
Cloning and functional expression of an acyl-ACP thioesterase FatB type from Diploknema (Madhuca)...
11
Research article Cloning and functional expression of an acyl-ACP thioesterase FatB type from Diploknema (Madhuca) butyracea seeds in Escherichia coli J.K. Jha a , M.K. Maiti a, * , A. Bhattacharjee a,1 , A. Basu a , P.C. Sen b , S.K. Sen a a IIT-BREF Biotek, Indian Institute of Technology, Kharagpur-721302, India b Department of Chemistry, Bose Institute, Kolkata-700009, India Received 25 March 2006; accepted 22 September 2006 Available online 13 October 2006 Abstract A cDNA of fatty acyl- acyl carrier protein (ACP) thioesterase (Fat) from developing seed of Madhuca butyracea has been cloned. The deduced amino acid sequence of the cDNA corresponding to the mature polypeptide showed 30–40% and 60–75% identity to the reported FatA and FatB class of plant thioesterases, respectively. This gene, MbFatB, is present as a single copy in M. butyracea genome and the MbFatB protein was detected clearly in seed tissues of this plant but not in that of Indian mustard (Brassica juncea). Heterologous expression of the MbFatB gene driven by different promoters in E. coli wild type and fatty acid β-oxidation mutant (fadD88) strains resulted production of the recombinant protein with various fusion tags either as biologically inactive (insoluble) or functionally active forms. Expression of functionally active recombinant MbFatB in E. coli affected bacterial growth and cell morphology as well as changed the fatty acid profiles of the membrane lipid and the culture supernatant. Alteration of the fatty acid composition was directed predominantly towards palmitate and to a lesser extent myristate and oleate due to acyl chain termination activity of plant thioesterase in bacteria. Thus, this new MbFatB gene isolated from a non- traditional oil-seed tree can be used in future for transgenic development of oil-seed Brassica, a widely cultivated crop that expresses predomi- nantly oleoyl-ACP thioesterase (FatA) in its seed tissue and has high amount of unwanted erucic acid in edible oil in order to alter the fatty acid profile in a desirable way. © 2006 Elsevier Masson SAS. All rights reserved. Keywords: Acyl-acyl carrier protein (ACP) thioesterase; FatB cDNA; Fatty acid profile analysis; Heterologous expression in E. coli; Diploknema (Madhuca) butyracea 1. Introduction The structural and functional organizations of the enzymes involved in de novo fatty acid synthesis in plant plastids and bacteria are similar and the multiple enzyme complex is known as fatty acid synthase (FAS). The reactions of the type-II FAS, unlike the type-I FAS of animals and yeast, are catalyzed by discrete and monofunctional enzymes through transfer of a growing acyl chain bound to the soluble cofactor, acyl carrier protein (ACP). For chain elongation the growing acyl-ACP is added to a primer of two-carbon unit by a cycle of four enzy- matic steps and the cycle continues until the desired carbon length of the fatty acid is achieved. The carbon chain elonga- tion stops by either of the following two pathways—‘prokar- yotic or plastidial’ and ‘eukaryotic or cytoplasmic’. By acyl- transferase enzyme, the acyl moiety from acyl-ACP is incorporated into a variety of glycerolipids essential for all bio- logical membranes including the plastidial one. This is known as ‘prokaryotic or plastidial pathway’ of lipid biosynthesis. Alternately, a specialized plastidial enzyme known as fatty acyl- acyl carrier protein (ACP) thioesterase (Fat) can also ter- minate the acyl chain elongation during FAS by hydrolyzing the thioester bond of acyl-ACP, releasing free fatty acids and www.elsevier.com/locate/plaphy Plant Physiology and Biochemistry 44 (2006) 645–655 Abbreviations: ACP, acyl carrier protein; FAME, fatty acid methyl ester; FAS, fatty acid synthase; Fat, fatty acyl-acyl carrier protein (ACP) thioesterase; GC-FID, gas chromatography-flame ionization detection; MbFatB, Madhuca (Diploknema) butyracea fatty acyl-acyl carrier protein (ACP) thioesterase of B class; TLC, thin layer chromatography; UTR, untranslated transcribed region. * Corresponding author. Tel.: +91 3222 281 084/281 086; fax: +91 3222 277 890. E-mail address: [email protected] (M.K. Maiti). 1 Present address: The Cleveland Clinic, 9500 Euclid Avenue, Cleveland, OH 44195, USA. 0981-9428/$ - see front matter © 2006 Elsevier Masson SAS. All rights reserved. doi:10.1016/j.plaphy.2006.09.017
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Cloning and functional expression of an acyl-ACP thioesterase FatB type from Diploknema (Madhuca)...

www.elsevier.com/locate/plaphy
Plant Physiology and Biochemistry 44 (2006) 645–655
Research article
Cloning and functional expression of an acyl-ACP thioesterase FatB typefrom Diploknema (Madhuca) butyracea seeds in Escherichia coli
J.K. Jhaa, M.K. Maitia,*, A. Bhattacharjeea,1, A. Basua, P.C. Senb, S.K. Sena
a IIT-BREF Biotek, Indian Institute of Technology, Kharagpur-721302, IndiabDepartment of Chemistry, Bose Institute, Kolkata-700009, India
Received 25 March 2006; accepted 22 September 2006Available online 13 October 2006
Abstract
A cDNA of fatty acyl-acyl carrier protein (ACP) thioesterase (Fat) from developing seed of Madhuca butyracea has been cloned. Thededuced amino acid sequence of the cDNA corresponding to the mature polypeptide showed 30–40% and 60–75% identity to the reportedFatA and FatB class of plant thioesterases, respectively. This gene, MbFatB, is present as a single copy in M. butyracea genome and the MbFatBprotein was detected clearly in seed tissues of this plant but not in that of Indian mustard (Brassica juncea). Heterologous expression of theMbFatB gene driven by different promoters in E. coli wild type and fatty acid β-oxidation mutant (fadD88) strains resulted production of therecombinant protein with various fusion tags either as biologically inactive (insoluble) or functionally active forms. Expression of functionallyactive recombinant MbFatB in E. coli affected bacterial growth and cell morphology as well as changed the fatty acid profiles of the membranelipid and the culture supernatant. Alteration of the fatty acid composition was directed predominantly towards palmitate and to a lesser extentmyristate and oleate due to acyl chain termination activity of plant thioesterase in bacteria. Thus, this new MbFatB gene isolated from a non-traditional oil-seed tree can be used in future for transgenic development of oil-seed Brassica, a widely cultivated crop that expresses predomi-nantly oleoyl-ACP thioesterase (FatA) in its seed tissue and has high amount of unwanted erucic acid in edible oil in order to alter the fatty acidprofile in a desirable way.© 2006 Elsevier Masson SAS. All rights reserved.
Keywords: Acyl-acyl carrier protein (ACP) thioesterase; FatB cDNA; Fatty acid profile analysis; Heterologous expression in E. coli; Diploknema (Madhuca)butyracea
1. Introduction
The structural and functional organizations of the enzymesinvolved in de novo fatty acid synthesis in plant plastids andbacteria are similar and the multiple enzyme complex is knownas fatty acid synthase (FAS). The reactions of the type-II FAS,
Abbreviations: ACP, acyl carrier protein; FAME, fatty acid methyl ester;FAS, fatty acid synthase; Fat, fatty acyl-acyl carrier protein (ACP)thioesterase; GC-FID, gas chromatography-flame ionization detection;MbFatB, Madhuca (Diploknema) butyracea fatty acyl-acyl carrier protein(ACP) thioesterase of B class; TLC, thin layer chromatography; UTR,untranslated transcribed region.
* Corresponding author. Tel.: +91 3222 281 084/281 086; fax: +913222 277 890.
E-mail address: [email protected] (M.K. Maiti).1 Present address: The Cleveland Clinic, 9500 Euclid Avenue, Cleveland,
OH 44195, USA.
