
Compared effects of Mpl ligand and other cytokines on human MK differentiation
15
Compared Effects of Mpl Ligand and Other Cytokines on Human MK Differentiation NATACHA VITRAT,~ KARINE COHEN-SOLAL; FRANCOISE NOROL,~ JOSETTE GUICHARD,~ ELISABETH CRAMER,~ WILLIAM VAINCHENKER,~ FRANCOISE WENDLING: NAJET DEBILI~ "INSERM U362, Institut Gustave Roussy, Villejuif, France; bINSERM U474, Hapital Henri Mondor, CrCteil, France Key Words. Endomitosis Platelet shedding Mpl ligand PEG-rHuMGDF Cytokine . Stroma Soluble Mpl ABSTRACT The discovery of the Mpl ligand (Mpl-L), also called thrombopoietin (TPO), has facilitated in vitro investigation of human megakaryocy- topoiesis. By confocal microscopy, endomitosis appeared as abortive mitosis skipping late stages of mitosis. No telophase and cytokinesis were observed. A spherical multipolar spindle which limits chromatid segregation was observed. The nuclear envelope subsequently reformed isolat- ing all chromatids in a single nucleus. Platelet shedding was ultrastructurally studied. Platelet release occurred after formation of long cyto- plasmic extensions (proplatelet formation), constriction areas delineating platelet territo- ries. Heterogeneity in platelet size may be determined by the length of these extensions. Pegylated-recombinant human megakaryocyte growth and development factor, a truncated form of Mpl-L, was the most efficient cytokine to produce proplatelet-hearing megakaryocytes (MKs) and platelets in vitro. However, functional platelets with a normal ultrastructure could he produced in the presence of a combination of other cytokines. Finally, we investigated whether the induction of MK differentiation by the MS-5 stromal cell lines is due to Mpl-L. MS-5 cells synthesized Mpl-L transcripts and a biologi- cally active protein. When human CD34' cells were grown in contact or noncontact cultures with MS-5 cells, MK differentiation was observed. Soluble Mpl receptor (sMpl-Fc) addi- tion inhibited MK growth, suggesting that the MK-promoting activity was due to Mpl-L pro- duction. Marrow stromal cell lines derived from TPO-'. mice were also able to sustain MK growth. Despite the absence of any production of Mpl-L, the sMpl-Fc continued to inhibit MK differentia- tion. This result suggests that the sMpl has a direct inhibitory effect and may explain the divergent results in the literature concerning the precise role of Mpl-L on the MK terminal differentiation. Stem CeUs 1998;I6(suppl2):37-51 INTRODUCTION Platelets play a crucial role in hemostasis, and their number must be kept constant. Megakaqocytopoiesis, which leads to platelet production, is a complex cellular phenomenon. Indeed, this regulation takes place at different cellular levels [I]. At the level of megakaqocyte (MK) progenitors, proliferation regulates the number of marrow MKs. During terminal differentiation, two additional parameters regulate platelet production: MK polyploidization and cytoplasmic maturation. A process called endomitosis, which theo- retically implicates the absence of nuclear envelope breakdown, is responsible for MK polyploidization Thrombopoietin: From Molecule to Medicine STEM CELLS 1998;16(suppl2):37-51 OAlphaMed Press. All rights reserved.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Compared effects of Mpl ligand and other cytokines on human MK differentiation

Compared Effects of Mpl Ligand and Other Cytokines on Human MK Differentiation NATACHA VITRAT,~ KARINE COHEN-SOLAL; FRANCOISE NOROL,~
JOSETTE GUICHARD,~ ELISABETH CRAMER,~ WILLIAM VAINCHENKER,~ FRANCOISE WENDLING: NAJET DEBILI~
"INSERM U362, Institut Gustave Roussy, Villejuif, France; bINSERM U474, Hapital Henri Mondor, CrCteil, France
Key Words. Endomitosis Platelet shedding Mpl ligand PEG-rHuMGDF Cytokine . Stroma Soluble Mpl
ABSTRACT The discovery of the Mpl ligand (Mpl-L),
also called thrombopoietin (TPO), has facilitated in vitro investigation of human megakaryocy- topoiesis. By confocal microscopy, endomitosis appeared as abortive mitosis skipping late stages of mitosis. No telophase and cytokinesis were observed. A spherical multipolar spindle which limits chromatid segregation was observed. The nuclear envelope subsequently reformed isolat- ing all chromatids in a single nucleus. Platelet shedding was ultrastructurally studied. Platelet release occurred after formation of long cyto- plasmic extensions (proplatelet formation), constriction areas delineating platelet territo- ries. Heterogeneity in platelet size may be determined by the length of these extensions. Pegylated-recombinant human megakaryocyte growth and development factor, a truncated form of Mpl-L, was the most efficient cytokine to produce proplatelet-hearing megakaryocytes (MKs) and platelets in vitro. However, functional
platelets with a normal ultrastructure could he produced in the presence of a combination of other cytokines. Finally, we investigated whether the induction of MK differentiation by the MS-5 stromal cell lines is due to Mpl-L. MS-5 cells synthesized Mpl-L transcripts and a biologi- cally active protein. When human CD34' cells were grown in contact or noncontact cultures with MS-5 cells, MK differentiation was observed. Soluble Mpl receptor (sMpl-Fc) addi- tion inhibited MK growth, suggesting that the MK-promoting activity was due to Mpl-L pro- duction. Marrow stromal cell lines derived from TPO-'. mice were also able to sustain MK growth. Despite the absence of any production of Mpl-L, the sMpl-Fc continued to inhibit MK differentia- tion. This result suggests that the sMpl has a direct inhibitory effect and may explain the divergent results in the literature concerning the precise role of Mpl-L on the MK terminal differentiation. Stem CeUs 1998;I6(suppl2):37-51
INTRODUCTION Platelets play a crucial role in hemostasis, and their number must be kept constant. Megakaqocytopoiesis,
which leads to platelet production, is a complex cellular phenomenon. Indeed, this regulation takes place at different cellular levels [I]. At the level of megakaqocyte (MK) progenitors, proliferation regulates the number of marrow MKs. During terminal differentiation, two additional parameters regulate platelet production: MK polyploidization and cytoplasmic maturation. A process called endomitosis, which theo- retically implicates the absence of nuclear envelope breakdown, is responsible for MK polyploidization
Thrombopoietin: From Molecule to Medicine STEM CELLS 1998;16(suppl2):37-51 OAlphaMed Press. All rights reserved.

38 Mpl-L and MK Differentiation
[2, 31 and leads to a single polylobulated nucleated cell with a 2”N DNA content [4, 51. It is associated with an increase in MK cytoplasmic volume and thus indirectly regulates platelet production. The process of MK polyploidization is poorly characterized. It is still a matter of debate as to whether polyploidization is due to a DNA endoreduplication in the absence of mitosis entry or an incomplete mitosis shutting out cytokinesis [6]. Platelet shedding by itself is directly dependent on cytoplasmic maturation, i. e., develop ment of demarcation membranes and cytoskeleton. It is generally considered that platelet shedding requires the formation of long cytoplasmic extensions termed proplatelets [7-111. However, it is also pos- sible that demarcation membranes directly delineate platelet territories and that future platelets are released to subsequent membrane fragmentation [ 12- 141.
