
Circalunare Steuerung der Populationsdynamik der Süß- wassermeduse Craspedacusta sowerbii (=...
34
197 Lauterbornia 67: 197-230, D-86424 Dinkelscherben, 2009-08-17 Circalunare Steuerung der Populationsdynamik der Süß- wassermeduse Craspedacusta sowerbii (= sowerbyi) (Lankester, 1880) Circalunar control of population dynamics of the freshwater jellyfish Cras- pedacusta sowerbii (= sowerbyi) (Lankester, 1880) Detlef E. Peukert Mit 7 Abbildungen und 4 Tabellen Schlagwörter: Craspedacusta, Hydrozoa, Cnidaria, Westerwald, Hessen, Deutschland, See, Lunar- periodik, Populationsdynamik, Strobilation, Verbreitung Keywords: Craspedacusta, Hydrozoa, Cnidaria, Westerwald, Hesse, Germany, lake, lunar period- icity, population dynamics, strobilation 2004 bis 2008 wurde das plötzliche Auftreten der Süßwassermeduse Craspedacusta sowerbii im Steinbruchsee Winkel bei Herborn-Schönbach in Hessen bei regelmäßigen Tauchgängen un- tersucht. Dabei zeigte sich, dass C. sowerbii ab dem ersten Vollmondtermin nach Sommerson- nenwende erschien und an den folgenden drei Vollmondterminen wiederholt in Massen auf- trat. Eine Metauntersuchung von Fundmeldungen im Internet bestätigte dieses Ergebnis. Es wird diskutiert, dass die im gleichen Abstand auftretende viermalige Massenproduktion von der Crustacea-Planktonpopulation zunächst wohl quantitativ allerdings nicht qualitativ ver- kraftet werden kann, da offenbar zur Populationsregeneration gleichartige Stadien selektiv der Prädation des Polypen und der Meduse zum Opfer fallen, so dass nach wenigen Jahren wiederholten Massenauftretens deren Abundanz wahrnehmbar zurückgeht, was sich in den Abundanz-Werten sowie im Ausbleiben der sommerlichen Klarphase zeigt. Eine Betrachtung der globalen Ausbreitung unter evolutivem Aspekt schließt die Untersuchung ab. Es wird an- geregt, die Folgen der Prädation der betreffenden Taxa unter dem circalunaren Aspekt weiter zu untersuchen. Between 2004 and 2008 the author was examining the sudden appearance of the freshwater medusa Craspedacusta sowerbii in the quarry pond Winkel near Herborn Schönbach in Hesse/Germany by scuba diving, in the course of which C. sowerbii was found out to appear from the first full moon after the summer solstice on, its number increasing in the following three full moon periods, which is indicated by four peaks of its abundance. This observation was confirmed by a meta research of findings published on the internet. The article discusses that the large number of C. sowerbii occurring four times at regular intervals may be coped with by the Crustacea plankton population as regards the quantity though not the quality as, supposedly, the same stages necessary for the regeneration of the population are consumed se- lectively by both the polyp and the medusa. Thus, after few years of repeated mass occurrence of C. sowerbii, its abundance is declining perceivably, which shows by its abundance as well as the absence of the summer clear water stage in phytoplankton. Finally, the author looks at the global spread from the evolutionary aspect. He suggests that the implications of the predation of the taxa concerned should be examined more closely, taking the circa-lunar aspect into account.
-
Upload
uni-marburg -
Category
Documents
-
view
1 -
download
0
Transcript of Circalunare Steuerung der Populationsdynamik der Süß- wassermeduse Craspedacusta sowerbii (=...

197
Lauterbornia 67: 197-230, D-86424 Dinkelscherben, 2009-08-17
Circalunare Steuerung der Populationsdynamik der Süß-wassermeduse Craspedacusta sowerbii (= sowerbyi)(Lankester, 1880)
Circalunar control of population dynamics of the freshwater jellyfish Cras-pedacusta sowerbii (= sowerbyi) (Lankester, 1880)
Detlef E. Peukert
Mit 7 Abbildungen und 4 Tabellen
Schlagwörter: Craspedacusta, Hydrozoa, Cnidaria, Westerwald, Hessen, Deutschland, See, Lunar-periodik, Populationsdynamik, Strobilation, VerbreitungKeywords: Craspedacusta, Hydrozoa, Cnidaria, Westerwald, Hesse, Germany, lake, lunar period-icity, population dynamics, strobilation
2004 bis 2008 wurde das plötzliche Auftreten der Süßwassermeduse Craspedacusta sowerbii imSteinbruchsee Winkel bei Herborn-Schönbach in Hessen bei regelmäßigen Tauchgängen un-tersucht. Dabei zeigte sich, dass C. sowerbii ab dem ersten Vollmondtermin nach Sommerson-nenwende erschien und an den folgenden drei Vollmondterminen wiederholt in Massen auf-trat. Eine Metauntersuchung von Fundmeldungen im Internet bestätigte dieses Ergebnis. Eswird diskutiert, dass die im gleichen Abstand auftretende viermalige Massenproduktion vonder Crustacea-Planktonpopulation zunächst wohl quantitativ allerdings nicht qualitativ ver-kraftet werden kann, da offenbar zur Populationsregeneration gleichartige Stadien selektivder Prädation des Polypen und der Meduse zum Opfer fallen, so dass nach wenigen Jahrenwiederholten Massenauftretens deren Abundanz wahrnehmbar zurückgeht, was sich in denAbundanz-Werten sowie im Ausbleiben der sommerlichen Klarphase zeigt. Eine Betrachtungder globalen Ausbreitung unter evolutivem Aspekt schließt die Untersuchung ab. Es wird an-geregt, die Folgen der Prädation der betreffenden Taxa unter dem circalunaren Aspekt weiterzu untersuchen.
Between 2004 and 2008 the author was examining the sudden appearance of the freshwatermedusa Craspedacusta sowerbii in the quarry pond Winkel near Herborn Schönbach inHesse/Germany by scuba diving, in the course of which C. sowerbii was found out to appearfrom the first full moon after the summer solstice on, its number increasing in the followingthree full moon periods, which is indicated by four peaks of its abundance. This observationwas confirmed by a meta research of findings published on the internet. The article discussesthat the large number of C. sowerbii occurring four times at regular intervals may be copedwith by the Crustacea plankton population as regards the quantity though not the quality as,supposedly, the same stages necessary for the regeneration of the population are consumed se-lectively by both the polyp and the medusa. Thus, after few years of repeated mass occurrenceof C. sowerbii, its abundance is declining perceivably, which shows by its abundance as well asthe absence of the summer clear water stage in phytoplankton. Finally, the author looks at theglobal spread from the evolutionary aspect. He suggests that the implications of the predation ofthe taxa concerned should be examined more closely, taking the circa-lunar aspect into account.

198
1 EinleitungDer Autor ist Sporttaucher und führt im Steinbruchsee Winkel bei Herborn inOberhessen, einem Gewässer des Hessischen Tauchsportverbandes, regelmäßigTauchgänge durch. Dabei wurde eine Population der Süßwassermeduse Craspe-dacusta sowerbii entdeckt und im Rahmen der Neobiota-Watcher Aktion 2005(vgl. Fritz & al, 2007: Quarry pond Winkel Gießen 2005 - Verband DeutscherSporttaucher 2006) gemeldet. Das unregelmäßige Auftreten der Medusen führtezu der Frage, ob und welche zeitlichen Muster hierfür verantwortlich sind. Da-zu wurden wesentliche Umstände ihres Vorkommens über fünf Jahre beobach-tet und protokolliert, soweit dies während der Tauchgänge ohne apparativenAufwand möglich war.Bearbeitet wurden folgende Fragestellungen:1. Wann im Jahresverlauf und unter welchen Umständen treten die Medusenauf?- Jahreszeitliches Vorkommen, Wassertemperatur, Mondphasen, Sonnenstandund Tageslänge2. Welchen Teil des Wasserkörpers nutzen die Medusen?- räumliche Besiedlung des Wasserkörpers3. Zu welcher Zeit halten sich die Medusen wo aus welchem Grund auf?- Tagesrhythmik der Medusen im Wasserkörper- Wanderbewegungen der Medusen im Wasserkörper4. Welches sind Faktoren für die Ausbreitung von Craspedacusta sowerbii?5. Wie lassen sich diese bezogen auf die Evolution der Species einordnen?
Im folgenden werden die Ergebnisse der Untersuchungen vorgestellt; sie be-leuchten die Steuerung der Populationsdynamik durch abiotische Faktoren.Die offensichtlichen trophischen Beziehungen im See zwischen Phytoplankton,Zooplankton und Medusen konnten noch nicht ausreichend untersucht wer-den, es werden daher nur die Grundtendenzen erörtert.
Zur Einordnung der Ergebnisse wird eine Übersicht über den Stand der For-schung zur Biologie von Craspedacusta gegeben.
2 ForschungsstandBei Craspedacusta sowerbii handelt es sich gemäß breiter Auffassung um ein Neozo-on, das sich inzwischen fest etabliert hat. Fritz & al. 2007 fassen den aktuellenUntersuchungsstand zur Verbreitung der Art in Deutschland zusammen undgeben eine Übersicht zur Nomenklatur.
Bei der Feststellung der Besiedlung des Wasserkörpers von der Oberflächebis in eine Tiefe von 20 Fuß (= 6,10 m) beruft sich Scott (1963) auf Apnoetau-cher. Hubschmann & Kishler (1972) beschreiben ein weit verbreitetes Vorkom-

199
men der Polypen im westlichen Eriesee. Sie siedeln auf Felsproben von allenuntersuchten Stationen in einer Tiefe von 2,50-9,00 m.
Craspedacusta sowerbii bevorzugt oligotrophe Seen, ist aber auch in mesotro-phen Gewässern zu finden. An ihren Biotop stellt sie den Anspruch ausreichen-der Sauerstoffversorgung aufgrund windbedingter Strömungen die den Wasser-körper durchmischen, Reichtum an Zooplankton als Ernährungsgrundlage undFels als Siedlungssubstrat das nicht versanden kann für die Polypenstadien. Dieoptimale Temperatur für die Polypen, aus denen sich die Medusen entwickeln,liegt zwischen 19 und 25 °C. Fällt die Temperatur unter 10 °C wechseln diePolypen zu Überdauerungskörpern, indem sie sich zu einer kompakten Kugelzusammenziehen (Schill 2007). Unterhalb von 16 °C verschwinden die Medu-sen (Acker & Muscat 1976, Jung Hee Park 1998). Tappenbeck (2003) nennt fürdie Entwicklung der Medusen eine Temperatur von mindestens 19-20 °C, amgünstigsten 25-27 °C. Lundberg & Svensson (2003) gehen auf das Medusenvor-kommen in warmen Seen Schwedens näher ein. Arbačiauskas & Lesutienė(2005) betonen ebenfalls, dass die Medusenproduktion an hohe Wassertempera-tur gekoppelt ist, räumen aber ein, dass auch andere Umweltfaktoren wichtigsind.
Viele Autoren liefern Fundmeldungen und Hypothesen zur Verbreitung derArt rund um den Globus. Unter den Publikationen seien pars pro toto die fol-genden Forschungsfelder aufgeführt:
Dodson & Cooper (1983), Jankowski & al. (2005) und Pérez Bote & al.(2006) und andere untersuchen die trophischen Bedürfnisse. Jung Hee Park(1998) stellt fest, dass die relative Abundanz von Craspedacusta sowerbii als Ernäh-rungmuster der Zooplanktonblüte wie ein Zeitschatten folgt. Trotz grundle-gender Arbeiten von Dodson & Cooper (1983) zur Rolle von Craspedacusta sower-bii als invasive Art im Nahrungsnetz der neuen Lebensgemeinschaften, diefeststellen, dass die hohe Abundanz der Prädatoren einen starken Effekt auf dasherbivore Crustacea-Zooplankton hat (was Jankowski & al. 2005 bestätigen),scheint die Aussage von Arbačiauskas & Lesutienė (2005) weiterhin gültig zusein, dass die Rolle der Süßwassermeduse in Nahrungsnetzen noch nicht hinrei-chend untersucht ist, trotz der differenzierten Untersuchungen zum Fraßver-halten der Medusen von Pérez Bote & al. (2006) unter Einbezug von 6 Copepo-da-, 5 Cladocera- und 8 Rotatoria-Arten.
Kuhl & Kuhl (1956) zeigen Polypenkolonien von Craspedacusta sowerbii und dieBildung von Wanderfrusteln im Zeitrafferfilm. Gen-Yu Sasaki (1999) klärt diesexuelle Reproduktion der Süßwassermeduse im Detail auf. Farkašova & Stlou-kal (2007) schließen sich der Vermutung von Ludwig & al. (1979) an, nach derstrobilierende Polypen durch Hormonabgabe ihre Nachbarpolypen zur Strobi-lation stimulieren, wodurch das Massenauftreten der Medusen zu erklären ist.

