
Chronic exposure of rats to noise: Relationship between long-term memory deficits and slow wave...
10
Behavioural Brain Research 171 (2006) 303–312 Research report Chronic exposure of rats to noise: Relationship between long-term memory deficits and slow wave sleep disturbances A. Rabat a,b,∗ , J.J. Bouyer a,b , O. George a,b , M. Le Moal a,b , W. Mayo a,b a Inserm, U588, Physiopathologie du Comportement, Institut Fran¸ cois Magendie, Bordeaux F-33000, France b Universit´ e Victor Segalen Bordeaux II, 146 rue L´ eo Saignat, 33077 Bordeaux Cedex F-33000, France Received 11 January 2006; received in revised form 4 April 2006; accepted 7 April 2006 Available online 22 May 2006 Abstract Noise is now recognized as a serious health problem in our modern societies. Although its deleterious and direct effects on cognitive tasks (long-term memory, mental arithmetic activity, visual tasks, etc.) are clearly admitted, no studies have determined a delayed indirect effect of noise on cognitive processes. Furthermore, the link between sleep disturbances related to environmental noise (EN) exposure and these indirect deteriorations of human performances has never been demonstrated. This could be due to inappropriate evaluation of sleep as well as to uncontrolled and confounding factors such as sex, age, and also inter-individual vulnerability. Based on a recently validated animal model [Rabat A, Bouyer JJ, Aran JM, Le Moal M, Mayo W. Chronic exposure to an environmental noise permanently disturbs sleep in rats: inter-individual vulnerability. Brain Res 2005;1059:72–82], aims of the present study were (i) to determine long-term memory (LTM) deficits following a chronic exposure to EN and (ii) to link these behavioral problems to sleep disturbances related to EN. For this purpose in a first experiment, LTM performances were evaluated before and following 9 days of EN. Results show LTM deficits following a chronic exposure to EN with inter-individual vulnerability. Vulnerability profile was related to the psychobiological profile of rats. Results of the second experiment show LTM deficits correlated to both debt of slow wave sleep (SWS) and to daily decrease of SWS bout duration. Our results demonstrate that chronic exposure to noise indirectly disturbs LTM possibly through SWS disturbances and suggest a possible role of the stress hormonal axis in these biological effects of noise. © 2006 Elsevier B.V. All rights reserved. Keywords: Long-term memory; Environmental Noise; Chronic exposure; Sleep; Inter-individual vulnerability; Rat 1. Introduction Noise is now recognized as a serious health problem in our modern societies [30,45]. Among non-auditory deleterious effects of noise, sleep disturbances are clearly documented (for review see Refs. [48,60]). Moreover, in humans, direct effects of noise on various cognitive abilities such as long-term mem- ory, mental arithmetic activity, visual tasks, etc. (for review see Refs. [13,27,32,59]) have been demonstrated. Convergent data indicate (1) that the relationship between noise effects and human performances follows a U-inverted shape with a facil- itating effect of noise until 70 dB and a deleterious one above ∗ Corresponding author. Present address: Inserm EMI 0216, Laboratoire de Neurobiologie de la Douleur Trig´ eminale, Facult´ e Dentaire, 13 Boulevard Charles de Gaulle, 63000 Clermont Ferrand, France. Tel.: +33 4 73 17 73 16; fax: +33 4 73 17 73 06. E-mail address: [email protected] (A. Rabat). this level, and (2) that unpredictable intermittent noises are more noxious than continuous ones [23,45]. In a recent review, Griefahn [23] highlighted the crucial question of a link between sleep disturbances related to environ- mental noise exposure and human performances deterioration the next day. Some studies, focused on this question, were field studies [24,66], known to be less sensitive than laboratory studies [23]. All these studies have evaluated human performances with simple or four choice reaction time tests [24,40,41,46,47,66], which assess psychomotor performances but not cognitive dis- turbances [65]. Moreover some of these studies have analyzed sleep disturbances by using either desynchronization level of EEG [40,41] or subjective sleep questionnaires [46,47], two parameters often unrelated to sleep disturbances [1,33,37], or have gathered sleep data from subjects of heterogeneous age [24,66], an exogenous factor known to interfere with effects of noise on sleep [22,23]. While some field studies have reported difficulties in school learning for children living in noisy areas (for review see Ref. 0166-4328/$ – see front matter © 2006 Elsevier B.V. All rights reserved. doi:10.1016/j.bbr.2006.04.007
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Chronic exposure of rats to noise: Relationship between long-term memory deficits and slow wave...

Behavioural Brain Research 171 (2006) 303–312
Research report
Chronic exposure of rats to noise: Relationship between long-termmemory deficits and slow wave sleep disturbances
A. Rabat a,b,∗, J.J. Bouyer a,b, O. George a,b, M. Le Moal a,b, W. Mayo a,b
a Inserm, U588, Physiopathologie du Comportement, Institut Francois Magendie, Bordeaux F-33000, Franceb Universite Victor Segalen Bordeaux II, 146 rue Leo Saignat, 33077 Bordeaux Cedex F-33000, France
Received 11 January 2006; received in revised form 4 April 2006; accepted 7 April 2006Available online 22 May 2006
Abstract
Noise is now recognized as a serious health problem in our modern societies. Although its deleterious and direct effects on cognitive tasks(long-term memory, mental arithmetic activity, visual tasks, etc.) are clearly admitted, no studies have determined a delayed indirect effect ofnoise on cognitive processes. Furthermore, the link between sleep disturbances related to environmental noise (EN) exposure and these indirectdeteriorations of human performances has never been demonstrated. This could be due to inappropriate evaluation of sleep as well as to uncontrolledaJBEeVoL©
K
1
oeroosdhi
NCf
0d
nd confounding factors such as sex, age, and also inter-individual vulnerability. Based on a recently validated animal model [Rabat A, BouyerJ, Aran JM, Le Moal M, Mayo W. Chronic exposure to an environmental noise permanently disturbs sleep in rats: inter-individual vulnerability.rain Res 2005;1059:72–82], aims of the present study were (i) to determine long-term memory (LTM) deficits following a chronic exposure toN and (ii) to link these behavioral problems to sleep disturbances related to EN. For this purpose in a first experiment, LTM performances werevaluated before and following 9 days of EN. Results show LTM deficits following a chronic exposure to EN with inter-individual vulnerability.ulnerability profile was related to the psychobiological profile of rats. Results of the second experiment show LTM deficits correlated to both debtf slow wave sleep (SWS) and to daily decrease of SWS bout duration. Our results demonstrate that chronic exposure to noise indirectly disturbsTM possibly through SWS disturbances and suggest a possible role of the stress hormonal axis in these biological effects of noise.2006 Elsevier B.V. All rights reserved.
eywords: Long-term memory; Environmental Noise; Chronic exposure; Sleep; Inter-individual vulnerability; Rat
. Introduction
Noise is now recognized as a serious health problem inur modern societies [30,45]. Among non-auditory deleteriousffects of noise, sleep disturbances are clearly documented (foreview see Refs. [48,60]). Moreover, in humans, direct effectsf noise on various cognitive abilities such as long-term mem-ry, mental arithmetic activity, visual tasks, etc. (for reviewee Refs. [13,27,32,59]) have been demonstrated. Convergentata indicate (1) that the relationship between noise effects anduman performances follows a U-inverted shape with a facil-tating effect of noise until 70 dB and a deleterious one above
∗ Corresponding author. Present address: Inserm EMI 0216, Laboratoire deeurobiologie de la Douleur Trigeminale, Faculte Dentaire, 13 Boulevardharles de Gaulle, 63000 Clermont Ferrand, France. Tel.: +33 4 73 17 73 16;
ax: +33 4 73 17 73 06.E-mail address: [email protected] (A. Rabat).
this level, and (2) that unpredictable intermittent noises are morenoxious than continuous ones [23,45].
