
Combined supplementation of vanadium and beta-carotene suppresses placental glutathione...
11
Journal of Gastroenterology and Hepatology (2004) 19, 683–693 Blackwell Science, LtdOxford, UKJGHJournal of Gastroenterology and Hepatology0815-93192004 Blackwell Publishing Asia Pty LtdJune 2004196683693Original Article Vanadium and beta-carotene in liver carcinogenesisMB Chattopadhyay Correspondence: Professor Malay Chatterjee, PO Box 17028, Jadavpur University, Kolkata 700 032, India. E-mail: [email protected] or [email protected] Accepted for publication 1 September 2003. HEPATOLOGY Combined supplementation of vanadium and beta-carotene suppresses placental glutathione S-transferase-positive foci and enhances antioxidant functions during the inhibition of diethylnitrosamine-induced rat liver carcinogenesis MITALI BASU CHATTOPADHYAY, MAHENDRA KUMAR CB, P SURESH KANNA, RAJARSHI SANKAR RAY, SOUVIK ROY AND MALAY CHATTERJEE Division of Biochemistry, Department of Pharmaceutical Technology, Jadavpur University, Kolkata, India Abstract Background and Aim: The present study was designed to investigate the chemopreventive effects of combined vanadium (V; 0.5 p.p.m.) and beta-carotene (BC; 120 mg/kg of basal diet) on diethylni- trosoamine (DEN)-induced and phenobarbital (PB)-promoted rat hepatocarcinogenesis. Methods: All rats were subjected to two-thirds partial hepatectomy (PH) at the fourth week. After PH they were administered either trioctanoin alone (groups A¢, B¢, C¢ and D¢) or a single injection of DEN in trioctanoin at a dose of 10 mg/kg of body weight (groups A, B, C and D). Two weeks after the DEN treatment PB was administered (0.05% in basal diet) to all the DEN-treated rats and continued until the end of the experiment. Supplementation of V (groups B and B¢), BC (groups C and C¢) or both V and BC (groups D and D¢) at the doses stated previously were started 4 weeks before DEN administration (at week 0) and continued until the 16th week. Results: It was observed that in the DEN-treated and PB-promoted group (group A) the expression of the numbers and areas of the placental form of glutathione S-transferase (GST-P)-positive altered hepatic foci (AHF) was maximum. Treatment with V (group B) and BC (group C) significantly reduced the expression of GST-P-positive AHF by 29.5% and 42.8%, respectively. An additive protection action (65.7%) was noticed in group D, which received both V and BC for the entire period of the experiment. It was also observed that supplementation of V and BC for the entire period of the experiment signifi- cantly reduced the number and size of the hyperplastic nodules, while the combination treatment worked as an additive effect, reducing the number and size of the hyperplastic nodules to 22% from 89%. More- over, a significantly reduced level of cytosolic glutathione (P < 0.001) and glutathione-S-transferase (P < 0.001) activity and stabilization of aerobic metabolism and hepatic architecture of the cells as com- pared with carcinogen control were observed in the V + BC-treated group. Conclusion: The present study suggests that V, an essential trace element, may be useful in combina- tion with BC, an antioxidant, in the inhibition of experimentally induced rat hepatocarcinogenesis. © 2004 Blackwell Publishing Asia Pty Ltd Key words: beta-carotene, diethylnitrosamine, glutathione, glutathione S-transferase, placental glu- tathione S-transferase, rat hepatocarcinogenesis, vanadium. INTRODUCTION Hepatocarcinogenesis induced by various carcinogens is a multistep and complex process and is a favored model in the rat that facilitates the study of the mech- anism of transformation from a normal cell to a malig- nancy. 1 The liver is often the first organ to be affected by metastasizing cancer. 2 Diethylnitrosamine (DEN) is a well-known liver carcinogen in rats, forming DNA adducts in the liver and inducing hepatocellular carci- nomas without cirrhosis through the development of putative preneoplastic enzyme-altered focal lesions. 3
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Combined supplementation of vanadium and beta-carotene suppresses placental glutathione...

Journal of Gastroenterology and Hepatology (2004) 19, 683–693
Blackwell Science, LtdOxford, UKJGHJournal of Gastroenterology and Hepatology0815-93192004 Blackwell Publishing Asia Pty LtdJune 2004196683693Original ArticleVanadium and beta-carotene in liver carcinogenesisMB Chattopadhyay
Correspondence: Professor Malay Chatterjee, PO Box 17028, Jadavpur University, Kolkata 700 032, India. E-mail:[email protected] or [email protected]
Accepted for publication 1 September 2003.
HEPATOLOGY
Combined supplementation of vanadium and beta-carotene suppresses placental glutathione S-transferase-positive foci and
enhances antioxidant functions during the inhibition of diethylnitrosamine-induced rat liver carcinogenesis
MITALI BASU CHATTOPADHYAY, MAHENDRA KUMAR CB, P SURESH KANNA, RAJARSHI SANKAR RAY, SOUVIK ROY AND MALAY CHATTERJEE
Division of Biochemistry, Department of Pharmaceutical Technology, Jadavpur University, Kolkata, India
AbstractBackground and Aim: The present study was designed to investigate the chemopreventive effects ofcombined vanadium (V; 0.5 p.p.m.) and beta-carotene (BC; 120 mg/kg of basal diet) on diethylni-trosoamine (DEN)-induced and phenobarbital (PB)-promoted rat hepatocarcinogenesis.Methods: All rats were subjected to two-thirds partial hepatectomy (PH) at the fourth week. After PHthey were administered either trioctanoin alone (groups A¢, B¢, C¢ and D¢) or a single injection of DENin trioctanoin at a dose of 10 mg/kg of body weight (groups A, B, C and D). Two weeks after the DENtreatment PB was administered (0.05% in basal diet) to all the DEN-treated rats and continued until theend of the experiment. Supplementation of V (groups B and B¢), BC (groups C and C¢) or both V andBC (groups D and D¢) at the doses stated previously were started 4 weeks before DEN administration(at week 0) and continued until the 16th week.Results: It was observed that in the DEN-treated and PB-promoted group (group A) the expression ofthe numbers and areas of the placental form of glutathione S-transferase (GST-P)-positive alteredhepatic foci (AHF) was maximum. Treatment with V (group B) and BC (group C) significantly reducedthe expression of GST-P-positive AHF by 29.5% and 42.8%, respectively. An additive protection action(65.7%) was noticed in group D, which received both V and BC for the entire period of the experiment.It was also observed that supplementation of V and BC for the entire period of the experiment signifi-cantly reduced the number and size of the hyperplastic nodules, while the combination treatment workedas an additive effect, reducing the number and size of the hyperplastic nodules to 22% from 89%. More-over, a significantly reduced level of cytosolic glutathione (P < 0.001) and glutathione-S-transferase(P < 0.001) activity and stabilization of aerobic metabolism and hepatic architecture of the cells as com-pared with carcinogen control were observed in the V + BC-treated group.Conclusion: The present study suggests that V, an essential trace element, may be useful in combina-tion with BC, an antioxidant, in the inhibition of experimentally induced rat hepatocarcinogenesis.© 2004 Blackwell Publishing Asia Pty Ltd
Key words: beta-carotene, diethylnitrosamine, glutathione, glutathione S-transferase, placental glu-tathione S-transferase, rat hepatocarcinogenesis, vanadium.
