
Introduction to Statistical Quality Control - Food QA Project

REVIEW
Chlorophyll a fluorescence: beyond the limits of the QA model
Gert Schansker • Szilvia Z. Toth • Alfred R. Holzwarth •
Gy}oz}o Garab
Received: 27 September 2012 / Accepted: 18 February 2013
� Springer Science+Business Media Dordrecht 2013
Abstract Chlorophyll a fluorescence is a non-invasive
tool widely used in photosynthesis research. According to
the dominant interpretation, based on the model proposed
by Duysens and Sweers (1963, Special Issue of Plant and
Cell Physiology, pp 353–372), the fluorescence changes
reflect primarily changes in the redox state of QA, the
primary quinone electron acceptor of photosystem II
(PSII). While it is clearly successful in monitoring the
photochemical activity of PSII, a number of important
observations cannot be explained within the framework of
this simple model. Alternative interpretations have been
proposed but were not supported satisfactorily by experi-
mental data. In this review we concentrate on the processes
determining the fluorescence rise on a dark-to-light tran-
sition and critically analyze the experimental data and the
existing models. Recent experiments have provided addi-
tional evidence for the involvement of a second process
influencing the fluorescence rise once QA is reduced. These
observations are best explained by a light-induced con-
formational change, the focal point of our review. We also
want to emphasize that—based on the presently available
experimental findings—conclusions on a/ß-centers, PSII
connectivity, and the assignment of FV/FM to the maximum
PSII quantum yield may require critical re-evaluations. At
the same time, it has to be emphasized that for a deeper
understanding of the underlying physical mecha-
nism(s) systematic studies on light-induced changes in the
structure and reaction kinetics of the PSII reaction center
are required.
Keywords Chl a fluorescence �Fluorescence yield changes � Light-induced conformational
changes � OJIP transient � Thermal phase
Abbreviations
Chl Chlorophyll
CP43, CP47 Core antenna proteins of photosystem II
of 43 and 47 kDa, respectively
DCMU 3-(30,40-Dichlorophenyl)-1,1-
dimethylurea
ETC Electron transport chain
FCCP Carbonylcyanide-p-
trifluoromethoxyphenylhydrazone
F0 Minimum chl a fluorescence yield in the
dark-adapted state
FM Maximum chl a fluorescence yield in the
dark-adapted state
FV Variable fluorescence defined as FM-F0
LED Light emitting diode
P680 Reaction center pigments of photosystem II
PFD Photon flux density
Pheo Pheophytin
PMS Phenazine methosulfate
PQ Plastoquinone
PSII and PSI Photosystem II and I, respectively
QA and QB Primary and secondary quinone electron
acceptors of photosystem II, respectively
RC Reaction center
S-states S0, S1,
S2, S3, and S4
Different redox states of the oxygen-
evolving complex
G. Schansker (&) � S. Z. Toth � G. Garab (&)
Institute of Plant Biology, Biological Research Center Szeged,
Hungarian Academy of Sciences, Szeged 6701, Hungary
e-mail: [email protected]
G. Garab
e-mail: [email protected]
A. R. Holzwarth
Max-Planck-Institute for Chemical Energy Conversion, 45470
Mulheim an der Ruhr, Germany
123
Photosynth Res
DOI 10.1007/s11120-013-9806-5
SSTF Saturating single turnover flash
TMPD N,N,N’,N’-tetramethyl-
p-phenylenediamine dihydrochloride
TRIS Tris(hydroxymethyl)aminomethane
TyrZ, TyrD Tyrosine donors to P680 localized on the
PSII reaction center D1 and D2 proteins,
respectively
Introduction
Measurements of chlorophyll a fluorescence transients are
used as a non-invasive tool to probe the activity of the
photosynthetic apparatus. It is undoubtedly the most widely
applied technique in photosynthesis research (reviewed
e.g., in Papageorgiou and Govindjee 2004). Upon exposing
a dark-adapted leaf or algal cells to a strong saturating light
pulse (3,000–12,000 lmol photons m-2 s-1, 200–1,000 ms),
the fluorescence increases from a minimum level F0 to the
maximum level FM, which is reached in about 200 ms. If
presented at a high time resolution a polyphasic OJIP
transient is observed (e.g., Strasser et al. 2004; see Fig. 1).
On a logarithmic time scale (Strasser and Govindjee 1991)
three rise phases can be identified: OJ (0–3 ms), JI
(3–30 ms), and IP (30–200 ms) (Neubauer and Schreiber
1987; Strasser and Govindjee 1991). When using the phase
amplitude modulation (PAM) technique, the fluorescence
induced by a saturating pulse is generally presented as a
spike (e.g., Schreiber 2004).
According to the most widely accepted interpretation,
hereafter called the QA model, which was proposed in 1963
by Duysens and Sweers and reviewed recently by Stirbet
and Govindjee (2012), the fluorescence rise from F0 to FM
is a reflection of the reduction of QA. This forms the basis
of nearly all theoretical treatments of fluorescence mea-
surements (Butler and Kitajima 1975; Paillotin 1976;
Strasser 1978; Trissl and Lavergne 1994; Kramer et al.
2004) including the quenching analysis (Schreiber 2004)
and the JIP test, a mathematical framework for the analysis
of OJIP transients (Strasser and Strasser 1995; Strasser
et al. 2004) and is the main explanation model in most
reviews (e.g., Maxwell and Johnson 2000; Baker 2008).
The QA model has been clearly successful in monitoring
the photochemical activity of photosynthetic samples.
However, it has also been recognized that a number of
important observations, related to the time and temperature
dependencies of the fluorescence rise, cannot be explained
within the framework of this simple model (e.g., Samson
et al. 1999; Schansker et al. 2011).
The QA model is, implicitly or explicitly, based on two
assumptions: (1) the fluorescence rise between F0 and FM is
a reflection of the progressive reduction of QA, i.e., the
transition from open to closed PSII reaction centers, and
(2) no changes in the fluorescence yield of open or closed
centers during the transition between the two states F0 and
FM occur. Assumption 2 means that the rate constants for
fluorescence emission and heat dissipation of neither open
nor closed centers change during the transition from F0 to
FM. With the introduction of the quenching analysis, a
method to separate photochemical (electron transport
related) and non-photochemical (fluorescence yield change
related) quenching of the fluorescence intensity, this
assumption was formulated more explicitly (van Kooten
and Snel 1990).