0981-9428/$ - see front matter © 2006 Elsevier Masson SAS. All rights reserved.doi:10.1016/j.plaphy.2006.09.017
unlike the type-I FAS of animals and yeast, are catalyzed bydiscrete and monofunctional enzymes through transfer of agrowing acyl chain bound to the soluble cofactor, acyl carrierprotein (ACP). For chain elongation the growing acyl-ACP isadded to a primer of two-carbon unit by a cycle of four enzy-matic steps and the cycle continues until the desired carbonlength of the fatty acid is achieved. The carbon chain elonga-tion stops by either of the following two pathways—‘prokar-yotic or plastidial’ and ‘eukaryotic or cytoplasmic’. By acyl-transferase enzyme, the acyl moiety from acyl-ACP isincorporated into a variety of glycerolipids essential for all bio-logical membranes including the plastidial one. This is knownas ‘prokaryotic or plastidial pathway’ of lipid biosynthesis.Alternately, a specialized plastidial enzyme known as fattyacyl-acyl carrier protein (ACP) thioesterase (Fat) can also ter-minate the acyl chain elongation during FAS by hydrolyzingthe thioester bond of acyl-ACP, releasing free fatty acids and

Fig. 1. Alignment of partial amino acid sequences from two classes of acyl-ACP thioesterase showing conserved regions used to design the primes forcloning the part of M. butyracea thioesterase cDNA. FatN and FatC are twodegenerate oligonucletides shown in lowercase. The four each of FatAs andFatBs and their GenBank accession numbers are as follows: AtFatA1,A. thaliana Z36912; BnFatA1 B. napus X73849; CtFatA1, C. tinctoriusM96569; GmFatA1, G. mangostana U92876; AtFatB1, A. thaliana Z36911;ChFatB1, C. hookeriana U17076; CwFatB1, C. wrightii U56103; GmFatB1,G. mangostana U92878.
J.K. Jha et al. / Plant Physiology and Biochemistry 44 (2006) 645–655646
ACP. The substrate specificities of Fat enzymes play crucialrole in determining the pool of free fatty acids that are conse-quently re-esterified with Coenzyme A to form acyl-CoA fortransport from plastid to endoplasmic reticulum (ER) and arerequired for glycerolipid synthesis. This is known as ‘eukaryo-tic or cytoplasmic pathway’ of lipid biosynthesis. In all plantcells, the glycerolipids generated in ER may shuttle betweenthe two organelles for necessary modifications during mem-brane lipid biosynthesis. However, in fat-storage tissues (e.g.developing seeds) most of the ER glycerolipids are convertedinto triacylglycerol for oil biosynthesis and only some parts areshuttled back into plastid for biosynthesis of membrane lipid.Thus, plant Fat is the key enzyme regulating the carbon fluxbetween the ‘prokaryotic’ and ‘eukaryotic’ pathways of lipidbiosynthesis, and not only it control the quality and quantityof fatty acid composition of the seed oil but also to some extentof the membrane lipid.
The plant Fat enzymes, encoded by nuclear genes, are tar-geted into the plastid. Based on sequence identity and substratespecificity, plant thioesterases have been classified into twodistinct but related (paralogous) gene families—FatA andFatB [17]. FatA is the housekeeping enzyme and ubiquitouslypresent in all plant tissues. The FatA orthologues reported sofar, have the highest activity towards unsaturated oleoyl(C18:1)-ACP and several fold lower specificity for saturatedacyl-ACPs [23]. However, the FatA1 of mangosteen (Garci-nia mangostana), a tropical plant having 56% stearic acid inseed oil, was found to be partly responsible for that [14]. Onthe other hand, FatB thioesterases have heterogeneous specifi-city towards various saturated acyl-ACPs including certainactivity for unsaturated C18:1-ACP [23]. Initially, FatB wasconsidered to be a medium (C12:0-ACP) chain specific thioes-terase found in developing seed tissues of California bay(Umbellularia californica), which is involved in the productionof oil rich in lauric acid [7]; but later several FatB orthologuesdiscovered in seeds of different plants to have thioesterase spe-cificity for various saturated acyl-ACPs, ranging from C8:0-ACP to C18:0-ACP [4,6,8,9,11,23,31,34]. Similar to FatA,FatB is also regarded to be present in all plant tissues but pre-dominantly expressing in developing seeds. This is in consis-tence with the observation that plant membrane (cellular andorganellar) lipids are rich in 16 and 18 carbon fatty acids,and different seed-oils/fats contain C18:1 and/or its derivativesalong with various saturated fatty acids (short, medium andlong carbon chain). In many seed tissues (e.g. Brassica, Arabi-dopsis), FatA is more abundant than FatB due to down regula-tion of the later thioesterase; where as in some other seeds(such as California bay, Cuphea, coconut, palm and cotton)the reverse is true. Experimental evidences in vitro demon-strated the close functional relationships among the membersof FatA and FatB class of thioesterases, i.e. FatAs are activepredominantly towards unsaturated acyl-ACPs (mainly C18:1-ACP); whereas FatBs have preferential specificity for one par-ticular saturated acyl-ACPs. Recent molecular modelingexperiments in silico and by site directed mutagenesis demon-strated their predicted 3-D structures are conserved [18,26].
Previously, we identified through protein purification and invitro enzyme assay a heat-stable palmitoyl/oleoyl specific acyl-ACP thioesterase having activity primarily in developing seedsas well as in young leaves and flower buds of Madhuca butyr-acea [2]. The seed-fat (called Phulwara fat) of this non-traditional oil-seed tree growing in sub-Himalayan region con-tains saturated and unsaturated fatty acids [2,25], the C16:0(palmitic acid) being the predominant constituent (65%).Indian mustard (Brassica juncea) and rapeseed (Brassica cam-pestris) are two widely cultivated oil-seed crops in Indian sub-continent and oil-seed Brassicas predominantly express oleoyl-ACP thioesterase (FatA) in seed tissues [19,22]. These Brassi-cas are having high amount (40–55%) of C18:1 derived verylong chain unsaturated fatty acids (e.g. C22:1 erucic acid) intheir seed oils, which are unwanted for human edible purpose.We were interested to characterize the gene encoding the novelthioesterase enzyme of M. butyracea and for its potential appli-cation in metabolic engineering of seed oil biosynthesis inBrassica in order to alter the fatty acid profile in a desirableway. We report here the cloning of a seed-expressed thioester-ase cDNA from M. butyracea and its functional characteriza-tion through heterologous expression driven by different pro-moters in various Escherichia coli strains followed by fattyacid profile analysis.
2. Results
2.1. Sequence analysis of a cloned thioesterase gene fromM. butyracea
By combining the reverse transcriptase-polymerase chainreaction (RT-PCR) and rapid amplification of cDNA end(RACE) techniques using degenerate primers (Fig. 1) andgene-specific primers (Fig. 2), we cloned the near-full lengthcDNA sequence of a Fat from the seed tissues of M. butyracea.The 1.218 kb DNA sequence has been deposited in GenBank

Fig. 2. Alignment of predicted secondary structures derived from the amino acid sequences of mature FatB thioesterases having diverse substrate specificity. Aminoacid sequence derived from M. butyracea thioesterase (MbFatB AY835984) depicted in first row followed by predicted secondary structures of MbFatB, GmFatB1(G. mangostana U92878), CcFatB1 (C. camphora U31813) and UcFatB1 (U. californica M94159). The later three FatBs have the thioesterase specificitypredominantly towards C16:0-ACP, C14:0-ACP and C12:0-ACP, respectively. The putative N terminus of the mature MbFatB thioesterase indicated by verticalarrowhead as proposed for ChFatB1 [17] and AtFatB1 [10]. The two conserved amino acid domains containing the two essential residues (H232 and C267 of matureMbFatB) for thioesterase catalytic activity are boxed. The similar and dissimilar amino acid replacements in MbFatB are indicated by bold and bold-underlined. Theamino acid sequence corresponding to the four gene-specific primers (GSP1, 2, 3 and 4) used for cloning are shown by horizontal arrows. The consensus secondarystructure of individual FatB was predicted by using Network Protein Sequence Analysis software [5] and the codes are: C random coiled; H α-helix, E extended β-strand. The multiple alignment was performed by ClustalW.