INTRODUCTION
Hepatocarcinogenesis induced by various carcinogensis a multistep and complex process and is a favoredmodel in the rat that facilitates the study of the mech-anism of transformation from a normal cell to a malig-
nancy.1 The liver is often the first organ to be affected bymetastasizing cancer.2 Diethylnitrosamine (DEN) is awell-known liver carcinogen in rats, forming DNAadducts in the liver and inducing hepatocellular carci-nomas without cirrhosis through the development ofputative preneoplastic enzyme-altered focal lesions.3

684 MB Chattopadhyay
Experimental analyses and epidemiological studieshave proved that natural dietary constituents are capa-ble of favorably modulating carcinogenic and mutagenicprocesses.4 Vanadium (V), an endogenous constituent ofall or most mammalian tissue, is believed to have a reg-ulatory role in biological systems.5 Previous studies fromthe authors’ laboratory6 and other laboratories7–9 haveestablished V as a significant biological regulator inassessing the physiological and biochemical state ofanimals in a dose-related manner.6 It has been shownthat V at 0.5 p.p.m. in drinking water was effective ininhibiting chemically induced two-stage rat hepatocar-cinogenesis without any toxic manifestation.10
In experimental rat liver carcinogenesis, beta-caro-tene (BC) has shown promise in inhibiting the neoplas-tic process at the stage of initiation induced by severalhepatocarcinogens, as reported from the authors’laboratory11,12 and elsewhere.13,14 There is good epide-miological evidence that carotenoid intake from fruitsand vegetables is linked with reduced cancer risk.Dietary intervention with supplements of BC has notproved to be effective against cancer,15 and someinstances, such as the Finnish trial in 199416 and theCARET trial17 have observed increased risk. A review byCollins discusses this dilemma and highlights the pau-city of data relating to the effect of carotenoids at rea-sonable in vivo concentrations with respect to DNAdamage and repair, cell cycle control and xenobioticmetabolism.18 This review ends with the recommenda-tion that the lack of evidence for the toxicity of BC19
allows its safe use as a dietary supplement and states thatwhen administered for a long time at high doses BC doesnot cause toxicity.20,21 This current study is directed atelucidating the efficacy of both the trace element V andthe antioxidant BC, which, when given together, caneffectively prevent the transformation of neoplastic cellsof liver treated with a single necrogenic dose of DEN.
The initiation of hepatocytes to give rise to foci ofaltered gene expression by genotoxic carcinogens suchas the placental form of glutathione S-transferase,(GST-P), has been well documented.22 In the DEN–phenobarbital (PB) model of rat hepatocarcinogenesis,GST-P has been described as the most effective singlemarker of preneoplasia,23,24 because it scores morealtered hepatic foci (AHF) than other markers in mostcases of hepatocarcinogenesis. GST-P is commonlyused as a method for the identification, quantitationand assessment of rat liver cancer development.25
The tumor marker enzymes such as glutathione(GSH) and glutathione S-transferase (GST) have beenrecognized as important markers in characterizing themetabolic patterns of preneoplastic cells and in thedevelopment of strategies for prevention of carcinogen-esis and early treatment of cancer.26 Reduced GSH, acystein-containing tripeptide, is involved in intracellularprotection from free radical damage and has a definiterole in detoxifying different xenobiotics, includingchemical carcinogens.27,28 GST, with the help of GSH,plays an important role in the detoxification of xenobi-otic compounds.29 The assessment of the toxic responseis mainly based on various detoxification pathways, andof these GSH and GST are the most important mole-cules.30 Thus the study of these biotransforming
enzymes has been found to be very useful and par-ticularly relevant in assessing the chemopreventive effi-cacy of combined supplementation of V + BC against acarcinogenic assault. To evaluate the extent of oxidativephosphorylation in the present study the authors deter-mined the mitochondrial O2 consumption in all groups.Mitochondrial O2 consumption is one of the best suit-able parameters to determine the extent of aerobicmetabolism in the cell.
The purpose of the present study is essentially two-fold. First, to demonstrate the protective response ofcombined supplementation of V + BC in modulatingmorphological, histopathological and biochemicalchanges in rat hepatocarcinogenesis; second to evaluatethe combined effects of V and BC on the developmentof enzyme-altered lesions, their positive phenotype forGST-P and to study the extent of aerobic metabolism bydetermining the mitochondrial O2 consumption of thehepatic cells.
METHODS
Animals and diet
Male Sprague–Dawley rats, obtained from the IndianInstitute of Chemical Biology, Kolkata, India, were usedfor the experiment. Rats were acclimatized for 1 weekbefore the commencement of the experiment. All therats were given a semipurified basal diet (Lipton, Mum-bai, India). Rats weighing 80–100 g were housed in wire-mesh cages (4–5 rats/cage). They had free access to waterand food under controlled environmental conditions ofhumidity (60 ± 5%), lighting (12-h light/dark cycle) andtemperature (23 ± 2∞C). Rats were maintained accord-ing to the NIH guidelines of Care and Use of LaboratoryAnimals throughout the experimental period.
Carcinogen
Diethylnitrosamine was purchased from Sigma Chem-ical (St Louis, MO, USA).
Experimental procedure
A total number of 80 4-week-old rats were divided intoeight groups. All the rats were subjected to two-thirdspartial hepatectomy (PH) at 8 weeks of age. Half of therats (groups A, B, C and D) were administered a singleinjection of DEN in trioctanoin at a dose of 10 mg/kgbodyweight. Following 2 weeks recovery from the sur-gery, the DEN-treated rats were given PB in the basaldiet (at a dose of 0.05% PB per rat) up to the 16th week.Group A rats were the DEN and PB control whilegroup A¢ was the respective normal vehicle control(these rats received a single dose of solvent trioctanoinand 0.2 mL of corn oil per oral, 6 days per week up tothe 16th week). Supplementation of V (0.5 p.p.m. inwater ad libitum; E Merk, Kolkata, India),10 BC(120 mg/100 g of basal diet)31 or both V and BC ingroup B, C and D rats, respectively, were started

Vanadium and beta-carotene in liver carcinogenesis 685
4 weeks before DEN administration (week 0) and con-tinued thereafter up to the 16th week. Groups B¢, C¢and D¢ served as the respective control of groups B, Cand D, respectively, and received V, BC or both V andBC, respectively, at the same dose mentioned previouslythroughout the experimental period. Treatment of V,BC and PB was withdrawn at week 16 and at the ter-mination of the experiment all the rats were killed byexposure to ether [Fig. 1].
