
CNTF-mediated protection of photoreceptors requires initial activation of the cytokine receptor...
18
CNTF-mediated protection of photoreceptors requires initial activation of the cytokine receptor gp130 in Müller glial cells Kun Do Rhee a , Steven Nusinowitz a , Kevin Chao a , Fei Yu a , Dean Bok a,b , and Xian-Jie Yang a,c,1 a Jules Stein Eye Institute, Department of Ophthalmology, b Department of Neurobiology, and c Molecular Biology Institute, University of California, Los Angeles, CA 90095 Edited by Constance L. Cepko, Harvard Medical School/Howard Hughes Medical Institute, Boston, MA, and approved October 11, 2013 (received for review February 28, 2013) Ciliary neurotrophic factor (CNTF) acts as a potent neuroprotective agent in multiple retinal degeneration animal models. Recently, CNTF has been evaluated in clinical trials for the inherited de- generative disease retinitis pigmentosa (RP) and for dry age- related macular degeneration (AMD). Despite its potential as a broad-spectrum therapeutic treatment for blinding diseases, the target cells of exogenous CNTF and its mechanism of action remain poorly understood. We have shown previously that constitutive expression of CNTF prevents photoreceptor death but alters the retinal transcriptome and suppresses visual function. Here, we use a lentivirus to deliver the same secreted human CNTF used in clinical trials to a mouse model of RP. We found that low levels of CNTF halt photoreceptor death, improve photoreceptor morphol- ogy, and correct opsin mislocalization. However, we did not detect corresponding improvement of retinal function as measured by the electroretinogram. Disruption of the cytokine receptor gp130 gene in Müller glia reduces CNTF-dependent photoreceptor sur- vival and prevents phosphorylation of STAT3 and ERK in Müller glia and the rest of the retina. Targeted deletion of gp130 in rods also demolishes neuroprotection by CNTF and prevents further activation of Müller glia. Moreover, CNTF elevates the expression of LIF and endothelin 2, thus positively promoting Müller and pho- toreceptor interactions. We propose that exogenous CNTF initially targets Müller glia, and subsequently induces cytokines acting through gp130 in photoreceptors to promote neuronal survival. These results elucidate a cellular mechanism for exogenous CNTF-triggered neuroprotection and provide insight into the com- plex cellular responses induced by CNTF in diseased retinas. C iliary neurotrophic factor (CNTF) belongs to a small sub- family of cytokines and has long been recognized as a po- tent neuroprotective molecule in the vertebrate retina (1). Enhancement of photoreceptor survival by CNTF has been demonstrated in multiple animal models of retinal degeneration, ranging from zebrafish to canine (2). Interestingly, CNTF is ef- fective in rescuing retinal degeneration due to various causes, including mutations in genes expressed by photoreceptors or the retinal pigment epithelium (RPE), as well as those induced by strong light, neurotoxins, or antibodies. In addition, CNTF prolongs the survival of retinal ganglion cells (3–5) and promotes axonal growth in optic nerve crush or transection models (6–8). Furthermore, CNTF may affect the physiology and survival of RPE cells (9, 10), which are critical in normal vision and retinal degenerative diseases. Based on its significant and broad neu- roprotective effects in damaged retinas, a secreted form of hu- man CNTF delivered from an encapsulated cell device has been tested in clinical trials and approved by the Food and Drug Administration (FDA) to treat retinitis pigmentosa (RP) and geographic atrophy (GA), a subset of age-related macular de- generation (AMD) (11–14). However, despite its clinical signif- icance, the mechanisms of CNTF-elicited neuroprotection in diverse retinal pathological conditions remain unclear. CNTF interacts with a tripartite receptor complex consisting of two single-pass membrane spanning receptor subunits (gp130 and LIFRβ) and an additional ligand-specific α receptor (CNTFRα) (15). Binding of CNTF to its receptor complex activates the Jak-STAT (16), the ERK (17), and the PI3K (18) pathways. In developing and mature rodent retinas, the CNTF family of cyto- kines stimulates Jak-STAT and ERK signaling (19–23). Previous studies have revealed that the dosage and duration of CNTF treatment critically affect the function of rescued neurons. Ro- dent retinal degeneration mutants treated with recombinant adeno-associated virus (rAAV) constitutively expressing CNTF show suppression of retinal function as measured by electro- retinogram (ERG) despite the robust preservation of photo- receptors (24–26). Exposure of normal rabbit retinas with high- dose CNTF-secreting capsules similarly led to diminished cone ERGs (27). Molecular and cellular analyses have demonstrated that persistent CNTF-induced signaling alters the retinal tran- scriptome, including reduced expression of genes involved in phototransduction and severe suppression of cone opsins (25, 28). However, single intravitreal injection of CNTF results in transient but reversible shortening of the photoreceptor outer segments and perturbation of phototransduction genes (29). To understand the mechanism of CNTF-triggered neuropro- tection, we used a lentivirus to deliver human CNTF (hCNTF) to a transgenic mouse model carrying the rds/peripherin P216L mutation (30), which causes dominant RP in humans. We show that low levels of hCNTF not only prolong cell survival but also Significance The cytokine CNTF has been approved by the FDA as a neuro- protective treatment for major retinal degenerative diseases. However, the mechanism of CNTF-triggered protection and CNTF-responsive cells in the retina remains unknown. Using molecular genetic analyses in a retinal degeneration mouse model, we identify the Müller glial cell as the direct initial target of exogenous CNTF signals. We provide evidence that CNTF signals stimulate a Müller glia and photoreceptor in- tercellular signaling loop involved in neuronal survival. None- theless, despite the marked improvement in photoreceptor morphology and survival, CNTF treatment does not result in a corresponding improvement of visual function. These results provide insight into the mechanism of CNTF action in the dis- eased retinas relevant to its clinical applications. Author contributions: K.D.R. and X.-J.Y. designed research; K.D.R., S.N., K.C., and D.B. performed research; K.D.R., S.N., F.Y., D.B., and X.-J.Y. analyzed data; and K.D.R. and X.-J.Y. wrote the paper. The authors declare no conflict of interest. This article is a PNAS Direct Submission. 1 To whom correspondence should be addressed. E-mail: [email protected]. This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10. 1073/pnas.1303604110/-/DCSupplemental. E4520–E4529 | PNAS | Published online November 4, 2013 www.pnas.org/cgi/doi/10.1073/pnas.1303604110
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of CNTF-mediated protection of photoreceptors requires initial activation of the cytokine receptor...

CNTF-mediated protection of photoreceptors requiresinitial activation of the cytokine receptor gp130 inMüller glial cellsKun Do Rheea, Steven Nusinowitza, Kevin Chaoa, Fei Yua, Dean Boka,b, and Xian-Jie Yanga,c,1
aJules Stein Eye Institute, Department of Ophthalmology, bDepartment of Neurobiology, and cMolecular Biology Institute, University of California, LosAngeles, CA 90095
Edited by Constance L. Cepko, Harvard Medical School/Howard Hughes Medical Institute, Boston, MA, and approved October 11, 2013 (received for reviewFebruary 28, 2013)
Ciliary neurotrophic factor (CNTF) acts as a potent neuroprotectiveagent in multiple retinal degeneration animal models. Recently,CNTF has been evaluated in clinical trials for the inherited de-generative disease retinitis pigmentosa (RP) and for dry age-related macular degeneration (AMD). Despite its potential as abroad-spectrum therapeutic treatment for blinding diseases, thetarget cells of exogenous CNTF and its mechanism of action remainpoorly understood. We have shown previously that constitutiveexpression of CNTF prevents photoreceptor death but alters theretinal transcriptome and suppresses visual function. Here, we usea lentivirus to deliver the same secreted human CNTF used inclinical trials to a mouse model of RP. We found that low levels ofCNTF halt photoreceptor death, improve photoreceptor morphol-ogy, and correct opsin mislocalization. However, we did not detectcorresponding improvement of retinal function as measured bythe electroretinogram. Disruption of the cytokine receptor gp130gene in Müller glia reduces CNTF-dependent photoreceptor sur-vival and prevents phosphorylation of STAT3 and ERK in Müllerglia and the rest of the retina. Targeted deletion of gp130 in rodsalso demolishes neuroprotection by CNTF and prevents furtheractivation of Müller glia. Moreover, CNTF elevates the expressionof LIF and endothelin 2, thus positively promoting Müller and pho-toreceptor interactions. We propose that exogenous CNTF initiallytargets Müller glia, and subsequently induces cytokines actingthrough gp130 in photoreceptors to promote neuronal survival.These results elucidate a cellular mechanism for exogenousCNTF-triggered neuroprotection and provide insight into the com-plex cellular responses induced by CNTF in diseased retinas.
Ciliary neurotrophic factor (CNTF) belongs to a small sub-family of cytokines and has long been recognized as a po-
tent neuroprotective molecule in the vertebrate retina (1).Enhancement of photoreceptor survival by CNTF has beendemonstrated in multiple animal models of retinal degeneration,ranging from zebrafish to canine (2). Interestingly, CNTF is ef-fective in rescuing retinal degeneration due to various causes,including mutations in genes expressed by photoreceptors orthe retinal pigment epithelium (RPE), as well as those inducedby strong light, neurotoxins, or antibodies. In addition, CNTFprolongs the survival of retinal ganglion cells (3–5) and promotesaxonal growth in optic nerve crush or transection models (6–8).Furthermore, CNTF may affect the physiology and survival ofRPE cells (9, 10), which are critical in normal vision and retinaldegenerative diseases. Based on its significant and broad neu-roprotective effects in damaged retinas, a secreted form of hu-man CNTF delivered from an encapsulated cell device has beentested in clinical trials and approved by the Food and DrugAdministration (FDA) to treat retinitis pigmentosa (RP) andgeographic atrophy (GA), a subset of age-related macular de-generation (AMD) (11–14). However, despite its clinical signif-icance, the mechanisms of CNTF-elicited neuroprotection indiverse retinal pathological conditions remain unclear.
CNTF interacts with a tripartite receptor complex consistingof two single-pass membrane spanning receptor subunits (gp130and LIFRβ) and an additional ligand-specific α receptor (CNTFRα)(15). Binding of CNTF to its receptor complex activates theJak-STAT (16), the ERK (17), and the PI3K (18) pathways. Indeveloping and mature rodent retinas, the CNTF family of cyto-kines stimulates Jak-STAT and ERK signaling (19–23). Previousstudies have revealed that the dosage and duration of CNTFtreatment critically affect the function of rescued neurons. Ro-dent retinal degeneration mutants treated with recombinantadeno-associated virus (rAAV) constitutively expressing CNTFshow suppression of retinal function as measured by electro-retinogram (ERG) despite the robust preservation of photo-receptors (24–26). Exposure of normal rabbit retinas with high-dose CNTF-secreting capsules similarly led to diminished coneERGs (27). Molecular and cellular analyses have demonstratedthat persistent CNTF-induced signaling alters the retinal tran-scriptome, including reduced expression of genes involved inphototransduction and severe suppression of cone opsins (25,28). However, single intravitreal injection of CNTF results intransient but reversible shortening of the photoreceptor outersegments and perturbation of phototransduction genes (29).To understand the mechanism of CNTF-triggered neuropro-
tection, we used a lentivirus to deliver human CNTF (hCNTF)to a transgenic mouse model carrying the rds/peripherin P216Lmutation (30), which causes dominant RP in humans. We showthat low levels of hCNTF not only prolong cell survival but also
Significance
The cytokine CNTF has been approved by the FDA as a neuro-protective treatment for major retinal degenerative diseases.However, the mechanism of CNTF-triggered protection andCNTF-responsive cells in the retina remains unknown. Usingmolecular genetic analyses in a retinal degeneration mousemodel, we identify the Müller glial cell as the direct initialtarget of exogenous CNTF signals. We provide evidence thatCNTF signals stimulate a Müller glia and photoreceptor in-tercellular signaling loop involved in neuronal survival. None-theless, despite the marked improvement in photoreceptormorphology and survival, CNTF treatment does not result ina corresponding improvement of visual function. These resultsprovide insight into the mechanism of CNTF action in the dis-eased retinas relevant to its clinical applications.
Author contributions: K.D.R. and X.-J.Y. designed research; K.D.R., S.N., K.C., and D.B.performed research; K.D.R., S.N., F.Y., D.B., and X.-J.Y. analyzed data; and K.D.R. andX.-J.Y. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.1To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1303604110/-/DCSupplemental.
E4520–E4529 | PNAS | Published online November 4, 2013 www.pnas.org/cgi/doi/10.1073/pnas.1303604110