In a recent review, Griefahn [23] highlighted the crucialquestion of a link between sleep disturbances related to environ-mental noise exposure and human performances deteriorationthe next day. Some studies, focused on this question, were fieldstudies [24,66], known to be less sensitive than laboratory studies[23]. All these studies have evaluated human performances withsimple or four choice reaction time tests [24,40,41,46,47,66],which assess psychomotor performances but not cognitive dis-turbances [65]. Moreover some of these studies have analyzedsleep disturbances by using either desynchronization level ofEEG [40,41] or subjective sleep questionnaires [46,47], twoparameters often unrelated to sleep disturbances [1,33,37], orhave gathered sleep data from subjects of heterogeneous age[24,66], an exogenous factor known to interfere with effects ofnoise on sleep [22,23].
While some field studies have reported difficulties in schoollearning for children living in noisy areas (for review see Ref.
166-4328/$ – see front matter © 2006 Elsevier B.V. All rights reserved.oi:10.1016/j.bbr.2006.04.007

304 A. Rabat et al. / Behavioural Brain Research 171 (2006) 303–312
[60]), no relationships between learning problems and chronicnoisy exposures have been really demonstrated [60]. A delayedindirect effect of noise on human performances through sleepdisturbances remains an unanswered question.
Animal models are a useful strategy to address such complexquestions. Recent studies, using different animal models, havetested effects of noise on behavioral performances [3,42,54,62].However, the purpose of these experiments were different fromours. Indeed they have tested direct effects of noise on learningand memory processes [3,42,54,62]. Besides, these studies haveused white noise, which is not representative of a human noisyenvironment. We have recently developed and validated a reli-able experimental situation in which a human noisy environmentwas adapted to rat’s audition [55] and using this experimentalmodel we have demonstrated deleterious effects of an acute anda chronic noise exposure on sleep [55,56].
Aims of the present study were thus to determine (i) a pos-sible delayed effect of a chronic exposure to an environmentalnoise (EN) exposure on long-term memory and (ii) a relation-ship between long-term memory deficits and sleep disturbancesinduced by EN. Experiments 1 and 2 addressed respectivelythese two objectives.
2. Methods
2.1. Animals and housing conditions
tioaa
2
s(wemc(r
The behavioral procedure consisted of two phases. In the first phase, rats wereintroduced in the central alley (starting box) for 30 s upon which the sliding doorswere removed and the rats were allowed to explore the “acquisition” configura-tion for 30 min. Immediately after completion of the acquisition trial, animalswere replaced in their home cages for an inter-trial interval of 4 h. During thesecond trial, rats were placed in the starting box and allowed to explore the“restitution” configuration for 20 min. These conditions, i.e., 30 min for acqui-sition trial and 4 h for inter-trial interval were determined previously as adequatefor the evaluation of the noise effects on long-term memory since naıve rats per-formed correctly. A recognition score was computed for each rat as the mean ofthree behavioral parameters: locomotor activity, number of visits and time spentin the modified compartment during the “restitution”, expressed as percentageof those recorded in the familiar compartment. A recognition score of 100% wasobserved when animals explored equally the two compartments.
2.3. Behavioral reactivity to novelty
Locomotor reactivity to novelty was evaluated in a novel environment con-sisting of a circular corridor (10 cm wide and 70 cm in diameter). Four photo-electric cells placed at the perpendicular axes of this apparatus automaticallyrecorded locomotion. The locomotor response was measured over a period of2 h between 16:00 and 18:00 h immediately after the animals were placed in thecorridor. The score of each animal, i.e., the mean number of photocell countscumulated over the 2 h of the behavioral test, reflects individual reactivity intothe novel environment [51]. Based on the median of the population, animalswere divided into two subgroups. The first subgroup defined as “high locomo-tor reactive (HR)” included rats presenting scores above the median value andthe second defined as “low locomotor reactive (LR)” the remaining rats [51].Locomotor reactivity to novelty was conducted 8 days after rats arrival at thelaboratory and 2 days before the spatial long-term memory test.
2
(pafetnd
2
ier
F the ta obile( “restitc latede
Thirty-seven male Long Evans rats (Janvier, France) 3 months old at theime of their arrival were used. Immediately upon arrival, animals were housedndividually in a temperature (22 ± 2 ◦C) and humidity (60%) controlled roomn a 12:12 light/dark schedule (lights on at 08:00) with ad libitum access to foodnd water. All experiments were carried out in accordance with the guidelinespproved by European Communities Council Directive (89/606/EEC).
.2. Spatial long-term memory test (Fig. 1)
Long-term memory performances were evaluated using a two-trial long-termpatial memory task [50]. The apparatus consisted of two equal compartments30 cm × 30 cm × 45 cm) separated by a central alley (30 cm × 10 cm × 45 cm)ith two opposite sliding doors (8 cm) connecting the central alley with
ach compartment. Two mobile square parallelepipeds (12 cm × 12 cm × 45 cm)ade of gray Plexiglas were used in each box to create either two different spatial
onfigurations or identical spatial environments, with two equal compartments“acquisition” configuration). Boxes were equipped with photoelectric beams toecord locomotor activity, number of visits and time spent in each compartment.
ig. 1. Two-trial recognition task. During 30 min, rats were allowed to explorefter, animals were replaced in their home cages for an inter-trial of 4 h and one m“restitution” configuration). During 20 min, rats were allowed to explore thisompartment/exploratory behavior in the non modified compartment) was calcuqually the two compartments.
.4. Surgery
Surgery was carried out at 14 weeks of age under pentobarbital anaesthesia330 mg/kg, i.p.). To record cortical EEG, four miniature screw electrodes werelaced through the skull 1.5 mm either side of the central suture, 1.5, 3.0, 4.5nd 6.0 mm anterior to lambda. Electrode wires were soldered to miniature plugsastened to the skull in a conventional set-up. In order to avoid for a detrimentalffect of surgery upon the auditory capacities of the animals, we have used round-ipped ear rods. In comparison to the classical ones these adapted ear rods areot deleterious to the auditory capacities of the animals as it was previouslyemonstrated [55].
