
Clonal propagation and production of cichoric acid in three species of Echinaceae
10
PLANT TISSUE CULTURE Clonal propagation and production of cichoric acid in three species of Echinaceae Anca-Livia Butiuc-Keul & Laurian Vlase & Cornelia Crăciunaş Received: 28 July 2011 / Accepted: 6 March 2012 / Published online: 23 March 2012 / Editor: J. Forster # The Society for In Vitro Biology 2012 Abstract In vitro tissue culture protocols were tested for propagation of Echinacea purpurea, Echinacea pallida and Echinacea angustifolia in order to obtain biomass for the production of cichoric acid, which is the major active com- pound in the Echinacea extracts. The in vitro culture process was initiated by seed germination on half-strength Murashige and Skoog (MS) medium. Multiplication was achieved on MS medium supplemented with naphthaleneacetic acid (NAA), indole-3-butyric acid (IBA), 2-iso-pentenyladenine (2iP), and N 6 -benzyladenine (BA) in different concentrations. Shoot explants produced the highest number of shootlets on MS medium, which was supplemented with 0.1 mg/l 2iP and 0.1 mg/l IBA. RAPD markers revealed genetic polymorphism in some instances between in vitro generated plantlets such as for E. purpurea plantlets analyzed with the OPO-8 primer. RAPD markers generated with the primer 4A-29 revealed low levels of genetic variation between in vitro plantlets for all three species of Echinacea, while remaining RAPD markers revealed no variation. Content of cichoric acid in leaves, shoots, and callus was analyzed by high-performance liquid chromatography/MS and was identified in all studied samples, independent of species or tissue type. Highest levels (0.39– 0.73 mg/g dw) were observed in shoots and leaves. Keywords Echinacea . Multiplication . Secondary metabolites . Cichoric acid . Medicinal plant Introduction Plant species have provided important medicinal remedies for thousands of years. The rapid growth of the marketplace for plant-based health products has been associated with an in- creased variety of medicinal plant preparations and with a continuous broadening of the categories driven by consumer interest in such products. Even today, the World Health Orga- nization estimates that up to 80 % of people still mainly rely on traditional remedies such as herbs for their medicines (Baum et al. 2006). During the 1990s, popularity of the genus Echinacea Moench (Asteraceae family) as a dietary supple- ment increased markedly, as the general public learned of its possible efficacy for fighting colds and other illnesses (Baum et al. 2006). Echinacea, also referred to as purple coneflower, is geographically confined to North America and is distributed in dry prairies from Texas to Saskatchewan and from West of the Rocky mountains to Minnesota (Binns et al. 2004). The major producers of Echinacea in Europe are in Germany, Switzerland, The Netherlands, Italy, and Spain (Galambosi 2004). In 1998, Echinacea was the tenth most important medicinal plant sold in Europe with annual sales of about $120 USD million. In North America, Echinacea ranked second in sales in the mainstream market in 2004. Retail sales of Echinacea products are worth more than $158 million annually in the USA, and worldwide, they have been estimat- ed at $1,300 million annually (Blumenthal 2005). Three species of Echinacea are generally used in medic- inal plant preparations (Perry et al. 2001): Echinacea pur- purea Moench (roots and shoots), Echinacea angustifolia DC (roots), and Echinacea pallida Nutt (roots). Echinacea A.-L. Butiuc-Keul (*) : C. Crăciunaş Department of Molecular Biology and Biotechnology, Babeş-Bolyai University, 400084, Cluj-Napoca, Romania e-mail: [email protected] L. Vlase Faculty of Pharmacy, Department of Pharmaceutical Technology and Biopharmaceutics, University of Medicine and Pharmacy “Iuliu Hatieganu”, 400023, Cluj-Napoca, Romania In Vitro Cell.Dev.Biol.—Plant (2012) 48:249–258 DOI 10.1007/s11627-012-9435-2
Transcript of Clonal propagation and production of cichoric acid in three species of Echinaceae

PLANT TISSUE CULTURE
Clonal propagation and production of cichoric acidin three species of Echinaceae
Anca-Livia Butiuc-Keul & Laurian Vlase &
Cornelia Crăciunaş
Received: 28 July 2011 /Accepted: 6 March 2012 /Published online: 23 March 2012 / Editor: J. Forster# The Society for In Vitro Biology 2012
Abstract In vitro tissue culture protocols were tested forpropagation of Echinacea purpurea, Echinacea pallida andEchinacea angustifolia in order to obtain biomass for theproduction of cichoric acid, which is the major active com-pound in the Echinacea extracts. The in vitro culture processwas initiated by seed germination on half-strength Murashigeand Skoog (MS) medium.Multiplication was achieved onMSmedium supplemented with naphthaleneacetic acid (NAA),indole-3-butyric acid (IBA), 2-iso-pentenyladenine (2iP), andN6-benzyladenine (BA) in different concentrations. Shootexplants produced the highest number of shootlets on MSmedium, which was supplemented with 0.1 mg/l 2iP and0.1 mg/l IBA. RAPDmarkers revealed genetic polymorphismin some instances between in vitro generated plantlets such asfor E. purpurea plantlets analyzed with the OPO-8 primer.RAPDmarkers generated with the primer 4A-29 revealed lowlevels of genetic variation between in vitro plantlets for allthree species of Echinacea, while remaining RAPD markersrevealed no variation. Content of cichoric acid in leaves,shoots, and callus was analyzed by high-performance liquidchromatography/MS andwas identified in all studied samples,independent of species or tissue type. Highest levels (0.39–0.73 mg/g dw) were observed in shoots and leaves.