The vast literature related to fluorescence changes on a
dark-to-light transition has recently been thoroughly
reviewed by Stirbet and Govindjee (2012). According to
that review, the QA model, i.e., the model of Duysens and
Sweers (1963) is ‘‘the most credible theory.’’ The authors
suggest that the whole rise kinetics, in particular including
the IP phase, originates essentially from a gradual photo-
reduction of QA only. At the same time, however, Stirbet
and Govindjee (2012) also point out that the complex
fluorescence rise kinetics can only be understood by
modifying the Duysens and Sweers theory. They state that
during the fluorescence rise ‘‘… the presence of several
quenchers and modifiers that influence the fluorescence
yield during the O–J–I–P transient …’’ are also to be taken
into account. This actually implies that neither of the two
basic assumptions of the Duysens and Sweers model is
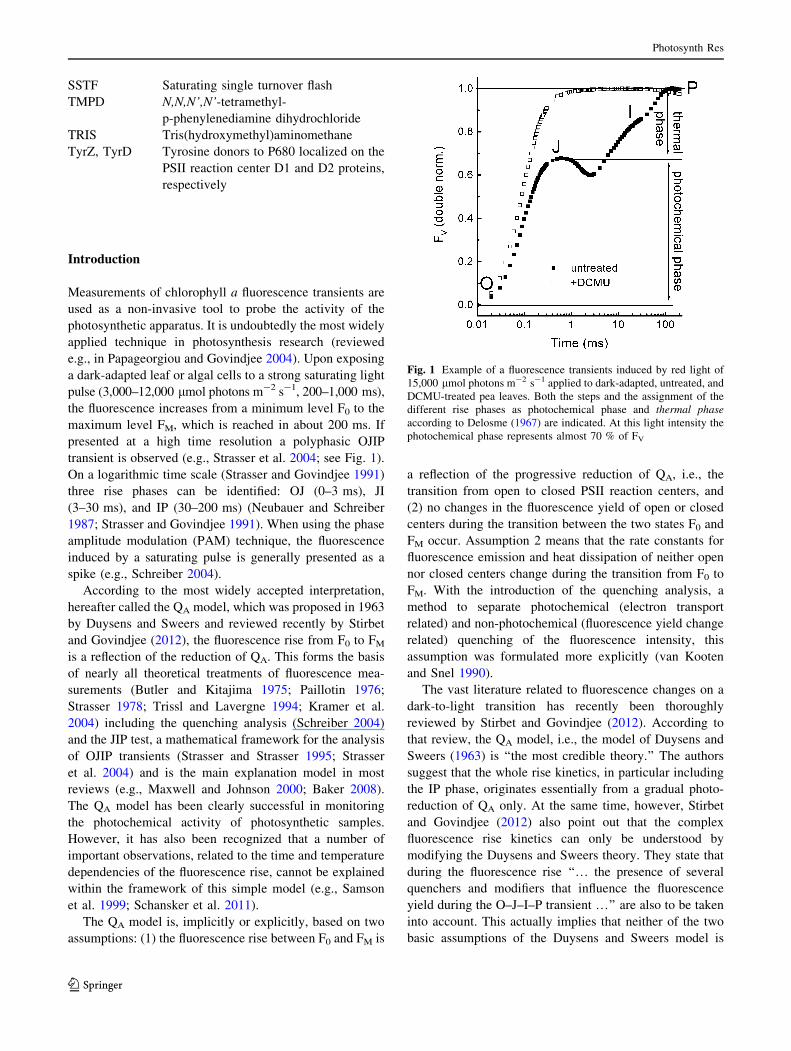
Fig. 1 Example of a fluorescence transients induced by red light of
15,000 lmol photons m-2 s-1 applied to dark-adapted, untreated, and
DCMU-treated pea leaves. Both the steps and the assignment of the
different rise phases as photochemical phase and thermal phaseaccording to Delosme (1967) are indicated. At this light intensity the
photochemical phase represents almost 70 % of FV
Photosynth Res
123

accepted. The implied ‘‘modifiers’’ in some cases form the
basis for alternative models presented in the literature;
some of these alternative models will be discussed below.
Most ‘‘modifiers,’’ e.g., P680?, TyrZ?, and TyrD, carot-
enoid triplets, or the potentially functioning alternative
reaction pathways, such as cyclic PSII and PSI electron
transfers, do affect (or may affect) part of the initial kinetic
phases of the fluorescence rise. However, none of them
offer an explanation for the crucial slow fluorescence rise
phase, the IP phase.
Experimental observations which are difficult
to reconcile with the QA model
A saturating single turnover flash (SSTF) induces at best
60–65 % of FM
The fluorescence intensity induced by a single SSTF in
intact spinach chloroplasts does not surpass the fluores-
cence intensity of the J step (Schreiber 1986) which is
approx. 60–65 % of FM. On the other hand, it has been
shown that an SSTF can reduce all or nearly all QA at room
temperature, as shown by the fact that it induces a nearly
maximum DA320nm signal (Joliot and Joliot 1981), which is
a direct measure for reduced QA, and a nearly maximum S2
multiline signal, arising from the manganese cluster of the
water splitting complex (Brudvig et al. 1983). Also, the
analysis of period-4 oscillations in the flash-oxygen yield
indicates, based on the miss factor, that the fraction of
unreduced QA on each single turnover flash is less than
7–8 % (Shinkarev (2005) re-analyzing the data of Forbush
et al. (1971)).
FM is reached only after many excitations and multiple
PSII turnovers
A photon flux density (PFD) of 3,000 lmol photons
m-2 s-1 white or red light means that each PSII becomes
excited once every *200 ls (cf. Neubauer and Schreiber
1987; Lazar and Pospısil 1999). This equates *1,000
excitations per 200 ms (the time needed to reach FM). It
has also been shown that to reach FM, the photosynthetic
electron transport chain (ETC) has to become reduced
(Schansker et al. 2005, 2006, 2008); in particular, it has
been shown by Joliot and Joliot (2002) that in leaves, under
their experimental conditions, about 20 electrons had to be
transported, i.e., 20 stable charge separations had to take
place, to reach FM. In addition, it has been demonstrated
that electron flow through PSI (Kautsky et al. 1960;
Munday and Govindjee 1969; Schreiber et al. 1989;
Harbinson and Hedley 1993; Schansker et al. 2005) in
combination with inactive FNR (Kautsky et al. 1960;
Munday and Govindjee 1969; Satoh 1981; Carillo et al.
1981; Schansker et al. 2005, 2006, 2008) affects the fluo-
rescence rise in leaves and intact chloroplasts.
J step (2–3 ms) also requires multiple charge separations
It follows (see point 2) that during the time needed to reach
the J step C10 excitations of each PSII occur. This multiple
excitation also induces multiple charge separations, which
is demonstrated by at least two observations: (i) In heat-
treated leaves as well as in TRIS-washed thylakoids, where
only one electron is available for donation to P680? in this
time range (Toth et al. 2007a), a fluorescence peak
(K) appears after *300 ls of illumination, which is fol-
lowed by a fluorescence decline due to forward electron
transport to QB. (ii) Experiments with SSTFs indicate that a
pair of electrons needs about 2 ms to reach the PQ-pool
(Petrouleas and Crofts 2005).
Kinetics in the presence of DCMU
In isolated thylakoid membranes in the presence of DCMU
(3-(3,4-dichlorophenyl)-1,1-dimethylurea), which blocks
the electron transport between QA and QB, an SSTF
induces about 70–75 % of FM (Joliot and Joliot 1979;
Samson and Bruce 1996). Joliot and Joliot (1979) observed
for spinach thylakoid membranes that while DA550, indi-
cating the local field due to the reduction of QA, reached
almost its highest value already after one saturating flash in
the presence of DCMU; the fluorescence yield was only
about 70–75 % of the maximum variable fluorescence.
Further flashes, usually 3–5, are needed to reach the
maximum fluorescence intensity. In leaves in continuous
strong light FM is reached after about 10 ms (Toth et al.
2005a), i.e., also only after multiple excitations.
The sigmoidicity of the fluorescence rise in the presence
of DCMU
Traditionally, the sigmoidicity of the fluorescence rise in the
presence DCMU is explained in terms of the connectivity
between PSII antennae (Joliot and Joliot 1964; Paillotin
1976; Strasser 1978). However, according to Vredenberg
(2004, 2008) this feature is difficult to explain within a QA
model. The kinetic analysis of Vredenberg (2008) suggests
that the sigmoidicity is related to the double excitation nature
of the fluorescence rise in the presence of DCMU.