improve the morphology of photoreceptors harboring the rds/P216L mutation. In addition, using conditional gene deletions,we identified the target cell type for the exogenous hCNTF andthe cell-specific requirements for the cytokine receptor gp130.We provide evidence that exogenous CNTF initially signalsthrough gp130 in Müller glia, which subsequently triggers theproduction of neuroprotective signal(s) that require gp130 in rodphotoreceptors for prevention of cell death.
ResultsLentiviral-Mediated hCNTF Expression from RPE. We constructeda CNTF-expressing lentiviral vector (LV-hCNTF) that carriesthe cytomegalovirus (CMV) immediate-early promoter followedby a human CNTF cDNA encoding a secreted and modifiedCNTF identical to that used in the clinical trials (12) (Fig. 1A).The control lentivirus (LV-IG) encodes only the internal ribo-somal entry site (IRES) and GFP downstream of the CMVpromoter (31) (Fig. 1A). Western blots confirmed that hCNTFwas indeed secreted into the culture media of HEK cells trans-fected with the LV-hCNTF plasmid (Fig. 1B). Immunolabelingshowed that LV-hCNTF-transduced cells coexpressed CNTFand GFP, in contrast to LV-IG–transduced cells, which ex-pressed only GFP (Fig. 1C). To measure the levels of CNTFproduction, ARPE19 cells infected by lentiviruses were analyzedby ELISA (Fig. 1D). Within a 24-h period, 170.0 ± 44.6 pg ofhCNTF was produced from 1.5 × 105 cells, with infection ratesat 90%.To examine lentiviral-mediated CNTF expression in vivo, ma-
ture eyes were injected subretinally with LV-IG or LV-hCNTF.Immunolabeling detected GFP expression as early as 72 h afterinfection (Fig. S1 and Table S1). By day 7 after injection, GFP
expression was localized to the RPE for both LV-IG– and LV-hCNTF–injected eyes, whereas low levels of widespread CNTFsignals were detected in LV-hCNTF–injected eyes only (Fig. 1 Eand F). ELISA analysis showed that 10 d after subretinal in-jection, LV-hCNTF–infected rds/P216L eyes contained 19.3 ±5.0 pg/eye of hCNTF at postnatal day 35 (P35) (Fig. 1D),whereas control virus LV-IG–infected eyes did not havedetectable hCNTF.
Effects of hCNTF on Photoreceptor Survival and Morphology. Wenext examined whether CNTF secreted from viral infected RPEcould protect photoreceptors from degeneration in rds/P216Lmutant mice (30). At P25, the rds/P216L retina displayed 20–30% fewer nuclei in the outer nuclear layer (ONL) with shorterinner and outer segments compared with the WT retina (Fig. 2 Aand B). By P52, only two to three rows of photoreceptor nucleiwith severely deteriorated outer segments remained in the mu-tant retinas (Fig. 2 C and G). Strikingly, rds/P216L retinas in-fected with LV-hCNTF at P25 maintained the thickness of theONL by P52 (Fig. 2 B and E). Morphometric quantificationconfirmed a pan-retinal rescue of the ONL in LV-hCNTF–injectedeyes compared with LV-IG–injected eyes (Fig. 2J). Moreover, bothhistological and transmission electron microscopy (TEM) anal-yses revealed a significant lengthening of both the inner andouter segments in retinas expressing hCNTF (Fig. 2 E, I, and K).Interestingly, injection of the control LV-IG virus also resultedin a detectable but statistically nonsignificant enhancement ofthe inner and outer segments (Fig. 2 C, D, G, H, and K). Theimprovement of photoreceptor morphology was also observed inmutant retinas analyzed at P108 (Fig. S2A). These results dem-onstrate that low levels of exogenous CNTF protect photo-
Fig. 1. Lentivirus-mediated expression of human CNTF in vitro and in vivo. (A) Schematics of lentiviruses encoding the CMV promoter followed by IRES-GFP(LV-IG) or human CNTF with a signal peptide (SP) and IRES-GFP (LV-hCNTF). (B) Western blot of cell extracts or culture media from HEK 293T cells transfectedwith LV-IG or LV-hCNTF probed for CNTF. The left lane was loaded with 100 ng of recombinant human CNTF. (C) Immunolabeling of 293T cells infected withLV-IG or LV-hCNTF for 48 h against GFP (green) and CNTF (red). (D) ELISA for hCNTF output from infected ARPE-19 cells (1.5 × 105 cells, between 48 and 72 hafter infection) and hCNTF levels in rds/P216L eyes 10 d after subretinal injection with LV-IG or LV-hCNTF at P25. (E and F) Immunolabeling for GFP and CNTFof rds/P216L eyes 5 d after LV-IG or LV-hCNTF injections at P27. [Scale bars, (E) 500 and (F) 50 μm.] gcl, ganglion cell layer; inl, inner nuclear layer; onl, outernuclear layer; rpe, retinal pigment epithelium.
Rhee et al. PNAS | Published online November 4, 2013 | E4521
NEU
ROSC
IENCE
PNASPL
US

receptors from degeneration and improve the morphology ofphotoreceptors carrying the rds/P216L mutation.
Effects of CNTF on Opsin Localization, Photoreceptor Gene Expression,and ERG. Previous studies showed that constitutive production ofhCNTF from a strong promoter encoded by rAAV suppressedcone opsin expression and the ERG (25, 28). Therefore, we ex-amined whether lentivirus-mediated hCNTF expression from RPEhad similar effects. Immunocytochemistry showed that rhodopsinwas mislocalized to the cell somata in the ONL of the LV-IG–
injected rds/P216L retinas (Fig. 3 B and C). Similarly, M-opsin wasmislocalized in the cone inner segments, somata, and the synapticlayer of the mutant retinas (Fig. 3 E and F). In contrast, LV-hCNTF–transduced rds/P216L mutant eyes not only retainedrhodopsin and M-opsin expression, but also showed a surprisingcorrection of opsin mislocalization found in the mutant (Fig. 3 DandG). Furthermore, labeling of whole mount retinas revealed anoverall increase in peanut agglutinin (PNA) signals and elongationof cone outer segments under the influence of hCNTF (Fig. S2 Band C ). However, we still observed a local reduction of M-opsinexpression in the temporal and ventral quadrant near the injectionsite (Fig. 3 I–P). Furthermore, real-time PCR analyses showed
that compared with LV-IG–injected controls, LV-hCNTF signifi-cantly reduced expression of cone-specific genes M-opsin, S-opsinand Gnat2, whereas no significant CNTF effects were found forrod-specific genes Rhodopsin and Gnat1, as well as genes ex-pressed by both rod and cone cells, including Crx, Nrl, and Nr2e3in the rds retinas (Fig. 3Q and Tables S2 and S3).We performed ERG assays to determine how LV-hCNTF
affected retinal function. Representative ERG waveforms ob-tained from noninjected WT and rds/P216L, as well as virus-injected, mice are shown in Fig. S3A. Under scotopic conditions,LV-IG– and LV-hCNTF–injected eyes showed dramatic andhighly significant reductions (up to 40–50%) in response ampli-tude compared with noninjected control eyes at both the level ofthe a- (Fig. 4 B and D and Tables S4 and S7) and b- (Fig. 4 A andD and Tables S5 and S7) waves. However, differences in a- or b-wave amplitudes between LV-IG– and LV-hCNTF–injected eyeswere not statistically significant, although there was a trend in thedirection of weaker signals in LV-hCNTF–injected eyes com-pared with the LV-IG–injected control eyes (Fig. 4 A, B, and Dand Tables S4, S5, and S7). Correspondingly, b-wave amplitudesunder photopic conditions were also reduced in LV-IG– and LV-hCNTF–injected eyes compared with noninjected rds/P216L eyes
Fig. 2. Lentiviral-mediated CNTF expression prevents degeneration and improves the morphology of mutant photoreceptors. (A–E) Histological staining bytoluidine blue shows degeneration caused by the rds/P216L mutation and the protective effect of LV-hCNTF. Viral infections were performed at P25 and eyesanalyzed by P52. (F–I) TEM analysis reveals improved photoreceptor morphology of LV-hCNTF infected rds/P216L retinas. (J) Quantification of the ONLthickness at P45 from the optic nerve head (ONH) to the ciliary margin for the WT and rds/P216L mutant injected with LV-IG or LV-hCNTF at P25 (n = 3 for all).(K) Quantification of IS/OS lengths at P45 for WT and rds/P216L mutant photoreceptors with or without lentivirus infections. Statistically significant samplepairs are indicated. ***P < 0.001. [Scale bars, (A–E) 50 and (F–I) 10 μm.] is, inner segment; olm, outer limiting membrane; onl, outer nuclear layer; os, outersegment; rpe, retinal pigment epithelium.
E4522 | www.pnas.org/cgi/doi/10.1073/pnas.1303604110 Rhee et al.