Keywords Echinacea . Multiplication . Secondarymetabolites . Cichoric acid .Medicinal plant
Introduction
Plant species have provided important medicinal remedies forthousands of years. The rapid growth of the marketplace forplant-based health products has been associated with an in-creased variety of medicinal plant preparations and with acontinuous broadening of the categories driven by consumerinterest in such products. Even today, the World Health Orga-nization estimates that up to 80 % of people still mainly relyon traditional remedies such as herbs for their medicines(Baum et al. 2006). During the 1990s, popularity of the genusEchinacea Moench (Asteraceae family) as a dietary supple-ment increased markedly, as the general public learned of itspossible efficacy for fighting colds and other illnesses (Baumet al. 2006). Echinacea, also referred to as purple coneflower,is geographically confined to North America and is distributedin dry prairies from Texas to Saskatchewan and from West ofthe Rocky mountains to Minnesota (Binns et al. 2004). Themajor producers of Echinacea in Europe are in Germany,Switzerland, The Netherlands, Italy, and Spain (Galambosi2004). In 1998, Echinacea was the tenth most importantmedicinal plant sold in Europe with annual sales of about$120 USD million. In North America, Echinacea rankedsecond in sales in the mainstream market in 2004. Retail salesof Echinacea products are worth more than $158 millionannually in the USA, and worldwide, they have been estimat-ed at $1,300 million annually (Blumenthal 2005).
Three species of Echinacea are generally used in medic-inal plant preparations (Perry et al. 2001): Echinacea pur-purea Moench (roots and shoots), Echinacea angustifoliaDC (roots), and Echinacea pallida Nutt (roots). Echinacea
A.-L. Butiuc-Keul (*) : C. CrăciunaşDepartment of Molecular Biology and Biotechnology,Babeş-Bolyai University,400084, Cluj-Napoca, Romaniae-mail: [email protected]
L. VlaseFaculty of Pharmacy,Department of Pharmaceutical Technology and Biopharmaceutics,University of Medicine and Pharmacy “Iuliu Hatieganu”,400023, Cluj-Napoca, Romania
In Vitro Cell.Dev.Biol.—Plant (2012) 48:249–258DOI 10.1007/s11627-012-9435-2

species have been increasingly studied for different applica-tions in biotechnology, especially in vitro culture and prep-aration of natural extracts (Abbasi et al. 2007). Plant andmedical scientists responded to this demand by concentrat-ing their efforts to understand the biology, cultivation, andpharmacology of these plants. Echinacea species, especiallyE. purpurea contain a variety of chemical compounds (216different medicinally active compounds) that are structurallydiverse (for example, cichoric acid, alkylamides, echinaco-side, cinarine, and polysaccharides) and perform multipleactivities that are beneficial for health (Murch et al. 2006).Known phenolic compounds in Echinacea species includecaffeic acid derivatives such as cichoric acid in E. purpureaand E. pallida, and echinacoside in E. angustifolia(Harborne and Williams 2004). Cichoric acid is the mainactive compound of Echinacea extract. It is known that natu-ral extracts ofEchinacea have significant immunomodulatory,antibacterial, antiviral, antifungal properties (Barrett 2003),and also antioxidant activities (Thygesen et al. 2007).
Commercial production ofEchinacea has been limited by arange of issues including contamination of plant materials bymicroorganisms, pollution from the environment, variabilityof active components, and lack of pure, standardized plantmaterial for biochemical analyses (Raman et al. 2004). Toaddress these issues, in vitro tissue culture techniques forEchinacea represent a desirable alternative to harvestingplants from natural or cultivated habitats. Echinacea specieshave been regenerated from a range of tissue types varyingfrom in vitro seedlings to mature field-grown plants, with anumber of studies having described the biomass productionby in vitro culture of Echinacea species (Lata et al. 2004). Inaddition, several studies have focused on the isolation andcharacterization of different classes of compounds responsiblefor the multiple activities of Echinacea extracts (Currier andMiller 2000; Wills and Stuart 2000; Merali et al. 2003).
In the present study, an improvedmethod is reported for thein vitro multiplication of E. purpurea, E. pallida, andE. angustifolia. This study is part of a broader research planaimed at development of alternative methods (such as in vitroculture) for harvesting of plants from natural and cultivatedhabitats (Butiuc-Keul and Deliu 2001; Butiuc-Keul et al.2002). In addition to the in vitro multiplication of the threeEchinacea species, the amount of cichoric acid in regeneratedplantlets and callus tissue was quantified and randomly am-plified polymorphic DNA (RAPD) markers were used toassess the genetic variability of the in vitro plantlets and toidentify potential somaclonal variants.
Materials and Methods
Plant material and in vitro culture. In vitro cultures wereinitiated from seeds of E. purpurea, E. pallida, and
E. angustifolia and were immersed in 70 % (v/v) ethanolfor 30 s and 15 min immersion in 50 % (v/v) Domestos®
(sodium hypochlorite at 5 % v/v). Seeds were collected fromfield plants from a single batch for each species and wereprovided by Alexandru Borza Botanical Garden, Cluj-Napoca, Romania. After sterilization with Domestos solution,seeds were washed three times (5 min) with sterile, distilledwater prior to placing on medium consisting of half-strengthMurashige and Skoog (MS) salts (Murashige and Skoog1962), 0.1 g/l myo-inositol, 30 g/l sucrose, and agar 0.8 %w/v (micro-agar, Duchefa, Haarlem, The Netherlands).Culture media were sterilized by autoclaving at 121°C and103 kPa (1.1 kg cm−2) for 20min after adjustment of the pH to5.8 with 1 N KOH. Cultivation was done in 100-ml glass jarscontaining 15 ml of medium. Cultures were grown for 30 d atconstant 22°C, at photoperiod regime of 16 h light (cool-whitefluorescent lights, 50 μmol s−1 m−2 photosynthetic photonflux density). Seedlings were placed onMSmedium, and after30 d, 1 cm microcuttings were excised and cultured on MSmedium supplemented 1.0 mg/l N6-benzylaminopurine (BA)and 0.1 mg/l α-naphthaleneacetic acid (NAA) in order toobtain plant material for further experiments. In vitro cultureof Echinacea shoots was previously achieved on MS mediumsupplemented with 1.0 mg/l indole-acetic acid (IAA) and 1.0–5.0 mg/l BA (Taha et al. 2009).
In order to improve the multiplication rate of these species ofEchinacea, in our experiment nodal explants of 1 cm obtainedon MS medium supplemented with 1.0 mg/l BA and0.1 mg/l NAA were cultured on MS medium supplementedwith indole-3-butyric acid (IBA), NAA, 2-iso-pentenyladenine(2iP), or BA in different concentrations as follows: MS1—without growth regulators; MS2—MS+0.1 mg/l BA+0.1mg/l NAA; MS3—MS+1.0 mg/l BA+0.1 mg/l NAA;MS4—MS+0.1 mg/l 2iP+0.1 mg/l IBA; MS5—MS+1.0 mg/l 2iP+1.0 mg/l IBA; and MS6—MS+2.0 mg/l 2iP+0.1 mg/l IBA. Leaf and shoot explants were also placed on MSmedium supplemented with 2.0 mg/l 2,4 dichlorophenoxyace-tic acid (2,4-D)+0.1 mg/l BA, for callus induction.