.5. EEG recordings and signal processing
Sleep states were monitored by recording EEG activity from chronicallymplanted cortical electrodes. Individual recordings were made from the twolectrodes providing the largest theta rhythm amplitude. Such derivation-ecorded signals depend on the vigilance level of the rat [2,55]. Signal of the
wo identical spatial configurations (“acquisition” configuration). Immediatelysquare parallelepiped was moved in order to create a new spatial configuration
ution” configuration. A recognition score (% exploratory behavior in the newfor each rat. A recognition score of 100% was observed when animals explored

A. Rabat et al. / Behavioural Brain Research 171 (2006) 303–312 305
Fig. 2. Design of experiment 1. Long-term memory performances were evaluated before (day 5) and following 9 days of EN exposure (day 10). After four days ofhabituation to the environment conditions of the noisy boxes, they were allowed a 24 h period (Control Day) with a LAeq, 24 h of 40 dB. Noise exposure began thefollowing day and lasted 9 days (Day 1 to 9). An additional control group was added in which long-term memory performances were evaluated before and after aperiod of 9 days with no noise events (Control group).
EEG was band-pass filtered (1–200 Hz) with an additional band-stop at 50 Hzand was amplified by 20,000. This automated EEG recording is able to discrimi-nate the vigilance level without any requirement for electromyogram recordings.The method has been detailed in previous studies [2,43,55].
EEG recordings began two weeks after the surgery on freely moving rats intheir home cage in a soundproof, temperature and humidity controlled room. Allrecordings began at 8:00 a.m. and lasted 10 days. All rats were submitted to thesame schedule. After three days of habituation to the recording environment anddevice, EEGs were recorded for a 24 h period (Control) with a global intensity ofnoise (Leq 24 h) of 40 dB. Animals were then exposed to 9 days of noise startingthe following day (days 1–9). Noise exposure began at 8:00 a.m.
2.6. Noise exposure conditions
In this study it was decided to adapt to the rat a noise representative of anoisy environment, i.e., the noise of a French aircraft carrier boat, similar tothat recorded from a noisy Airport environment. Since the audiogram of rats,compared to the human one, is characterized by a lower capacity for detectinglow frequencies, under 500 Hz, and a better capacity for detecting high, over8000 Hz, frequencies [26], the physical characteristics of the noise were adaptedto the rat’s audiogram in order to be more relevant to the auditory capacities ofrats. The technical adaptation has been detailed in a previous work [55]. Brieflyan audio software (Wave Lab 3.0, Steinberg, Germany) which enables to translateall the frequencies of the noise from the human audiogram to the rat’s one hasbeen used.
This noise is designated environmental noise since it has major octave bandfrequencies ranging from 125 to 2000 Hz, which is similar to environmentalnoises such as aircraft noise and road traffic noise [35,49]. Environmental noise(EN) was composed of a background noise (70 dB) and of several unpredictablenoise events with levels ranging from 85 to 103 dB and a duration ranging from1
of EN is expressed by the equivalent sound level (LAeq, T) which determines theglobal level of noise over a period of time, T, according to this formula: LAeq,T =10 × log(1/T )
∫ T
010L(t)/10 dt[49]. This acoustic parameter is representative of
the effects of noise on sleep [38]. The global intensity of EN, expressed by anacoustical parameter, Leq 24 h, is 88 dB. EN was played by a mixer softwarewhich was connected to 16 tweeters (two loudspeakers for each box, as small asa rat’s individual box) by four amplifiers. These amplifiers produced the sameintensity from 0.5 to 20 kHz.
2.7. Experimental design
2.7.1. Experiment 1 (Fig. 2, 27 rats)Long-term memory performances were evaluated before (Pre-EN) and fol-
lowing 9 days of EN exposure (Post-EN). An additional control group wasadded in which long-term memory performances were evaluated before andafter a period of 9 days with no noise events (Control group). Three weeks aftertheir arrival, animals were tested in the behavioral task. The following day, ratswere placed in the noisy boxes. After four days of habituation to the environ-ment conditions, they were allowed for a 24 h period (Control Day) with a LAeq,24 h of 40 dB. Noise exposure began the following day at 08:00 h and lasted 9days (days 1–9). The day after (day 10) rats were retested in the behavioral task.For the control group the experiment design was similar except a lack of noiseexposure.
2.7.2. Experiment 2 (Fig. 3, 10 rats)Experimental design for sleep recordings and EN exposure were similar to
the one previously described [56] and long-term memory performances wereevaluated in an experimental design similar to experiment 1. Briefly, following1 week of habituation to the housing conditions of the laboratory rats wereo
F viousa ontrol
8 s to 39 s (spaced by silent epochs ranging from 20 to 165 s). The noise level
ig. 3. Design of experiment 2. Experimental design was similar to that of the prend sleep recordings conditions, rats were sleep recorded during 10 days, one c
perated for sleep recordings. After 9 days, rats were tested in the behavioral
experiment. For sleep recordings, after 4 days of habituation to the environmentday and 9 days of environmental noise exposure.

306 A. Rabat et al. / Behavioural Brain Research 171 (2006) 303–312
task. The following day, they were habituated to the sleep recordings conditionsduring 4 days and then were sleep recorded during 10 days, one control dayand 9 days of noise. The following day they were retested in the behavioraltask.
2.8. Statistical analysis
Data were analyzed using a statistical software (Statistica®, USA). In allcases, a normality test and an equal variance test were carried out. T-test was usedto compare recognition score to chance score. One-way ANOVA with repeatedmeasures was used to compare recognition scores between Pre-EN (the situa-tion before noise exposure) and Post-EN (after a chronic environmental noiseexposure) and also between Pre-Control and Post-Control. One-way ANOVAwas used to compare variation of long-term memory performances between“long-term memory vulnerable rats” (“V” rats) and “long-term memory resis-tant rats” (“R” rats) and to compare locomotor reactivity to novelty betweenthese same two experimental groups of rats. When the normality test failed, aKruskall–Wallis test and an U Mann–Whitney test were used to compare num-ber, duration of bouts and total amount of slow wave sleep and paradoxical sleepbetween “V” and “R” rats during control day and the 9 days of environmentalnoise.
3. Results
3.1. Experiment 1: effect of a chronic exposure toenvironmental noise on long-term memory: inter-individualdifferences in long-term memory deficits are related to thepsychobiological profile of rats
3c
ioflt
Fig. 4A) and especially not significantly different from the pre-vious one (F(1,9) = 0.27, p > 0.61, Fig. 4A).
3.1.2. LTM performances of rats exposed 9 days toenvironmental noise (EN)
Before noise exposure, rats explored significantly morethe novel compartment compared to the familiar one (t-test:p < 0.001, Fig. 4B). Thus they were able to remember what theyhave seen 4 h before and did not differ from the control group(F(1,25) = 0.11, p > 0.74). After being exposed to EN, a detec-tion of the novelty was still observed (t-test: p < 0.05, Fig. 4B).However the LTM score was significantly lower than the previ-ous one (F(1,16) = 6.06, p < 0.05, Fig. 4B).
3.1.3. Locomotor activity of the two experimental groups inthe behavioral test
The global locomotor activity recorded during the acquisi-tion session was not significantly different between the situa-tion before and the situation after the 9 days of EN exposure(F(1,16) = 0.58, p > 0.45). The same conclusion was obtainedfor the control group (F(1,9) = 2.38, p > 0.15).
