple saltbridges are found in intersubunit interactions couldbe that in a partially buried surrounding, such as aninterface, a residue can not form a hydrogen bondwith the solvent and must therefore satisfy itselectrostatic potential with a network of hydrogenbonds to allow their burial. Our findings do notsupport this hypothesis, however, because the
Table 3. Relation between salt-bridge type and thenumber of secondary structures
Number of secondary structuresSalt-bridge type involved in salt-bridge interactions
1 2 3 4 5Network of 5 residues 0 0 2 7 6Network of 4 residues 2 18 30 21Network of 3 residues 7 51 98Network of 2 residues 193 652
Table 4. Distribution of simple and complex salt bridgesbetween intrasubunit and intersubunit interactions
No. amino acid residues(%)
Interaction Salt-bridge typeSimple Complex
Intrasubunit 1560 (72%) 615 (28%)Intersubunit 138 (30%) 322 (70%)
Complex Salt Bridging in Proteins764
Table 5. Number of H-bonds associated with differentamino acid residues that participate in simple versuscomplex salt-bridge formation
Salt-bridge type Wilcoxon
Simple Complex (MW)
Residue Ave Cases Ave Cases 2-Tailed P
Arginine 1.80 444 2.24 289 0.0000Aspartic acid 1.42 496 1.82 226 0.0000Glutamic acid 1.15 469 1.48 261 0.0000Lysine 0.80 522 1.07 149 0.0008
Ave, average number of H-bond interactions with all residuesof this type (not including interaction with water).
bridges. (Solvent accessibility is defined accordingto the Connolly (1983) contact area of the chargedatoms of each residue.) When grouped according tosolvent exposure, H-bond formation was similar insimple and complex salt bridges. The results shownin Table 8 indicate that surface exposure is criticalto the level of H-bond formation in the protein,which is reasonable because of the participation ofwater in hydrogen bonding.
Significance of arginine
Janin et al. (1988) found that Arg residues, whichare involved in H-bonds between subunits, areamong the most abundant charged residues atsubunit interfaces. Arginine is important in allos-teric transitions (Perutz, 1989) and in the thermosta-bility of certain proteins (Barlow & Thornton, 1983;Qaw & Brewer, 1986).
Figure 2A shows that in complex salt bridges,arginine occupies 43.3% of all connecting pos-itions, whereas lysine occupies only 11.6% of thepositions. Figure 2D shows that a residue thatparticipates in complex salt-bridge formation canbe located either at an edge position (at the end ofthe complex network connected to only oneoppositely charged residue) or at a central position(serving to connect several residues). The distri-bution of positive residues at the edge position incomplex salt bridges (Figure 2B) also favorsarginine, but to a lesser extent. The picture wasdifferent for simple salt bridges, however, wherethe distribution of all residue types (Asp-Glu,Lys-Arg) was roughly equal, even leaningslightly toward the opposite direction and show-ing a small preference for lysine over arginine(Figure 2C).
In all cases where one single residue interactssimultaneously with three others, the interactionsinvolved either arginine or glutamic acid (ratherthan aspartic acid, which was absent). Arginine ispreferred because its geometrical structure allows
complex salt-bridge formation might be importantfor allosteric regulation. The results of a separatestudy on the role of complex salt bridges in allosterywill be presented elsewhere.
Hydrogen-bonded salt bridges
We measured the number of H-bonds that areassociated with the residues that participate in saltbridges. The results presented in Table 5 show thatthe number of residues involved in hydrogenbonding was significantly higher (Wilcoxon, 2-tailedP < 0.001) in complex salt bridges relative to simplesalt bridges. Table 6 shows that whether connectingthe same or different secondary structures, complexsalt bridges had a significantly higher (P < 0.04)number of H-bonds. The same tendency occurred inintrasubunit interactions (Table 7). In intersubunitinteractions, the number of H-bonds formed by Aspor Lys in simple and complex salt bridges wassimilar. On the other hand, Arg and Glu formedsignificantly higher (P < 0.04) numbers of H-bondsin complex salt bridges, which reflects theirimportance as connecting residues.