Numerous cytokines act in vivo and in vitro on megakaryocytopoiesis. However, the recent isolation and cloning of the Mpl ligand (Mpl-L), also called thrombopoietin (TPO) or megakaryocyte growth and development factor (MGDF) for its truncated form, has shown that this cytokine was the predominent homeostatic regulator of platelet production [15-201. Unexpectedly, Mpl-L acts on all stages of MK dif- ferentiation including progenitor proliferation, MK polyploidization, and maturation [21,22]. Furthermore, it has been suggested that the in vitro action of several cytokines on MK maturation may require the pres- ence of Mpl-L since MK maturation was greatly impaired by the addition of a soluble Mpl receptor (sMpl) [18, 231. However, mice deficient in Mpl-L (TPO.’.) or in c-mpl (c-mpt’.) exhibit a parallel reduction in MK and platelet numbers, suggesting that Mpl-L is dispensable during terminal maturation [24-271. Mpl- L, as erythropoietin, is a humoral growth factor synthesized by the kidney and the liver [15, 16, 191. Nevertheless, Mpl-L is also synthesized by marrow stromal cells [28] and may be involved in a paracrine type of regulation of megakaryocytopoiesis and hematopoiesis [28-311. The precise role of the stroma- derived Mpl-L on megakaryocytopoiesis is still poorly understood and some authors have suggested that it plays a crucial role in the regulation of platelet production in thrombocytopenia [30, 311.
This manuscript summarizes recent results from our group concerning human megakaryocytopoiesis, i.e., characterization of endomitosis and platelet shedding [32,33] and studies on the precise effects of Mpl-L and different cytokines on these processes [34]. It could be demonstrated that Mpl-L plays an important role in vitro on MK terminal differentiation which can be nearly entirely replaced by a combination of pleiotrop- ic cytokines. Stroma derived from TPO’. mice was able to support human megakaryocytopoiesis as well as a normal stroma. Our results unexpectedly showed that a sMpl had an inhibitory effect on the MK proliferation independently of the presence of Mpl-L.
MATERIALS AND METHODS
Blood and Bone Marrow Cells Blood CD34’ cells were isolated from leukapheresis samples performed on patients undergoing autole
gous peripheral blood stem cell transplantation. Bone marrow cells were obtained from normal adult donors undergoing hip surgery. Informed consent was obtained from all donors. Mononuclear cells were separated on a Ficoll gradient (Lymphoprep; Nycomed Pharma; Norway) and used for isolation of CD34’ cells.
Isolation of CD34’ Cells CD34’ cells were purified using a magnetic cell sorting system (mini MACS; Miltenyi Biotec
GmbH; Bergisch Gladbach, Germany) in accordance with the manufacturer’s recommendations. Their purity was determined by flow cytometry using phycoeqthrim (PE)-HPCA2 and was over 80%.
Cell Sorting Bone marrow CD34’ cells were treated with neuraminidase to prevent binding of platelets [35]. Cells
were subsequently incubated with PE-HPCA2 (Becton Dickinson; Mountain View, CA) and fluorescein isothiocynate (FITC)-anti-CD4la or antiLCD38 monoclonal antibodies (mAbs) (Immunotech; Lumigny,

Vitrat, Cohen-Solal, Norol et al. 39
France) and sorted on a FACVantage flow cytometer (Becton Dickinson). Cells were sorted into CD34'CD41- and CD34'CD41' or CD34+CD38'ow and CD34TD38' cell fractions.
Murine Cell Lines MS-5 cells, a generous gift from K. J. Mori (Niigata University; Niigata, Japan), were grown in
a-minimal essential medium (a-MEM) and 10% fetal calf serum (FCS) (Dutscher; Brumath, France). Cells were trypsinized every 10 days and used between passages 7 to 15.
Stromal cell lines from TPO' mice were established as described by ltoh et al. [36] with some modifications. Briefly, bone marrow cells from one femur of a TPO ' mouse were directly flushed in a 25 cmz Costar tissue culture-treated flask containing 8 ml myeloid long-term culture medium for murine cells (Stem Cell Technologies, Inc.; Vancouver, Canada) supplemented with M freshly prepared hydrocortisone (Sigma; St. Louis, MO). Flasks were depopulated of the nonadherent cells twice a week during four weeks by a complete medium change. At week 4, the adherent cells were trypsinized (tryp- sinelEDTA, GIBCO BRL; Grand Island, NY) and seeded in six petri dishes (10-cm diameter) in the same culture medium. After one week, dishes were totally y-irradiated with 9 Gy (Cob" apparatus), and cells were then grown in a-MEM, GIBCO BRL) supplemented with only 10% FCS. Clones which developed (between two to six per dish) were individually picked up and transferred for amplification in 25 cm2 flasks in a-MEM plus 10% FCS. Out of 30 clones picked, only seven could establish permanent cultures. Two of them were used for subsequent experiments (TPO-KO 1 and TPO-KO 7).
BdF3 cells (parental and transfected with the murine c-mpl cDNA) were maintained in a-MEM supplemented with 10% FCS and 2.5% WEHI-3 conditioned medium (CM).
Human Cytokines and Cytokine Receptors Recombinant human (rHu) interleukin 3 (IL-3; a gift from Immunex; Seattle, WA) and rHuIL-6 were
both used at a final concentration of 100 U/ml (3 and 5 ng/ml, respectively). Recombinant human stem cell factor (rHuSCF) and PEG-rHuMGDF (gifts from Amgen; Thousand Oaks, CA) were usually used at a final concentration of 50 ng/ml and 10 ng/ml, respectively. In some experiments, a murine (m) sMpl- Fc (a gift from ZymoGenetics; Seattle, WA) was added at variable concentrations from 0.5 to SO pg/ml. The ms4-1-BB-Fc receptor was used as a control (generous gift from D. Cosmann, Immunex).
Cell Cultures CD34' cells were cultured in Iscove's modified Dulbecco's medium with penicillin/streptomycinlglu-
tamine and 11 .5 pmoVl a-thioglycerol (Sigma). Cultures were usually performed in serum-free conditions [22]. CD34' cells were cultured in 6 or 24-well tissue culture plates, in 3- or 1-ml volumes, respectively. CD34'CD41' cells were grown in 96-well plates in a 100 pl volume.
For coculture experiments, Ba/F3 cells were washed and deprived of WEHI-3 CM for 4 h before being plated at a concentration of 1,000 cells/ml on subconfluent MS-5 cell monolayers in 24-well plates (Costar; Cambridge, MA). Cell growth was examined with an inverted microscope. CD34' cells were plated on sub- confluent MS-5 or TPO' cell monolayers in 24-well plates at a concentration ranging from 2,000 to 15,000 cells in a medium containing 10% FCS and 1% bovine serum albumin or in serum-free conditions (see below) with or without sMpl-Fc, with or without 10 ng/ml PEG-rHuMGDF. In some experiments, a tran- swell (0.45 pm microporous membrane, cyclopore; Falcon, Becton Dickinson; Grenoble, France) was inserted between the stromal cell layer and target cells which were grown in the upper chamber of the insert.