(Fig. 4C and Table S6). However, only at the brightest stimulusintensities did the difference between noninjected rds/P216Leyes and the LV-hCNTF–injected eyes reach statistical signifi-cance (Fig. 4D and Tables S4–S7), which may reflect the sup-pression by hCNTF on cone gene expression (Fig. 3Q and TableS3). Because injection of the control LV-IG virus or the vehicleresulted in similar ERG reductions (Fig. S3B and Tables S8–S10), these findings suggest that the reduced response ampli-tudes in injected eyes were largely due to the traumatic effects ofthe subretinal injection itself and not from the effects of CNTF.
Nonetheless, hCNTF treatment did not result in an improvedERG response as one might have expected from the markedimprovement of photoreceptor morphology.
Effects of gp130 Deletion in Müller Cells on CNTF-MediatedNeuroprotection and Signaling. Intravitreal injection of recombi-nant CNTF resulted in rapid STAT3 phosphorylation in Müllerglia in the rds/P216L retina (Fig. 5 A and B), suggesting thatMüller cells could be direct targets for exogenous CNTF. To testthis hypothesis, we ablated the prerequisite cytokine receptor
Fig. 3. Exogenous CNTF corrects mislocalization of opsins and affects photoreceptor gene expression. (A) A diagram shows where images were obtainedfrom an adult retinal section. (B–G) Confocal micrographs of immunolabeled P45 retinas for rhodopsin (B–D) or M-opsin (E–G) merged with DAPI images. Viralinjections for rds retinas were performed at P25. Arrows and arrowheads indicate mislocalized opsins in cell soma and synapses, respectively. (H) A diagram ofa flat mount adult retina showing the injection site. (I–P) Images of flat mount rds/P216L retinas injected at P25 with LV-IG or LV-hCNTF and immunolabeledfor M-opsin at P45. (Q) Quantification of photoreceptor transcripts by real-time PCR at P52 (n = 3 for all). For statistics, see Table S3. [Scale bars, (B–G) 20 and(I–P) 100 μm.] is, inner segment; onl, outer nuclear layer; os, outer segment; rpe, retinal pigment epithelium.
Rhee et al. PNAS | Published online November 4, 2013 | E4523
NEU
ROSC
IENCE
PNASPL
US

gp130 (32) in Müller cells using a GFAP-Cre driver mouse (33).When crossed with ROSA.EYFP reporter mice (34), YFP andthe Müller marker CycD3 were coexpressed (Fig. 5C). FACSanalysis showed that 4.3 ± 0.3% of retinal cells expressed glutaminesynthetase (GS), a Müller glial marker, and 59.3 ± 0.5% of theGS-positive cells also expressed YFP.We delivered LV-hCNTF subretinally into the rds/P216L mice
with the genotype of gp130fl/+;GFAP-Cre, which served as con-trols, or gp130 Müller conditional KO (cKO) mutant with thegenotypes of gp130fl/fl;GFAP-Cre (gp130 Müller cKO). ROSA.YFP indicated extensive Cre activity in Müller cells in bothgp130+/− and gp130 Müller cKO retinas (Fig. 5 D, E, H, and I).Morphometric quantification detected significant reduction ofONL thickness in gp130 Müller cKO retinas compared with thegp130 heterozygous controls (Fig. 5L). Immunolabeling for themajor CNTF downstream signaling events, the phosphorylationof STAT3 and ERK, detected broad distribution of phospho-STAT3 (pSTAT3) and phospho-ERK (pERK) in the LV-hCNTF–injected gp130 heterozygous retinas (Fig. 5 F and G).However, in gp130 Müller cKO retinas, CNTF-induced STAT3and ERK activation was severely dampened (Fig. 5 J and K).Furthermore, Western blots showed that recombinant CNTF-induced phosphorylation of STAT3 was attenuated in gp130Müller cKO retinas compared with gp130 heterozygous rds/P216Leyes (Fig. 5M). Together, these results indicate that a functionalgp130 receptor is required in Müller glia for CNTF-mediatedneuroprotection of photoreceptors, and deletion of gp130 inMüller glia prevents activation of CNTF-triggered signalingpathways in the rest of the retina.
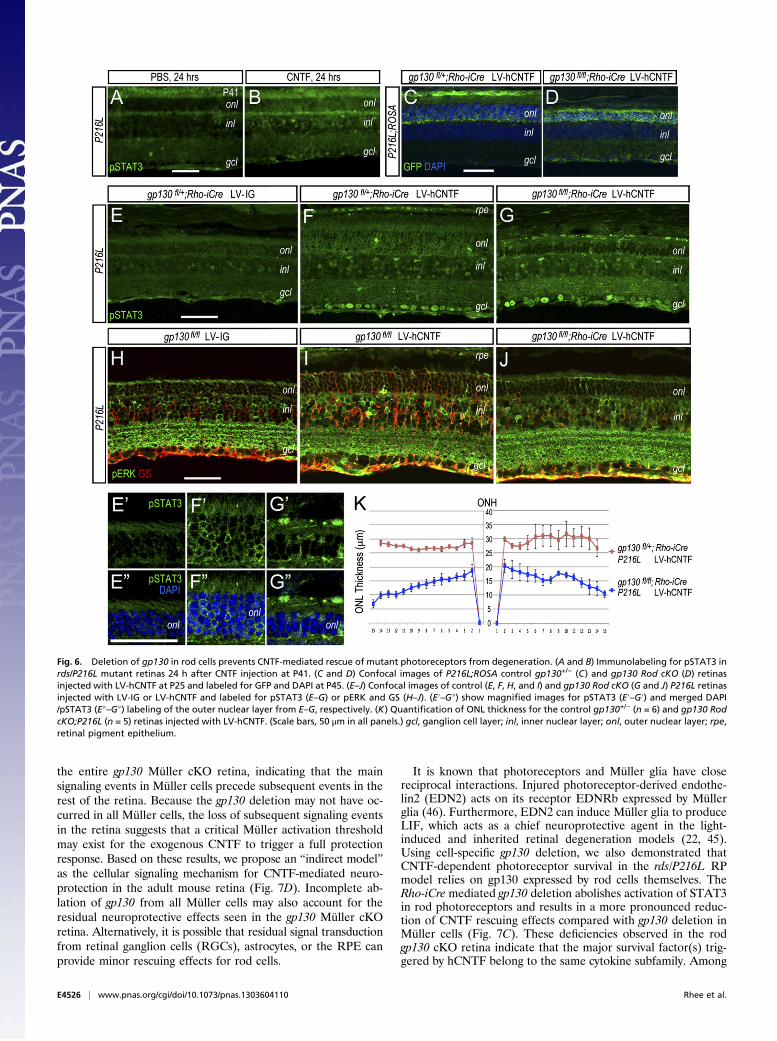
Requirement of gp130 in Rod Photoreceptors for Neuroprotection.Although CNTF initially up-regulated pSTAT3 in Müller glia,long-term CNTF expression via LV-hCNTF led to increasedpSTAT3 in the entire retina including the ONL (Fig. 5F). Inaddition, the rds/P216L retina exposed to CNTF for 24 h alsoshowed pSTAT3 distribution in the ONL (Fig. 6 A and B). Todetermine whether neuroprotection by CNTF required gp130function in rod photoreceptors, we analyzed CNTF effects in therds/P216L retina with gp130 deletion in rods using a transgenicline with the rhodopsin promoter driving Cre (Rho-iCre) (35). Inboth the gp130fl/+;Rho-iCre and gp130fl/fl;Rho-iCre (gp130 RodcKO) retinas injected with LV-hCNTF, the ROSA YFP reportershowed robust ONL expression (Fig. 6 C and D). However,CNTF-dependent photoreceptor rescue was severely diminishedin the gp130 Rod cKO rds/P216L retinas but remained intactin the gp130 heterozygous controls (Fig. 6 C, D, F, and G).Quantification further confirmed the significant reduction ofONL thickness in gp130 Rod cKO retinas compared with theheterozygous control mice injected with LV-hCNTF (Fig. 6K).Moreover, in contrast to the inner retina and RPE, the gp130Rod cKO retina did not show elevated pSTAT3 in ONL rod cellsin response to hCNTF (Fig. 6 E–G, E′–G′, and E″–G″). Theseresults indicate that the gp130 receptor is required in rod cells forSTAT3 activation and for CNTF-dependent neuronal survival.In the control rds retina, colabeling by pERK and GS detected
a further activation of Müller glia by hCNTF as revealed by morerobust Müller cell processes in the ONL, inner nuclear layer(INL), and in the ganglion cell layer (Fig. 6 H and I). In-terestingly, in gp130 Rod cKO retinas, the GS signals labelingMüller glia were substantially reduced in the ONL and INL (Fig.6 I and J), suggesting that gp130 is required in rod cells to sustain
Fig. 4. ERG analyses of lentiviral transduced rds/P216L mutant retinas. (A and B) Scotopic b-wave (A) and a-wave (B) amplitudes at P45 are shown fornoninjected rds/P216L eyes (n = 6), and rds/P216L eyes injected at P25 with LV-IG (n = 25) or LV-hCNTF (n = 37). (C) Photopic b-wave amplitudes recorded at P45are shown for noninjected rds/P216L eyes (n = 6), and rds/P216Lmutants injected at P25 with LV-IG (n = 17) or LV-hCNTF (n = 25). (D) Changes of ERG responsesrelative to noninjected rds/P216Lmutants at the highest scotopic or photopic stimuli. *P < 0.02; **P < 0.01; ***P < 0.001. Detailed statistics for ERG are shownin Tables S4–S7.
E4524 | www.pnas.org/cgi/doi/10.1073/pnas.1303604110 Rhee et al.