We asked whether solvent accessibility to theresidues influences the number of complex salt
Table 6. Number of H-bonds associated with different amino acid residues participating in simpleversus complex salt-bridge formation in the same or different secondary structures
Salt-bridge type Wilcoxon
Simple Complex (MW)
Type of interaction Residue Ave Cases Ave Cases 2-Tailed P
Same secondary structure Arginine 1.69 85 2.40 35 0.0070Aspartic acid 1.01 104 1.37 35 0.0365Glutamic acid 0.85 107 1.30 47 0.0082Lysine 0.64 126 0.98 43 0.0176
Different secondary structure Arginine 1.82 359 2.21 254 0.0001Aspartic acid 1.53 392 1.90 191 0.0001Glutamic acid 1.24 362 1.52 214 0.0011Lysine 0.85 369 1.11 106 0.0080
Ave, average number of H-bond interactions with all residues of this type (not including interaction withwater).
Complex Salt Bridging in Proteins 765
Table 7. Number of H-bonds associated with different amino acid residues participating in intra-or intersubunit simple versus complex salt-bridge formation
Salt-bridge type Wilcoxon
Simple Complex (MW)
Type of interaction Residue Ave Cases Ave Cases 2-Tailed P
Intrasubunit Arginine 1.78 402 2.13 196 0.0013Aspartic acid 1.39 451 1.78 161 0.0000Glutamic acid 1.15 436 1.32 170 0.0268Lysine 0.79 486 0.99 108 0.0330
Intersubunit Arginine 1.98 42 2.46 93 0.0478Aspartic acid 0.73 45 1.91 65 0.3905Glutamic acid 1.21 33 1.77 91 0.0027Lysine 1.00 36 1.29 41 0.1412
Ave, average number of H-bond interactions with all residues of this type (not including interaction withwater).
three possible directions of interactions. The highpreference for glutamic acid over aspartic acid maybe attributed to the longer chain length of glutamicacid, which allows more conformational freedomand reduces some steric hindrance from the bulkytriple-direction interaction.
The functional group of arginine is constructedfrom three asymmetrical nitrogen atoms (Ne, Nh1,Nh2) each of which can serve as a proton donor.Similar to the results of a study by Mitchell et al.(1992 ), we found that the carboxyl groups in simplesalt bridges showed a preference toward the Ne
atom. The carboxyl groups in complex salt bridges,however, were spread evenly around the guan-idinium group of arginine; with no preferencetoward any position of the N atom.
Spatial combinations in complex salt bridges
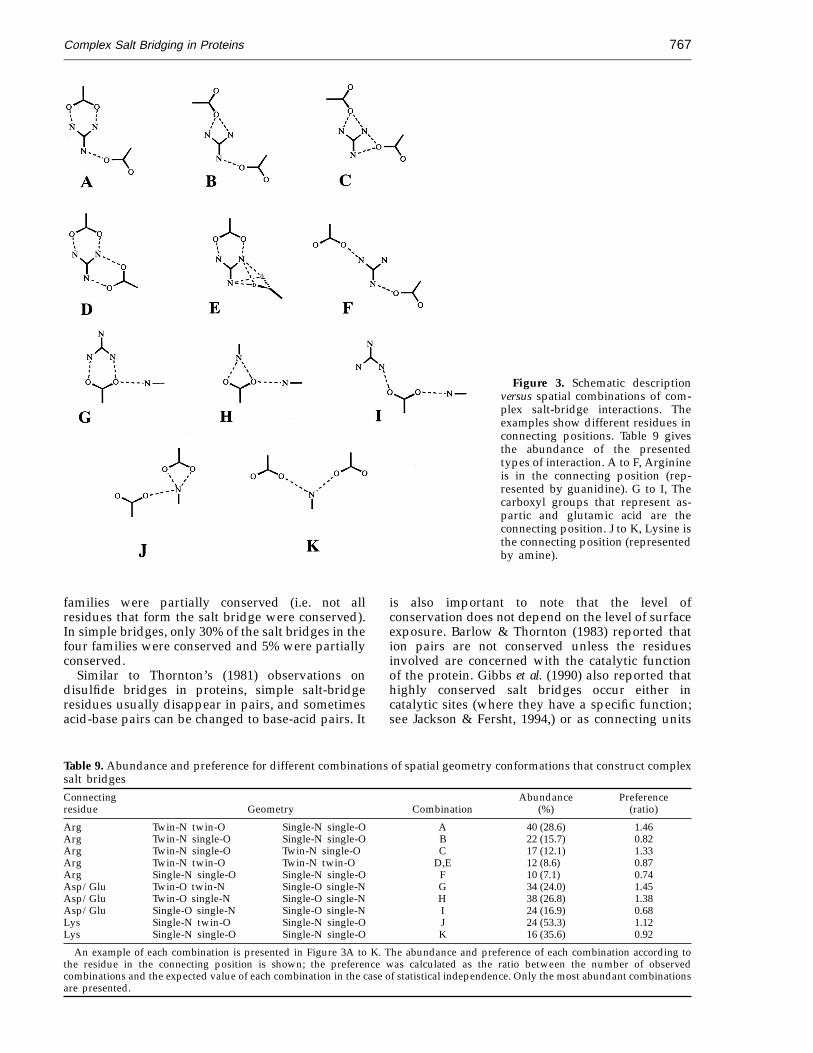