J.K. Jha et al. / Plant Physiology and Biochemistry 44 (2006) 645–655 647
(accession number AY835984). Although the 5′-end of thegene has not been cloned, we were able to obtain the 3′-untranslated transcribed region (3′-UTR) and coding DNAsequence corresponding to the mature polypeptide of theM. butyracea thioesterase based on the transit peptide cleavagesites of published thioesterases [14,16]. The derived aminoacid sequence of M. butyracea thioesterase mature polypeptide(Fig. 2) revealed ~60–75% identity with several reported FatBtype thioesterases, whereas only ~30–40% homology withFatA type thioesterases. It is interesting to note that this newthioesterase showed strong identity, i.e. 77% withG. mangostana FatB1 (GmFatB1). It was also found that a
few conserved amino acid residues essential for catalytic activ-ity of thioesterase [33] are distributed in the mature polypep-tide region. Based on sequence analysis, the M. butyraceathioesterase gene was assigned as MbFatB. Although thereare scattered but significant amino acid differences amongstthe set of four FatB thioesterase enzymes (MbFatB, GmFatB1,CcFatB1 and UcFatB1) having different substrate specificityand originated from different plants, the secondary structureof this class of FatB protein remain conserved (Fig. 2). Twocritical amino acid residues, histidine and cysteine essential forcatalytic activity of any FatB thioesterase, belonging to theconserved active-site motifs NQHVNN and YRRECG [33],
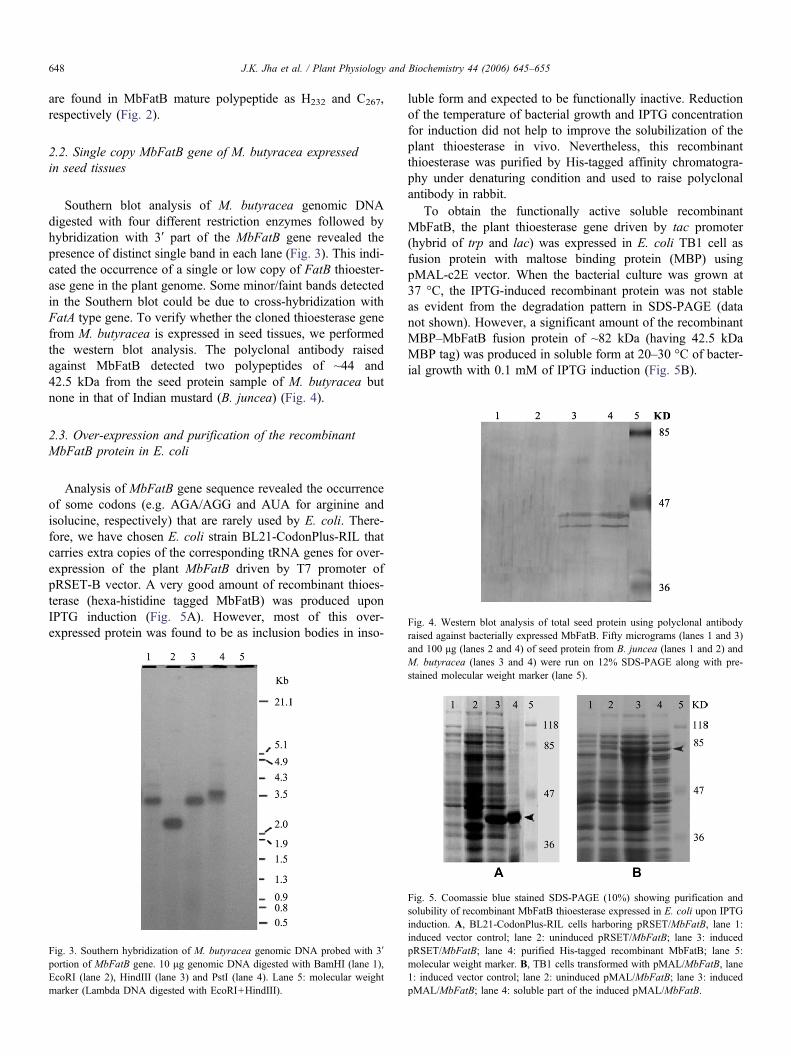
Fig. 4. Western blot analysis of total seed protein using polyclonal antibodyraised against bacterially expressed MbFatB. Fifty micrograms (lanes 1 and 3)
J.K. Jha et al. / Plant Physiology and Biochemistry 44 (2006) 645–655648
are found in MbFatB mature polypeptide as H232 and C267,respectively (Fig. 2).
2.2. Single copy MbFatB gene of M. butyracea expressedin seed tissues
Southern blot analysis of M. butyracea genomic DNAdigested with four different restriction enzymes followed byhybridization with 3′ part of the MbFatB gene revealed thepresence of distinct single band in each lane (Fig. 3). This indi-cated the occurrence of a single or low copy of FatB thioester-ase gene in the plant genome. Some minor/faint bands detectedin the Southern blot could be due to cross-hybridization withFatA type gene. To verify whether the cloned thioesterase genefrom M. butyracea is expressed in seed tissues, we performedthe western blot analysis. The polyclonal antibody raisedagainst MbFatB detected two polypeptides of ~44 and42.5 kDa from the seed protein sample of M. butyracea butnone in that of Indian mustard (B. juncea) (Fig. 4).
2.3. Over-expression and purification of the recombinantMbFatB protein in E. coli
Analysis of MbFatB gene sequence revealed the occurrenceof some codons (e.g. AGA/AGG and AUA for arginine andisolucine, respectively) that are rarely used by E. coli. There-fore, we have chosen E. coli strain BL21-CodonPlus-RIL thatcarries extra copies of the corresponding tRNA genes for over-expression of the plant MbFatB driven by T7 promoter ofpRSET-B vector. A very good amount of recombinant thioes-terase (hexa-histidine tagged MbFatB) was produced uponIPTG induction (Fig. 5A). However, most of this over-expressed protein was found to be as inclusion bodies in inso-
Fig. 3. Southern hybridization of M. butyracea genomic DNA probed with 3′portion of MbFatB gene. 10 μg genomic DNA digested with BamHI (lane 1),EcoRI (lane 2), HindIII (lane 3) and PstI (lane 4). Lane 5: molecular weightmarker (Lambda DNA digested with EcoRI+HindIII).
luble form and expected to be functionally inactive. Reductionof the temperature of bacterial growth and IPTG concentrationfor induction did not help to improve the solubilization of theplant thioesterase in vivo. Nevertheless, this recombinantthioesterase was purified by His-tagged affinity chromatogra-phy under denaturing condition and used to raise polyclonalantibody in rabbit.
To obtain the functionally active soluble recombinantMbFatB, the plant thioesterase gene driven by tac promoter(hybrid of trp and lac) was expressed in E. coli TB1 cell asfusion protein with maltose binding protein (MBP) usingpMAL-c2E vector. When the bacterial culture was grown at37 °C, the IPTG-induced recombinant protein was not stableas evident from the degradation pattern in SDS-PAGE (datanot shown). However, a significant amount of the recombinantMBP–MbFatB fusion protein of ~82 kDa (having 42.5 kDaMBP tag) was produced in soluble form at 20–30 °C of bacter-ial growth with 0.1 mM of IPTG induction (Fig. 5B).
and 100 μg (lanes 2 and 4) of seed protein from B. juncea (lanes 1 and 2) andM. butyracea (lanes 3 and 4) were run on 12% SDS-PAGE along with pre-stained molecular weight marker (lane 5).
Fig. 5. Coomassie blue stained SDS-PAGE (10%) showing purification andsolubility of recombinant MbFatB thioesterase expressed in E. coli upon IPTGinduction. A, BL21-CodonPlus-RIL cells harboring pRSET/MbFatB, lane 1:induced vector control; lane 2: uninduced pRSET/MbFatB; lane 3: inducedpRSET/MbFatB; lane 4: purified His-tagged recombinant MbFatB; lane 5:molecular weight marker. B, TB1 cells transformed with pMAL/MbFatB, lane1: induced vector control; lane 2: uninduced pMAL/MbFatB; lane 3: inducedpMAL/MbFatB; lane 4: soluble part of the induced pMAL/MbFatB.
