
Climate, fire, farming and the recent vegetation history of subantarctic Campbell Island
15
Earth and Environmental Science Transactions of the Royal Society of Edinburgh http://journals.cambridge.org/TRE Additional services for Earth and Environmental Science Transactions of the Royal Society of Edinburgh: Email alerts: Click here Subscriptions: Click here Commercial reprints: Click here Terms of use : Click here Climate, re, farming and the recent vegetation history of subantarctic Campbell Island Matt McGlone, Janet Wilmshurst and Colin Meurk Earth and Environmental Science Transactions of the Royal Society of Edinburgh / Volume 98 / Issue 01 / March 2007, pp 71 - 84 DOI: 10.1017/S1755691007000060, Published online: 05 October 2007 Link to this article: http://journals.cambridge.org/abstract_S1755691007000060 How to cite this article: Matt McGlone, Janet Wilmshurst and Colin Meurk (2007). Climate, re, farming and the recent vegetation history of subantarctic Campbell Island. Earth and Environmental Science Transactions of the Royal Society of Edinburgh, 98, pp 71-84 doi:10.1017/S1755691007000060 Request Permissions : Click here Downloaded from http://journals.cambridge.org/TRE, IP address: 130.216.158.78 on 06 Nov 2014
-
Upload
landcareresearch -
Category
Documents
-
view
3 -
download
0
Transcript of Climate, fire, farming and the recent vegetation history of subantarctic Campbell Island

Earth and Environmental Science Transactions of the RoyalSociety of Edinburghhttp://journals.cambridge.org/TRE
Additional services for Earth and Environmental Science Transactions of theRoyal Society of Edinburgh:
Email alerts: Click hereSubscriptions: Click hereCommercial reprints: Click hereTerms of use : Click here
Climate, re, farming and the recent vegetation history of subantarcticCampbell Island
Matt McGlone, Janet Wilmshurst and Colin Meurk
Earth and Environmental Science Transactions of the Royal Society of Edinburgh / Volume 98 / Issue 01 / March 2007, pp 71 - 84DOI: 10.1017/S1755691007000060, Published online: 05 October 2007
Link to this article: http://journals.cambridge.org/abstract_S1755691007000060
How to cite this article:Matt McGlone, Janet Wilmshurst and Colin Meurk (2007). Climate, re, farming and the recent vegetation history ofsubantarctic Campbell Island. Earth and Environmental Science Transactions of the Royal Society of Edinburgh, 98, pp71-84 doi:10.1017/S1755691007000060
Request Permissions : Click here
Downloaded from http://journals.cambridge.org/TRE, IP address: 130.216.158.78 on 06 Nov 2014

Climate, fire, farming and the recent vegetationhistory of subantarctic Campbell Island
Matt McGlone, Janet Wilmshurst and Colin MeurkLandcare Research, PO Box 40, Linden 7640, New Zealand.E-mail: [email protected]
ABSTRACT: Campbell Island is a small, uninhabited peat-covered island lying in the coolsouthern ocean 600 km south of the New Zealand mainland. Dracophyllum scrub is the main coverfrom sea level to 200 m, above which tussock grassland, macrophyllous forbs and tundra dominate.Seven peat profiles from sea level to the tundra zone provide an elevational transect for pollen andcharcoal records spanning the last 500 years. Scrub density was relatively low between 200 and400 cal yrs BP, possibly due to Little Ice Age cooling, but had recovered by the time Europeansdiscovered the island in AD 1810. Burning and grazing during a brief farming episode (AD 1895–1931) severely reduced scrub and palatable grasses and forbs. Vegetation recovery is now welladvanced following cessation of farming and the later elimination of all feral grazing animals, catsand rats. Climates were cool in the southwest Pacific during the farming period, and since AD 1970the island has warmed by c. 0·5(C. However, there has been no upwards movement of the scrublinedespite vigorous regeneration of scrub at lower altitudes. The island’s cloudy, highly oceanic climateappears to offset increasing summer warmth, and scrubline is likely to rise only if clearer and lesswindy, as well as warmer, summers eventuate.
KEY WORDS: charcoal, Dracophyllum, Holocene, Little Ice Age, pollen analysis, subantarctic
A scatter of six, small, isolated island groups lie to the south ofNew Zealand from 48(S to 55(S in the cool ocean watersbetween the subtropical convergence and the Antarctic conver-gence (Fig. 1). These islands are at the southern limit forwoody growth and thus span a major biome transition other-wise not represented in this sector of the Southern Ocean.The stormy, windy conditions make the islands inhospitableand difficult of access. All islands were exploited for marinemammals, and the larger two islands (Campbell andAuckland) had short episodes of farming in the 19th century.Military coast-watching stations were set up on Campbell andAuckland islands during World War II and staffed in part byscientists. The unusual vegetation and extraordinary wildlifeon these island groups has attracted many scientific expedi-tions since the middle of the 19th century and now regular visitsby conservation management teams. Despite their relativeinaccessibility the islands have a remarkably well documentedhistory. Early visitors often published their observations andtook photographs. The islands are covered with thick peats,from which a substantial number of peat cores have beencollected. These peat cores have been analysed for pollen andcharcoal and provide a detailed record of post-glacial veg-etation and climate change for the islands (McGlone 2002).Campbell and Auckland islands thus provide the opportunityto combine well-documented histories of human activitieswith vegetation and climate sequences, allowing us to developour understanding of the complex interactions occurring atsensitive ecotones.
This paper looks at the combined effects of climate change,grazing and fire on Campbell Island. One of the advantages ofworking on a peat-covered island is that palaeoecologicalprofiles can placed precisely at ecologically and climaticallysensitive locations. New palynological and charcoal evidence ispresented from a series of seven peat profiles positioned closeto ecotone boundaries from sea level to 450 m on leeward
slopes of the island. Using this new information, along withpreviously published historical observations, photographs, anddemographic studies of scrub spread, the following questionsare addressed: What was the prehuman vegetation of theisland? How did it change in the course of the 19th and20th centuries? How have fire, grazing and climate changeinteracted? And finally, what is likely to happen to thevegetation cover in the future? In particular, it is hoped toresolve the issue of whether or not the current marked climatewarming in the Southern Ocean region is having a significanteffect on the vegetation independent of human and animalimpacts.
1. Campbell Island: setting, environment andvegetation cover
Campbell Island (52(34#S, 169(09#E) covers 113 km2, and isthe eroded eastern segment of a Pliocene volcanic complex(Campbell 1981). Basaltic flows and volcanic intrusions covertwo-thirds of the island, and schists, sedimentary conglomer-ates, mudstones, limestones, tuffs and breccias make up theremaining area. Vigorous wave action has eroded steep cliffs incoasts exposed to the prevailing westerly weather. The easternlowlands have gentle to moderately steep slopes and broadvalleys. Basaltic and gabbro intrusions form prominent conicalhills, and resistant basaltic and andesitic flows form steepridges and peaks. Some peaks are over 500 m (the highest isMt Honey at 570 m). The island, on the evidence of U-shapedvalleys, tills and cirque-like landforms in the uplands, appearsto have been heavily glaciated in the past (McGlone et al.1997).
The island lies within a zone of persistent, strong westerlywinds (70% of days with surface winds >7·5 m s�1), and has acool, moist, cloudy climate (Table 1). The climate is highlyoceanic (�2·4 on the Conrad Continentality Index for the
Earth and Environmental Science Transactions of the Royal Society of Edinburgh, 98, 71–84, 2007
� 2007 The Royal Society of Edinburgh. doi:10.1017/S1755691007000060

Fig
ure
1(L
eft)
Cam
pbel
lIsl
and
loca
tion
s,st
udy
area
show
nby
rect
angl
e.In
set:
loca
tion
ofC
ampb
ellI
slan
dre
lati
veto
New
Zea
land
mai
nlan
d.(R
ight
)st
udy
area
show
ing
cori
ngsi
tes
and
1970
fenc
elin
e.
72 MATT MCGLONE ET AL.

years 1941–1970) due to the small island size, persistent cloud,low sunshine hours and high humidity (Meurk & Blaschke1990).
Peats cover nearly the entire landscape, and only fresh slips,cliffs, steep slopes >30(, outcrops and exposed tops above400 m have mineral soils (Campbell 1981). Widespread, well-humified peat soils (0·5–4·0 m deep; >95% organic) blanketmoderately to well-drained slopes. Water tables in the peatsoils are low, and shallow holes (c. 50 cm) dug in them do notfill with water except immediately after rain. Nevertheless, peatsoils are continuing to grow vertically as litter breakdown isslow under year-round cool temperatures and constantly moistsurface layers (McGlone 2002). Wetlands have high watertables (<5 cm). Oligotrophic peat bogs up to 10 m deep andconsisting of coarse, fibrous peat tend to form where drainageis sluggish or impeded, mainly on broad interfluves or in gentlysloping valleys. Fens of poorly consolidated peat occupydrainage runnels and flushes. On steep slopes, shallow peat orpeaty loams form over the regolith, and landslides after heavyrain are relatively common (Campbell 1981).