We found that the spatial geometry of simple andcomplex salt bridges was very similar. This sectionexplains the combinations and geometries aroundthe connecting residues in complex salt bridges.Thornton’s group (Singh et al., 1987; Mitchell et al.,1992) has thoroughly analyzed the geometries ofinteracting simple ion pairs, but to our knowledge,a description of the geometry around the connectingresidues in complex salt bridges has not beenreported. The statistical survey in the present studysearched for all possible combinations around eachcentral residue in connecting positions (Arg, Lys,Asp, or Glu). Figure 3 shows three structuralcategories, A, B, and C. The abundance of each
Table 8. Number of H-bonds associated with different amino acid residues participating in buriedor exposed simple versus complex salt-bridges
Salt-bridge type Wilcoxon
Simple Complex (MW)
Exposure Residue Ave Cases Ave Cases 2-Tailed P
0.00 A2 Arginine 2.85 26 2.97 32 0.3864Buried Aspartic acid 2.82 38 2.53 34 0.2350
Glutamic acid 2.35 26 3.21 19 0.0550Lysine 1.76 41 1.72 29 0.8173
0 < A2 < 5.0 Arginine 3.41 22 2.79 43 0.0424Partially Aspartic acid 1.81 42 2.57 47 0.0003buried Glutamic acid 1.69 32 1.60 40 0.7259
Lysine 1.48 40 1.10 20 0.2174
1.5 < A2 < 5.0 Arginine 2.48 64 2.63 62 0.5884Exposed Aspartic acid 2.01 84 1.82 56 0.4430
Glutamic acid 1.70 81 1.63 75 0.8553Lysine 0.93 86 1.20 45 0.0495
5.0 < A2 Arginine 1.49 315 1.76 152 0.0065Strongly Aspartic acid 1.04 313 1.15 89 0.1802exposed Glutamic acid 0.88 322 1.09 127 0.0317
Lysine 0.57 343 0.62 55 0.7235
Ave, average number of H-bond interactions with all residues of this type (not including interaction withwater).

Complex Salt Bridging in Proteins766
Figure 2. The distribution of amino acid types in saltbridges. A, Amino acid type distribution in complex saltbridges at connecting positions. B, Amino acid typedistribution in complex salt bridges at edge positions. C,Amino acid type distribution in simple salt bridge. D,Illustration of complex salt bridge and its possiblepositions. A residue in complex salt bridges can be at aconnecting position or at an edge position, as can be seenin D. At a connecting position (A) arginine plays a keyrole (43%), lysine on the other hand has a minor part asa connector in complex salt bridges (12%). At the edgeposition, acidic groups, especially glutamic acid (30%),have an enhanced importance (P = 0.000), and arginine isslightly preferred to lysine (25% and 18%, respectively).No preference for any amino acid was seen in simple saltbridges (C). We suspect that the preference for Arg as aconnector is due to the structure of guanidinum, whichenables interactions in three directions, and its long,flexible residue hydrocarbon chain.