CNTF-triggered Müller activation. To examine whether CNTFaltered expression of signaling molecules in the retina, we per-formed real-time PCR analyses 27 d after viral injection. Asexpected, glial fibrillary acidic protein (GFAP) mRNA was ele-vated in the rds/P216L retinas (Fig. 7A and Table S11). Com-pared with age-matched WT retinas, the noninjected rds/P216Lretinas expressed higher levels of transcripts for CNTF and car-diotrophin-1 (CT-1), but without significant changes for leukemiainhibitory factor (LIF), cardiotrophin-like cytokine (CLC) andoncostatin M (OSM) (Fig. 7A and Table S11). In viral injectedrds retinas, LV-hCNTF transduction further increased ex-pression of endothelin 2 (EDN2) and brain-derived neuro-trophic factor (BDNF), compared with LV-IG injection (Fig. 7Aand Table S11). Interestingly, intravitreal injections of recombinantCNTF caused a rapid but transient increase of LIF transcripts,whereas a further increase of endogenous CNTF was detected24 h later (Fig. 7B and Table S12).
DiscussionMüller glial cells have the same developmental origin as retinalneurons and share many transcripts expressed by retinal pro-genitor cells (36, 37). In the mature retina, Müller glia play im-portant roles in maintaining retinal network homeostasis andintegrity and undergo reactive gliosis in response to stress anddamage (38, 39). In the zebrafish and chicken, Müller glia havebeen identified as a source of endogenous stem cells able torepair damaged retinas (40–42). It is also known that Müller gliain the mature retina produce multiple signaling molecules thatcould critically influence retinal neuronal activity, survival, andrepair (22, 43–45). In this study, we provide genetic evidence thatthe protection of photoreceptors by exogenous hCNTF relies oninitial signaling through the gp130 receptor in Müller glia. Con-ditional deletion of gp130 in Müller glia significantly decreasesCNTF-dependent protection (Fig. 7C). Importantly, CNTF-inducedSTAT3 and ERK activation is severely attenuated throughout
Fig. 5. Deletion of gp130 in Müller glia reduces neuroprotection and blocks activation of STAT and ERK by CNTF. (A and B) Immunolabeling for pSTAT3 inrds/P216Lmutant retinas 1 h after PBS (A) or CNTF (B) injection at P41. (C) Immunolabeling of ROSA-YFP retinas for GFP and Cyclin D3 at P81 in the presence orabsence of GFAP-Cre. (D–K) Confocal images of P216L; ROSA retinas injected with LV-hCNTF at P25 and labeled at P45 for DAPI (D and H), GFP (E and I),pSTAT3 (F and J), and pERK (G and K). (L) Quantification of ONL thickness for the control rds/P216L eyes injected with LV-hCNTF (n = 3) and gp130 Müller cKOeyes injected with LV-IG (n = 2) or LV-hCNTF (n = 3). (M) Western blot of retinal extracts from rds/P216L eyes that were gp130 Müller cKO or gp130+/− injectedwith PBS or CNTF for 1 h and probed for pSTAT3, STAT3, and γ Tubulin. Except for noninjected WT, extracts from two individual injected eyes are shown.[Scale bars, (A, C, and D), 50 μm.] gcl, ganglion cell layer; inl, inner nuclear layer; onl, outer nuclear layer; rpe, retinal pigment epithelium.
Rhee et al. PNAS | Published online November 4, 2013 | E4525
NEU
ROSC
IENCE
PNASPL
US
the entire gp130 Müller cKO retina, indicating that the mainsignaling events in Müller cells precede subsequent events in therest of the retina. Because the gp130 deletion may not have oc-curred in all Müller cells, the loss of subsequent signaling eventsin the retina suggests that a critical Müller activation thresholdmay exist for the exogenous CNTF to trigger a full protectionresponse. Based on these results, we propose an “indirect model”as the cellular signaling mechanism for CNTF-mediated neuro-protection in the adult mouse retina (Fig. 7D). Incomplete ab-lation of gp130 from all Müller cells may also account for theresidual neuroprotective effects seen in the gp130 Müller cKOretina. Alternatively, it is possible that residual signal transductionfrom retinal ganglion cells (RGCs), astrocytes, or the RPE canprovide minor rescuing effects for rod cells.
It is known that photoreceptors and Müller glia have closereciprocal interactions. Injured photoreceptor-derived endothe-lin2 (EDN2) acts on its receptor EDNRb expressed by Müllerglia (46). Furthermore, EDN2 can induce Müller glia to produceLIF, which acts as a chief neuroprotective agent in the light-induced and inherited retinal degeneration models (22, 45).Using cell-specific gp130 deletion, we also demonstrated thatCNTF-dependent photoreceptor survival in the rds/P216L RPmodel relies on gp130 expressed by rod cells themselves. TheRho-iCre mediated gp130 deletion abolishes activation of STAT3in rod photoreceptors and results in a more pronounced reduc-tion of CNTF rescuing effects compared with gp130 deletion inMüller cells (Fig. 7C). These deficiencies observed in the rodgp130 cKO retina indicate that the major survival factor(s) trig-gered by hCNTF belong to the same cytokine subfamily. Among
Fig. 6. Deletion of gp130 in rod cells prevents CNTF-mediated rescue of mutant photoreceptors from degeneration. (A and B) Immunolabeling for pSTAT3 inrds/P216L mutant retinas 24 h after CNTF injection at P41. (C and D) Confocal images of P216L;ROSA control gp130+/− (C) and gp130 Rod cKO (D) retinasinjected with LV-hCNTF at P25 and labeled for GFP and DAPI at P45. (E–J) Confocal images of control (E, F, H, and I) and gp130 Rod cKO (G and J) P216L retinasinjected with LV-IG or LV-hCNTF and labeled for pSTAT3 (E–G) or pERK and GS (H–J). (E′–G″) show magnified images for pSTAT3 (E′–G′) and merged DAPI/pSTAT3 (E″–G″) labeling of the outer nuclear layer from E–G, respectively. (K) Quantification of ONL thickness for the control gp130+/− (n = 6) and gp130 RodcKO;P216L (n = 5) retinas injected with LV-hCNTF. (Scale bars, 50 μm in all panels.) gcl, ganglion cell layer; inl, inner nuclear layer; onl, outer nuclear layer; rpe,retinal pigment epithelium.
E4526 | www.pnas.org/cgi/doi/10.1073/pnas.1303604110 Rhee et al.

the likely candidates is LIF, because it similarly requires thegp130 receptor complex for signaling. Our data show that CNTFcan cause a rapid and transient increase of LIF transcripts. Thetransient nature of LIF induction may reflect a negative feedbackregulation. Interestingly, we observed lower levels of CNTF-triggered Müller cell activation when gp130 was deleted from rodcells. This result suggests a positive feedback loop wherebygp130-mediated cell signaling events in rod cells furtherstrengthen the initial CNTF-dependent Müller cell activation.Our quantitative PCR (qPCR) results show that hCNTF signif-icantly enhances EDN2 mRNA expression, thus supporting theparticipation of exogenous hCNTF in this positive feed-forwardsignaling loop between Müller glia and photoreceptors (22, 45,46). In addition to promoting EDN2 expression, hCNTF alsostimulated expression of BDNF and possibly fibroblast growthfactor 2 (FGF2), both of which can act as trophic factors onmultiple cell types. Although exogenous CNTF does not directlysignal to rod cells initially, it is possible that endogenous CNTFsignals elevated in injured retinas (47–49) (Fig. 7 A and B) maysynergize with other induced factors to promote survival. It is worthnoting that in contrast to our findings here, it has been reportedthat light preconditioning-induced protection of photoreceptorsrequires gp130 in rod cells, but not in Müller glia (23, 50).Despite promoting neuronal survival, prolonged exposure to
high levels of CNTF can cause detrimental effects, includingsuppression of both cone opsin expression and ERG in the rds/
P216L model (25, 28). This phenomenon raises the questionwhether lower-dose CNTF treatments can avoid these harmfuleffects. Although it is difficult to directly compare the levels ofCNTF produced in the current study with the doses produced byencapsulated cells in the human trials, we detected hCNTF levelsat around 20 pg/eye in the LV-hCNTF–infected mouse eyes.Under these conditions, we observed a remarkable improvementin photoreceptor morphology represented by the lengthening ofboth inner and outer segments. This observation is reminiscentof the cone outer segment regeneration promoted by CNTF ina rat rhodopsin mutant (51). Furthermore, we also observeda correction of mislocalized rod and cone opsins in CNTF-treated retinas carrying the dominant P216L mutation, suggest-ing that CNTF is able to enhance the WT peripherin functionand to partially overcome the dominant mutant effect in thismouse model. Although the exact protective mechanism ofCNTF remains obscure, the benefits of low exogenous CNTF onpreventing further degeneration and promoting photoreceptorstructural recovery have clear clinical implications. In the light-damaged zebrafish retina, activated ERK but not STAT3 is re-quired for neuroprotection by exogenous CNTF (52). Futureinvestigations are necessary to determine the specific downstreamsignaling modalities required for low-level CNTF-dependent cellsurvival in the mammalian retina.Paradoxically, the remarkable improvement of photoreceptor
morphology and survival are not accompanied by any significant
Fig. 7. Summary of CNTF triggered protection in the rds/P216L model of photoreceptor degeneration. (A) Real-time PCR quantification of GFAP and growthfactor transcripts. WT and P216L are noninjected samples at P52; P216L, LV-IG and P216L, LV-hCNTF were injected with viruses at P25 and analyzed at P52. n =3 for all. For statistics, see Table S11. (B) Real-time PCR quantification of transcript levels for LIF, CNTF, and CT-1 in rds/P216L retinas. Injections of CNTF or PBSwere performed at P25, and RNAs were collected at 3 or 24 h after injections. n = 3 for all. For statistics, see Table S12. (C) Comparison of ONL thickness for WTand rds/P216L mutant retinas under the influence of hCNTF in gp130+/+, gp130 Müller or rod heterozygous, and gp130 Müller or Rod cKO genetic back-grounds. Significant P values (Student t test) are indicated. (D) A proposed indirect model of exogenous CNTF-induced neuroprotection of rod photoreceptorsin the rds/peripherin mutant retina. The secreted human CNTF derived from LV-transduced RPE initially activates signaling events in Müller glia through thegp130 cytokine receptor. This Muller glia response triggers the release of other signaling molecules, which subsequently activate gp130 expressed by rodphotoreceptors to enhance cell survival. Loss of gp130 in Müller glia prevents the activation of the entire signaling cascade, whereas removal of gp130 fromrod cells prevents rod photoreceptor rescue and diminishes sustained activation of Müller glia.
Rhee et al. PNAS | Published online November 4, 2013 | E4527
NEU
ROSC
IENCE
PNASPL
US