After seven to 12 days nonadherent cells were recovered and stained by an FITC anti-CD4lb mAb (TAB, a generous gift from R. Mac Ever; Oklahoma City, OK). Each sample was diluted in 300 p1 of phosphate-buffered saline (PBS) and analyzed on a flow cytometer (FACSort, Becton Dickinson) (see below). The absolute number of MK in each sample was determined. Adherent cells were fixed for 1 min with methanol and stained with the FITC antLCD41 mAb. Cultures were examined under an inverted

40 Mpl-L and MK Differentiation
microscope equipped for fluorescent microscopy (Diaphot, Nikon; Tokyo, Japan). In order to compare the number of MK grown in cultures performed with and without transwells by means of flow cytometry, adherent cells were detached by trypsinization and treated as above.
Determination of MKs and Platelet Numbers Produced in Culture The total numbers of MKs and platelets were determined by flow cytometry after FITC-TAB
(anti-CD4lb) labeling. Cells from each culture condition were distributed in the same volume. For each sample, the acquisition rate was the same (100 seconds). MKs were defined as brightly-positive CD41 cells with scatter properties of nucleated cells. Culture-derived platelets were enumerated as CD4 1' events with the same scatter properties as blood platelets. Samples were analyzed with a FACSort flow cytometer (Becton Dickinson). Proplatelet-displaying MKs were defined as cells exhibiting one or more cytoplasmic processes with areas of constriction.
Determination of MK Ploidy MK ploidy was measured by a double-staining technique and flow cytometry [37]. MKs were
identified after labeling with FITC-TAB, while DNA staining was performed by incubating the cells in a solution of propidium iodide (50 pg/ml in isotonic NaCl containing 100 pg/ml RNAse; Merck; Darmstadt, Germany) and 0.1% Tween 20 (Sigma) for 1 h. Control cells were stained with an irrelevant FITC-IgG, mAb and propidium iodide.
Detection of the Activation-Dependent Antigen P-Selectin (CD62) on Platelets Cultured cells were stimulated for 10 min at 37°C with 2 U/ml thrombin (Stago; Asnikres, France) added
directly to the culture well. Activated platelets were incubated for 30 min with both PE-auti-CD62 (2 pg per ml) and FITC-TAB. Cells were then fixed for 1 h with an equal volume of 0.5% paraformaldehyde. Control cells were fixed in the same manner prior to activation. Cells were subsequently resuspended in PBS.
Ultrastructural Studies Cultured cells were examined by electron microscopy. Cells were fixed with 1.25% glutaraldehyde
in 0.1 mM phosphate buffer for 1 h at 22"C, washed, post-fixed with osmium tetroxide, dehydrated, and embedded in Epon. Thin sections were examined with a Philips CM 10 electron microscope (Philips; Eindoven, Netherlands) after lead citrate staining.
Immunolabeling for Confocal Microcopy Cells were centrifuged at 300 g for 10 min at 37°C onto slides covered with polylysine (1 mg/ml,
Sigma), fixed for 5 min with a microtubule stabilizing buffer MTSB (80 mM K-PIPES, pH 6.5, 5 mM EGTA and 2 mM MgC12 for a 5 X solution) containing 3% paraformaldehyde and then permeabilized for 15 min with the same buffer containing 0.2% saponin at room temperature. Antibodies used included mouse antibodies (a mixture of two mAbs directed towards a and p tubulin), a rabbit polyclonal anti- body against von Willebrand factor (vWF) and a human antikinetochore antibody. DNA was labeled by Hoechst 33258 at 7.5 ng/ml (Hoechst 33258, Sigma) for 15 min in the dark.
Ribonuclease Protection Assay A XhoI-PvuII fragment containing nucleotides 1 to 540 of the Mpl-L coding sequence was inserted
into the polylinker site of the Bluescript M13 vector (Stratagene; La Jolla, CA) and a 32P-labeled anti- sense riboprobe was transcribed from the T7 promoter. As an internal control for each reaction sample, a '*P-labeled antisense murine actin riboprobe transcribed from the pBACT5 plasmid (kindly provided by F. Dautry, Villejuif, France) was used [38]. Total RNA (20 pg) prepared from MS-5 cells was hybridized with radioactive probes at 55°C overnight. Nonhybridizing RNA was digested with RNase A

Vitrat, Cohen-Solal, Norol et al. 41
(10 pg/ml) and RNase T1 (1,000 U/ml) for 1 h at 37°C. To stop RNase action SDS (0.6 %) and proteinase K (145 yg/ml) were added for 15 min at 37°C. Protected fragments were extracted in the presence of 15 pg of carrier transfer RNA with phenol/chloroform/isoamyl alcohol and precipitated with absolute ethanol at -20°C. Fragments were resolved on a 4% polyacrylamide, 7 M urea gel and autoradiographed on hyperfilmm MP (Amersham; Les Ulis, France). The size of the protected fragments was determined using labeled Mspl-digested pBR322 (New England Biolabs; Beverly, MA).
RESULTS AND DISCUSSION Cultures of marrow or blood CD34' cells, stimulated with PEG-rHuMGDF alone or in combination
with SCF, generate a large proportion of MKs. MKs are fully mature at day 12 in culture of marrow CD34' cells and slightly later in culture of blood CD34' cells (14-16 days). These cultures, therefore, facilitate the observation of the endomitotic and platelet shedding processes.
Endomitosis Is Abortive Mitosis MKs started to polyploidize after about six days of culture. These cultured cells were examined to study
whether MKs enter mitosis during polyploidization. At day 9, three-color immunofluorescence labeling was performed with antibodies against tubulin (FITC) to visualize the mitotic spindle, antibodies against vWF (identification of MKs) or kinetochores (localization of chromosome centromere) (TRITC) and the Hoechst dye (labeling of DNA). Another labeling was also performed with an antilamin B antibody to visualize the nuclear envelope. A total of more than 300 endomitotic MKs were observed. This approach allowed us to demonstrate that MKs entered into mitosis with a breakdown of the nuclear envelope and a condensation of the chromosomes. Endomitosis processed normally until anaphase. As illustrated in Figure 1, the mitotic spin- dle was complex and contained several poles. Their numbers increased with ploidy. These spindles were spherical with polar microtubules extending from one pole to another. Each pole involved several individual spindles. Metaphases with an alignment of sister chromatids were observed. Thereafter, chromatids migrated towards the centrosomes and kinetochores were located close to each aster (Fig. 1). Next, chromatids segre- gated but remained close to each other. This was essentially due to both the short length and complexity of the spindles. Indeed, chromatids segregated all along each spindle pig. 2) but, due to their intrication, remained close to each other. At the end of the anaphase, chromosomes decondensed. No telophase and cytokinesis were observed, but the nuclear envelope reformed, confining all chromatids into a single nucleus.
These data confirm a recent report in which the same strategy was used [39]. The authors also observed that sister chromatids were located in close proximity. They interpreted this result as a consequence of the absence
Figure 1. Presence of a spherical mitotic spindle and an anaphase in an endomitotic MK. Cells were centrifuged on a slide, fixed with a microtubule stabilizing buffer containing 3% paraformaldehyde and then permeabilized with 0.2% saponin. They were labeled with an antitubulin antibody and an antikinetochore antibody and then examined by confocal microscopy. A) antitubulin staining shown alone (FITC). Note that the complex mitotic spindle has a spherical conformation; B ) staining with the antikinetochore antibody (TRITC); C ) superimposition of a double-stain- ing with the antitubulin mAb (FITC) and the antikinetochore (TRITC). This clearly demonstrates that sister chromatids segregate to each pole of the spindle. Note that no nuclear envelope is observed.