3.1.4. Inter-individual vulnerability in LTM deficitsinduced by EN
Variation of LTM performances showed inter-individual vari-a(Lna““pr
F (Post-w scores re (PrE he recC (Pos
.1.1. Long-term memory (LTM) performances in theontrol group
Before the period of 9 days of silence, rats explored signif-cantly more the novel compartment compared to the familiarne (t-test: p < 0.01, Fig. 4A). Thus their long-term memory per-ormances were intact. After being exposed to 9 days of silence,ong-term memory performances were not modified as the long-erm memory score was highly significant (t-test: p < 0.001,
ig. 4. (A) Recognition score before (Pre-Control) and after 9 days of silenceas not disturbed. Comparison between the recognition score and the chance
cores of Pre-Control and Post-Control: p > 0.61. (B) Recognition scores befoN induced a delayed long-term spatial memory deficit. Comparison between tomparison between recognition scores before (Pre-EN) and after 9 days of EN
bility (Fig. 5A). Indeed some rats had few or no LTM deficitsLTM-resistant rats or “R” rats) whereas other ones had strongTM disturbances (LTM-vulnerable rats or “V” rats) followingoise exposure (Fig. 5A). After noise exposure, there was thussignificant difference for LTM disturbances between “V” andR” rats (V–R effect: F(1,15) = 35.08, p < 0.001, Fig. 5B) withV” rats showing significant LTM deficits (V rats-basal value:< 0.001, Fig. 5B) and “R” rats showing no such deficits (R
ats-basal value: p > 0.23, Fig. 5B).
Control). After 9 days without environmental noise long-term spatial memory(rectangular bar): **p < 0.01; ***p < 0.001. Comparison between recognitione-EN) and just after 9 days of environmental noise (Post-EN). The 9 days ofognition score and the chance score (rectangular bar): *p < 0.05; ***p < 0.001.t-EN): #p < 0.05.

A. Rabat et al. / Behavioural Brain Research 171 (2006) 303–312 307
Fig. 5. (A): Total (grey bar) and individual (points) variation of long-term spatial memory (LTM) performance (% (Post-EN–Pre-EN)/Pre-EN). Results show a greatvariation of LTM performances between resistant rats (white points), which had few or no LTM deficits, and vulnerable rats (black points) with high of very high LTMdeficits. (B) Variation of LTM performances for vulnerable (V) and resistant (R) rats. Statistical differences with basal value: ***p < 0.001. Statistical differencesbetween vulnerable and resistant rats: ###p < 0.001.Vulnerable rats showed LTM deficits whereas resistant rats had no such deficits.
3.1.5. Significant correlation between the psychobiologicalprofile of rats and their individual LTM disturbancesfollowing noise exposure
Locomotor reactivity to novelty differed significantlybetween “V” and “R” rats (V–R effect: F(1,15) = 19.44p < 0.001; Fig. 6A). A significant and negative correlation wasfound between individual variation of LTM performances after9 days of EN and individual locomotor reactivity to novelty(r = −0.59; p = 0.012; Fig. 6B). Subjects with a high reactivityto novelty were the more affected in LTM following chronicnoise exposure; conversely subjects with low reactivity to nov-elty were the less affected in LTM.
3.2. Experiment 2: relationship between long-term memorydeficits and slow wave sleep disturbances induced by achronic exposure to EN
3.2.1. Effects of a chronic noise exposure on LTMperformances
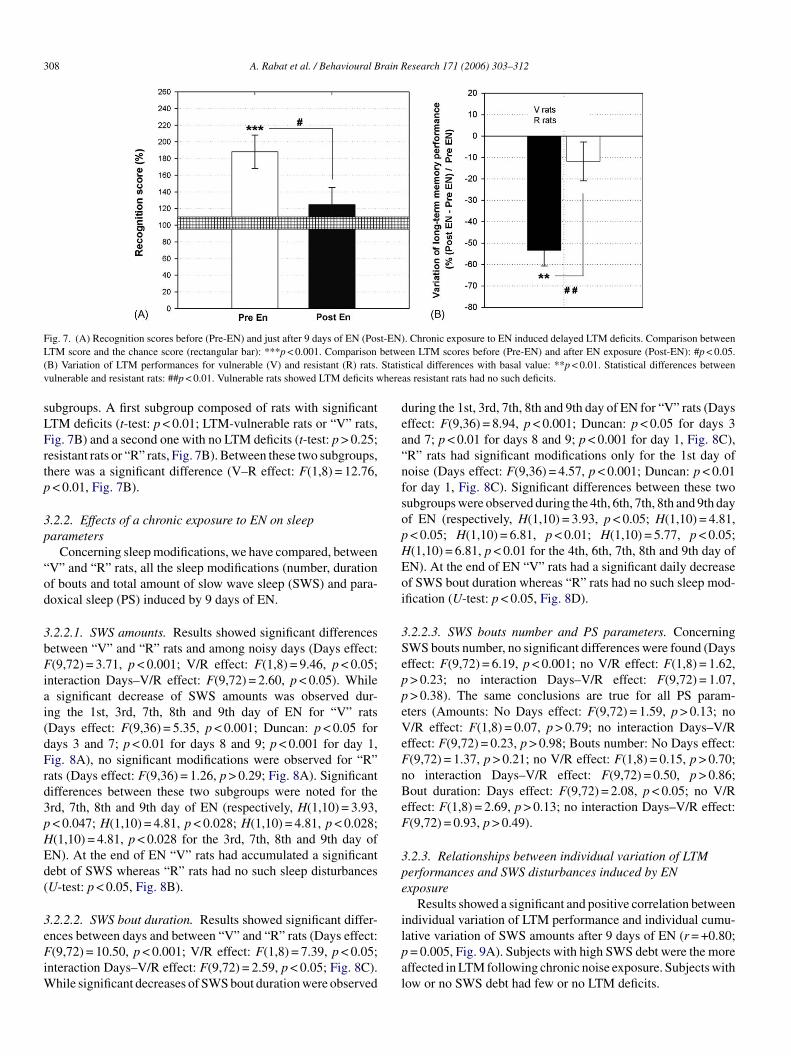
As in experiment 1, rats explored significantly more the novelcompartment than the familiar one (t-test: p < 0.001, Fig. 7A)before a chronic exposure to EN. Following EN exposure, recog-nition score was not significantly different from chance (t-test:p > 0.28, Fig. 7A) but was significantly inferior to the previ-ous one (F(1,9) = 9,85 p < 0.05, Fig. 7A). We thus defined two
F ) rats.a tion bl ere re
ig. 6. (A) Locomotor reactivity to novelty for vulnerable (V) and resistant (Rctive than R rats in the new environment. (B) A significant and negative correlaong-term spatial memory performance (r = −0.59, p = 0.012). The more rats w
Statistical comparison between V and R rats: ###p < 0.001. V rats were moreetween individual locomotor reactivity of rats to novelty and variation of their
active to novelty the more they had LTM deficits after 9 days of EN.