enhancement in retinal function as indicated by the ERG. Thereason for this finding is not understood but likely involvesCNTF-induced alteration of transcription, which ultimately leadsto the suppression of visual function. As we have shown pre-viously (28), exposure to high levels of CNTF down-regulates anumber of photoreceptor specific genes, particularly those as-sociated with phototransduction. Consistent with our findingsthat cone cells are more sensitive to CNTF signaling, we alsodetected mild suppression on cone opsin and Gnat2 expressioneven under low CNTF expression conditions, which may con-tribute to the mild suppression in photopic b-wave amplitudes.Intriguingly, in high-dose CNTF–treated eyes, the nuclear mor-phology of rod photoreceptors loses the stereotypical organiza-tion and resembles those of cone cells with less heterochromatinand more euchromatin (25), indicating altered chromatin orga-nization. Accumulating evidence indicates that mature retinalneurons are plastic and can undergo fate reprogramming. Forexample, ablating the Nrl gene in mature rod photoreceptorsresults in partial transformation of rod cells toward a cone cellfate, accompanied by altered rod nuclear morphology and en-hanced cell survival (53). It is possible that Jak-STAT signalingdirectly influences the chromatin structure of photoreceptor cellsin a cell autonomous fashion. However, because exogenousCNTF induces the release of a myriad of growth factors in theretina, it remains to be determined which signaling pathwaycontributes to the epigenetic changes observed in CNTF-treatedretinas. Furthermore, intravitreal injection of LIF causes a down-regulation of RPE65 protein, the key isomerase required in theRPE to restore the 11-cis-retinal chromophore, leading to aslower regeneration of rhodopsin (10). We also observed CNTF-induced STAT3 phosphorylation in RPE cells (Figs. 5 and 6).Because inhibition of RPE65 activity is known to prevent light-induced degeneration (54, 55), CNTF-like cytokine-inducedmodulation of the visual cycle could in part account for thesuppression of ERG and enhanced survival of photoreceptors.Therefore, current data indicate that CNTF treatment of matureretinas causes complex molecular and cellular responses inmultiple cell types, some of which may profoundly influenceneuronal characteristics and retinal function.Together, the results presented in this study demonstrate that
exogenous CNTF initially targets Müller glia to trigger a cascadeof signaling events leading to photoreceptor survival. Futureinvestigations focusing on the downstream signaling events in-duced by CNTF and the mechanisms involved in its beneficialand detrimental impacts on cellular physiology will enhance ourability to develop more effective neuroprotective therapies in themammalian retina.
Materials and MethodsAnimals. The rds/peripherin P216L transgenic mice (30) were maintained ona WT CD1 background. Mice with a floxed allele of gp130 (32) on a mixedbackground were crossed with transgenic mouse lines GFAP-Cre (33) or Rho-iCre75 (35) on the C57BL/6J background to generate gp130 deletions inMüller glia or rod photoreceptors, respectively. In some experiments, therds/P216L transgene and/or the ROSA.EYFP Cre reporter (34) were alsopresent in the gp130 retinal mutant background. Genotyping primers aresummarized in SI Materials and Methods. All animal procedures followed
National Institutes of Health guidelines and were approved by the AnimalResearch Committee at University of California–Los Angeles.
Lentiviral Vector Construction, Production, and Injection. LV-IG (31) encodesthe CMV promoter followed by the IRES-GFP cassette. LV-hCNTF was con-structed by inserting the cDNA of a secreted form of human CNTF withS166D and G167H substitutions (25) between the CMV promoter andIRES-GFP cassette. The production, titer determination, and subretinal in-jection of lentiviruses were performed as previously described (56). Lenti-viral stocks with titers of 1 × 107 CFU/mL were used for in vivo subretinalinjections, unless otherwise specified. For in vivo injections, 1 μg of recom-binant human CNTF (PeproTech) in 1 μL volume per eye was injectedintravitreously.
FACS, Immunohistochemistry, Histology, and TEM. FACS analysis was per-formed as described in ref. 57 and in SI Materials and Methods. For immu-nofluorescent labeling, cells or tissues were fixed with 4% (wt/vol)paraformaldehyde in PBS and processed as described previously (28). Wholemount retinas were incubated overnight with primary and secondary anti-bodies at 4 °C, followed by extensive washes. Fluorescent images werecaptured using a Spot II camera and Nikon E800 or Olympus FluoView 1000confocal microscopes. Antibodies used are summarized in Table S1. For 1-μmsemithin sections and TEM, eyes were fixed in 2% (wt/vol) formaldehyde and2.5% (wt/vol) glutaraldehyde in 0.1 M sodium phosphate buffer and pro-cessed as described previously (58).
Morphometrics. A minimum of three sections from each eye was used toacquire digital images of the retina. The ONL thickness as demarcated byDAPI-positive photoreceptor nuclei was determined at 15 equal spatialintervals from the optic nerve head to the ciliary margin. The lengths of innerand outer segments of photoreceptors were measured using 1-μm semithinsections stained with toluidine blue and ImagePro Plus software (MediaCybernetics).
Western Blot, ELISA, and Real-Time PCR.Western blots of cell lysates, medium,or retinal extracts were processed as described previously (21, 28). Theamounts of viral produced hCNTF in vitro and in vivo were determinedwith a human CNTF Quantikine ELISA kit (R&D Systems) as described in SIMaterials and Methods. Real-time PCRs were performed as described in ref.28 and SI Materials and Methods, and primers used for qPCR are shown inTable S2.
Electroretinogram. Detailed procedures for ERG are described in SI Materialsand Methods and ref. 59.
Statistics. One-way ANOVA was performed for the comparison with multiplesubgroups, and pairwise comparisons within ANOVA were conducted withTukey corrections if the ANOVAwas statistically significant. Values of P < 0.02were considered statistically significant. For the pairwise analysis presentedin Fig. 7C, the Student t test was used and values of P < 0.01 were consideredstatistically significant.
ACKNOWLEDGMENTS. We thank Jane Coffman, Shannan Eddington, SamirHabib, Tania Hioe, Paul Kim, Josephine Lee, Marcia Lloyd, Hitomi Suzuki,and Carrie Zhao for excellent technical support. We thank Dr. WilliamW. Hauswirth for the secretable human CNTF plasmid, Dr. Jason C.-K. Chenfor the Rho-iCre mouse, and Dr. Robert Molday for the rhodopsin antibody.The work was in part supported by grants from National Institutes of HealthEY019052, the Oppenheimer Family Foundation, and the Ernest G. HermanEndowed Chair in Ophthalmology (to X.-J.Y.), Macula Vision ResearchFoundation and Dolly Green Endowed Chair in Ophthalmology (to D.B.),and National Eye Institute Core Grant EY00331 for vision research at UCLA.
1. LaVail MM, et al. (1992) Multiple growth factors, cytokines, and neurotrophins rescue
photoreceptors from the damaging effects of constant light. Proc Natl Acad Sci USA
89(23):11249–11253.2. Wen R, Tao W, Li Y, Sieving PA (2012) CNTF and retina. Prog Retin Eye Res 31(2):
136–151.3. Meyer-Franke A, Kaplan MR, Pfrieger FW, Barres BA (1995) Characterization of the
signaling interactions that promote the survival and growth of developing retinal
ganglion cells in culture. Neuron 15(4):805–819.4. Takahata K, et al. (2003) Retinal neuronal death induced by intraocular administra-
tion of a nitric oxide donor and its rescue by neurotrophic factors in rats. Invest
Ophthalmol Vis Sci 44(4):1760–1766.
5. van Adel BA, Kostic C, Déglon N, Ball AK, Arsenijevic Y (2003) Delivery of ciliary
neurotrophic factor via lentiviral-mediated transfer protects axotomized retinal
ganglion cells for an extended period of time. Hum Gene Ther 14(2):103–115.6. Cui Q, Lu Q, So KF, Yip HK (1999) CNTF, not other trophic factors, promotes axonal
regeneration of axotomized retinal ganglion cells in adult hamsters. Invest Oph-
thalmol Vis Sci 40(3):760–766.7. Jo SA, Wang E, Benowitz LI (1999) Ciliary neurotrophic factor is an axogenesis factor
for retinal ganglion cells. Neuroscience 89(2):579–591.8. MacLaren RE, et al. (2006) CNTF gene transfer protects ganglion cells in rat reti-
nae undergoing focal injury and branch vessel occlusion. Exp Eye Res 83(5):
1118–1127.
E4528 | www.pnas.org/cgi/doi/10.1073/pnas.1303604110 Rhee et al.