200
Der Vorgang alternierender Polypen- und Medusengenerationen wird als Meta-genese bezeichnet.
Fritz & al. (2007) und Petrusek & al. (2005) messen einer notwendigen sexu-ellen Vermehrung für die Verbreitung der Art eine untergeordnete Bedeutungzu, was an dem Auftreten meist nur eines Geschlechts aufgrund eines Einwan-derungsereignisses im gleichen Land bei gleichzeitig hoher Mobilität festge-macht wird. Das nehmen Lundberg & Svensson (2003) als Grundlage zur Dis-kussion einer "Gründereffekt-Hypothese" als Ausbreitungsstrategie der Art.Trotz der Anregung zur Re-Evaluation der Taxa von Craspedacusta scheint diemorphologische wie auch die genetische Plastizität hoch zu sein, was neuereUntersuchungen von Fritz et al. (2009) belegen.
Die Medusen-Saison fällt bei nahezu allen Autoren in die von Dexter & al.(1949) für das Vorkommen in Ohio und Pennsylvania formulierte Zeitspannevon Juli bis Oktober mit der höchsten Abundanz im September, obwohl auchFunde schon im Mai und erst im November getätigt wurden.
Bezogen auf Autoren der Südhalbkugel bezieht sich die Saison entsprechendauf die Monate Januar bis April. Das bestätigen Silva & Roche (2007) auchwenn in Brasilien etwa Meldungen für Juli und September bis Januar vorliegen.Entsprechend wie Dexter & al. (1949) für die Nordhalbkugel nennt Fish (1975)die Monate Februar und März als Funddaten am Beispiel der Nord-Insel vonNeuseeland und beschreibt für Februar eine hohe Frequenz an vielen Stellendes Sees auf der Südhalbkugel.
Dennoch wird das Auftreten der Süßwassermeduse als sporadisch und un-vorhersagbar angegeben (Peard 2002) sowie (New Hampshire Department ofEnvironmental Services 2004).
3 Untersuchungsort und Methoden3.1 Der SeeDer Steinbruchsee Winkel liegt zwischen Herborn-Schönbach und Driedorf-Roth im Westerwald (Landkreis Lahn-Dill/Hessen), 465 m über NN. Der Seeerstreckt sich zwischen 50°39’24.30’’N (Nordseite) und 50°39’19.90’’N (Südsei-te) sowie 8°13’07.60’’E (Westseite) und 8°13’33.35’’E (Ostseite), ermittelt nachGoogle Earth (2009). In dem ehemaligen Steinbruch wurde zwischen 1920 und1979 Säulenbasalt abgebaut. Der See ist heute Gewässer des Hessischen Tauch-sportverbandes.
Die größte Ausdehnung beträgt in der Länge 510 m, in der Breite 130 m.Der schmale, längliche östliche Teil, der überwiegend von Anglern der Angel-sportgruppe 1976 Schönbach e.V. genutzt wird, besitzt eine Länge von 310 mund eine Breite von 40 m. Der von Tauchern genutzte fast ovale westliche Teil

201
ist 200 m lang und 130 m breit. Der See hat eine Tiefe von 10 m imAnglerbereich bis 32 m im Taucherbereich.
Der betauchbare Westteil des Sees weist ein nahezu senkrecht abfallendesSteilufer auf. Nur in den oberen 10 m hat der Abhang eine Neigung durch Ab-raum und Blockhalden. Auf etwa 18 m Tiefe breitet sich ein horizontales Ni-veau bis zu einer zentralen Kante aus, von der aus ein weiterer Steilabfall bis ins30-m-"Loch" führt. Die Grundfläche des "Lochs" stellt dann das tiefste Niveaudar. Eine ehemalige Fahrstraße führt vom Einstieg (Abb. 1: vor "b") bis an die10-m-Kante (Abb. 1: bei "9 m"). Eine weitere steigt aus dem Loch in Richtungdes lang gestreckten Ostteils auf.
Abb. 1: Lageplan des Steinbruchsees Winkel (unten Norden, oben Süden)
3.2 Untersuchungsmethoden3.2.1 Erfassung von Lebensraum und Abundanz von CraspedacustasowerbiiDie Tauchgänge wurden während der Öffnungszeiten des Sees mittwochs zwi-schen 19 und 20 h und am Wochenende zwischen 10 und 11 h durchgeführt.Beobachtungszeitraum war die Tauchsaison der Jahre 2004 bis 2008. DieTauchgänge verteilten sich wie folgt: 2004: 47, 2005: 44, 2006: 40, 2007: 35,2008: 43. Das Jahr 2008 wurde zur Kontrolle für die bis dahin gewonnenen Er-kenntnisse genutzt. Der Einstieg geschah immer von der Nordseite an der ehe-maligen Fahrstraße aus in Richtung Süden bis zur Lochkante auf 18 m. Vonhier aus verliefen die Tauchgänge entweder in Richtung Osten oder Westen, ei-nige auch noch weiter nach Süden auf der 30 m Sohle durch das Loch. Die Stre-cken fielen unterschiedlich weit aus bis die Austauchphase begann, in der die

202
beim Abtauchen notierten Beobachtungen geprüft wurden. Auf diese Weisekonnten durch die Frequenz der Taucher am Einstieg bedingte Beeinflussungenausgeschlossen und subjektive Wahrnehmungen zu Tauchbeginn und nach un-ten zu Tauchende hin in die andere Richtung nach oben relativiert werden. JeTauchgang lag den Beobachtungen somit eine Kontrollbeobachtung an einerweiteren Stelle zugrunde. Notiert wurde jeweils entweder von oben oder vonunten kommend das erste zu beobachtende Individuum und die Wassertiefemit Hilfe des Tiefenmessers festgestellt. In dem Bereich, in dem die Individuen-zahl offensichtlich am höchsten war, wurde die Bonitierung vorgenommen.
Die Schätzung der Abundanz der Individuen wurde ebenfalls in zwei Stufenin horizontaler Richtung gegen den Wasserkörper vorgenommen. Zur Orien-tierung wurden die Individuen in einem Wasserkörper von 50 x 50 x 50 cm aus-gezählt, der an Hand von Körpermaßen (Armlänge und weitere) abgestecktwurde; in einem zweiten Schritt wurde die Zählung in einem Kubikmeter veri-fiziert und diente als Bezugsvolumen für ein Bonitierungs(Schätz)verfahren.Nachdem sich nach einem Jahr herausstellte, dass die Bonitierung gegenüberder Auszählung eines Viertel Kubikmeters nur unwesentlich mit Fehlernbehaftet war, wurde auf das aufwändige Auszählen verzichtet und nur noch dasBonitierungsverfahren angewandt. Dieses beruht auch auf eigenen Erfahrungen(Peukert 1982, 1985 und 1988). Für die Kennzeichnung der Abundanz vonCraspedacusta sowerbii wird die Skala von Stark (2005) in abgewandelter Formverwendet (Tab. 1), ein Verfahren ähnlich der Abundanzbestimmung nach DIN38 410-1 (2004). Nominale Abundanzangaben der Autoren im Rahmen derMetauntersuchung wurden den 5 unterschiedenen Abundanzstufen zugeordnet.
Tab. 1: Definition der Abundanzstufen für das Vorkommen von Craspedacusta sower-bii, verändert nach Stark (2005)Stufe Grau-
stufe (%)Abudanz(Ind./m³)
Definition
1 6,25 1-3 sehr selten, vereinzelt, 1-3 Individuen oder eine kleine Individuengruppe2 12,5 4-10 selten, 4-10 Individuen oder kleine Individuengruppen3 25 10-30 verbreitet, regelmäßig in mäßiger Individuenzahl anzutreffen; höchstens 25 % des Maximal-
vorkommens4 50 30-60 häufig, überall in großer Zahl, teilweise dichte Bestände; etwa 50% des Maximalvorkommens5 75 > 60 sehr häufig, massenhaft, zusammenhängende Bestände; mehr als 75% des Maximalvorkom-
mens
3.2.2 Erfassung von Kleinkrebsen als Hauptnahrung der MedusenDie quantitative Erfassung der Abundanz der Copepoda und Cladocera folgtTabelle 1 (Stark 2005), wobei die geschätzte Individuenzahl auf einen Liter be-zogen wurde. Die Volumenschätzung wurde anhand des Lichtkegels derTauchlampe in Verbindung ebenfalls mit Körpermaßen vorgenommen. Mitunter Wasser geöffneten Plastikflaschen wurden zur Bestimmung der Hauptar-ten Wasserproben entnommen.

203
3.2.3 Überprüfung der Hypothese der circalunaren Häufung vonCraspedacusta sowerbii durch eine orientierende MetauntersuchungZur Überprüfung des gehäuften Auftretens der Medusen um den Vollmondter-min herum, wurden in einer Metauntersuchung die Fundmeldungen in der Li-teratur in Bezug zu ihren nächstliegenden Vollmonddaten gesetzt und als Dia-gramm aufbereitet (Tab. 2, Abb. 7). Von den im Internet recherchiertenFunddaten haben 81 Autoren aus aller Welt - auch von der Südhalbkugel - eineTagesangabe ihres Fundes gemacht. Allerdings nennen viele Autoren entwedernur den Fundmonat oder beschreiben den Zeitpunkt ihrer Funde innerhalb desgenannten Monats nur allgemein; diese konnten nicht in die Metauntersuchungeinbezogen werden. Bei den exakten Datierungen kann man allerdings davonausgehen, dass ein repräsentativer Querschnitt der Funddaten erreicht wurde,um einen Trend zu ermitteln. Da keiner der Autoren einen circalunarenRhythmus weder in Bezug auf das Massenauftreten noch die Strobilationäußert, sind Datenverfälschungen unwahrscheinlich. Vielmehr wurden dieFunde fast immer zufällig gemacht: "Usually the medusa was discovered bychance or when looking for someone other organism of interest" (Fritz et al.2007). Wegen des häufig geäußerten Erstaunens über die Funde wurde in derRegel zeitnah publiziert.

Tab.
2:
Fund
mel
dung
en v
on C
rasp
edac
usta
sow
erbi
i in
Bez
ug z
umn
näc
hste
n V
ollm
ondt
erm
in (
in T
agen
) in
der
Lite
ratu
r. D
ieni
chtn
umer
isch
en M
enge
nang
aben
wur
den
Abu
ndan
zstu
fen
nac
h Ta
b. 1
zug
eord
net.
Bei
feh
lend
en A
ngab
en,
wur
de d
ie A
bun-
danz
stuf
e 1
ange
nom
men
. Pub
likat
ions
datu
m =
Fun
ddat
umFu
nd-
datu
mVo
ll-m
ond
Diff
e-re
nz (d
)Fu
ndor
tG
eogr
aphi
sche
Lage
Auto
r/Beo
bach
ter
Men
gena
ngab
eAb
un-
danz
Inte
rnet
quel
le(le
tzte
r Zug
riff a
m 3
0.4.
2009
)19
95-
08-2
419
95-
08-1
0+1
4D
e Za
an b
ei ‘t
Kal
f:D
e P
ool
Nie
derla
nde
Ed Z
ijpen
kele
exe
mpl
aren
1ht
tp://
ww
w.k
nnv.
nl/z
aanw
ater
/kw
alle
n.ht
m
1928
-09
-14
1928
-08
-31
+14
Stal
lwor
th L
ake
bei
Tusc
aloo
sa
Ala
bam
a U
SA
Willi
am E
. Whi
teke
ine
Ang
abe
1ht
tp://
ww
w.js
tor.o
rg/p
ss/1
5369
92 N
otes
on
a Fr
esh-
Wat
er M
edus
aFo
und
in S
tallw
orth
Lak
e, T
usca
loos
a, A
laba
ma
Willi
am E
. Whi
teB
iolo
gica
l Bul
letin
, Vol
. 59,
No.
2 (O
ct.,
1930
), pp
. 222
-232
(arti
cle
cons
ists
of 1
1 pa
ges)
Pub
lishe
d by
: Mar
ine
Bio
logi
cal L
abor
ator
y(fi
rst o
bser
ved)
1949
-08
-20
1949
-08
-08
+12
Rou
nd L
ake
Ohi
o U
SA
Dex
ter,
Surra
rrer &
Dav
isgr
eat a
bund
ance
4ht
tps:
//kb.
osu.
edu/
dspa
ce/b
itstre
am/1
811/
3726
/1/V
49N
06_2
35.p
df
1985
-08
-12
1985
-07
-31
+12
Als
dorfe
r Wei
her
Deu
tsch
land
Wol
fgan
g V
oigt
meh
rere
hun
dert
Exe
mpl
are
5ht
tp://
ww
w.n
abu-
aach
en-
land
.de/
doku
men
tatio
nen/
arch
iv_n
eubu
erge
r.htm
#sue
ss19
99-
09-0
819
99-
08-2
7+1
2Au
hafe
n H
ard
Bode
nsee
,Ö
ster
reic
hin
atur
a Er
lebn
isN
atur
scha
u G
mbH
ein
Indi
vidu
um1
http
://in
atur
a.at
/wis
sen/
gem
_941
5.sh
tm
2007
-09
-27
2007
-09
-15
+12
Löbe
jün
Sach
sen-
Anha
ltD
euts
chla
ndS
imon
iske
ine
Ang
abe
1ht
tp://
ww
w.a
ktiv
-urla
ub-m
ache
n.de
/vid
eos/
hydr
ozoe
nen/
tauc
hen-
loeb
ejue
n-27
0920
0820
02-
08-0
320
02-
07-2
4+1
0D
onca
ster
’sH
atfie
ld W
ater
Par
kYork
shire
Gro
ßbrit
anni
enC
olin
How
es27
Exe
mpl
are
3ht
tp://
ww
w.y
nu.o
rg.u
k/je
llyfis
h.ht
m
2002
-10
-01
2002
-09
-21
+10
Nyk
öpin
gsån
Söde
rman
land
Fred
rik L
undb
erg
kein
e A
ngab
e1
http
://w
ww
.nrm
.se/
utst
alln
inga
rcos
mon
ova/
jour
hava
ndef
orsk
are/
jou
rhav
ande
biol
og/ry
ggra
dslo
sadj
ur/s
otva
ttens
man
eter
.125
8.ht
ml
2008
-07
-28
2008
-07
-18
+10
Štěr
kovně
v H
lučí
nTs
chec
hien
Kubačk
ake
ine
Ang
abe
1
2002
-09
-01
2002
-08
-23
+9H
jorte
n B
otor
p-st
röm
men
, Sto
raVä
llan
Dal
älve
n,R
ösjö
n N
orrs
tröm
,Lå
ngba
n G
öta
älv
Smål
and,
Dal
ar-
na, U
ppla
nd,
Värm
land
Sch
wed
en
Lund
berg
&S
v ens
son
män
gder
av
med
usor
3ht
tp://
ww
w.n
rm.s
e/ut
stal
lnin
garc
osm
onov
a/jo
urha
vand
efor
skar
e/jo
urh
avan
debi
olog
/rygg
rads
losa
djur
/sot
vatte
nsm
anet
er.1
258.
htm
l
2004
-10
-07
2004
-09
-28
+9P
rose
rpin
aR
eser
voir
Span
ien
José
Lui
s P
érez
-Bo
te e
t al.
76 in
divi
dual
s/m
2 of
sur
face
wat
er /
net s
ampl
es 1
72in
divi
dual
s
4ht
tp://
ww
w.k
bini
rsnb
.be/
en/in
stitu
te/a
ssoc
iatio
ns/rb
zs_w
ebsi
te/b
jz/b
ack/
pdf/B
JZ%
2013
6(2)
/136
_2_1
63_1
66ht
ml/V
olum
e%20
136(
2),
%20
pp.%
2016
3-16
6.ht
ml
2002
-08
-15
2002
-08
-23
+8H
över
n Sö
derk
ö-pi
ngså
n un
d G
er-
sken
Mot
ala
Strö
m
Öst
ergö
tland
,S
chw
eden
Stef
an L
undb
erg
&Ja
n-E
rik S
vens
son
i sto
ra m
ängd
er4
http
://w
ww
.nrm
.se/
utst
alln
inga
rcos
mon
ova/
jour
hava
ndef
orsk
are/
jou
rhav
ande
biol
og/ry
ggra
dslo
sadj
ur/s
otva
ttens
man
eter
.125
8.ht
ml
2002
-08
-01
2002
-07
-24
+8A
jeR
öder
mar
k/ M
es-
sel/D
iebu
rg,
Deu
tsch
land
Tapp
enbe
ckke
ine
Ang
abe
1ht
tp://
ww
w.lt
sv-s
a.de
/Sei
ten/
Um
wel
t/qua
llen2
.htm
1948
-08
-26
1948
-08
-19
+7Fe
uerlö
scht
eich
bei
Nel
a Pa
rkC
leve
land
Hei
ghts
Ohi
o U
SA
Dex
ter,
Thom
asSu
rrarre
r & D
avis
4 Ex
empl
are
2ht
tps:
//kb.
osu.
edu/
dspa
ce/b
itstre
am/1
811/
3726
/1/V
49N
06_2
35.p
df
204