Alternative interpretations assuming
that the fluorescence yield is determined by both QA
and (an) additional process(es)
Morin (1964) and Delosme (1967)—using a special setup,
with a shutter opening time of 30 and 5 ls, respectively,
Photosynth Res
123

which also allowed the use of high light intensities—were
the first to observe an intermediate step in the fluorescence
rise (without inhibitors). Their recorded fluorescence rise
had, therefore, more complex kinetics than those published
by Duysens and Sweers (1963). It should be noted here that
such measurements could only be done routinely 25 years
later with the introduction of fast LED-based systems.
Delosme (1967) proposed that only the first 2 ms of the
fluorescence rise was due to the reduction of QA. The
nature of the remaining part, called the ‘‘thermal phase’’
due to its temperature dependence, was not identified.
Since then different models and interpretations have been
proposed. They all agree on the crucial involvement of QA-
and at least one further component but differ from each
other concerning the nature of this additional process. They
also differ from each other whether or not they accept one
or the other of the two basic assumptions of the QA model.
(It is to be noted here that the term ‘‘thermal phase’’ as
introduced by Delosme in 1967 is somewhat misleading. It
has been thoroughly documented that this ‘‘thermal
phase,’’ i.e., the slow rise of the fluorescence induction is
also a light-induced reaction. We adhere to this terminol-
ogy throughout this review, but in order to avoid any
misunderstanding we italicize it).
Redox reactions
Most of the authors who studied the thermal phase, starting
with Delosme (1967), proposed the involvement of a light-
induced redox reaction. In these models, Assumption 2, but
not 1, was considered valid. Joliot and Joliot (1979, 1981)
proposed a redox factor called Q2, involved in a non-
electrogenic reaction; the identity of which, however,
remained elusive. Recently, Boussac et al. (2011) found
that in Thermosynechococcus elongatus QB– could extract
an electron from the non-heme iron leading to oxidized
non-heme iron in a fraction of PsbA3-PSII (a stress
inducible cyanobacterial gene product) but not in PsbA1-
PSII. They proposed that Q2 might be identical with the
non-heme iron. This interpretation, however, has not been
confirmed by studies monitoring the redox state of the non-
heme iron and of the chl a fluorescence in green leaves.
Vredenberg (2000) suggested that the redox component
was pheophytin (Pheo), and that both QA- and Pheo- are
required for the full closure of the PSII reaction center
(RC) and for reaching FM. However, accumulation of
Pheo- was not demonstrated (for extensive literature on
Pheo- and chl a fluorescence quenching see Stirbet and
Govindjee 2012). Schreiber (2002) proposed a reduction of
the inactive branch of PSII but, again, experimental evi-
dence was not presented. We also stress that low-temperature
fluorescence induction data suggest that the process associated
with the thermal phase cannot be a redox factor on the
acceptor side of PSII (Schansker et al. 2011).
Delosme (1971) noted that the process reflected by the
thermal phase must be closely linked to the reduction of QB
and the PQ-pool. Later it was suggested that the occupancy
state of the QB-site (the redox state and/or the presence of a
PQ molecule bound to the QB-site) determines the thermal
phase (Vasil’ev and Bruce 1998; Samson et al. 1999;
Yaakoubd et al. 2002). Amesz and Fork (1967) showed that
oxidized artificial quinones can quench fluorescence and
subsequently it was shown that oxidized PQ is also a fluo-
rescence quencher in isolated thylakoid membranes (Joliot
1974; Vernotte et al. 1979) and especially in PSII membranes
(Kurreck et al. 2000). However, in intact leaves PQ-pool
quenching does not occur (Toth et al. 2005a). Further, the
observation that FM can only be reached following the
reduction of the ETC (Schansker et al. 2005, 2006, 2008)
could in principle be explained by assuming that the thermal
phase is determined by the QB-site occupancy state (Vasil’ev
and Bruce 1998; Samson et al. 1999; Yaakoubd et al. 2002),
since it depends on the PQ-pool redox state, which will, to
some extent, be sensitive to the electron flow through PSI.
However, there are three problems with the QB-site occu-
pancy hypothesis: (i) there is no clear link between the
QB-site occupancy state and processes that could lead to an
increase in the fluorescence yield; (ii) it cannot explain
measurements made on DCMU-inhibited samples, where
the QB-site occupancy state does not play a role; and (iii) the
thermal phase can relax and be re-induced without changes
in the QB-site occupancy state (Schansker et al. 2011).
a/ß-centers
The concept of PSII a/ß heterogeneity was introduced to
explain the biphasicity of the fluorescence rise in the
presence of DCMU (Melis and Homann 1975, 1976).
According to this model, which is based on the validity of
both Assumptions 1 and 2, but operates with two QA
molecules in two different types of RCs, the two rise
phases originate from two different populations of PSII
with different antenna sizes. The sigmoidal fluorescence
rise was proposed to arise from a-centers located in the
grana; they were thought to possess large light-harvesting
antennae interconnected via energy migration. In contrast,
the ß-centers, which were proposed to be located in the
stroma lamellae and to exhibit an exponential fluorescence
rise, explained by their small antenna sizes exhibiting no
connectivity (Melis and Homann 1975). In later work
Melis (1985) proposed overlap between ß-centers and
QB-non-reducing centers (Lavergne 1982). However, the
a/ß-center model does not provide a satisfactory explanation
for the complexity of the fluorescence rise under different
Photosynth Res
123

conditions: (i) the model does not explain the observation
that the slow fluorescence rise in the presence of DCMU
exhibits a strong temperature dependence (Schansker et al.
2011)—which cannot be explained either by antenna or RC
heterogeneity; (ii) the model cannot account for the
observations of France et al. (1992) that biphasicity arises
only after double (or multiple) excitations with (a) short
dark interval(s) between the excitations. This does not
mean that there is no (antenna size) heterogeneity of
PSII—for which there exists extensive biochemical evi-
dence (e.g., Danielsson et al. 2006)—it only suggests that
antenna size heterogeneity of PSII does not necessarily
induce a distinguishable kinetic phase in the fluorescence
rise. Instead, antenna heterogeneities may exert diffuse
effects on the OJ rise.
Vredenberg (2004) noted that the interpretation of the
a/ß-concept is model dependent, having a different meaning
within his three-state model. As shown by Vredenberg
(2008), the sigmoidicity of the fast fluorescence rise phase
in the presence of DCMU—an important feature of the a/ß-
concept—can mathematically be described by two kineti-
cally overlapping exponential functions if the two rate
constants differ by less than one order of magnitude.
Hence, the wealth of experimental observations obtained
and interpreted within the framework of a/ß heterogeneity
require a critical re-evaluation.
The transmembrane uniform electric field
The role of the light-induced transmembrane uniform
electric field, induced by a primary charge separation and
consecutive ‘‘delocalization’’ of the field (Junge 1977), as a
co-determinant of the fluorescence rise kinetics has been
considered in several studies (Pospısil and Dau 2002;
Bulychev and Vredenberg 2001; Vredenberg and Bulychev
2003; Rubin and Riznichenko 2009). Electrochemical
effects on the fluorescence yield have been shown under
special conditions like a salt jump or redox titrations (e.g.,
Dau and Sauer 1991; Krieger and Rutherford 1997).
According to this interpretation, Assumption 2 is not valid.