The indigenous vascular flora of the island consists of132 species (Meurk et al. 1994a) including two small trees(Dracophyllum longifolium and D. scoparium) and five shrubs(Myrsine divaricata, Coprosma cuneata, C. ciliata, Hebebenthamii, and H. elliptica). D. longifolium and D. scopariumhave introgressed on the island, and many specimens cannot bereliably attributed to either species (Wardle 1987). A strikingelement of the flora is the seven macrophyllous forb (hereafter‘macroforb’) species (Anisotome antipoda, A. latifolia,Bulbinella rossii, Pleurophyllum criniferum, P. hookeri, P.speciosum and Stilbocarpa polaris), all but the last endemic tothe southern islands. Although a large range of vegetationcommunities have been identified (Meurk & Given 1990;Meurk et al. 1994a), a few broad groupings adequatelycharacterise the island for the purpose of the present paper.
The heavy ocean swell, strong persistent salt-laden winds(Meurk et al. 1994b), and high usage by marine mammals haveinduced a maritime turf and grassland vegetation along theexposed coasts. Maritime turfs occur in the most exposedconditions, with macroforb–grassland communities and tallPoa-dominated tussock grasslands extending up steep slopes.At low altitudes and sheltered sites, Dracophyllum spp. formlow forest (up to 5 m tall). Myrsine divaricata and Coprosmaspp. occur within the forest and as fringing thickets in exposedlocalities, and the ferns Polystichum vestitum and Histiopterisincisa are common in canopy gaps and gullies. With increasingaltitude and lowering canopy height, Myrsine divaricata,Coprosma spp. and Polystichum vestitum play increasinglydominant roles especially in the face of increased exposure.Tussock grassland (before farming mainly Chionochloa
antarctica, and latterly browse-resistant Poa litorosa) becomeincreasingly prominent from around 100 m in altitude,especially on exposed faces. Tussock grassland is dominantfrom 200 m, which is approximately the elevational limit forcontinuous woody cover, although patches of low-growingscrub may extend on sheltered sites up to 250 m. The altititu-dinal limits (reached on sheltered north-facing slopes) forstunted individuals of D. scoparium and D. longifolium are396 m and 308 m respectively. The tussock grassland becomeslower and sparser with altitude and interfingers with macro-forb stands and Marsippospermum gracile (Juncaceae) turfmeadows. Above 300–400 m, alpine tundra with rushes andstunted macroforbs replaces grassland and, on the most ex-posed and highest sites, stony fellfields with rosette herbs andscattered stunted macroforbs and graminoids occur. There arearound 39 adventive vascular plant species, and ten widespreadnaturalised species, Cerastium fontanum, Poa annua, Stellariamedia, Sagina procumbens, Poa pratensis, Festuca rubra andAgrostis capillaris being the most consistently encountered(Meurk 1977).
Peat soils have a cover of scrub, grassland, macroforbor tundra vegetation. Oligotrophic bogs have mosaics ofdwarf shrubs (Dracophyllum scoparium, Myrsine divaricata,Coprosma spp.), tussocks and cushions (Centrolepis ciliata,Coprosma perpusilla, Astelia antarctica, Oreobolus pectinatus,etc). Fens are characterised by Carex appressa, Pleurophyllumcriniferum, Hierochloe redolens, ferns and Coprosma spp.
2. Campbell Island history
Exploitation of the island for its abundant marine mammalsbegan immediately after its discovery in 1810, but large haulsof seal skins ceased by around 1820 (Chilton 1909) and sealing,as a profitable activity, was over by 1830 (Redwood 1950).During this time Perseverance Harbour, being the safestanchorage, was the temporary home for a number of sealinggangs and the Ross Expedition in 1840 noted the remains ofhuts at the head of the harbour. Gangs would occasionallyremain for months at a time. A four-member sealing gang hadbeen on the island four years when it was picked up in January1839 at the very end of the sealing era (Eden 1955). Localisedoccupation during the 19th century is indicated by the remainsof huts and grave stones at Perseverance Harbour, NortheastHarbour and Northwest Bay (Meurk 1977). The island wasalso an occasional port of call for ships to take on water andwood (Ross 1847; McNab 1907). A number of scientificexpeditions visited for short periods, and a French expeditionmade a four-month visit in 1874 (Godley 1970).
Norway rats (Rattus norvegicus) probably establishedduring the sealing era, and flourished, reaching populationsthat fluctuated seasonally between 50 000 and 200 000(McClelland & Tyree 2002). A few sheep (Ovis aries) and goats(Capra domestis) were liberated on Campbell Island between1865 and 1895 to provide food for castaways, but these soondied out (Cockayne 1909). In November 1894, James Gordonacquired the New Zealand Government farming lease for theisland, and a small flock of sheep was established in 1895, butmajor importations were not made until 1901 (SouthlandTimes: 24 April 1895; 3 January 1901). Half the island wasgrazed by 1903 (Cockayne 1903) and two-thirds by 1909(Laing 1909). Sheep farming was ultimately uneconomic andthe farm was abandoned in August 1931, leaving behind4000 sheep and about 20 cattle (Bos taurus) and cats (Felisdomesticus) (Rudge 1986). Whaling had begun in early 1909 inNorthwest Bay, and in Northeast Harbour from 1911 to 1914(Kerr 1976), but by 1955 the whaling had ceased (Eden 1955).
Table 1 Climate statistics 1941–1970 (NIWA 2003). Campbell Island(Mt Beeman 15 m asl)
Mean annual temperature 6·8(CMean temperature warmest month (January) 9·3(CMean temperature coldest month (July) 4·6(CMean daily temperature range 4·7(CExtreme maximum temperature 20·8(CExtreme minimum temperature �6·7(CDays per annum with screen frost 23·6Average annual rainfall 1404 mmRaindays per annum (>1 mm) 252Average relative humidity (9 am) 87%Sunshine hours per annum (% possible) 664 (17%)Overcast days at 9 am (1942–1962) 291
RECENT VEGETATION HISTORY, CAMPBELL ISLAND 73
Military coast-watchers were stationed in PerseveranceHarbour on Campbell Island during 1941–1945 and, besidestheir military duties, undertook scientific observations(Sorensen 1950). Coast-watching ceased in 1944, but an officialMeteorological Station at the foot of Mt Beeman has beenin continuous operation since 1941 (staffed until 1990, butautomated since then).
Campbell Island became a nature reserve in 1954 (Bell1970), and by the 1960s the increasing sheep numbers wererecognised as a threat to natural values (Moore & Moffat1990). In 1970 a fenceline was erected across the centre of theisland, from Tucker Cove to the cliffs west of Col Peak, by theEcology Division of the New Zealand Department of Scientificand Industrial Research (Fig. 1) to evaluate the effects of sheepon the flora and fauna by removing them from the northernhalf of the island. Monitoring in 1981 and 1984 of plotsestablished in 1970 showed that removal of grazing pressure inthe north led to rapid regeneration of palatable large forbs andgrasses on fertile soils. Change was slower on infertile, cold orexposed sites and those that were naturally occupied by shrubsor browse-resistant forbs and grasses (Meurk 1982, 1989). Thesheep population south of the fence increased from 2500 to3500 in 1983. A second fenceline was erected in 1984, restrict-ing sheep to the southwestern part of the island. In 1987, allremaining sheep were shot. Cats died out through naturalcauses in the 1990s possibly because the elimination of sheepincreased tussock growth thereby reducing their preferredshort-grass habitat. Norway rats were eliminated by poisoningin the winter of 2001 and Campbell Island is the largest ever tobe freed of rats (McClelland & Tyree 2002).
3. Palynological and charcoal analyses
A modern pollen survey consisting of samples from 46 sitescovering all the major vegetation types has been publishedfor Campbell Island (McGlone & Meurk 2000), and all pollenand spore taxa percentages were shown to have a significantpositive correlation with their abundance in the vegetation,with the exception of the widespread ferns Polystichumvestitum and Blechnum spp., and the ubiquitous macroforbBulbinella rossii, which are over-represented in the pollen rain.In most cases the pollen and spore spectra permitted confidentidentification of the local vegetation cover, although thestatistical relationships were not strong enough to attemptquantitative reconstruction. A further 16 surface samples havesince been added to the modern pollen data set.