Fund
-da
tum
Voll-
mon
dD
iffe-
renz
(d)
Fund
ort
Geo
grap
hisc
heLa
geAu
tor/B
eoba
chte
rM
enge
nang
abe
Abun
-da
nzIn
tern
etqu
elle
(letz
ter Z
ugrif
f am
30.
4.20
09)
1970
-01
-29
1970
-01
-22
+7La
ke M
arae
tai
N N
ew Z
eala
ndG
.R. F
ish
2 sp
ecim
ens
1ht
tp://
ww
w.rs
nz.o
rg/p
ublis
h/nz
jmfr/
1971
/5.p
hp
2005
-07
-29
2005
-07
-21
+7P
rivat
e Po
nd S
ani-
tariu
m R
d., M
erlin
Jose
phin
e C
oun-
ty O
rego
n U
SABe
nt C
row
eth
ousa
nds
5ht
tp://
ww
w.c
lr.pd
x.ed
u/ne
ws/
hotto
pics
.php
2004
-07
-09
2004
-07
-02
+7G
rüne
r See
Rat
in-
gen
Deu
tsch
land
BSW
R D
ietm
arFa
hls
Dut
zend
e3
http
://w
ww
.bsw
r.de/
Faun
a/su
essw
asse
rqua
lle.h
tm
2006
-08
-16
2006
-08
-09
+7un
beka
nnt
Deu
tsch
land
Uw
e O
hse
kein
e A
ngab
e1
http
://na
turfo
togr
afen
-foru
m.d
e/o1
3524
0/S
%C
3%B
C%
C3%
9Fw
asse
rqua
lle+g
ewis
cht
2007
-10
-02
2007
-09
-26
+6Lö
bejü
nSa
chse
n-An
halt
Deu
tsch
land
Sim
onis
kein
e A
ngab
e1
http
://w
ww
.akt
iv-u
rlaub
-mac
hen.
de/v
ideo
s/hy
droz
oene
n/ta
uche
n-lo
ebej
uen-
2709
2008
1949
-08
-13
1949
-08
-08
+5R
ound
Lak
eO
hio
US
AD
exte
r, Su
rrarre
r &D
avis
kein
e A
ngab
e1
http
s://k
b.os
u.ed
u/ds
pace
/bits
tream
/181
1/37
26/1
/V49
N06
_235
1997
-09
-21
1997
-09
-16
+5Bo
b R
ice
Pond
Idah
o U
SA
Jess
ica
Lopo
ur5-
6 In
divu
duen
2ht
tp://
ww
w.g
arf.o
rg/fr
eshj
elly
.htm
l
2002
-08
-28
2002
-08
-23
+5H
aage
ner B
agge
r-se
eH
aag/
Am
per
Deu
tsch
land
Tapp
enbe
ckke
ine
Ang
abe
1ht
tp://
ww
w.lt
sv-s
a.de
/Sei
ten/
Um
wel
t/qua
llen2
.htm
2003
-08
-17
2003
-08
-12
+5P
rivat
see
bei H
of-
heim
Sü
dhes
sen
Deu
tsch
land
Mel
(Ben
utze
r-na
me
im F
orum
)se
hr v
iele
Indi
vidu
en4
http
://sc
uba-
boar
d.de
/wbb
2/th
read
1217
.htm
l
1970
-01
-26
1970
-01
-22
+4La
ke M
arae
tai
N N
ew Z
eala
ndG
.R. F
ish
5 sp
ecim
ens
2ht
tp://
ww
w.rs
nz.o
rg/p
ublis
h/nz
jmfr/
1971
/5.p
hp
1999
-09
-29
1999
-09
-25
+4ni
cht a
ngeg
eben
Japa
nG
en-y
u-S
asak
itw
o fe
mal
es...
and
two
wild
mal
es1
http
://w
ww
.mic
rosc
opy-
uk.o
rg.u
k/m
ag/in
dexm
ag.h
tml?
http
://w
ww
.mic
rosc
opy-
uk.o
rg.u
k/m
ag/a
rtnov
99/fw
jelly
.htm
l20
05-
07-2
420
05-
07-2
1+4
Maa
rsse
veen
sepl
asse
nU
trech
tN
iede
rland
eV
ince
nt24
mas
saal
5ht
tp://
ww
w.d
uike
rslo
g.nl
/pub
lic/in
dex.
php?
uid=
Vin
cent
24
1976
-09
-11
1976
-09
-08
+3P
ine
Lake
bei
One
ida
Wis
cons
in U
SA
Dod
son
& C
oope
rse
vera
l hun
dred
5ht
tp://
ww
w.a
slo.
org/
lo/to
c/vo
l_28
/issu
e_2/
0345
1999
-10
-27
1999
-10
-24
+3N
agoy
a C
ityJa
pan
Gen
-yu-
Sas
aki
kein
e A
ngab
e1
2002
-07
-27
2002
-07
-24
+3O
bern
dorfe
r Wei
-he
rM
öhre
ndor
f bei
Erla
ngen
Deu
tsch
land
Tapp
enbe
ck, L
utz
kein
e A
ngab
e1
http
://w
ww
.ltsv
-sa.
de/S
eite
n/U
mw
elt/q
ualle
n2.h
tm
2002
-08
-26
2002
-08
-23
+3C
anyo
nSü
plin
gen
Deu
tsch
land
Tapp
enbe
ck, L
utz
kein
e A
ngab
e1
http
://w
ww
.ltsv
-sa.
de/S
eite
n/U
mw
elt/q
ualle
n2.h
tm
1947
-07
-05
1947
-05
-05
+2La
ke M
ilton
Ohi
o U
SA
Dex
ter,
Surra
rrer &
Dav
ism
any
spec
imen
s3
http
s://k
b.os
u.ed
u/ds
pace
/bits
tream
/181
1/37
26/1
/V49
N06
_235
2002
-08
-25
2002
-08
-23
+2Be
licke
r See
Gen
thin
Deu
tsch
land
Tapp
enbe
ckke
ine
Ang
abe
1ht
tp://
ww
w.lt
sv-s
a.de
/Sei
ten/
Um
wel
t/qua
llen2
.htm
2005
-07
-19
2005
-07
-21
+2M
aars
seve
ense
plas
sen
Utre
cht
Nie
derla
nde
Vin
cent
24al
gem
een
3ht
tp://
ww
w.d
uike
rslo
g.nl
/pub
lic/in
dex.
php?
uid=
Vin
cent
24
1992
-08
-14
1992
-08
-13
+1Sh
effie
ld C
anal
Basi
nSh
effie
ldG
roßb
ritan
nien
How
es &
Ste
phen
Bla
kebo
roug
h5
Exem
plar
e2
http
://w
ww
.ynu
.org
.uk/
jelly
fish.
htm
2002
-07
-25
2002
-07
-24
+1K
iesg
rube
Kaat
sche
nD
euts
chla
ndTa
ppen
beck
kein
e A
ngab
e1
http
://w
ww
.ltsv
-sa.
de/S
eite
n/U
mw
elt/q
ualle
n2.h
tm
205

Fund
-da
tum
Voll-
mon
dD
iffe-
renz
(d)
Fund
ort
Geo
grap
hisc
heLa
geAu
tor/B
eoba
chte
rM
enge
nang
abe
Abun
-da
nzIn
tern
etqu
elle
(letz
ter Z
ugrif
f am
30.
4.20
09)
2002
-08
-24
2002
-08
-23
+1St
avsj
ön, K
ilaån
Söde
rman
land
, S
chw
eden
Lund
berg
&S
vens
son
män
gder
av
med
usor
3ht
tp://
ww
w.n
rm.s
e/ut
stal
lnin
garc
osm
onov
a/jo
urha
vand
efor
skar
e/jo
urh
avan
debi
olog
/rygg
rads
losa
djur
/sot
vatte
nsm
anet
er.1
258.
htm
l20
02-
08-2
420
02-
08-2
3+1
Stei
nbru
ch I,
II u
ndIII
Löbe
jün
bei H
alle
Deu
tsch
land
Tapp
enbe
ck, L
utz
kein
e A
ngab
e1
http
://w
ww
.ltsv
-sa.
de/S
eite
n/U
mw
elt/q
ualle
n2.h
tm
2006
-02
-14
2006
-02
-13
+1ni
cht a
ngeg
eben
Indi
enR
ajes
hR
adha
kris
hnan
kein
e A
ngab
e1
http
://w
ww
.indi
anat
urew
atch
.net
/vie
w_c
at.p
hp?t
ag=%
20?
%20
Cra
sped
acus
ta%
20so
wer
bii
2002
-08
-23
2002
-08
-22
+1H
över
n un
d G
ers-
ken
Öst
ergö
tland
,S
chw
eden
Lund
berg
&S
vens
son
kein
e A
ngab
e1
Faun
a &
flor
a 98
: 1, 2
003
1947
-08
-31
1947
-08
-31
0C
ryst
aL L
ake
bei
Rav
enna
Ohi
o U
SA
Dex
ter,
Surra
rrer &
Dav
isse
vera
l spe
cim
ens
2ht
tps:
//kb.
osu.
edu/
dspa
ce/b
itstre
am/1
811/
3726
/1/V
49N
06_2
35.p
df
2007
-09
-26
2007
-09
-26
0H
ambu
rg L
ake
Ham
burg
Tow
n-sh
ip, M
ichi
gan
US
A
Behn
an/C
iam
paco
mm
only
, abu
ndan
t in
lake
sfo
r som
e tim
e, th
ree
of th
eje
llyfis
h
3w
ww
.ham
burg
.mi.u
s/ne
ws%
20ite
ms/
0926
2007
-Cat
ch
2002
-08
-22
2002
-08
-23
-1ni
cht a
ngeg
eben
nich
t ang
egeb
enG
roßb
ritan
nien
How
es &
Har
dyke
ine
Ang
abe
1ht
tp://
ww
w.y
nu.o
rg.u
k/je
llyfis
h.ht
m
2008
-08
-15
2008
-08
-16
-1Ö
reg-
Dun
ában
(Don
au)
Bécs
(Wie
n)U
ngar
nAn
onym
uske
ine
Ang
abe
1w
ww
.met
ropo
l.hu/
nagy
vila
g/ci
kk/2
8501
9
2006
-04
-12
2006
-04
-13
-1La
go T
anga
nyik
aTa
ngan
yika
Olg
aen
gra
ndes
can
tidad
es4
http
://w
ww
.acu
aris
tas.
cl/v
iew
topi
c.ph
p?f=
22&
t=60
21
1949
-09
-05
1949
-09
-07
-2La
ke W
alle
npau
-pa
ck b
ei S
cran
ton
Penn
sylv
ania
US
AD
exte
r, Su
rrarre
r &D
avis
abun
danc
e3
http
s://k
b.os
u.ed
u/ds
pace
/bits
tream
/181
1/37
26/1
/V49
N06
_235
1973
-03
-17
1973
-03
-19
-2La
ke M
arae
tai
N N
ew Z
eala
ndG
.R. F
ish
25 s
peci
men
s3
http
://w
ww
.rsnz
.org
/pub
lish/
nzjm
fr/19
75/5
2.ph
p
2001
-08
-02
2001
-08
-04
-2N
iede
rneu
endo
rfer
See
Span
daue
r Zita
d-el
le D
euts
chla
ndTa
ppen
beck
kein
e A
ngab
e1
http
://w
ww
.ltsv
-sa.
de/S
eite
n/U
mw
elt/q
ualle
n2.h
tm
2002
-08
-21
2002
-08
-23
-2D
onca
ster
’s H
at-
field
Wat
er P
ark
York
shire
Gro
ßbrit
anni
enH
owes
&S
cow
crof
tke
ine
Ang
abe
1ht
tp://
ww
w.y
nu.o
rg.u
k/je
llyfis
h.ht
m
2007
-07
-27
2007
-07
-30
-3Ü
ttels
heim
er S
eeD
uisb
urg
Deu
tsch
land
Uw
e O
hse
Ein
zelfu
nd1
http
://na
turfo
togr
afen
-foru
m.d
e/o8
7043
/S%
C3%
BC
%C
3%9F
was
serq
ualle
+1+2
007
1949
-09
-03
1949
-09
-07
-4R
ound
Lak
eO
hio
US
AD
exte
r, Su
rrarre
r &D
avis
only
a fe
w s
peci
men
s2
http
s://k
b.os
u.ed
u/ds
pace
/bits
tream
/181
1/37
26/1
/V49
N06
_235
2000
-05
-14
2000
-05
-18
-4G
en-y
u S
asak
i1
Stro
bila
tion
im E
xper
imen
t 1
ww
w.P
lana
rian.
net/g
f/sow
erby
i/bud
ding
2002
-09
-17
2002
-09
-21
-4Pe
truse
kke
ine
Ang
abe
1ht
tp://
ww
w.ia
bc.c
z/sc
ripts
/det
ail.p
hp?i
d=41
20
2007
-07
-26
2007
-07
-30
-4W
iene
r Bad
ege-
wäs
ser
Wie
n, Ö
ster
reic
hA
g./R
ed.
die
erst
en Q
ualle
n1
http
://di
epre
sse.
com
/hom
e/pa
nora
ma/
wel
t/319
437/
inde
x.do
2007
-07
-26
2007
-07
-30
-4Fi
scht
eich
bei
Tös
-sr
iede
ren
Egl
isau
, Sch
wei
zS
chne
iter/K
och
Ansa
mm
lung
von
min
dest
ens
1000
Stü
ck5
ww
w.n
bt.c
h/st
orys
/sto
rys_
arch
iv
2003
-09
-06
2003
-09
-10
-4St
einb
ruch
see
bei
Hal
leH
alle
, Deu
tsch
-la
ndH
ambu
rger
Aben
dbla
ttke
ine
Ang
abe
1ht
tp://
ww
w.a
bend
blat
t.de/
chl/3
9.ht
ml
2003
-09
-05
2003
-09
-10
-5H
auba
chse
e D
uis-
burg
Deu
tsch
land
Bio
logi
sche
Sta
-tio
n W
estli
ches
Ruh
rgeb
iet e
.V
2 Ex
empl
are
1ht
tp://
ww
w.b
swr.d
e/Fa
una/
sues
swas
serq
ualle
.htm
206