42 Mpl-L and MK Differentiation
Figure 2. Presence of an anaphase in an endomitotic MK. Same labeling as in Figure I ; the double-staining has been superimposed at diferent levels. It showed that chromatids seg- regated along the complex spindle toward the diferent poles.
of anaphase B due to a lack of outward move- ment of the spindle poles. From the present result, it seems likely that these outward move- ments cannot occur because of the complex spindle organization with centrosomes involved in different spindles and polar micro- tubules extending from each pole to another. This may also explain the differences in size of asters which may attract different chromatid numbers. Further experiments will be required to demonstrate this hypothesis.
Together, these results demonstrate that endomitosis is true but abortive mitosis, skip- ping the telophase and cytokinesis, and is not endoreduplication. It was also assumed that MKs do not enter mitosis because they lack cyclin B1 or have a defective cdc2 activity 140- 431. We have also demonstrated that cyclin B1 is present and colocalizes with the spindle in endomitotic MKs [33]. In addition, a normal histone H1 kinase activity associated with
cyclin B1 is present in polyploid MKs. It remains to be determined whether the histone H1 kinase activity is related to an association of cyclin B1 with cdc2.
Presently, the mechanisms responsible for the switch from a mitotic to an endomitotic process during MK differentiation are still unknown. Two different mechanisms which may be associated could be involved: A) a modification in the regulation of the cell cycle. Indeed, it has recently been suggested that p21 may play an important role in the polyploidization of MKs 1441; B) an abnormality in structural proteins that are involved in migration of organelles or cytokinesis such as actin, myosin, or kinesin.
Proplatelet Formation is Associated with Platelet Shedding The quality of the maturation of cultured MKs has been greatly improved with the use of purified
CD34' cells, serum-free medium and stimulation by Mpl-L. Under an inverted microscope, it was pos- sible to observe a large fraction of MKs becoming filamentous (proplatelet formation). These long pseudopods elongated and broke irregularly to give rise to detached fragments (platelets). Therefore, these cultures allowed examination of platelet shedding at the ultrastructural level.
On an electron microscope, cytoplasmic expansions could be observed and each MK may have sev- eral cytoplasmic extensions. These were apparently preceded by changes in the chromatin appearance which became condensed as well as by the alignment and dilatation of the demarcation membranes located at the periphery of the cytoplasm. Along these cytoplasmic extensions, constriction points appeared which delimited platelet territories. As shown in Figure 3A and B, the size of platelet territo- ries was much smaller at the periphery of the extension than at its origin. Normal-sized platelets con- taining organelles (a-granules, open canalicular system and microtubules) were observed (Fig. 3). By immunofluorescence, discoid platelets were surrounded by a microtubule coil and filamentous actin. These platelets spread onto polylysine-coated glass slides and reorganized their cytoskeleton. Platelets produced in culture could also be activated by thrombin as demonstrated by the expression of CD62 on their surface.
This part of the work confirms the results obtained by Choi et a.2. 145,461, showing that it is possible to obtain platelet production in vitro without a stromal layer, and that platelet release does not require

Vitrat, Cohen-Solal, Norol et al. 43
Figure 3. Ultrastructure of platelet-shedding MKs and platelets obtained in the presence of PEG- MGDF from CD3& cell. A ) A mature MKpresenting signs of platelet formation. This M K displays a long cytoplasmic extension with dilated demarcation membranes (dm). The constriction areas delineate platelet territories (pp ) of much smaller size at the distal than at the proximal part of the pseudopod. Apparently detached platelets are present (p). Magnification X4100. B ) A smaller mature MK presenting signs of platelet formation. Proplatelet for-
mation not yet detached can be observed (pp) as well as true detachedplatelets. The extension of the cytoplasmic exten- sion is much smaller than on the previous MK (A) and platelets released are of bigger size. Magnification X4100.
blood flow forces. It also confirms previous reports showing that platelet shedding occurs after the formation of long cytoplasmic extensions (proplatelet formation) arising from the unfolding and dilata- tion of the demarcation membranes. These data, however, do not exclude that other cellular mechanisms for platelet production may occur, although this process was the only one observed in cultured human MKs. This mechanism of platelet formation may explain the heterogeneity in platelet size in normal physiology and pathology. Indeed, platelets which arose from the more distal part of the extensions were of much smaller size than those arising from the proximal part. It can be hypothesized that the size of the future platelets may directly depend on the length of the cytoplasmic extension (see Fig. 3A and B) [32]. Characterization of the different cellular molecules, which directly regulate proplatelet formation, will allow a more complete understanding of platelet shedding.
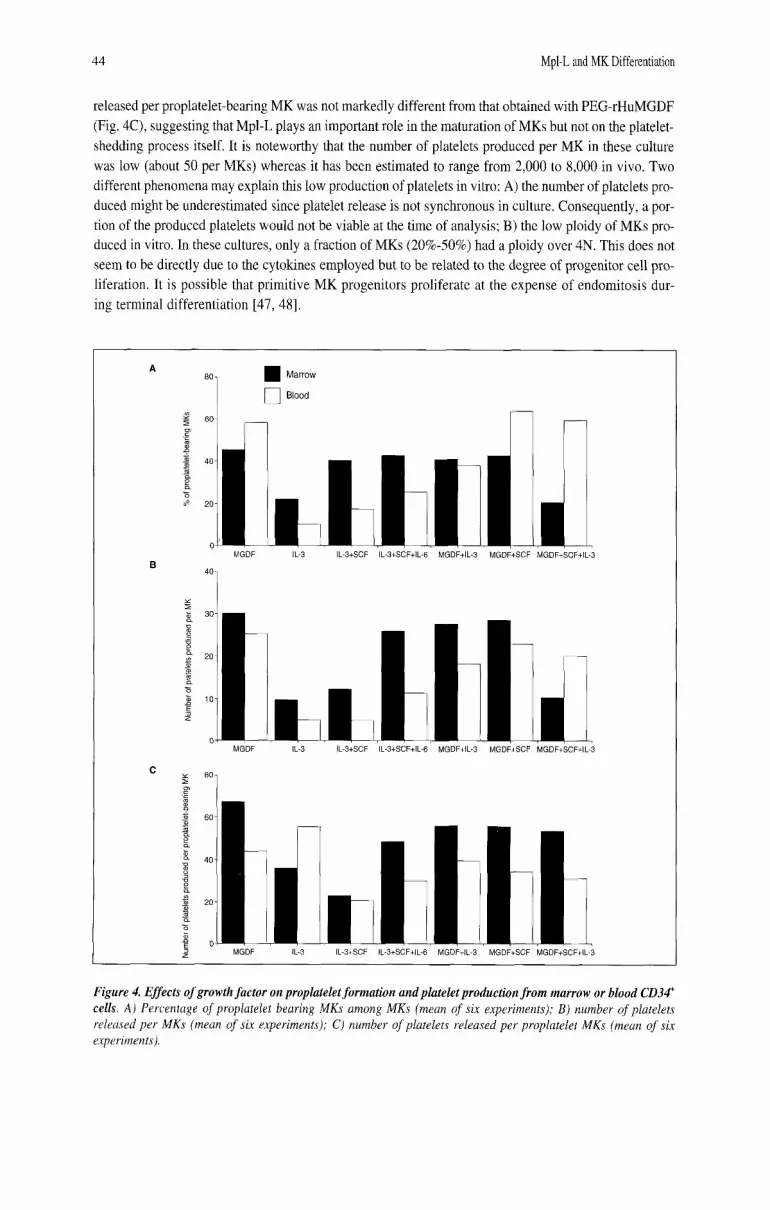
Effects of Mpl-L and Other Cytokines on the Production of Proplatelet-Bearing MKs and Platelets We subsequently examined the effects of Mpl-L and other cytokines on the platelet production in
vitro. Platelets were quantitated by flow cytometry as CD41' elements with the same scatter properties as blood platelets. In a first series of experiments, cytokines were tested on blood and marrow-derived CD34' cells. PEGrHuMGDF was used alone or in different combinations with IL-3 plus SCF, or SCF plus IL-6. IL-3 was the only cytokine other than PEG-rHuMGDF, which induced MK differentiation from CD34' cells. However, the percentage of proplatelet-bearing MKs was 2.5 to fourfold less than with PEG-rHuMGDF, suggesting that IL-3 was not able to alone induce a complete MK maturation (Fig. 4A). As a consequence, the number of platelets produced by IL-3 was diminished in the same order of mag- nitude as the number of proplatelet bearing MKs (data not shown). The number of platelets released by MK in the presence of IL-3 was similarly diminished (Fig. 4B). In contrast, the number of platelets