A total of three pollen diagrams from Campbell Island havebeen published from exposures in marine cliffs and a peat scarp
(Moar 1973; McGlone et al. 1997). They are all low resolutionand poorly dated, but show that peat growth on the islandbegan in the early late glacial 15 000–16 000 cal yrs BP with atundra-like macrophyllous forb and grassland vegetationdominating, and that the current scrub-tussock landscapeswere formed by c. 8000 cal yrs BP. The two cliff sites register amore exposed environment after 3000–4000 cal yrs BP.
3.1. SitesSeven new pollen diagrams have been completed for this studyranging in altitude from near sea level to 450 m, and coveringthe complete environmental gradient on the island with theexception of the coastal tussock–macroforb zone. Some sitesextend back to c. 15 000–16 000 cal yrs BP, but for the pur-poses of this study only the last 500 years will be considered.
Sites used in this study are from deep (>6 m) raised bogs,shallow (<4 m) peat soils and one from a high-altitude tundrasoil. The critical differences between the three substrate typesare: raised bogs have a high water table, typically a fewcentimetres below the surface, and thus often a vegetativecover of wetland plants; peat soils tend to sit above the watertable, and have exclusively shrub, grass or macroforb veg-etation cover; and tundra soils which occur on the exposed,high-altitude tops, tend to be very shallow with numerousstones, silt and sand (almost entirely lacking from the othertwo types) and largely vegetated with Marsippospermumrushes, low-growing graminoids and herbs, mosses and stuntedmacroforbs.
The sites are referred to in this paper by the suffix S (peatsoil), B (peat bog) or T (tundra soil) coupled with their altitudeabove sea level in metres. Collection numbers and basic siteinformation are given in Table 2.
3.2. MethodsCores from the deep bog sites (B-50 & B-120) were taken witha 1-m-long, 10-cm-diameter D-section peat corer. The S-30peat soil was sampled from an exposure. The other shallowpeat soils were sampled by inserting a longitudinally split10-cm-diameter PVC tube into the peat, and digging it outintact. Cores were wrapped in polythene film and stored at4(C in the laboratory. Measured samples of 1–2 cm3 weretaken from the cores in the laboratory and pollen and sporepreparation followed standard techniques of heating in 10%KOH, sieving (100 �m), acetolysis, and mounting in glycerinejelly for microscopic analysis (Moore et al. 1991). Sampleswere spiked during the initial processing step with tablets ofknown amounts of exotic Lycopodium for calculation ofpalynomorph concentrations. Chionochloa antarctica and Poa
Table 2 Pollen site details
Designation Field code and location name Alt (m) Site type Vegetation Total core depth (m)
S-30 X9911 (Homestead Scarp) 30 Peat soil Dracophyllum–Myrsine–Coprosma scrub(1·5–2·0 m height)
3·96
B-50 X9901 (Homestead Bog) 50 Oligotrophic raised bog Cushion bog with scattered dwarfDracophyllum and Chionochloa
9·60
S-115 X9906 (Fenceline transect) 115 Peat soil Dracophyllum (<1·2 m height), Myrsinedivaricata, Coprosma spp. scrub
0·50
B-120 X9903(Mt Honey Saddle) 120 Oligotrophic raised bog Cushion bog with 30% Dracophyllumshrub cover (<0·5 m height)
6·52
S-195 X9908 (Fenceline transect) 195 Peat soil Grassland–macroforbs; limit toDracophyllum – scattered shrubs(<0·4 m height)
0·84
S-360 X9914 (Mt Azimuth) 360 Peat soil Macroforbfield 0·50T-420 X99/12 (Mt Fitzeau saddle) 420 Tundra peat soil Sedge–rush–macroforb tundra 0·50
74 MATT MCGLONE ET AL.

litorosa are of a similar size (R50 �m) but largerthan other grass pollen. Under oil immersion they can bedistinguished by the tendency for the surface microspicules toform discrete islands in Poa but to be uniformly spreadin Chionochloa. Although not all grains can be definitivelyclassified as one or the other, and thus there is more uncer-tainty than for most palynomorphs, curves for these twospecies are presented because of their importance in signallingchanges in the browsing regime. The pollen sum consists of allpollen and spore types and for nearly all samples a sum >250palynomorphs was achieved. In the fast-growing bog peats,pollen concentrations were often very low and a sum of>150 palynomorphs was resorted to for some slides. Pollenconcentrations were calculated by the ratio of spike to otherpalynomorphs. Microscopic charcoal particles >50 �m lineardimension were scored along with the palynomorph count andare presented as a percentage of the pollen sum.
Chronology is provided by Accelerator Mass Spectrometryradiocarbon dating on peat (Table 3). Use of bulk samples wasunavoidable at these sites because of the well decomposednature of the peat. Additional chronology was provided bydetermination of the farming horizon (c. AD 1895) as revealedby the appearance of charcoal and the rise of unpalatablespecies and decline of browse-sensitive species. Pollen typesregarded as belonging to unpalatable or browse-resistantspecies are Bulbinella, Poa litorosa (although this species isgrazed in the absence of more palatable vegetation) andall shrubs. Anisotome, Chionochloa, Pleurophyllum, andStilbocarpa are classed as browse-sensitive species (Meurk1977).
Charcoal was also analysed at one site (S-115) by themacroscopic sieving method (Millspaugh & Whitlock 1995) inorder to compare results from pollen-slide charcoal analyses.From each 1-cm contiguous core interval, subsamples of 1 cm3
of wet sediment were soaked in a deflocculant for several days,and gently washed through a series of nested sieves (mesh sizes250, 125, and 63 �m). Counting was done under a stereo-microscope of sieved sediment dispersed as a shallow layer inwater in a gridded Petri dish. Charcoal particles can be reliablyrecognised by their dark, opaque appearance, angular planarshape, and brittleness under pressure.
4. Results
4.1. Macroscopic charcoal – S-115Charcoal of any size range is rare in Campbell Island peats,and in the S-115 profile the first trace of macroscopic charcoal
occurs at 16·5 cm (Fig. 2). The most abundant size category is125–250 �m (maximum count 195 particles 1 cm3) while thesmaller and larger size categories had maximum counts of<100. Because particles >200 �m tend to be deposited within25 m of a fire, and even small-sized charcoal (<20 �m) ismainly deposited less than 100 m from a fire (Clark 1988), it isapparent that most of the charcoal is derived from fires at thesite or in immediately adjacent areas. The macroscopic char-coal curves show early peaks of larger sized charcoal and alater surge in smaller particles (63–125 �m). There is noindication in the macroscopic charcoal curve of individual fireevents or phases. The relatively smooth unimodal curve isconsistent with regular burning, perhaps on a seasonal basis,rather than sporadic burns separated by long periods withoutfire. The macroscopic charcoal categories correlate well withthe microscopic pollen-slide charcoal particles, especially the125–250 �m fraction (r2=0·924). The first influx of macro-scopic charcoal in the 16–17 cm sample coincides well with thefirst trace of Bulbinella pollen at 18 cm and the major increaseof macroscopic charcoal in the 14–15 cm sample with apronounced rise of microscopic charcoal and Poa litorosaat 14 cm. Macroscopic charcoal input collapses rapidlybetween the 6–7 and 5–6 cm samples and Dracophyllumrises to a peak at 6 cm. If this Dracophyllum peak is, asseems probable, part of the post-farming recovery (Bestic et al.2005), the continuing influx of charcoal particles, althoughlow, is problematic, as there was no further burning after 1931.The most probable explanations are continued flaking ofcharcoal fragments from charred stumps, or erosion andredeposition from charcoal-rich exposed peat surfaces (poss-ibly by sheep).
4.2. Pollen and microscopic charcoal: Pre-farming:500–55 cal yrs BP (cal AD 1450–1895)The palynological indications of pre-farming vegetation of theseven sites is consistent with the current vegetation zonation ofthe leeward side of the island. The below-shrubline peat soilsites (S-30 (Fig. 3), S-115 (Fig. 5)) have 50–80% shrub pollenconsistent with a local shrub cover of 40–80% (McGlone &Meurk 2000). The total contribution of shrub pollen (includingDracophyllum, Myrsine and Coprosma undiff.) falls from 20%at S-195 (Fig. 7), which lies at the shrub line, to low percent-ages of Coprosma undiff. only at S-360 (Fig. 8), and to noshrub representation at the highest-altitude site, T-420 (Fig. 9).S-195 is dominated by grass but with some macroforb rep-resentation; S-360 by macroforbs and grass; and T-420 bymacroforbs with little grass. Compared with their present
Table 3 Accelerator mass spectrometry radiocarbon ages with calibrated age ranges reported in cal yrs BP. Radiocarbon ages were calibrated withCALIB 5.1 radiocarbon calibration program (Stuiver et al. 2005) using the Southern Hemisphere calibration dataset shcal04.14c (McCormac et al.2004). All samples submitted were of highly humified bulk peat
Site name Depth (cm) Lab code 14C years BP Median calibrated age BP One sigma distribution of range
Lower calibrated age range Upper calibrated age range
S-30 28 Wk-17095 574�32 537 522 551B-50 28 Wk-17690 279�36 294 154 322B-50 50 Wk-13468 797�41 690 666 721S-115 20 Wk-14719 439�43 459 337 505B-120 21 Wk-13456 313�42 369 298 441B-120 36 Wk-13457 839�43 711 678 734S-195 10·5 Wk-14722 510�43 512 497 533S-360 20·5 Wk-14734 106·8%M* — — —S-360 26·5 Wk-17693 82�37 71 -5 240T-420 17 Wk-14731 486�48 498 472 531
* Modern sample calibrated against CALIBOMB (Wellington) to November 1957–March 1958 (�8 cal BP).