Fund
-da
tum
Voll-
mon
dD
iffe-
renz
(d)
Fund
ort
Geo
grap
hisc
heLa
geAu
tor/B
eoba
chte
rM
enge
nang
abe
Abun
-da
nzIn
tern
etqu
elle
(letz
ter Z
ugrif
f am
30.
4.20
09)
2003
-09
-05
2003
-09
-10
-5St
einb
ruch
gew
äs-
ser a
m P
eter
sber
gbe
i Hal
le (S
aale
)
Sach
sen-
Anha
ltD
euts
chla
ndG
roße
&S
chne
ider
kein
e A
ngab
e1
http
://w
ww
.uni
-pro
toko
lle.d
e/na
chric
hten
/id/2
1972
/
1947
-09
-24
1947
-09
-30
-6C
ryst
al L
ake
bei
Rav
enna
Ohi
o U
SA
Dex
ter,
Surra
rrer &
Dav
isst
ill ab
unda
nt3
http
s://k
b.os
u.ed
u/ds
pace
/bits
tream
/181
1/37
26/1
/V49
N06
_235
1975
-02
-20
1975
-02
-26
-6La
ke R
otot
iN
New
Zea
land
G.R
. Fis
hla
rge
num
bers
3ht
tp://
ww
w.rs
nz.o
rg/p
ublis
h/nz
jmfr/
1975
/52.
php
2002
-08
-17
2002
-08
-23
-6D
onca
ster
’s H
at-
field
Wat
er P
ark
York
shire
Gro
ß-br
itann
ien
How
es &
Farn
ingh
amth
ousa
nds(
!)5
http
://w
ww
.ynu
.org
.uk/
jelly
fish.
htm
2002
-09
-15
2002
-09
-21
-6D
onca
ster
’s H
at-
field
Wat
er P
ark
York
shire
Gro
ß-br
itann
ien
How
es &
Alle
n40
Exe
mpl
are
4ht
tp://
ww
w.y
nu.o
rg.u
k/je
llyfis
h.ht
m
2008
-09
-09
2008
-09
-15
-6La
c d’
Ann
ecy
Fran
krei
chR
oche
fort/
Cor
olla
en c
as d
e fo
rte a
bond
ance
5ht
tp://
doris
.ffes
sm.fr
/fich
e2.a
sp?f
iche
_num
ero=
442
2002
-08
-16
2002
-08
-23
-7E
inla
ufw
eihe
rAm
per W
erke
Irsch
ing
Deu
tsch
-la
nd
Tapp
enbe
ckke
ine
Ang
abe
1ht
tp://
ww
w.lt
sv-s
a.de
/Sei
ten/
Um
wel
t/qua
llen2
.htm
2008
-08
-09
2008
-08
-16
-7V
itkov
ický
chTs
chec
hien
Kapl
erke
ine
Ang
abe
1
1974
-02
-27
1974
-03
-08
-8La
ke M
arae
tai
N N
ew Z
eala
ndG
.R. F
ish
25 s
peci
men
s3
http
://w
ww
.rsnz
.org
/pub
lish/
nzjm
fr/19
75/5
2.ph
p
2003
-08
-04
2003
-08
-12
-8D
ix É
tang
s à
Cuy
Fran
krei
chD
obri
& C
alda
ra in
:D
anie
l Nar
din
les
prem
iers
spé
cim
ens
ont
été
déco
uver
ts1
http
://w
ww
.che
z.co
m/m
onam
iph/
cras
peda
.htm
2002
-06
-15
2002
-06
-24
-9Fi
scht
eich
Nie
derp
örin
g/D
egge
ndor
fD
euts
chla
nd
Tapp
enbe
ckke
ine
Ang
abe
1ht
tp://
ww
w.lt
sv-s
a.de
/Sei
ten/
Um
wel
t/qua
llen2
.htm
1961
-01
-22
1961
-01
-31
-9M
inin
g po
ol n
ear
Ampa
ng S
elan
gar
Mal
aya
Hoh
Lia
n Yo
ng &
Won
g Ph
ui W
eng
seve
ral h
undr
ed w
ithin
a fe
wm
inut
es, v
ery
larg
e nu
mbe
rs5
Bull.
Nat
iona
l Mus
. 30
1961
38-
39
2003
-07
-04
2003
-07
-13
-9N
euen
burg
erse
erBe
i Est
avay
erS
chw
eiz
Stei
nman
n1-
2 St
ück
pro
Kubi
kmet
er2
http
://w
ww
.pst
einm
ann.
net/n
eo_c
rasp
edac
usta
.htm
l
1949
-08
-30
1949
-09
-07
-10
Rou
nd L
ake
Ohi
o U
SA
Dex
ter,
Surra
rrer &
Dav
ism
any
spec
imen
s3
http
s://k
b.os
u.ed
u/ds
pace
/bits
tream
/181
1/37
26/1
/V49
N06
_235
2005
-09
-08
2005
-09
-18
-10
Töpp
erse
e/Te
gge
Dui
sbur
gD
euts
chla
ndBS
WR
Uw
e O
hse
meh
r als
50
Indi
vidu
en4
http
://w
ww
.bsw
r.de/
Faun
a/su
essw
asse
rqua
lle.h
tm
2003
-07
-03
2003
-07
-13
-10
Neu
enbu
rger
see
Sch
wei
zPa
trick
Ste
inm
ann
kein
e A
ngab
e1
http
://w
ww
.pst
einm
ann.
net/n
eo_c
rasp
edac
usta
.htm
l
2008
-08
-06
2008
-08
-16
-10
Etan
g en
Als
ace
Fran
krei
chE
lsaß
Sand
ra C
lub
depl
ongé
e de
Sar
re-
bour
g
kein
e A
ngab
e1
http
://cp
s57.
free.
fr
2008
-07
-08
2008
-07
-18
-10
Lac
le L
éman
Fran
krei
chN
yon
Hae
ner,
Rod
olph
eils
ne
sont
pas
nom
breu
x2
http
://w
ww
.lem
antin
es.c
h/ar
ticle
s_di
vers
/med
use_
lem
aniq
ue/m
edu
se_l
eman
ique
.htm
l20
08-
07-0
820
08-
07-1
8-1
0Le
Lém
an,
Fran
krei
chH
aene
rIls
ne
sont
pas
nom
breu
x à
les
avoi
r vue
s. P
ourta
nt, e
lles
sont
bel
et b
ien
là.
2ht
tp://
ww
w.le
man
tines
.ch/
artic
les_
dive
rs/m
edus
e_le
man
ique
/med
use
_lem
aniq
ue.h
tml
207
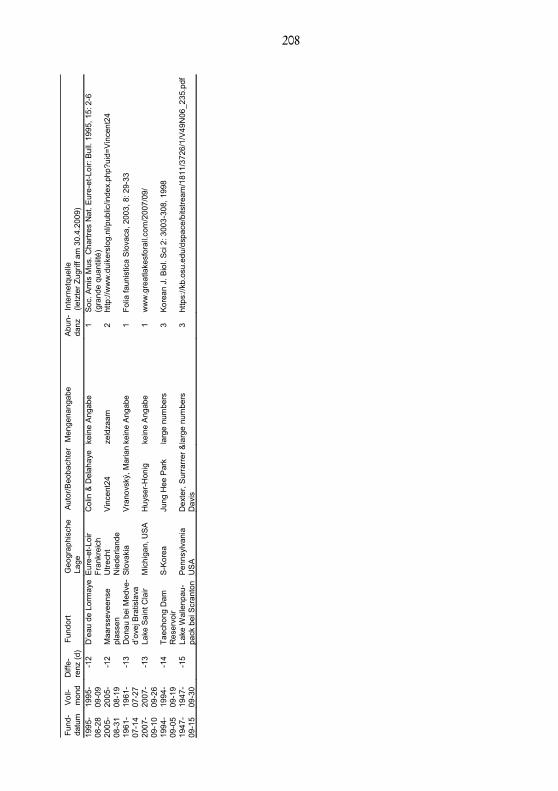
Fund
-da
tum
Voll-
mon
dD
iffe-
renz
(d)
Fund
ort
Geo
grap
hisc
heLa
geAu
tor/B
eoba
chte
rM
enge
nang
abe
Abun
-da
nzIn
tern
etqu
elle
(letz
ter Z
ugrif
f am
30.
4.20
09)
1995
-08
-28
1995
-09
-09
-12
D’e
au d
e Lo
rmay
eEu
re-e
t-Loi
rFr
ankr
eich
Col
in &
Del
ahay
eke
ine
Ang
abe
1So
c. A
mis
Mus
. Cha
rtres
Nat
. Eur
e-et
-Loi
r: Bu
ll. 1
995,
15:
2-6
(gra
nde
quan
tiité
)20
05-
08-3
120
05-
08-1
9-1
2M
aars
seve
ense
plas
sen
Utre
cht
Nie
derla
nde
Vin
cent
24ze
ldza
am2
http
://w
ww
.dui
kers
log.
nl/p
ublic
/inde
x.ph
p?ui
d=V
ince
nt24
1961
-07
-14
1961
-07
-27
-13
Don
au b
ei M
edve
-d’
ovej
Bra
tisla
vaS
lova
kia
Vra
novs
ký, M
aria
nke
ine
Ang
abe
1Fo
lia fa
unis
tica
Slo
vaca
, 200
3, 8
: 29-
33
2007
-09
-10
2007
-09
-26
-13
Lake
Sai
nt C
lair
Mic
higa
n, U
SA
Huy
ser-
Hon
igke
ine
Ang
abe
1w
ww
.gre
atla
kesf
oral
l.com
/200
7/09
/
1994
-09
-05
1994
-09
-19
-14
Taec
hong
Dam
Res
ervo
irS
-Kor
eaJu
ng H
ee P
ark
larg
e nu
mbe
rs3
Kore
an J
. Bio
l. Sc
i 2: 3
003-
308,
199
8
1947
-09
-15
1947
-09
-30
-15
Lake
Wal
lenp
au-
pack
bei
Scr
anto
nPe
nnsy
lvan
iaU
SA
Dex
ter,
Surra
rrer &
Dav
isla
rge
num
bers
3ht
tps:
//kb.
osu.
edu/
dspa
ce/b
itstre
am/1
811/
3726
/1/V
49N
06_2
35.p
df
208

209
4 ErgebnisseDie Ergebnisse berücksichtigen die Jahre 2004 bis 2008. Das Medusenvorkom-men wurde, wie oben erwähnt, erstmals für diesen See (vgl. Neobiota-WatcherAktion 2005) gemeldet. Im Rahmen der Fragestellung wurden erstmals für denSteinbruchsee Winkel die Schichtungsverhältnisse des Wasserkörpers im Jahres-verlauf untersucht. Diese Ergebnisse werden allerdings nur insoweit wiederge-geben, als die für das Medusenvorkommen relevant sind.
4.1 Schichtung des WasserkörpersFast während der ganzen Saison ist ab April bis September eine meist ausge-prägte Trübschicht (Metalimnion) zwischen etwa 4 und 12 m zu beobachten,die mit der physikalischen Sprungschicht, die durch die Wassertemperatur defi-niert ist, korrelliert (Abb. 2).
Abb. 2: Temperaturprofil imSee Winkel zur Bestimmungder Sprungschicht
4.2 Saisonale Phasen im See Winkel (Tab. 3)Ungefähr zwischen Frühlings- und Herbstanfang weist die zwischen 4 und 12m Tiefe befindliche Sprungschicht in der Regel eine Sichtweite von 0,5 bis ma-ximal 3 m auf. Während dieser Zeit sind zwei Trübphasen zu beobachten, dievon Phasen geringerer Trübung (Klarphasen) mit einer Sichtweite von 3 m undmehr unterschieden werden können. Das zwischen der Wasseroberfläche und

210
einer Tiefe von 4 m liegende Epilimnion weist in diesen Phasen eine Sichtweitevon 4-5 m und mehr auf. Während der beiden Trübphasen sind auch das Epi-limnion bis zur Oberfläche und das Hypolimnion bis auf 20 m Tiefe stärkereingetrübt.
Tab. 3: Steinbruchsee Winkel. Saisonale Phasen der Sichtweite (von Jahr zu Jahrleicht schwankend)
Phase Dauer (Termin) Dauer (Wo-chen)
Kalenderwoche (KW) Bezug zum Sonnenstand
1. Klarphase(Winter)
von Jahresbeginn bisAnfang April
14 1.-14. bis 1-2 Wochen nach Früh-lingsanfang
1. Trübphase(Frühjahr)
von Anfang April bis An-fang der 2. Juniwoche
9 14.-23. von 1-2 Wochen nach Früh-lingsanfang bis 3 Wochen vorSommeranfang
2. Klarphase(Sommer)
von Anfang der 2. Juni-woche bis Anfang der 4.Juliwoche
6 23.-29. von 3 Wochen vor bis 3 Wo-chen nach Sommeranfang
2. Trübphase(Sommer)
von Anfang der 4. Juliwo-che bis Anfang der 3.Septemberwoche
9 29.-38. von 3 Wochen nach Sommer-anfang bis 1 Woche nachHerbstanfang
3. Klarphase(Herbst)
von Anfang der 3. Sep-temberwoche bis Jahres-ende, dann in die 1. Klar-phase des nächsten Jah-res übergehend
14 39.-52. 1 Woche nach Herbstanfangbis Jahresende (Winteranfang)
Ab drei Wochen vor Herbstanfang ist bis Dezember in der Tiefe von 17-20 mzugleich der für Schwefelwasserstoff charakteristische Geruch festzustellen, derselbst durch die Tauchmaske den durch Abbauprozesse bedingten Sauerstoff-mangel (Anoxie) im Hypolimnion anzeigt. Sauerstoffmessungen am 19.04.2009ergaben die folgenden Werte: Oberflächenwasser: 10,5 mg/l; 5 m Tiefe 10mg/l; 15 m 7 mg/l; und 29 m 7 mg/l, die die Durchmischung des Sees bestäti-gen (Schmidt 2009). Der Steinbruchsee Winkel zählt somit zu den dimiktischenSeen, die wie die meisten Seen in unseren gemäßigten Breiten zweimal im Jahrim Frühjahr und im Herbst voll zirkulieren.
4.3 Lebensraum und Abundanz von Craspedacusta sowerbiiDie Süßwassermedusen-Saison fällt in die Zeit der zweiten Trübwasserphaseund dauert bis Oktober an. In den Jahren 2004 bis 2008 erschien Craspedacusta so-werbii im Steinbruchsee regelmäßig zu Beginn der 2. Trübphase, die in der 29.Kalenderwoche einsetzte. Das Auftreten der Population hielt über den Herbst-anfang hinaus noch eine Mondphase länger als die Dauer der 2. Trübphase biszur 41. Kalenderwoche an und brach dann zusammen. Das Gesamtauftretender Süßwassermeduse umfasste somit einen Zeitraum von 12 Wochen mit vierMaxima ihrer Abundanz entsprechend den vier Vollmondphasen in dieser Zeit.In den Jahren ihres Vorkommens wies die Individuendichte jeweils wenige Ta-ge um den Vollmond herum meist über 50 Individuen/m³ auf. In den dazwi-schen liegenden Zeitabschnitten war die Individuendichte deutlich geringer.