308 A. Rabat et al. / Behavioural Brain Research 171 (2006) 303–312
Fig. 7. (A) Recognition scores before (Pre-EN) and just after 9 days of EN (Post-EN). Chronic exposure to EN induced delayed LTM deficits. Comparison betweenLTM score and the chance score (rectangular bar): ***p < 0.001. Comparison between LTM scores before (Pre-EN) and after EN exposure (Post-EN): #p < 0.05.(B) Variation of LTM performances for vulnerable (V) and resistant (R) rats. Statistical differences with basal value: **p < 0.01. Statistical differences betweenvulnerable and resistant rats: ##p < 0.01. Vulnerable rats showed LTM deficits whereas resistant rats had no such deficits.
subgroups. A first subgroup composed of rats with significantLTM deficits (t-test: p < 0.01; LTM-vulnerable rats or “V” rats,Fig. 7B) and a second one with no LTM deficits (t-test: p > 0.25;resistant rats or “R” rats, Fig. 7B). Between these two subgroups,there was a significant difference (V–R effect: F(1,8) = 12.76,p < 0.01, Fig. 7B).
3.2.2. Effects of a chronic exposure to EN on sleepparameters
Concerning sleep modifications, we have compared, between“V” and “R” rats, all the sleep modifications (number, durationof bouts and total amount of slow wave sleep (SWS) and para-doxical sleep (PS) induced by 9 days of EN.
3.2.2.1. SWS amounts. Results showed significant differencesbetween “V” and “R” rats and among noisy days (Days effect:F(9,72) = 3.71, p < 0.001; V/R effect: F(1,8) = 9.46, p < 0.05;interaction Days–V/R effect: F(9,72) = 2.60, p < 0.05). Whilea significant decrease of SWS amounts was observed dur-ing the 1st, 3rd, 7th, 8th and 9th day of EN for “V” rats(Days effect: F(9,36) = 5.35, p < 0.001; Duncan: p < 0.05 fordays 3 and 7; p < 0.01 for days 8 and 9; p < 0.001 for day 1,Fig. 8A), no significant modifications were observed for “R”rats (Days effect: F(9,36) = 1.26, p > 0.29; Fig. 8A). Significantdifferences between these two subgroups were noted for the3pHEd(
3eFiW
during the 1st, 3rd, 7th, 8th and 9th day of EN for “V” rats (Dayseffect: F(9,36) = 8.94, p < 0.001; Duncan: p < 0.05 for days 3and 7; p < 0.01 for days 8 and 9; p < 0.001 for day 1, Fig. 8C),“R” rats had significant modifications only for the 1st day ofnoise (Days effect: F(9,36) = 4.57, p < 0.001; Duncan: p < 0.01for day 1, Fig. 8C). Significant differences between these twosubgroups were observed during the 4th, 6th, 7th, 8th and 9th dayof EN (respectively, H(1,10) = 3.93, p < 0.05; H(1,10) = 4.81,p < 0.05; H(1,10) = 6.81, p < 0.01; H(1,10) = 5.77, p < 0.05;H(1,10) = 6.81, p < 0.01 for the 4th, 6th, 7th, 8th and 9th day ofEN). At the end of EN “V” rats had a significant daily decreaseof SWS bout duration whereas “R” rats had no such sleep mod-ification (U-test: p < 0.05, Fig. 8D).
3.2.2.3. SWS bouts number and PS parameters. ConcerningSWS bouts number, no significant differences were found (Dayseffect: F(9,72) = 6.19, p < 0.001; no V/R effect: F(1,8) = 1.62,p > 0.23; no interaction Days–V/R effect: F(9,72) = 1.07,p > 0.38). The same conclusions are true for all PS param-eters (Amounts: No Days effect: F(9,72) = 1.59, p > 0.13; noV/R effect: F(1,8) = 0.07, p > 0.79; no interaction Days–V/Reffect: F(9,72) = 0.23, p > 0.98; Bouts number: No Days effect:F(9,72) = 1.37, p > 0.21; no V/R effect: F(1,8) = 0.15, p > 0.70;no interaction Days–V/R effect: F(9,72) = 0.50, p > 0.86;Bout duration: Days effect: F(9,72) = 2.08, p < 0.05; no V/Reffect: F(1,8) = 2.69, p > 0.13; no interaction Days–V/R effect:F
3pe
ilpal
rd, 7th, 8th and 9th day of EN (respectively, H(1,10) = 3.93,< 0.047; H(1,10) = 4.81, p < 0.028; H(1,10) = 4.81, p < 0.028;(1,10) = 4.81, p < 0.028 for the 3rd, 7th, 8th and 9th day ofN). At the end of EN “V” rats had accumulated a significantebt of SWS whereas “R” rats had no such sleep disturbancesU-test: p < 0.05, Fig. 8B).
.2.2.2. SWS bout duration. Results showed significant differ-nces between days and between “V” and “R” rats (Days effect:(9,72) = 10.50, p < 0.001; V/R effect: F(1,8) = 7.39, p < 0.05;
nteraction Days–V/R effect: F(9,72) = 2.59, p < 0.05; Fig. 8C).hile significant decreases of SWS bout duration were observed
(9,72) = 0.93, p > 0.49).
.2.3. Relationships between individual variation of LTMerformances and SWS disturbances induced by ENxposure
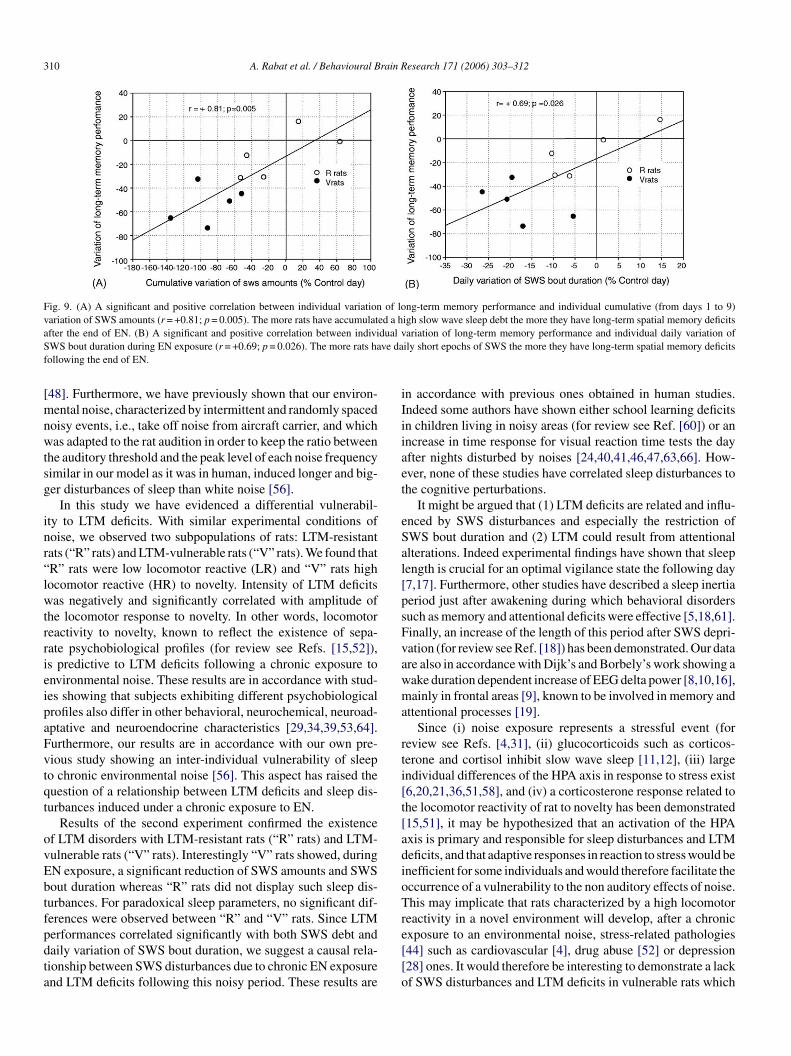
Results showed a significant and positive correlation betweenndividual variation of LTM performance and individual cumu-ative variation of SWS amounts after 9 days of EN (r = +0.80;= 0.005, Fig. 9A). Subjects with high SWS debt were the moreffected in LTM following chronic noise exposure. Subjects withow or no SWS debt had few or no LTM deficits.