Cultures were evaluated after 30 d of culture. The numberof shoots and roots/explant and shoot length were measured.Ten replicates were used per treatment, and the experimentswere repeated twice. After 30 d, the shoots from all 10replicates grown on MS4 medium (which proved to besuperior for multiplication) were collected, and fresh weight(fw) was recorded. Plants were dried at room temperature,and after 1 wk, the dry weight (dw) was also measured.Similar biomass calculations were performed for callus.One-way analysis of variance (ANOVA) and Tukey testswere used to confirm differences between means. MiniTabsoftware was used for statistical analysis.
RAPD analysis. Genomic DNA was isolated from leavesusing the cetyl trimethylammonium bromide method as
250 BUTIUC-KEUL, VLASE ET AL.

described by Doyle and Doyle (1987). DNA mplification byPCR was performed using 10 nucleotides primers (OperonTechnologies) and the specific conditions that were previ-ously established in our laboratory (Crăciunaş et al. 2007).Reactions were performed in 0.2-ml Eppendorf tubes con-taining 2 mM MgCl2, 1 μM of each primer, 200 μM of eachdNTP, 1.5 U of Taq (Fermentas), and 25 ng of genomicDNA in a final volume of 25 μl. Steps of the thermocyclingreaction conditions were as follows: (1) T095°C for 5 min,(2) T094°C for 45 s, (3) primer annealing at 36°C for 45 s,(4) elongation T072°C for 1.5 min; steps 2–4 were repeated35 times. A total of 28 random primers were tested, but dataare only presented for six primers which generated the bestresults (Table 1). Amplicons were separated on 1.5 % (w/v)agarose gels and stained with 0.5 μg/ml ethidium bromide.
Quantification of cichoric acid. Analysis of cichoric acidfrom Echinacea extracts was performed using a newly de-veloped procedure of liquid chromatography coupled withmass spectrometry detection. This method provided a high-throughput determination, having a major advantage of rap-id analysis (<1 min per sample), and high specificity due tomass spectrometry detection.
Plant material was dried in vegetation chamber at 22°Cuntil dw was constant. Dried material was finely ground usingmortar and pestle, and 1 g was mixed with 10 ml of 50 % (v/v)methanol and left for 24 h. Samples were then centrifuged at3,000 rpm for 10 min. The supernatant was used for analysisof cichoric acid by high performance liquid chromatography(HPLC) coupled with mass spectrometry detection (Agilent1100 series HPLC system, Agilent Technologies, Darmstadt,Germany), coupled with an Agilent Ion Trap SL mass spec-trometer equiped with an electrospray ion source. A referencestandard of cichoric acid (Dalton Chemical Laboratories, Tor-onto, ON, Canada, batch AI-01-119A-020201) was used forquality control. The stock solution of cichoric acid (1 mg/ml)was prepared by dissolving the appropriate quantity of refer-ence standard in methanol and was used to prepare calibrationstandards with concentrations of 0.75, 1.5, 3.0, 7.5, and
15 μg/ml. The calibration standards were prepared in bidis-tilled water. To verify the precision and accuracy of method,three control standards (0.75, 3.0, and 15 μg/ml) were used. Inbrief, chromatographic separation of cichoric acid wasobtained using a Zorbax SB-C18 100 mm×3.0 mm i.d.,3.5 μm column (Agilent Technologies). The mobile phaseconsisted in 95/5 (v/v) ammonium acetate, 1 mM in waterand acetonitrile, isocratic elution, and mobile phase flow rateof 1 ml/min. The mass spectrometer operated in negativemultiple reaction monitoring mode, using nitrogen as nebulis-ing and dry gas. The nebulizer was set at 65 psi, and the drygas flow was 12 l/min at 350°C. The mass spectrometer wasset to record the transition m/z 473>m/z 293+m/z 311, whichis specific to cichoric acid. The retention time of cichoric acidwas 0.51 min. The concentration of cichoric acid wasdetermined automatically by the instrument data system(QuantAnalysis 1.7 software, Brucker Daltonics, Darmstadt,Germany) using peak area and the external standard method.The calibration curve model was determined by the linearregression, using an 1/Y2 weighting scheme. The lower limitof quantification was established at 0.75 μg/ml. At quantifi-cation limit, the method precision (expressed as coefficient ofvariation, CV %) was 6.8 %, and accuracy (expressed asrelative difference between obtained and theoretical concen-tration, bias %) was 5.3 %, respectively.
Results
Shoot induction and multiplication. In vitro cultures wereestablished by aseptic germination of seeds on half-strengthMSmedium. The rate of seed germination was 80–90 % (datanot shown). Previous studies revealed a high level of infectionof seeds with microorganisms (Mechanda et al. 2003), andseveral researchers have proposed removal of the seed coatlayers to prevent contamination (Harbage 2001). However, asequential process of surface sterilization proved to be highlyefficient in ensuring visually contamination-free seed germi-nation. Surface sterilization of seeds was also accomplishedby other authors (Taha et al. 2009).