Category B: central aspartic or glutamic acid(Figure 3, combinations G through I)
Combination G, where the two carboxyl oxygenatoms are bivalently bound to arginine, and oneoxygen atom is additionally bound to another base(either arginine or lysine) (twin-O twin-N; single-Osingle-N), was the most abundant (28%) in thiscategory. The statistical occurrence of combinationG was 1.75 higher than expected in the case ofindependence.
Category C: central lysine (Figure 3,combinations J and K)
The high abundance (48%) of combination J isexpected by random occurrence. Each carboxylicresidue of combination K contributes one valence tothe interaction.
Distance variables between atoms
Among other factors, the measure of the strengthof salt-bridge interaction is a function of (1) theinteratomic distances, and (2) the angles betweenthe interacting units. We measured the followingvariables that would help in characterizing thesalt-bridge interaction: the N–O distance, whichreflects the salt-bridge interaction.
Analysis of the interatomic distances presented inTable 10 clearly shows that in both simple andcomplex salt bridges, the distance between the N–Oatoms mediating the interaction is almost the same.On the other hand, except for Lys–Glu, the distancebetween the Ca atoms is significantly different(two-tailed P > 0.0162). From this observation, wecan conclude the following: (1) simple and complexsalt bridges share the same chemical bond becausethe N–O distances were maintained; and (2) thedistances between Ca atoms in the protein backboneconnected by complex salt bridges are larger thanthose connected by simple salt bridges, except in thecase of Lys–Glu interactions. Hence in complexsalt-bridge interactions, the flexible hydrocarbonpart of the residue must expand to reach andconnect the Ca atoms in the backbone.
Salt bridge conservation in proteins
We defined conserved residues as those that areconserved at least down to 50% identity of proteins.We performed multiple-sequence alignment of theprotein families† 1ypi, 1wsy, 2hla, and 3csc, whichcontained 39 simple and 20 complex salt bridgescomposed of 155 amino acid residues. Although thesalt bridges were generally not highly conserved,the residues in complex salt bridges were moreconserved than those in simple salt bridges.Forty-seven percent of the complex salt bridges inall four protein families were conserved. Twenty-one percent of the salt bridges in all four protein
combination (shown schematically in Figure 3)within each category is presented in Table 9.
Category A: central arginine (Figure 3,combinations A through F)
In this category the preferred combination (by afactor of 1.4) was twin-N twin-O; single-N single-O(combination A) where two nitrogen atoms ofarginine are twin-bound to the carboxyl group, andthe third nitrogen atom is monovalently bound toan additional carbonyl group. When one valence ofthe carboxyl group in a four-residue salt bridgemust remain open for additional interactions,however, the system clearly favors combination C(by a factor of 1.36).
† Key to protein families: 1ypi, triose phosphateisomerase (yeast); 1wsy, tryptophan synthase(Salmonella typhimurium); 2hla, human class Ihistocompatibility antigen; 3csc, citrate synthase(chicken).
Complex Salt Bridging in Proteins 767
Figure 3. Schematic descriptionversus spatial combinations of com-plex salt-bridge interactions. Theexamples show different residues inconnecting positions. Table 9 givesthe abundance of the presentedtypes of interaction. A to F, Arginineis in the connecting position (rep-resented by guanidine). G to I, Thecarboxyl groups that represent as-partic and glutamic acid are theconnecting position. J to K, Lysine isthe connecting position (representedby amine).
families were partially conserved (i.e. not allresidues that form the salt bridge were conserved).In simple bridges, only 30% of the salt bridges in thefour families were conserved and 5% were partiallyconserved.