44 Mpl-L and MK Differentiation
released per proplatelet-bearing MK was not markedly different from that obtained with PEG-rHuMGDF (Fig. 4C), suggesting that Mpl-L plays an important role in the maturation of MKs but not on the platelet- shedding process itself. It is noteworthy that the number of platelets produced per MK in these culture was low (about 50 per MKs) whereas it has been estimated to range from 2,000 to 8,000 in vivo. Two different phenomena may explain this low production of platelets in vitro: A) the number of platelets pro- duced might be underestimated since platelet release is not synchronous in culture. Consequently, a por- tion of the produced platelets would not be viable at the time of analysis; B) the low ploidy of MKs pro- duced in vitro. In these cultures, only a fraction of MKs (20%-50%) had a ploidy over 4N. This does not seem to be directly due to the cytokines employed but to be related to the degree of progenitor cell pro- liferation. It is possible that primitive MK progenitors proliferate at the expense of endomitosis dur- ing terminal differentiation [47, 481.
Marrow
0 Blood
MGDF
I MGDF
I MGDF
iL-3
t IL-3
I IL-3
IL-S+SCF IL-3+SCFtIL-6 MGDF+IL-3 MGDFtSCF
L IL-3tSCF
r IL-S+SCF
I WGDF+SCF+IL-3
IL-3+SCF+lL-6 MGDF+IL-3 MGDFtSCF MGDF+SCFtlL-3
I L-S+SCF+IL t MGDF+IL-3
Figure 4. Effects of growth factor on proplatelet formation and platelet production from marrow or blood CD34' cells. A) Percentage of proplatelet bearing MKs among MKs (mean of six experiments); B ) number of platelets released per MKs (mean of six experiments); C) number of platelets released per proplatelet MKs (mean of six experiments).

Vitrat, Cohen-Solal, Norol et al. 45
Addition of SCF plus IL-6 to IL-3 allowed the growth of similar numbers of MKs compared to PEG-rHuMGDF, but the production of platelets was 2.5- to threefold lower. Addition of PEG- rHuMGDF to all the different cytokine combinations increased the production of MKs and platelets. However, the percentages of proplatelet-bearing MKs or the number of platelets released per MKs were not significantly different from cultures stimulated by PEG-rHuMGDF alone (Fig. 4A, B, and C). The percentage of MKs produced in the different culture conditions was markedly dependent upon the com- bination of cytokines used (from 75% in the presence of PEG-rHuMGDF alone to 7% in the presence of IL-3, IL-6 and SCF); thus differences in the cell populations present in culture may interfere with MK maturation and platelet production.
In a second series of experiments, we compared the effects of PEG-rHuMGDF to the combination of IL-3, SCF and IL-6 on proplatelet formation and platelet shedding using a population of more mature MK progenitors (CD34'CD41' cells) which synchronously differentiate within five to seven days into mature MKs. Contaminant cells were rare (<lo%). PEG-rHuMGDF and the combination of the three cytokines (SCF, IL-3, and IL-6) gave a similar number of MKs. However, the number of proplatelet- bearing MKs was higher in the presence of PEG-rHuMGDF, as well as the platelet production (46 or 11 1 platelets per 1 X lo3 CD34'CD41' cells, for IL-3+SCF+IL-6 versus PEG-rHuMGDF) (Fig. 5) . Ploidy distribution and mean ploidy value (MPV: 8) were similar in both conditions and much higher than those observed with the total CD34' cell population. This directly demonstrates that ploidy of mature MKs in culture is mainly dependent on the differentiation stage of the MK progenitors from which they derive.
No differences in the ultrastmcture of MK or platelets produced in the presence of PEG-rHuMGDF or a combination of three cytokines were observed [34].
These results suggest that Mpl-L plays a key role in platelet production by regulating the MK number, but not by having a specific role in MK maturation. Other cytokines, especially in combination, have similar effects on MK maturation. This result is in agreement with the recent results obtained in c-mpl'. or TPO' mice in which the numbers of MKs and platelets are decreased in parallel [24-271. In addition, the ultrastructure of MKs and platelets from these mice is similar to those of wild-type mice [25]. In con- trast, mice deficient for the transcriptional factor NF-E2 have a defect in the maturation of MKs with an abnormal demarcation membrane system leading to an ineffective platelet production [49]. Therefore, it
PEG-rHuMGDF
Proplatelet-bearing MKs Platelets
Figure 5. Effects of growth factor on proplatelet formation and platelet production from marrow CD34' CD41' cells. A ) Number of proplatelet bearing MKs per plated CD34' CD41' cell (mean of four experiments); B ) number of platelets released per plated CD34' CD41' cell (mean offour experiments).

46 Mpl-L and MK Differentiation
seems likely that NF-E2 transcriptionally regulates some genes responsible for platelet shedding, pre- sumably genes implicated in the organization of the cytoskeleton. In contrast, the transduction signal induced by Mpl-Llc-mpl does not specifically regulate this cellular aspect of MK maturation.
Our results are opposite of those obtained by Kuushansky et ul. who found that Mpl-L was necessary to obtain a full MK development [14, 231. The authors blocked the in vitro MK maturation induced by cytokines such as IL-3, IL-6, IL-11, and SCF with a sMpl. They concluded that these cytokines affect terminal maturation through induction of Mpl-L synthesis. However, using an enzyme-linked immunosorbent assay or a sensitive biological assay, we were unable to detect Mpl-L in the supernatant of our MK cultures stimulated by a combination of three cytokines. In favor of this result, it has been recently shown that IL-11 directly acts on MK differentiation in vitro [50]. This raises the possibility that the sMpl affects MK maturation in vitro independently of its Mpl-L-binding capacity (see below).