A. Rabat et al. / Behavioural Brain Research 171 (2006) 303–312 309
Fig. 8. (A) Variation of slow wave sleep (SWS) amounts, expressed as percentage of control day, during control day and noisy days. While “V” rats have significantdecrease of SWS amounts during several days, “R” rats have no SWS perturbations. Statistical comparison between “V” rats and basal value: *p < 0.05, **p < 0.01,***p < 0.001. Statistical comparison between “V” rats and “R” rats: #p < 0.05. (B) Cumulative (from days 1 to 9) variation of SWS amounts, expressed as percentageof control day, for “V” and “R” rats. Statistical comparison between “V” rats and “R” rats: #p < 0.05. Only “V” rats showed a significant debt of SWS. (C) Variationof SWS bout duration, expressed as percentage of control day, during control day and the 9 days of EN. While “V” rats have significantly shorter epochs of SWSduring several days, “R” rats display shorter SWS epochs for only one day with. Statistical comparison between “V” rats and basal value: **p < 0.01, ***p < 0.001.Statistical comparison between “R” rats and basal value: **p < 0.01. Statistical comparison between “V” and “R” rats: #p < 0.05, ##p < 0.01. (D) Daily variation ofSWS bout duration amounts, expressed as percentage of control day, for “V” and “R” rats. Statistical comparison between “V” rats and “R” rats: #p < 0.05. Only“V” rats showed shorter SWS epochs under EN exposure.
Individual variation of LTM performance was significantlyand positively correlated with individual daily variation ofSWS bout duration during noise exposure (r = +0.69; p = 0.026,Fig. 9B). While subjects with short SWS epochs during noiseexposure showed LTM deficits at the end of noise, subjects withno or few reduction of SWS epochs had no LTM deficits.
No significant correlations have been observed between indi-vidual variation of LTM performance and SWS bouts num-ber (r = −0.30; p > 0.391) or PS parameters (PS amounts:r = −0.386; p > 0.27; PS bouts number: r = −0.40; p > 0.24; PSbout duration: r = −0.16; p > 0.65).
4. Discussion
Using a validated rat model [56], aims of this study were toinvestigate: (1) long-term memory deficits following a chronic
exposure to an environmental noise, and (2) the link betweenmemory deficits and sleep disturbances observed during thenoisy period.
Our results demonstrate that a chronic exposure to an environ-mental noise induces significant delayed LTM deficits after theend of noise exposure. Memory disorders could not result from“physical” tiredness since locomotor activity in the testing boxwas not significantly different before and after noise exposure.These results are partially in accordance with other studies usinganimal model. Indeed and independently of noise level, someauthors found a deleterious effect of noise on memory [3,62]whereas others found an improving effect [42,54]. Though itis, our study is original since in the other studies behavior wastested during noise exposure whereas in our study, behavior wastested after noise exposure. Besides these authors have usedwhite noise, not representative of human noisy environment
310 A. Rabat et al. / Behavioural Brain Research 171 (2006) 303–312
Fig. 9. (A) A significant and positive correlation between individual variation of long-term memory performance and individual cumulative (from days 1 to 9)variation of SWS amounts (r = +0.81; p = 0.005). The more rats have accumulated a high slow wave sleep debt the more they have long-term spatial memory deficitsafter the end of EN. (B) A significant and positive correlation between individual variation of long-term memory performance and individual daily variation ofSWS bout duration during EN exposure (r = +0.69; p = 0.026). The more rats have daily short epochs of SWS the more they have long-term spatial memory deficitsfollowing the end of EN.
[48]. Furthermore, we have previously shown that our environ-mental noise, characterized by intermittent and randomly spacednoisy events, i.e., take off noise from aircraft carrier, and whichwas adapted to the rat audition in order to keep the ratio betweenthe auditory threshold and the peak level of each noise frequencysimilar in our model as it was in human, induced longer and big-ger disturbances of sleep than white noise [56].
In this study we have evidenced a differential vulnerabil-ity to LTM deficits. With similar experimental conditions ofnoise, we observed two subpopulations of rats: LTM-resistantrats (“R” rats) and LTM-vulnerable rats (“V” rats). We found that“R” rats were low locomotor reactive (LR) and “V” rats highlocomotor reactive (HR) to novelty. Intensity of LTM deficitswas negatively and significantly correlated with amplitude ofthe locomotor response to novelty. In other words, locomotorreactivity to novelty, known to reflect the existence of sepa-rate psychobiological profiles (for review see Refs. [15,52]),is predictive to LTM deficits following a chronic exposure toenvironmental noise. These results are in accordance with stud-ies showing that subjects exhibiting different psychobiologicalprofiles also differ in other behavioral, neurochemical, neuroad-aptative and neuroendocrine characteristics [29,34,39,53,64].Furthermore, our results are in accordance with our own pre-vious study showing an inter-individual vulnerability of sleepto chronic environmental noise [56]. This aspect has raised thequestion of a relationship between LTM deficits and sleep dis-t
ovEbtfpdta
in accordance with previous ones obtained in human studies.Indeed some authors have shown either school learning deficitsin children living in noisy areas (for review see Ref. [60]) or anincrease in time response for visual reaction time tests the dayafter nights disturbed by noises [24,40,41,46,47,63,66]. How-ever, none of these studies have correlated sleep disturbances tothe cognitive perturbations.
It might be argued that (1) LTM deficits are related and influ-enced by SWS disturbances and especially the restriction ofSWS bout duration and (2) LTM could result from attentionalalterations. Indeed experimental findings have shown that sleeplength is crucial for an optimal vigilance state the following day[7,17]. Furthermore, other studies have described a sleep inertiaperiod just after awakening during which behavioral disorderssuch as memory and attentional deficits were effective [5,18,61].Finally, an increase of the length of this period after SWS depri-vation (for review see Ref. [18]) has been demonstrated. Our dataare also in accordance with Dijk’s and Borbely’s work showing awake duration dependent increase of EEG delta power [8,10,16],mainly in frontal areas [9], known to be involved in memory andattentional processes [19].