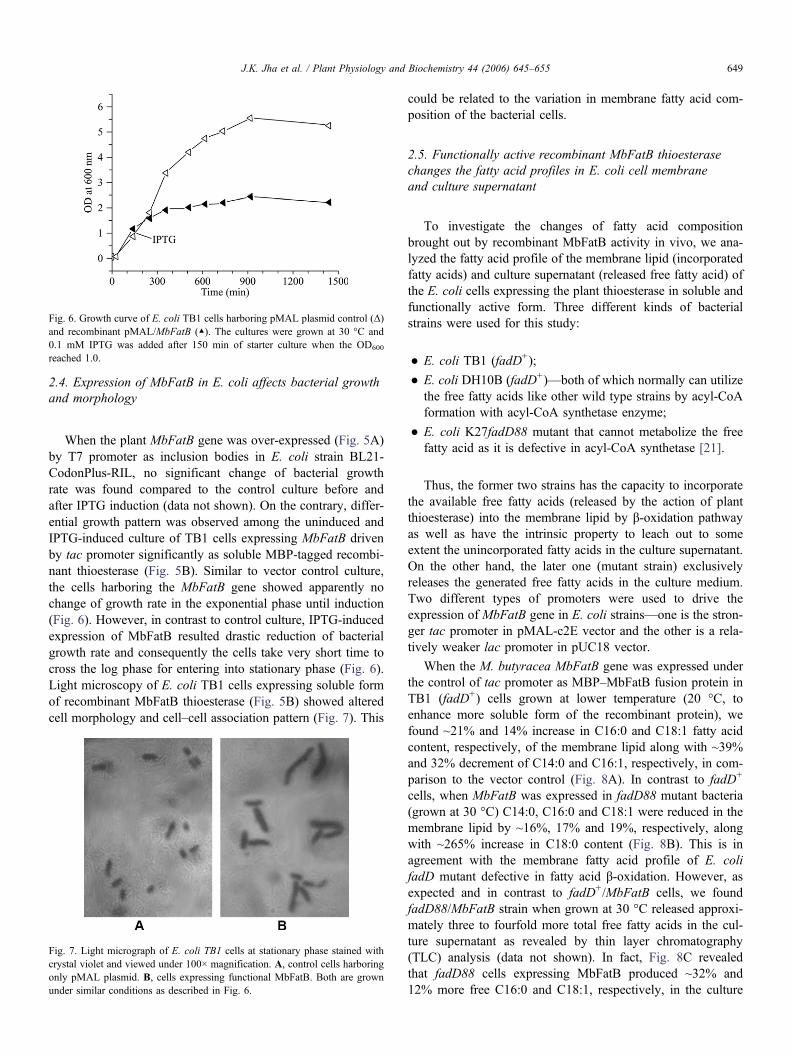
Fig. 6. Growth curve of E. coli TB1 cells harboring pMAL plasmid control (Δ)and recombinant pMAL/MbFatB (▲). The cultures were grown at 30 °C and0.1 mM IPTG was added after 150 min of starter culture when the OD600
reached 1.0.
J.K. Jha et al. / Plant Physiology and Biochemistry 44 (2006) 645–655 649
2.4. Expression of MbFatB in E. coli affects bacterial growthand morphology
When the plant MbFatB gene was over-expressed (Fig. 5A)by T7 promoter as inclusion bodies in E. coli strain BL21-CodonPlus-RIL, no significant change of bacterial growthrate was found compared to the control culture before andafter IPTG induction (data not shown). On the contrary, differ-ential growth pattern was observed among the uninduced andIPTG-induced culture of TB1 cells expressing MbFatB drivenby tac promoter significantly as soluble MBP-tagged recombi-nant thioesterase (Fig. 5B). Similar to vector control culture,the cells harboring the MbFatB gene showed apparently nochange of growth rate in the exponential phase until induction(Fig. 6). However, in contrast to control culture, IPTG-inducedexpression of MbFatB resulted drastic reduction of bacterialgrowth rate and consequently the cells take very short time tocross the log phase for entering into stationary phase (Fig. 6).Light microscopy of E. coli TB1 cells expressing soluble formof recombinant MbFatB thioesterase (Fig. 5B) showed alteredcell morphology and cell–cell association pattern (Fig. 7). This
Fig. 7. Light micrograph of E. coli TB1 cells at stationary phase stained withcrystal violet and viewed under 100× magnification. A, control cells harboringonly pMAL plasmid. B, cells expressing functional MbFatB. Both are grownunder similar conditions as described in Fig. 6.
could be related to the variation in membrane fatty acid com-position of the bacterial cells.
2.5. Functionally active recombinant MbFatB thioesterasechanges the fatty acid profiles in E. coli cell membraneand culture supernatant
To investigate the changes of fatty acid compositionbrought out by recombinant MbFatB activity in vivo, we ana-lyzed the fatty acid profile of the membrane lipid (incorporatedfatty acids) and culture supernatant (released free fatty acid) ofthe E. coli cells expressing the plant thioesterase in soluble andfunctionally active form. Three different kinds of bacterialstrains were used for this study:
● E. coli TB1 (fadD+);
● E. coli DH10B (fadD+)—both of which normally can utilizethe free fatty acids like other wild type strains by acyl-CoAformation with acyl-CoA synthetase enzyme;
● E. coli K27fadD88 mutant that cannot metabolize the freefatty acid as it is defective in acyl-CoA synthetase [21].
Thus, the former two strains has the capacity to incorporatethe available free fatty acids (released by the action of plantthioesterase) into the membrane lipid by β-oxidation pathwayas well as have the intrinsic property to leach out to someextent the unincorporated fatty acids in the culture supernatant.On the other hand, the later one (mutant strain) exclusivelyreleases the generated free fatty acids in the culture medium.Two different types of promoters were used to drive theexpression of MbFatB gene in E. coli strains—one is the stron-ger tac promoter in pMAL-c2E vector and the other is a rela-tively weaker lac promoter in pUC18 vector.
When the M. butyracea MbFatB gene was expressed underthe control of tac promoter as MBP–MbFatB fusion protein inTB1 (fadD+) cells grown at lower temperature (20 °C, toenhance more soluble form of the recombinant protein), wefound ~21% and 14% increase in C16:0 and C18:1 fatty acidcontent, respectively, of the membrane lipid along with ~39%and 32% decrement of C14:0 and C16:1, respectively, in com-parison to the vector control (Fig. 8A). In contrast to fadD+
cells, when MbFatB was expressed in fadD88 mutant bacteria(grown at 30 °C) C14:0, C16:0 and C18:1 were reduced in themembrane lipid by ~16%, 17% and 19%, respectively, alongwith ~265% increase in C18:0 content (Fig. 8B). This is inagreement with the membrane fatty acid profile of E. colifadD mutant defective in fatty acid β-oxidation. However, asexpected and in contrast to fadD+/MbFatB cells, we foundfadD88/MbFatB strain when grown at 30 °C released approxi-mately three to fourfold more total free fatty acids in the cul-ture supernatant as revealed by thin layer chromatography(TLC) analysis (data not shown). In fact, Fig. 8C revealedthat fadD88 cells expressing MbFatB produced ~32% and12% more free C16:0 and C18:1, respectively, in the culture
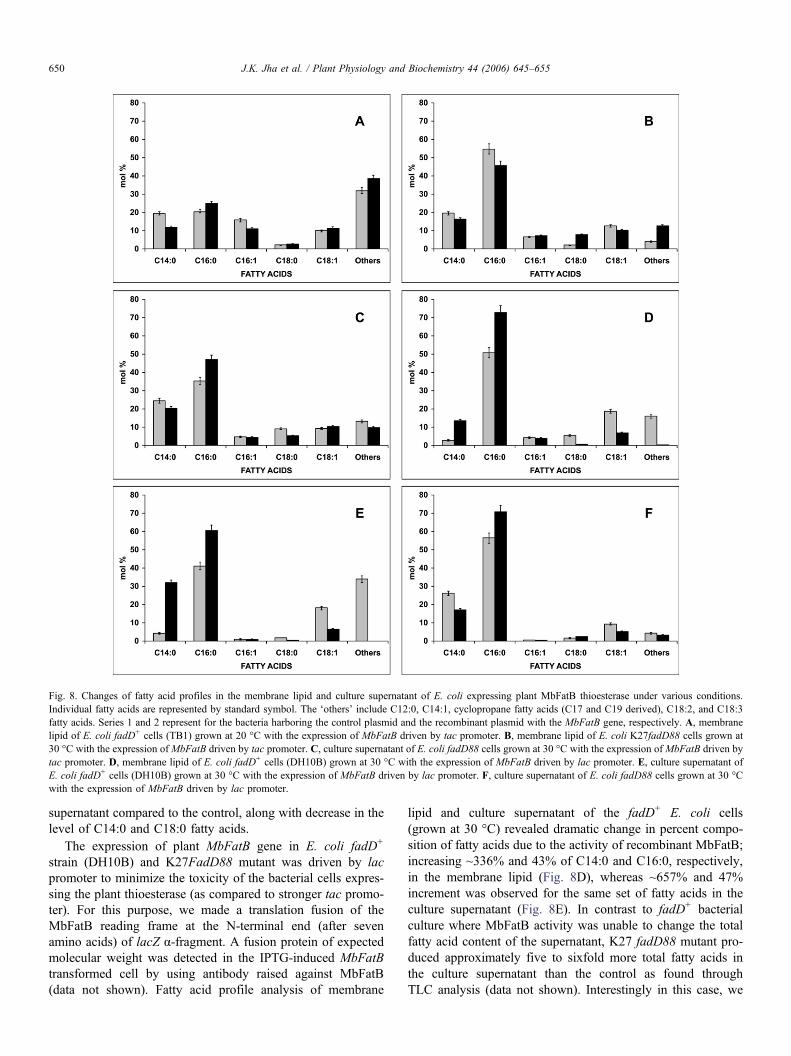
Fig. 8. Changes of fatty acid profiles in the membrane lipid and culture supernatant of E. coli expressing plant MbFatB thioesterase under various conditions.Individual fatty acids are represented by standard symbol. The ‘others’ include C12:0, C14:1, cyclopropane fatty acids (C17 and C19 derived), C18:2, and C18:3fatty acids. Series 1 and 2 represent for the bacteria harboring the control plasmid and the recombinant plasmid with the MbFatB gene, respectively. A, membranelipid of E. coli fadD+ cells (TB1) grown at 20 °C with the expression of MbFatB driven by tac promoter. B, membrane lipid of E. coli K27fadD88 cells grown at30 °C with the expression ofMbFatB driven by tac promoter. C, culture supernatant of E. coli fadD88 cells grown at 30 °C with the expression ofMbFatB driven bytac promoter. D, membrane lipid of E. coli fadD+ cells (DH10B) grown at 30 °C with the expression of MbFatB driven by lac promoter. E, culture supernatant ofE. coli fadD+ cells (DH10B) grown at 30 °C with the expression of MbFatB driven by lac promoter. F, culture supernatant of E. coli fadD88 cells grown at 30 °Cwith the expression of MbFatB driven by lac promoter.