Especially in the model of Vredenberg the electric field is
an important driving force for the fluorescence rise,
determining the IP phase. Vredenberg (2000) showed that
following a pre-flash the F0 of the OJIP transient measured
shortly afterward was increased by 18 % and showed a
relaxation time in darkness of 105 s. Vredenberg linked
this to changes in the lateral resistance of the chloroplast
compartments showing similar relaxation kinetics. How-
ever, as shown by Schansker and Strasser (2005), QB-
induced by a 10-s FR light pulse also causes an increase of
F0 and relaxation time of 100 s. This effect—also induced
by an STF—is due to the equilibrium between QA and QB-,
where the electron at pH 7 spends 5 % of its time on QA
and 95 % of its time on QB (Diner 1977) and the equilib-
rium shifts further to QA as the stromal pH increases
(Robinson and Crofts 1984). Bulychev and Vredenberg
(2001) worked with samples with a destroyed PSII donor
side. They used N,N,N’,N’-tetramethyl-phenylenediamine
dihydrochloride (TMPD) and phenazine methosulfate
(PMS) as electron donors, assuming that these compounds
only donated electrons to PSI and showed that under these
conditions TMPD and PMS stimulate the fluorescence rise.
The authors interpreted this as an influence of the PSI-
generated electrical field on PSII fluorescence. However,
both ascorbate (midpoint potential of 58 mV (Loach
1976)) and TMPD (midpoint potential of 240 mV (Prince
et al. 1981)) can donate electrons to the donor side of Mn-
depleted PSII (see Mano et al. (2004) and Toth et al. (2009)
for ascorbate and Tamura et al. (1990) for TMPD), making
it likely that PMS with a midpoint potential of 85 mV
(Prince et al. 1981) can do this as well. In addition, PMS is
also a fluorescence quencher (Wientjes and Croce 2012).
Vredenberg et al. (2006) observed that a train of 40 STFs
leads to a gradual increase of both the initial and maximum
flash-induced fluorescence intensity. In the presence of
FCCP (carbonylcyanide-p-trifluoromethoxy-phenylhy-
drazone) this increase is no longer observed. The authors
assumed that FCCP acting as a protonophoric uncoupler
abolishes the alkalinization of the stroma and that, there-
fore, the observed flash-induced fluorescence increase was
related to the alkalinization of the stroma and an accom-
panying shift of the equilibrium between QA and QB-
toward QA (Diner 1977; Robinson and Crofts 1984).
However, FCCP is also an ADRY reagent (McCauley et al.
1987) and as shown by de Wijn and van Gorkom (2001),
under flash conditions FCCP mediates electron transport
between the PQ-pool and the S2 and S3 states of the Mn-
cluster of PSII—possibly via cyt b559, which has been
shown to undergo autooxidation and fast, flash-induced
reduction in the presence of ADRY reagents (Barabas et al.
1993)—keeping the PQ-pool oxidized. This interpretation
is also much more in line with the work of Schansker et al.
(2005) and Toth et al. (2007b) on the relationship between
the PQ-pool redox state and the kinetic properties of OJIP
transients. The inactive centers that Vredenberg et al.
(2006) describe in their paper are in this context substrate
(PQ)-limited PSII reaction centers. Vredenberg and Bulychev
(2003) observed that valinomycin (VMC) infiltrated in
leaves suppresses the IP phase of the fluorescence rise.
VMC can dissipate the electric field and the authors
interpreted this result to indicate that the electric field
drives the IP phase. This interpretation is, however, in
disagreement with the measurements of Schreiber and
Neubauer (1990), who observed that the electric field
decreases during the IP phase. In addition, the effect of the
oxygen concentration on the fluorescence decline beyond
Photosynth Res
123

the P-step in the presence of VMC observed by Sokolove
and Marsho (1979) can also be interpreted as a secondary
effect of VMC on the acceptor side of PSI. This is con-
firmed by the observation that the suppression of the IP
phase in the presence of VMC is accompanied by an
inhibition of the 820-nm transmission increase normally
observed during the IP phase (G Schansker and SZ Toth,
unpublished data). That VMC may have multiple effects on
the ETC is also suggested by the observation that it has an
effect on the JI rise in PSII and thylakoid membranes
(Pospısil and Dau 2002). Vredenberg (2011) and Vreden-
berg et al. (2012) proposed that CET is the electric field
creating driving force during the IP phase. However, there
is consensus in the literature that under high light condi-
tions, under which OJIP transients are measured, the
PQ-pool is largely reduced at the I step, defined as the
fluorescence intensity reached after 20–30 ms of illumi-
nation by e.g., 3,000 lmol photons m-2 s-1 (Lazar 2003,
Zhu et al. 2005, Toth et al. 2007b), and as a consequence
the redox poise necessary for CET (reviewed by Heber
2002) is missing. In addition, it is in that case difficult to
explain why the FM in the presence of DCMU, which has
the same value as in the absence of DCMU (Toth et al.
2005a), can be reached within 10 ms.
In summary, the transmembrane uniform electric field
may very well have a small effect on the fluorescence rise;
however, unambiguous experimental data supporting such
a role, for the moment, are missing. We also want to point
out that the local field within PSII is two-to-three orders of
magnitude more intense than the uniform transmembrane
electric field (Zimanyi and Garab 1989); local fields may
play a significant role in the fluorescence rise, an effect
which will be discussed below.
Conformational changes
A fluorescence yield change induced by a light-driven
conformational change within the RC is another possible
determinant of the fluorescence rise; in that case both
Assumptions 1 and 2 of the QA model must be considered
to be invalid. Moise and Moya (2004a, b) were the first to
consider conformational changes as an explanation for the
thermal phase in detail. Using multi-frequency phase and
modulation fluorometry, they determined the fluorescence
lifetimes and fluorescence yields along the fluorescence
rise (induced by a light intensity of 70 lmol photons
m-2 s-1). A non-linear relationship between lifetime and
yield was observed, which was proposed to reflect a con-
formational change within PSII, in particular, in CP47
(Moise and Moya 2004a, b). These conformational changes
were observed both in the physiological temperature range
and at cryogenic temperatures, and both in the absence and
presence of DCMU, but did no longer occur below
-50 �C. Based on a study of the temperature dependences
and induction/relaxation kinetics of fluorescence transients
in a wide temperature interval Schansker et al. (2011) came
to the same conclusion as Moise and Moya (2004a, b): the
process associated with the thermal phase is related to a
conformational change. However, these authors were still
able to detect dark relaxation of the fluorescence intensity
attributable to conformational changes at temperatures as
low as -60 �C. The difference between the two sets of
low-temperature data, criticized by Stirbet and Govindjee
(2012), can readily be explained by the different light
intensities used in the two studies and the fact that charge
stabilization (Brudvig et al. 1983; Schlodder 2008) and
probably also the conformational changes (e.g., via protein
flexibility) become hindered at low temperatures. Schansker
et al. (2011) used not only much stronger light, 3,500 lmol
red photons m-2 s-1, but also gradually increased the pulse
length when decreasing the temperature, from 80 ms at
-20 �C to 300 ms at -60 �C. Further, Schansker et al.
(2011) showed that at cryogenic temperatures the two
fluorescence rise components exhibited substantial activa-
tion energies, and only one process occurred in a temper-
ature independent, nearly activationless manner (as
expected for the primary charge separation within PSII (cf.
Dekker and van Grondelle 2000)). Also, the fast relaxation
of the rise at physiological temperatures could not be
attributed to known redox changes. These observations are
consistent with the involvement of conformational changes
in the fluorescence rise as proposed by Moise and Moya
(2004a, b).
Nature, mechanism, and significance
of the conformational changes
In this section we provide an overview on additional
experimental evidences demonstrating the occurrence of
conformational changes and we will discuss their signifi-
cance as far as the interpretation of the fluorescence rise
parameters is concerned. First, however, we have to
address the problem related to the thermal phase in the
presence of DCMU—mainly because its occurrence is
doubted by some researchers. We also comment on pub-
lished model calculations, which show that the pure QA
model—despite many efforts—cannot offer an appropriate
explanation for the slow fluorescence rise kinetics.