Morphology and morphometry of liver
After the animals were killed, the livers were promptlyexcised from all the treated and control rats, weighedand examined on the surface for subcapsular macro-scopic lesions (hyperplastic nodules [HN]). The nod-ules with approximate spheres were measured in two
perpendicular directions to the nearest millimeter intothree categories, ≥3, <3–≥1 and <1 mm, according tothe published criteria of Moreno et al.13
For histological studies, representative longitudinallycut blocks were taken from the left, right-median andright-anterior lobes and the samples were fixed imme-diately in 10% buffered formalin to prevent deforma-tion. At least three slices per liver were embedded in lowmelting point paraffin. Sections of 5 mm thick weredeparaffinized in xylene and rehydrated through 100%,90% and 70% alcohol. Five contiguous sections weremade from each liver slice for routine histopathologicalevaluation using HE staining. Specific hepatocellularlesions observed in HE staining were recognized usinglight microscopy (Model no. 5591; Adcon) according topublished criteria.32,33 The histological slides werecoded so that the particular treatment was unfamiliar tothe individual performing these studies.
Figure 1 The basic ex-perimental protocol. BC,beta-carotene; DEN, dieth-ylnitrosoamine; PB, pheno-barbital; V, vanadium.

686 MB Chattopadhyay
Biochemical estimations
Preparation of liver cytosolic microsomal fractionAfter the animals were killed, livers of either lobe wereexcised, minced and homogenized with ice-cold 1.15%(w/v) KCl solution (pH 7.4) in a Teflon-coated glasshomogenizer to make a 10% (w/v) homogenized solu-tion. The cytosolic fraction was prepared using differ-ential centrifugation. The homogenized liver wascentrifuged at 9000 g for 30 min and the resultantsupernatant fraction was centrifuged at 105000 g for90 min. The supernatant of the 105000 g representedthe cytosolic fraction.
The level of GSH was quantified using the methoddescribed by Ellman.34 For this, 1 mL of cytosol wasmixed with 1 mL of 4% 5-sulfosalicylic acids and themixture was centrifuged at 1500 g for 15 min. Thesupernatant was reacted with 0.1 mM 5,5¢-dithio-bis(2-nitrobenzoic acid) (Sigma). The solution was kept atroom temperature for 10 min and read at 412 mm byHitachi U-2000 spectrophotometer.
The activity of cytosolic GST was determined usingan adaptation of the method described by Habig et al.using 1-chloro-2,4-dinitrobenzene (CDNB) and GSH(Sigma) as substrates.35 The reaction mixture (1 mL)consisted of 0.1 M phosphate buffer (pH 6.5), 1 mMGSH, 1 mM CDNB and a suitable amount of cytosol(1.2–1.6 mg protein/mL). The reaction mixture wasincubated at 37∞C for 5 min and the reaction wasinitiated by the addition of the CDNB. The increase inoptical density at 340 nm was measured spectro-photometrically with a temperature-controlled cellcompartment.
Total protein content
The cytosolic protein and the amount of mitochondrialprotein were estimated using the method described byLowry et al. using bovine serum albumin (BSA) as astandard.36
Mitochondrial isolation
The liver tissue was homogenized in ice-cold 0.25 mol/L sucrose using a Teflon pestle tissue homogenizer. Thehomogenate was then centrifuged (800 g, 4∞C, 10 min).The supernatant was decanted and centrifuged(14000 g, 4∞C, 30 min). The sediment mitochondrialfraction was collected and washed in cold sucrose solu-tion. The mitochondrial pellet was weighed, resus-pended in sucrose solution, sonicated and used forfurther determinations.
Determination of O2 consumption
Mitochondrial O2 consumption was determined polaro-graphically using Clark-type oxygen electrodes. Thetemperature of the system was maintained using an iso-therm circular water bath. The oxygen electrodes and
monitor were calibrated using distilled water at thedesired temperature (25∞C). Adenosine diphosphatewas added to a respiration medium to a final volume of3 mL. The rate of O2 consumption was normalized on aper milligram of mitochondrial protein basis. Data arereported as nmol O2/mg protein/min.
Immunohistochemical staining of placental glutathione S-transferase expression
The liver slices placed into acetone were fixed overnightat -20∞C with one change of fresh acetone.37 Sectionswere placed into poly L-lysine-coated slides (0.5% inwater; Sigma), deparaffinized in xylene and rehydratedwith graded alcohol to water. The liver sections wereacetone-fixed for 5 min, allowed to dry and then rehy-drated in phosphate-buffered saline. Endogenous biotinwas blocked by incubation with avidin solution (50 mg/mL) for 30 min. Non-specific protein binding was min-imized by use of 5% normal serum. The rabbit antiratGST-P antibody was diluted 1:700 in 1% BSA andincubated with the tissue sections overnight at 4∞C.Next, the sections were incubated with a biotinylatedsecondary antibody (goat antirabbit immunoglobulin G[Boehringer Mannheim, Germany] diluted 1:200 in 1%BSA) for 30 min at room temperature. This was fol-lowed by incubation with streptavidin bound to b-galactosidase (Sigma) and subsequent chromogendevelopment with 5-bromo-4-chloro-3-indoyl b-galac-tosidase (Boehringer Mannheim) was carried out asdescribed by Bondi et al.38 The sections were counter-stained with Mayer’s hematoxylin and then coverslipped with Crystal mount. The areas of liver sectionswere determined with a planimeter and the number ofsingle hepatocytes expressing GST-P as well as GST-P-positive foci were determined per cm2 of hepatic tissue.The percentage size distributions of different foci (i.e.<0.05, 0.05–0.20 and >0.2 mm2) were also determinedin all DEN-treated rats. GST-P-positive lesions com-posed of >11 cells were considered as altered liver cellfoci.
Statistical analysis
Significance of difference between mean values of dif-ferent groups of rats was assessed using Student’s t-test.Additive effects of treatment of V and BC were assessedusing two-way23 and three-way24 ANOVA.
RESULTS
Body and liver weights
The final bodyweights of group A rats (DEN + PB) weresignificantly less (P < 0.001) than those of group A¢(normal vehicle control group) at the end of the study.Treatment with V and BC in group D rats effectivelyincreased this fall in bodyweight and the effect is addi-tive in nature because the 23 ANOVA analysis showed that
Vanadium and beta-carotene in liver carcinogenesis 687
the interaction effect between them is not significant(Table 1). The liver weights did not vary predominantlybetween different rat groups (Table 1). The mean rela-tive liver weight was found to be decreased significantly(P < 0.001) in group D rats (which received both V andBC) compared with group A¢ (normal vehicle controlgroup) rats. The 23 ANOVA (tables not shown) alsoshowed that V and BC produce an effect that was greaterthan the individual effect of V and BC because the inter-action effect between them is not significant.