Since (i) noise exposure represents a stressful event (forreview see Refs. [4,31], (ii) glucocorticoids such as corticos-terone and cortisol inhibit slow wave sleep [11,12], (iii) largeindividual differences of the HPA axis in response to stress exist[6,20,21,36,51,58], and (iv) a corticosterone response related tot[adioTre[[o
urbances induced under a chronic exposure to EN.Results of the second experiment confirmed the existence
f LTM disorders with LTM-resistant rats (“R” rats) and LTM-ulnerable rats (“V” rats). Interestingly “V” rats showed, duringN exposure, a significant reduction of SWS amounts and SWSout duration whereas “R” rats did not display such sleep dis-urbances. For paradoxical sleep parameters, no significant dif-erences were observed between “R” and “V” rats. Since LTMerformances correlated significantly with both SWS debt andaily variation of SWS bout duration, we suggest a causal rela-ionship between SWS disturbances due to chronic EN exposurend LTM deficits following this noisy period. These results are
he locomotor reactivity of rat to novelty has been demonstrated15,51], it may be hypothesized that an activation of the HPAxis is primary and responsible for sleep disturbances and LTMeficits, and that adaptive responses in reaction to stress would benefficient for some individuals and would therefore facilitate theccurrence of a vulnerability to the non auditory effects of noise.his may implicate that rats characterized by a high locomotor
eactivity in a novel environment will develop, after a chronicxposure to an environmental noise, stress-related pathologies44] such as cardiovascular [4], drug abuse [52] or depression28] ones. It would therefore be interesting to demonstrate a lackf SWS disturbances and LTM deficits in vulnerable rats which

A. Rabat et al. / Behavioural Brain Research 171 (2006) 303–312 311
have received glucocorticoids antagonists before a chronic expo-sure to EN.
In conclusion this work demonstrates that LTM deficits fol-lowing a chronic exposure to an environmental noise are relatedto SWS disturbances in rats. It suggests that this kind of envi-ronment would have the same effect on human physiology andbehavior. It would be of interest to know whether people whostrongly react to stress will be more sensitive to these non-auditory effects of noise on sleep and behavior than low reactivesubjects. Moreover, this experimental approach provides theopportunity to clarify the neuroanatomical networks and neuro-transmissions involved and influenced by endocrine responsesdue to stress [14,25,51,57].
Acknowledgements
The authors thank M. Kharouby for her technical assistance.This work was supported by grants from INSERM, Universitede Bordeaux II and DGA.
References
[1] Akerstedt T. Do subjective sleep reports describe objective sleep? IntNews Sleep Optimized Manage 2003;1:11–5.
[2] Ambrosini MV, Mariucci G, Colarieti L, Bruschelli G, Carobi C, Giu-ditta A. The structure of sleep is related to the learning ability of rats.
[
[
[
[
[
[
the sensation-seeking trait in man. Neuropsychobiology 1996;34:136–45.
[16] Dijk DJ, Brunner DP, Beersma DG, Borbely AA. Electroencephalogrampower density and slow wave sleep as a function of prior waking andcircadian phase. Sleep 1990;13:430–40.
[17] Dinges DF, Pack F, Williams K, Gillen KA, Powell JW, Ott GE, et al.Cumulative sleepiness, mood disturbance, and psychomotor vigilanceperformance decrements during a week of sleep restricted to 4–5 h pernight. Sleep 1997;20:267–77.
[18] Ferrara M, De Gennaro L. How much sleep do we need? Sleep MedRev 2001;5:155–79.
[19] Fuster JM. Executive frontal functions. Exp Brain Res 2000;133:66–70.[20] Garcia A, Armario A. Individual differences in the recovery of the
hypothalamic-pituitary-adrenal axis after termination of exposure to asevere stressor in outbred male Sprague–Dawley rats. Psychoneuroen-docrinology 2001;26:363–74.
[21] Goldstein MJ. Individual differences in response to stress. Am J CommPsychol 1973;1:113–37.
[22] Griefahn B. Environmental noise and sleep. Review-need for furtherresearch. Appl Acoust 1991;32:255–68.
[23] Griefahn B. Sleep disturbances related to environmental noise. NoiseHealth 2002;4:57–60.
[24] Griefahn BGE. Noise and sleep at home, a field study on primary andafter-effects. J Sound Vib 1986;105:373–83.
[25] Hanley NR, Van de Kar LD. Serotonin and the neuroendocrine regulationof the hypothalamic-pituitary-adrenal axis in health and disease. VitamHorm 2003;66:189–255.
[26] Heffner HE, Heffner RS, Contos C, Ott T. Audiogram of the hoodedNorway rat. Hear Res 1994;73:244–7.
[27] Hockey GR. Effect of loud noise on attentional selectivity. Q J ExpPsychol 1970;22:28–36.
[
[
[
[
[
[
[
[
[
[
[
[
[
[
Eur J Neurosci 1993;5:269–75.[3] Arnsten AF, Goldman-Rakic PS. Noise stress impairs prefrontal corti-
cal cognitive function in monkeys: evidence for a hyperdopaminergicmechanism. Arch Gen Psychiatry 1998;55:362–8.
[4] Babisch W. Stress hormones in the research on cardiovascular effects ofnoise. Noise Health 2003;5:1–11.
[5] Balkin TJ, Badia P. Relationship between sleep inertia and sleepi-ness: cumulative effects of four nights of sleep disruption/restrictionon performance following abrupt nocturnal awakenings. Biol Psychol1988;27:245–58.
[6] Berger M, Bossert S, Krieg JC, Dirlich G, Ettmeier W, Schreiber W,et al. Interindividual differences in the susceptibility of the cortisol sys-tem: an important factor for the degree of hypercortisolism in stresssituations? Biol Psychiatry 1987;22:1327–39.
[7] Bonnet MH, Arand DL. Clinical effects of sleep fragmentation versussleep deprivation. Sleep Med Rev 2003;7:297–310.
[8] Borbely AA, Baumann F, Brandeis D, Strauch I, Lehmann D. Sleepdeprivation: effect on sleep stages and EEG power density in man.Electroencephalogr Clin Neurophysiol 1981;51:483–95.
[9] Cajochen C, Foy R, Dijk DJ. Frontal predominance of a relative increasein sleep delta and theta EEG activity after sleep loss in humans. SleepRes Online 1999;2:65–9.
10] Cajochen C, Wyatt JK, Czeisler CA, Dijk DJ. Separation of circa-dian and wake duration-dependent modulation of EEG activation duringwakefulness. Neuroscience 2002;114:1047–60.
11] Chang FC, Opp MR. Blockade of corticotropin-releasing hormonereceptors reduces spontaneous waking in the rat. Am J Physiol1998;275:R793–802.
12] Chang FC, Opp MR. Role of corticotropin-releasing hormone in stressor-induced alterations of sleep in rat. Am J Physiol Regul Integr CompPhysiol 2002;283:R400–7.
13] Cohen SWN. Nonauditory effects of noise on behavior and health. JSocial Issues 1981;37:36–70.
14] Czyrak A, Mackowiak M, Chocyk A, Fijal K, Wedzony K. Role ofglucocorticoids in the regulation of dopaminergic neurotransmission. PolJ Pharmacol 2003;55:667–74.
15] Dellu F, Piazza PV, Mayo W, Le Moal M, Simon H. Novelty-seekingin rats—biobehavioral characteristics and possible relationship with
28] Holsboer F. The corticosteroid receptor hypothesis of depression. Neu-ropsychopharmacology 2000;23:477–501.