The occurrence of the thermal phase in the presence
of DCMU
When considering the role of conformational changes on
the fluorescence yield of closed centers, we implicitly
assume that the thermal phase is also expressed in the
Photosynth Res
123

presence of PSII inhibitors such as DCMU, and the phys-
ical mechanism(s) underlying the quantitative fluorescence
rise are the same both in the presence or absence of
DCMU, and that the difference is confined to the kinetics,
due to the absence of electron acceptors beyond QA. We
must note, however, that this view is not shared by all
researchers. Stirbet and Govindjee (2012), considering the
arguments in favor or disfavor of either side, suggest that
the thermal phase is absent in the presence of DCMU. In
their view, the ‘‘theories supporting [the idea of its pres-
ence] do not have sufficient experimental support.’’
Concerning the occurrence of thermal phase in the
presence of DCMU, first, we would like to point out that, as
discussed above, the widely accepted model on the fluo-
rescence rise in the presence of DCMU, i.e., explaining the
fluorescence rise in terms of a/ß heterogeneity, is difficult
to reconcile with the very different temperature depen-
dences of the two fluorescence rise phases in the presence
of DCMU. We would also like to stress that, to our
knowledge, there is no experimental evidence supporting
the (in our view much stronger) assumption that the basic
processes and effects related to the fluorescence induction
within PSII in the presence of DCMU are principally dif-
ferent from those in its absence. In fact, the experimental
observation that neither the F0 nor the FM intensities
depend on the presence of DCMU in intact systems (Toth
et al. 2005a) argues against such differences due to the
presence of DCMU in the same way as it argues against
PQ-pool quenching of PSII fluorescence in intact systems.
Also, at low temperature (below -20 �C) fluorescence
properties appear to be identical in the presence and
absence of DCMU. Minor differences, due to differences
e.g., in the recombination kinetics, might be induced by the
shift in the redox potential of QA in the presence of DCMU
(Krieger-Liszkay and Rutherford 1998). Also, the induc-
tion of a further oxidation of non-heme iron upon the
addition of DCMU, in a fraction of PsbA3 PSII, but not of
PsbA1 PSII, as shown by Boussac et al. (2011), to some
extent might pose the possibility of differences between the
nature of fluorescence inductions in untreated and DCMU-
treated samples. On the other hand, Samson and Bruce
(1996) studying the effect of a pre-oxidized non-heme iron
in the presence of DCMU on the flash-induced fluorescence
intensity concluded that the non-heme iron did not signif-
icantly contribute to the low fluorescence yield induced by
an SSTF. It thus seems more likely that a partial oxidation
of the non-heme iron, if it occurs in higher plant leaves
under physiological conditions, will affect only the shape
of the fluorescence rise to some extent but not the final
fluorescence level(s).
Hence, we believe that there exists no convincing evi-
dence that DCMU suppresses the thermal phase. There is,
however, an apparent discrepancy, which requires an
explanation: an SSTF induces a larger FV in the presence of
DCMU than in its absence, *85 versus 65 % in leaves
(e.g., Schansker et al. 2011), and 75 versus 50 % in isolated
thylakoid membranes (Joliot and Joliot 1979; Samson and
Bruce 1996; Vredenberg 2000). (This difference between
leaves and isolated thylakoid membranes can be explained
by the occurrence of PQ-pool quenching in isolated thy-
lakoid membranes (cf. Samson and Bruce 1996) but not in
leaves (Toth et al. 2005a) (see above). Alternatively, a
structural re-organization of the entire thylakoid membrane
and/or changes in the extent of stacking of the thylakoid
membranes during isolation could as well have an effect on
the ratio between the photochemical and thermal phase).
This apparent discrepancy between the data in the presence
and absence of DCMU can be resolved, however, by
analyzing the dark relaxation kinetics instead of the
induction kinetics (Schansker et al. 2011). In that case
*30 % of FV is associated with the thermal/slow rise
phase—both in the presence and absence of DCMU.
Schansker et al. (2011) argued that an explanation for the
observation that only *15 % of FV is observed for the
slow rise phase in the presence of DCMU is a kinetic
overlap between the photochemical phase and the thermal
phase.
Taken together, the above-mentioned data and argu-
ments strongly suggest that the thermal phase is present
both in the absence and presence of DCMU, and thus there
is no need to assume that the physical background of the
phenomena is substantially different in the presence and
absence of DCMU.
Simulations
Stirbet and Govindjee (2012) in their argumentation
emphasized the role of simulations as tools to characterize/
explain the OJIP transient but did not discuss the literature
in detail. Researchers simulating OJIP transients based on
the QA model are faced with simulated fluorescence tran-
sients that rise much faster than the experimentally
observed transients. Processes introduced in the models to
slow the fluorescence rise down mentioned by Lazar and
Schansker (2009) are an electron acceptor X that accepts
electrons from QB at a high rate, PQ-pool quenching, slow
and fast reducing PQ-pools, and extremely slow PSII RCs.
The two models that consist of the most comprehensive
description of the ETC are the models of Lazar (2003,
2009) and Zhu et al. (2005). These are actually two com-
ponent models in the sense that QA as well as the oxidized
PQ-pool are considered to be fluorescence quenchers. In
both cases the maximum fluorescence level is simulated
after *20 ms of illumination. This indicates clearly that
OJIP transients measured at high light intensities PQ-pool
quenching, if it did exist, would not provide an explanation
Photosynth Res
123

for a theoretical OJIP transient reproducing the correct
experimental time course. As demonstrated by Lazar
(2009) the problem is not so much the kinetic model, since
the PSI reactions can be modeled with a correct time
dependence, but the fact that this does not translate into the
right fluorescence amplitudes of the fluorescence rise
phases. In the model of Vredenberg, which is kinetically
simpler, the kinetic phase assigned by Lazar (2003, 2009)
and Zhu et al. (2005) to PQ-pool quenching is attributed to
the release of Pheo quenching (Vredenberg 2000) [or to
active RCs that turn during illumination into inactive RCs
that can then release Pheo quenching (Vredenberg et al.
2006)]. On the basis of QA and Pheo only the OJI rise can
be simulated, whereas the IP phase is simulated assuming
an effect of the electric field on the fluorescence yield
(Vredenberg and Bulychev 2002, 2003; Vredenberg et al.
2007).
The fact that on the basis of realistic, experimentally
proven parameters (see I.2.4 for a discussion of the electric
field), no satisfactory simulation has been obtained so far,
suggests that (i) either of the basic assumptions are not
valid and/or (ii) additional factors must be taken into
consideration. A simulation of the OJIP transient using a
conformational change model as proposed by Schansker
et al. (2011) has not been tried yet; it definitely will require
a deeper understanding of the nature of the changes and the
underlying physical mechanism(s).
The nature and mechanism of the conformational
changes
The temperature dependence of the fast and slow rise
phases in the presence of DCMU yields important infor-
mation on the character of these phases. Schansker et al.
(2011) showed that the fast rise phase was nearly temper-
ature independent, whereas the slow rise phase had a strong
temperature dependence, strongly suggesting that the fast
and the slow rise phases represent two fundamentally dif-
ferent processes. As a consequence, the kinetic overlap
observed at room temperature disappears, leading to a
disappearance of the sigmoidicity of the fluorescence rise
(Schansker et al. 2011). We do stress, however, that
(i) these observations are unrelated to sigmoidicity that
may occur in the absence of DCMU and (ii) this does not
necessarily imply that connectivity between PSII antennae
does not exist. In fact, recent quenching analyses based on
fluorescence lifetime measurements show that in isolated
thylakoid membranes at least four PSII supercomplexes
appear to form connected domains (Lambrev et al. 2011).