RECENT VEGETATION HISTORY, CAMPBELL ISLAND 75

vegetation cover, the two bog sites (B-50 (Fig. 4), B-120(Fig. 6)) have relatively low representation of wetland pollentypes and high scrub values.
Density of scrub (in particular, Dracophyllum) may havedeclined between 400 and 200 cal yrs BP (c. cal AD 1550–1750). This is most clearly shown in B-50 (Fig. 4), B-120
Figure 2 Site S-115. Macroscopic sieved charcoal, pollen slide charcoal and browse-sensitive (Chionochloa) andbrowse-resistant (Poa litorosa and Bulbinella) pollen types.
Figure 3 Site S-30, percentage diagram. Pollen sum: total pollen and spores. Charcoal fragments as percentageof sum. Dotted line: commencement of farming.
76 MATT MCGLONE ET AL.

(Fig. 6) and S-195 (Fig. 7), where Dracophyllum is leastabundant between 400 and 300 cal yrs BP, but recoversbetween 200 and 100 cal yrs BP. These three sites are at thelimit for Dracophyllum (the bog sites because of high water
tables; the soil site because it is at the scrub altitudinal limit)and thus might be predicted to be most sensitive. The lowest-altitude site, S-30, reveals a fall in Dracophyllum percentagesfrom c. 500 cal yrs BP, but no subsequent recovery before
Figure 4 Site B-50, percentage diagram. Pollen sum: total pollen and spores. Charcoal fragments as percentageof sum. Dotted line: commencement of farming.
Figure 5 Site S-115, percentage diagram. Pollen sum: total pollen and spores. Charcoal fragments as percentageof sum. Dotted line: commencement of farming.
RECENT VEGETATION HISTORY, CAMPBELL ISLAND 77

Figure 6 Site B-120, percentage diagram. Pollen sum: total pollen and spores. Charcoal fragments as percentageof sum. Dotted line: commencement of farming.
Figure 7 Site S-195, percentage diagram. Pollen sum: total pollen and spores. Charcoal fragments as percentageof sum. Dotted line: commencement of farming.
78 MATT MCGLONE ET AL.
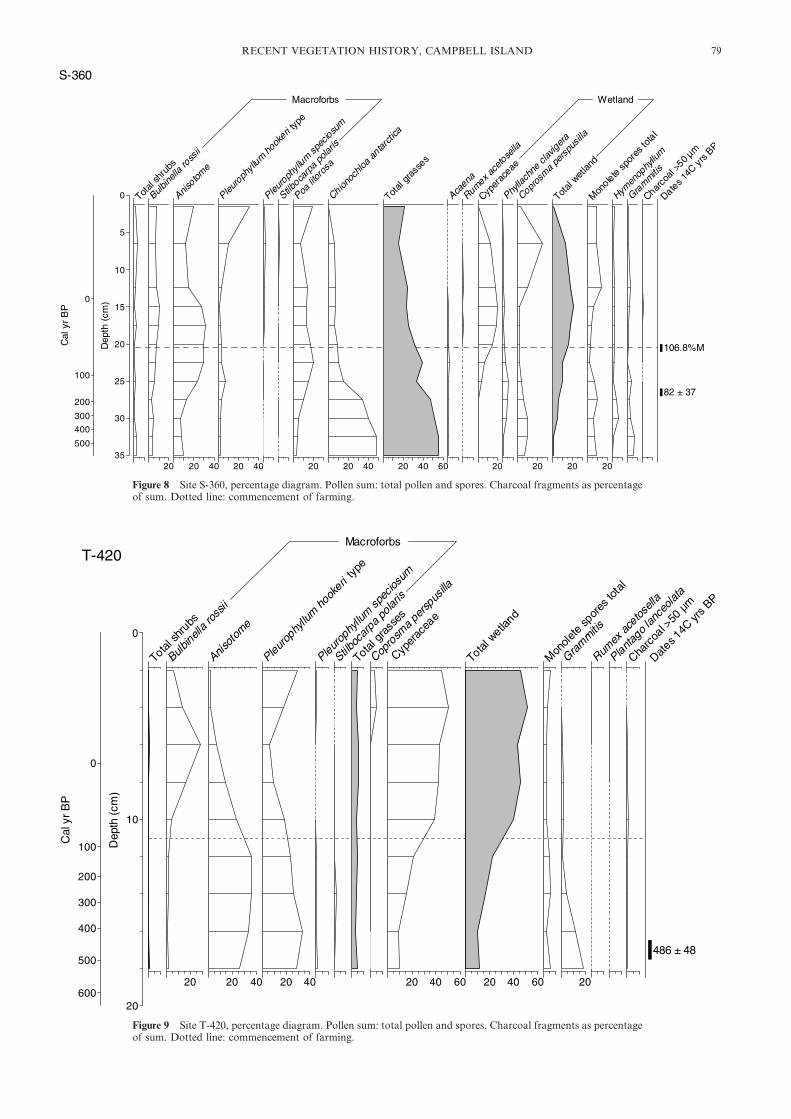
Figure 8 Site S-360, percentage diagram. Pollen sum: total pollen and spores. Charcoal fragments as percentageof sum. Dotted line: commencement of farming.
Figure 9 Site T-420, percentage diagram. Pollen sum: total pollen and spores. Charcoal fragments as percentageof sum. Dotted line: commencement of farming.
RECENT VEGETATION HISTORY, CAMPBELL ISLAND 79

farming (Fig. 3). Macroforbs have increasing representation atS-360 from 200 cal yrs BP, which probably indicates increasingvegetation density at the site (Fig. 8).
4.3. Farming era (55–19 cal yrs BP: cal AD 1895–1931)The beginning of the farming era is marked in all but the twohigh altitude sites by a strong increase in the amount of>50 �m charcoal as a percentage of the total pollen sum. Themacroscopic sieving analysis (see section 4.1) suggests thatthe microscopic pollen-slide charcoal results adequately reflectmacroscopic charcoal concentrations in the sediments andtherefore the occurrence of local fires. The upland bog (B-120)shows several peaks in microscopic charcoal, suggesting thatfiring was more episodic here than elsewhere (Fig. 6). The twohigh altitude sites (S-360 and T-420) have only traces ofcharcoal, suggesting that airborne charcoal was dispersed onlyto a very limited extent (Figs 8 & 9).
Burning is accompanied by major vegetation change. At thescrub-covered sites, scrub abundance declined relative to itsimmediate pre-fire state. Scrub pollen percentages at the twopeat soil sites (S-30 and S-115) indicate a decline of 46% and33% respectively (Figs 3 & 5), but a greater impact occurred atthe two peat bog sites (B-50 and B-120) with declines of 81%and 76% respectively (Figs 4 & 6). There is no significantchange in Dracophyllum abundance at S-195 (Fig. 7). Thepalatable macroforbs Pleurophyllum spp., Anisotome spp. andtussock Chionochloa antarctica declined and the unpalatablemacroforb Bulbinella rossii and the relatively unpalatabletussock Poa litorosa increased. This is strong evidence forgrazing and, as these changes coincide with charcoal evidencefor burning, the fires must have been lit during the farming era.On the peat soil sites, Bulbinella and grass (mainly Poalitorosa) increased as the scrub declined, whereas on the bogsites Chionochloa tussock also declined (although Poa litorosamade up an increasing proportion of what remained) andwetland elements increased strongly.
The two highest altitude sites do not have a pronouncedcharcoal signal. However, at S-360 Bulbinella, Poa litorosa andwetland elements increase indicating the onset of anthropo-genic disturbance. Anisotome percentages fall towards the endof this period (Fig. 8). At T-420 the decline of palatablePleurophyllum hookeri type and Anisotome and the rise ofunpalatable Bulbinella is evidence of grazing even at thehighest altitudes (Fig. 9).
