
Behavioural effects of receptor-specific substance P agonists
Upload
neurocrineCategory
view
0download
0
Journul of Ncurothemistry Raven Press, New York 0 1985 International Society for Neurochemistry
Complete Conversion of Brain D, Dopamine Receptors from the High- to the Low-Affinity State for Dopamine Agonists, Using Sodium Ions and Guanine Nucleotide
Dimitri Grigoriadis and Philip Seeman
Department of Pharmacology, Faculty of Medicine, University of Toronto, Toronto, Ontario, Cunudri
Abstract: Since previous work had shown that brain D, 3,4-dihydroxyphenylethylamine (dopamine) receptors were only partly converted from their high-affinity state to their low-affinity state, we here tested whether it was possible to obtain a complete 100% conversion of these receptors into their low-affinity state. It was first essential to resolve the components of ['Hlspiperone binding to dopaminergic sites and nondopaminergic sites in rat striatal homogenates. In the presence of 50 pM S-sul- piride (to occlude the dopaminergic sites), therefore, we first determined that the residual binding of [3Hl~p ipe r~ne (approximately 20%) was inhibited by serotonergic ago- nists much more effectively than dopamine or noradren- aline, thus identifying the serotonergic component of [3H]spiperone binding. Thus, dopamine (or ADTN) in- hibited the binding of [3H]spiperone at a high-affinity site (with dissociation constant of 10 nM dopamine), at a low- affinity site (with dissociation constant of 2,000 nM do- pamine), and at the serotonergic site (with dissociation constant of 50,000 nM dopamine). In the absence of so- dium ions, the high-affinity site was about 50% occupied
by ['Hlspiperone, and guanine nucleotide had no effect on this proportion. In the presence of 120 mM NaCI, however, the high-affinity site was reduced to 15% and guanine nucleotide completely eliminated this high-af- finity site, 100% of the sites having been completely con- verted to their low-affinity state. Using ['HIN-propyl-nor- apomorphine to label the high-affinity state of the do- pamine receptor, 50% conversion into the low-affinity state occurred at 45 mM LiCI, 69 mM NaCI, and 202 rnM KCI. We conclude that it is possible to convert brain D2 dopamine receptors completely into their low-affinity state, in the presence of NaCl and a guanine nucleotide, providing that appropriate allowance is made for the se- rotonergic component of ['Hlspiperone binding. Key Words: D, dopamine receptors-Binding affinity- Guanine nucleotide-Sodium i~ns-[~H]Spiperone- Serotonin-Rat striatum. Grigoriadis D. and Seeman P. Complete conversion of brain D2 doparnine receptors from the high- to the low-affinity state for dopamine ag- onists, using sodium ions and guanine nucleotide. J . Nerr- rochem. 44, 1925-1935 (1985).
This paper reports a complete 100% conversion of brain 3,4-dihydroxyphenylethylamine (dopa- mine) receptors from their high-affinity state for do- pamine to their low-affinity state for dopamine. Hitherto, it had been well known that guanine nu- cleotides lowered the affinity of dopamine receptors for dopamine agonists (Zahniser and Molinoff, 1978; Creese et al., 1979u,b; Goldstein et al., 1980; Di Paolo et a]., 1982; Battaglia and Titeler, 1982; Lazareno, 1983). A major difficulty with brain tissue, however, has been to convert all the dopa- mine receptors from their state of high affinity for dopamine to a state of low affinity for dopamine. A complete conversion of this sort has been readily
Received July 10, 1984; accepted December 13, 1984. Address correspondence and reprint requests to Dr. P. Seeman
at Department of Pharmacology, Faculty of Medicine, University of Toronto, Toronto, Ontario MSS lA8, Canada.
obtained for the dopamine receptors in anterior pi- tuitary tissue (Sibley et al., 1982; De Lean et al., 1980; George et al., 1983tr,h).
In brain tissue, however, such cwnplete conver- sion has not been possible. For example, Huff and Molinoff (1982) found that even extremely high con- centrations of guanine nucleotide (300 p M GTP) still left 23% of the ['Hlspiperone binding sites in their high-affinity state for dopamine (130 nM do- pamine). Wreggett et al. (1982) and Wreggett and Seeman (1984) also found that 21% of the high-af- finity sites remained in the presence of excess nu- cleotide. Although Leff and Creese (1982) men- tioned that complete conversion had occurred in
Ahhrc'viu/iorrs I I S C ~ : ADTN. ( 2 )-6.7-dihydroxy-2-arninotet- ralin; doparnine, 3.4-dihydroxyphenylethylamine; Gpp(NH)p, 5'-guanylylimidodiphosphate: ['HIN PA. I7H]N-n-propyl-nora- pomorphine.
1925

I926 D . GRIGORIADIS AND P . SEEMAN
canine brain tissue, their incubation conditions of 8-12 h at 4°C were extremely unusual. Our objec- tive was to obtain complete conversion using the standard incubation period of 60 min at 37°C. The present study describes the complete conversion of brain dopamine receptors from their high affinity for dopamine to their low-affinity state, by using both the customary NaCl and guanine nucleotide and by also making the appropriate computer-as- sisted allowance for the presence of 5-hydroxytryp- tamine (serotonin) receptors in the rat striatum.
MATERIALS AND METHODS
Materials [3H]Spiperone (22.6 Ciimmol) and [3HlN-n-propyl-nor-
apomorphine ([3H]NPA, 53.1 Ci/mmol) were obtained from New England Nuclear (Boston, MA, U.S.A.). Do- pamine, norepinephrine, tryptamine, serotonin, and 5'- guanylylimidodiphosphate [Gpp(NH)p] were obtained from Sigma Chemical, St. Louis, MO, U.S.A. The fol- lowing drugs were from the companies listed: (?)-6,7- dihydroxy-2-aminotetralin (ADTN), Research Biochem- icals, Wayland, MA, U.S.A.; (+)-butaclamol, Ayerst Re- search Laboratories, Montreal, Canada; ketanserin (R41468), Janssen Pharmaceuticals, Beerse, Belgium; S - sulpiride, Chernische Fabrik Schurholz, Koln, F.R.G.
Preparation of striatal homogenates Male Sprague-Dawley rats (Charles River, 200-225 g)
were killed by cervical dislocation. The brains were re- moved immediately and placed in ice-cold buffer (50 mM Tris-base, 5 mM KCI, 1.5 mM CaCI2, 4 mM MgCI2, 1 mM EDTA, pH 7.4, 0°C). The striata were dissected out, placed in Eppendorf microcentrifuge tubes, quick-frozen in liquid nitrogen, and stored at - 70°C for no longer than I week. On the day of assay, the striata were weighed and homogenized in the above buffer ( I 0 ml) by a Poly- tron (Brinkmann PT-lo), setting 7.0 for 20 s, and centri- fuged for 10 min at 480 g. The supernatant was saved and the pellet was resuspended in buffer (10 ml) and centri- fuged again for 10 min at 480 g. The final pellet was dis- carded, and the supernatants were combined and centri- fuged for 30 min at 48,000 g. The pellet was resuspended in buffer (same as above but pH 7.4, 37°C) to a final concentration of 4.5 mg original wet weighthi buffer, Polytron-homogenized at setting 7 for 5 s, preincubated for 10 rnin at 3 7 T , and cooled on ice for 30 min before addition to the incubation tubes.
[ 3H]Spiperone binding assays The buffer (pH 7.4, 37°C) used in the assays was the
same as described above with 12 pM nialamide and 0.1% ascorbic acid added. Binding assays were done in tripli- cate in disposable glass test tubes (12 X 75 mm).
For competition assays, using Eppendorf polypro- pylene pipette tips, test tubes received in order: 0.25 ml buffer (with or without test ions), 0.25 ml competing drug, 0.25 ml buffer [with or without Gpp(NH)p, 0.1 mM final concentration], 0.25 ml [3H]spiperone (80 pM final con- centration), 0.5 ml striatal homogenate (with a final con- centration of I .S mg original wet weight/ml) to give a total reaction volume of 1.5 ml. In all studies where the effects
of ions were to be studied, choline chloride was used as a control for ionic strength.