Concerning the nature of the conformational change,
another feature of the fluorescence rise in the presence of
DCMU is important. The initial part of the fluorescence
rise is sigmoidal. This is traditionally interpreted as a
consequence of energy transfer between PSII antennae
(Joliot and Joliot 1964). However, Vredenberg (2004,
2008) showed that sigmodicity can also be observed if
kinetic overlap between two consecutive processes with
exponential kinetics occurs. As noted above, a precondition
for this sigmoidicity is that the two rate constants differ by
less than one order of magnitude. Schansker et al. (2011)
proposed that the overlapping process originated from a
light-induced conformational change. Both France et al.
(1992) and Steffen et al. (2001) observed that the fluores-
cence rise induced by a single excitation is exponential.
This suggests that a single excitation induces only a single
process and not both a stable charge separation and the
additional process in parallel: a conformational change
according to the interpretation of Schansker et al. (2011).
Vredenberg (2004) also interpreted the observation of
Steffen et al. (2001) in sequential terms. In addition, France
et al. (1992) using a pump-probe approach observed that
short light pulses of 2 ls or less induced an exponential
fluorescence rise, irrespective of the intensity of the pulse
(the number of excitations per pulse). Sigmoidicity could
only be achieved with two short flashes spaced by a dark
interval of some tens of microseconds or by a sufficiently
long (C50 ls) light pulse. This confirms the involvement
of two sequential reactions (first the reduction of QA and
only then the induction of the conformational change), of
which the second can only occur following a short dark
interval. These results were contested by Hemelrijk and
van Gorkom (1992); however, the latter authors used
continuous measuring light instead of a pump-probe
approach. Thus the comparability of the two sets of
experiments is not straightforward.
The experiments discussed above demonstrate that there
exist two processes involved in the fluorescence rise that
differ in their properties and of which only one can be
assigned to the reduction of QA. Presently, it is unclear why
two excitations of a PSII RC occurring within 2 ls fail to
induce the conformational change(s). A possible explana-
tion is that the stabilization of the charge separated state, or
just the stabilization of the QA- state requires 2–50 ls. As
argued by Schansker et al. (2011) and discussed in more
detail in the following section, the redox state of QA seems
to be important for the stability of the conformational
changes and the amplitude of the associated fluorescence
change.
The stability of QA- determines the stability
of the conformational changes in PSII
Although experimental observations inconsistent with the
pure QA model already exist for many years, alternative
interpretations never gained wider popularity because a
credible mechanism for such an alternative interpretation
Photosynth Res
123

was missing. The QB-site occupancy theory (Vasil’ev and
Bruce 1998; Samson et al. 1999; Yaakoubd et al. 2002)
provides a good starting point for an argument that allows
us to identify the factor that determines the stability of the
conformational changes and how the light-induced con-
formational changes can follow the reduction of QA.
As the ETC becomes reduced, fewer oxidized PQ mole-
cules will be bound to the QB-sites and the re-oxidation rate
of the QA molecules will decrease, i.e., QA will become more
stably reduced. At a light intensity of 15,000 lmol photons
m-2 s-1, the excitation rate (*1 excitation per PSII per
40 ls) is much higher than the forward electron transfer from
QA to QB (halftime of 100–200 ls for the first and
400–600 ls for the second electron (Petrouleas and Crofts
2005). After 2 ms of illumination [99 % of QA may be
reduced whereas the fluorescence intensity is still 30–40 %
below FM (cf. Schansker et al. 2006). However, the lifetime
of QA- in each PSII RC does not depend on the light intensity
but on the rate of forward electron transport, which in turn
depends on the redox state of the ETC. In other words, if the
induction and stability of the conformational changes would
depend on the stability of QA- in each PSII RC the accom-
panying fluorescence yield changes would be sensitive to the
reduction of the ETC (Schansker et al. 2011). It is possible
that as the electron transport chain becomes gradually
reduced there is a continuous induction and relaxation of the
proposed conformational changes and that the fluorescence
yield change associated with these conformational changes
also makes a small contribution to the OJ rise. The rela-
tionship between ETC redox state and thermal phase can also
explain the dependence of the area above the OJIP transient
on the number of electrons needed to reduce the ETC
(Strasser and Strasser 1995; Joliot and Joliot 2002; Toth et al.
2007b).
The above concept linking the stability of QA-, in broad
terms, to the light-induced conformational changes
(Schansker et al. 2011), offers an explanation for the cor-
relation of the JIP phase with the ETC redox state. In order
to explain the observation that the thermal phase and the
slow fluorescence rise in the presence of DCMU, as
stressed above, can only be induced by light following the
reduction of QA, further assumptions must be made. We
must assume that stabilization of the charge separated state
or that of QA- occurs with some delay relative to the
reduction step. It can be speculated that electrostatic cou-
pling between the donor and acceptor sides can account for
such an effect. The existence of such an effect can be
inferred from data showing that treatments affecting the
donor side of PSII like Ca2? depletion can exert an effect
on the redox potential of QA (e.g., Krieger and Rutherford
1997). Because of the presence of strong local fields, the
role of ionic movements should also be considered in these
processes (see below).
Experimental evidence for the occurrence
of conformational changes in PSII and purple bacteria
Several independent observations suggest the occurrence of
conformational changes in PSII, and also in purple bacte-
rial RCs. It has been shown that external electric fields,
applied during illumination and cooling, can induce chan-
ges in the thermoluminescence and thermodepolarization
currents of isolated thylakoid membranes, suggesting
transitions between conformational states of PSII RCs
(Knox et al. 1984). Delosme et al. (1994) studying spinach
PSII membranes and Chlamydomonas reinhardtii core
particles reported that charge separations induce volume
changes (contractions) in PSII. This was confirmed for
manganese-depleted Synechocystis PCC6803 core parti-
cles (Hou et al. 2001), showing that the conformational
changes do not depend on the activity of the oxygen-
evolving complex. In addition, a secondary reaction lead-
ing to an expansion of the sample (5–10 ls of illumination)
was observed; an expansion missing in bacterial RCs and
PSI (Hou et al. 2001).
There exists a vast literature demonstrating light-
induced conformational changes in purple bacterial RCs
(e.g., Abgaryan et al. 1998; McMahon et al. 1998;
Goushcha et al. 2000; Barabash et al. 2002). All these
observations strongly suggest a close association between
conformational transitions and the functioning of PSII-type
RCs. The light-induced conformational changes in bacte-
rial RCs are often rather short-lived. Nagy et al. (2008)
ascribed fluorescence changes in Rhodobacter sphaeroides
with a lifetime of 30–40 ls to a conformational change. In
purple bacteria a change in the protein environment of the
bacteriochlorophyll dimer P has been proposed to occur in
response to these conformational changes (Deshmukh et al.
2011a, b). Of particular interest to the present discussion of
the thermal phase in the PSII fluorescence induction are,
however, studies on bacterial RCs which demonstrate that
(i) the effects of light-induced conformational changes on
the fluorescence parameters (and their relaxation) depend
critically on the lifetimes of the internal electric fields
related to the type and lifetime of the various radical pair
species as intermediates of the charge separation/recom-
bination process (Muller et al. 1995, Holzwarth and Muller
1996), (ii) that the light-induced conformational changes
can be accumulated by subsequent multi-excitations
(Goushcha et al. 1997a; Abgaryan et al. 1998), and (iii) are
principally following a highly non-linear system behavior
(Christophorov et al. 2000, 2003; Goushcha et al. 1997b,
1999, 2000). It should be noted that the term ‘‘light-
induced conformational change’’ used here explicitly
includes not only changes in the conformational backbone
of the protein surrounding the cofactors, but also—and
very likely—different protonation states of amino acid side
Photosynth Res
123

chains which can be brought about by small conforma-
tional changes of the backbone, by a strong electric field
and modified redox states of cofactors. Quite generally, the
implied ‘‘conformational changes’’ are caused by the light-
induced stimulation of charge species (either radical pairs,
charged states, or explicit proton movements) that move or
migrate through the RC protein complex.