Based on ANOVA analysis and the Tukey test, culturemedia was shown to significantly influence both shoot mul-tiplication and root induction rates in all three species ofEchinacea. The best media for shoot multiplication wereMS4 for E. purpurea, for which on average 7.1 shoots/nodalexplant and 4.6 roots/nodal explant were obtained, and MS5for E. pallida (5.4 shoots/nodal explant) and E. angustifolia(3.9 shoots/nodal explant). The highest numbers of shoots/nodal explant were obtained in E. purpurea, then E. pallida,and E. angustifolia. MS4 medium was also superior for rootinduction in E. purpurea (4.6 roots/nodal explant) andE. pallida (4.5 roots/nodal explant). In E. angustifolia, the
Table 1. Characteristics of primers used for DNA amplification
Primername
Nucleotide sequence Number ofnucleotides
sm (°C)
OPE-3 5′’-CCAGATGCAC-3′ 10 37
OPO-4 5′-AAGTCCGCTC-3′ 10 37
OPO-7 5′-CAGCACTGAC-3′ 10 37
OPO-8 5′- CCTCCAGTGT-3′ 10 37
4A-27 5′-CAATCGCCGT-3′ 10 37
4A-28 5′-CAAACGTCGG-3′ 10 37
4A-29 5′-ACGTAGCGTC-3′ 10 37
4A-30 5′-TCAGAGCGCC-3′ 10 37
CLONAL PROPAGATION AND PRODUCTION OF CICHORIC ACID 251

highest level of root induction was obtained on MS2 medium(3.8 roots/nodal explant). Shoot length was similar on MS3,MS4, MS5, andMS6 of the tested media, and plantlets were onaverage 2.1–3.2 cm long, depending on the plant species. TheMS2 medium supplemented with 0.1 mg/l BA+0.1 mg/l NAAensured superior shoot elongation, plantlets being 4.2 cm long
for E. purpurea, 3.6 cm for E. pallida, and 3.4 cm forE. angustifolia. The length of induced roots was similar on allmedia in E. purpurea and E. pallida (on average, 1.3–2.4 cm).For E. angustifolia, the length of induced roots was higher onMS1 medium (on average, 3.8 cm; Table 2). Several aspects ofthe in vitro multiplication process are shown in Fig. 1.
Table 2. Influence of culture medium on shoot multiplication and elongation of the three species of Echinacea after 30 d
Species Culture medium Number of plantlets/explant Length of plantlets (cm) Number of roots/explant Length of roots (cm)
E. purpurea MS1 1.4±0.52 a 4.1±0.54 e 1.9±0.57 fg 2.0±0.23 j
MS2 1.5±0.53 a 4.2±0.57 e 2.2±0.79 fg 2.0±0.28 j
MS3 3.1±0.88 b 2.6±0.29 d 1.7±0.67 f 1.4±0.18 i
MS4 7.1±0.99 c 2.5±0.17 d 4.6±0.84 h 2.4±0.16 k
MS5 2.6±0.7 b 2.4±0.17 d 2.2±0.92 fg 1.3±0.16 i
MS6 2.6±0.7 b 2.5±0.14 d 2.8±0.92 g 1.5±0.18 i
E. pallida MS1 1.0±0 a 3.4±0.59 f 1.7±0.67 g 1.9±0.27 jk
MS2 1.3±0.48 a 3.6±0.38 f 2.3±0.82 g 1.6±0.4 i, j
MS3 2.5±0.71 b 2.5±0.32 de 2.4±0.7 g 1.4±0.18 i
MS4 4.8±0.79 c 2.7±0.32 e 4.5±1.08 h 2.4±0.15 l
MS5 5.4±0.84 c 2.1±0.23 d 1.7±0.67 g 2.2±0.23 kl
MS6 3.2±0.63 b 2.2±0.26 d 2.3±0.82 g 1.3±0.21 i
E. angustifolia MS1 1.0±0 a 3.9±0.5 h 3.3±0.82 ij 3.8±0.29 o
MS2 1.4±0.52 b 3.4±0.25 g 3.8±0.79 j 3.3±0.2 n
MS3 3.6±0.7 c 3.2±0.22 fg 3.3±0.67 ij 1.9±0.16 k
MS4 3.7±0.67 c 2.9±0.28 ef 3.3±0.67 ij 2.6±0.18 m
MS5 3.9±0.74 c 2.6±0.23 d e 2.7±0.82 i 3.4±0.18 n
MS6 3.8±0.79 c 2.4±0.19 d 2.6±0.7 i 2.3±0.24 l
Nodal explants used for this experiment were generated from shoots produced on the same respective media (1.0 mg/l BA+0.1 mg/l NAA). Valuesrepresent means±SD, n010. Values with different letters within a column are significantly different (p<0.001) according to Tukey test
Figure 1. In vitro organogenesis of Echinacea species, after 30 d cul-ture. (a) E. purpurea on MS4 medium supplemented with 0.1 mg/l2iP+0.1 mg/l IBA, (b) E. pallida on MS2 medium supplemented with0.1 mg/l BA+0.1 mg/l NAA, (c) E. angustifolia on MS4 medium, (d)
E. purpurea, (e) E. pallida, (f) E. angustifolia, and (g) callus ofE. purpurea and E. pallida on MS medium supplemented with2.0 mg/l 2,4-D+0.1 mg/l BA.
252 BUTIUC-KEUL, VLASE ET AL.
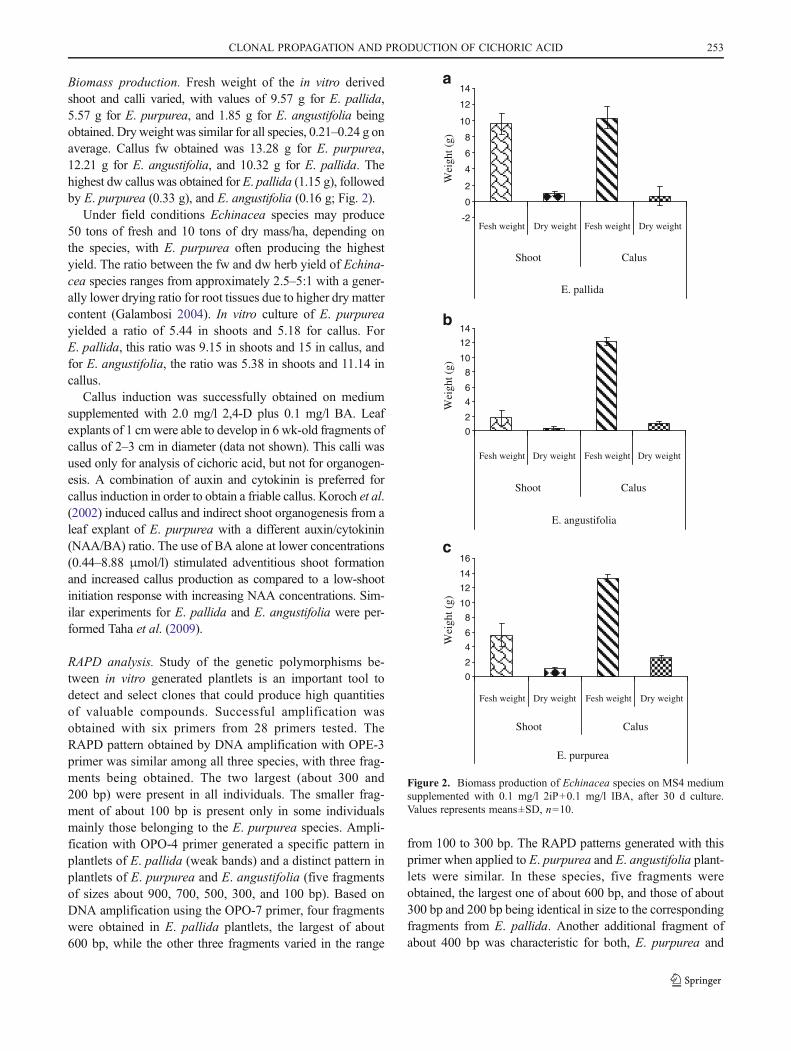
Biomass production. Fresh weight of the in vitro derivedshoot and calli varied, with values of 9.57 g for E. pallida,5.57 g for E. purpurea, and 1.85 g for E. angustifolia beingobtained. Dryweight was similar for all species, 0.21–0.24 g onaverage. Callus fw obtained was 13.28 g for E. purpurea,12.21 g for E. angustifolia, and 10.32 g for E. pallida. Thehighest dw callus was obtained for E. pallida (1.15 g), followedby E. purpurea (0.33 g), and E. angustifolia (0.16 g; Fig. 2).