For Scatchard-type saturation assays, the tubes re- ceived in order: 0.25 ml buffer [with or without test ions or Gpp(NH)p], 0.25 ml buffer (with or without (+)-bu- taclamol), and 0.5 ml [3H]spiperone (10 pM-2.0 nM), and 0.5 ml striatal homogenate (with a final concentration of 1.5 mg original wet weightiml). All assays were carried out at equilibrium at 37°C. The tubes were prewarmed in the bath for 5 min before the membrane homogenate was added to start the reaction. All tubes were incubated for 60 min and the reaction was terminated by separation of the free radioligand from bound by rapid vacuum filtra- tion using a 12-well Cell Harvester (Skatron, Lier, Norway). The filters were washed for 20 s (approximately 10 ml fluid) with 50 mM Tris buffer (pH 7.4 at 22°C) and then placed in liquid scintillation mini-vials along with 4.5 ml of scintillation cocktail (Scint-A, United Technologies Packard, Downers Grove, IL, U.S.A.). After 12 h of shaking (100 rpm at 0-4"C), the samples were counted for bound radioactivity in a refrigerated Packard 460 C liquid scintillation counter at 45% efficiency.
Specific binding of [3H]~piper~ne was defined as that displaced by 1 .0 pM ( + )-butaclamol; as detailed in Re- sults, this includes binding to both dopaminergic and se- rotonergic components.
For the study of the serotonergic component of the striatum, 50 FM (-)-sulpiride was used to preclude ['Hlspiperone binding to D, sites. This concentration was chosen from List and Seeman (1981), where it was re- ported that 1-10 pM (-)-sulpiride was sufficient to in- hibit >95% of [3H]spiperone binding to D, sites (List and Seeman, 1981).
[3H]NPA binding assay The buffer used in this assay was identical to that of
the [3H]spiperone binding assay. In 12 x 75 mm dispos- able glass test tubes, the following ingredients were added in order: 0.5 ml buffer (with or without test ions, final concentration, 120 mM), 0.25 ml buffer (with or without (+)-butaclaniol, final concentration I pM), 0.25 ml ['HINPA (final concentration in test tube, 300 pM), and 0.5 ml membrane suspension (final concentration, I .5 mg original wet weightiml, prepared as previously de- scribed), to yield a total reaction volume of 1.5 ml. A time course experiment in the presence of all ions was performed on the same tissue pool on the same day and samples were incubated from 2 min to 4 h. The reaction was stopped again by rapid vacuum filtration and the samples treated and counted for radioactivity in precisely the same manner as those of the ['Hlspiperone binding assay.
The [3H]NPA ion inhibition data were done using NaCI, KCl, LiC1, and choline-CI, and again carried out in trip- licate on the same pool of tissue on a given day. The reaction took place at 37"C, in the presence of 300 pM [3H]NPA and incubated for 45 rnin. This time period was chosen from the time course experiment as the time when [3H]NPA, in the presence of all the ions, reached equilib- rium. The rest of this procedure was carried out in exactly the same way as the [3H]spiperone binding assay.
Data analysis A Scatchard-type saturation analysis was done for each
different condition that was to be examined by agonist/
1. Neurochem., Vol. 44, No. 6, 1985

CONVERSION OF D, RECEPTORS FROM HIGH TO LOW AFFINITY 1927
[)H]spiperone competition. The measured affinity of ['Hlspiperone for the D, dopamine receptor was subse- quently used for analysis of the ag~nist/[~H]spiperone competition. All data, Scatchard-type or competition- type, were analyzed using the iterative nonlinear least squares curve-fitting program "LIGAND" (Munson and Rodbard, 1980) run on a DEC-system 10 computer. The program provides objective data analysis of ligand binding giving estimates of the binding parameters. Curves can be fit to single- or multi-site models using the "extra sum of squares" principle to allow the user to decide statistically whether a fit to a more complex model i s justified and differs significantly from a fit to a simpler model. The analysis, as an additional index, uses an "F test" to determine whether adding parameters to a model significantly improves the fit to any given set of data.
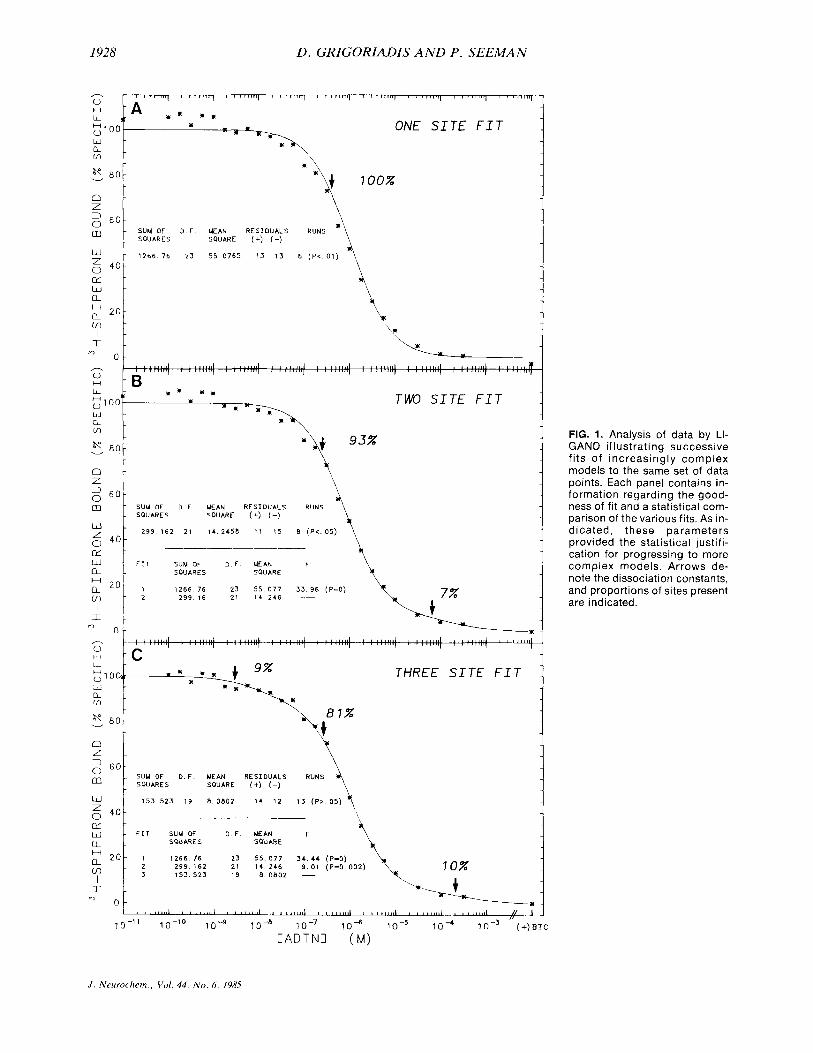
Figure 1 demonstrates the use of this analytical tech- nique. In Fig. lA, a set of data were fit to a singlesite model, that is, two parameters plus the nonspecific binding. The fit of the line (and hence the estimated pa- rameters), was significantly different from the data points at the p < 0.01 level. This fit was obviously rejected in favor of a more-than-one-site model.
Figure 1B shows the data fitted to a two-site model. Again the line and the data points were significantly dif- ferent from each other but this time only at the p < 0.05 level of significance. Looking at the comparison of these two fits, it is clear that with an F value of 33.77, the second fit was significantly better than the first (even though it does not significantly approximate the data points itself). We were, therefore, again justified in choosing a more-than-two-site model to compare these fits.
Figure IC shows the fit to three sites, one of the sites being given the affinity of spiperone at the serotonergic site. The fit of the parameters was highly significant and the line approximated the data at the p > 0.05 level of significance. The data, therefore, fitted a three-site model best (p > 0.05), since there was no significant difference between the fitted line (hence, the parameters estimated) and the experimental data. In addition, the F value in- dicated that the three-site fit was better than a two-site fit by a level of significance of p = 0.002.