Effect of vanadium and beta-carotene on hepatic nodulogenesis
Table 2 summarizes the effect of V and BC on the inci-dence and multiplicity of hepatic nodular hyperplasia
treated in rats with DEN and promoted using PB. Theappearance of hepatic HN, as judged by their percent-age incidence, total number and multiplicity was pro-nounced in group A rats (DEN + PB control). Themaximum beneficial effect was observed in group Danimals that received both V and BC; the reduction ofincidence of nodules was 40% in this group whereas thereduction percentages were 60% and 70% in group Cand B animals, respectively, which received only V andBC, respectively, when compared with group A (100%),the DEN + PB control group. Again the nodule multi-plicity was reduced significantly in group D rats(P < 0.001) when compared with that of group A.Moreover, treatment with both V and BC reduced thelarger nodules (≥3 mm) most effectively (22.7%) than V(36.5%) and BC (24.4%) alone treated rats when com-pared with group A (47.7%). The 22 ANOVA (table not
Table 1 Final bodyweight, final liver weight and relative liver weight of different groups of rats at the end of the study
Group Treatment Final bodyweight (g) Liver weight (g)Relative liver weight
(g liver/100 g bodyweight)
A¢ Normal control 309.83 ± 7.47† 9.01 ± 1.02 2.90 ± 0.32A DEN + PB control 262.10 ± 9.85‡ 10.34 ± 0.73 3.92 ± 0.33††
B DEN + PB + V 271.72 ± 9.02§ 10.04 ± 0.93 3.68 ± 0.30‡‡
B¢ V control 286.80 ± 8.00 9.76 ± 0.78 3.40 ± 0.33C DEN + PB + BC 281.90 ± 8.46§ 9.48 ± 0.85 3.35 ± 0.28‡‡
C¢ BC control 290.80 ± 7.07 9.39 ± 0.81 3.22 ± 0.25D DEN + PB + V + BC 288.60 ± 8.92¶ 9.25 ± 0.79 3.10 ± 0.40§§
D¢ V + BC control 295.70 ± 8.32 9.05 ± 1.07 3.04 ± 0.37
†Values are mean ± SD for 10 rats. ‡P < 0.001 when compared with group A¢ (normal vehicle control). §P < 0.05 when com-pared with group A (DEN control). ¶P < 0.001 when compared with group A (DEN control). ††P < 0.05 when compared withgroup A¢ (normal vehicle control). ‡‡P < 0.01 when compared with group A (DEN control). §§P < 0.001 when compared withgroup A (DEN control). BC, biliary cirrhosis; DEN, diethylnitrosoamine; PB, phenobarbital; V, vanadium.
From 23 ANOVA analysis for bodyweight. The main effect of A, B and C and the interaction effect of A–B and A–C are significant(P < 0.001). The interaction effect between B and C is not significant, which signifies that the combined effect of V and BC wasadditive in nature. Source: A = DEN; B = V; C = BC.
From 23 ANOVA analysis for liver weight and relative liver weight. The main effects of A and C are significant (P < 0.001) in thecase of liver weight and relative liver weight. The interaction effect between B and C is significant (2.5 for liver weight and 1.68for relative liver weight), which signifies that the combined effect of V and BC was additive in nature. Source: A = DEN; B = V;C = BC.
Table 2 Effect of vanadium and beta-carotene in combination on hepatic nodulogenesis
Group Treatment
No. ofrats with
nodules/totalno. of rats
Nodule incidence
(%)Total no.
of nodules
Nodules relative to size (%)
Average no. ofnodules/nodule-
bearing liver(nodule
multiplicity)≥3 mm <3 ≥ 1mm <1mm
A DEN + PB 10/10 100 89 42 (47.1) 24 (26.9) 23 (26.1) 8.9 ± 1.20†
B DEN + PB + V 7/10 70 52 19 (36.5) 23 (44.2) 10 (19.2) 7.4 ± 0.83‡
C DEN + PB + BC 6/10 60 49 12 (24.4) 11 (22.4) 26 (53.0) 8.1 ± 0.91‡
D DEN + PB + V + BC 4/10 40 22 5 (22.7) 10 (45.4) 7 (31.8) 5.5 ± 0.77§
†Values are mean ± SD. ‡P < 0.01 when compared with groups B and A and groups C and A. §P < 0.001 when compared withgroups D and A using Student’s t-test. BC, biliary cirrhosis; DEN, diethylnitrosoamine; PB, phenobarbital; V, vanadium.
From 23 ANOVA analysis for nodule multiplicity. The interaction effect between A and B is not significant (0.13), which signifiesthat the combined effect of V and BC is additive in nature. Source: A = DEN + V; B = DEN + BC.

688 MB Chattopadhyay
shown) analysis of the data of nodule multiplicityshowed that V and BC when supplemented togetherhad an additive effect in inhibiting the nodule multiplic-ity, because the interaction effect between them is notsignificant.
Effect of vanadium and beta-carotene on hepatic histopathology
Histopathological examination in liver sections from thenormal untreated group, group A¢ (Fig. 2a), as well asother control groups, groups B¢, C¢ and D¢ (figures notshown) which received V or BC alone or both in com-bination, respectively, revealed normal liver parenchy-mal cells with granulated cytoplasm and small uniformnuclei radially arranged around the central vein. In con-trast, gross structural alterations were seen in group Arats (DEN + PB control) with predominantly baso-philic, eosinophilic and clear cell foci (Fig. 2b). Exten-sive vacuolation was observed in the cytoplasmencircling the nucleus with masses of acidophilic mate-rials. Some nuclei in the cells were large and hyperchro-matic with prominent and centrally located nucleoli(Fig. 2b). Phenotypically altered hepatocyte popula-tions in the form of altered liver cell foci and nodules invarying extent were noticeable throughout the hepaticparenchyma of all groups treated with DEN, that is,groups A, B, C, and D. The cellular architecture ofhepatic lobules in group D rats that received both V andBC in combination for 16 weeks seemed to be almostcomparable to normal (Fig. 2c). Group D elicited themaximum protection against DEN-induced hepatocar-cinogenesis of both V and BC for the entire period ofstudy, which was reflected in the almost normal hepa-tocellular architecture. Liver cells from this group werefound to contain compact cytoplasmic material withonly clear cell foci (Fig. 2c). The nucleocytoplasmicratio was decreased considerably as compared withgroup A (DEN + PB control). The configuration ofsinuses appeared normal within Kupffer cells. The sizeof the nuclei resembled that of normal cells and thebinucleated cells were extremely less frequent. However,a moderate improvement in hepatic cellular architecturewas observed in groups B and C rats receiving V and BCalone, respectively (figures not shown). A considerablevacuolation was still observed in the cytoplasm, and thecompactness of the hepatocytes was not like the normaluntreated control group (group A¢). The liver sectionsfrom these groups presented a predominance of clearcell foci rather than eosinophilic and/or basophilic foci.The numbers of binucleated cells were less frequent ascompared with group A¢ with normal sized nuclei.