211
2008 stellte bezüglich der Medusen-Abundanz eine Ausnahme dar (Abb. 3, 4,5). Die Medusen bewegten sich nicht unter die 6-m-Grenze.
Abb. 3: Zum zweiten Vollmondtermin nach Sommersonnenwende setzt ab Mitte Juliim See Winkel das erste Massenauftreten von Craspedacusta sowerbii im Jahr ein.Die Tiere kommen bis zu einer Tiefe von 5-6 m und bei einer Temperatur >17 °C vor.Pfeile = Vollmond, TG = Tauchgang
Abb. 4: Das Massenauftreten von Craspedacusta sowerbii im See Winkel wiederholtsich vier Mal im Jahr mit Beginn des zweiten Vollmonds nach Sommersonnenwendeund liegt stets in einer Woche mit dem jeweiligen Vollmondtermin (Pfeile)

212
Abb. 5: 2008 war die Abundanz von Craspedacusta sowerbii im See Winkel um denFaktor 10 geringenr als in den vorangegangenen Jahren ihres Massenauftretens. DasAuftreten wurde ebenfalls um den Vollmondtermin (Pfeile) herum beobachtet,lediglich das letzte Auftreten im Jahr war um 11 Tage vor dem Eintritt des Vollmondsvorgezogen. Ein Individuum wurde vor der Sommersonnenwende registriert
4.4 Circalunares Massenauftreten von Craspedacusta sowerbiiWie es die Diagramme Abbildung 3 bis 5 der Jahre 2005, 2006 und 2008 der er-hobenen Daten belegen, findet in der zweiten Jahreshälfte in etwa zum zweitenVollmondtermin nach Sommersonnenwende das erste Massenauftreten vonMedusen statt, gefolgt von weiteren drei Massenauftreten zu den jeweils darauffolgenden Vollmondterminen. Die Abundanz am ersten Termin ist in der Re-gel noch deutlich niedriger als die des Folgeauftretens. Im Jahr 2007 blieb Cras-pedacusta sowerbii völlig aus, ebenso wie die in den Vorjahren zu beobachtende 2.Klarphase, d.h. dass das Phytoplankton in diesem Jahr nicht abgeerntet wurde.Der jeweilige Vollmondtermin liegt in der Woche auf die in den Abbildungen3, 4, 5, 6 ein Pfeil zeigt. In der 33. und 34. und nach der 40. Kalenderwochewurden 2005 keine Daten erhoben. Als Hauptarten, die in der Population auf18 m Tiefe am häufigsten vertreten waren, konnten die folgenden Taxa be-stimmt werden: Daphnia longispina (06.08.2008), Cyclops strenuus (13.08.2008), Ecto-cyclops phaleratus (02.05.2009), Daphnia longispina, Daphnia pulex und Cryptocyclops bicolor(17.06.2009). Die in Abbildung 6 beobachtete Beziehung zwischen Medusen-und Crustacea-Auftreten kann dennoch zunächst nur als Hinweis gewertetwerden, der durch weitere Untersuchungen überprüft werden muss.

213
Abb. 6: Einem vor der Sommersonnenwende liegenden Populationsmaximum anCrustacea folgen zwei weitere nach der Sommersonnenwende. Die sich zu den Voll-mondterminen (Pfeile) einstellende Populationsentwicklung von Craspedacusta so-werbii legt eine Korrelation zwischen Medusen und Crustacea Taxa durch Prädationgemäß den Lotka-Volterra-Regeln nahe (vgl. auch Abschnitt 5.4). Beachte die unter-schiedlichen Dimensionen des Bezugsvolumens
4.5. Ergebnisse der Metauntersuchung zur circalunaren Rhythmikvon Craspedacusta sowerbiiDie zur Verfizierung der Beobachtungen angestellte Metauntersuchung vonMeldungen aus der im Internet nachgewiesenen Literatur zeigt, dass sich Fundevon Craspedacusta sowerbii auffällig um ihre jeweiligen Vollmonddaten herum häu-fen. Von einer Woche vor bis zu einer Woche nach Vollmond wird ein meistals gehäuft bezeichnetes Auftreten angegeben (Abb. 7). In dieser Zeitspanne be-findet sich der Mond in seinem letzten zunehmenden sowie in seinem erstenabnehmenden Viertel, den hellsten Vierteln des Mondzyklus. Somit deckt sichdie Häufigkeit des Auftretens von Craspedacusta sowerbii im See Winkel mit denBefunden der Literatur. Diese werden hier erstmalig in einer Metauntersu-chung unter biorhythmischem Aspekt ausgewertet.
Offensichtlich sind Vollmond und die beiden hellen Mondviertel ein exter-ner Zeitgeber für die Knospung (Strobilation) der Polypen. Angesichts des se-xuellen Zwecks der Medusengeneration mit der Entlassung der Geschlechtspro-dukte ins Wasser wird somit ein relativ gleichartiger Reifegrad vieler Tieresicher gestellt. Aufgrund der circalunaren und räumlichen Synchronisation desabendlichen Aufsteigens der Tiere unter die Oberfläche erhöht sich deren Ab-undanz und damit die Chance, dass sich im dreidimensionalen Wasserkörper

214
die Geschlechtsprodukte gehäuft treffen und der Erfolg einer Befruchtungwahrscheinlicher wird.
Abb. 7: Häufung der Fundmeldungen von Craspedacusta sowerbii in der Literatur umihre nächsten Vollmondtermine herum (Tab. 2)
Geht man von den Ergebnissen von Gen-Yu Sasaki (1999) aus, der die Ent-wicklung von der Geschlechtszellenabgabe, der Befruchtung über die Phase alsPlanula-Larve bis zum Festsetzen des Polyps untersucht hat und bezieht mandie genannte Gesamtentwicklungszeit ein, so benötigt der Vorgang acht Tage:"(Eggs) were spawned in 20th Oct." und dann "near the midnight of the 27thOct. 1999, I found two embryos" "embryos that were in the two-cell stage ofcleavage!" In der anschließenden Entwicklung über die Planula-Larve bis zumFestsetzen des Polyps vergeht ein weiterer Tag. Drei Tage nach Vollmond, deram 24.10.1999 lag, beobachtete Gen-Yu Sasaki die Embryos, die dann allerdingsnoch Wachstumszeit bis zur makroskopisch beim Tauchen zu beobachtendenMeduse benötigen. Der Zuwachs ihres Körpers liegt dann während des zweitenund dritten dunklen Mondviertels. Die reife sexuelle Funktion der Medusen istaufgrund der Synchronisation ja auch frühestens erst ab dem Eintritt in dasletzte Mondviertel vor Vollmond vonnöten. Gen-Yu Sasaki (1999) stellt als

215
Entwicklungszeit fest: "I had reared the medusae for about one month frommid October 1999". Auch Lundberg et al. (2007: 4) geben eine entsprechendeEntwicklungsdauer an: "Förutsatt att tillgången på zooplankton är tillräckligkommer de unga medusorna att växa och utvecklas till könsmogna individerinom ca. 5 veckor". [Vorausgesetzt, dass die Versorgung mit Zooplanktonausreichend ist, erscheinen die jungen Medusen, um heran zu wachsen und sichinnerhalb von 5 Wochen zu geschlechtsreifen Individuen zu entwickeln]ebenso wie Pérez-Bote et al. (2006: 165) "Craspedacusta sowerbyi medusaereach sexual maturity after 5-6 weeks".
Wird dieser Zeitbedarf der Entwicklung berücksichtigt, lässt sich auch derBeginn des ersten Auftretens der Süßwassermeduse vier Wochen nach Sommer-anfang (21.-22.06.) etwa in der zweiten bis zur dritten Juliwoche nach dem ers-ten Juli-Vollmond erklären.
Der Höchststand der Sonne mindestens als Voraussetzung für die nachfol-gend zu erwartende höchste Temperatur im Jahr, wenn nicht auch das Lichtdes Langtags, in Kombination mit einem Vollmonddatum um Sommeranfangherum, so die hier ausgesprochene Hypothese, induziert die Strobilation vonMedusen durch die Polypen. Dadurch gerät der Start des ersten Massenaufkom-mens der Tiere außerdem in die 2. Trübphase, in der sich die Zooplanktonpo-pulation als Nahrungsgrundlage für die Medusen erneut aufbaut. In den Unter-suchungsjahren 2005 und 2006 lagen die Vollmonddaten im Juni und im Julientsprechend zeitlich günstig um die Sonnenwende herum.
5 DiskussionBezüglich Neozoen ergibt sich stets die Frage nach dem Einfluss der eingewan-derten Art auf autochthone Lebensgemeinschaften. Andererseits ist im Erfolgder Neozoen, sich in dem neuen Lebensraum auszubreiten und fortpflanzen zukönnen auch eine Evolutionsstrategie zu sehen, die für die betreffende Arteinen Selektionsvorteil hat. Daher werden beide Aspekte vor dem Hintergrundder Untersuchung diskutiert.
5.1 Ökologischer Einfluss von Craspedacusta sowerbii auf den Le-bensraum: TemperaturDie "unvorhersagbare" Massenentwicklung erscheint auf der Basis der vorlie-genden Ergebnisse durchaus berechenbarer. In der Regel nehmen Organismeneine ökonomische Ressourcennutzung vor. Unter diesem Aspekt ist die Anpas-sung an eine Mindesttemperatur zweifelsfrei für die Entwicklung der poikilo-thermen Polypen aus den Dauerstadien des Winters und die Medusensaisonbiologisch sinnvoll. Die die Erst-Strobilation auslösende Mindesttemperatur istgleichzeitig für die globale Verbreitung der Art in kältere Regionen limitierend.

216
Nach dem Ansteigen des Sonnenstands bis zur Sommersonnenwende, dem einehöhere Wassertemperatur im Epi- und Metalimnion folgen, leitet dann derSommeranfangs-Vollmond als Zeitgeber die erste Strobilation der Polypen imJahr ein und synchronisiert die Reife der Medusen. Er koordiniert außerdem inKombination mit der positiv fototaktischen Reaktion der Medusen, die sich inVollmondnächten an der Wasseroberfläche konzentrieren, die (potenzielle) se-xuelle Fortpflanzung. Außerdem hat er dirigierende Wirkung in Bezug auf einegünstige Ernährungslage für die Medusen während der zweiten Trübwasserpha-se. Die darauf folgenden vier Massenauftreten von Medusen werden ebenfallsdurch den Vollmond der folgenden Monate gesteuert. Die sinkende Wassertem-peratur führt schließlich zum Einstellen weiterer Strobilation und den Über-gang der Polypen in die Winterstadien.
In diesem Zusammenhang kommen Farkašova & Stloukal (2007: 3) auf-grund ihrer Beobachtungen zu der Hypothese "The strobilation could happenwhen the temperature of the water is lower then mentioned in the literature".Dazu führen sie Fleming & Hazelwood (1967) an "who observed medusaewhen the temperature of the water was only 16 °C", was von Sládek & Šraier(2005: 3) bestätigt wird: "the occurence of medusae in abandoned stone pit Ko-jetice, Czech republic overwhelm with water of temperature from 8 to 16 °C.".Auch Danli Wang et. al. (2006) stellen fest: "The results indicated that the suit-able temperature (...) of C. sowerbii xinyangensis (is) 15 degrees C - 25 degreesC (...). The average oxygen consumption amount and rate of C. sowerbiixinyangensis increased with increasing temperature". Dabei ist nicht nur dieOberflächentemperatur von Bedeutung, sondern auch diejenige in größererTiefe, in der die Polypen zur Entwicklung kommen.
So macht Wieser (1993) eine um 2-4 °C höhere Temperatur in der Wasser-säule zwischen 0-6 m im Weizelsdorfer Baggersee (Kärnten) für das Auftretenvon Craspedacusta sowerbii verantwortlich. "Noch markanter erscheint der Ver-gleich der Temperaturkurve in 7 m Tiefe über Grund im Vergleich zum Jahr1991, wo eine markant höhere Temperatur von Mai bis hin zum Septemberverfolgt werden kann (7 m Tiefe: August 1991 17,8 °C, August 1992 24,3 °C)".Dass die Temperatur in dieser Tiefe eine Rolle spielt, lässt sich aus den Fund-orten der Polypen in einer Tiefe von 2,50-9,00 m im Eriesee ableiten, die Hub-schmann & Kishler (1972) feststellen.
Die beobachtete Temperatur der Oberfläche im Mai von 12 °C, im Juni von14 °C und im Juli von 17-23 °C sowie solche von 9-15 °C im Juni-Juli im Meta-limnion haben im See Winkel das Medusenauftreten in den Jahren 2004 bis2006 nicht verhindert, so dass die von vielen Autoren als Voraussetzung vermu-tete Grenztemperatur von deutlich über 20 °C, die für die Strobilation gefor-dert wird, zu Sommeranfang in unserem See nicht erreicht wurde, da dieser 465

217
m ü.M. liegt. Dennoch scheint es evident zu sein, dass wenn die Temperaturunter einer Grenztemperatur bleibt, die niedriger sein wird, als bislang ange-nommen, bleibt auch die Entwicklung der Medusen aus.
Mit diesem Temperaturfenster, das Anfang bis Mitte Juni während der 2.Klarphase die entsprechende Entwicklungstemperatur für die Medusenproduk-tion anbietet, wird gleichzeitig genügend Zooplankton zur Verfügung gestellt,das die Polypen als Energielieferanten zur Strobilation nutzen können.
Ist eine Mindesttemperatur in der Wassertiefe, in der die Polypen siedeln,nicht gegeben, könnten sie Energie für die Entwicklung der Medusen einspa-ren. Immerhin können sie für die im Winter folgende Temperatur unter 10 °CÜberdauerungskörper bilden, und sich erst in der nächsten Saison vegetativfortpflanzen.
Die Evidenzen sind angesichts der Ergebnisse äußerst schwach, in Mitteleu-ropa die Temperatur als limitierenden Faktor für das Ausbleiben der Medusen-entwicklung verantwortlich zu machen. Da auch die Jahre, in denen kein Me-dusenvorkommen im Steinbruchsee beobachtet wurde (z.B. 2007)Temperaturwerte aufwiesen, bei denen die Entwicklung in den Jahren vorherstattfand, kann vielmehr davon ausgegangen werden, dass andere Faktoren fürdie Medusenentwicklung verantwortlich sind.
5.2 Circalunare SteuerungNeben die notwendige Basistemperatur tritt der Mondzyklus zur circalunarenSteuerung des Massenauftretens der Medusen. Dass der Mond und die Gezeitenbedeutende Zeitgeber sind, ist schon lange bekannt. So fasst Fautin (2002) zu-sammen, dass die massenhafte Abgabe von Gameten zuerst von Marshall & Ste-phenson (1933) beschrieben und von Harrison et al. (1984) intensiver an 32 Ar-ten von Steinkorallen (Scleractinia) des Großen Barriere Riffs dokumentiertwurde. Die nahe der Meeresoberfläche vorkommenden Organismen und jene,die im Gezeitenbereich leben, beeinflusst der Mond unter anderem durch dasEntlassen von Eiern und Spermien ins Wasser. Hinsichtlich der SteinkorallePocillopora damicornis von West Australien berichtet sie, dass sie die meistenihrer Planulalarven während der Neumondtermine im Sommer abgibt, wäh-rend die meisten Planulae im nördlichen Großen Barriere Riff monatlich beiVollmond entlassen werden. Bezüglich der Massenentlassung von Gameten beiCnidaria stellt sie fest, dass sie außer für Anthozoa keine Daten gefunden hat:"I found no data, however, for cnidarians other than anthozoans" Fautin (2002:1738). Auch für Craspedacusta sowerbii, ebenfalls zu den Cnidaria zählend, wurdenkeine Angaben über die Massenabgabe von Geschlechtsprodukten gefunden,die ja dann von den Medusen synchronisiert ausgehen müsste, obwohl dies beieinigen marinen Hydrozoa beschrieben ist.