Under field conditions Echinacea species may produce50 tons of fresh and 10 tons of dry mass/ha, depending onthe species, with E. purpurea often producing the highestyield. The ratio between the fw and dw herb yield of Echina-cea species ranges from approximately 2.5–5:1 with a gener-ally lower drying ratio for root tissues due to higher dry mattercontent (Galambosi 2004). In vitro culture of E. purpureayielded a ratio of 5.44 in shoots and 5.18 for callus. ForE. pallida, this ratio was 9.15 in shoots and 15 in callus, andfor E. angustifolia, the ratio was 5.38 in shoots and 11.14 incallus.
Callus induction was successfully obtained on mediumsupplemented with 2.0 mg/l 2,4-D plus 0.1 mg/l BA. Leafexplants of 1 cmwere able to develop in 6 wk-old fragments ofcallus of 2–3 cm in diameter (data not shown). This calli wasused only for analysis of cichoric acid, but not for organogen-esis. A combination of auxin and cytokinin is preferred forcallus induction in order to obtain a friable callus. Koroch et al.(2002) induced callus and indirect shoot organogenesis from aleaf explant of E. purpurea with a different auxin/cytokinin(NAA/BA) ratio. The use of BA alone at lower concentrations(0.44–8.88 μmol/l) stimulated adventitious shoot formationand increased callus production as compared to a low-shootinitiation response with increasing NAA concentrations. Sim-ilar experiments for E. pallida and E. angustifolia were per-formed Taha et al. (2009).
RAPD analysis. Study of the genetic polymorphisms be-tween in vitro generated plantlets is an important tool todetect and select clones that could produce high quantitiesof valuable compounds. Successful amplification wasobtained with six primers from 28 primers tested. TheRAPD pattern obtained by DNA amplification with OPE-3primer was similar among all three species, with three frag-ments being obtained. The two largest (about 300 and200 bp) were present in all individuals. The smaller frag-ment of about 100 bp is present only in some individualsmainly those belonging to the E. purpurea species. Ampli-fication with OPO-4 primer generated a specific pattern inplantlets of E. pallida (weak bands) and a distinct pattern inplantlets of E. purpurea and E. angustifolia (five fragmentsof sizes about 900, 700, 500, 300, and 100 bp). Based onDNA amplification using the OPO-7 primer, four fragmentswere obtained in E. pallida plantlets, the largest of about600 bp, while the other three fragments varied in the range
from 100 to 300 bp. The RAPD patterns generated with thisprimer when applied to E. purpurea and E. angustifolia plant-lets were similar. In these species, five fragments wereobtained, the largest one of about 600 bp, and those of about300 bp and 200 bp being identical in size to the correspondingfragments from E. pallida. Another additional fragment ofabout 400 bp was characteristic for both, E. purpurea and
-2
0
2
4
6
8
10
12
14
E. pallida
Wei
ght (
g)0
2
4
6
8
10
12
14
E. angustifolia
Wei
ght (
g)
0
2
4
6
8
10
12
14
16
Fesh weight Dry weight Fesh weight Dry weight
Fesh weight Dry weight Fesh weight Dry weight
Fesh weight Dry weight Fesh weight Dry weight
Shoot Calus
Shoot Calus
Shoot Calus
E. purpurea
Wei
ght (
g)
a
b
c
Figure 2. Biomass production of Echinacea species on MS4 mediumsupplemented with 0.1 mg/l 2iP+0.1 mg/l IBA, after 30 d culture.Values represents means±SD, n010.
CLONAL PROPAGATION AND PRODUCTION OF CICHORIC ACID 253

E. angustifolia (Fig. 3). RAPD markers generated with theseprimers did not reveal genetic polymorphism between thein vitro plantlets.
RAPD markers generated with OPO-8 primer showedminimal diferences between the three species of Echinacea.In plantlets of E. pallida, four fragments were obtained,three larger of about 400, 300, and 200 bp and the latter ofabout 80 bp. For this latter species, genetic polymorphismcould not be detected with these RAPD markers. For theE. purpurea-derived plantlets, RAPD patterns generatedwith OPO-8 primer displayed one to four polymorphic frag-ments in the range of 80–500 bp. For E. angustifolia, theRAPD pattern generated with the OPO-8 primer containedthree fragments of about 80–300 bp for all plantlets, and sono genetic variation between samples was detected. RAPDpatterns generated with the primer 4A-27 exhibited fourfragments of about 100–500 bp in E. pallida plantlets andfour fragments of about 100–400 bp in E. purpurea andE. angustifolia plantlets, none being polymorphic. Based onDNA amplification with the 4A-28 primer, five low poly-morphic fragments of about 100–700 bp with low variability
were obtained in plantlets of E. pallida, and four identicalfragments of about 100–600 bp were obtained for E. purpureaand E. angustifolia plantlets (Fig. 4).