Potential processes connecting conformational changes
to a fluorescence yield change causing the additional
fluorescence
When searching for the nature and mechanism of the
conformational changes in PSII responsible for the addi-
tional fluorescence rise component, there are several key
observations which must be taken into consideration.
From the data discussed above, it seems clear that the
conformational changes are primarily induced after the
reduction of QA (Schansker et al. 2011); also, it has been
shown that the additional fluorescence rise, which is
proposed to be associated with the conformational chan-
ges, can only be induced by an additional flash following
the first SSTF with a lag phase, in the 2–50 ls range,
rather than by increasing the intensity of the second flash
fired with a shorter dark interval (France et al. 1992); the
analysis of the fluorescence rise kinetics with long flashes
also indicates that the activationless process attributable
to a charge separation occurs first and it is followed by
the slower rise phase(s) assigned to conformational
changes (Schansker et al. 2011). These findings are in line
with the notion discussed above, that the extent of con-
formational changes induced in the RC complex is con-
trolled primarily by the strength and lifetime of electric
fields and charges moving through or being localized
within the RC. For the present discussion the QA- state is
the most long-lived charged state in the RC. Using the
same arguments, similar effects as caused by internal
electric fields, but perhaps of smaller magnitude, can then
be also caused by external electric fields across the
membrane in which the RC is located.
The lag phase and the above sequence of events may
give us a hint concerning the process(es) occurring after the
reduction of QA. One can speculate that this might be
correlated with (subsequent) charge redistributions in the
aqueous (electrolyte) phases following the generation of
vectorial charge separation in the membrane (Fowler and
Kok 1974; Junge 1977). Indeed, Osvath et al. (1994), by
investigating the light gradient electric signal induced by
short laser flashes on magnetically aligned thylakoid
membranes, have shown that the ionic equilibration in the
lateral direction occurs with a time constant of about 20 ls.
This may bring about conformational changes in the
vicinity of the local charges.
Considering closed RCs (QA reduced), an additional
light reaction and the mechanism responsible for the light-
induced conformational changes, the most likely hypothe-
sis is that they can be induced by the multiple, transient
formation of a P?Pheo- charge separations, which also
occur in closed RCs (Szczepaniak et al. 2008, 2009;
Martinez-Junza et al. 2008) and which can transiently
induce a very strong electrical field, that persists longer
than in open RCs. Such conformational changes do not
necessarily relax fully and can be accumulated by multiple
turnovers. It cannot be ruled out either that protein con-
formational changes are induced or driven locally by the
ultrafast local heat packages produced via (nonradiative)
dissipative processes due to recombination of the charges
in the RC and/or due to dissipation of the excited triplet
states of chl molecules in the core; these, in turn, can
induce elementary structural changes (Cseh et al. 2000;
Gulbinas et al. 2006). The generation of local electric field
and/or thermal transients may facilitate the conformational
adjustment of the reaction center proteins to the local field
due to the presence of QA- coupled to D?, where D is an
electron donor molecule.
Conformational changes may readily explain (light-
induced) changes in the fluorescence yield by affecting the
redox potential of for example the P680/P680? couple.
A similar process has been proposed in photosynthetic
bacteria (Deshmukh et al. 2011a, b). Most likely, a variety
of molecular localizations and conformationally induced
changes of internal rates or redox potentials in the RCs add
up to provide the overall effects of light-induced confor-
mational changes on the observed fluorescence yields. The
picture is complex and at present the various effects are not
studied in detail. Nevertheless, the principal effects are
sufficiently clear to conclude that light-induced confor-
mational changes do have significant effects on fluores-
cence yields in RCs. It is important to note that these
effects are intimately related to the functioning of RCs.
Suffice it to note that it is very clear now that energetic
relaxations of the radical pair energies do occur after
charge separation in RCs (both in open as well as closed
RCs) without which photosynthetic primary processes
would not be efficient and stable charge separation would
not occur over sufficiently long time scales to allow the
subsequent catalytic processes for energy storage (Holzwarth
2008a, b).
The data presented in Schansker et al. (2011) are also
compatible with the suggestion that the fluorescence yield
increase following the reduction of QA reflects a molecular
switch between a photosynthetic and a dissipative state as
earlier suggested by Raszewski and Renger (2008) for the
entire FV. Experimental support for a change in the prop-
erties of PSII RCs following the induction of the confor-
mational changes was obtained by Martinez-Junza et al.
Photosynth Res
123

(2008) and Braslavsky and Holzwarth (2012). Again, it is
quite conceivable that this dissipative state helps—inter
alia—to protect PSII RCs in vivo from damage due to over-
excitation in situations where PSII RCs are closed. The
induction of this putative dissipative state takes less than
10 ms in the presence of stably reduced QA and the
reversal to the energy conversion state takes less than
100 ms once the light is turned off and may even be faster
following the oxidation of QA (Schansker et al. 2011). The
kinetic analysis of these authors also indicates that in a
small fraction of PSII reaction centers QA- becomes re-
oxidized by recombination with the donor side within the
first 100 ms of darkness (relaxation time of 0.4–0.6 s
(Schansker et al. 2005, 2011)) but this does not affect the
analysis and interpretation of the data. The idea is illus-
trated in Fig. 2.
Interpretation of the FV/FM value
The FV/FM ((FM-F0)/FM) is undoubtedly the most widely
used parameter in photosynthesis research. Kitajima and
Butler (1975) using a theoretical treatment of energy fluxes
in PSII based on the concept of Duysens and Sweers (1963)
showed that FV/FM is an expression of the maximum
quantum yield of primary photochemistry of PSII. How-
ever, this analysis is only correct if the redox state of QA is
the only determinant of the fluorescence yield. Within the
context of the conformational change concept, as well as
with the other alternative models, this derivation is not
valid. Another argument that has been put forward against
the FV/FM has to do with the FM value. Neubauer and
Schreiber (1987) suggested that the IP phase was due to a
release of PQ-pool quenching. Schreiber et al. (1995)
considered on this basis the use of the fluorescence
intensity at the I step for the calculation of the FV/FM.
Falkowski and co-workers (e.g., Prasil et al. 1996) argued
that the FM induced by an SSTF was the correct FM value
for the calculation of the FV/FM and that the rest was the
release of PQ-pool quenching. However, as discussed
above, PQ-pool quenching is not observed in leaves.
Recently, Vredenberg and Prasil (Vredenberg et al. 2012)
argued that the IP phase was due to a CET-induced fluo-
rescence yield change and, therefore, in their view, the FM
was problematic for this reason (however, see I.2.4 for a
discussion on the role of CET in the fluorescence rise).
It may be noted here that in the case of photoinhibition,
where there is a linear correlation between the FV/FM value
and the severity of the photoinhibition treatment, it has to
be assumed that the maximum quantum yield of each PSII
RC can change gradually. Although redox titrations of QA
show that potential and variable fluorescence correlate
(e.g., Krieger-Liszkay and Rutherford 1998), this has not
been shown to occur.