4.4. Post-farmingAll sites show recovery after fire (ceased AD 1931) and grazing(ceased AD 1970–1984). The peat soil sites show the largestresponse to the cessation of fire and grazing. At S-30,Bulbinella and Poa litorosa decline with reducing charcoalinput, and Dracophyllum, Myrsine divaricata and Coprosmaundiff. all recover and total scrub pollen is presently 84% of itspre-fire level (Fig. 3). A more marked response occurs at S-115where the current scrub pollen is now 146% of its pre-fire level(Fig. 5). The bog sites in contrast have recovered only slightly:scrub pollen at B-50 is now 15% of its pre-fire level (Fig. 4) andat B-120, 8% (Fig. 6). The two highest altitude sites show aresurgence of Pleurophyllum but Anisotome has only a moder-ate response and grass levels are essentially unchanged,although composition has altered. At S-360, wetland elementsretreat (Fig. 8) but not at the highest-altitude site T-420(Fig. 9).
5. Discussion
The pollen and charcoal results taken in isolation suggestunequivocally that scrub was abundant up to 200 m on the
sheltered eastern side of the island before human occupation,and fire was rare. Macroforbs were only abundant in thetundra zone and exposed western sites (McGlone 2002) andBulbinella rossii much less common than it was to be duringthe height of the farming era. Scrub, and in particularDracophyllum, was less dense 300–400 cal yrs BP, but itincreased to peak immediately before the farming era. Thereduction of scrub by fire and grazing and the replacement ofpalatable species by grazing-resistant species during the farm-ing era on Campbell Island are clearly reflected in the pollenand microscopic charcoal results. Most pollen sites also showrecovery of the scrub vegetation, which corresponds with thewell-documented increase in density of scrub over the past70 years (Wilmshurst et al. 2004; Bestic et al. 2005) and laterresurgence of palatable macroforbs and Chionochloa antarcticaafter elimination of sheep (Meurk 1982, 1989, and unpublishedobservations).
While the pollen and charcoal results concur with documen-tary evidence for the farming era and scientific observationsmade since, the same can not be said for the status of the islandvegetation before the farming era. It has been proposed thatthe scrub cover of the island was greatly reduced either by cooltemperatures in the late 18th or early 19th century or by earlyactivities of sealers then prevented from recovering by ongoingcool conditions in the late 19th century (Wilmshurst et al. 2004;Bestic et al. 2005). Thus, the subsequent spread of scrubafter the farming era has been at least partially attributed towarming in the 20th century. To resolve this question it isnecessary to examine carefully the different lines of evidencefor the restriction of scrub in the pre-farming era.
5.1. Scrub distribution before farmingThe earliest descriptions of the vegetation are brief impression-istic comments based on a four-day visit of the Erebus andTerror in December 1840 in the course of the Ross Expedition.Hooker noted ‘‘. . . the broader faces of the hills muchexposed, and hence bare of any but a grassy vegetation. . . Inthe narrow sinuous bays, however, the scene is quite changed,for they are often margined by a slender belt of brushwood,with an abundant undergrowth of Ferns, stretching up thesteep and confined gullies.’’ (Hooker 1844, p. 3) and ‘‘. . . a beltof brushwood, composed of some of the tree [Dracophyllum]mentioned as inhabitants of the last visited island, but in a verystunted state form a verdant line close to the beach. This issucceeded by bright green slopes, so studded with Chrysobac-tron [Bulbinella rossii] as to give them a yellow tinge, visible afull mile from the shore.’’ (Ross 1847, pp. 158–9) M’Cormickmade two excursions to different parts of the island mentioningthat there was dense scrub near the coast and in some valleys,with shorter scattered woody vegetation in among the tussockgrassland at altitude, and also the presence of grassy valleysand hollows running down to the coast (M’Cormick 1884, pp.139–43).
Observations were scarce from 1840 until the late19th century (Godley 1970). The French scientific expedition in1874 gave no account of the vegetation but mentioned a largefire (Filhol 1885, quoted in Godley 1970). Armstrong (1868)reported the ‘‘making of fires everywhere’’ in the tussock toattract attention during a visit to determine if any castawayswere on the island. Kirk (1891) visited in the summer of 1890and commented: ‘‘all through the island the ligneous veg-etation presents a starved and stunted appearance, except in afew sheltered positions. It ceases at low altitudes, in manyplaces not rising above 100 ft [30 m]; the highest point I foundit troublesome was under 600 ft [180 m]; . . .’’ (Kirk 1891, p.223). If ‘troublesome’ indicates dense scrub above knee height,this coincides with the current height and density of scrub on
80 MATT MCGLONE ET AL.

the island, which falls below 0·5–m average height at around150 m altitude and is increasingly interspersed with tussock(Bestic & Meurk unpublished observations).
The brief 19th century descriptions can be interpreted asindicating that tall, dense scrub was limited to areas close tothe shoreline or in the valley bottoms, although low sparsescrub was also present near the current scrubline. There can beno doubt that there was some reduction of scrub abundancefrom human activities, as wood cutting would have ac-companied occupation by sealing gangs in the early 19th
century and provisioning ships. Both sealers and members oflater expeditions were accustomed to setting fires to improveaccess (Wilmshurst et al. 2004). However, the early occupationphase was short (AD 1810 to c. AD 1839) and concentrated atthe head of Perseverance Harbour. Although localised anddiscontinuous brief occupations continued, along with visits byships throughout the 19th century, this seems insufficient tohave maintained tussock grassland vegetation below thescrubline.
The pollen-diagram evidence appears to contradict thedocumentary evidence as it reveals scrub was of a similardensity to the present day before the farming era began, evennear the current scrubline at c. 200 m, and extended up to thesame altitudinal limit. There is no evidence from any of ourpeat profiles for early disturbance of the vegetation before theincrease of unpalatable herbs, as might have been expected ifthere was widespread burning of the vegetation during thesealing era. While most sites are not within the zone ofmaximum human impact, sites S-30 and B-50 are at lowaltitude close to the epicentre of human impact and yet show afull scrub cover until the farming era.
There is some indication that the scrub vegetation wasrecovering from a low density state between c. 400–200 cal yrs BP or cal AD 1550–1750, which spans the Little IceAge cooling between AD 1630 and 1680 as inferred frommainland New Zealand tree-ring and speleothem-oxygen-isotope chronologies (Palmer & Xiong 2004; Williams et al.2005). However, it is important to note that although scrubcover decreased, there is no evidence that scrub altitudinallimits changed. Scrub remained common at its current altitu-dinal limit. The effects of the Little Ice Age cooling, if that is infact what is being registered in the pollen profiles, appears tohave been a reduction in scrub dominance across a widealtitudinal and edaphic range rather than an altitudinal depres-sion in scrubline. In other words, during this cool period, scrubwas less able to compete with grasses and macroforbs from sealevel to the scrubline, especially on unfavourable sites, butcapable of maintaining itself on favourable sites.
The apparent abundance of Bulbinella rossii on the upperslopes recorded in the earliest historical account (Hooker 1844)is of interest, as the pollen diagrams presented here and inprevious work (McGlone 2002) do not show Bulbinella domi-nance before settlement. Given that grazing had yet to reducethe tussock cover and encourage the Bulbinella, it seemsimprobable that it was as common as it later became. The‘bright green’ slopes described in any case are not likely to referto the tawny tussock cover, but rather to high-altitudemacroforb-dominant tundra. Bulbinella would have been infull flower at the time of the visit, and the large, brightly yellowracemes may have led to an overestimate of its density.
5.2. Scrub and farmingScrub was massively reduced by fires and woodcutting duringthe farming era (Meurk 1977). Shepherds burnt extensivelywhenever possible to increase the palatability of the tussockand permit better access for sheep (Cockayne 1903; Spence1968; Dingwall & Gregory 2004). Most of the burning
occurred in the southern part of the island where farmingactivities were concentrated and sheep populations highest,particularly in the Perseverance Harbour area (Timms 1978).The shepherds based at the head of Perseverance Harbourneeded large-diameter trunks for firewood, and were accus-tomed to go by boat along the north side of the harbour to cutfrom tall stands (Dingwall & Gregory 2004). Burning stoppedwith the end of farming in 1931.
Fire and sheep grazing rapidly reduced Chionochloaantarctica, the palatable dominant tussock, and the manypalatable herbs, most noticeably the macroforbs Pleuro-phyllum spp., Anisotome spp. and Stilbocarpa polaris. In theirplace, the less-palatable tussock Poa litorosa and fleshy, bittermacroforb Bulbinella rossii vastly extended their range. Theweedy native species Acaena became very common in heavilygrazed areas and exotics spread.