RESULTS
Resolution of the dopaminergic and serotonergic components of [3H]spiperone binding
Approximately 80% of the total 13H]spiperone binding to rat striatum is to dopaminergic sites, and it has been presumed that the remaining 20% was serotonergic in nature (List and Seeman, 1981). Thus, we found it first necessary t o determine whether the 20% component had a true serotonin agonist rank order of sensitivity.
We first determined the absolute density (Bmax) of the serotonin S, component. Figure 2 illustrates Scatchard-type saturation analyses of [3H]spi- perone binding to rat striatal homogenates in the presence and absence of a potent serotonergic an- tagonist, ketanserin (R41468). The affinity for ['Hlspiperone was slightly altered in the presence of 50 nM ketanserin and ketanserin decreased the
receptor density (Bmax) by 12% (from 26 2 2 pM to 23 +- 2 pM; n = 3.' Neither of these changes was statistically significant. However, in the pres- ence of 50 p M sulpiride (to preclude all ['HI- spiperone binding to D, sites), the total binding of [3H]spiperone was markedly reduced, the Scat- chard plot was linear, and saturable binding ob- tained with a K , of 220 pM and the receptor density (Bmax) 3.8 ? 0.9 pM (n = 6). The decrease in the receptor density observed in the presence of ketan- serin (although not statistically significant) was equal to the receptor density in the presence of sul- piride; this was compatible with the premise, but did yet not prove, that the 3.8 pM difference was serotonergic.
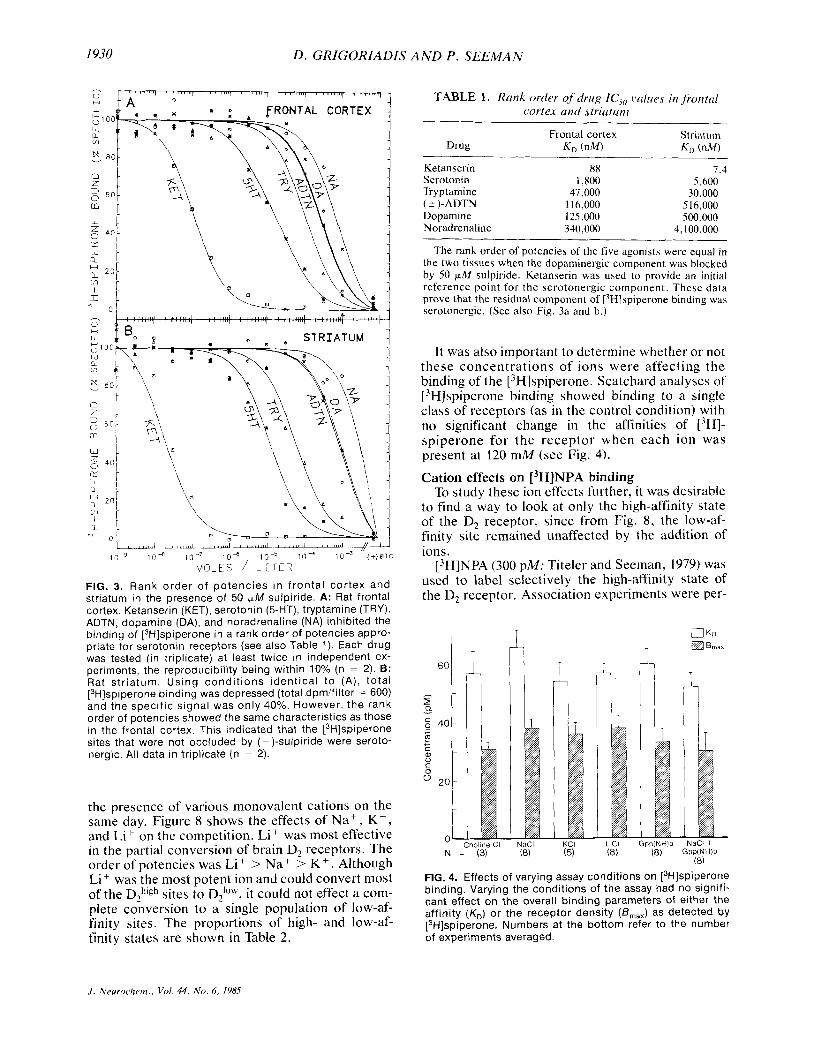
In experiments using 50 p M sulpiride to preclude [3H]~piperone binding to D, receptors, Fig. 3 and Table 1 illustrate that serotonin was the most potent agonist in both the frontal cortex and the striatum. This indicated that about 20% of [3H]spiperone binding was to serotonergic sites.
Once the serotonergic site had been identified, the effects of ions and guanine nucleotide could then be studied on the D, high- and low-affinity states. We first attempted to block the serotonergic component using 50 nM ketanserin but found we could only block <50% of the serotonergic sites. Ketanserin (250 nM) was also tested but this mark- edly reduced the total binding of ['Hlspiperone (data not shown). We decided, therefore, that the S, site would be included in all the calculations and analyses. Knowing the affinity of [3H]spiperone at that site (Fig. 2; in the presence of 50 p M sulpiride), we could analyze for two more parameters, those being the affinity and receptor concentration of that very low affinity serotonergic site.
Effects of ions and guanine nucleotide on [3H]spiperone binding to striatal homogenates
To analyze any [3H]spiperone/agonist competi- tion curve under different conditions, it was first essential to measure the K , and B,;,, of ['HI- spiperone under those conditions. For this reason, saturation experiments of ['Hlspiperone binding were done in the presence of ions and guanine nu- cleotide. The results are shown in Fig. 4. Addition of 120 mM NaCI, KCI, LiCI, or choline chloride did not change the affinity or the apparent receptor den-
' Receptor densities (B,,,, values) are expressed as pmol of bound ['HIspiperoneiL of final incubation medium. This can easily be converted to pmolig original wet weight or pmol mg protein as follows: We routinely use a wet weight tissue concen- tration of 1.5 mg ml in a final incubation volume of 1.5 ml. The conversion is applied, i.e.: 25 pmol/L x 1.5 m112.25 mg = 16.67 pmolig (original wet weight).
In our studies, approximately 7- 10% of the original wet weight (8 ) of the tissue is equal to the amount of protein (mg) in the sample: therefore, the concentration of receptors is 16.67 pmol g or 1.21-1.73 pmol mg protein.
.I. Nrirroclirm., Vol . 44, N o . 6 , 1985
1928
- ' G
Z l O O - w - R - m
K 8 0 -
n -
v
z - 2 6 0 -
m -
w - 5 4 0 - c - w - H
m -
I -
a - a 2 0 -
I
o 0 - - H G - 100 w - a - m
LT 8 0 - v
a - 2 . 2 6 0 -
D . GRIGORIADIS A N D P . SEEMAN
- , ,
3 i A - ~ K K l
I ' r ' ' " ' l 8 ' ,-I r ' ~ 1 ~ ' ~ f a ~~~~~~l I """I ' ' 1 ' 1 1 2 1
K - I ~ ONE S I T E F I T
SUM OF D F MEAN R E S I D U A L S RUNS . SOUARES SOUARE (+) (-)
1266 76 23 55 0765 13 13 6 ( P < 01)
\* d- it4
I m TWO S I T E F I T :: I' s' X i
SUM OF D F MEAN R E S I D U A L S RUNS SOUARE (+) (-) SOUARES
w - 6 4 0 - w - u F I T S U M OF D . F MEAN
SOUARES SOUARE
1 2 6 6 . 7 6 23 5 5 . 0 7 7 3 3 . 9 6 (P-0) m 299 1 6 21 1 4 . 2 4 6 -
I
299 162 21 14 2458 1 1 15 8 ( P C 05)
_____-_____
L L - ; 1 O03r l d - R - m
8 0 v
n L - ' "1 SUM OF D.F. MEAN R E S I D U A L S RUNS \
SOU ARES SOUARE (+) (-) m
:: Ut
X K THREE S I T E F I T
8 7 % r - = ? t 153 523 19 8 . 0 8 0 2 14 12 13 ( P Z 05) 4
\ - - - _ _ _ _ _ _ ~
7 0%
I l i - w - FIT SUM OF D F M A N a - SOU ARE S SQUARE
H a 2 0 - 1 1266 76 23 55 077 34 44 (P-0) 299 162 21 14 246 9 01 (P-0 002) 153 523 19 8 0802 - I
I . " 0 -
FIG. 1. Analysis of data by LI- GANO illustrating successive fits of increasingly complex models to the same set of data points. Each panel contains in- formation regarding the good- ness of fit and a statistical com- parison of the various fits. As in- dicated, these parameters provided the statistical justifi- cation for progressing to more complex models. Arrows de- note the dissociation constants, and proportions of sites present are indicated.