J.K. Jha et al. / Plant Physiology and Biochemistry 44 (2006) 645–655650
supernatant compared to the control, along with decrease in thelevel of C14:0 and C18:0 fatty acids.
The expression of plant MbFatB gene in E. coli fadD+
strain (DH10B) and K27FadD88 mutant was driven by lacpromoter to minimize the toxicity of the bacterial cells expres-sing the plant thioesterase (as compared to stronger tac promo-ter). For this purpose, we made a translation fusion of theMbFatB reading frame at the N-terminal end (after sevenamino acids) of lacZ α-fragment. A fusion protein of expectedmolecular weight was detected in the IPTG-induced MbFatBtransformed cell by using antibody raised against MbFatB(data not shown). Fatty acid profile analysis of membrane
lipid and culture supernatant of the fadD+ E. coli cells(grown at 30 °C) revealed dramatic change in percent compo-sition of fatty acids due to the activity of recombinant MbFatB;increasing ~336% and 43% of C14:0 and C16:0, respectively,in the membrane lipid (Fig. 8D), whereas ~657% and 47%increment was observed for the same set of fatty acids in theculture supernatant (Fig. 8E). In contrast to fadD+ bacterialculture where MbFatB activity was unable to change the totalfatty acid content of the supernatant, K27 fadD88 mutant pro-duced approximately five to sixfold more total fatty acids inthe culture supernatant than the control as found throughTLC analysis (data not shown). Interestingly in this case, we

J.K. Jha et al. / Plant Physiology and Biochemistry 44 (2006) 645–655 651
found only enhancement of C16:0 ~25% more in the culturesupernatant, but the level of C14:0 and C18:1 decreased uponexpression of MbFatB in E. coli K27 fadD88 (Fig. 8F).
3. Discussion
Propensity and diversity of fatty acids in seed fats/oils arelargely controlled by the substrate specificity of FatB thioester-ase enzyme active in seed tissues of the respective plant spe-cies. These genes from few plants have been used for trans-genic approach [8,11,14] to produce tailor-made fatty acids inthe cultivated crop and model plant where they are normallyabsent or poorly expressed. In certain cases, protein engineer-ing has been utilized to alter the substrate specificity of theselected Fats to improve the desired quality of the oil [13,15,34]. Thus, functional characterization of newly identifiedthioesterase from any plant is the primary requirement toincrease the gene pool necessary for metabolic engineering offatty acid biosynthesis in crop plants to make desirable fats/oilsfor edible or industrial purpose.
In our previous report [2], we identified a heat-stable palmi-toyl/oleoyl specific Fat in developing seeds of M. butyracea. Inthe present study, we cloned the cDNA corresponding to thatthioesterase gene, named as MbFatB. The derived amino acidsequence and predicted secondary structure (Fig. 2) of MbFatBthioesterase mature peptide were found to be highly homolo-gous with G. mangostana FatB1 (GmFatB1), which has beenreported to be preferably C16:0-ACP specific with some activ-ities towards C18:0-ACP and C18:1-ACP [14]. Earlier studieshave shown that, even though the amino acid sequences ofUcFatB1 and CcFatB1 are 91% identical, the former isC12:0-ACP specific while the later one is specific for C14:0-ACP [34]. The MbFatB shares 77%, 57% and 56% amino acididentity with GmFatB1, CcFatB1 and UcFatB1, respectively.Although MbFatB differs significantly in amino acid sequencefrom the representative of three different substrate specificFatB thioesterases, its predicted secondary structure is highlyconserved with that of GmFatB1, CcFatB1 and UcFatB1(Fig. 2), showing 84%, 76% and 81% homology, respectively.There are several amino acids (similar or dissimilar replace-ment with other three FatBs) uniquely present in MbFatB(Fig. 2), leading to such a secondary structure of this thioester-ase, which resulted in more closeness in some regions to any ofthe three FatBs having diverse substrate specificity.
Although there is report of 8–10 copies of FatA gene inBrassica napus [19] and one gene of FatB exists in Arabidop-sis [10]; we found M. butyracea genome contains only singlecopy of MbFatB gene as evident from Southern analysis(Fig. 3). The expression of this gene product could be easilydetected in seed tissues of M. butyracea but not in that ofIndian mustard (B. juncea) (Fig. 4). Presence of two polypep-tide bands (Fig. 4 lanes 3 and 4) in seed sample ofM. butyracea was also noted in our earlier report of enzymati-cally active thioesterase sample preparation [2]. Finding ofmore than one thioesterase polypeptide in seed protein couldbe due to the presence of isoforms or different N-terminal clea-
vage or partial proteolysis activity; and has been noted earlierby several other groups for Arabidopsis and Garcinia thioes-terase [10,14]. Our previous study on in vitro determination ofa heat-stable enzyme activity and substrate specificity ofM. butyracea seed-specific thioesterase, combined with thepresent finding of seed-specific expression of the clonedMbFatB gene demonstrate that we have successfully clonedthe novel thioesterase gene from a non-traditional oil-seedtree of sub-Himalayan region. Additionally, we have alsoshown that this type of FatB thioesterase is absent (Fig. 4lanes 1 and 2) or poorly expressed in seed tissues of oil-seedBrassica, which predominantly expresses FatA thioesterase, ashas been documented in earlier studies [19,22].
For functional characterization of M. butyracea thioesterasein vivo, we expressed the MbFatB gene in various E. colistrains using different promoters and fusion tags to influencesolubility and functionality of the recombinant protein. High-level expression of MbFatB driven by T7 promoter in E. coliBL21-CodonPlus-RIL, produced recombinant thioesterase asinclusion bodies (Fig. 5A). However, this insoluble hexa-histi-dine-tagged MbFatB, expected to be functionally inactive as inother cases [29,32], was used to raise polyclonal antibody inrabbit.
For obtaining the soluble recombinant MbFatB, the genewas expressed by tac promoter in E. coli TB1 cells as MBP-tagged MbFatB (Fig. 5B). In contrast to control culture, IPTG-induced expression of MbFatB resulted drastic reduction ofbacterial growth rate (Fig. 6). This metabolic burden of theexpressed MbFatB on bacterial growth could be due to severalreasons, such as accumulation of foreign protein or alterationof lipid metabolism in E. coli by plant thioesterase as notedearlier also [30]. Moreover, we could find that there arechanges in the cell–cell association pattern of those bacteriawhere growth rate of the culture is retarded as evident fromthe light microscopy of bacterial cells expressing the solublerecombinant MbFatB (Fig. 7). Thus, functionality of theMbFatB in heterologous system was evident from the growthbehavior and altered cell morphology of E. coli expressingactive plant thioesterase. This is related to the new fatty acidprofile generated in the bacteria by recombinant MbFatBthioesterase and reflected in the altered fatty acid compositionof the bacterial membrane lipid as discussed below.