We subsequently investigated whether the effects of stromal cells on MK differentiation and maturation were related to the synthesis of Mpl-L or other cytokines.
Effects of Stromal Cells on MK Maturation Are Not Entirely Related to the Synthesis of Mpl-L In a first series of experiments, we used the MS-5 cell line which is capable of sustaining MK
differentiation in short-term liquid cultures [5 I]. Since it was demonstrated that stromal cells synthesize Mpl-L [28,29], we investigated whether Mpl-L mRNA could be detected in MS-5 cells. Total RNAs pre- pared from subconfluent cultures were subjected to RNAse protection experiments. The riboprobe used in these experiments was constructed to protect mRNAs corresponding to wild-type and various trun- cated Mpl-L isoform transcripts [52-541. Two major Mpl-L transcripts were protected in MS-5 cells. The upper band corresponds to mRNA encoding the wild-type, secreted Mpl-L, while the lower band might correspond to a splice variant probably coding for a nonsecreted protein.
To determine if MS-5 cells secreted biologically active Mpl-L protein in the culture supernatants, conditioned medium (CM) was prepared and assayed using the BalF3-c-mpl proliferation assay. CM col- lected after four or eight days slightly stimulated the proliferation of the BalF3-c-mpl cells, but not the parental Ba/F3 cells, when used at a final concentration of 50% (volhol). This activity corresponded to about 5 pg/ml Mpl-L which is the threshold of our assay. In contrast, when BalF3-c-mpl cells were direct- ly cocultivated on MS-5 monolayers, they adhered to the MS-5 cells and actively proliferated whereas parental Ba/F3 cells did not survive for more than two days. Furthermore, addition of 5 pg/ml sMpl-Fc at the initiation of the culture completely inhibited the growth of the BaR3 transfectants. This inhibitory effect was reversed by addition of 1 ng/mL rmuMpl-L. These results indicate that bone marrow-derived MS-5 stromal cells produce small amounts of biologically active Mpl-L in the culture supernatant.
CD34' cells were subsequently cocultured on the MS-5 cell line. As observed under an inverted microscope, large MKs were first noted at day 5. Their number increased until day 7 for the CD34TD38' and day 10-12 for the CD34+CD38'OW subsets. Most MKs with a round morphology were adherent to MS-5 cells during the first days of culture. When MK became fully mature and had morphological changes asso- ciated with proplatelet formation, some of them detached from the stromal cell layer and floated in the supernatant. In order to precisely quantitate the number of MKs produced, a direct enumeration of MKs was performed either with an inverted microscope or flow cytometry on the nonadherent cells or by in situ fluorescence microscopy on the adherent cell fraction after labeling with an FITC anti-CD41 antibody. MK represented 2%-4% of the cells present in supernatants, the other cells being CD34' and CD15' cells. In contrast, MKs comprised the majority of the adherent cells. The addition of PEG-rHuMGDF greatly increased (about fivefold) the number of MKs (Fig. 6; p<0.05). In contrast, the addition of 50 pg/ml of sMpl-Fc in the cocultures nearly completely abolished the MK growth (98% inhibition) in a dose-depen- dent manner (0.5 pg/ml to 50 pg/ml). Further experiments were performed at 5 pg per ml of sMpl-Fc which produced a 60%-80% (average: 74%; p = 0.05) inhibition of MK growth. A partial abrogation of the inhibitory effect mediated by sMpl was observed after addition of 10 ng/ml PEG-rHuMGDF, a dose

Vitrat, Cohen-Solal: Nor01 et al. 41
10,000-
1,000
100
10
1
0 large^^
Total MK
[I Adherent MK
Nonadherent MK
MS-5 MS-5+AMpl-Fc MS-5+PEG-rHuMGDF MS-5+PEG-rHuMGDF+sMpI-Fc
Figure 6. Effects of MS-5 cells on the MK differentiation. Results show the mean offive experiments (four from CD34'CD38' cells, one from CD3@CD3@" cells). The data are expressed as a percentage (by comparing the num- ber of M K observed in cultures with MS-5 alone (100%) to that of other conditions of cultures). CD34'CD3Si cells (10,000) or CD34'CD38'"" cells (2,000, Exp. 5) were grown in 96-well plates for seven or 10 days. Large MKs in each well were enumerated under an inverted microscope. Adherent MKs and nonadherent MKs were identifed and quantitated by in situ irnmunofluorescent labeling or flow cytometry, respectively. Total amount of MKs is the sum of adherent and nonadherent MKs. MS-5 cells were present as a confluent layer on the bottom of the well. PEG- rHuMGDF (10 ng/ml) andor sMpl-Fc (5 pg/ml) were added at the outset of culture. The data were analyzed by a Student's paired t-test.
which completly reverted the growth inhibition mediated by the sMpl-Fc on the Ba/F3 c-mpl cells (Fig. 6). Cultures were also performed in conditions where MS-5 and CD34' cells were separated by a filter
(0.45 pm). As shown by morphology and expression of CD41, MKs were present among the cells growing in the transwell even though they were not in close contact with MS-5 cells present on the bottom of the plate (Table 1). Elongated MKs were present suggesting that platelet production may occur. MK differen- tiation was also inhibited (66%) by the addition of sMpl-Fc (Table 1). Of note, when PEG-rHuMGDF was added in the cultures, the number of MKs produced was about twofold higher than in control cultures with- out MS-5 cells suggesting that MS-5 cells must synthesize other cytokines synergizing with Mpl-L [21-231. In contrast, no MKs were detected when MS-5 cells were omitted from these cultures.
Altogether, these experiments demonstrate that MS-5 cells synthesize Mpl-L, but the role of this stroma-derived Mpl-L on human MK differentiation is not totally straightforward since: A) the inhibitory effect of the sMpl-Fc could not be totally reverted by addition of PEG-rHuMGDF and B) an only weak inhibition of MS-5-induced MK differentiation was observed by addition of an antimurine Mpl-L neutralizing polyclonal antibody (data not shown).