218
Der Steinbruchsee Winkel liegt gegenüber seiner Umgebung in einer kraterarti-gen Grube in der die Sonne erst spät am Morgen erscheint und am Abend be-reits früh hinter dem Kraterrand verschwindet. Daher finden sich die Medusen,obgleich im ganzen See vorkommend, gehäuft auf der besonnten Nord- undeinem Teil der Westseite. Vorausgesetzt die Polypen besiedeln dort das Litoralim Epilimnionbereich bevorzugt (vgl. allerdings Hubschmann & Kishler 1972),könnte die positive fototaktische Reaktion der Medusen nach der mitter-nächtlichen Strobilation (vgl. Gen-Yu Sasaki 1999) dazu führen, dass die Medu-sen bei Vollmond, bei dem der Mond um Mitternacht stets im Süden steht, inRichtung Süden ins Pelagial wandern, wo bei Tagesanbruch nach der Strobilati-on eher mit Crustaceen als Nahrung zu rechnen ist (Steiner 2002: 36) und diesedadurch für die Medusen besonders gesichert ist.
Als Ursache der beobachteten und durch die Metauntersuchung bestätigtenSynchronisation der Geschlechtsreife der Tiere ist durchaus auch eine circalu-nare Stimulation der Hormonausschüttung denkbar, wie sie Farkašová &Stloukal (2007) annehmen.
5.3 Räumliches Vorkommen und AbundanzZum Vergleich seien die Abundanzangaben (ausschließlich weiblicher Tiere)von Strauß (1996) mit "einer maximalen Populationsdichte von 384 Medusenpro m³" genannt, während Arbačiauskas & Lesutienė (2005) von einem Massen-auftreten von 20-30 Individuen/m² in einem isolierten Altarm-See des FlussesNemunas in Litauen im heißen Sommer 2002 berichten. Sie waren weder imPelagial noch in tieferen Schichten zu finden. Die beobachtete Häufigkeit imSee Winkel kann in den Jahren, in denen Medusen auftreten, bis zu vier Mal jeSaison diese Dichte erreichen bzw. überschreiten, wie es zuletzt im Jahr 2006zu beobachten war (vgl. Abb. 4, 5, 6).
Fish (1975) beobachtet die Medusen etwa 20 m vor dem Ufer und in 2-5 mWassertiefe. Auch Pérez Bote & al. (2006) haben ihre Medusen nur in offenemWasser gefunden: "C. sowerbyi medusae were found only in open water in theProserpina Reservoir. This is a common feature of C. sowerbyi medusae,which occur in surface and deeper waters as reported by Byers (1945) and Ziser& Burke (1984). Kimmel et al. (1980: 165) found C. sowerbyi mid-lake inBroken Bow Reservoir with no additional sightings near shore, while in theOhio River medusae were recorded at the surface and as deep as 3 m both dayand night (Beckett & Turanchik, 1980)....Angradi (1998) found a highly local-ized distribution of medusae in a reservoir, and suggests that the distributionfrom a location determined by the habitat requirement of the polyp". Diese Be-obachtung kann bestätigt werden, wobei die Medusen im See Winkel amAbend (bei Nachttauchgängen) durchaus am Ufer gehäuft auftraten.

219
Boothroyd et al. (2002) beobachten Craspedacusta sowerbii in einer Dichte von 12,7Individuen/100 l im Oberflächengewässer des Lake Kainui in der Waikato-Region in Neuseeland. Dort weisen sie eine unregelmäßige fleckige Verteilungim offenen Wasser von der Litoralzone entfernt auf. Es schien, dass die Medu-sen selektiv das Vorkommen der beweglichen Copepoda Ceriodaphnia aufsuch-ten, was aus der Untersuchung der Nahrungszusammensetzung hervorging.Dabei erlitt die Copepoda-Population eine geschätzte Mortalitätsrate von 3,41% je Tag im Gegensatz zur Mortalitätsrate der übrigen Zooplankton-Taxa, diebei 0,5 % je Tag lag. Das veranlasst die Autoren zur Schlussfolgerung, dass dieseSelektivität in der Nahrungsaufnahme von Craspedacusta sowerbii die Zooplank-ton-Gemeinschaft qualitativ und quantitativ prägt.
5.4 NahrungDie am Abend zur Nahrungsaufnahme in das Epilimnion aufsteigenden sichnegativ fototaktisch verhaltenden Crustacea treffen besonders um den Voll-mond herum auf die hohe Individuenkonzentration von Craspedacusta sowerbii,deren Population sich in Massen lunarsynchronisiert zur Abgabe der Ge-schlechtsprodukte in den oberen Schichten des Epilimnions versammelt. Da-durch erhält die Medusenpopulation optimale Nahrungsbedingungen, die zuLasten der Entwicklungsstadien der Crustacea geht, die um den Vollmond her-um auftreten.
Da nach der 2. Klarphase das Nahrungsangebot - das Phytoplankton - für dieCrustacea knapp geworden ist, müssen die sich wieder aufbauenden Populatio-nen höhere Positionen als die im Hypolimnion des Frühsommers aufsuchen(vgl. Steiner 2002: Abb. 11). Allerdings bleiben die Crustacea dann im oberenBereich des Metalimnions, um den im Epilimnion jagenden Fraßfeiden unterden Fischen, die in hellen Vollmondnächten des Sommers dann auch nochdurch die in Massen vorkommenden Medusen verstärkt werden, zu entgehen.Immerhin können sich die Medusen "vertikal im Wasser fortbewegen und sindabhängig vom Nahrungsangebot oder Tageszeit gerne in tieferen Wasserschich-ten (...) oder steigen (...) gegen Abend (Spadinger & Maier, 1999) oder bei gu-tem Nahrungsangebot in höhere Schichten auf." (VDST & BfN (Hsg.): www.-neobiota.info, letzter Zugriff: 4.6.2008).
Es ist davon auszugehen, dass die Entwicklungsstadien bevorzugter Arten,die aufgrund des Massenauftretens der Medusen und dem von ihnen ausgehen-den Fraßdruck reduziert werden, dann in der Folge zum Aufbau der neuenCrustacea-Population fehlen. Während das Auftreten der beiden Trübphasen(viel Phytoplankton, wenig Zooplankton) mit den auf sie jeweils folgendenKlarphasen (viel Zooplankton wenig Phytoplankton) mit den ersten beidenLotka-Volterra-Regeln zu erklären sind, überlagert der sich zyklisch zu vier

220
aufeinander folgenden Vollmondterminen einstellende Effekt des Fraßdrucksdie Effekte der Populationsentwicklung, der unter den Crustacea immer wiedergleiche Stadien trifft, wenn sich diese ebenfalls lunarzyklisch entwickeln. Craspe-dacusta sowerbii hat durch Fraßselektion, welche besonders Copepoda betrifft, of-fensichtlich nicht nur qualitativ sondern durch ihr lunarzyklisch wiederholtesMassenauftreten auch quantitativ selektiven Einfluss auf diejenigen Copepodit-Stadien, die seit dem jeweils vergangenen Vollmond den Wiederaufbau der Po-pulation vorantreiben könnten. Daher ist es sehr wahrscheinlich, dass beihoher Individuenabschöpfung durch den Fraß der Medusen und Polypen derübliche Aufbau einer sich in dynamischer Balance mit den einheimischen Fress-feinden befindlichen Population aus dem Gleichgewicht gerät, was im Folge-jahr Auswirkungen auf die Medusenpopulation haben dürfte. Diese Hypothesesollte durch Untersuchungen geprüft werden.
In direkter Folge ist zu erwarten, dass die wiederholt dezimierte Populationan Crustacea auch das Nahrungsangebot für die tiefer sitzenden Polypen beein-trächtigt, deren Beitrag für die potenzielle Fortpflanzung der Art allerdingsdurch die Medusenbildung zu dem Zeitpunkt geleistet ist. Da die Neozoa unse-re Seen allerdings mit nur einem, in Deutschland dem weiblichen Geschlechtbesiedelt haben (Tappenbeck 2008), können die Medusen die sexuelle Fort-pflanzung nicht erfolgreich umsetzen. Die Reproduktion bleibt also auf die Po-lypen beschränkt, die unter Mangelbedingungen Ausdauerformen bilden kön-nen.
Die stark dezimierte Crustacea-Population hat sich im Jahr 2007 nicht rege-nerieren können, was am Ausfall der sommerlichen Klarphase und der starkenTrübung durch die Entwicklung planktischer Algen augenfällig wurde. Zudemkonnte im gleichen Jahr ein Ausbleiben von Medusen festgestellt werden. DiesePopulationsdynamik ist offensichtlich auf das Massenauftreten von Medusen inden Vorjahren zurückzuführen.
5.5 Ausbleibendes Medusenauftreten im Jahr 2007Während sich der Polyp von Craspedacusta sowerbii "gegenüber ungünstigen Um-weltbedingungen als sehr widerstandsfähig (zeigt)" und "sehr gut große Tempe-raturschwankungen und Hungerperioden" übersteht, sterben junge Medusen infrühen Entwicklungsstadien ab, ohne die Geschlechtsreife zu erlangen, wennsie kein ausreichendes Nahrungsangebot haben (Fohrmann 1985: 4). Daher be-fürchtet Dumont (1994) einen dramatischen Einfluss auf die Planktongesell-schaft ab einer Medusendichte von 30 Individuen/m³. Es ist somit offensicht-lich, dass die reduzierte Phytoplanktonproduktion und in der Folge auch diereduzierte Zooplanktonproduktion während der 2. Trübphase sowie eventuellauch zu geringe Temperatur im Hypolimnion als Hauptfaktoren für das Aus-

221
bleiben der Medusen im Jahr 2007 verantwortlich sind. So stellt Steinmann(2008) fest: "In den Jahren 2007 und 2008 wurden keine Quallenaufkommen imZürichsee beobachtet". Selbst das ausführlich geführte Internet-Logbuch vonMannak (2009), das frühere Funde meldet, weist für die Jahre 2007-2008 die Be-merkung "geen" (= keine) Funde für Craspedacusta sowerbii aus.
Folgt man dieser Hypothese, hätte im dynamischen Flusssystem des Jangtse-kiang mit seinen Überschwemmungen diese Prädator-Beziehung eventuelldurch Zufuhr von Nahrungsorganismen aus dem Mittel- und Oberlauf ergänztwerden können. Im Fluss ist eine kontinuierlicher verlaufende Populationsdy-namik zu erwarten als in unseren Seen. Im stagnierenden See hingegen ist miteiner stärkeren Schwankung der Beute-Population zu rechnen, da diese zu ihrerRegeneration Zeit braucht und dieser Prozess überdies zum nächsten Voll-mondtermin erneut beeinträchtigt wird. Darüber hinaus sind die Rest-Populati-on bzw. die sich regenerierenden und in Entwicklung befindlichen Jugendstadi-en durch die im Verlauf der vier aufeinander folgenden Mondzyklen beiVollmond auftretenden Massenvorkommen so starkem Fraßdruck ausgesetzt,dass das Ökosystem wohl nur wenige Jahre hintereinander diesem rhythmi-schen Fraßdruck widerstehen und die Neozoen integrieren kann. Aus der Ar-gumentation folgt, dass die Medusen von Craspedacusta sowerbii trotz der häufiggegenteiligen Ansicht starken Einfluss auf die Zooplanktonentwicklung im Seenehmen.
5.6 Folgenbeurteilung des Auftretens von Craspedacusta sowerbiiRicciardi & Cohen (2007) führen als Beispiel für ihre Untersuchung der Wir-kungsgradkriterien für eingeführte Arten an (Tab. 4): Viele Arten haben keinesubstanzielle ökologische Wirkung in ihrem Gebiet verursacht, in das sie einge-wandert sind, wie z.B. die Süßwassermeduse Craspedacusta sowerbii, die globaleVerbreitung bekommen hat. Andere allerdings, die kaum verbreitet sind, ob-wohl sie genügend Möglichkeiten dazu, hätten, haben starken Einfluss auf dieneuen Populationen ausgeübt, wie z.B. die Asiatische Muschel Potamocorbula amu-rensis, die 1987 in der nördlichen San Francisco Bucht verbreitet wurde und we-sentliche Effekte auf das Plankton hat, indem die Algenblüte im Frühjahr aus-blieb und die Chlorophyllkonzentration seitdem um das Fünffache niedrigerist als vorher. In der Folge sind einige Zooplanktonten, darunter drei Copepo-da-Arten und die Schwebgarnele Neomysis mercedis stark zurückgegangen.