It has been noted earlier by different researchers that func-tional expression of plant thioesterase in bacteria changes thefatty acid profile significantly [10,16,27,30] in the membraneand or culture supernatant depending upon the substrate speci-ficity of the thioesterase and the genetic background of theE. coli strains used as heterologous host.
Upon over-expression of the MbFatB gene by strong pro-moter, tac, in E. coli TB1 (fadD+) cells, we found fatty acidprofile of the membrane lipid shifted more towards C16:0 andC18:1 (Fig. 8A). Enhancement of C16:0 and C18:1 fatty acidsis in conformity with our previous study [2] demonstratingdual-substrate (C16:0-ACP and C18:1-ACP) specificity ofthioesterase purified from M. butyracea seeds carried out invitro. It should be noted here that E. coli C18:1(C18:1Δ11cis)

J.K. Jha et al. / Plant Physiology and Biochemistry 44 (2006) 645–655652
is structurally different from plant C18:1(C18:1Δ9cis). The sig-nificant increase in unsaturated fatty acids (Fig. 8, grouped as‘others’ with C12:0, C14:1, cyclopropane fatty acid, C18:2 andC18:3) revealed from Fig. 8A is not unusual as the bacterialculture was grown at low temperature (20 °C). In order tomaintain the membrane fluidity, increase in the amount ofC18:1 was partially nullified by the consequent increase inother unsaturated fatty acids as has been reported earlier also[20]. Increase in the content of several fatty acids in bacterialmembrane lipid is compensated by concomitant decrease inC14:0 and C16:1 amount (Fig. 8A) due to more carbon fluxtowards C16:0-ACP and subsequent acyl chain terminationpreferably at C16:0-ACP by MbFatB before the desaturationstep.
In contrast to the above situation, C16:0 and C18:1 fattyacid level of the membrane lipid of fadD88 mutant expressingMbFatB had been decreased significantly compared to control(Fig. 8B) and the profile is in agreement with the nature oflipid metabolism of the E. coli mutant defective in β-oxidation. This discrepancy between fadD+/MbFatB andfad88/MbFatB could be due to the fact that although C16:0and C18:1 were produced in a significant extent in free formdue to the activity of MbFatB, these free fatty acids are knownnot to serve as the substrates for membrane lipid biosynthesisof E. coli fadD88 strain [21]. On the other hand, in fadD88/MbFatB culture supernatant, C16:0 fatty acid increased signif-icantly along with augmentation of C18:1 (Fig. 8C).
It is known that C16:0 is the predominant fatty acid in themembrane phospholipid of any E. coli cell grown at 30 °C andincreasing the C16:0 fatty acid content beyond a thresholdlevel is extremely difficult as this can alter the membrane fluid-ity and affect bacterial growth. However, drastic enhancementof C12:0 has been achieved through expression of C12:0-ACPspecific plant thioesterase [30], as this fatty acid is the minorcomponent of bacterial membrane. On the other hand, it is pos-sible to accumulate significantly any fatty acid released(depending upon the substrate specificity of thioesterase) intothe culture medium as has been demonstrated in other cases[10,16,30].
A clearer picture of MbFatB activity in vivo was foundwhen the gene expression was driven by lac, a normal promo-ter of E. coli, where a smaller (only seven amino acid fromlacZ α-fragment) tagged-thioesterase was produced at lowlevel yet mostly in active form, unlike the situation with stron-ger tac promoter expression system, in which a larger MBP-tagged recombinant thioesterase produced partly in insoluble(inactive) form. In fadD+/MbFatB cells, the C14:0 and C16:0fatty acids were increased in considerable amount in both themembrane lipid (Fig. 8D) and culture supernatant (Fig. 8E).However, to our surprise, we found approximately three andsixfold increment of C14:0 in the same membrane and super-natant, respectively (Fig. 8D, E). This dramatic increment inC14:0 level indicated that the MbFatB might have specificityas well towards C14:0-ACP and this has been reflected only inthese two samples of DH10B (fadD+) cells because the respec-tive controls have very less amount (~3–4%) of C14:0 fatty
acid (Fig. 8D, E). All other control samples had very high(~20–26%) background level of this particular fatty acid andthe effect of MbFatB activity on C14:0-ACP could not appar-ently observed. Moreover, in fadD88/MbFatB culture superna-tant, predominantly C16:0 fatty acid was found because thecarbon flux during fatty acid synthesis was terminated byMbFatB preferentially at C16:0-ACP and the free fatty acidreleased into the medium (Fig. 8F).
Our previous report on enzymatic assay in vitro of the pur-ified native thioesterase from seed tissues of M. butyraceademonstrated almost equal substrate specificity towardsC16:0-ACP and C18:1-ACP [2]. In the present study, predomi-nant increase in C16:0, along with significant enhancement ofC14:0 and C18:1 content of either membrane lipid or mediumfatty acid pool in various conditions of heterologous expres-sion, is the most prominent feature in E. coli expressing func-tional MbFatB thioesterase. This apparent discrepancy in spe-cificity of M. butyracea thioesterase between the two studiescould be possibly due to the cognate substrate available (acyl-ACPs supplied in vitro vs. synthesized de novo) or severalother reasons as has been suggested by Salas and Ohlrogge[23]. Realizing to the facts that we did not check the substratespecificity of MbFatB on C14:0-ACP earlier [2] and a smallpart of acyl-ACP pool in E. coli expressing functional MbFatBcan further participate in chain-elongation process beyondC16:0-ACP, which is the most preferred substrate for acylchain termination by this plant thioesterase, the present find-ings reconfirm and extend the previous observation.
In conclusion, we could clone the cDNA corresponding tothe mature polypeptide of FatB thioesterase from the develop-ing seed tissues of M. butyracea, a non-traditional oil-seed treeand characterize functionally the gene product by heterologousexpression in E. coli. We are now engaged in the process ofdeveloping transgenic Indian mustard (B. juncea), a widelycultivated oil-seed crop for reducing its erucic acid content byearly termination of fatty acyl chain-elongation with this newthioesterase. We believe that will open up diverse utility of theseed oil in future.
4. Materials and methods
4.1. Plant material
Seeds of Diploknema (Madhuca) butyracea, a sub-Himalayan plant species were collected from the trees located1000 m above the sea level from Darjeeling district, in India.Seed kernels collected from immature fruits of 57–65 daysafter flowering [2] and immediately frozen in liquid N2, kept at–80 °C freeze until use.
4.2. Preparation of mRNA and synthesis of first strand cDNA
Total cellular RNA was isolated from frozen seed tissuesusing hot phenol extraction method [28]. The RNA samplewas treated with RNase-free DNase I (Boehringer Mannheim)

J.K. Jha et al. / Plant Physiology and Biochemistry 44 (2006) 645–655 653
and PolyA+ RNAs was purified from total RNA preparationfollowing the standard method [1]. The first strand cDNAwas synthesized using a modified oligo-dT primer[5′-GCGGCCACGGATCCGTCGAC(T)15VN-3′] and reversetranscriptase Superscript-II RT (GIBCO-BRL) followingmanufacturer’s instructions.