Effect of vanadium and beta-carotene on hepatic cytosolic glutathione level and glutathione S-transferase activity in different groups
A single intraperitoneal injection of DEN in group Arats resulted in a significant increase (P < 0.001) in the
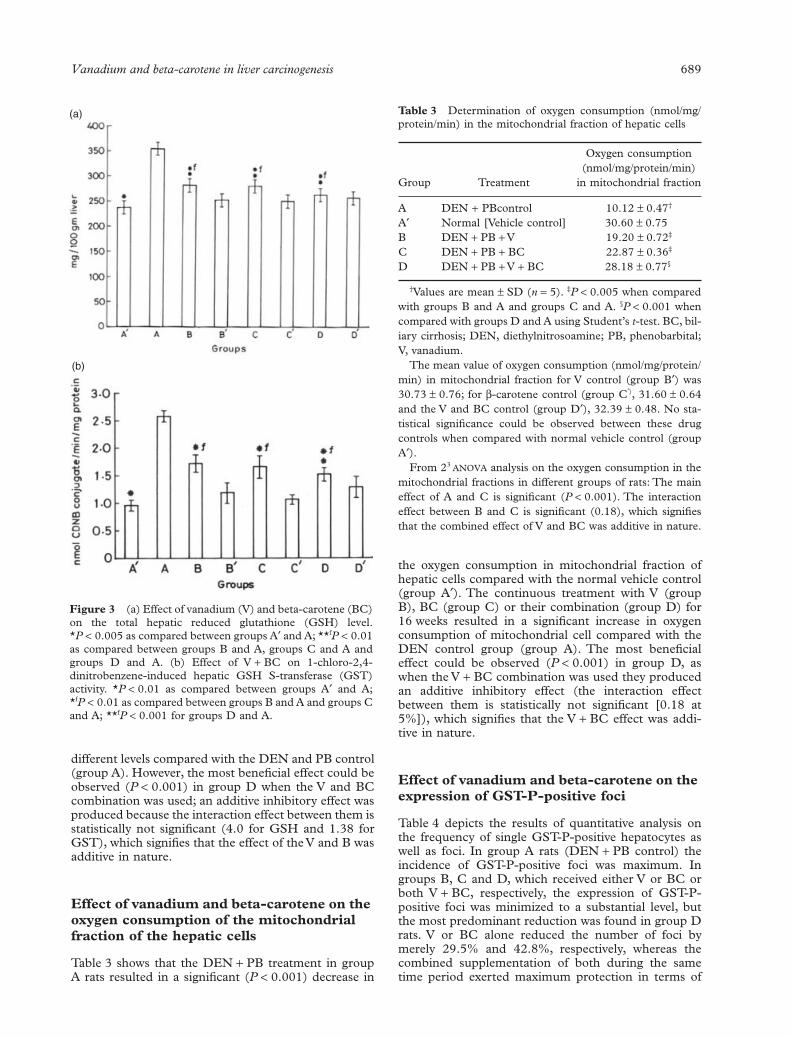
total hepatic GSH pool (Fig. 3a) and GST activity(Fig. 3b) in the hepatic cytosol compared with thenormal vehicle control (group A¢). The continuoustreatment with V (group B), BC (group C) or their com-bination (group D) for 16 weeks resulted in a significantdecrease in both the total GSH pool and GST activity at
Figure 2 (a) Thin section of the normal untreated rat liver(group A¢) showing normal hepatocellular architecture (HE¥400). (b) Section of the rat liver (group A) showing abnormalarchitecture after initiation with diethylnitrosoamine (DEN;10 mg/kg bodyweight) and promoted by phenobarbital (PB)(0.05%) for 16 weeks (HE ¥400). (c) Section of theDEN + PB-treated rat liver (group D) showing almost normalarchitecture after continuous supplementation of vanadium(0.5 p.p.m.) and beta-carotene (120 mg/kg of basal diet) for16 weeks (HE ¥400).
b
c
Vanadium and beta-carotene in liver carcinogenesis 689
different levels compared with the DEN and PB control(group A). However, the most beneficial effect could beobserved (P < 0.001) in group D when the V and BCcombination was used; an additive inhibitory effect wasproduced because the interaction effect between them isstatistically not significant (4.0 for GSH and 1.38 forGST), which signifies that the effect of the V and B wasadditive in nature.
Effect of vanadium and beta-carotene on the oxygen consumption of the mitochondrial fraction of the hepatic cells
Table 3 shows that the DEN + PB treatment in groupA rats resulted in a significant (P < 0.001) decrease in
the oxygen consumption in mitochondrial fraction ofhepatic cells compared with the normal vehicle control(group A¢). The continuous treatment with V (groupB), BC (group C) or their combination (group D) for16 weeks resulted in a significant increase in oxygenconsumption of mitochondrial cell compared with theDEN control group (group A). The most beneficialeffect could be observed (P < 0.001) in group D, aswhen the V + BC combination was used they producedan additive inhibitory effect (the interaction effectbetween them is statistically not significant [0.18 at5%]), which signifies that the V + BC effect was addi-tive in nature.
Effect of vanadium and beta-carotene on the expression of GST-P-positive foci
Table 4 depicts the results of quantitative analysis onthe frequency of single GST-P-positive hepatocytes aswell as foci. In group A rats (DEN + PB control) theincidence of GST-P-positive foci was maximum. Ingroups B, C and D, which received either V or BC orboth V + BC, respectively, the expression of GST-P-positive foci was minimized to a substantial level, butthe most predominant reduction was found in group Drats. V or BC alone reduced the number of foci bymerely 29.5% and 42.8%, respectively, whereas thecombined supplementation of both during the sametime period exerted maximum protection in terms of
Figure 3 (a) Effect of vanadium (V) and beta-carotene (BC)on the total hepatic reduced glutathione (GSH) level.*P < 0.005 as compared between groups A¢ and A; **fP < 0.01as compared between groups B and A, groups C and A andgroups D and A. (b) Effect of V + BC on 1-chloro-2,4-dinitrobenzene-induced hepatic GSH S-transferase (GST)activity. *P < 0.01 as compared between groups A¢ and A;*fP < 0.01 as compared between groups B and A and groups Cand A; **fP < 0.001 for groups D and A.
(a)
(b)
Table 3 Determination of oxygen consumption (nmol/mg/protein/min) in the mitochondrial fraction of hepatic cells
Group Treatment
Oxygen consumption(nmol/mg/protein/min)
in mitochondrial fraction
A DEN + PBcontrol 10.12 ± 0.47†
A¢ Normal [Vehicle control] 30.60 ± 0.75B DEN + PB + V 19.20 ± 0.72‡
C DEN + PB + BC 22.87 ± 0.36‡
D DEN + PB + V + BC 28.18 ± 0.77§
†Values are mean ± SD (n = 5). ‡P < 0.005 when comparedwith groups B and A and groups C and A. §P < 0.001 whencompared with groups D and A using Student’s t-test. BC, bil-iary cirrhosis; DEN, diethylnitrosoamine; PB, phenobarbital;V, vanadium.