~, C A D T N I (M)
J . Neurochem., Vol. 44, No. 6, 19x5

CONVERSION OF D, RECEPTORS FROM HIGH TO LOW AFFINITY 1929
0. 2 5 i FIG. 2. Scatchard analysis of [3H]spiperone binding for determining the density of binding sites. [3H]Spiperone binding done in the presence of 120 mM NaCI. The control [3H]spiperone binding was saturable with a K, of 90 pM (in this experiment) and a re- ceptor density (BmaX) of 25.0 pM (pmoliL of final incubation suspension). In the presence of 50 nM ketanserin, the K, decreased to 80 pM and the B,,, decreased slightly to 21.6 pM. In the presence of 50 p,M sulpiride (to occlude the D, component), [3H]spiperone still exhibited saturable binding to a different site (serotonergic) with a K, of 210 pM and a B,,, of 3.4 pM. Each point is the mean of triplicate determinations. This is a represen- tative Scatchard from five experiments with the means quoted in the text.
0 0 4 0 8 0 12 0 16 0 20 0 24 0 20 0
sity significantly from that of the control situation (no ions or nucleotide). Similarly, the addition of guanine nucleotide [ 100 FM Gpp(NH)pl with or without 120 mM NaCl also had little or no effect on the binding parameters of [3H]spiperone for the D2 receptor (see Fig. 4).
All Scatchard plots were linear, indicating that [3H]spiperone had equal affinity for the DZhigh and DZIow states of the receptor. In all subsequent anal- yses of competition data, the dissociation constant (K,) of [3H]spiperone used was 55 k 12 pM. An average of eight independent Scatchard experi- ments done on different days showed that the KD values were between 37 and 90 pM, the mean being 55 F 12 SEM (n = 8 ) , and the receptor density was consistently 32 +_ 8 pM (n = 8). Effects of guanine nucieotide and sodium ions on ag~nist/[~H]spiperone competition
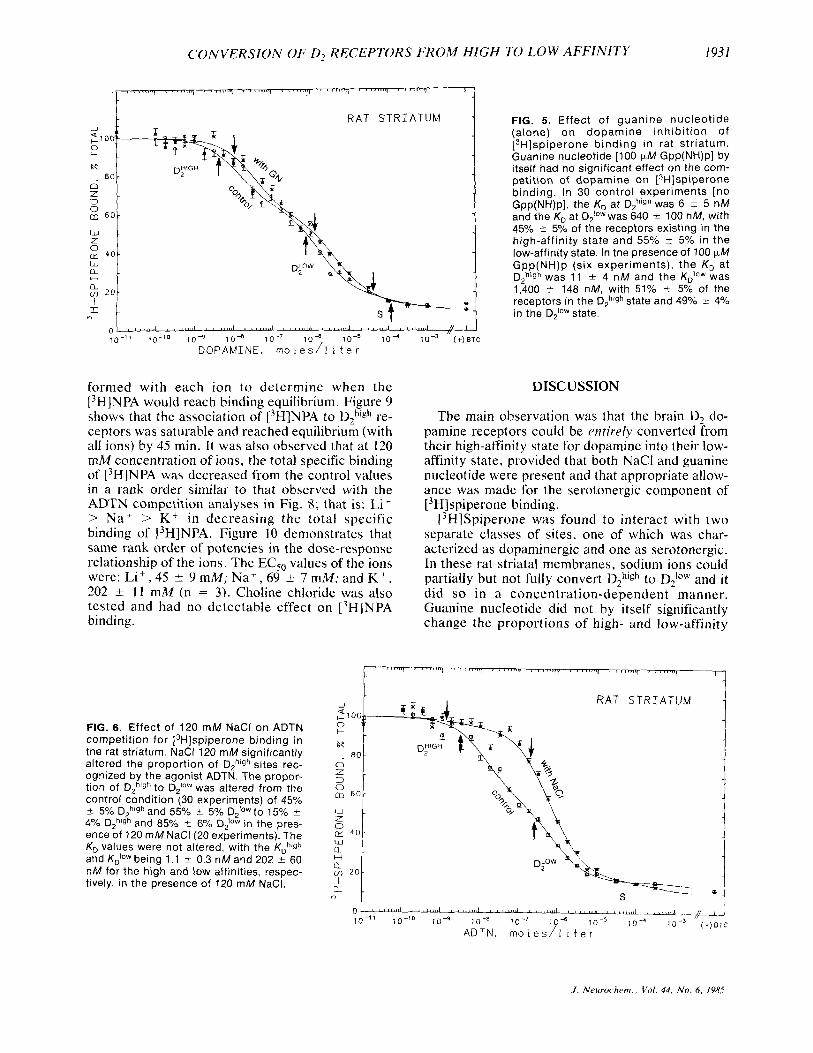
In the control buffer medium (no NaCl), guanine nucieotide had no significant effect on agonist com- petition of [3H]~piper~ne binding. Figure 5 shows that 100 F M Gpp(NH)p did not alter either the pro- portions of high- and low-affinity states or the af- finities of the agonist €or these states. These data agree with earlier findings (Wreggett et al., 1982; Wreggett and Seeman, 1983) where the incubation buffer routinely omitted sodium. As will be shown later, the nucleotide regulation of high- and low- affinity states was sensitive to the presence of monovalent cations.
NaCl (120 mM> had a dramatic effect on the rela- tive proportions of the dopamine D, receptor states. Figure 6 shows that in the absence of NaCl the ag- onist differentiated two distinct populations of D2 receptor states in approximately 5050 proportions. When 120 mM sodium was added to the medium, we observed a partial conversion of the high-affinity
state to the low-affinity state; the proportion of the high-affinity state decreased from 50% to approxi- mately 15%. Sodium ions did not change the dis- sociation constant of either the high- or low-affinity state for the agonist.