In order to better understand the role of Mpl-L synthesized by the stroma, we derived several stromal cell lines from TP0-I- mice and used two of them for subsequent experiments. These cell lines like MS- 5 cells had the phenotype of immortalized myofibroblastic cells (P. Charbord, personal communication). TPOI- stromal cell lines sustained MK differentiation as well as the MS-5 cells either in coculture or after interposition of a membrane insert (Fig. 7). Proplatelet formation was also observed. Surprisingly, the sMpl-Fc and not a control irrelevant soluble receptor (4-1 BB Fc) inhibited MK differentiation both in TPO.'. or MS-5 coculture. Two hypotheses could explain these unexpected results: A) it is possible that an induction of Mpl-L synthesis by human hematopoietic cells could occur following adhesion or stim- ulation by a stroma-derived cytokine. By means of reverse transcriptase-polymerase chain reaction, however, we were unable to detect human Mpl-L transcripts in the cultured cells; B) an effect of sMpl-Fc

48 Mpl-L and MK Differentiation
Table 1. Comparison 0 1 MK growrh irom CDXCD3d ' i A in dirzct coculturch or in ininswcll plat:s
Megakaryocyte number
Experiment 1 MS-5 2,991 MS-5tPEG-rHuMGDF 8,076 (270%) MS-5tTW 912 (31%) MS-5tTWtsMPL-Fc 110 (4%) MS-5tTWtPEG-rHuMGDF 6,085 (230%) MS-5tTWtPEG-rHuMGDFtsMPL-Fc 4,657 (152%)
Experiment 2 MS-5 519 MS-5tTW 198 (38%) MS-5tTWtPEG-rHuMGDF 14,605 (2,794%) TWtPEG-rHuMGDF 8,136 (1568%)
CD34'CD3Xt cells (10,000) from cytapheresis (Exp. 1) or marrow (Exp. 2) were grown in 24-well plates in the presence or absence of a transwell plate in serum or serum-free conditions (Exp. 2). MS-5 cells were present as a subconfluent layer on the bottom of the well. The sMpl-Fc (5 mglml) and PEG-rHuMGDF (SO ng/ml) were added at the onset of the culture. MKs were enumerated by flow cytometry. TW: transwell plate.
10,000
1,000
t 100
E
Y 10 E
13
3 C
1
0.1
M~I-L-'-
O M
I 0
5
I ~Mpl-FC I PEGMuMGDF I 'EG-rHuMGDF+sMI r Fc Transwell
answellcPEG-rHuMGDF
Figure 7. Effects of MS-5 and TPOI' stromal cells on the MK differentiation. The experiment was performed as in Figure 6. Results show the mean of one experiment pergc"rmed in triplicate from blood CD34' cells. The data are expressed as the number of MKs grown from I X Id CD34' cells (mean ? SE). MKs from the adherent and non- adherent fractions were pooled and studied by flow cytometry after labeling with an anti-CD41 antibody. MS-5 or TPO" (TPO-K07) cells were present as a confluent layer on the bottom of the well. PEG-rHuMGDF (10 ng/ml) and/or sMpl-Fc ( 5 pg/ml) were added at the ontset of culture. Two other experiments with marrow CD34' cells gave identical results.
on MK differentiation by trapping a yet undefined cytokine or by directly binding to cells. This last hypothesis seems more likely since it has been recently reported that sMpl had a synergistic effect with SCF and Flt3 ligand on primitive hematopoiesis [%I. It was hypothesized that this effect was mediated through a direct interaction with the cell membrane [55] .

Vitrat, Cohen-Solal, Norol et al. 49
Our data demonstrate that stromal cells from TPO-'. mice synthesize a cytokine or a combination of cytokines which is able to induce a complete MK differentiation. This result explains why TPO-'- mice have a basal megakaryocytopoiesis (15% of wild-type mice). The present coculture system may facilitate the characterization of molecules, other than Mpl-L, which regulate platelet production in basal conditions.
ACKNOWLEDGMENT We are grateful to J.L. Nichol (Amgen) for providing the rHuSCF and PEG-rHuMGDF, D. Foster
(ZymoGenetics) for the murine sMpl-Fc, D. Cosman (Immunex) for IL-3 and the murine s4-1BBFc fusion protein, and F. de Sauvage (Genentech) for the TPO~'~ mice. This work was supported by the INSERM, the Institut Gustave Roussy, and by grants from la Ligue de Paris contre le Cancer.
REFERENCES
1 Hoffman R. Regulation of megakaryocytopoiesis.
2 Ebbe S, Yee T, Carpenter D et al. Megakaryocytes increase in size within ploidy groups in response to the stimulus of thrombocytopenia. Exp Hematol
3 Therman E, Sarto GE, Stubblefled PA. Endomitosis: a reappraisal. Hum Genet 1983:63:13-18.
4 Odell TT, Jackson CW Jr, Reiter RS. Generation cycle of rat megakaryocytes. Exp Cell Res 1968;
5 Odell TT, Jackson CW, Friday TJ. Megakaryo- cytopoiesis in rats with special reference to polyploidy. Blood 1970;35:775-782.
6 Ebbe S. Biology of megakaryocytes. Prog Hemost Thromb 1976;3:211-229.
7 Becker RP, De Bruyn PP. The transmural passage of blood cells into myeloid sinusoids and the entry of platelets into the sinusoidal circulation; a scan- ning electron microscopic investigation. Am J Anat
8 Radley JM, Scurfield G. The mechanism of platelet release. Blood 1980;56:996-999.
9 Radley JM, Haller CJ. The demarcation membrane system of the megakaryocyte : a misnomer? Blood
10 Thiery JB, Bessis M. La genkse des plaquettes 2 partir des mkgacaryocytes observCs sur la cellule vivante. C R Acad Sci 1956;242:290-292.
1 1 Thiery JB, Bessis M. MCcanisme de la plaquetto- genkse. Etude in vitro par la inicrocinkmatographie. Rev Hemat 1956;II: 162-176.
12 Behnke 0. An electron microscope of the megakary ocyte of the rat bone marrow. I. The development of the demarcation membrane system and the platelet surface coat. J Ultrastruct Res 1968;24:412-433.
13 Zucker-Franklin D, Petursson S. Thrombo- cytopoiesis: analysis by membrane tracer and
Blood 1989;74 1196-121 2.
1988;16:55-61.
53:321-328.
1976; 145: 182-205.
1982;60:213-219.
freeze-fracture on fresh human and cultured mouse megakaryocytes. J Cell Biol 1984;99:390-402.
14 Zucker-Franklin D, Kaushansky K. Effect of thrombopoietin on the development of megakary- ocytes and platelets: an ultrastructural analysis.
15 Bartley TD, Bogenberger J, Hunt P et al. Identification and cloning of a megakaryocyte growth and development factor that is a ligand for the cytokme receptor Mpl. Cell 1994;77:1117-1124.
16 de Sauvage FJ, Hass PE, Spencer SD et al. Stimulation of megakaryocytopoiesis and thrombopoiesis by the c-Mpl ligand. Nature 1994;369:533-538.
17 Kaushansky K, Lok S, Holly RD et al. Murine thrombopoietin: the ligand for c-mpl expands megakaryocyte progenitors, induces their differen- tiation and stimulates platelet production. Nature
18 Kaushansky K. Thrombopoietin: the primary regu- lator of platelet production. Blood 1995;86:419-431.
19 Lok S, Kaushansky K, Holly RD et al. Murine thrombopoietin: expression cloning, cDNA sequence and stimulation of platelet production in v i v a Nature 1994;369:565-568.
20 Wendling F, Maraskovsky E, Debili N et al. The Mpl ligand is a humoral regulator of megakaryocy- topoiesis. Nature 1994;369:571-574.
21 Broudy V, Lin NL, Kaushansky K. Thrombopoietin (c-mpl Ligand) acts synergistically with erythropoi- etin, stem cell factor, and interleukin 11 to enhance murine megakaryocyte colony growth and increas- es megakaryocyte ploidy in vitro. Blood 1995; 85:1719-1726.