The mean value of oxygen consumption (nmol/mg/protein/min) in mitochondrial fraction for V control (group B¢) was30.73 ± 0.76; for b-carotene control (group C¢), 31.60 ± 0.64and the V and BC control (group D¢), 32.39 ± 0.48. No sta-tistical significance could be observed between these drugcontrols when compared with normal vehicle control (groupA¢).
From 23 ANOVA analysis on the oxygen consumption in themitochondrial fractions in different groups of rats: The maineffect of A and C is significant (P < 0.001). The interactioneffect between B and C is significant (0.18), which signifiesthat the combined effect of V and BC was additive in nature.

690 MB Chattopadhyay
minimizing the expression of GST-P-positive foci by66.7%. A 22 ANOVA (table not shown) analysis of theGST-P-positive foci data showed that the interactioneffect between V and BC is not significant, signifying theadditive nature of V and BC, which minimizes theexpression of these foci more effectively than V or BCalone. A significant decrease (P < 0.001) in the averagearea (mm2) of GST-P-positive foci was observed in therats treated with V + BC (group D) compared withgroup A. Results of the 22 ANOVA analysis of the datarevealed that V + BC additively inhibit the average areaof foci, as shown by the interaction being not signifi-cant. Rats in groups A¢, B¢, C¢ and D¢ (control groups ofA, B, C, and D) did not show any detectable foci.Figure 4 shows the positive and negative immunostain-ing of GST-P foci.
DISCUSSION
The findings of the present investigation demonstratedthat the supplementation of V and BC in combinationgreatly inhibited DEN-initiated and PB-promoted rathepatocarcinogenesis by stabilizing the levels of hepaticantioxidant enzymes as well as by preventing the
changes to the hepatic architecture induced by DEN.Such a protective effect, which could be envisagedthrough inhibition of expression of GST-P-positive foci,was maximum in group D, in which V and BC were sup-plemented in combination.
The present findings regarding oxygen uptake inmitochondrial cells suggest there is re-establishment ofaerobic metabolism in the treated groups B, C and D(supplemented with either V or BC or V + BC, respec-tively) compared with the DEN control group. Thisfinding is in line with previous observations39 that thisre-establishment of aerobic metabolism not only limitsprogression of uncontrolled division and growth of can-cer cells but also, if held in this state long enough,induces programmed cell death. If the cancer cellsrestart aerobic metabolism, they revert the cells back tobeing quasi-normal cells again. However, genetic dam-age is not corrected, but if the cells are held in a normalstate of aerobic metabolism with time they will gothrough the normal process of programmed cell deathand the cancer cells will be permanently eliminated.39
Results of the present study showed that administra-tion of V and BC, either alone or in combination,throughout the experimental period reduces the inci-dence, total number, multiplicity and size of the HN
Table 4 Additive effect of vanadium (V) and beta-carotene (BC) on the inhibition of the placental form of glutathione S-transferase (GST-P)-positive foci in different groups of rats
Group TreatmentSingle GST-P-positive
cells/cm2
GST-P-positive foci Size distribution (%)
No./cm2 Average area (mm2) <0.05 0.05–0.20 >0.20
A DEN + PB 99.6 ± 6.05 12.68 ± 5.30 7.30 ± 2.90† 7.2 11.9 80.9B DEN + PB + V 66.9 ± 11.9 8.79 ± 5.86 4.20 ± 2.50‡ 16.8 12.2 71.0C DEN + PB + BC 49.1 ± 12.8 7.25 ± 2.90 3.60 ± 3.10‡ 25.8 13.4 60.8D DEN + PB + V + BC 29.3 ± 3.32 4.34 ± 2.56 2.04 ± 1.50§ 32.1 14.7 46.8
†Values represented mean ± SD for 10 rats. ‡P < 0.01 as compared with group A (DEN control). §P < 0.001 as compared withgroup A (DEN control). DEN, diethylnitrosoamine; PB, phenobarbital.
From 23 ANOVA analyses for GST-P-positive foci (no./cm2 )and area (mm2): The interaction effect between A and B is not sig-nificant (0.17 for no. of foci and 0.21 for area of foci), which signifies the combined effect of V and BC was additive in nature.
Figure 4 (a) Immunohistochemical staining of placental glutathione-S-transferase (PGST)-positive foci (¥100). (b) Negativecontrol of PGST-positive foci (¥100).
a b

Vanadium and beta-carotene in liver carcinogenesis 691
induced by DEN. The authors observed from thepresent study that V + BC demonstrated a pronouncedeffect in inhibiting the appearance of hepatic nodules of>3 mm in size from 47% to approximately 23%.Although V or BC alone could inhibit the average num-ber of nodules in groups B and C rats, respectively, anadditive inhibitory effect was observed when theseagents were supplemented together (in group D rats).In view of this, inhibition and regression of nodulargrowth by the combined actions of V + BC observed inthe present study may be important for cancer chemo-prevention. A striking observation, however, was theinhibition of GST-P in single hepatocytes (Fig. 4b).These cells accumulate in dense clusters mainly aroundthe central veins, clearly distinguished for foci (Fig. 4a).GST-P is a valuable marker of preneoplastic foci, nod-ules and tumors26 and is not expressed in normal hepa-tocytes.41 The decrease in the total number of singleGST-P-positive foci hepatocytes in group D rats wassignificant when analyzed using Student’s t-test. Liter-ature describing the in vivo effects of V + BC on thedevelopment of enzyme-altered lesions, phenotype pos-itive for GST-P, is scarce. The results of this investiga-tion confirm the data of a recent study in rats in whichsimultaneous vitamin E and BC supplementationcaused suppression of the enzyme-altered GST-P-posi-tive foci.42 A similar pattern was observed with plasmag-glutamyl transpeptidase-positive foci. The highestdose of BC at 50 mg/day/rat provided the greatestreduction in severity.42 Moreover, also in agreementwith the present study, BC with V significantly reducedthe nodular incidence of nodule and inhibited GST-P-positive foci expression in a rat hepatocarcinogenesismodel.