Most of the vegetation photographs (1888–1998) for theisland are derived from sites at the head of PerseveranceHarbour, and they show that scrub was highly restricted at theend of the 19th century. However, late in the farming period(1929–1931) over a third of the island, and all the lower slopes,were estimated by one of the shepherds to have been ‘‘coveredin dense scrub’’ (Spence 1968), which may either be a faultyrecollection or one based on the northern part of the islandwhere scrub may have remained denser. An analysis of changein sequences of photographs showed that scrub increased afterthe end of farming in 1931, particularly in the years before1960/61, and that scrub was not extending above current scrublimits (Wilmshurst et al. 2004).
5.3. Scrub expansionExactly when the scrub cover began to expand and why hasbeen debated. A detailed account of the vegetation fromobservations made in 1941–1945 (Oliver & Sorenson 1951)does not explicitly refer to scrub expansion, although referenceis made to juvenile Dracophyllum shrubs in tussock grassland.Zotov (1965) made field observations and tree-ring counts in1961 and was the first to comment on the spread of Dracophyl-lum scrub. He argued Dracophyllum expansion had begun withgrassland fires set by sealers and whalers, and later by farmers,which gave opportunities for it to enter dense tussock standsbut, with the cessation of fire in 1931, it had stopped spreading.
Meurk argued ‘‘the climate amelioration since the Little IceAge . . . may cause vegetation zones to climb perhaps 100 mabove former levels so that woody vegetation will becomemore prominent than in the immediate past. The destruction ofthe low alpine snow-grass has merely accelerated this trend.’’(Meurk 1977, p. 113). Rudge (1986) also claimed thatDracophyllum scrub was extending altitudinally and becomingdenser as a result of warming climates since the turn of the 20th
century. McGlone et al. (1997) suggested that rising meanannual temperatures since 1950, combined with the drier soilsunder the open fire- and grazing-affected vegetation, hadpromoted the rapid spread of scrub. Wilmshurst et al. (2004)modified this conclusion, suggesting that since the cessation offarming, scrub had spread by infilling its former range butwithout extending above its previous altitudinal limit, butargued that warmer, drier climates had assisted this process.However, Bestic et al. (2005) cast doubt even on this limitedrole for changing climate. They demonstrated through demo-graphic analysis of Dracophyllum scrub on 17 plots acrossenvironmental gradients on the island no change in its altitu-dinal range, and showed that its resurgence followed closely onthe cessation of farming in 1931 and bore no relationship toany warming trend.
Dracophyllum is probably the most warmth-dependent ofthe scrub types but the two lower peat soil sites (S-115 and
RECENT VEGETATION HISTORY, CAMPBELL ISLAND 81

S-30), although showing a clear decline with the onset of fire,also show increasing abundance during the farming era. AtS-115, Dracophyllum, after an initial slight decline, rapidlybecame more abundant than it previously had been (Fig. 5).The pollen and charcoal data thus support the resultsfrom demographic analysis that Dracophyllum regenerationoccurred throughout the fire period and that Zotov (1965)was correct in suggesting that fire opened opportunities forDracophyllum as well as destroying it. As mean annualtemperatures were low during the farming period (c. 0·8(Cbelow the 1951–1980 average for the New Zealand region atthe end of the 19th century; Sturman & Tapper 2006), and mostof the 19th century was warmer (Palmer & Xiong 2004), theseresults suggest that the apparent restriction of scrub noted bypre-farming observers is unlikely to be a result of lowertemperatures, as concluded by Bestic et al. (2005).
5.4. Present vegetation changesAlthough the vegetation of the island has not recovered to itsoriginal state following human modification, successions arewell advanced. Scrub on the peat soils has nearly recovered atthe lower site (S-30), has rebounded to higher densities at themid-elevation site (S-115) and has recovered at the upper site(S-195). The relative rapidity of scrub recovery on CampbellIsland peat soils can be contrasted with the often slow andunpredictable scrub invasion of fire-induced tussock grass-lands in the South Island of New Zealand after their retirementfrom grazing and protection from fire (Mark & Dickinson2003). This may reflect the more complete elimination of scrubseed sources and greater loss of nutrients in comparableinduced tussock grasslands on the mainland. At the twohighest altitude sites, whilst Pleurophyllum spp. have made astrong recovery, Anisotome spp. have yet to make similargains. Pleurophyllum spp. have light, plumed, wind-borneseeds, while Anisotome spp. have heavier, barely winged seedsand this discrepancy in recovery could mainly be due todiffering dispersal ability. However, until 2001 Norway ratswere present in large numbers and it is highly likely that thisrodent consumes seeds of both P. hookeri and Anisotome spp.on Campbell Island, as it is known to have significant effectson plant regeneration through seed predation on a wide rangeof species on New Zealand offshore islands (Towns & Broome2003). Ship rats (Rattus rattus) are known to destroy a highpercentage of Pleurophyllum hookeri racemes on MacquarieIsland (Shaw et al. 2005). If rats preferentially ate the largerand probably more nutritious Anisotome seeds, it is possiblethat they may have retarded the recovery rate of that species.
Post-farming successions have been slow on the two bogsites. While the burning-and-grazing phase initiated a declineof Dracophyllum and tussock, neither recovered rapidly afterthe burning ceased, and both Dracophyllum and tussocksremain sparse on the bog surfaces. Re-establishment of tussockand scrub on bogs is clearly slower than on more fertile peatsoils but vegetation plots established in the 1970s demonstratea slow spread of scrub on the bogs not registered yet in thepollen profiles (CDM unpublished results). On the other hand,burning and grazing seem to have stimulated wetland plants.The phase switch from a dry, scrub-tussock cover to wetlandseen in the pollen results almost certainly resulted from afeedback loop of reduced evapotranspiration, increased sur-face wetness, and less suitable conditions for scrub persistenceor establishment. The tall-tussock grasses and Dracophyllumkeep bog surfaces drier than they otherwise would be throughtranspiration and evaporation of water intercepted by theirleaf surfaces. Work at an experimental catchment in southeast-ern New Zealand showed that tussock cover has evaporativecharacteristics similar to those of forest (Fahey & Rowe 1992),
and palaeoecological work at the same site demonstrated thatthe burning and grazing that accompanied settlement in the1860s led to a substantial increase in surface wetness anddominance by wetland plants of a previously tussock andscrub covered bog (McGlone & Wilmshurst 1999). Slow scruband grassland recovery on the Campbell Island bogs cantherefore be attributed mainly to the increased wetness persist-ing after removal of tall vegetation. The demographic study ofDracophyllum regeneration on Campbell Island supports thisconclusion, as it showed expansion of scrub into wet fen sitesin the years AD 1970–1990 (some 30 years later than inwell-drained sites) coinciding with a prolonged period of lowwinter rainfall (Bestic et al. 2005).
The trajectory of the vegetation seems set to continuetowards increased woodiness below scrubline, increasing domi-nance by Chionochloa and reduced macroforb representationin the tussock–grassland zone, and increased density of macro-forbs above the tussock–grassland zone and in maritime areas.However, the rate of scrub increase outside of the areas whereit is already well established is likely to slow as the cover oftussock and macroforbs thickens reducing the patches ofwell-lit, short moss and turf carpets where Dracophyllumseedlings prefer to establish. Scrub is now likely to spreadmainly through marginal accretion to existing stands, whereasduring the grazing era expansion was possible by means ofseed dispersal into open grassland.
5.5. Future vegetation and climateA major concern worldwide is the biological consequences ofanthropogenic warming. This concern is heightened at sensi-tive ecosystem boundaries such as alpine treeline, and in small,vulnerable, endemic-rich ecosystems. Halloy & Mark (2003)suggested that climate warming over the next century couldput at risk many such herbaceous ecosystems on the NewZealand mainland through rising alpine treelines. CampbellIsland is an interesting case in this regard as it lies at theregional tree limit.
The New Zealand region has experienced marked warmingover the 20th century. There was a period of much coolertemperatures around AD 1900 (c. 0·8(C lower than the 1951–1980 average from 1898 to 1904; Sturman & Tapper 2006) andmean annual temperatures were about 0·5(C lower than the1951–1980 average from 1910 to 1950 in the southwest Pacific(Salinger et al. 1996; Sturman & Tapper 2006), and 0·7(Clower than those of the 21st century. For the first 30 years(1941–1971) of the Campbell Island weather station record,there was no trend in winter or summer mean temperatures.However, since then, mean summer temperatures haveincreased by c. 0·6(C and mean winter temperatures byc. 0·4(C (NIWA 2003; Wilmshurst et al. 2004). Therefore,since the beginning of farming in 1895 there has been at least a0·5(C and possibly as much as a 1(C warming, a trendconsistent with that from other subantarctic islands (Adamsonet al. 1988; Smith & Steenkamp 1990). In terms of vegetationzonation, 0·5(C is equivalent to a 100 m altitudinal shift. Nosuch altitudinal shift has been noted, despite monitoring of thevegetation over the entire period of the instrumental climaterecord. Moreover, as noted in section 5.1, previous coolingseems not to have resulted in an altitudinal shift. This phenom-enon requires explanation as altitudinal limits to woodygrowth are on good evidence believed to be controlled bysummer temperatures (Korner & Paulsen 2004).