4.3. Isolation, cloning and sequencing of MbFatB gene
For isolation of a part of the thioesterase gene, we designedtwo degenerate oligonucleotide primers—FatN [5′-CN TCNRWN TGG GYN ATG ATG AA-3′] and FatC [5′-AC RTTRTT NAC RTG YTK RTT-3′], corresponding to the aminoacid sequences (T/S)S(K/V)W(V/A)MMN and N(Q/K)HVNNV that are conserved amongst the two members ofFatA and FatB sequences in many plant species (Fig. 1). ThePCR product generated from the seed-specific cDNA sampleusing these two primers resulted a DNA band of ~220 bp,which was cloned in pUC18 by blunt-end ligation and severalrandomly selected clones were sequenced double-strand fol-lowing Sanger’s dideoxy method using T7 Sequenase kit(USB). Since, bioinformatic analysis revealed that this partialcDNA is highly homologous to reported FatB class of plantthioesterases, a new degenerate primer FatN0 [5′-ggtctagatctATH AAY CAR YTN CCN GAY TGG-3′] was designedbased on the N-terminal conserved region INQLPDW, foundin Arabidopsis thaliana (Z36911), Cuphea hookeriana(U17076), Garcinia mangostana (U92878), Gossypium hirsu-tum (AF076535) and Helianthus annuus (AF036565). Here,Leu (bold) is the first amino acid of mature peptide proposedby Jones et al. for ChFatB1 [17] and Dormann et al. forAtFatB1 [10]. Next, two gene-specific primers—GSP1 [5′-AAGAGACACGGAGGTTATCTA-3′] and GSP2 [5′-GTTGACATGTTGGTTGACATC-3′] were designed from the knownsequence of the gene fragment (Fig. 2) for subsequent 3′-RACE and 5′RT-PCR using the same first strand cDNA sam-ples. The primer pairs—GSP1 and RACE primer [5′-GCGGCCACGGAT CCGTCGAC-3′] yielded a DNA bandof ~645 bp for 3′-RACE, where as GSP2 and FatN0 primerresulted ~708 bp product for 5′-RT-PCR. The DNA fragmentswere cloned separately in pUC18 by blunt-end ligation andrandomly selected clones from both events were sequenced.The near-full length cDNA (without 5′-UTR) of 1218 bp wasre-cloned in pBlueScript using the HindIII site present in theoverlapping fragments of 5′ and 3′ part of the gene. TheM. butyracea thioesterase gene was named as MbFatB andthe recombinant plasmid designated as pBS/MbFatB. TheDNA sequence analyses were carried out by BLAST searchand Bioware Jellyfish software.
4.4. Southern hybridization
The genomic DNA was isolated from seed tissue ofM. butyracea following cetyltriethyl ammonium bromide(CTAB) extraction method as described by Doyle and Doyle[12]. Ten micrograms of genomic DNA was digested with dif-
ferent restriction enzymes and electrophoresed on 0.8% agar-ose gel followed by transfer onto nylon membrane (Amer-sham) according to Sambrook et al. [24]. The blot washybridized with 708 bp 3′ portion of the MbFatB gene labeledwith (32P)dCTP and washing was performed according to Sam-brook et al. [24].
4.5. Recombinant MbFatB expression in E. coli
The 1.1 kb BglII–XhoI DNA fragment encoding the maturepolypeptide of MbFatB gene from pBS/MbFatB was re-clonedat the same sites of pRSET-B (T7 promoter based vector, Invi-trogen), generating pRSET/MbFatB. The DNA sequence of thecorrect reading frame of the His-tagged fusion was confirmedby sequencing. The control plasmid and the recombinant onewere transformed into E. coli BL21-CodonPlus-RIL (Strata-gene). The same gene as BglII–HindIII (partial) fragmentfrom pBS/MbFatB was re-cloned in pMAL-c2E (tac promoterbased vector, New England Biolab) using BamHI and HindIIIsites of the vector; and resulted the recombinant pMAL/MbFatB. A set of control and recombinant plasmids was trans-formed into E. coli strains TB1 (New England Biolab) andK27fadD88, a fatty acid β-oxidation mutant [21], obtainedfrom Dr. K.D. Chapman (Department of Biological Science,Division of Biochemistry and Molecular Biology, Universityof North Texas, Denton, USA).
E. coli BL21-CodonPlus-RIL cells harboring the controlplasmid pRSET-B and pRSET/MbFatB were grown at 30 °Cin LB broth containing 200 μg/ml ampicillin and 30 μg/ml ofchloramphenicol. Cells were induced at OD600 0.6 with0.4 mM IPTG and growth was continued for next 4 hours.Induction experiments were also performed with the same cul-tures at lower temperature (20 °C) and IPTG concentration(0.1 mM). Similar expression studies were performed withthe cultures of TB1 cells transformed with pMAL-c2E andpMAL/MbFatB, except that LB media contained only200 μg/ml ampicillin.
To express the MbFatB gene under lac promoter of pUC18in frame with the N-terminal of lacZ α-fragment, we havedesigned two gene-specific primers—GSP3 [5′-atggaattcgATGATCAACCAGCTTCCGGACT-3′] and GSP4 [5′-cagatc-tagaTTAAGCACTCTCTGCTGGAAGC-3′] (Fig. 2). The PCRproduct was cloned as EcoRI–XbaI fragment at the same sitesgenerating pUC/MbFatB, and the correct reading frame of thefusion gene with the expected tag of seven amino acids fromlacZ α-fragment was confirmed by DNA sequencing. A set ofcontrol and recombinant plasmids was transformed into E. coliDH10B and K27fadD88 cells, grown in LB media containing500 μg/ml ampicillin to prevent rescuing of the recombinantplasmid. The respective cultures were induced at an OD600
0.6 with 0.4 mM IPTG.
4.6. Antibody production and immunoblotting
Two hundred and fifty milliliters culture of pRSET/MbFatBin E. coli strain BL21-CodonPlus-RIL cell was induced as

J.K. Jha et al. / Plant Physiology and Biochemistry 44 (2006) 645–655654
mentioned above. The harvested cell pellet was resuspended insolubilization buffer (50 mM Tris–HCl, pH 8.0), and was dis-rupted by sonication. Almost all of the recombinant proteinwas accumulated as inclusion bodies, which were collectedby centrifugation at 10,000 × g for 10 min, dissolved in 1×his binding buffer (20 mM Tris–HCl, pH 7.5, 5 mM imidazole,500 mM NaCl). The purification of recombinant protein wascarried out using Ni–NTA (Ni–nitrilotriacetic acid) spin col-umn (Qiagen) under denatured condition as per manufacturer’sinstruction. Two hundred and fifty micrograms of the purifiedprotein was used to raise polyclonal antibody against MbFatB.
Total protein from the seeds of M. butyracea and B. junceawere extracted by homogenizing the seed tissues in a buffercontaining 50 mM potassium phosphate (pH 6.9), 5 mMEDTA, 1 mM sodium ascorbate, 2 mM 2-marcaptoethanoland 1 mM PMSF. The sample was sonicated followed by cen-trifugation at 15,000×g for 10 min. The clarified solubleextract was loaded in 10% SDS-PAGE, blotted onto PVDFmembrane and probed with MbFatB antiserum at 1:2500 dilu-tions. Rest of the steps of Alkaline Phosphatase method wasperformed following the instruction manual of BM Chromo-genic Western Blotting Kit (Roche Applied Science).
4.7. Lipid extraction and fatty acid methyl ester (FAME)analysis
Total lipid was extracted from the bacterial cell pellet usingthe modified protocol of Bligh and Dyer [3]. The wet cell pasteof 50 mg was diluted with 1 ml of deionized water and to thatsuspension 3.75 ml of chloroform/methanol (1:2, v/v) wasadded. The mixture was shaken for several hours with intermit-tent shaking. After centrifugation at 6000×g for 5 min thesupernatant extract was decanted in another closed vial, theresidue was resuspended in 4.75 ml of methanol/chloroform/water (2:1:0.8, v/v). The mixture was then shaken and centri-fuged at 10,000×g for 10 min. Two supernatants were mixedtogether, and to the resultant supernatant 2.5 ml each of metha-nol and chloroform added and centrifuged at 10,000×g for10 min. Lower chloroform layer was withdrawn and broughtto dryness in a rotary evaporator. The residue was dissolved in5 ml of dry methanol containing 5% concentrated H2SO4. Thismixture was then refluxed at 90 °C for 2 h. Thereafter, themethanol layer was evaporated in rotary evaporator; the residuewas dissolved in standard saline water followed by shakingwith n-hexane. The hexane layer was evaporated to dryness.TLC was performed with that lipid extract in a solvent mixturecontaining hexane/diethyl ether/acetic acid (80:20:1.5, v/v/v).Finally the FAME band was excised and eluted with hexane.The small FAME eluate was subjected to gas liquid chromato-graphy (GLC) using a DEGS column in Hewlett Packardinstrument (model 5840A, Hewlett Packard Inc., Palo Alto,CA, USA). The flame ionization detection (FID)-temperaturewas 200 °C and the operating temperature was maintained at180 °C.
Lipid from the bacterial culture supernatant was extractedfollowing the protocol of Voelker and Davies [30], and
FAME analysis was performed as mentioned above. Data pre-sented in this study are the average of the three experiments ineach case.
Acknowledgements
We sincerely thank Dr. K.D. Chapman for providing theE. coli K27fadD88 strain used in this study. The authors aregrateful to Professor Sudhamoy Ghosh for critical commentsand suggestions. Financial assistance in the form of researchfellowship to JKJ from University Grant Commission, Govern-ment of India and grant support to this laboratory from theDepartment of Biotechnology, Government of India areacknowledged.