ADTN was allowed to compete against [3H]- spiperone in the presence of 120 mM and 240 mM NaCl. The 240 mM NaCl converted even more of the D,high receptors to D21"W (10% 2 4% high and 90% k 9% low sites remaining; n = 3) . We found, therefore, that even at very high concentrations of NaCl we could not completely convert all of the DZhigh receptors to their D 2 * O W state (data not shown),
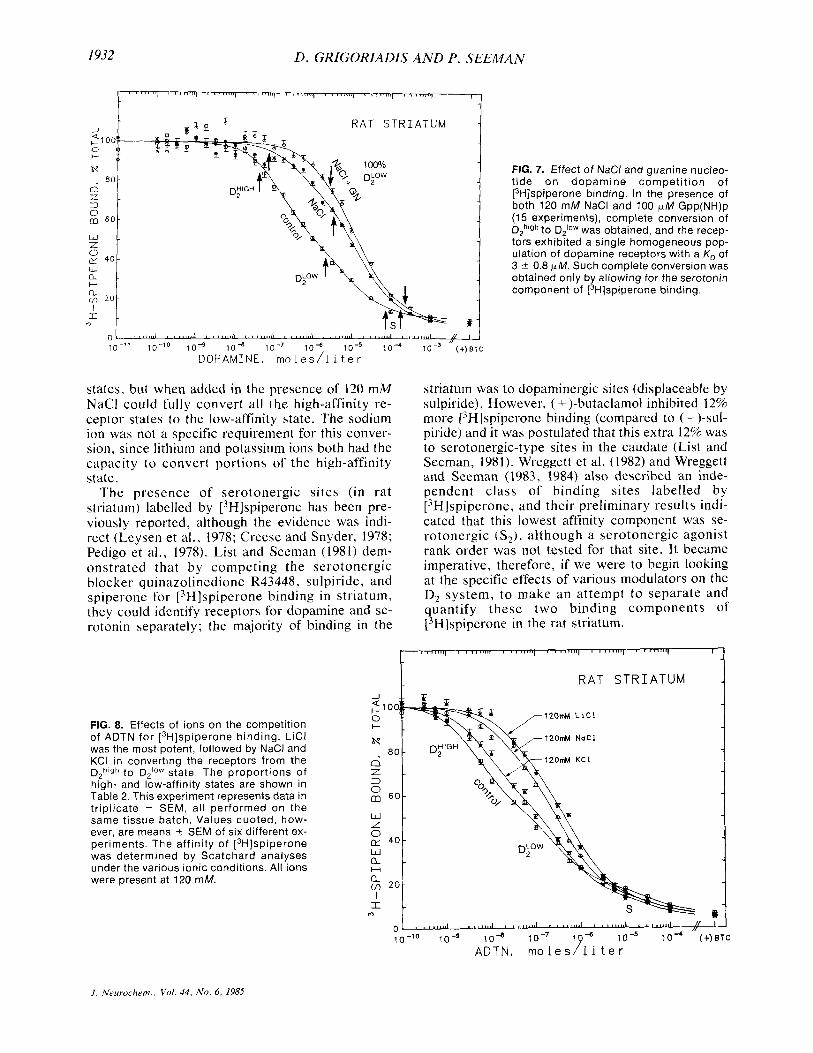
By adding guanine nucleotide in the presence of NaCl, we could demonstrate complete conversion of all the D,high receptor sites to their DZioW state. Figure 7 shows dopamine competition for [3H]- spiperone binding under three conditions (per- formed simultaneously). In the control conditions (no ions or nucleotide), dopamine inhibited ['Hlspiperone binding in three phases, the dopa- minergic component present in 50: 50 proportions of DZhigh and D2IoW, and the serotonergic component. In the presence of 120 mM NaCl, there was partial conversion and the proportions were shifted to 15% ? 4% in the DZhigh state and 85% -C 6% in the DPW state. Concomitant addition of 120 mM NaCl and 100 F M Gpp(NH)p was sufficient to convert com- pletely all the high-affinity state receptors to their low-affinity state, yielding a single homogeneous population of dopamine receptors in the low-affinity state (Fig. 7).
Cation selectivity for receptor regulation We next examined whether the sodium-induced
conversion was specific for this ion. (_+)-ADTN competition for [3H]spiperone binding was done in
J . Neurochem., Vol. 44, No. 6, 198.5
I930 D. GRIGORIADIS AND P . SEEMAN
l o - ' 10" lo-' l o d lo- ' lo - ' ( + ) a r c M O L E S / L I T E R
FIG. 3. Rank order of potencies i n f ronta l cortex and striatum in the presence of 50 pM sulpiride. A: Rat frontal cortex. Ketanserin (KET), serotonin (5-HT), tryptamine (TRY), ADTN, dopamine (DA), and noradrenaline (NA) inhibited the binding of [3H]spiperone in a rank order of potencies appro- priate for serotonin receptors (see also Table 1). Each drug was tested (in triplicate) at least twice in independent ex- periments, the reproducibility being within 10% (n = 2). B: Rat str iatum. Using condi t ions identical t o (A ) , total [3H]spiperone binding was depressed (total dpmlfilter = 600) and the specific signal was only 40%. However, the rank order of potencies showed the same characteristics as those in the frontal cortex. This indicated that the [3H]spiperone sites that were not occluded by (-)-sulpiride were seroto- nergic. All data in triplicate (n = 2).
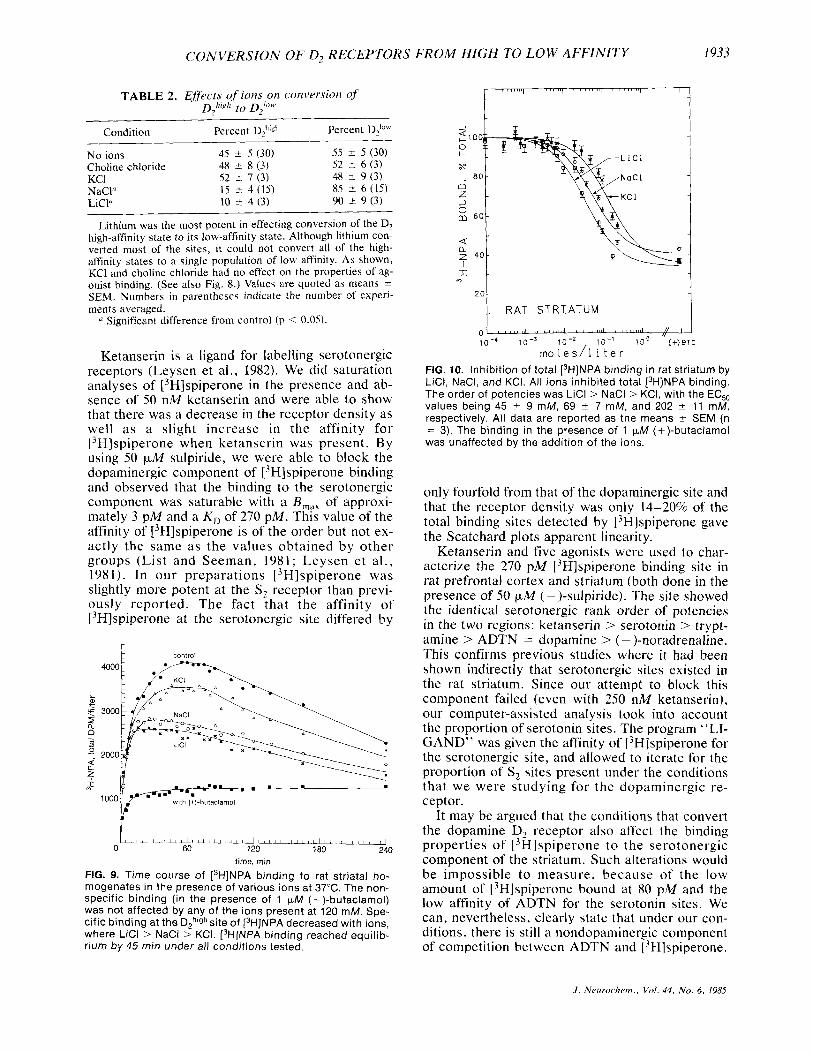
the presence of various monovalent cations on the same day. Figure 8 shows the effects of Na+ , Kf, and Li+ on the competition. Li' was most effective in the partial conversion of brain D2 receptors. The order of potencies was Li+ > Na' > K + . Although Li+ was the most potent ion and could convert most of the D2h'gh sites to D2low, it could not effect a com- plete conversion to a single population of low-af- finity sites. The proportions of high- and low-af- finity states are shown in Table 2.
TABLE 1. Rank order o j drug IC,, values in frontal cortex and striaturn
Frontal cortex Striaturn Drug K , (nM) KD (nM)
Ketanserin 88 7.4 Serotonin 1,800 5,600 Tryptamine 47,000 30,000 ( 2 )-ADTN 116,000 5 16,000
Noradrenaline 340,000 4,100,000 Dopamine 125,000 500,000
The rank order of potencies of the five agonists were equal in the two tissues when the dopaminergic component was blocked by 50 p M sulpiride. Ketanserin was used to provide an initial reference point for the serotonergic component. These data prove that the residual component of [3H]spiperone binding was serotonergic. (See also Fig. 3a and b.)