From the present work, it is evident that althoughcellular damage and architecture alteration haveoccurred during DEN treatment (group A), V + BCsupplementation seemed to diminish these changes.Although there was slight cellular and nuclear change,the arrangement of the hepatocytes appeared normal.This hypothesis is further strengthened by the resultsobtained that showed significant decreases in the levelof GSH and GST in V + BC-supplemented rats (groupD) compared with the carcinogen control group(group A). The cellular GSH, alone or with GST, canfunction as a non-critical neutrophil for conjugationreactions and plays an important role in the deactiva-tion of electrophilic compounds.13,43 Therefore, anincrease in the GSH level along with GST activity ingroup D rats (V + BC-supplemented) indicates theability of the cells to detoxify electrophilic compounds.A concurrent increase in the GSH level in group D isin agreement with the increased conjugational capacityof GST, because of increased availability of both bio-molecules as a result of combined treatment withV + BC. Group B and C rats, which, received V andBC alone, respectively, showed a decreased level ofGSH, but an additive effect was found in V + BCtreated rats (Group D) (Fig. 3a). This may be as aresult of the interaction of these compounds with reac-tive oxygen species generated after metabolism ofDEN, leading to the formation of additional pyridinenucleotides and thereby abating the growth and spread
of neoplastic nodules. In most cases, a strong correla-tion is observed between an increase in GST activity,metabolic inactivation and resistance to several anti-cancer agents.44 In the present investigation, theauthors have found increased activity of the GSTenzyme in DEN control rats (group A) and subsequentdepletion of the enzyme activity in all treatmentgroups. But an additive inhibitory effect was found ingroup D rats that received V + BC for the entire study(Fig. 3b). In tumors where increased GST activity hasbeen implicated in drug resistance, the use of combi-nation chemotherapy with GST inhibitors has been asuccessful approach to achieving apoptosis.45
Based upon the present study and those reported pre-viously from the authors’ laboratory10 as well as anotherlaboratory,8 it has been confirmed that V when admin-istered alone at low concentration acts as an anticarci-nogenic agent in experimental animals. The mechanismof the anticarcinogenic effect of V may involve inhibitionof metabolic activation of the procarcinogen, leading toreduced generation and/or binding of the ultimate car-cinogen to DNA.46 The possible effects of BC on initi-ation have been supported by the demonstration of theirantigenotoxic properties, both in vivo and in vitro,47,48
but little is known about how they act. Recently,carotenoid-rich extracts of carrots, tomato paste ororange juice have been shown to reduce the number ofliver preneoplastic foci induced by AFB1 in rats, whenadministered together with the carcinogen.49 Althoughthe primary mechanism of action of BC remains elu-sive, its chemopreventive effect might be mediatedthrough its effect on epithelial cell proliferation anddifferentiation.50
So, by combining these two chemopreventive agentsat the apparently non-toxic levels of the doses underconsideration, the authors have shown that they signif-icantly inhibit rat liver carcinogenesis by influencing thedifferent antioxidant enzymes and stabilization of theaerobic metabolism. The combined efficacy of V + BCresults in the maintenance of the hepatic architecturewith protection against the expression of GST-P-positive foci as induced by a potent carcinogen (DEN)in the presence of PB administration. Regardless of themechanism, the results of the present study provide evi-dence that combination therapy with V + BC has a bet-ter protective effect than a single entity in the inhibitionof rat liver carcinogenesis induced by DEN. Theobserved biological response of V + BC in combinationmay also have a broader implication as a chemopreven-tive agent.
ACKNOWLEDGMENTS
The authors like to thank Professor M Manson, MRCToxicology Unit, University of Leicester, UK, for pro-viding the antiserum for GST-P immunostaining. MBChattopadhyay is indebted to the Indian Council ofMedical Research, Government of India (Grant no. 45/2/2001/BMS/TRM) for the financial support to carryout this work. We are also grateful to Dr PK Sen, SeniorReader, Department of Mathematics, Jadavpur Univer-sity, Calcutta, India, for his excellent guidance in sta-

692 MB Chattopadhyay
tistical interpretation of the data. The authors aregrateful to Dr Amitabha Chatterjee, Calcutta UniversityCollege of Medicine, Calcutta, India, for his help ininterpretation of the histopathological study. Theauthors thank Dr Ranjan Basak and Dr Barun KantiSaha of Jadavpur University for technical advice andsupport.
REFERENCES
1 Farber E. The sequential analysis of liver cancer induc-tion. Biochim. Biophys. Acta 1980; 605: 149–66.
2 Elizabeth M, Fauziah O, Asmah R, Maznah I, Asmah Y,Siti M. Changes in morphology of induced liver cancertreated with Strobilanthes crispus. Anal. Microsc. 2000; 1:23–32.
3 Singer B, Grunberger D. Molecular Biology and Mutagensand Carcinogens. New York, NY: Plenium Press, 1984;431–50.
4 Cozzi R, Ricordy R, Agilitti T, Gatta V, Perticono P,Desalvia R. Ascorbic acid and beta-carotene as modula-tors of oxidative damage. Carcinogenesis 1997; 18: 223–8.
5 Boone CW, Kellof GJ, Malone WE. Identification of can-didate cancer chemopreventive agents and their evalua-tion in animal models and human clinical trials: a review.Cancer Res. 1990; 50: 2–9.
6 Bishayee A, Chatterjee M. Inhibition of altered liver cellfoci and persistent nodule growth by vanadium duringdiethylnitrosamine-induced hepatocarcinogenesis in rats.Anticancer Res. 1995a; 15: 455–62.
7 Chakraborty A, Selvaraj S. Differential modulation ofxenobiotic metabolizing enzymes by vanadium duringDEN-induced hepatocarcinogenesis in Sprague-Dawleyrats. Neoplasma 2000; 47: 81–9.
8 Djordjevic C. Antitumor activity of vanadium com-pounds. Met. Ions Biol. Syst. 1995; 31: 595–616.
9 Sakurai H, Tamura H, Okatawi K. Mechanism for a newantitumor vanadium complex: hydroxyl radical dependentDNA cleavage by 1,10-phenanthroline-vandyl complexesin the presence of hydrogen peroxide. Biochem. Biophys.Res. Commun. 1995; 206: 113–37.
10 Bishayee A, Chatterjee M. Inhibitory effect of vanadiumon rat liver carcinogenesis initiated with diethyl nitro-samine and promoted by Phenobarbital. Br. J. Cancer1995b; 71: 1214–20.
11 Krinsky NJ. Carotenoids and cancer in animal models. J.Nutr. 1989; 119: 123–6.
12 Sarkar A, Mukherjee B, Chatterjee M. Inhibition of3¢-methyl-4-dimethyl-aminobenzene-induced hepatic car-cinogenesis in rats by dietary beta-carotene: changes inhepatic antioxidant defense enzyme levels. Int. J. Cancer1995; 61: 799–805.
13 Moreno FS, Rizzi MBSI, Dagli MLZ, Penteado MVC.Inhibitory effect of beta-carotene on preneoplastic lesionsinduced in Wister rats by the resistant hepatocyte model.Carcinogenesis 1991; 12: 1817–22.
14 Tsuda H, Uehara N, Iwahori Y et al. Chemopreventiveeffects of beta-carotene, alpha-tocopherol and five natu-rally occurring antioxidants on initiation of hepatocar-cinogenesis by 2-amino-3-methylimidazo [4,5-f] quilinein the rat. Jpn. J. Cancer Res. 1994; 85: 1214–19.
15 Collins AR. Carotenoids and genomic stability. Mutat.Res. 2001; 475: 21–8.
16 Alpha-tocopherol, b-carotene cancer prevention StudyGroup. The effect of vitamin E and beta-carotene on theincidence of lung cancer and other cancers in male smok-ers. N. Engl. J. Med. 1994; 330: 1029–35.