29] Hooks MS, Jones GH, Smith AD, Neill DB, Justice Jr JB. Responseto novelty predicts the locomotor and nucleus accumbens dopamineresponse to cocaine. Synapse 1991;9:121–8.
30] Ising H, Kruppa B. Health effects caused by noise: evidence in theliterature from the past 25 years. Noise Health 2004;6:5–13.
31] Ising HBC. Acute and chronic endocrine effects of noise: review of theresearch conducted at the institute for water, soil and air hygiene. NoiseHealth 2000;7:7–24.
32] Jerison HJ. Effects of noise on human performance. J Appl Psychol1959;43:96–101.
33] Job RF. Noise sensitivity as a factor influencing human reaction to noise.Noise Health 1999;1:57–68.
34] Kabbaj M, Devine DP, Savage VR, Akil H. Neurobiological correlates ofindividual differences in novelty-seeking behavior in the rat: differentialexpression of stress-related molecules. J Neurosci 2000;20:6983–8.
35] Kawada T, Suzuki S. Change in rapid eye movement (REM) sleep inresponse to exposure to all-night noise and transient noise. Arch EnvironHealth 1999;54:336–40.
36] Kirschbaum C, Prussner JC, Stone AA, Federenko I, Gaab J, Lintz D,et al. Persistent high cortisol responses to repeated psychological stressin a subpopulation of healthy men. Psychosom Med 1995;57:468–74.
37] Kuroiwa MXP, Suzuki S, Sasazawa Y, Kawada T, Tamura Y. Habitua-tion of sleep to road traffic noise observed not by polygraphy but byperception. J Sound Vib 2002;250:101–6.
38] Kuwano Smtnsmm. The effect of different kinds of noise on the qualityof sleep under the controlled conditions. J Sound Vib 2002;250:83–90.
39] Lemaire V, Aurousseau C, Le Moal M, Abrous DN. Behavioural traitof reactivity to novelty is related to hippocampal neurogenesis. Eur JNeurosci 1999;11:4006–14.
40] LeVere TE, Bartus RT, Hart FD. Electroencephalographic and behav-ioral effects of nocturnally occurring jet aircraft sounds. Aerosp Med1972;43:384–9.
41] LeVere TEMG, Hart FD. Waking performance decrements followingminimal sleep disruption: the effects of habituation during sleep. PhsyiolPsychol 1975;3:147–54.

312 A. Rabat et al. / Behavioural Brain Research 171 (2006) 303–312
[42] Maes JH, de Groot G. Effects of noise on the performance of rats in anoperant discrimination task. Behav Processes 2003;61:57–68.
[43] Mandile P, Giuditta A, Romano F, Montagnese P, Piscopo S, CotugnoM, et al. Waking EEG power spectra in the rat: correlations with trainingperformance. Brain Res Cogn Brain Res 2003;17:94–105.
[44] McEwen BS. Mood disorders and allostatic load. Biol Psychiatry2003;54:200–7.
[45] Muzet A. The need for a specific noise measurement for populationexposed to aircraft noise during night-time. Noise Health 2002;4:61–4.
[46] Ohrstrom E. Effects of low levels of road traffic noise during the night: alaboratory study on number of events, maximum noise levels and noisesensitivity. J Sound Vib 1995;179:603–15.
[47] Ohrstrom ERR. Sleep disturbance effects of traffic noise. A laboratorystudy on after effects. J Sound Vib 1982;84:87–103.
[48] Ouis D. Exposure to nocturnal road traffic noise: sleep disturbance andits after effects. Noise Health 1999;1:11–36.
[49] Ouis d. Annoyance from road traffic noise: a review. J Environ Psychol2001;21:101–20.
[50] Pallares M, Darnaudery M, Day J, Le Moal M, Mayo W. The neuros-teroid pregnenolone sulfate infused into the nucleus basalis increasesboth acetylcholine release in the frontal cortex or amygdala and spatialmemory. Neuroscience 1998;87:551–8.
[51] Piazza PV, Deminiere JM, Le Moal M, Simon H. Factors that pre-dict individual vulnerability to amphetamine self-administration. Science1989;245:1511–3.
[52] Piazza PV, Le Moal ML. Pathophysiological basis of vulnerability todrug abuse: role of an interaction between stress, glucocorticoids, anddopaminergic neurons. Annu Rev Pharmacol Toxicol 1996;36:359–78.
[53] Piazza PV, Rouge-Pont F, Deminiere JM, Kharoubi M, Le Moal M,Simon H. Dopaminergic activity is reduced in the prefrontal cortexand increased in the nucleus accumbens of rats predisposed to develop
[54] Prior H. Effects of predictable and unpredictable intermittent noise onspatial learning in rats. Behav Brain Res 2002;133:117–24.
[55] Rabat A, Bouyer JJ, Aran JM, Courtiere A, Mayo W, Le Moal M.Deleterious effects of an environmental noise on sleep and contributionof its physical components in a rat model. Brain Res 2004;1009:88–97.
[56] Rabat A, Bouyer JJ, Aran JM, Le Moal M, Mayo W. Chronic exposureto an environmental noise permanently disturbs sleep in rats: inter-individual vulnerability. Brain Res 2005;1059:72–82.
[57] Rosario LA, Abercrombie ED. Individual differences in behavioral reac-tivity: correlation with stress-induced norepinephrine efflux in the hip-pocampus of Sprague–Dawley rats. Brain Res Bull 1999;48:595–602.
[58] Singh A, Petrides JS, Gold PW, Chrousos GP, Deuster PA. Differen-tial hypothalamic-pituitary-adrenal axis reactivity to psychological andphysical stress. J Clin Endocrinol Metab 1999;84:1944–8.
[59] Smith A. A review of the effects of noise on human performance. ScandJ Psychol 1989;30:185–206.
[60] Stansfeld S, Haines M, Brown B. Noise and health in the urban envi-ronment. Rev Environ Health 2000;15:43–82.
[61] Tassi P, Muzet A. Sleep inertia. Sleep Med Rev 2000;4:341–53.[62] Thiel CM, Muller CP, Huston JP, Schwarting RK. Auditory noise can
prevent increased extracellular acetylcholine levels in the hippocampusin response to aversive stimulation. Brain Res 2000;882:112–9.
[63] Vallet MGJMBV, Favre B, Labiale G. Long term sleep disturbance dueto traffic noise. J Sound Vib 1983;90:173–91.
[64] Wesensten NJ, Balkin TJ, Belenky G. Does sleep fragmentation impactrecuperation? A review and reanalysis. J Sleep Res 1999;8:237–45.
[65] Wetherell A. Cognitive and psychomotor performance tests and exper-iment design in multiple chemical sensitivity. Environ Health Perspect1997;105(Suppl 2):495–503.
[66] Wilkinson RT, Campbell KB. Effects of traffic noise on quality of sleep:assessment by EEG, subjective report, or performance the next day. J
amphetamine self-administration. Brain Res 1991;567:169–74.
Acoust Soc Am 1984;75:468–75.



