It was also important to determine whether or not these concentrations of ions were affecting t h e binding of the L3H]spiperone. Scatchard analyses of ['HJspiperone binding showed binding to a single class of receptors (as in the control condition) with no significant change in the affinities of ['HI- spiperone for the receptor when each ion was present at 120 mM (see Fig. 4).
Cation effects on [3H]NPA binding To study these ion effects further, it was desirable
to find a way to look at only the high-affinity state of the D, receptor, since from Fig. 8, the low-af- finity site remained unaffected by the addition of ions.
[3H]NPA (300 pM; Titeler and Seeman, 1979) was used to label selectively the high-affinity state of the D, receptor. Association experiments were per-
I
5 40 r
L c C a,
C
20
CholineCI NaCl KCI LiCI Gpp(NHlp NaCl I N = (3) (8) (5) (8) (8) GPP(NH)P
(8)
FIG. 4. Effects of varying assay conditions on [3H]spiperone binding. Varying the conditions of the assay had no signifi- cant effect on the overall binding parameters of either the affinity (K,) or the receptor density (5max) as detected by [3H]spiperone. Numbers at the bottom refer to the number of experiments averaged.
J . Nerirochem., Vol . 44. No. 6 , 1985
CONVERSION OF D2 RECEPTORS FROM HIGH TO LOW AFFINITY 1931
I
R A T S T R I A T U M
t a -
FIG. 5. Effect of guanine nucleotide (alone) on dopamine inhibit ion of [3H]spiperone binding in rat striatum. Guanine nucleotide [lo0 pM Gpp(NH)p] by itself had no significant effect on the com- petition of dopamine on [3H]spiperone binding. In 30 control experiments [no Gpp(NH)p], the K, at Dghlgh was 6 4 5 nM and the KD at Diowwas 640 2 100 nM, with 45Y0 ? 5% of the receptors existing in the high-affinity state and 55% ? 5% in the low-affinity state. In the presence of 100 (*M Gpp(NH)p (six experiments), the KD at DZhlgh was 11 f 4 nM and the K i o w was 1,400 r 148 nM, with 51% ? 5% of the receptors in the D,h'gh state and 49% ? 4% in the D;Ow state.
formed with each ion to determine when the ['HINPA would reach binding equilibrium. Figure 9 shows that the association of 13H]NPA to D,high re- ceptors was saturable and reached equilibrium (with all ions) by 45 min. It was also observed that at 120 mM concentration of ions, the total specific binding of ['HIINPA was decreased from the control values in a rank order similar to that observed with the ADTN competition analyses in Fig. 8; that is: Li t > N a + > K + in decreasing the total specific binding of [3H]NPA. Figure 10 demonstrates that same rank order of potencies in the dose-response relationship of the ions. The EC,, values of the ions were: Li', 45 t_ 9 mM; Na', 69 5 7 mM; and K + , 202 k I 1 mM (n = 3 ) . Choline chloride was also tested and had no detectable effect on ['HINPA binding.
DISCUSSION
The main observation was that the brain D, do- pamine receptors could be entirely converted from their high-affinity state for dopamine into their low- affinity state, provided that both NaCl and guanine nucleotide were present and that appropriate allow- ance was made for the serotonergic component of [3H]spiperone binding.
[3H]Spiperone was found to interact with two separate classes of sites, one of which was char- acterized as dopaminergic and one as serotonergic. In these rat striatal membranes, sodium ions could partially but not fully convert D,high to D2IoW and it did so in a concentration-dependent manner. Guanine nucleotide did not by itself significantly change the proportions of high- and low-affinity
I
R A T S T R I A T U M
FIG. 6. Effect of 120 mM NaCl on ADTN competition for [3H]spiperone binding in the rat striatum. NaCl 120 mM significantly altered the proportion of Dph'gh sites rec- ognized by the agonist ADTN. The propor- tion of DZh1gh to D2IaW was altered from the control condition (30 experiments) of 45% 2 5% DPhlgh and 55% t 5% D,'OW to 15% 2 4% DZhlgh and 85% 2 6% Diow in the pres- ence of 120 mM NaCl(20 experiments). The K, values were not altered, with the KDh'Qh and KAow being 1.1 t 0.3 nM and 202 2 60 nM for the high and low affinities, respec- tively, in the presence of 120 mM NaCI. .
S
J . Neurochem.. Vol . 14, No. 6, 1985
1932 D . GRIGORIADIS AND P. SEEMAN
1 '"'n"( ' """" ' ""'? ' """" ' " 1 " " 1 ' """'1 ' """1 ' "Irn"
I
O A T '=TRIATUM 2 1 0 0 0 I- + H I '"TI
2 1 6 0
K 4 0
l \ M l J
nLOW H I -2
# -
FIG. 7. Effect of NaCl and guanine nucleo- tide on dopamine competition of [3H]spiperone binding. In the presence of both 120 mM NaCl and 100 pM Gpp(NH)p (15 experiments), complete conversion of DZh'gh to D210w was obtained, and the recep- tors exhibited a single homogeneous pop- ulation of dopamine receptors with a K, of 3 1 0.8 pM. Such complete conversion was obtained only by allowing for the serotonin component of [3H]spiperone binding.
states, but when added in the presence of 120 mM NaCl could fully convert all the high-affinity re- ceptor states to the low-affinity state. The sodium ion was not a specific requirement for this conver- sion, since lithium and potassium ions both had the capacity to convert portions of the high-affinity state.
The presence of serotonergic sites (in rat striatum) labelled by ['Hlspiperone has been pre- viously reported, although the evidence was indi- rect (Leysen et al., 1978; Creese and Snyder, 1978; Pedigo et al., 1978). List and Seeman (1981) dem- onstrated that by competing the serotonergic blocker quinazolinedione R43448, sulpiride, and spiperone for [3H]spiperone binding in striaturn, they could identify receptors for dopamine and se- rotonin separately; the majority of binding in the
striatum was to dopaminergic sites (displaceable by sulpiride). However, ( + )-butaclamol inhibited 12% more ['Hlspiperone binding (compared to ( - )-sul- piride) and it was postulated that this extra 12% was to serotonergic-type sites in the caudate (List and Seeman, 1981). Wreggett et al. (1982) and Wreggett and Seeman (1983, 1984) also described an inde- pendent class of binding sites labelled by ['Hlspiperone, and their preliminary results indi- cated that this lowest affinity component was se- rotonergic (S2), although a serotonergic agonist rank order was not tested for that site. It became imperative, therefore, if we were to begin looking at the specific effects of various modulators on the D2 system, to make an attempt to separate and quantify these two binding components of ['Hlspiperone in the rat striaturn.
FIG. 8. Effects of ions on the competition of ADTN for [3H]spiperone binding. LiCl was the most potent, followed by NaCl and KCI in converting the receptors from the Dph'gh to D,Iow state. The proportions of high- and low-affinity states are shown in Table 2. This experiment represents data in triplicate 2 SEM, al l performed on the same tissue batch. Values quoted, how- ever, are means % SEM of six different ex- periments. The affinity of [3H]spiperone was determined by Scatchard analyses under the various ionic conditions. All ions were present at 120 mM.
J . Neurochem.. Voi. 44, No. 6 , 1985
CONVERSION OF D, RECEPTORS FROM HIGH TO LOW AFFINITY
01 ' ' * l l " J ' " m i " d ' L c m ' ' u L ' ' 1 1 1 1 " 1 " '
1933
TABLE 2. EjfecIs of ions on conversion of DZhi,?/l to D /OM.
2
Condition Percent DZh'gh Percent D P w N o ions 45 * 5 (30) 55 i 5 (30) Choline chloride 48 ? 8 (3) 52 I 6 (3) KCI 5 2 -+ 7 (3) 48 t 9 (3) NaCI" 15 -+ 4 (15) 85 +- 6 (15) LiCI" 10 -+ 4 (3) 90 2 9 (3)
Lithium was the most potent in effecting conversion of the D2 high-affinity state to its low-affinity state. Although lithium con- verted most of the sites, it could not convert all of the high- affinity states to a single population of low affinity. As shown, KCl and choline chloride had no effect on the properties of ag- onist binding. (See also Fig. 8.) Values are quoted as means 2 SEM. Numbers in parentheses indicate the number of experi- ments averaged.