Similar to Thornton’s (1981) observations ondisulfide bridges in proteins, simple salt-bridgeresidues usually disappear in pairs, and sometimesacid-base pairs can be changed to base-acid pairs. It
is also important to note that the level ofconservation does not depend on the level of surfaceexposure. Barlow & Thornton (1983) reported thation pairs are not conserved unless the residuesinvolved are concerned with the catalytic functionof the protein. Gibbs et al. (1990) also reported thathighly conserved salt bridges occur either incatalytic sites (where they have a specific function;see Jackson & Fersht, 1994,) or as connecting units
Table 9. Abundance and preference for different combinations of spatial geometry conformations that construct complexsalt bridgesConnecting Abundance Preferenceresidue Geometry Combination (%) (ratio)
Arg Twin-N twin-O Single-N single-O A 40 (28.6) 1.46Arg Twin-N single-O Single-N single-O B 22 (15.7) 0.82Arg Twin-N single-O Twin-N single-O C 17 (12.1) 1.33Arg Twin-N twin-O Twin-N twin-O D,E 12 (8.6) 0.87Arg Single-N single-O Single-N single-O F 10 (7.1) 0.74Asp/Glu Twin-O twin-N Single-O single-N G 34 (24.0) 1.45Asp/Glu Twin-O single-N Single-O single-N H 38 (26.8) 1.38Asp/Glu Single-O single-N Single-O single-N I 24 (16.9) 0.68Lys Single-N twin-O Single-N single-O J 24 (53.3) 1.12Lys Single-N single-O Single-N single-O K 16 (35.6) 0.92
An example of each combination is presented in Figure 3A to K. The abundance and preference of each combination according tothe residue in the connecting position is shown; the preference was calculated as the ratio between the number of observedcombinations and the expected value of each combination in the case of statistical independence. Only the most abundant combinationsare presented.

Complex Salt Bridging in Proteins768
Table 10. Interatomic average distance (A)N–O distance Ca distance
(charged atoms) (backbone atoms)Connectedresidues Simple Complex 2-Tailed P Simple Complex 2-Tailed P
Arg-Asp 2.93 3.05 0.0270 7.57 8.42 0.0010Arg-Glu 2.94 3.02 0.1014 7.93 8.46 0.0098Lys-Asp 3.08 2.85 0.2250 7.27 7.80 0.0162Lys-Glu 3.10 3.01 0.7644 7.80 8.08 0.9811
between different protein subunits, such as inchloramphenicol acetyl transferase.
Conclusions
Our broad statistical study, which was executedusing the GOMLEEN program, revealed thatcomplex salt bridges are not only common inproteins but also important for protein function.Multiple-sequence alignment showed that mostcomplex salt bridges in specific positions are notconserved throughout evolution but rather can bereplaced either by other neighboring salt bridges orby other interactions. The complex salt bridge isonly one of several options that can be used by aprotein for its proper folding and function. We choseto define the characteristics of complex salt bridgesas an example of a clustered structural unit.Analysis of the interacting residues at the level ofprimary, secondary, and quaternary structures ledto the following conclusions. (1) The abundance ofcomplex salt bridges is high. One-third of allresidues participating in salt-bridge formation werepart of complex salt bridges, implying that this typeof salt bridge is a common structural unit. (2) Thegeometry of interaction between acidic and basicresidues is very similar in simple and complex saltbridges. Adding one residue to a simple interactionrepresents a minor change in the geometry butprovides the molecule with a more complexinteraction, a phenomenon that may explain thecooperative effect of salt bridges in proteins. Suchmoderate changes in salt-bridge networks can begenerated stepwise and reversibly without trappingthe protein in a local energetic minimum. (3) Themain role of complex salt bridges is connectingprotein subunits or joining two secondary struc-tures to form quaternary structures, where they canconnect as many as five secondary structure units.(4) The number of salt-bridge residues involved inhydrogen bonding was significantly higher incomplex salt bridges relative to simple salt bridges,whether the complex salt bridge connects the sameor different secondary structures. When grouped
according to solvent exposure, however, H-bondformation was similar in simple and complex saltbridges.
At the residue level of analysis, we found that themost common residue in complex salt bridges,arginine, plays a key role not only in theirconstruction, serving both as a connecting unit anda branching element, but also in their function. Atthe atomic level of analysis, we found that the mostcommon organization of hydrogen bonds for aresidue at a connecting position in complex saltbridges is two hydrogen bonds on one side of theresidue and one hydrogen bond on the other.