The vigorous spread of Dracophyllum scrub below its altitu-dinal limit since the end of farming operations (Wilmshurstet al. 2004; Bestic et al. 2005) took place largely before themid-20th century warming began. Maximum Dracophyllumspread coincided with a decade-long (1957–1967) fall of mean
82 MATT MCGLONE ET AL.

summer temperatures by about 1(C within the longer-termwarming trend (Wilmshurst et al. 2004). Climate change hastherefore only played a minor role in the post-farmingexpansion of scrub (Bestic et al. 2005). Tussock grassland abovescrubline was severely depleted by grazing and burning, andkept low and open by grazing for up to 56 years after farmingceased, thus maintaining opportunities for Dracophyllum spreadabove its current limits. Sheep can have only minor browsingeffects on Dracophyllum as witnessed by its explosive spreadbelow the scrub limit over this time. How can this failure toextend altitudinally be reconciled with the post-1970 warming?
Wilmshurst et al. (2004) point out that a feature of the late20th century warming has been the occurrence of exceptionallywarm winters. Warm winters can be deleterious to woodyplants through increasing their metabolic activity and thusdepleting carbohydrate reserves and de-hardening shoots(Crawford 2000). Warm summers will not compensate for thiswinter depletion if they are not accompanied by decreasedcloud and fog, as the region has very low sunshine hours.Increased summer temperatures may only serve to increaserespiratory losses if the shrubs can make only limited photo-synthetic gain because of low light levels. Finally, wind isclearly a factor in controlling the growth of scrub on the island.The windward side of the island is virtually scrub-free andaverage wind speed increases strongly with altitude. It is poss-ible therefore that the degree of climate change experiencedsince AD 1900 has not yet been sufficiently radical to affect inany significant way the altitude of the scrubline. A sustainedswitch to warmer summers and more summer sunshine hourscertainly would lead to a rise in scrubline. However, if summerdays remain cloudy and windy, and winters become warmer,the most likely result is no altitudinal increase in scrubline.
6. Conclusions
Campbell Island, because of the ready availability of pollen-based histories from virtually every point on the island,historical accounts, and careful recent vegetation and feralanimal monitoring, offers unusual opportunities to examinethe effects of climate and humans on its vegetation cover.Palynological and charcoal analyses from seven peat soiland peat bog sites presented here, combined with previousobservations, permit the following conclusions:
1. Scrub density had recovered by the early 19th century froma low point that probably coincided with a Little Ice Agecooling between cal AD 1630 and 1680.
2. Although early brief accounts of the vegetation andpre-farming photographs from the head of PerseveranceHarbour could be interpreted as implying major restrictionof scrub during the 19th century, palynological analysesshow no evidence for less scrub cover than present in thedecades before the farming era (AD 1895–1931).
3. Widespread burning during the farming era led to a majorreduction in scrub cover. A combination of fire and grazingled to massive increases in the unpalatable macroforbBulbinella rossii and the browse-resistant tussock Poalitorosa, and major declines of the palatable Anisotome spp.,Pleurophyllum spp. Stilbocarpa polaris and Chionochloaantarctica.
4. Scrub has expanded vigorously on drier peat soils since theend of farming activity regaining most of its previousdensity. Peat bogs have had a slower response because withdestruction of their previous woody cover their surfacesbecame much wetter which in turn has inhibited regenera-tion of scrub and tussock.
5. The altitudinal limit to scrub has been remarkably resistantto change. Scrubline was not lower during the Little Ice Agealthough scrub density was somewhat reduced. Nor has thescrubline risen despite at least a 0·5(C rise in mean annualtemperature in the 20th century and severe reduction ofcompeting grasses by grazing.
6. It is likely that future warming will not greatly affect thealtitude of the scrubline on the island unless maximumsummer warmth and sunshine hours increase.
Because islands attract disproportionate attention, theyare often very well documented. It is important to notethat without the multiplicity of evidence available (historicaldocuments, photographic sequences, tree-ring chronologies,detailed local climate records and palynological studies) it islikely that effects of climate change would have been over-stated (as the present authors have done in the past) and thoseof direct human and animal impacts under-emphasised. Mostsimilar regions on mainland New Zealand and elsewhere havefar less documentation, and certainly not all focused on onesmall area. Nevertheless, wide-ranging inferences are oftendrawn from short-term studies and pattern analysis. Thepresent results suggest conclusions as to long-term trajec-tories and their drivers derived from such data will often bemisleading, and more caution is needed in their interpretation.
7. Acknowledgements
We thank our co-workers in this ongoing project, RichardDuncan and Kim Bestic, for discussions and provision ofadditional information. Alison Watkins prepared the pollenand charcoal samples. The New Zealand Department ofConservation provided logistic support while our team workedon Campbell Island. We thank the journal referees RewiNewnham and Dmitri Mauquoy for their helpful suggestions.Funding for this research was provided by the Foundation forResearch, Science and Technology, New Zealand.
8. References
Adamson, D. A., Whetton, P. & Selkirk P. M. 1988. An analysis of airtemperature records for Macquarie Island: decadal warming,ENSO cooling, and Southern Hemisphere circulation patterns.Papers and Proceedings of the Royal Society of Tasmania 122,107–112.
Armstrong, H. 1868. Cruise of the brig Amherst. Official Report. NewZealand Government Gazette, Province of Southland 6, 51–56.
Bell, B. D. 1970. The wild sheep of Campbell Island. Forest and Bird178, 6–10.
Bestic, K. L., Duncan R. P., McGlone M. S., Wilmshurst J. M.& Meurk C. D. 2005. Population age structure and recentDracophyllum spread on subantarctic Campbell Island. NewZealand Journal of Ecology 29, 291–7.
Campbell, I. B. 1981. Soil pattern of Campbell Island. New ZealandJournal of Science 24, 111–35.
Chilton, C. 1909. The subantarctic islands of New Zealand and thehistory of their investigation. In Chilton, C. (ed.) The sub-antarctic islands of New Zealand, Vol. I, XIV–XXXV. Wellington:Government Printer, Philosophical Institute of Canterbury.
Clark, J. S. 1988. Particle motion and the theory of stratigraphiccharcoal analysis: source area, transport, deposition andsampling. Quaternary Research 30, 67–80.
Cockayne, L. 1903. A botanical excursion during midwinter to theSouthern Islands of New Zealand. Transactions and Proceedingsof the New Zealand Institute 36, 225–333.
Cockayne, L. 1909. The ecological botany of the subantarctic islandsof New Zealand. In Chilton, C. (ed.) The subantarctic islands ofNew Zealand, Vol. I, 182–235. Wellington: Government Printer,Philosophical Institute of Canterbury.
Crawford, R. M. M. 2000. Ecological hazards of oceanic environ-ments. New Phytologist 147, 257–281.
Dingwall, P. R. & Gregory, J. G. (eds) 2004. A musterer’s sojournon Campbell Island: the diary of Alfred Austin, 1919–1921.Wellington: Department of Conservation.
RECENT VEGETATION HISTORY, CAMPBELL ISLAND 83

Eden, A. W, 1955. Islands of despair. London: Andrew Melrose.Fahey, B. D. & Rowe, L. K. 1992. Land-use impacts. In Mosely, M. P.
(ed.) Waters of New Zealand, 265–84. Wellington: New ZealandHydrological Society.
Godley, E. J. 1970. Botany of the Southern Zone: Exploration,1847–1891. Tuatara 18, 49–93.
Halloy, S. R. P. & Mark, A. F. 2003. Climate-change effects on alpineplant biodiversity: A New Zealand perspective on quantifying thethreat. Arctic, Antarctic and Alpine Research 35, 248–54.
Hooker, J. D. 1844. The botany of the Antarctic voyage of H.M.Discovery ships Erebus and Terror in the years 1839–43, Vol. 1.London: Reeve.
Kerr, I. S. 1976. Campbell Island. A history. Wellington: A. H. & A. W.Reed.
Kirk, T. 1891. On the botany of the Antarctic islands. Report of theThird Meeting of the Australasian Association for the Advancementof Science, 213–31. Sydney: AAAS.