RAPD patterns generated with the primer 4A-29 permitteddiscrimination of plantlets belonging to the three species ofEchinacea. For E. pallida plantlets, three identical fragmentsof 250–500 bp were obtained, while four polymorphic frag-ments of about 150–500 bp in plantlets of E. purpurea andtwo identical fragments of 400 and 250 bpwere obtained fromplantlets of E. purpurea and E. angustifolia, respectively.DNA amplification with the primer 4A-30 generated a RAPDpattern that was similar in all plantlets and independent ofspecies, such that five identical fragments of about 120–500 bp were obtained for all plantlets (Fig. 5).
Quantification of cichoric acid. Cichoric acid from Echina-cea extracts were identified on the basis of MS/MSspectrum (Fig. 6), revealing presence in all three speciesof Echinacea, within shoots, leaves, and callus. Highestconcentrations were identified in the shoots and leaves ofE. pallida, followed by the shoots and leaves of E.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28
Figure 3. RAPD pattern of the in vitro plantlets of different species ofEchinacea. Lanes 1–3, E. pallida with OPE-3 primer; lanes 4–6, E.purpurea with OPE-3 primer; lanes 7–9, E. angustifolia with OPE-3primer; lanes 10–12, E. pallida with OPO-4 primer; lanes 13–15, E.purpurea with OPO-4 primer; lanes 16–18, E. angustifolia with OPO-4 primer; lane 19, molecular weight marker; lanes 20–22, E. pallida
with OPO-7 primer; lanes 23–25, E. purpurea with OPO-7 primer;lanes 26–28, E. angustifolia with OPO-7 primer. Molecular weightmarker band sizes, 3,000, 2,000, 1,500, 1,200, 1,000, 900, 800, 700,600, 500, 400, 300, 200, and 100 bp (strong bands, 1,000 and 500 bp;O’Gene Ruler, 100 bp; DNA Ladder Plus, 0.1 μg/μl, Fermentas).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28
Figure 4. RAPD pattern of the in vitro plantlets of different species ofEchinacea. Lanes 1–3, E. pallida with OPO-8 primer; lanes 4–6, E.purpurea with OPO-8 primer; lanes 7–9, E. angustifolia with OPO-8 primer; lanes 10–12, E. pallida with 4A-27 primer; lanes 13–15, E.
purpurea with 4A-27 primer; lanes 16–18, E. angustifolia with 4A-27primer; lane 19, molecular weight marker (see Fig. 3); lanes 20–22, E.pallida with 4A-28 primer; lanes 23–25, E. purpurea with 4A-28primer; lanes 26–28, E. angustifolia with 4A-28 primer.
254 BUTIUC-KEUL, VLASE ET AL.
purpurea (Table 3). Interestingly, the in vitro plantlets ofE. angustifolia contained high concentrations of cichoricacid, whereas field-grown plants contained especiallyechinacoside (Harborne and Williams 2004). It is note-worthy that cichoric acid was identified also in callus,with the highest concentrations observed in the callus ofE. purpurea and the lowest concentrations in the callusof E. angustifolia.
Discussion
In vitro propagation of the three species of Echinacea wassuccessfully obtained in all media studied, but the bestresults were obtained on MS medium supplemented with0.1 mg/l 2iP plus 0.1 mg/l IBA, leading to shoot inductionand multiplication from nodal explants. Several methods ofin vitro culture have been previously reported for
1000 bp
500 bp
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20
Figure 5. RAPD pattern of the in vitro plantlets of different species ofEchinacea. Lane 1, molecular weight marker; lanes 2–4, E. pallidawith 4A-29 primer; lanes 5–7, E. purpurea with 4A-29 primer; lanes8–10, E. angustifolia with 4A-29 primer; lanes 11–13, E. pallida with
4A-30 primer; lanes 14–16, E. purpurea with 4A-30 primer; lanes 17–19, E. angustifolia with 4A-30 primer; 20, molecular weight marker(see Fig. 3).
Figure 6. Typical images of chromatograms and mass spectra (insert) of cichoric acid from reference standard solution (upper panel) and fromEchinacea extracts (lower panel).
CLONAL PROPAGATION AND PRODUCTION OF CICHORIC ACID 255

commercially relevant Echinacea species (Choffe et al.2000a,b; Harbage 2001; Koroch et al. 2002, 2003;Lakshmanan et al. 2002; Zobayed and Saxena 2003; Cokerand Camper 2004; Sauve et al. 2004; Jones et al. 2007; Tahaet al. 2009), and nearly all of these protocols have utilizedembryonic or in vitro grown seedling explants. Such choicesof explant material reflect a preference for juvenile tissuesthat generally have high organogenic competence such thatin vitro seed germination is not difficult to perform (Korochet al. 2003). In other experiments, a range of tissues, includ-ing anther, mesophyll protoplast, petiole, stem, seed, flowerstalks, leaf sections, hypocotyls, cotyledons, and roots havebeen used as explants for in vitro culture and induction ofcallus (Abbasi et al. 2007). Choice of explant varies withspecies and plays an important role in determining theefficiency of propagation.
Somaclonal variation of in vitro plantlets was studiedthrough the use of RAPD markers. A total of 28 primerswere tested, but successful amplification was obtained onlywith six primers. These primers revealed low levels ofgenetic polymorphism between in vitro generated plantlets.OPO-8 primer revealed genetic polymorphism between invitro plantlets of E. purpurea, while the primer 4A-29revealed limited genetic polymorphisms between in vitroplantlets of all Echinacea species. RAPD patterns generatedwith the primer 4A-29 also allowed the discrimination ofplantlets belonging to the three species of Echinacea. ForE. pallida plantlets, three identical fragments were obtainedafter amplification, as compared to four fragments fromplantlets of E. purpurea, and two fragments from plantletsof E. angustifolia. The pattern obtained from use of theprimer 4A-30 was the same for all plantlets, independentof species origin. The limited genetic polymorphism of invitro generated plantlets revealed by RAPD markers may bedue to the genetic variability of selected seeds that wereused to initiate the cultures rather than somaclonal variationinduced by in vitro propagation. Previous genetic character-ization of the three medicinal Echinacea species usingRAPD analysis indicated a high degree of differentiationbetween species with a relatively lower difference between
E. angustifolia and E. pallida. Species-specific markerswere identified from amplicons obtained with four of thetwenty decamer primers contained in the Operon RAPD kitA. In particular, one marker was identified as diagnostic forE. angustifolia (OPA 20, 1,800 bp) and E. pallida (OPA 10,600 bp) and three markers for E. purpurea (OPA11:1250 bp; OPA 17:750, 1800 bp; Nieri et al. 2003). Asthese markers are species specific, they are not useful forclonal variation analysis at the intraspecies level, as com-pared to the informative markers reported in this study.