222
Tab. 4: Rangkriterien für die Wirkung eingeführter Arten nach Ricciardi & Cohen(2007)
Ranking Qualification Fähigkeit7 Caused severe (>80 %) declines in the populations
of at least two native species in multiple regionsVerursacht starke (>80 %) Rückgänge von mindes-tens zwei einheimischen Arten in mehreren Gebie-ten
6 Caused severe (>80 %) declines in the populationsof at least two native species at one site, or one na-tive species in multiple regions
Verursacht starke (>80 %) Rückgänge von mindes-tens zwei einheimischen Arten an einem Ort oderbei einer einheimischen Art in mehreren Gebieten
5 Caused a severe (>80 %) decline in the populationof a single native species at one site
Verursacht einen starken (>80 %) Rückgang in derPopulation einer einheimischen Art an einem Ort
4 Caused a significant (but <80 %) decline in at leasttwo native species at one site, or of one native spe-cies in multiple regions
Verursacht einen signifikanten (aber <80 %) Rück-gang von mindestens zwei einheimischen Arten aneinem Ort oder bei einer einheimischen Art in meh-reren Gebieten
3 Caused a significant (but <80 %) decline in one na-tive species at one site
Verursacht einen signifikanten (aber <80 %) Rück-gang in der Population einer einheimischen Art aneinem Ort
2 Had a negative effect on the fitness or survival ofmultiple native species in one or more geographi-cally-distinct regions
Hatte einen negativen Effekt auf die Fitness (An-passungsfähigkeit) oder das Überleben mehrerereinheimischer Arten in einem oder mehreren geo-grafisch unterschiedlichen Gebieten
1 Had a negative effect (e.g. through competition) onthe fitness or survival of individuals of one nativespecies at one site
Hatte einen negativen Effekt (z.B. durch Konkur-renz) auf die Fitness (Anpassungsfähigkeit) oderdas Überleben von Individuen einer einheimischenArt an einem Ort
0 No demonstrable negative impact on native species Keine nachweisbare negative Wirkung auf einheimi-sche Arten
Wenn der Zusammenbruch der Zooplanktonpopulation von adulten bzw. ju-venilen Stadien der Arten im Jahre 2007 sowie das geringe Vorkommen vonCraspedacusta sowerbii auf die übermäßige Prädation der Süßwassermeduse inklusi-ve ihres Polypen in den Vorjahren zurückzuführen ist, kann ihr Einfluss nachder Liste der Rangkriterien für die Wirkung eingeführter Arten nach Ricciardi& Cohen (2007) mit dem Rang 3 bezeichnet werden, wenn angenommen wird,dass nur die Population einer Art, z.B. Cerodaphnia (Boothroyd et al. 2002) unterder Prädation leidet. Unter dem Aspekt, dass mehrere Arten betroffen sind,wie Jankowski et al. (2005) berichten: "The results showed a significant de-crease of small herbivorous crustaceans, i.e. Bosmina longirostris and juvenilecyclopoid copepods, in the medusa treatments of the microcosms and the en-closure experiments" und Pérez Bote & al. (2006) durch ihre differenziertenUntersuchungen zum Fraßverhalten der Medusen 6 Copepoden-, 5 Cladoceren-und 8 Rotatorienarten bestätigen, ist auch der Rang 4 in Erwägung zu ziehen.Folglich lässt sich die Wirkung von Craspedacusta sowerbii auf einheimische Artenkeineswegs als untergeordnet bezeichnen. Vielmehr kann Dodson & Cooper(1983) zugestimmt werden, dass die Art einen starken Effekt auf dieHerbivoren hat ("strong effect on the herbivores") den auch Jankowski et al.(2005) bestätigigen.
Bei vielen grundsätzlich für viele Seen geltenden Gesetzmäßigkeiten, mussjedoch bedacht sein, dass jeder See eine spezifische Artenzusammensetzung be-sitzt, so dass von einem ökologischen "Seeindividuum" gesprochen werden

223
kann. Das führt demnach auch zu einer individuellen Antwort eines Gewässersauf solche Einflüsse, wie es die Besiedlung von Craspedacusta sowerbii hat. Dieseseespezifische Antwort, wie sie es z.B. im Steinbruchsee Winkel durch das Aus-bleiben der Crustacea-Population im Jahre 2007 nach dem dreijährigen Massen-auftreten der Medusen und dem allmählichen Wiederaufbau der Medusenpopu-lation nach einem Jahr der Regeneration der Crustacea-Population im Jahr2008 gezeigt hat, kann in anderen Gewässern durchaus anders ausfallen. Darinliegen wohl auch Gründe, die zu der vielfach geäußerten Unberechenbarkeitdes Medusenauftretens beitragen.
6 Schlussfolgerungen im Blick auf Anpassungspotenzial und Beur-teilung der evolutiven Strategie von Craspedacusta sowerbiiFish (1970: 68) vermutet: "Optimum conditions are likely to be provided by afluvial lake system where the polyps develop mainly in the flowing water andthe medusae in the more static areas (Lytle, 1960), and this may account for thefrequent reports of medusae in artificially impounded waters such as thepresent record for Lake Maraetai".
So ist der biologische Sinn des Verhaltens der Polypen, sich auf einen exter-nen Zeitgeber - Sonnenstand und Mondphase - zu beziehen, in seinem vollenUmfang nur zu verstehen, wenn der ursprüngliche Lebensraum berücksichtigtwird, der in dem Flusssystem des Jangtsekiang liegt (Kimura 1937, Uchida1955). In diesem sollen sich die Gezeiten bis 600 km flussaufwärts bemerkbarmachen, da das Gefälle von Itschang bis zum Meer nur 2–4 cm auf 1 km be-trägt. Regelmäßiges Hochwasser steigt im Mittellauf bis zu 20 m und füllt beimÜbertritt eine Vielzahl großer und kleiner Seen. Die mitgeführte Sediment-fracht hat fruchtbare Marschböden abgelagert, wie sie z.B. in der Dongting-Ebene zu finden sind. Die bei uns zu beobachtende Toleranz für oligotrophebis mesotrophe Gewässerverhältnisse hat dieser Art daher diesen Lebensraummöglich gemacht. Zudem stellt auch Fohrmann (1985: 4) fest, dass der Polypvon Craspedacusta sowerbii "vorwiegend in schnellfließenden Gewässern" vor-kommt. Tappenbeck (2008) sieht darin sogar eine wesentliche Ursache für dieVerbreitung der Art: "Demnach kommt dem ca. 2 mm großen Polypen sicher-lich die größere Bedeutung zu und seine Vorkommen z.B. in einem großenFließgewässer im Substrat der Sohlbereiche sind die Ausgangsorte der Verbrei-tung der Art im gesamten Gewässersystem, einschließlich der Staubereiche, derparallelen Kiesungen in der Aue, u.a. anthropogener Gewässer und auch techni-schen Einrichtungen. Die Migration im Fließgewässer und von Fließgewässerzu Fließgewässer ohne (Kanal-) Verbindungen und durch ganz Europa könnennicht nur mit Drift und Wasservogelübertragung erklärt werden, sie sind unbe-kannt".

224
Die Fähigkeit des circalunaren Rhythmus dürfte bei Craspedacusta sowerbii nurdeswegen überraschen, weil sie im Süßwasser lebt, das üblicherweise keine Ge-zeiten aufweist. Doch die weit ins Land hinein spürbaren Süßwassertiden desJangtsekiang unterliegen den gleichen Zyklen wie die im Meer. D.h. auch hiertreten Nipp- und Springtiden auf. Die bei Vollmond (und Neumond) auftreten-den Springtiden im Flusssystem des Jangtsekiang sychronisieren nicht nur dieAbgabe der Geschlechtsprodukte sondern sorgen unter Umständen auch fürden Wasserübertritt in ufernahe Seen und Tümpel. Dadurch könnte die Aus-breitung über Gameten, befruchtete Eizellen oder Planulalarven erfolgen, dieinnerhalb einiger Tage zu Polypen heranwachsen. Von der Mondphase unab-hängige Überschwemmungen unterstützen die Verbreitung in Auseen undTümpel zusätzlich. Bei dem erwähnten Pegelstand von 20 m ist dann auch dieVerbreitung über die Medusen denkbar, deren Erfolg allerdings aufgrund ihrerrelativ kurzen Lebensdauer und mechanischen Empfindlichkeit vermutlich hin-ter dem der Polypen zurücktritt.
Bei der Frage, ob das Licht oder die Schwerkraft des Vollmonds für die Syn-chronisation der Strobilation ursächlich ist, können nur Mutmaßungen ange-stellt werden. Allerdings müsste bezüglich der Schwerkraft dann auch der Neu-mond zur Synchronisation führen, da es dabei auch zu einer Springtidekommt. Das ist jedoch aufgrund der Untersuchungen im See Winkel nicht zubeobachten.
Möglicherweise sind auch bei Craspedacusta sowerbii die im Genom der Steinko-ralle Acropora millepora gefundenen stammesgeschichtlich ursprünglichen Crypto-chrome anzunehmen, die mithilfe blauen Lichts die Information des ultravio-letten Lichtes weitergeben. Levy et al. vom Zentrum für Meeresstudien an derUniversität von Queensland St. Lucia, Australien, sehen die Funktion der bei-den zu verschiedenen Tages- und Nachtzeiten unterschiedlich aktiven Korallen-gene cry1 und cry2 in der Signalisierung des Anteils schädlichen ultraviolettenLichtes der oberen Wasserschichten für die Organismen. Dabei fand die For-schergruppe heraus, dass das Gen cry2 auf Mondlicht reagiert. Damit könntendie beiden Erbanlagen zu den Auslösern der spektakulären Massenvermehrunggehören, heißt es in "Science". http://www.n-tv.de/870053.html
Fautin (2002) bemerkt zur Bedeutung reproduktiver Fähigkeiten, dass repro-duktive Eigenarten vermutlich die evolutive Entwicklung spiegeln, weshalbviele zunächst auch taxonomische Relevanz hatten. Allerdings wurdezunehmend festgestellt, dass einige weit mehr variieren als erwartet. Je mehrdie Lebenszyklen auf Umweltbedingungen zu reagieren vermögen, destoweniger lassen sich Rückschlüsse aus ihnen zur Evolution ziehen. Und weiterstellt sie fest: "Wie ich bereits schrieb (Fautin 1991), sind die Lebenszyklus-Muster schwierig zu finden, vielleicht weil die anatomische und physiologische

225
Schlichtheit der Cnidaria sie in ihrer Evolution plastisch macht" (Fautin (2002:1741).
Zur Querteilung (Strobilation) stellt Fautin (2002) fest, dass diese eine repro-duktive Form ist, die für vergleichende Analysen äußerst interessant und aus-sichtsreich ist und Aufschluss über die Evolution und Ökologie gibt. Diese be-sondere Art vegetativer Fortpflanzung ist für alle drei Hauptklassen derCnidaria dokumentiert. Dabei stellt sie die Frage, ob deren Auftreten in ande-ren Taxa der Beleg für ein grundlegendes Potenzial für größere reproduktiveFlexibilität innerhalb der Cnidaria ist als es typischerweise offensichtlich ist.Die Umstände unter denen Strobilation auftritt und wie sie gesteuert wirdkönnten wichtig für das Verständnis der Ökologie und der Fortpflanzungsbio-logie sein.
In diesem Zusammenhang vermag Craspedacusta sowerbii mit ihrem Lebenszy-klus (Gen-Yu Sasaki 1999 a und b), die der r-Lebenszyklusstrategie zuzuordnenist, besonders neu bzw. schnell entstehende – allerdings auch ebenso schnellwieder vergehende – Lebensräume zu besiedeln, wie es die Überschwemmungs-auen mit ihren Seen und temporären Tümpeln des Flusssystems des Jangtseki-ang darstellen – und vielen in der Literatur berichteten Gewässern in denen dieArt eingewandert ist. Sowohl die sexuelle Vermehrung der Medusen aber be-sonders die vegetative durch Strobilation in Verbindung mit Frustelbildung umden Winter zu überdauern und die euryöke Potenz, sind die Erfolgsfaktorenfür eine schnelle, erfolgreiche Besiedlung sowie eine zügige vollständige Nut-zung des Lebensraumes. Das Ausbleiben von Konkurrenz im neu besiedeltenGewässer wirkt sich für die erfolgreiche Besiedlung und Wachstumsrate der Po-pulation ebenfalls günstig aus.
Bei Craspedacusta sowerbii findet die Begrenzung des Populationsansatzes durchSemelparitie statt. In semelparen Organismen, die nur einen Vermehrungsvor-gang in ihrem Leben durchführen, finden die üblichen evolutionärenBedingungen für die Evolution der Seneszenz keine Anwendung, weil es kei-nen Raum für mehrmalige Fortpflanzung gibt (Semelparitie-Hypothese vonTallamy & Brown 1999). Daher kommt der Betrachtung der elterlichen Inves-tition eine besondere Bedeutung zu.
Bei begrenzten Ressourcen reduzieren Investitionen für eine Leistung dieMöglichkeiten in weitere Leistungen zu investieren. Das führt zu negativenphäno- und/oder genotypischen Korrelationen zwischen Leistungen (tradeoffs,Stearns 1992). Diese negative Korrelation besteht zwischen der gegenwärtigenReproduktionsleistung eines Organismus einerseits und den zukünftigen Re-produktionsleistungen (= residueller Reproduktionswert) andererseits (Trivers1972). Das Ausmaß, mit dem Elternorganismen ihren residuellen Reprodukti-onswert durch Unterstützung der Nachkommenschaft verringern, wird als el-

226
terliche Investition ("parental investment") bezeichnet. Dieses sind die Kostender Unterstützung (Clutton-Brock 1991, Stearns 1992). Da die Selektion denGesamt-Reproduktionswert erhöht, sollte sich Unterstützung der Nachkom-men nur dann evolutiv durchsetzen, wenn ihre Vorteile die Kosten übersteigen(Clutton-Brock 1991). Ein solcher Netto-Nutzen der Unterstützung entstehtentweder durch relativ große Vorteile oder durch relativ niedrige Kosten (nachTallamy & Brown 1999 in: Stegmann 2002). Indem die Meduse von Craspedacus-ta sowerbii ihre Geschlechtsprodukte ins Wasser entlässt, entsteht somit keine el-terliche Investition über deren Entwicklung hinaus. Allerdings ist die Synchro-nisierung des Reifegrades der Elterntiere vor der Abgabe der Gameten durchausals Investition in o.g. Sinne aufzufassen. Es ist anzunehmen, dass sich für dieseder biologische Kostenaufwand niedrig hält und sich der Gesamt-Reprodukti-onswert vergleichsweise erhöht. Daher hat die circalunare Synchronisation derStrobilation einen erhöhten Selektionswert, der sich u.a. in der großen Fähig-keit zeigt, neue Gebiete weltweit zu besiedeln.
Tappenbeck (2008) bezweifelt den Status von Craspedacusta sowerbii als Neozo-on und nimmt vielmehr an, dass sie als "Agriozoon", d.h. als etabliertes Neozo-on zu bezeichnen ist. Für die hier betrachteten evolutiven Prozesse erscheintdiese Frage daher untergeordnet zu sein. Allerdings wird Tappenbeck (2008)zugestimmt, dass die in den letzten Jahren beobachtete Verbreitung der Art imHinblick auf die niedrige Temperatur im See Winkel (bezogen auf die Litera-turdaten), nicht unmittelbar auf den Klimawandel zurückzuführen ist.
Das zur Diskussion gestellte Faktorenbündel ist in seiner differenziertenökologischen Wirkung neben den in der Metagenese des circalunar gesteuertenLebenszyklus der Art liegenden Sicherungen optimal vorbereitet, neue Lebens-räume zu erobern, nachdem einmal die Süßwassertoleranz zur Einwanderungin Flüsse erworben worden ist. Dieser Prozess wurde möglicherweise im Jang-tsekiang durch das geringe Gefälle, das Auftreten der besonders weit ins Landreichenden Gezeiten und die regelmäßigen Überschwemmungen der Flussauebegünstigt. Insgesamt ist daher Craspedacusta sowerbii innerhalb ihrer Sippe als er-folgreiche progressive Species zu beurteilen.
DankIch möchte mich bei meinen Tauchlehrern des Tauchclubs Wetzlar (TCW) für meine auf Sicher-heit bedachte Ausbildung im Gerätetauchen bedanken. Ich danke auch meinen Tauchpartnerinnenund -partnern des TCW und des Herborner Unterwasserclubs (HUC), die mich auf vielenTauch-gängen begleitet und zu einem sicheren Wiederauftauchen beigetragen haben. Besonderer Dank giltmeinem Sohn Robert Peukert, der auf vielen Tauschgängen ein interessierter Partner war und michbei den Untersuchungen unterstützte. Frau Annelies Kolbe nahm freundlicherweise die englischeÜbersetzung der Zusammenfassung vor; ihr gilt mein herzlicher Dank.