9. Li R, Wen R, Banzon T, Maminishkis A, Miller SS (2011) CNTF mediates neurotrophicfactor secretion and fluid absorption in human retinal pigment epithelium. PLoS ONE6(9):e23148.
10. Chucair-Elliott AJ, et al. (2012) Leukemia inhibitory factor coordinates the down-regulation of the visual cycle in the retina and retinal-pigmented epithelium. J BiolChem 287(29):24092–24102.
11. Zhang K, et al. (2011) Ciliary neurotrophic factor delivered by encapsulated cell in-traocular implants for treatment of geographic atrophy in age-related macular de-generation. Proc Natl Acad Sci USA 108(15):6241–6245.
12. Sieving PA, et al. (2006) Ciliary neurotrophic factor (CNTF) for human retinal de-generation: Phase I trial of CNTF delivered by encapsulated cell intraocular implants.Proc Natl Acad Sci USA 103(10):3896–3901.
13. Talcott KE, et al. (2011) Longitudinal study of cone photoreceptors during retinaldegeneration and in response to ciliary neurotrophic factor treatment. Invest Oph-thalmol Vis Sci 52(5):2219–2226.
14. Birch DG, Weleber RG, Duncan JL, Jaffe GJ, Tao W; Ciliary Neurotrophic Factor Reti-nitis Pigmentosa Study Groups (2013) Randomized trial of ciliary neurotrophic factordelivered by encapsulated cell intraocular implants for retinitis pigmentosa. Am JOphthalmol 156(2):283–292, e1.
15. Ip NY (1998) The neurotrophins and neuropoietic cytokines: Two families of growthfactors acting on neural and hematopoietic cells. Ann N Y Acad Sci 840:97–106.
16. Bonni A, Frank DA, Schindler C, Greenberg ME (1993) Characterization of a pathwayfor ciliary neurotrophic factor signaling to the nucleus. Science 262(5139):1575–1579.
17. Boulton TG, Stahl N, Yancopoulos GD (1994) Ciliary neurotrophic factor/leukemiainhibitory factor/interleukin 6/oncostatin M family of cytokines induces tyrosinephosphorylation of a common set of proteins overlapping those induced by othercytokines and growth factors. J Biol Chem 269(15):11648–11655.
18. Oh H, et al. (1998) Activation of phosphatidylinositol 3-kinase through glycoprotein130 induces protein kinase B and p70 S6 kinase phosphorylation in cardiac myocytes.J Biol Chem 273(16):9703–9710.
19. Peterson WM, Wang Q, Tzekova R, Wiegand SJ (2000) Ciliary neurotrophic factorand stress stimuli activate the Jak-STAT pathway in retinal neurons and glia.J Neurosci 20(11):4081–4090.
20. Wahlin KJ, Campochiaro PA, Zack DJ, Adler R (2000) Neurotrophic factors cause ac-tivation of intracellular signaling pathways in Müller cells and other cells of the innerretina, but not photoreceptors. Invest Ophthalmol Vis Sci 41(3):927–936.
21. Rhee KD, Goureau O, Chen S, Yang XJ (2004) Cytokine-induced activation of signaltransducer and activator of transcription in photoreceptor precursors regulates roddifferentiation in the developing mouse retina. J Neurosci 24(44):9779–9788.
22. Joly S, Lange C, Thiersch M, Samardzija M, Grimm C (2008) Leukemia inhibitoryfactor extends the lifespan of injured photoreceptors in vivo. J Neurosci 28(51):13765–13774.
23. Ueki Y, Le YZ, Chollangi S, Muller W, Ash JD (2009) Preconditioning-induced pro-tection of photoreceptors requires activation of the signal-transducing receptorgp130 in photoreceptors. Proc Natl Acad Sci USA 106(50):21389–21394.
24. Schlichtenbrede FC, et al. (2003) Intraocular gene delivery of ciliary neurotrophicfactor results in significant loss of retinal function in normal mice and in the Prph2Rd2/Rd2 model of retinal degeneration. Gene Ther 10(6):523–527.
25. Bok D, et al. (2002) Effects of adeno-associated virus-vectored ciliary neurotrophicfactor on retinal structure and function in mice with a P216L rds/peripherin mutation.Exp Eye Res 74(6):719–735.
26. Liang FQ, et al. (2001) AAV-mediated delivery of ciliary neurotrophic factor prolongsphotoreceptor survival in the rhodopsin knockout mouse. Mol Ther 3(2):241–248.
27. Bush RA, et al. (2004) Encapsulated cell-based intraocular delivery of ciliary neuro-trophic factor in normal rabbit: Dose-dependent effects on ERG and retinal histology.Invest Ophthalmol Vis Sci 45(7):2420–2430.
28. Rhee KD, et al. (2007) Molecular and cellular alterations induced by sustained ex-pression of ciliary neurotrophic factor in a mouse model of retinitis pigmentosa. In-vest Ophthalmol Vis Sci 48(3):1389–1400.
29. Wen R, et al. (2006) Regulation of rod phototransduction machinery by ciliary neu-rotrophic factor. J Neurosci 26(52):13523–13530.
30. Kedzierski W, Lloyd M, Birch DG, Bok D, Travis GH (1997) Generation and analysis oftransgenic mice expressing P216L-substituted rds/peripherin in rod photoreceptors.Invest Ophthalmol Vis Sci 38(2):498–509.
31. Pfeifer A, Brandon EP, Kootstra N, Gage FH, Verma IM (2001) Delivery of the Crerecombinase by a self-deleting lentiviral vector: Efficient gene targeting in vivo. ProcNatl Acad Sci USA 98(20):11450–11455.
32. Betz UA, et al. (1998) Postnatally induced inactivation of gp130 in mice results inneurological, cardiac, hematopoietic, immunological, hepatic, and pulmonary de-fects. J Exp Med 188(10):1955–1965.
33. Zhuo L, et al. (2001) hGFAP-cre transgenic mice for manipulation of glial and neu-ronal function in vivo. Genesis 31(2):85–94.
34. Srinivas S, et al. (2001) Cre reporter strains produced by targeted insertion of EYFPand ECFP into the ROSA26 locus. BMC Dev Biol 1:4.
35. Li S, et al. (2005) Rhodopsin-iCre transgenic mouse line for Cre-mediated rod-specificgene targeting. Genesis 41(2):73–80.
36. Turner DL, Cepko CL (1987) A common progenitor for neurons and glia persists in ratretina late in development. Nature 328(6126):131–136.
37. Blackshaw S, et al. (2004) Genomic analysis of mouse retinal development. PLoS Biol2(9):E247.
38. Dyer MA, Cepko CL (2000) Control of Müller glial cell proliferation and activationfollowing retinal injury. Nat Neurosci 3(9):873–880.
39. Bringmann A, et al. (2009) Cellular signaling and factors involved in Müller cell gliosis:Neuroprotective and detrimental effects. Prog Retin Eye Res 28(6):423–451.
40. Fischer AJ, Reh TA (2001) Müller glia are a potential source of neural regeneration inthe postnatal chicken retina. Nat Neurosci 4(3):247–252.
41. Bernardos RL, Barthel LK, Meyers JR, Raymond PA (2007) Late-stage neuronal pro-genitors in the retina are radial Müller glia that function as retinal stem cells.J Neurosci 27(26):7028–7040.
42. Ramachandran R, Fausett BV, Goldman D (2010) Ascl1a regulates Müller glia de-differentiation and retinal regeneration through a Lin-28-dependent, let-7 microRNAsignalling pathway. Nat Cell Biol 12(11):1101–1107.
43. Yang XJ, Cepko CL (1996) Flk-1, a receptor for vascular endothelial growth factor(VEGF), is expressed by retinal progenitor cells. J Neurosci 16(19):6089–6099.
44. Wan J, Ramachandran R, Goldman D (2012) HB-EGF is necessary and sufficient forMüller glia dedifferentiation and retina regeneration. Dev Cell 22(2):334–347.
45. Bürgi S, Samardzija M, Grimm C (2009) Endogenous leukemia inhibitory factor pro-tects photoreceptor cells against light-induced degeneration. Mol Vis 15:1631–1637.
46. Rattner A, Nathans J (2005) The genomic response to retinal disease and injury: Evi-dence for endothelin signaling from photoreceptors to glia. J Neurosci 25(18):4540–4549.
47. Valter K, et al. (2005) Time course of neurotrophic factor upregulation and retinalprotection against light-induced damage after optic nerve section. Invest OphthalmolVis Sci 46(5):1748–1754.
48. Honjo M, et al. (2000) Expression of ciliary neurotrophic factor activated by retinalMüller cells in eyes with NMDA- and kainic acid-induced neuronal death. InvestOphthalmol Vis Sci 41(2):552–560.
49. Chun MH, et al. (2000) Upregulation of ciliary neurotrophic factor in reactive Müllercells in the rat retina following optic nerve transection. Brain Res 868(2):358–362.
50. Ueki Y, Chollangi S, Le YZ, Ash JD (2010) gp130 activation in Müller cells is not es-sential for photoreceptor protection from light damage. Adv Exp Med Biol 664:655–661.
51. Wen R, et al. (2012) Regeneration of cone outer segments induced by CNTF. Adv ExpMed Biol 723:93–99.
52. Kassen SC, et al. (2009) CNTF induces photoreceptor neuroprotection and Müller glialcell proliferation through two different signaling pathways in the adult zebrafishretina. Exp Eye Res 88(6):1051–1064.
53. Montana CL, et al. (2013) Reprogramming of adult rod photoreceptors preventsretinal degeneration. Proc Natl Acad Sci USA 110(5):1732–1737.
54. Mandal MN, et al. (2011) Alpha-phenyl-N-tert-butylnitrone (PBN) prevents light-induced degeneration of the retina by inhibiting RPE65 protein isomerohydrolaseactivity. J Biol Chem 286(37):32491–32501.
55. Li S, et al. (2013) Fatty acid transport protein 4 (FATP4) prevents light-induced de-generation of cone and rod photoreceptors by inhibiting RPE65 isomerase. J Neurosci33(7):3178–3189.
56. Hashimoto T, et al. (2007) Lentiviral gene replacement therapy of retinas in a mousemodel for Usher syndrome type 1B. Gene Ther 14(7):584–594.
57. Sakagami K, Gan L, Yang XJ (2009) Distinct effects of Hedgehog signaling on neu-ronal fate specification and cell cycle progression in the embryonic mouse retina.J Neurosci 29(21):6932–6944.
58. Jin M, et al. (2009) The role of interphotoreceptor retinoid-binding protein on thetranslocation of visual retinoids and function of cone photoreceptors. J Neurosci29(5):1486–1495.
59. Nusinowitz S, et al. (2003) Electroretinographic evidence for altered phototrans-duction gain and slowed recovery from photobleaches in albino mice with a MET450variant in RPE65. Exp Eye Res 77(5):627–638.
Rhee et al. PNAS | Published online November 4, 2013 | E4529
NEU
ROSC
IENCE
PNASPL
US