Although the size of Echinacea plants is determinedmainly by environmental factors such as water stress, fertil-ization, or light exposure, the concentrations of alkylamidesand cichoric acid are predominantly under genetic control,but not exclusively. It would therefore seem commerciallyadvantageous to maximize plant growth, knowing that alky-lamide and cichoric acid concentrations in the plant wouldnot be reduced (Wills and Stuart 1999). That was the reasonfor in vitro multiplication focused on biomass production. Itis well known that efficient and consistent production ofsecondary metabolites and other bioactive natural productsmay be achieved more easily with in vitro cultured cells andorgans than with field-grown plants (Wang et al. 2006).Hydrophilic pharmacological components such as cichoricacid and echinacoside were also detected from cell culturesof E. angustifolia (Smith et al. 2002; Pellati et al. 2005;Wang et al. 2006), but not in E. purpurea and E. pallida.Other studies analyzed the content of cichoric acid in dif-ferent organs of Echinacea (Qu et al. 2005). Ligulate floretsshowed the highest concentration of cichoric acid, whileendosperm and seed coat did not contain this metabolite.For some selected red or pink-flowered E. purpurea clones,cichoric acid content reached 12 %, but was lower in white-flowered E. purpurea (White Swan) and E. pallida. Among12 different lines of white-flowered E. purpurea ligulate flor-ets, cichoric acid reached only 2.6 % (Qu et al. 2005). Thelevel of cichoric acid for in vitro generated plantlets wasinferior to the content of field-grown plants. The content ofcichoric acid identified in shoots and leaves of E. angustifoliaand E. pallida is inferior to that reported by other authors(Taha et al. 2009), but different culture media was used in thepresent study for biomass production. It is not clear why shoottissues produced more cichoric acid than callus, but previousstudies on the extracts from axillary proliferation shoots of E.angustifolia revealed significant production of caffeic acidderivatives, while leaf callus accumulated mainly alkylamides(Lucchesini et al. 2009). These results showed that the choiceof procedures for in vitro multiplication permitted plant bio-mass to be obtained in order produce the active compoundstypical of Echinacea species.
The overall progress made in Echinacea biotechnology islikely to accelerate the industry trend toward controlledcultivation of plants rather than collection from the wild.
Table 3. Concentration of cichoric acid in plantlets and callus ofEchinacea obtained in vitro
Sample Concentration in extract (μg/g dw)
E. purpurea shoots 576.2±0.99
E. purpurea callus 110.1±0.13
E. pallida shoots 730.6±0.38
E. pallida callus 65.1±0.27
E. angustifolia shoots 389.3±0.33
E. angustifolia callus 23.8±0.21
Values represent means±SD, n010
256 BUTIUC-KEUL, VLASE ET AL.

There is an increased recognition in the natural product indus-try of the requirement for new standards of the quality relatedto the raw and processed plant material (Busse 2000; Barnes2003). In vitro cultures could provide plant material free ofpathogens and xenobiotics benefiting extract preparation. Theresults from this study are important because they show that invitro plantlets of Echinacea produce important quantities ofcichoric acid, as do field-grown plants; in addition theseextracts are free of xenobiotics and other pollutants.
Acknowledgments This work was supported by CNCSIS grantnumber 1338/2005, financed by Ministry of Education and Research,Romania. Eppendorf thermocycler and electrophoresis devices wereprovided by the Alexander von Humboldt Foundation.
References
Abbasi BH, Saxena PK, Murch SJ, Liu C-Z (2007) Echinacea bio-technology: challenges and opportunities. In Vitro Cell Dev BiolPlant 43:481–492
Barnes J (2003) Quality, efficacy and safety of complementary medi-cines: fashions, facts and the future. Part I. Regulation and quality.Br J Clin Pharmacol 55:226–233
Barrett B (2003) Medicinal properties of Echinacea: a critical review.Phytomedicine 10:66–86
Baum BR, Binns SE, Arnason JT (2006) Integrating recent knowledgeabout the genus Echinacea: morphology, molecular systematics,phytochemistry. Herbal Gram 72:32–46
Binns SE, Arnason JT, Baum BR (2004) Taxonomic history andrevision of the genus Echinacea. In: Miller SC (ed) Echinacea.The genus Echinacea. CRC, Boca Raton, pp 3–11
Blumenthal M (2005) Herb sales down 7.4 percent in mainstreammarket. Herbal Gram 66:63
Busse W (2000) The significance of quality for efficacy and safety ofherbal medicinal products. Drug Inf J 34:15–23
Butiuc-Keul AL, Deliu C (2001) Clonal propagation of Arnica montanaL.,a medicinal plant. In Vitro Cell Dev Biol Plant 37:581–585
Butiuc-Keul AL, Suteu A, Keul M, Deliu C (2002) The role of naturalextracts on the in vitro multiplication of Arnica montana. ContribBot, Cluj-Napoca 37:183–190
Choffe KL, Murch SJ, Saxena PK (2000a) Regeneration of Echinaceapurpurea: induction of root organogenesis from hypocotyls andcotyledon explants. Plant Cell Tissue Organ Cult 62:227–234
Choffe KL, Victor JMR, Murch SJ, Saxena PK (2000b) In vitroregeneration of Echinacea purpurea L.: direct somatic embryo-genesis and indirect shoot organogenesis in petiole culture. InVitro Cell Dev Biol Plant 36:30–36