Looking at complex salt bridges as an assembly ofpositive–negative interactions, we found that thespatial geometry of simple and complex salt bridgeswas very similar. Therefore, the metamorphosisfrom a simple to a complex salt bridge is probablynot dramatic† because half of the salt-bridgeinteraction is already constrained in a geometry thatsatisfies complex salt-bridge interactions.‡ Thus, avery small step for a single residue represents alarge step for the protein. Moreover, changes in thedegrees of freedom, which are limited in the simplesalt bridge, will not be excessive when the bridgebecomes complex.
Methods
Definitions
Before the salt bridges could be analyzed, it was firstnecessary to choose a criterion that would provide aworking definition for a salt bridge. For the purpose ofthis study, the term ‘‘salt bridge’’ denotes a non-bondedor hydrogen-bonded paired electrostatic interactionbetween acidic carboxyl groups and basic amino groupsin one or more protein chains. A simple salt bridge joinsa single pair of charged amino acid residues in single oradjacent protein chains, whereas a complex salt bridgejoins more than two residues (such as Asp-Arg-Asp). Inchoosing the cases for our study, we used the followingbroad definition of salt bridges: E4 A between N–O atompairs, independent of the angle of interaction (Barlow &Thornton, 1983). We analyzed both the total number ofresidues and the number of H-bonded residues thatparticipate in simple and complex salt-bridge formation.For the purpose of this study, H-bonds were definedaccording to the following criteria as defined for theprogram QUANTA8 (Molecular Simulations, Inc., 1983).(1) The distance between the donor and acceptor heavyatoms E 3 A; (2) the C–N–O and N–O–C angles e 90°;(3) each residue type (Asp, Glu, Arg, Lys) is consideredindividually; (4) water molecules from the crystal are notincluded.
† In CAT, for example, creating a complex salt bridgefrom a simple one barely changes the existinginteraction (Gibbs et al., 1990).
‡ Dao-Pin et al. (1991) proposed that creation of acomplex salt bridge shows a cooperative effect becausethe constraint of the first interaction limits the degreeof freedom of the system.

Complex Salt Bridging in Proteins 769
Data
We chose 94 distinct proteins, each with fullcrystallographic data, from the Brookhaven Protein DataBank (PDB; Bernstein et al., 1977); for protein names, seethe list in Table 1. To minimize redundancy, only proteinshaving less than 30% identity homology with any otherprotein in the set were included (Hbohum et al., 1992).When proteins having less than 65% homology wereincluded, the results and the conclusions were similar tothose obtained using 30% homology.
Programs
QUANTA
A library of secondary structures for each amino acidin the proteins was generated using the programQUANTA8 (Molecular Simulations, Inc., 1983). In thisprogram, the recognized secondary-structure types arethose based on hydrogen bonding patterns (defined byKabsch & Sander, 1983) and beta bulges (described byRichardson, 1981). Two additional secondary-structuretypes, extended and folded conformations, are deter-mined solely on the basis of the pseudo-torsion angles ofa-carbon atoms in four consecutive residues. In caseswhere not all subunit structures were listed in the PDB,we generated the full structure from symmetry con-ditions.
MS
The solvent exposure surface area of the functionalatoms in the residues was defined by the MS program(Connolly, 1983; available as QCPE No. 429). For thisstudy, we used the contact surface area that was generatedwith a 1.5 A probe.
GOMLEEN
Simple and complex salt bridges were identified andcharacterized by the program GOMLEEN, which wedeveloped for this study. GOMLEEN is a FORTRAN 77program that recognizes and analyzes simple andcomplex salt-bridge interactions in three-dimensionalprotein structures. The algorithm can be used foranalyzing other clustering interactions in proteins. Theinput for GOMLEEN is a file containing the list of all theselected proteins for analysis. GOMLEEN needs access tothe relevant Brookhaven PDB coordinate files and to thesecondary structures library of proteins that QUANTA8
generates. The six main output files contain both theextracted and calculated data about each salt bridge. Eachfile accumulates data about a different salt-bridge type(salt bridges having either two, three, four, five, six, ormore than six residues). The output files contain thefollowing variables on each residue. Each residue iscounted as a case, and the data for each case are in onerow. The information for each residue was obtained onthree levels: (1) general data on the protein; (2)information about the salt bridge as a whole; and (3)information about each individual residue, including itsinteraction with neighboring residues.