22 Debili N, Wendling N, Katz A et al. The Mpl-Ligand or thrombopoietin or megakaryocyte growth and dif- ferentiative factor has both direct proliferative and differentiative activities on human megakaryocyte progenitors. Blood 1995;86:25 16-2525.
Blood 1996;88: 1632-1638.
1994;369:568-571.

Mpl-L and MK Differentiation so
23
24
25
26
27
28
29
Kaushansky K, Broudy VC, Lin N et al. Thrombopoietin, the Mpl ligand, is essential for full megakaryocyte development. Proc Natl Acad Sci USA 1995;92:3234-3238.
Alexander WS, Roberts AW, Nicola NA et al. Deficiencies in progenitor cells of multiple hematopoietic lineages and defective megakaryocy- topoiesis in mice lacking the thrombopoietin receptor
Bunting S, Widmer R, Lipari T et al. Normal platelets and megakaryocytes are produced in vitro in the absence of thrombopoietin. Blood 1997; 903423-3429.
de Sauvage FJ, Carver-Moore K, Luoh S-M et al. Physiological regulation of early and late stages of megakaryocytopoiesis by thrombopoietin. J Exp Med 1996;183:65 1-656.
Gurney AL, Carver-Moore K, de Sauvage FJ et al. Thrombocytopenia in c-mpl-deficient mice. Science
Nagahisa H, Nagata Y, Ohnuki T et al. Bone marrow stoma1 cells produce thrombopoietin and stimulate megakaryocyte growth and maturation but suppress proplatelet formation. Blood 1996; 87:1309-1316.
Guerriero A. Worford L. Holland HK et al.
c-Mpl. Blood 1996;87:2 162-21 70.
I994;26S: 1445- 1447.
37 Jackson CW, Brown LK, Sommerville BC et al. Two-color flow cytometric measurement of DNA distributions of rat megakaryocytes in unfixed, unfractionated marrow cell suspensions. Blood
38 Cohen-Solal K, Villeval JL, Titeux M et al. Constitutive expression of Mpl ligand transcripts dur- ing thrombocytopenia or thrombocytosis. Blood
39 Nagata Y, Muro Y, Todokoro K. Thrombopoietin- induced polyploidization of bone marrow megakary- ocytes is due to a unique regulatoly mechanism in late mitosis. J Cell Biol 1997;139:449-457.
40 Datta NS, Williams JL, Caldwell J et al. Novel alter- ations in CDKlKyclin B 1 kinase complex formation occur during the acquisition of a polyploid DNA con- tent. Mol Cell Biol 1996;7:209-223.
41 Garcia P, Calks C. Endoreplication in megakq- oblastic cell lines is accompanied by sustained expression of GUS cyclins and downregulation of cdc25C. Oncogene 1996; 13:695-703.
42 Wang Z, Zhang Y, Kamen D et al. Cyclin D3 is essential for megakaryocytopoiesis. Blood 199.5;
43 Zhang Y, Wang Z, Ravid K. The cell cycle in
1984;63:768-778.
1996;88:2578-2584.
86~3783-3788.
ThromboDoietin is svnthesized bv bone marrow polyploid megakaryocytes is associated with stromal cells. Blood 1$97;90:3444-?+45S.
30 McCarthy JM, Sprugel KH, Fox NE et al. Murine thrombopoietin mRNA levels are modulated by platelet count. Blood 1995;86:3668-3675.
31 Sungaran R, Markovic B, Chong BH. Localization and regulation of thrombopoietin mRNA expression in human kidney, liver, bone marrow, and spleen using in situ hybridization. Blood 1997;89:101-107.
32 Cramer E, Norol F, Guichard J et al. Ultrastructure of platelet formation by human megakaryocytes cultured with the Mpl ligand. Blood 1997;
33 Vitrat N, Cohen-Solal K, Pique C et al. Endomitosis of human megakaryocytes are due to abortive mitosis. Blood 1998 (in press).
34 Norol F, Vitrat N, Cramer E et al. Effects of cytokines on platelet production from blood and marrow CD34 positive cells. Blood 1998 (in press).
35 Dercksen MW, Weismar IS, Richel DJ et al. The value of flow cytometric analysis of platelet glyco- protein on CD34 positive cells measured under conditions that prevent P-selectin-mediated binding of platelets. Blood 1995;86:3771-3782.
36 Itoh K, Tezuka H, Sakoda H et al. Reproducible establishment of hemopoietic supportive stromal cell lines from murine bone marrow. Exp Hematol 1989; 17: 145-153.
8912336.2346.
reduced activity of cyclin-B I-dependent cdc2 kinase. J Biol Chem 1996;271:4266-4272.
44 Matsumura I, Isbikawa J, Nakajima K et al. Thrombopoietin-induced differentiation of a human megakaryoblastic leukemia cell line, CMK, involves transcriptional activation of p21wAf’’c’p’ by STATS. Mol Cell Biol 1997;17:2933-2943.
45 Chen J, Herced-Harjacek L, Groopman JE et al. Regulation of platelets activation in vitro by the c-Mpl ligand, thrombopoietin. Blood 1995;
46 Choi ES, Hokom MM, Chen JL et al. The role of megakaryocyte growth and development factor in terminal stages of thrombopoiesis. Br J Haematol 1996;95:227-233,
47 Amaga M, South K, Cohen JL et al. Interrelation- ship between mitosis and endomitosis in culture of human megakaryocyte progenitor cells. Blood
48 Debili N, Mass6 J, Katz A et al. Effects of recom- binant hematopoietic growth factors (IL-3, IL-6, SCF, LIF) on the megakaryocyte differention of CD34 positive cells. Blood 1993;82:84-95.
49 Shivdasani RA, Rosenblatt MF, Zucker-Franklin D et al. Transcription factor NF-E2 is required for platelet formation independent of actions of throrn- bopoietinlMGDF in megakaryocyte development. Cell 19958 1 :69S-704.
86:4054-4062.
1987;69:486-496.

Vitrat, Cohen-Solal, Norol et a1 51
50
51
52
53
Weich NS, Wang A, Fizgerald M et al. Recombinant human interleukin 11 directly promotes megakary- ocytopoiesis in vitro. Blood 1997;90:3893-3902.
Debili N, Coulombel L, Croisille L et al. Characterization of a bipotent erythro-megakaryocytic progenitor in human bone marrow. Blood 1996;
Chang M-S, McNinch J, Basu R et al. Cloning and characterization of the human megakqocyte growth and development factor (MGDF) gene. J Biol Chem
Gurney AL, Kuang W-J, Xie M-H et al. Genomic structure, chromosomal localization, and conserved
88: 1284-1296.
1995;270:5 1 1-5 14.
54
55
alternative splice forms of thrombopoietin. Blood 1995;85:981-988.
Wada T, Nagata Y, Nagahisa H et al. Characterization of the truncated thrombopoietin variants. Biochem Biophys Res Commun 1995; 2 13: 1091 - 1098.
Ku H, Hirayama F, Kato T et al. Soluble thrombopoietin receptor (Mpl) and granulocyte colony-stimulating factor receptor directly stimulate proliferation of primitive hematopoietic progenitors of mice in synergy with Steel factor or the ligand for Flt3/Flk2. Blood 1996;88:4124-413 1.




