17 Omenn GS, Goodman GE, Thornquist MD et al. Effectsof a combination of beta-carotene and vitamin A on lungcancer and cardiovascular disease. N. Engl. J. Med. 1996;334: 1150–5.
18 Collins AR. Carotenoids and genomic stability. Mutat.Res. 2001; 475: 21–8.
19 Bendich A. The safety of beta-carotene. Nutr. Cancer1988; 11: 207–14.
20 Hennekens CH, Mayrent SL, Willet W. Vitamin A, car-otenoids and retinoids. Cancer 1986; 58: 1837–41.
21 Hathcock JN, Hatton DJ, Jenkins MY, McDonald JT,Sunderesan PR, Wilkening VL. Evaluation of vitamin Atoxicity. Am. J. Clin. Nutr. 1990; 52: 183–202.
22 Ito N, Tsuda H, Tatematsu M, Hasegawa R, Tsuda H.Medium-term bioassay system for detection of carcino-gens and modifiers of hepatocarcinogenesis utilizing theGST-positive liver cells focus as an endpoint marker. Tox-icol. Pathol. 1989; 17: 630–41.
23 Hendrich S, Cambell H, Pitot H. Quantitative stereolog-ical evaluation of four histochemical markers of alteredfoci in multistage hepatocarcinogenesis in the rat. Carcino-genesis 1987; 8: 1245–50.
24 Tatamatsu M, Mera Y, Ito N, Satoh K, Sato K. Relativemerits of immunohistochemical demonstrations of pla-cental, A, B and C forms of glutathione S transferase andhistochemical demonstration of g-glutamyltransferase asmarkers of altered foci during liver carcinogenesis in rats.Carcinogenesis 1985; 6: 1621–6.
25 Pitot H. Altered hepatic foci: their role in murine hepato-carcinogenesis. Annu. Rev. Pharmacol. Toxicol. 1990; 30:465–500.
26 Sato K. Glutathione transferase as markers of preneo-plasia and neoplasia. Adv. Cancer Res. 1990; 52: 205–55.
27 Chesseaud LF. The role of glutathione and glutathioneS-transferase in the metabolism of chemical carcinogensand other electrophilic agents. Adv. Cancer Res. 1979; 29:175–274.
28 Wattenberg LW. Inhibition of carcinogenesis by minordietary constituents. Cancer Res. 1992; 52: 2085–91.
29 Nijhoff WA, Groen GM, Peters WHM. Induction of rathepatic and intestinal glutathione S-transferase and glu-tathione by dietary naturally occurring anticarcinogens.Int. J. Oncol. 1993; 3: 1131–9.
30 Bhattacharya S. The genesis, status and future of envi-ronmental toxicology. Indian J. Exp. Biol. 1996; 34:1131–5.
31 Sarkar A, Basak R, Bishayee A, Basak J, Chatterjee M. b-carotene inhibits rat liver chromosomal aberrations andDNA chain break after a single injection of diethylni-trosmine. Br. J. Cancer 1997; 76: 855–61.
32 Bannasch P. Cytology and cytogenetics of neoplastic(hyperplastic) hepatic nodules. Cancer Res. 1976; 36:2555–62.
33 Stewart HL, Williams GM, Keysser CH, Lombard LS,Montail RJ. Histological typing of liver tumors of the rat.J. Natl Cancer Inst. 1980; 64: 117–207.

Vanadium and beta-carotene in liver carcinogenesis 693
34 Ellman GE. Tissue sulfhydryl groups. Arch. Biochem. Bio-phys. 1959; 82: 70–7.
35 Habig WH, Pabst MJ, Jacoby WB. Glutathione S-transferase, the first enzymatic step in mercapturic acidformation. J. Biol. Chem. 1974; 249: 7130–9.
36 Lowry OH, Rosebrough NJ, Farr AL, Randall RL. Pro-tein measurement with the folin phenol reagent. J. Biol.Chem. 1951; 193: 265–75.
37 Kaku T, Ekem J, Lindayen C, Bailey D, van Nostrand A,Farber E. Comparison of formalin and acetone fixationfor immunohistochemical detection of carcinoembryonicantigen (CEA) and keratin. Am. J. Clin. Pathol. 1983; 80:806–15.
38 Bondi A, Chieregatti G, Eusebi U, Fulcheri E, BussolatiG. The use of b-galactosidase stain as a tracer in immu-nocytochemistry. Histochemistry 1982; 76: 153–8.
39 Warburg OA. On the origin of cancer cells. Science 1956;123: 309–15.
40 Boik JC. Cancer and Natural Medicine: a Textbook of BasicScience Research and Clinical Research. Oregon: OregonMedical Press, 1996.
41 Satoh K, Kitahara A, Soma Y, Inaba Y, Hatayama I, SatoK. Purification, induction and distribution of placentalglutathione transferase: a new marker enzyme for preneo-plastic cells in the rat chemical hepatocarcinogenesis.Proc. Natl Acad. Sci. 1985; 82: 3964–8.
42 Moreno FS, Ton SW, Naves MMV et al. Inhibitory effectsof beta-carotene and vitamin A during the progressionphase of hepatocarcinogenesis involve of cell proliferation
but not alterations in DNA methylation. Nutr. Cancer2002; 44: 80–9.
43 Uhlig S, Wendel A. The physiological consequences ofglutathione variations. Life Sci. 1992; 51: 1083–94.
44 Kulkarni AA, Kulkarni AP. Retionoids inhibit mamma-lian glutathione transferase. Cancer Lett. 1995; 91: 335–84.
45 Mannervik B, Danielson UH. Glutathione transferases –structure and catalytic activity. CRC Crit. Rev. Biochem.1988; 23: 283–7.
46 Chatterjee M, Bishayee A. Vanadium – a new tool in can-cer chemoprevention: Recent progress and future oppor-tunities. In: Nriagu JO, ed. Vanadium in the Environment.Part 2: Health Effects. New York: John Wiley, 1998; 347–89.
47 Krinsky NI. Effects of carotenoids on cellular and animalsystems. Am. J. Clin. Nutr. 1991; 53: 238–46S.
48 Krinsky NI. Micronutrients and their influence onmutagenicity and malignant transformation. Ann. NYAcad. Sci. 1993; 686: 229–42.
49 He RK, Gascon-Barré M. Influence of the Vitamin Dstatus on the early hepatic response to carcinogenexposure in rats. J. Pharmacol. Exp. Ther. 1997; 281:464–9.
50 Muto Y, Fuji J, Moriwaki H, Kawaguchi T, Noda T.Growth retardation in human cervical dysplasia-derivedcell lines by beta-carotene through down-regulation ofepidermal growth factor receptor. Am. J. Clin. Nutr.1995; 62: 1535–40S.




