Protein level:
PDB protein name and a remark if necessary.Existence of protein subunits.
Resolution of the crystal.Number of arginine, lysine, aspartic and glutamic acidsin the protein.
Salt-bridge level:
Number of Arg, Lys, Asp, and Glu residues in the saltbridge.Total number of amino acid residues in the salt bridge.Topological arrangement of the salt bridge.Interaction between subunits via the specific residue.The number of the residue and its subunit name.
Residue level:
Name of the residue.Number of residues connected to the specific residue.
For each interaction between a specific residue and anyother residue, the following information is also providedin the same row:
Number of the interacting residue and its subunitname.Azimuthal and equatorial angles of the basic groupsrelative to the carboxyl groups.Azimuthal and equatorial angles of the carboxyl groupsof Glu and Asp relative to the guanidinium of Arg.The staggered angle in an Arg–carboxyl interaction.Geometry of the spatial conformation of interaction.Indication about intersubunit interaction in the relevantpair.Distances between the interacting atoms and theirnames.Distance between the Ca atoms.Distance between the carbon atoms that are adjacent tothe functional groups.Number of amino acid residues separating each saltbridge according to the primary structure.
The data from GOMLEEN were then entered into thefollowing three different database sets: (1) proteins (eachprotein is a case); (2) salt bridges (each salt bridge is acase); and (3) residues (each residue is a case).
SSPS
We used the SPSS8 program for Windows PC forstatistical analysis of the salt-bridge populations andamino acid compositions of the interfaces of allosteric andnon-allosteric proteins.
All programs except SSPS were run on a Crimson workstation of Silicon Graphics.
Statistics
The SPSS program for Windows (5.0.1) was used for thestatistical analysis of the GOMLEEN program’s output.Statistical significance was calculated by the Wilcoxontwo-tailed test and the Kendall correlation coefficient.
AcknowledgementsWe thank Dr Amnon Horovitz for stimulating
discussions and for reviewing the preprint.

Complex Salt Bridging in Proteins770
References
Arad, D., Musafia, B., Aharoni, O. & Mannor, S. (1992).Multiple-center Salt Bridges and Protein–Protein Inter-action. Israel Chemical Society, 57th Annual Meeting, TelAviv, Israel.
Argos, P. (1988). An investigation of protein subunit anddomain interfaces. Protein Eng. 2, 101–113.
Baker, D. F. & Kantrowitz, E. R. (1993). The conservedresidues Glu37, Asp100, Arg269, are important forstructural stabilization of E. coli aspartate transcar-bamylase. Biochemistry, 32, 10150–10158.
Barlow, D. J. & Thornton, J. M. (1983). Ion-pairs inproteins. J. Mol. Biol. 168, 867–885.
Bernstein, F. C., Koetzle, T., Williams, G., Meyer, E., Brice,M., Rogers, J., Kennard, O., Shimanouchi, T. &Tasumi, M. (1977). The Protein Data Bank: acomputer-based archival for macromolecular struc-tures. J. Mol. Biol. 112, 535–542.
Connolly, M. L. (1983). Analytical molecular surfacecalculation. J. Appl. Crystallog. 16, 548–558.
Dao-Pin, S., Suer, U., Nicholson, H. & Matthews,B. W. (1991). Contributions of engineered surfacesalt bridges to stability of T4 lysozyme deter-mined by direct mutagenesis. Biochemistry, 30,7142–7153.
Dym, O., Mevarech, M. & Sussman, J. L. (1995). Structuralfeatures that stabilize halophilic malate-dehydrogen-ase from an archaebacterium. Science, 267, 1344–1346.