Supporting InformationRhee et al. 10.1073/pnas.1303604110SI Materials and MethodsMouse Genotyping. Mice were kept on a 12/12-h light/dark cycle.Genomic DNAs from the tail were used for genotyping using thefollowing PCR primers: Cre (forward: 5′GAC GAT GCA ACGAGT GAT GA, reverse: AGC ATT GCT GTC ACT TGG TC),gp130 (forward: 5′ACG TCA CAG AGC TGA G TG ATGCAC, reverse: 5′GGC TTT TCC TCT GGT TCT TG), iCre(forward: 5′ TCA GTG CCT GGA GTT GCG CTG TGG, re-verse: 5′ CTT AAA GGC CAG GGC CTG CTT GGC), rdsP216L
(forward: 5′CCT GGA GTT GCG CTG T, reverse: 5′GTC TTTTTC ATG AAG CAC C), and ROSA.EYFP (Primer 1: AAGACC GCG AAG AGT TTG TC, primer 2: GGA GCG GGAGAA ATG GAT ATG, primer 3: AAA GTC GCT CTG AGTTGT TAT).
FACS. To determine the percentage of Müller cells or rod cellsthat express Cre, mouse linesGFAP-Cre (1) or Rho-iCre75 (2) onthe C57BL/6J background were crossed to the ROSA.EYFP Crereporter (3). Dissociated adult retinal cells were immunolabeledfor GFP and glutamine synthetase (GS) and analyzed using anLSRII flow cytometer (BD Biosciences) as described (4). Flowcytometry data (n = 4) were analyzed with FlowJo (Tree Star).
ELISA. To determine human ciliary neurotrophic factor (hCNTF)produced by lentiviral vector (LV)-hCNTF in vitro, 1.5 ×105 APRE-19 cells were infected by 107 CFU/mL viruses for48 h. The media were changed at 48 h after viral infection, andcells were incubated for another 24 h before collection at 72 hafter infection for ELISA. To determine LV-hCNTF–mediatedCNTF production in vivo, protein extracts of individual P35eyecups, including retina, retinal pigment epithelium (RPE), andchoroid, were used for ELISA 10 d after lentiviral infection. Thecontrol lentivirus (LV-IG) infection results in nondetectablelevels of hCNTF either in vitro or in vivo as shown in Fig. 1D.
Real-Time PCRs.Total RNAs (n = 3) were isolated using RNA-Bee(TEL-Test) and converted to ssDNA using oligo dT and Su-perScript III reverse transcriptase (Invitrogen). PCR primer se-quences and references are summarized in Table S2. Real-timePCRs were carried out in duplicates using 1 ng cDNA per 20-μLreaction with SYBR Green PCR Master Mix and the AppliedBioscience StepOne real-time PCR system. Real-time PCR datawere normalized with GAPDH transcript levels.
Electroretinogram. Following overnight dark adaptation, micewere anesthetized with an i.p. injection of saline containingketamine (150 mg/kg body weight) and xylazine (5 mg/kg bodyweight). Electroretinograms (ERGs) were recorded from thecorneal surface after pupil dilation (1% atropine sulfate) usinga gold loop corneal electrode together with a mouth referenceand tail ground electrode as described (5). A drop of methyl-cellulose (2.5%, wt/vol) on the corneal surface was used to en-sure electrical contact and to maintain corneal integrity. Bodytemperature was maintained at 38 °C with a heated water pad.All stimuli were presented in a large integrating sphere coatedwith highly reflective white matte paint (#6080; Eastman KodakCorp.). Responses were amplified (Grass CP511 AC amplifier,×10,000) and digitized using a data acquisition board (PCI-1200;National Instruments) in a personal computer. Signal processingwas performed with custom software (LabWindows/CVI; Na-tional Instruments). For each stimulus condition, responseswere computer-averaged with up to 50 records averaged forthe weakest signals. A signal rejection window was adjustedonline to eliminate artifacts. Dark-adapted ERGs were recordedto blue (Kodak Wratten 47A) light flashes up to the maximumintensity of 0.42 cd.s/m2. Cone-mediated responses were ob-tained with white flashes up to a maximum of 4.35 cd.s/m2 ona rod-saturating background (32 cd.s/m2). All stimuli were pre-sented at 1 Hz except for the brightest flashes, where the pre-sentation rate was slowed to 0.2 Hz.
1. Zhuo L, et al. (2001) hGFAP-cre transgenic mice for manipulation of glial and neuronalfunction in vivo. Genesis 31(2):85–94.
2. Li S, et al. (2005) Rhodopsin-iCre transgenic mouse line for Cre-mediated rod-specificgene targeting. Genesis 41(2):73–80.
3. Srinivas S, et al. (2001) Cre reporter strains produced by targeted insertion of EYFP andECFP into the ROSA26 locus. BMC Dev Biol 1:4.
4. Sakagami K, Gan L, Yang XJ (2009) Distinct effects of Hedgehog signaling on neuronalfate specification and cell cycle progression in the embryonic mouse retina. J Neurosci29(21):6932–6944.
5. Nusinowitz S, et al. (2003) Electroretinographic evidence for altered phototransductiongain and slowed recovery from photobleaches in albino mice with a MET450 variant inRPE65. Exp Eye Res 77(5):627–638.
Rhee et al. www.pnas.org/cgi/content/short/1303604110 1 of 8

Fig. S1. Range of infection and time course of gene expression mediated by lentiviruses in vivo. (A) Live fluorescent images of flat mount WT CD1 eyecups 5 dafter subretinal delivery of lentiviruses. Note that the ventral temporal quadrum consistently show intense GFP signals for two LV-IG and two LV-hCNTF in-jected eyes. (B) Merged immunofluorescent GFP labeling and DAPI images of the entire rds/P216L eyecup sections show that viral infection reaches up to 40%of the RPE 10 d after lentiviral infection. (C) Merged immunofluorescent images for Sox9 and GFP shows RPE expression of LV-encoded GFP at 3 and 5 d aftersubretinal injection of rds/P216L mutant eyes. [Scale bars, (A and B) 500 and (C) 50 μm.] gcl, ganglion cell layer; inl, inner nuclear layer; onl, outer nuclear layer;rpe, retinal pigment epithelium.
Rhee et al. www.pnas.org/cgi/content/short/1303604110 2 of 8

Fig. S2. Influence of hCNTF on morphology of rds/P216L mutant photoreceptors. (A) Histological staining of semithin rds/P216L retinal sections are shown.Viral infections were performed at P25 and eyes harvested at P108. Insets show the inner and outer segments of the corresponding images at 3×magnification.(B) Images of peanut agglutinin (PNA)-labeled whole retinas infected with LV-IG or LV-hCNTF at P25 and analyzed at P45. The LV-hCNTF–treated retinas showhigher signals of PNA. (C) Images of PNA-labeled cone cell sheath in different retinal quadrum. [Scale bars, (A and C) 50 and (B) 500 μm.] inl, inner nuclear layer;onl, outer nuclear layer; rpe, retinal pigment epithelium.
Rhee et al. www.pnas.org/cgi/content/short/1303604110 3 of 8

Fig. S3. Effects of subretinal injection on ERG responses. (A) Representative scotopic and photopic ERG waveforms for WT, rds/P216L mutant, and lentiviral-infected rds/P216L eyes are shown. Infection of rds/P216L retinas with LV-IG or LV-hCNTF was carried out at P25, and ERGs of all animals were recorded at P45.(B) Effects of subretinal injections on ERG of rds/P216L eyes were measured at P45. Two LV-IG stocks with different titers and cell culture medium CD293, fromwhich the lentiviruses produced from HEK 293T cells were harvested, were used for injections at P25. Effects of medium or LV injection on the amplitudes ofscotopic a- and b-waves and a photopic b-wave are shown. Statistical analysis (Tables S8–S10) indicates that injection of the CD293 medium and the two LV-IGstocks results in similar suppression on scotopic a-waves, and the CD293 medium causes more significant perturbation on scotopic b-waves. With the exceptionfor the high titer LV-IG (107 CFU/mL), no significant disruption on photopic b-wave was detected.
Rhee et al. www.pnas.org/cgi/content/short/1303604110 4 of 8

Table S1. Summary of antibodies
Cell type Antibody Type Dilution* Source Catalog number or reference
Primary antibodiesMuller glia Cyclin D3 mAb 1:100 Cell Signaling Inc. 2936
Glutamine synthetase (GS) mAb 1:50 Chemicon MAB302Sox9 rAB 1:500 Millipore AB5535
Photoreceptor M-opsin rAb 1:200 Chemicon AB5405PNA 1:250 Vector Laboratories FL-1071Rhodopsin (Rho4D2) mAb 1:250 Dr. R. Molday (1)
Signaling molecules pERK1/2 (Thr202/Tyr204) rAb 1:100 Cell Signaling Inc. 9101pSTAT3 (Tyr705) rAb 1:100, 1:1,000 (wb) Cell Signaling Inc. 9171STAT3 (K-15) rAb 1:1,000 (wb) Cell Signaling Inc. 9132
Miscellaneous CNTF hAb 1:100, 1:1,000 (wb) R&D Systems MAB257GFP rAb 1:200 Invitrogen A6455γ Tubulin mAb 1:5,000 (wb) Sigma T6557
Secondary antibodiesAlexa 488 conjugated gAb 1:500, 1:1,000 (wm) Invitrogen/Molecular Probes A11008Alexa 568 conjugated gAb 1:500, 1:1,000 (wm) Invitrogen/Molecular Probes A11004Alexa 647 (anti-mouse) gAb 1:500 (f) Invitrogen A21235HRP (anti-mouse) shAb 1:1,000 (wb) GE Healthcare NA931HRP (anti-rabbit) dAb 1:1,000 (wb) GE Healthcare NA934
*Dilutions are for immunolabeling of sections, except for flow cytometry (f), whole mount labeling (wm), or Western blot (wb).
1. Laird DW, Molday RS (1988) Evidence against the role of rhodopsin in rod outer segment binding to RPE cells. Invest Ophthalmol Vis Sci 29(3):419–428.
Table S2. Primers for real-time PCR
Type Gene Forward Reverse References
PhotoreceptorCrx GTTCAAGAATCGTAGGGCGAA TGAGATGCCCAAAGGATCTGTNr2e3 AGTCCCAGGTGATGCTAAGC TTCTAAGATGTGCTGCCCC (1)Nrl TTCTGGTTCTGACAGTGACTACG AAGGCTCCCGCTTTATTTC (1)M-opsin CAGTGGCAAAGCAACAGAAA AGGGTGGGCAGTAGCAAAGS-opsin TGGGATTTGTCTTCTTTGTGG CCGAGGGATACATTGACCAGRho TCTTCTTTGGGATGGTGAGC TGGTGGTCATTCTTCGTCCTGnat1 TGACCAATAATGAAGCCATGCT TCTTTGCCCATGGTTGATCA (2)Gnat2 TCCACCTCAGTATCTGTTTTCC TTGAGGTCAAGGAACTGACTCTT (2)
Growth factorsCNTF family CT-1 GAGGCCAAGATCCGCCAGAC TGCACGTATTCCTCCAGAAGTTG (3)
CLC GACTCGTGGGGGATGTTAGC CTAAGCTGCGGAGTTGATGCTCNTF TCTGTAGCCGCTCTATCTGG GGTACACCATCCACTGAGTCAA (4)LIF ATTGTGCCCTTACTGCTGCTG GCCAGTTGATTCTTGATCTGGT (4)OSM CCCTATATCCGCCTCCAAAACC GACTCTGTCCAGTGTGGTGTAC (3)
Others BDNF CAAAGCCACAATGTTCCACCAG GATGTCGTCGTCAGACCTCTCG (5)FGF2 GCGACCCACACGTCAAACTA TCCCTTGATAGACACAACTCCTC (4)EDN2 TGCGTTTTCGTCGATGCTC GTCTGTCCCGCAGTGTTCA (4)
MiscellaneousGAPDH AGGTCGGTGTGAACGGATTTG TGTAGACCATGTAGTTGAGGTCA (4)GFAP CCACCAAACTGGCTGATGTCTAC TTCTCTCCAAATCCACACGAGC (5)
1. Hennig AK, Peng GH, Chen S (2008) Regulation of photoreceptor gene expression by Crx-associated transcription factor network. Brain Res 1192:114–133.2. Montana CL, et al. (2013) Reprogramming of adult rod photoreceptors prevents retinal degeneration. Proc Natl Acad Sci USA 110(5):1732–1737.3. Quinton LJ, et al. (2008) Alveolar epithelial STAT3, IL-6 family cytokines, and host defense during Escherichia coli pneumonia. Am J Respir Cell Mol Biol 38(6):699–706.4. de Melo J, et al. (2012) Injury-independent induction of reactive gliosis in retina by loss of function of the LIM homeodomain transcription factor Lhx2. Proc Natl Acad Sci USA 109(12):
4657–4662.5. Joly S, Lange C, Thiersch M, Samardzija M, Grimm C (2008) Leukemia inhibitory factor extends the lifespan of injured photoreceptors in vivo. J Neurosci 28(51):13765–13774.
Rhee et al. www.pnas.org/cgi/content/short/1303604110 5 of 8