" Significant difference from control ( p < 0.05).
Ketanserin is a ligand for labelling serotonergic receptors (Leysen et al., 1982). We did saturation analyses of [3H]spiperone in the presence and ab- sence of S O nM ketanserin and were able to show that there was a decrease in the receptor density as well as a slight increase in the affinity for [3H]spiperone when ketanserin was present. By using 50 p M sulpiride, we were able to block the dopaminergic component of ['Hlspiperone binding and observed that the binding to the serotonergic component was saturable with a B,,, of approxi- mately 3 pM and a K , of 270 pM. This value of the affinity of [3H]spiperone is of the order but not ex- actly the same as the values obtained by other groups (List and Seeman, 1981; Leysen et a l . , 1981 1. In our preparations [3H]spiperone was slightly more potent at the S2 receptor than previ- ously reported. The fact that the affinity of [3Hlspiperone at the serotonergic site differed by
k control
1.. - . 101
v 0 60 120 180 240
time. rnin FIG. 9. Time course of [3H]NPA binding to rat striatal ho- mogenates in the presence of various ions at 37°C. The non- specific binding (in the presence of 1 KM (+)-butaclamol) was not affected by any of the ions present at 120 mM. Spe- cific binding at the D,h"Jhsite of (3H]NPA decreased with ions, where LiCl > NaCl 1 KCI. [3H]NPA binding reached equilib- rium by 45 rnin under all conditions tested.
R A T S T R I A T U M
only fourfold from that of the dopaminergic site and that the receptor density was only 14-20% of the total binding sites detected by [3H]spiperone gave the Scatchard plots apparent linearity.
Ketanserin and five agonists were used to char- acterize the 270 pM L3H]spiperone binding site in rat prefrontal cortex and striatum (both done in the presence of SO p M (-)-sulpiride). The site showed the identical serotonergic rank order of potencies in the two regions: ketanserin > serotonin > trypt- amine > ADTN = dopamine > (-)-noradrenaline. This confirms previous studies where it had been shown indirectly that serotonergic sites existed in the rat striatum. Since our attempt to block this component failed (even with 250 nM ketanserin), our computer-assisted analysis took into account the proportion of serotonin sites. The program "LI- GAND" was given the affinity of [3H]spiperone for the serotonergic site, and allowed to iterate for the proportion of S, sites present under the conditions that we were studying for the dopaminergic re- ceptor.
It may be argued that the conditions that convert the dopamine D, receptor also affect the binding properties of ['Hlspiperone to the serotonergic component of the striatum. Such alterations would be impossible to measure, because of the low amount of ['Hlspiperone bound at 80 pM and the low affinity of ADTN for the serotonin sites. We can, nevertheless, clearly state that under our con- ditions, there is still a nondopaminergic component of competition between ADTN and 13H]spiperone.
J . Neirrochem., Vol. 44. No. 6 , 1985

1934 D . GRIGORIADIS AND P . SEEMAN
The data for rank-order at that low-affinity binding site (Fig. 3) indicate that it is a serotonergic com- ponent in the rat striatum.
The K , of [3H]spiperone for the serotonergic site (220 pM; in the presence of SO ~ L M sulpiride), al- though based on low absolute cpm, was in agree- ment with previously published values by others using ['Hlketanserinispiperone for the study of se- rotonin receptors (530 pM; Leysen et al., 1978), and in agreement with the [3H]spiperone K , (200-400 pM) on serotonin S, receptors in rat frontal cortex, as determined in this laboratory under identical conditions to those used in the present study (un- published). There is a potential error in using this value as a constant in all our calculations. Such an error, however, would mostly or almost solely af- fect the quantification of the serotonergic compo- nent with which we are not primarily concerned. A main finding of the present experiments is that the ADTN/['H]spiperone competition data in the striatum could not be fitted to a two-site model. The "fit" was statistically significantly increased by using a third site for competition (see Fig. 1).
Using Monte Carlo simulations for ligand/re- ceptor interactions, Burgisser (1983) has shown that the successful resolution of separate binding sites depends on the number of concentrations tested, among other factors. We used 30 points (over a 10'"- fold range of concentration), for which there was <5% variation in the cpm/filter (even those at 125 cpmifilter). Under such conditions Burgisser esti- mates that i t is possible to resolve a component as small as 2 or 376, when the K , of this component differs from the other K , by a factor of 100-200 (which is the case in our experiments). Thus, the serotonin component of 7-20% in the present study was justifiably resolved by LICAND based on the Monte Carlo simulation studies of Burgisser.
As measured by ag~nist/[~H]spiperone competi- tion, guanine nucleotide by itself did not alter the states of the D, receptor significantly. This was sur- prising since other researchers have previously been able to obtain some conversion of the high- affinity state to the low-affinity state. For example, it has been reported that 300 IJ.M Cpp(NH)p could convert approximately 20% of the high-affinity sites to low-affinity sites (Huff and Molinoff, 1982; Wreg- gett and Seeman, 1983). Agonist binding to dopa- mine receptors in brain is guanine nucleotide-sen- sitive and the conversion of high-affinity sites to low-affinity sites has been shown by others (Zahn- iser and Molinoff, 1978; Creese et a l . , 1979u,h; Hamblin and Creese, 1982) to be guanine nucleo- tide-dependent. The common feature in these pre- vious studies is that the membrane preparation had been exposed to NaCl during the assay. We believe that it is this NaCl that permits the guanine nucleo- tide-induced conversion of the high-affinity state to be converted to low-affinity state.
NaCl by itself, when present at physiological con- centrations, could partially convert the D,high re- ceptors to D,low. Doubling the NaCl concentration further increased the conversion but still could not effect a complete shift in the affinity states. When guanine nucleotide was added to the incubation me- dium in the presence of sodium, however, complete conversion of the high-affinity state to the low-af- finity state occurred. The conclusion is that sodium and guanine nucleotides exert their actions by dif- ferent mechanisms, and that the effects of guanine nucleotides have a dependence for sodium; that is, in NaC1-free conditions in the striatum, guanine nu- cleotides alone cannot convert DZhigh to D21aw. These results are in good agreement with other systems in which the receptor exists in two states and is pos- tulated to mediate the inhibition of adenylate cy- clase (Michel et al., 1980; Rodbell, 1980; Limbird et al., 1982).
It is interesting to compare the striatal D, dopa- mine receptor with the D, receptor found in the anterior pituitary. Although many similarities exist between these two receptors, there are some very definite differences. Guanine nucleotides in porcine and rat anterior pituitary can decrease the fraction of receptors that exist in the high-affinity form. In the presence of only 50 F M Gpp(NH)p, in a sodium- free medium, full conversion can be obtained. Sim- ilarly the presence of 100 mM NaCl alone is suffi- cient to convert fully the high-affinity state to the low-affinity state (George et al., 1983a,h). This sug- gests that the regulation of the D, receptor in the pituitary differs from that of the striatal D, receptor.
Although ADTN competition for ['Hlspiperone binding in the presence of various ions showed lithium to be the most potent in converting D,high to DZloW (converting almost 90% of the high-affinity re- ceptors to their low state), concentration-response curves of lithium on [3H]NPA binding showed its potency to be marginally better than that of sodium (45 +- 9 mM vs. 69 +- 7 mM). [3H]NPA also has high affinity for the D, site in the striaturn. Since there is no specific D, antagonist available, the di- rect effect of the ions on the pure D,high state of the receptor remains to be examined in the future.
We had previously used the term "D," to signify DZhigh dopamine receptors that were resistant to conversion to the low-affinity state (see Seeman, 1980 for review; Seeman, 1982; Wreggett and Seeman, 1983). This term is no longer necessary since all the D, dopamine receptors in both brain and pituitary can now be fully converted from their state of high affinity for dopamine to their state of low affinity for dopamine.