Coker PS, Camper ND (2004) In vitro culture of Echinacea species. In:Miller SC (ed) Echinacea. The genus Echinacea. CRC, BocaRaton, pp 23–28
Crăciunaş C, Coste A, Vălimareanu S, Farago M, Iliescu M, Iuoraş R,Dordea M, Butiuc-Keul AL (2007) Molecular characterization oftwo grapevine cultivars from Romania. Sci Res Hort 463–472
Currier NL, Miller SC (2000) Natural killer cells from aging micetreated with extracts from Echinacea purpurea are quantitativelyand functionally rejuvenated. Exp Gerontol 35:627–639
Doyle JJ, Doyle JL (1987) A rapid DNA isolation procedure for smallquantities of freh leaf tissue. Phytochem Bull 19:11–15
Galambosi B (2004) Cultivation in Europe. In: Miller SC (ed) Echinacea.The genus Echinacea. CRC, Boca Raton, pp 29–52
Harbage JF (2001) Micropropagation of Echinacea angustifolia, E.pallida, and E. purpurea from stem and seed explants. HortS-cience 36:360–364
Harborne JB,Williams CA (2004) Phytochemistry of the genus Echinacea.In: Miller SC (ed) Echinacea. The genus Echinacea. CRC, BocaRaton, pp 55–71
Jones MPA, Yi Z, Murch SJ, Saxena PK (2007) Thidiazuron-inducedregeneration of Echinacea purpurea L.: micropropagation in solidand liquid culture systems. Plant Cell Rep 26:13–19
Koroch A, Juliani HR, Kapteyn J, Simon JE (2002) In vitro regenerationof Echinacea purpurea from leaf explants. Plant Cell Tissue OrganCult 69:79–83
Koroch AR, Kapteyn J, Juliani HR, Simon JE (2003) In vitro regener-ation of Echinacea pallida from leaf explants. In Vitro Cell DevBiol Plant 39:415–418
Lakshmanan P, Danesh M, Taji A (2002) Production of four commer-cially cultivated Echinacea species by different methods of invitro regeneration. J Hortic Sci Biotechnol 77:158–163
Lata H, Bedir E, Moraes RM, Andrade Z (2004) Mass propagation ofEchinacea angustifolia: a protocol refinement using shoot encap-sulation and temporary immersion liquid system. Acta Hortic629:409–414
Lucchesini M, Bertoli A, Mensuali-Sodi A, Pistelli L (2009) Establishmentof in vitro tissue cultures from Echinacea angustifolia D.C. adultplants for the production of phytochemical compounds. Sci Hortic122:484–490
Mechanda SM, Baum BR, Johnson DA, Arnason JT (2003) Direct shootregeneration from leaf segments of mature plants of Echinaceapurpurea (L.) Moench. In Vitro Cellular Dev Biol Plant 39:505–509
Merali S, Binns S, Paulin-Levasseur M, Ficker C, Smith M, Baum BR,Brovelli E, Arnason JT (2003) Antifungal and anti-inflammatoryactivity of the genus Echinacea. Pharm Biol 41:412–420
Murashige T, Skoog A (1962) A revised medium for rapid growth andbioassays with tobacco tissue cultures. Physiol Plant 15:473–483
Murch SJ, Peiris SE, Shi WL, Zobayed SMA, Saxena PK (2006)Genetic diversity in seed populations of Echinacea purpureacontrols the capacity for regeneration, route of morphogenesisand phytochemical composition. Plant Cell Rep 25:522–532
Nieri P, Andinolfi B, Morelli I, Breschi MC, Simoni G, Martinotti E(2003) Genetic characterization of the three medicinal Echinaceaspecies using RAPD analysis. Planta Med 69:685–686
Pellati F, Benvenuti S, Melegari M, Lasseigne T (2005) Variability inthe composition of anti-oxidant compounds in Echinacea speciesby HPLC. Phytochem Anal 16:77–85
Perry NB, Burges EJ, Glennie VL (2001) Echinacea standardization:analytical methods for phenolic compounds and typical levels inmedicinal species. J Agric Food Chem 49:1702–1706
Qu L, Chen Y, Wang X, Scalzo R, Davis JM (2005) Patterns of variationin alkamides and cichoric acid in roots and aboveground parts ofEchinacea purpurea (L.) Moench. HortScience 40:1239–1242
Raman P, Patino LC, Nair MG (2004) Evaluation of metal and micro-bial contamination in botanical supplements. J Agric Food Chem52:7822–7827
Sauve RJ, Mmbaga MT, Zhou S (2004) In vitro regeneration of theTennessee Coneflower (Echinacea tennesseensis). In Vitro CelDev Biol Plant 40:325–328
Smith MAL, Kobayashi H, Gawienowski M, Briskin DP (2002) An invitro approach to investigate medicinal chemical synthesis bythree herbal plants. Plant Cell Tissue Organ Cult 70:105–111
Taha HS, Abd El-Rahman RA, Maharik NT (2009) In vitro productionof caffic acid derivatives in calli and regenerate cultures of Ech-inacea angustifolia DC and Echinacea pallida Nutt. J Appl SciRes 5:13–20
Thygesen L, Thulin J, Mortensen A, Skibsted LH, Molgaard P (2007)Antioxidant activity of cichoric acid and alkamides from Echina-cea purpurea, alone and in combination. Food Chem 101:74–81
CLONAL PROPAGATION AND PRODUCTION OF CICHORIC ACID 257

Wang B, Zhang G, Zhu L, Chen L, Zhang Y (2006) The genetictransformation of Echinacea purpurea with Agrobacterium rhizo-genes and bioactive ingredients analysis in transformed cultures.Colloids Surf B 53:101–104
Wills RBH, Stuart DL (1999) Generation of high quality AustralianEchinacea purpurea products. Rural Industries Research andDevelopment Corporation, Barton, ACT, Australia, pp 3-9
Wills RBH, Stuart DL (2000) Effect of handling and storage onalkylamides and cichoric acid in Echinacea purpurea. J Sci FoodAgric 80:1402–1406
Zobayed SMA, Saxena PK (2003) In vitro regeneration of Echinaceapurpurea L.: enhancement of somatic embryogenesis by indole-butyric acid and dark pre-incubation. In Vitro Cell Dev Biol Plant39:605–612
258 BUTIUC-KEUL, VLASE ET AL.




