227
LiteraturAcker, T. S. & A. M. Muscat (1976): The ecology of Craspedacusta sowerbii Lankester, a freshwa-
ter hydrozoan.- The American Midland Naturalist 95: 323-336, Notre Dame, Ind.Arbačiauskas, K. & J. Lesutienė (2005): The Freshwater Jellyfish (Craspedacusta sowerbii) in
Lithuanian Waters.- Acta Zoologica Lituanica 15(1): 54-57, VilniusBoothroyd, Ian K.G., M. Kay Etheredge & John D. Green (2002): Spatial distribution, size struc-
ture, and prey of Craspedacusta sowerbyi Lankester in a shallow New Zealand lake.-Hydrobiologia 468: 23-32, Dordrecht
Clutton-Brock, T. H. & C. Godfray. (1991): Parental investment.- In: Krebs, J. R. & N. B. Davies(eds): Behavioural Ecology: An Evolutionary Approach: 234-262, (Blackwell) Boston
Clutton-Brock, T. H. (1991): The evolution of parental care.- 368 pp., (Princeton University Press)Princeton, N.J.
Danli Wang, Shanliang Xu, Honglei Jiang, Hua Yang (2006): Tolerance of Craspedacusta sowerbyixinyangensis to the stresses of some ecological factors.- Ying Yong Sheng Tai Xue Bao, June 17(6): 1103-1106
Dexter, R. W., T. C. Surrarer & C. W. Davis (1949): Some recent records of the Freshwater Jelly-fish Craspedacusta sowerbii from Ohio and Pensylvania.- The Ohio Journal of Science 49: 235-241, Columbus, Ohio
Dodson, S. I. & S. D. Cooper (1983): Trophic relationships of the freshwater jellyfish Craspedacus-ta sowerbyi Lankester 1880.- Limnology and Oceanography 28: 345-351, Lawrence, Ks.
Dumont, H.J. (1994): The distribution and ecology of the fresh- and brakish- water medusae of theworld.- Hydrobiologia 272: 1-12, Dordrecht
Farkašova, M. & E. Stloukal (2007): Distribution of the Freshwater Jellyfish Craspedacusta sower-byi at the Podunajská Nížina Lowland (Slovakia).- Acta Zoologica Universitatis Comenianae47: 1-6, Bratislawa
Fautin, D.G. (2002): Reproduction of Cnidaria.- Canadian Journal of Zoology 80: 1735-1754, Otta-wa
Fish, G. R. (1970): Craspedacusta sowerbyi Lankester (Coelenterata: Limnomedusae) in New Zea-land Lakes.- New Zealand Journal of Marine and Freshwater Research 5: 66-69, Wellington
Fish, G. R. (1975): Further Records of Craspedacusta sowerbyi Lankester in North Island Lakes(Note) New Zealand Journal of Marine and Freshwater Research 9: 573-574, Wellington
Fleming, W. R., & D. H. Hazelwood (1967): Ionic and osmoregulation in the freshwater medusaCraspedacusta sowerbyi.- Comparative Biochemistry and Physiology 23: 911-915, London
Fohrmann, U. (1985): Polypenkolonien, Wanderfrusteln und Entwicklungsstadien der Medusenvon Craspedacusta sowerbii.- Publikationen zu wissenschaftlichen Filmen, Sektion Biologie, Se-rie 17, Nummer 23, Film D 1167; Institut für den wissenschaftlichen Film, Göttingen
Fritz, G. B., R. O. Schill, M. Pfannkuchen & F. Brümmer (2007): The freshwater jellyfish Craspe-dacusta sowerbii Lankester, 1880 (Limnomedusae, Olindiidae) in Germany, with a brief noteon its nomenclature.- Journal of Limnology 66: 54-59, Verbania
Fritz, G. B., M. Pfannkuchen, A. Reuner, R. O. Schill & F. Brümmer (2009): Craspedacusta sower-bii, Lankester, 1880 – population dispersal analysis using COI and ITS sequences.- Journal ofLimnology 68: 46-52, Verbania
Gen-yu Sasaki (1999): Freshwater Jellyfish (Craspedacusta sowerbyi). Micscape Magazine Nov.http://www.microscopy-uk.org.uk
Gen-yu Sasaki (1999): Sexual Reproduction in Freshwater Jellyfish (Craspedacusta sowerbyi).Micscape Magazine Dec. http://www.microscopy-uk.org.uk
Google Earth 2009 Tele Atlas Image 2009 GeoContent http://www.google.com/intl/en/ Eye alt962 m
Harrison, P. I., R. C. Babcock, G. D. Bull, J. K. Oliver, C. C. Wallace & B. L. Willis (1984): Massspawning in tropical reef corals.- Science 223: 1186-1189, Washington, D.C.

228
Hubschmann, J. H. & W. J. Kishler (1972): Craspedacusta sowerbii Lankester 1880 and Cordylo-phora lacustris Allman 1871 in Western Lake Erie (Coelenterata).- The Ohio Journal of Science72: 318-321, Columbus, Ohio
Jankowski, T., T. Strauss & H. T. Ratte (2005): Trophic interactions of the freshwater jellyfishCraspedacusta sowerbii.- Journal of Plankton Research 27: 811-823, London
Jung Hee Park (1998): First Record of a Freshwater Jellyfish, Craspedacusta sowerbii Lankester1880 (Limnomedusae, Olindiidae) from Reservoirs in Korea.- Korean Journal of BiologicalSciences 2: 303-308, Seoul
Kimura, S. (1937): Distribution and some Notes on the Chinese Freshwater Medusa.- Bulletin ofthe Shanghai Science Institute 6: 319-342, Shanghai
Kuhl, W. & G. Kuhl (1956): Craspedacusta sowerbii - Polypenkolonien und Bildung von Wander-frusteln.- Film B 714 des IWF, Göttingen. Publikation von G. Kuhl, Göttingen 1958, 14 pp.
Levy, O. et al. (2007): Innere Uhr der Korallen.- Science 318: 467, Washington D.C. (zitiert nach:n-tv.de Fundsache Nr. 227 24.10.2007 http://www.n-tv.de/870053.html
Levy, O., L. Appelbaum, W. Leggat, Y. Gothlif, D. C. Hayward, D. J. Miller, O. Hoegh-Guldberg(2007): Light-Responsive Cryptochromes from a Simple Multicellular Animal, the Coral Acro-pora millepora.- Science 19 October 2007: Vol. 318. no. 5849, pp. 467 - 470 DOI: 10.1126/sci-ence.1145432
Ludwig, H. W., Becker, N. & Rambow, C. (1979): Die Süßwassermeduse Craspedacusta sowerbii(Hydrozoa: Limnomedudusae) im Rhein-Neckar-Gebiet (Heidelberger Beiträge zur Faunistikund Ökologie des Rhein-Neckar-Gebiets Nr. 1).- Beiträge zur Naturkundlichen Forschung inSüdwestdeutschland 38: 141-153, Karlsruhe
Lundberg, S. & J. E. Lundberg (2003): Medusainvasion i varma sjöar.- Fauna och Flora 98: 18-28,Göteborg
Lundberg, S., J. E. Svensson & A. Petrusek (2007): Sötvattensmaneten Craspedacusta - enkönandepopulationer brevis för upprepad invasion.- Fauna och Flora 102: 18-22, Göteborg
Lytle, C. F. (1960): A note on distribution patterns in Craspedacusta.- Transactions of the Americ-an Microscopical Society 79: 461-469, Columbus, Ohio
Mannak, E. alias Geckoman (2009): Duikerslog.nl. Letzter Zugriff: 30.4.2009 http://www.duikers-log.nl/public/detail.php?id=Geckoman&datum=1240132620
Marshall, S. M., & Stephenson, T. A. (1933): The breeding of reef animals. Part I. The corals.-Scientific Report of the Great Barriee Reef Expedition 1928-1929, 3: 219-245.- In: Fautin, D. G.(2002): Reproduction of Cnidaria.- Canadian Journal of Zoology 80: 1735-1754, Ottawa
DIN 38410-1 (2004): Deutsche Einheitsverfahren zur Wasser, Abwasser- und Schlammuntersu-chung – Biologisch-ökologische Gewässeruntersuchung (Gruppe M) – Teil 1: Bestimmung desSaprobienindex in Fließgewässern (M 1).-80 pp., (Beuth) Berlin
Neobiota-Watcher Aktion (2005) http://neobiota.umwelt.vdst.de Letzter Zugriff: 24.4.2009New Hampshire Department of environmental Services (Hg.) (2004): Freshwater Jellyfish in New
Hampshire - They do exist! Environmental Fact Sheet. http://des.nh.gov/organization/com-missioner/pip/factsheets/bb/documents/bb-51.pdf
Peard, T. (2002): Natural History of the the freshwater jellyfish http://www.jellyfish.iup.edu/his-tory.html. Last updated 2/08/08. Email: [email protected]
Pérez Bote, J., A. Muñoz, R. M. R. Roso & A. J. Romero (2006): First record of Craspedacustasowerbyi Lankester, 1880 (Cnidaria: Limnomedusae: Olindiidae) in the Proserpina Reservoir(Extremadura, SW Spain) with notes on their feeding habits.- Belgian Journal of Zoology 136:163-166, Diepenbeek
Petrusek, A., J. Šedivy, S. Lundberg & J.-E. Svensson (2005): Detecting multiple invasions of Cras-pedacusta sowerbyi by indentification of the sex of the medusae. INWAT: Biological invasionsin inland waters, Firenze, May 5-7
Peukert, D. (1982): Eine meeresbiologische Exkursion - Anregung für die Sekundarstufe II (Leis-tungskurs).- Unterricht Biologie 6(67): 29-37, Seelze

229
Peukert, D. (1985): Experimente im ökologischen Schulgarten während des Unterrichts in dergymnasialen Oberstufe.- TASPO-Magazin (5): 19-25, Braunschweig
Peukert, D. (1988): Ökologische Nischen im Wattenmeer - Unterrichtsmodell für die Sekundarstu-fe II.- Unterricht Biologie 12(136): 41-45; Seelze
Ricciardi, A. & J. Cohen (2007): The invasiveness of an introduced species does not predict its im-pact.- Biological Invasions 9: 309-315, Dordrecht
Schill, R. O.: Steckbrief Süsswassermeduse Craspedacusta sowerbii. http://neobiota.umwelt.vdst.de/pdf/Craspedacusta.pdf (Letzter Zugriff 4.6.2008)
Schmidt, B. (2009): VDST Sonderkurs Gewässeruntersuchung HTSV-See Schönbach (HessischerTauchsport Verband e.V.)
Scott, M. (1963): Fresh-water Jellyfish in Maine. http://www.state.me.us/dep/blwq/doclake/jel-ly.htm (Letzter Zugriff 14.7.2009).
Silva, W.M. & K. F. Roche (2007): Occurence of the freshwater jellyfish Craspedacusta sowerbii(Lancester, 1880) (Hydrozoa, Limnomedusae) in a calcareous lake in Mato Grosso do Sul, Bra-zil. Biota Neotropica, v7 (n1) – bn()210701 http://www.biotaneotropica.org.br
Sládek , T. & Šraier, Z, (2005): In: Farkašova, Miroslavá & Eduard Stloukal: Distribution of theFreshwater Jellyfish Craspedacusta sowerbyi at the Podunajská Nížina Lowland (Slovakia).-Acta Zoologica Universitatis Comenianae 47(1): 1-6, Bratislawa
Spadinger, R. & G. Maier (1999): Prey selection and diet feeding of the freshwater jellyfish,Craspedacusta sowerbyi.- Freshwater Biology 41: 567-573, Oxford
Stark, H. (2005): Das "Hausgewässer" beobachten. In: Tauchen in heimischen Seen - Ein kleinerBiologieführer für Sporttaucher - Informationen und Anregungen für bewussteres und erlebnis-reicheres Tauchen. http://suesswassertauchen.homepage.t-online.de/beobacht.htm Stand15.12.2005
Stark, H. (2005): Das "Hausgewässer" beobachten. Letzter Besuch der Homepage 24.4.2009http://suesswassertauchen.homepage.t-online.de/beobacht.htm
Stearns, S.C. (1992): The Evolution of Life Histories.- 248 pp., (Oxford University Press) OxfordStegmann, U. E. (2002): Brutpflege, Lebensgeschichte und Taxonomie südostasiatischer Membraci-
den (Insecta: Homoptera). http://www.opus-bayern.de/uni-wuerzburg/volltexte/2002/236/pdf/k1.pdf Letzter Zugriff: 24.4.2009
Steiner, S. (2002): Dynamik und Energietransfer einer planktischen Crustaceengemeinschaft in Ab-hängigkeit von der Nahrungsgrundlage und den Planktivoren.- Dissertation Fakultät für Forst-,Geo- und Hydrowissenschaften der Technischen Universität Dresden, 152 pp.
Steinmann, P. (2008): Aquatische Neozoen Süsswasserqualle: Süsswasserquallen – Craspedacusta so-werbyi. Gewässerbiologische Studien Wissenschaftliche Taucharbeiten, Stein am Rheinhttp://www.psteinmann.net/neo_craspedacusta.html
Strauss, T. (1996): Planktonökologie und aquatische Nahrungsnetze.- Forschungsinstitut für Öko-systemanalyse und –bewertung e.V. (gaiac) RTWH Aachen University http://www.gaiac.rwth-aachen.de/index.htm?http://www.gaiac.rwth-aachen.de/Forschung/Planktonoekologie/Plank-tonoekologie.html
Tallamy D. W. & W. P. Brown (1999): Semelparity and the evolution of maternal care in insects.-Animal Behaviour 57: 727-730, Amsterdam
Tappenbeck, L. (2008): Faunistik und Ökologie der Süßwasserqualle Craspedacusta sowerbyi (Lan-cester 1880) - Versuch einer aktuellen Bestandsanalyse für Deutschland.- Artenschutzreport 22:12-14, Jena
Tappenbeck, L. (4/2003): Ein Beitrag zur Autökologie und zum Vorkommen der SüßwasserqualleCraspedacusta sowerbyi Lancester, 1880 in Deutschland. Sporttaucher Nürnberg
Trivers, R. L. (1972): Parental investment and sexual selection.- In Campbell, B. (ed.): Sexualselection and the descent of man, 1871-1971: 136-179, Chicago

230
Uchida, T. (1955): Dispersal in Japan of the freshwater Medusa, Craspedacusta sowerbii Lankaster,with remarks on C. iseana (Oka and Hara).- In: Annotationes Zoologicae Japonese 28: 114-120,Tokyo
Verband Deutscher Sporttaucher e.V. (VDST) & Bundesamt für Naturschutz (BfN) (ed.) (2008):Projekt Neobiota - Steckbrief Süsswassermeduse Craspedacusta sowerbii www.neobiota.info
Wieser, Gabriele (1993): Zum Vorkommen von Medusen der Süßwasserpolypen Craspedacusta so-werbii (Limnomedusae, Olindiidae, Coelenterata) im Weizelsdorfer Baggersee (Kärnten).- Ca-rinthia II 183/103: 255-260, Klagenfurt
Anschrift des Verfassers: Dipl.-Biol. Detlef E. Peukert, Bergstraße 29, D-35582 Wetzlar,[email protected]
Manuskripteingang: 2009-06-08Angenommen: 2009-06-28




