Acknowledgment: We thank Dr. Susan R. George, Dr. Jim Wells, Dr. Masayuki Watanabe, and Dr. L. Endrenyi for valuable discussion and assistance. We thank Dr. P. Munson and Dr. D. Rodbard of the National Institutes of
J . Neurochem.. Vol, 44, N o . 6 , 1985

CONVERSION OF D, RECEPTORS FROM HIGH TO LOW AFFINITY 1935
Health, Bethesda, for providing the LIGAND program and for their advice. This work was supported by the Ontario Mental Heal th Foundation, and Canadian Psy- chiatric Research Foundation and the Medical Research Council of Canada.
REFERENCES Battaglia G. and Titeler M. (1982) [’H]-N-Propylapomorphine
and [iH]-spiperone binding in brain indicate two states of the D2-dopamine receptor. Erir. J . Pharmucol. 81, 493-498.
Burgisser E. (1983) Model testing in radioligand/receptor inter- action by Monte Carlo simulation. J . R e c ~ p t . Res. 3, 261 - 281.
Creese I. and Snyder S. H. (1978) 3H-Spiroperidol labels sero- tonin receptors in rat cerebral cortex and hippocampus. Eur. J . Phnrmucol. 49, 201-202.
Creese I. , Usdin T., and Snyder S. H. (1979~) Guanine nucleo- tides distinguish between two dopamine receptors. Nature 278, 577-578.
Creese I . . Usdin T. B. . and Snvder S. H. (19796) Dopamine re- ~
ceptor binding regulated b; guanine nucleotides. k o / . Phar- mwol . 16, 69-76.
De Camilli P., Macconi D., and Spada A. (1979) Dopamine in- hibits adenylate cyclase in human prolactin-secreting pitu- itary adenomas. Nutrrre 278, 151 - 154.
De Lean A , . Stadel J. M., and Lefkowitz R. J. (1980) A ternary complex model explains the agonist specific binding prop- erties of the adenylate cyclase coupled beta-adrenergic re- ceptors. J . B i d . Chcm. 255, 7108-7112.
Di Paolo T., Gagne B . , Poyet P., and Labrie F. (1982) [’ H] Apomorphine binding to a GTP-sensitive dopaminergic site in bovine anterior pituitary gland. Con. J . Biochrm. 60, 1043- 1049.
George S., Binkley K.. and Seeman P. (1983~) Dopamine re- ceptor sites and states in human brain. J . N~irrri l Trmsm. 18 (Suppl.), 149-156.
George S. R. . Watanabc M., and Seeman P. (1983h) Commen- tary: the dopamine receptor in the anterior pituitary gland, in Dop ( I in in r Re c,ep tors, A in ericci n Cli ern icul So cie t y Sy m- p o ~ i ~ i r n Series 224 (Kaiser C. and Kebabian J . W., eds), pp. 93-99. American Chemical Society. Washington. D.C.
Goldstein M., Lieberman A,, Lew J. Y., Asano T., Rosenfeld M. R., and Makman.M. H . (1980) Interaction of pergolide with central dopaminergic receptors. Proc . N d . Acud. %i.
Hamblin M. and Creese 1. (1982) ’H-Dopamine binding to rat striatal D-2 and D-3 sites: enhancement by magnesium and inhibition by guanine nucleotides and sodium. Lrjir Sci. 30, 1587- 1595.
Huff R. M. and Molinoff P. B. (1982) Quantitative determination of dopamine receptor subtypes not linked to activation of adenylate cyclase in rat striatum. Proc. Not/. A u I ~ . Sc.i. USA 79, 7561 -7565.
Lazareno S. (1983) Effects of GTP on ‘H-domperidone binding and its displacement by dopamine in rat striatal homoge- nates. J . Recrpt. Res. 3, 163-175.
Leff S. E. and Creese I . (1982) Solubilization of D-2 dopamine receptors from canine caudate: agonist-occupation stabil-
USA 77, 3725-3728.
izes guanine nucleotide sensitive receptor complexes. Biochem. Biophys. Res. Comrnmn. 108, 1150-1157.
Leysen J. E., Niemegeers C. J . E . , Tollenaere J . P., and Lad- uron P. M. (1978) Serotonergic component of neuroleptic receptors. Nuture 272, 168-171.
Leysen J . E., Awouters F., Kennis L., Laduron P. M.. Vanden- berk J . , and Janssen P. A. J. (1981) Receptor binding profile of R41 468, a novel antagonist at 5-HT2 receptors. Life Sci. 28, 1015-1022.
Leysen J. E., Niemegeers C. J. E., Van Nueten J. M., and Lad- uron P. M. (1982) LiH]-Ketanserin (R 41 468), a selective ’H- ligand for serotonin-2 receptor binding sites: binding prop- erties, brain distribution and functional role. M o l . Phur- mucol. 21, 301-314.
Limbird L . E., Speck J. L., and Smith S. K. (1982) Sodium ion modulates agonist and antagonist interactions with the human platelet alpha,-adrenergic receptor in membrane and solubilized preparations. Mol. Phnrmacol. 21, 609-617.
List S. J. and Seeman P. (1981) Resolution of dopamine and se- rotonin receptor components of 3H-spiperone binding to rat brain regions. Proc. Nut/. Acad. Sci. USA 78, 2610-2624.
Michel T., Hoffman B. B., and Lefkowitz R. J. (1980) Differ- ential regulation of the alpha?-adrenergic receptor by so- dium and guanine nucleotides. Nntrrre 288, 709-71 l .
Munson P. and Rodbard D. (1980) “Ligand”: a versatile com- puterized approach for characterization of ligand-binding systems. A n d . Biocherii. 107, 220-239.
Pedigo N. W., Reisine T. D. , Fields J. Z., and Yamamura H. I . (1978) ’H-Spiroperidol binding to two receptors in both the corpus striatum and frontal cortex of the rat brain. Errr. J . Phurmac,ol. 50, 45 1-453.
Rodbell M. (1980) The role of hormone receptors and GTP reg- ulatory proteins in membrane transduction. Nrrrrri.c> 284, 17- 22.
Seeman P. (1980) Brain dopamine receptors. Phcr~mrc.ol. Rev . 32, 229-313.
Seenian P. (1982). Nomenclature of central and peripheral do- paminergic sites and receptors. Bioc,hcwi. P/7ti!’/nfItd. 31, 2563-2568.
Sibley D. R., De Lean A . , and Creese 1. (1982) Anterior pituitary dopamine receptors. Demonstration of interconvertible high and low affinity states of the D2 dopamine receptor. J . B i d . Chem. 257, 6351-6361,
Titeler M . and Seeman P. (1979) Selective labelling of different dopamine receptors by a new ’H-ligand: ’H-N-propyl-nor- apomorphine. Errr. J . Phnrmucol. 56, 291 -292.
Wreggett K. A. and Seeman P. ( 1983) Classification and agonist requirements of dopamine receptors. Acfrr P h i r m . Siiec. 1 (SUPPI.). 30-30.
Wreggett K . A . and Seeman P. (1984) Agonist high- and low- affinity states of the D,-dopamine receptor in calf brain: partial conversion by guanine nucleotide. Mid. Phurincic~ol.
Wreggett K . A . . George S. R. . and Seeman P. (1982) Modulation of the Dz dopamine receptor in calf caudate by guanine nu- cleotide. Soc.. Nwro.cc,i. Ahsrr. 8, 719.
Zahniser N. K. and Molinoff P. B. (1978) Effect of guanine n u - cleotides on striatal dopamine receptors. Nurrrr~ 275, 453- 455.
25, 10-17.
J . Nertrocheitr.. Vo l . 44, N o . 6 , I985
Copyright © 2022 FDOKUMEN






![Solution Structure of the Na+ form of the Dimeric Guanine Quadruplex [d(G3T4G3)]2](https://static.fdokumen.com/doc/165x107/6318f44265e4a6af370f95cf/solution-structure-of-the-na-form-of-the-dimeric-guanine-quadruplex-dg3t4g32.jpg)