Gibbs, M. R., Moody, P. C. E. & Leslie, G. W. (1990).Crystal structure of aspartic acid-199 > asparaginemutant of chloramphenicol acetyltransferase to2.35 A resolution: structural consequences of disrup-tion of a buried salt bridge. Biochemistry, 29,11261–11265.
Hbohum U., Scharf, M., Schneider, R. & Sander, C. (1992)Selection of representative protein data sets. ProteinSci. 1, 409–417.
Hendsch, Z. S. & Tibor, B. (1994). Do salt bridges stabilizeproteins—a continuum electrostatic analysis. ProteinSci. 3, 211–226.
Horovitz, A. (1995). The relation between cooperativity inligand binding and intramolecular cooperativity inallosteric proteins. Proc. Roy. Soc. ser. B, 259, 85–87.
Horovitz, A., Serrano, L., Avron, B., Bycroft, M. &Fersht, A. (1990). Strength and cooperativity ofcontributions of surface salt bridges to proteinstability. J. Mol. Biol. 216, 1031–1044.
Jackson, S. E. & Fersht, A. R. (1994). Contribution ofresidues in the reactive site loop of chymotrypsininhibitor 2 to protein stability and activity. Biochem-istry, 33, 13880–13887.
Janin, J., Miller, S. & Chothia, C. (1988). Surface subunitinterfaces and interior of oligomeric proteins. J. Mol.Biol. 204, 1551–1564.
Kabsch, W. & Sander, C. (1983). Dictionary of proteinsecondary structure: pattern recognition of hydro-gen-bonded and geometrical features. Biopolymers,22, 2577–2637.
Kelly, C. A., Nishiyama, M., Shanishi, Y., Beppu, T. &Birktoft, J. J. (1993). Determinants of protein stabilityobserved in the 1.9 A crystal structure of malatedehydrogenase from the thermophilic bacteriumThermus flavus. Biochemistry, 32, 3913–3922.
Korn, A. P. & Burnett, R. M. (1991). Distribution andcomplementarity of hydropathy in multisubunitproteins. Proteins: Struct. Funct. Genet. 9, 37–55.
Marqusee, S. & Baldwin, R. L. (1987). Helix stabilizationby Glu− . . . Lys+ salt bridges in short peptides of denovo design. Proc. Natl Acad. Sci. USA, 84, 8898–8902.
Marqusee, S. & Sauer, R. T. (1994). Contributions of ahydrogen bond/salt bridge network to the stabilityof secondary and tertiary structure in l repressor.Protein Sci. 3, 2217–2225.
Mitchell, J. B. O., Thornton, J. M. & Singh, J. (1992).Toward the understanding of the arginine-aspartateinteraction. J. Mol. Biol. 226, 251–262.
Perutz, M. F. (1978). Electrostatic effects in proteins.Science, 201, 1187–1191.
Perutz, M. F. (1989). Mechanisms of cooperativity andallosteric regulation in proteins. Quart. Rev. Biophys.22, 139–236.
Perutz, M. F. (1994). Polar zippers: their role in humandisease. Protein Sci. 3, 1629–1637.
Pingchiang, C. L., Gans, P. J. & Kallenbach, N. R. (1992).Energetic contributions of solvent-exposed ion pairsto alpha-helix structure. J. Mol. Biol. 223, 343–350.
Qaw, F. S. & Brewer, J. M. (1986). Arginyl residues andthermal stability in proteins. Mol. Cell. Biochem. 71,121–127.
Richardson, J. S. (1981). The anatomy and taxonomy ofprotein structure. Advan. Protein Chem. 34, 167–218.
Singh, J., Thornton, J. M. & Campbell, S. F. (1987). Thegeometries of interacting arginine-carboxyls inproteins. FEBS Letters, 224, 161–171.
Thornton, J.M. (1981). Disulphide bridges in globularproteins. J. Mol. Biol. 151, 261–187.
Edited by F. E. Cohen
(Received 3 August 1995; accepted 31 August 1995)




