References
[1] J. Allen, Purification and fractionation of Poly(A)+ RNA, in, MethodsEnzymol. 152 (1987) 254–261.
[2] A. Bhattacharjee, S.K. Ghosh, D. Ghosh, S. Ghosh, M.K. Maiti, S.K.Sen, Identification of a heat-stable palmitoyl/oleoyl acyl-acyl carrier pro-tein thioesterase in developing seeds of Madhuca butyracea, Plant Sci.163 (4) (2002) 791–800.
[3] E.G. Bligh, W.J. Dyer, A rapid method of total lipid extraction and pur-ification, Can. J. Biochem. Physiol. 37 (1959) 911–917.
[4] G. Bonaventure, J.J. Salas, M.R. Pollard, J.B. Ohlrogge, Disruption ofFATB gene in Arabidopsis demonstrates an essential role of saturatedfatty acids in plant growth, Plant Cell 15 (2003) 1020–1033.
[5] C. Combet, C. Blanchet, C. Geourjon, G. Deléage, NPS@: Network Pro-tein Sequence Analysis, Trends Biochem. Sci. 25 (3) (2000) 147–150[291].
[6] H.M. Davies, Medium chain acyl-ACP hydrolysis activities of develop-ing oilseeds, Phytochemistry 33 (1993) 1353–1356.
[7] H.M. Davies, L. Anderson, C. Fan, D.J. Hawkins, Developmental induc-tion, purification, and further characterization of C12:0-ACP thioesterasefrom immature cotyledons of Umbellularia californica, Arch. Biochem.Biophys. 290 (1991) 37–45.
[8] K. Dehesh, A. Jones, D.S. Knutzon, T.A. Volelker, Production of highlevels of 8:0 and 10:0 fatty acids in transgenic canola by over expressionof ChFatB2, a thioesterase cDNA from Cuphea hookeriana, Plant J. 9(1996) 167–172.
[9] P. Dörmann, F. Spener, J.B. Ohlrogge, Characterization of two acyl-acylcarrier protein thioesterases from developing Cuphea seeds specific formedium-chain and oleoyl-acyl carrier protein, Planta 189 (1993) 425–432.
[10] P. Dörmann, T.A. Voelker, J.B. Ohlrogge, Cloning and expression inEscherichia coli of a novel thioesterase from Arabidopsis thaliana speci-fic for long-chain acyl-acyl carrier proteins, Arch. Biochem. Biophys.316 (1995) 612–618.
[11] P. Dormann, T.A. Voelker, J.B. Ohlrogge, Accumulation of palmitate inArabidopsis mediated by acyl-acyl carrier protein thioesterase FATB1,Plant Physiol. 123 (2000) 637–643.
[12] J.J. Doyle, J.L. Doyle, Isolation of plant DNA from fresh tissue, BRLFo-cus 12 (1990) 13–15.
[13] M.T. Facciotti, P.B. Bertain, L. Yaun, Improved stearate phenotype intransgenic canola expressing a modified acyl-acyl carrier protein thioes-terase, Nat. Biotechnol. 17 (1999) 593–597.
[14] D.J. Hawkins, J.C. Kridl, Characterization of acyl-ACP thioesterase ofmangosteen (Garcinia mangostana) seed and high level of stearate pro-duction in transgenic canola, Plant J. 13 (1998) 743–752.
[15] M.J. Hills, Improving oil functionality by tuning catalysis of thioesterase,Trends Plant Sci. 4 (1999) 421–422.

[16
[17
[18
[19
[20
[21
[22
[23
[24
[25
[26
[27
[28
[29
[30
[31
[32
[33
[34
J.K. Jha et al. / Plant Physiology and Biochemistry 44 (2006) 645–655 655
] T.T. Huynh, R.M. Pirtle, K.D. Chapman, Expression of a Gossypium hir-sutum cDNA encoding a FatB palmitoyl-acyl carrier protein thioesterasein Escherichia coli, Plant Physiol. Biochem. 40 (2002) 1–9.
] A. Jones, H.M. Davies, T.A. Voelker, Palmitoyl-acyl carrier protein(ACP) thioesterase and the evolutionary origin of plant acyl-ACP thioes-terases, Plant Cell 7 (1995) 359–371.
] M.M. Kimberly, J. Shanklin, A structural model of the plant acyl-acylcarrier protein thioesterase FatB comprises two helix/4-stranded sheetdomains, the N-terminal domain containing residues that affect specifi-city and the C-terminal domain containing catalytic residues, J. Biol.Chem. 280 (2005) 3621–3627.
] N.M. Loader, E.M. Woolner, A. Hellyer, A.R. Salbas, R. Safford, Isola-tion and characterization of two Brassica napus embryo acyl-ACPthioesterase cDNA clones, Plant Mol. Biol. 23 (1993) 769–778.
] A.G. Marr, J.L. Ingraham, Effect of temperature on the composition offatty acids in Escherichia coli, J. Bacteriol. 84 (1962) 1260–1267.
] P. Overath, G. Pauli, H.U. Schairer, Fatty acid degradation in Escheri-chia coli: an inducible acyl-CoA synthetase, the mapping of old muta-tions, and the isolation of regulatory mutants, Eur. J. Biochem. 7 (1969)559–574.
] M.K. Pathak, A. Bhattacharjee, D. Ghosh, S. Ghosh, Acyl-acyl carrierprotein (ACP)-thioesterase from developing seeds of Brassica campestriscv. B-54 (Agrani), Plant Sci. 166 (2004) 191–198.
] J.J. Salas, J.B. Ohlrogge, Characterization of substrate specificity of FatAand FatB acyl-ACP thioesterases, Arch. Biochem. Biophys. 403 (2002)25–34.
] J. Sambrook, E.F. Fritsch, T. Maniatis, Molecular Cloning: A LaboratoryManual, second ed, Cold Spring Harbor Laboratory, Cold Spring Harbor,NY, 1989.
] A. Sengupta, S.K. Roy Choudhury, Triglyceride composition of Mad-huca butyracea seed fat, J. Am. Oil Chem. Soc. 55 (1978) 621–624.
] M.J. Serrano-Vega, R. Garcés, E. Martinez-Force, Cloning, characteriza-tion and structural model of a FatA-type thioesterase from sunflowerseeds (Helianthus annuus L.), Planta 221 (2005) 868–880.
] M.J. Serrano-Vega, M. Venegas-Calerón, R. Garcés, E. Martinez-Force,Cloning and expression of fatty acids biosynthesis key enzymes fromsunflower (Helianthus annuus L.) in Escherichia coli, J. Chromatogr.B. 786 (2003) 221–228.
] T.C. Verwoerd, B.M.M. Dekker, A. Hoekema, A small-scale procedurefor the rapid isolation of plant RNAs, Nucleic Acids Res. 17 (1989)2362.
] A. Villaverde, M.M. Carrio, Protein aggregation in recombinant bacteria:biological role of inclusion bodies, Biotechnol. Lett. 25 (2003) 1385–1395.
] T.A. Voelker, H.M. Davies, Alteration of the specificity and regulationof fatty acid synthesis of Escherichia coli by expression of a plantmedium-chain acyl-acyl carrier protein thioesterase, J. Bacteriol. 176(1994) 7320–7327.
] T.A. Voelker, A. Jones, A.M. Cranmer, H.M. Davies, D.S. Knutzon,Broad-range and binary-range acyl-acyl carrier protein thioesterase sug-gest an alternative mechanism for medium chain production in seeds,Plant Physiol. 114 (1997) 669–677.
] E.A. Woestenenk, M. Hammarström, S. van den Berg, T. Härd, H. Ber-glund, His tag effect on solubility of human proteins produced in Escher-ichia coli: a comparison between four expression vectors, J. Struct.Funct. Genomics 5 (2004) 217–229.
] L. Yuan, B.A. Nelson, G. Caryl, The catalytic cysteine and histidine inthe plant acyl-acyl carrier protein thioesterases, J. Biol. Chem. 271(1996) 3417–3419.
] L. Yuan, T.A. Voelker, D.J. Hawkins, Modification of substrate specifi-city of an acyl-acyl carrier protein thioesterase by protein engineering,Proc. Natl. Acad. Sci. USA 92 (1995) 10639–10643.




