Korner, C. & Paulsen, J. 2004. A world-wide study of high altitudetreeline temperatures. Journal of Biogeography 31, 713–32.
Laing, R. M. 1909. The chief plant formations and associations ofCampbell Island. In Chilton, C. (ed.) The subantarctic islands ofNew Zealand, Vol. II, 482–92. Wellington: Government Printer,Philosophical Institute of Canterbury.
M’Cormick R. 1884. Voyages of discovery in the Arctic and AntarcticSeas and round the world: being personal narratives of attempts toreach the North and South Poles; and of an open-boat expedition upthe Wellington Channel in search of Sir John Franklin and HerMajesty’s Ships ‘Erebus’ and ‘Terror’, in Her Majesty’s Boat‘Forlorn Hope’, under the command of the author, Vol. 1. London:Sampson Low, Marston, Searle, and Rivington.
Mark, A. F. & Dickinson, K. J. 2003. Temporal responses over30 years to removal of grazing from a mid-altitude snow tussockgrassland reserve, Lammerlaw Ecological Region, New Zealand.New Zealand Journal of Botany 41, 655–68.
McClelland, P. & Tyree P. 2002. Eradication: the clearance ofCampbell Island. New Zealand Geographic 58, 86–94.
McCormac, F. G., Hogg, A. G., Blackwell, P. G., Buck, C. E., Higham,T. F. G. & Reimer P. J. 2004. SHCal04 Southern HemisphereCalibration 0–11.0 cal kyr BP. Radiocarbon 46, 1087–1092.
McGlone, M. S. 2002. The Late Quaternary peat, vegetation andclimate history of the Southern Oceanic Islands of New Zealand.Quaternary Science Reviews 21, 683–707.
McGlone, M. S., Moar, N. T., Wardle P. & Meurk, C. D. 1997. Thelate-glacial and Holocene vegetation and environmental history ofCampbell Island, far southern New Zealand. The Holocene 7,1–12.
McGlone, M. S. & Meurk, C. D. 2000. Modern pollen rain, sub-antarctic Campbell Island, New Zealand. New Zealand Journal ofEcology 24, 181–94.
McGlone, M. S. & Wilmshurst, J. M. 1999. A Holocene record ofclimate, vegetation change and peat bog development, east Otago,South Island, New Zealand. Journal of Quaternary Science 14,239–54.
McNab, R. 1907. Murihiku and the southern islands. Invercargill:William Smith.
Meurk, C. D. 1977. Alien plants in Campbell Island’s changingvegetation. Mauri Ora 5, 93–118.
Meurk, C. D. 1982. Regeneration of subantarctic plants on CampbellIsland following exclusion of sheep. New Zealand Journal ofEcology 5, 51–8.
Meurk, C. D. 1989. Vegetation monitoring, with special reference tothe subantarctic islands of New Zealand. In Craig, B. (ed.)Environmental monitoring in New Zealand, 209–19. Wellington:New Zealand Department of Conservation.
Meurk, C. D., Foggo, M. N. & Wilson, J. R. 1994a. The vegetation ofsubantarctic Campbell Island. New Zealand Journal of Ecology18, 123–68.
Meurk, C. D., Foggo, M. N., Thomson, B. M. & Bathhurst, E. T. J.1994b. Ion-rich precipitation and vegetation pattern on sub-antarctic Campbell Island. Arctic and Alpine Research 26, 281–9.
Meurk, C. D. & Blaschke, P. M. 1990. How representative canrestored islands really be? An analysis of climo-edaphic environ-ments in New Zealand. In Towns, D. R., Daugherty, C. H. &Atkinson, I. A. E. (eds) Ecological restoration of New Zealandislands. Conservation Sciences Publication 2, 252–72. Wellington:New Zealand Department of Conservation.
Meurk, C. D. & Given, D. R. 1990. Vegetation map of CampbellIsland. Christchurch, New Zealand: DSIR Land Resources, De-partment of Scientific and Industrial Research.
Millspaugh, S. H. & Whitlock C. 1995. A 750-year fire history basedon lake sediment records in central Yellowstone National Park,USA. The Holocene 5, 283–292.
Moar, N. T. 1973. Late Pleistocene vegetation and environment insouthern New Zealand. In van Zinderen Bakker, E. M. (ed.)Palaeoecology of Africa and the surrounding islands and Antarctica8, 179–98. Cape Town: Balkema.
Moore, P. D., Webb, J. A. & Collinson, M. E. 1991. Pollen analysis,2nd edition. Oxford: Blackwell.
Moore, P. J. & Moffat, R. D. 1990. Research and managementprojects on Campbell Island 1987–1988. Science and ResearchInternal Report 57. Wellington: New Zealand Department ofConservation.
NIWA (National Institute of Water and Atmospheric Research) 2003.National climate database (http://www.niwa.co.nz/services/clidb).
Oliver, R. L. & Sorenson, J. H. 1951. The vegetation. In Botanicalinvestigations on Campbell Island. Cape Expedition Series, Bulletin7 (1), 5–24. Wellington: New Zealand Department of Scientificand Industrial Research.
Palmer J. G. & Xiong L. M. 2004. New Zealand climate over the last500 years reconstructed from Libocedrus bidwillii Hook. f. tree-ring chronologies. Holocene 14, 282–9.
Redwood, R. 1950. Forgotten islands of the South Pacific: the story ofNew Zealand’s southern islands. Wellington: Reed.
Ross, J. C. 1847. A voyage of discovery and research in the Southern andAntarctic regions during the years 1838–43, Vol, I. London: JohnMurray.
Rudge, M. R. 1986. The decline and increase of feral sheep (Ovis ariesL.) on Campbell Island. New Zealand Journal of Ecology 9,89–100.
Salinger, M. J., Allan, R., Bindoff, N., Hannah, J., Lavery, B., Lin, Z.,Lindesay, J., Nicholls, N., Plummer, N. & Torok, S. 1996.Observed variability and change in climate and sea level inAustralia, New Zealand and the South Pacific. In Bouma, W. J.,Pearman, G. I. & Manning M. R. (eds) Greenhouse: coping withclimate change, 100–126. Melbourne: CSIRO.
Shaw, J. D., Hovenden, M. J. & Bergstrom, D. M. 2005. The impactof introduced ship rats (Rattus rattus) on seedling recruitment anddistribution of a subantarctic megaherb (Pleurophyllum hookeri).Austral Ecology 30, 118–25.
Smith, V. R. & Steenkamp, M. 1990. Climatic change and its ecologi-cal implications at a subantarctic island. Oecologia 85, 14–24.
Sorensen, J. H. 1950. The Royal Albatross. Cape Expedition SeriesBulletin No. 2. Wellington: Government Printer, Department ofScientific and Industrial Research.
Spence, A. 1968. A story of the Campbell Islands. Review: TussockGrasslands and Mountain Lands Institute 15, 63–75.
Stuiver, M., Reimer, P. J. & Reimer, R. W. 2005. CALIB 5.0. [wwwprogram and documentation].
Sturman, A. P. & Tapper, N. J. 2006. The weather and climate ofAustralia and New Zealand, 2nd edition. Oxford: Oxford Univer-sity Press.
Timms, J. 1978. Marlborough whalers at Campbell Island, 1909–1916:a narrative based on the recollections of J Timms, Kerr, I. S. &Judd, N. (eds). Wellington: Government Printer, Department ofLands and Survey.
Towns, D. R. & Broome, K. G. 2003. From small Maria to massiveCampbell: forty years of rat eradications from New Zealandislands. New Zealand Journal of Zoology 30, 377–98.
Wardle, P. 1987. Dracophyllum (Epacridaceae) in the Chatham andsubantarctic islands of New Zealand. New Zealand Journal ofBotany 25, 107–14.
Williams, P. W., King D. N. T., Zhao, J. X. & Collerson, K. D. 2005.Late Pleistocene to Holocene composite speleothem O-18 andC-13 chronologies from South Island, New Zealand – did a globalYounger Dryas really exist? Earth and Planetary Science Letters230, 301–17.
Wilmshurst, J. M., Bestic, K. L., Meurk, C. D. & McGlone, M. S.2004. Recent spread of Dracophyllum scrub on subantarcticCampbell Island, New Zealand: climatic or anthropogenic ori-gins? Journal of Biogeography 31, 401–413.
Zotov, V. D. 1965. Grasses of the subantarctic islands of the NewZealand region. Records of the Dominion Museum 5 (15), 101–146.
MS received 10 February 2006. Accepted for publication 9 March 2007.
84 MATT MCGLONE ET AL.




