Table S3. Statistics for real-time PCR of photoreceptor genes
Comparison group
Photoreceptor genes (P values)
Crx Nrl Nr2e3 Rho Gnat1 M-Opsin S-Opsin Gnat2
One-way ANOVA 0.000* 0.000* 0.000* 0.000* 0.000* 0.004* 0.011* 0.000*Pairwise comparisonWT vs. P216L 0.000* 0.000* 0.000* 0.000* 0.000* 0.188 0.598 0.004*WT vs. P216L, LV-IG 0.000* 0.000* 0.000* 0.000* 0.000* 0.881 0.355 0.903WT vs. P216L, LV-hCNTF 0.000* 0.000* 0.000* 0.000* 0.000* 0.051 0.131 0.002*P216L vs. P216L, LV-IG 0.333 0.995 0.914 0.605 0.224 0.470 0.961 0.002*P216L vs. P216L, LV-hCNTF 0.200 0.998 0.596 0.437 0.858 0.003* 0.022 0.000*P216L, LV-IG vs. LV-hCNTF 0.978 1.000 0.916 0.988 0.567 0.018* 0.011* 0.004*
*Statistically significant P values.
Table S4. Statistics for ERG shown in Fig. 4B
Comparison group
Scotopic a-wave intensity (P value)
4.37E06 8.43E06 4.37E05 8.43E05 0.00044 0.00084 0.00437 0.00843 0.04374 0.08429 0.10998 0.5104
One-way ANOVA 0.283 0.15 0.998 0.825 0.129 0.159 0.091 0.062 0.350 0.107 0.007* 0.008*Pairwise comparisonP216L vs. LV-IG 0.005* 0.006*P216L vs. LV-hCNTF 0.039 0.024LV-IG vs. LV-hCNTF 0.332 0.285
*Statistically significant P values.
Table S5. Statistics for ERG shown in Fig. 4A
Comparison group
Scotopic b-wave intensity (P value)
4.37E06 8.43E06 4.37E05 8.43E05 0.00044 0.00084 0.00437 0.00843 0.04374 0.08429 0.10998 0.5104
One-way ANOVA 0.095 0.003* 0.000* 0.000* 0.000* 0.000* 0.000* 0.000* 0.000* 0.000* 0.000* 0.000*Pairwise comparisonP216L vs. LV-IG 0.003* 0.000* 0.000* 0.000* 0.000* 0.000* 0.000* 0.000* 0.000* 0.000* 0.000*P216L vs. LV-hCNTF 0.053 0.000* 0.000* 0.000* 0.000* 0.000* 0.000* 0.000* 0.000* 0.000* 0.000*LV-IG vs. LV-hCNTF 0.062 0.997 0.946 0.398 0.395 0.232 0.192 0.094 0.192 0.107 0.090
*Statistically significant P values.
Table S6. Statistics for ERG shown in Fig. 4C
Comparison group
Photopic b-wave intensity (P value)
0.58246 0.74921 1.16721 1.45959 6.19011
One-way ANOVA 0.048 0.036 0.035 0.032 0.010*Pairwise comparison
P216L vs. LV-IG 0.827 0.768 0.876 0.759 0.451P216L vs. LV-hCNTF 0.116 0.086 0.110 0.078 0.019*LV-IG vs. LV-hCNTF 0.112 0.101 0.076 0.093 0.072
*Statistically significant P values.
Rhee et al. www.pnas.org/cgi/content/short/1303604110 6 of 8

Table S7. Statistics for ERG shown in Fig. 4D
Comparison group
Highest intensity (P value)
Scotopic a-wave Scotopic b-wave Photopic b-wave
One-way ANOVA 0.008* 0.000* 0.010*Pairwise comparisonP216L vs. LV-IG 0.006* 0.000* 0.451P216L vs. LV-hCNTF 0.024 0.000* 0.019*LV-IG vs. LV-hCNTF 0.531 0.072 0.072
*Statistically significant P values.
Table S8. Statistics for ERG scotopic a-wave impacted by injections shown in Fig. S3B
Comparison group
Scotopic a-wave intensity (P value)
4.37E05 8.43E05 0.00044 0.00084 0.00437 0.00843 0.04374 0.08429 0.10998 0.5104
One-way ANOVA 0.130 0.945 0.138 0.201 0.040 0.330 0.643 0.190 0.020 0.004*Pairwise comparisonP216L vs. LV-IG 107 0.029 0.021P216L vs. LV-IG 105 0.163 0.030P216L vs. CD293 0.053 0.007*LV-IG 107 vs. LVIG5 0.816 0.998LV-IG 107 vs. CD293 0.991 0.948LV-IG 105 vs. CD293 0.933 0.889
*Statistically significant P values.
Table S9. Statistics for ERG scotopic b-wave impacted by injections shown in Fig. S3B
Comparison group
Scotopic b-wave intensity (P value)
4.37E05 8.43E05 0.00044 0.00084 0.00437 0.00843 0.04374 0.08429 0.10998 0.5104
One-way ANOVA 0.010* 0.005* 0.003* 0.002* 0.002* 0.001* 0.013* 0.015* 0.014* 0.015*Pairwise comparisonP216L vs. LV-IG 107 0.100 0.096 0.039 0.043 0.027 0.029 0.069 0.091 0.075 0.065P216L vs. LV-IG 105 0.103 0.165 0.159 0.110 0.049 0.058 0.289 0.207 0.178 0.176P216L vs. CD293 0.008* 0.003* 0.002* 0.001* 0.002* 0.001* 0.011* 0.012* 0.013* 0.015*LV-IG 107 vs. LV-IG5 1.000 0.991 0.882 0.963 0.992 0.986 0.856 0.968 0.966 0.950LV-IG 107 vs. CD293 0.612 0.348 0.532 0.394 0.545 0.383 0.813 0.758 0.825 0.894LV-IG 105 vs. CD293 0.604 0.229 0.197 0.198 0.390 0.236 0.371 0.496 0.564 0.616
*Statistically significant P values.
Table S10. Statistics for ERG photopic b-wave impacted byinjections shown in Fig. S3B
Comparison group
Photopic b-wave intensity (P value)
0.58246 0.74921 1.16721 1.45959 6.19011
One-way ANOVA 0.236 0.046 0.194 0.100 0.015*Pairwise comparison
P216L vs. LV-IG 107 0.055 0.012*P216L vs. LV-IG 105 0.999 0.424P216L vs. CD293 0.691 0.106LV-IG 107 vs. LV-IG5 0.066 0.264LV-IG 107 vs. CD293 0.412 0.703LV-IG 105 vs. CD293 0.663 0.837
*Statistically significant P values.
Rhee et al. www.pnas.org/cgi/content/short/1303604110 7 of 8

Table S11. Statistics for real-time PCR of growth factor transcripts affected by viral infections
Comparison group
Growth factors (P values)
GFAP CNTF LIF CT-1 CLC OsM FGF-2 Edn-2 BDNF
One-way ANOVA 0.000* 0.000* 0.112 0.001* 0.024 0.496 0.042 00.000* 0.000*Pairwise comparison
WT vs. P216L 0.000* 0.001* 0.001* 0.815 0.519 0.005* 0.297WT vs. P216L, LV-IG 0.003* 0.001* 0.001* 0.020 0.757 0.704 0.699WT vs. P216L, LV-hCNTF 0.000* 0.000* 0.003* 0.664 0.032 0.000* 0.000*P216L vs. P216L, LV-IG 0.106 0.976 0.999 0.069 0.971 0.021 0.849P216L vs. P216L, LV-hCNTF 0.256 0.220 0.717 0.991 0.232 0.017* 0.000*P216L, LV-IG vs. LV-hCNTF 0.007* 0.127 0.781 0.104 0.129 0.000* 0.000*
*Statistically significant P values.
Table S12. Statistics for real-time PCR of cytokine transcriptsaffected by CNTF injection
Comparison group
Growth factors (P values)
LIF CNTF CT-1
One-way ANOVA 0.000* 0.003* 0.854Pairwise comparison
3H PBS vs. 3H CNTF 0.001* 1.0003H PBS vs. 24H PBS 0.090 0.0373H PBS vs. 24H CNTF 0.088 0.007*3H CNTF vs. 24H PBS 0.000* 0.0383H CNTF vs. 24H CNTF 0.000* 0.007*24H PBS vs. 24H CNTF 1.000 0.606
*Statistically significant P values.
Rhee et al. www.pnas.org/cgi/content/short/1303604110 8 of 8




