The meaning of the FV/FM value can also be defined in a
more phenomenological way. Measurements on protoplasts
of Valerianella locusta (Van Wijk and Krause 1991) and
pea leaves (Schansker and Van Rensen 1999) showed that
the decline of the FV/FM value induced by photoinhibition
paralleled the loss of the oxygen-evolving capacity of PSII,
making it a good parameter to follow the inactivation of
PSII RCs. In the case of heat stress a decline in the FV/FM
value was shown to be due to the increasing inability of the
remaining PSII RCs to reduce the ETC before FNR is
activated (Toth et al. 2005b). Other studies have demon-
strated that linear correlations can be obtained between the
parameter FV’/FM’ (determined in the light-adapted state)
and the quantum yield of CO2 fixation (e.g., Harbinson
et al. 1990; Oberhuber et al. 1993).
Fig. 2 Schematic
representation of the relaxation
and regeneration of the thermalphase. A first strong pulse of
light generates a full OJIP
transient in *200 ms; in
darkness the thermal phaserelaxes within 100 ms and can
be regenerated within 3 ms by a
second light pulse. For the
measurement pea leaves were
used. The chl a fluorescence was
induced by two 5,000 lmol
photons m-2 s-1 pulses of
700 ms and 2 s, respectively,
spaced 80 ms apart. The figure
is modified from Fig. 7 of
Schansker et al. (2011)
Photosynth Res
123

Flash-oxygen measurements may be more appropriate to
determine the PSII quantum yield of primary photochem-
istry. As pointed out above, Shinkarev (2005) calculated an
average miss factor of 7–8 % for the flash-oxygen yield,
implying that the PSII quantum yield of primary photo-
chemistry is[0.92/0.93. In other words, the PSII quantum
yield of primary photochemistry is considerably higher
than normally thought and probably also considerably
more invariable than FV/FM.
Conclusions
Although the QA model, proposed by Duysens and Sweers
nearly four decades ago, has proven to be very successful
and, in broad terms, provided a good model as far as the
photochemical activity of PSII is concerned, it is also
becoming more and more evident that (an) additional
process(es) has/have to be taken into account to understand
the complexity of the fluorescence rise kinetics, most
particularly the so-called thermal phase, the slowly rising
kinetic components of the fluorescence transient.
In this paper we have given an overview of the most
important observations that are difficult to understand
within the framework of the pure QA model and have
critically reviewed alternative explanations, paying partic-
ular attention to the role of light-induced conformational
changes. They appear to offer the most consistent and
convincing explanation for the wide body of presently
available experimental data both in the absence and pres-
ence of DCMU and both at physiological and cryogenic
temperatures. Although many open questions remain,
especially with regard to the precise molecular nature and
mechanism of these changes, the fact that conformational
changes have been observed in PSII reaction centers in
independent experiments, and also in purple bacterial RCs,
adds strong support to this proposal. The observation that
there exist important differences between the electron
transport rates and intermediates of open and closed RCs
points toward the crucial role of conformational changes in
controlling a molecular switch between photosynthetic and
dissipative states.
Amending the pure QA model by the QA plus confor-
mational change model has important consequences for the
interpretation of the parameter FV/FM. While it evidently
remains a sensitive parameter related to the photochemical
activity of PSII, its exact quantitative meaning will have to
be determined again. Further, since the sigmoidal and
biphasic rise kinetics of the chl a fluorescence (in the case
of DCMU blocked RCs) depend on the induction of this
second process, i.e., a superimposition of a fluorescence
yield change driven by light-induced conformational
changes on the reduction kinetics of QA, conclusions
regarding the connectivity of PSII units and on PSII het-
erogeneity, in terms of e.g., a/ß centers, will have to be re-
evaluated critically.
Acknowledgments The authors thank Drs. Petar Lambrev, Laszlo
Kovacs, and Fabrice Rappaport for useful discussions. This work was
supported by the Hungarian Research Foundation (OTKA, grant no.
MB08B82403, PD72718 and CNK80345 to G.S., S.Z.T. and G.G.,
respectively), and by NIH-A*STAR grant (TET-10-1-2011-0279) to
GG. S.Z.T. acknowledges financial support by the Bolyai Janos
Research Foundation of the Hungarian Academy of Sciences. This
work was also supported by the Marie Curie Initial Training Network
‘‘HARVEST’’ sponsored by the 7th Framework Program of the
European Union (grant number 238017 to G.G. and A.R.H.).
References
Abgaryan GA, Christophorov LN, Goushcha AO, Holzwarth AR,
Kharkyanen VN, Knox PP, Lukashev EA (1998) Effects of
mutual influence of photoinduced electron transitions and slow
structural rearrangements in bacterial photosynthetic reaction
centers. J Biol Phys 24:1–17
Amesz J, Fork DC (1967) Quenching of chlorophyll fluorescence by
quinones in algae and chloroplasts. Biochim Biophys Acta
143:97–107
Baker NR (2008) Chlorophyll fluorescence: a probe of photosynthesis
in vivo. Annu Rev Plant Biol 59:89–113
Barabas K, Kravcova T, Garab G (1993) Flash-induced reduction of
cytochrome b-559 by QB- in chloroplasts in the presence of
protonophores. Photosynth Res 36:59–64
Barabash YM, Berezetskaya NM, Christophorov LN, Goushcha AO,
Kharkyanen VN (2002) Effects of structural memory in protein
reactions. J Chem Phys 116:4339–4352
Boussac A, Sugiura M, Rappaport F (2011) Probing the quinone
binding site of photostem II from Thermosynechococcus elong-atus containing PsbA1 or PabA3 as the D1 protein through the
binding characteristics of herbicides. Biochim Biophys Acta
1807:119–129
Braslavsky SE, Holzwarth AR (2012) Role of carotenoids in
photosystem II (PS II) reaction centres. Int J Thermophys
33:2021–2025
Brudvig GW, Casey JL, Sauer K (1983) The effect of temperature on
the formation and decay of the multiline EPR signal species
associated with photosynthetic oxygen evolution. Biochim
Biophys Acta 723:366–371
Bulychev AA, Vredenberg WJ (2001) Modulation of photosystem II
chlorophyll fluorescence by electrogenic events generated by
photosystem I. Bioelectrochemistry 54:157–168
Butler WL, Kitajima M (1975) Fluorescence quenching in photosys-
tem II of chloroplasts. Biochim Biophys Acta 376:116–125
Carillo N, Arana JL, Vallejos RH (1981) Light modulation of
chloroplast membrane-bound ferredoxin-NADP? oxidoreduc-
tase. J Biol Chem 256:1058–1059
Christophorov LN, Holzwarth AR, Kharkyanen VN, van Mourik F
(2000) Structure-function self-organization in nonequilibrium
macromolecular systems. Chem Phys 256:45–60
Christophorov LN, Holzwarth AR, Kharkyanen VN (2003) Confor-
mational regulation in single molecule reactions. Ukrainian J
Phys 48:672–680
Cseh Z, Rajagopal S, Tsonev T, Busheva M, Papp E, Garab G (2000)
Thermooptic effect in chloroplast thylakoid membranes; Ther-
mal and light stability of pigment arrays with different levels of
structural complexity. Biochemistry 39:15250–15257
Photosynth Res
123

Danielsson R, Suorsa M, Paakkarinen V, Albertsson P-A, Styring S,
Aro E-M, Mamedov F (2006) Dimeric and monomeric organi-
zation of photosystem II; Distribution of five distinct complexes
in the different domains of the thylakoid membrane. J Biol Chem
281:14241–14249
Dau H, Sauer K (1991) Electric field effect on chlorophyll fluores-
cence and its relation to photosystem II charge separation
reactions studied by a salt-jump technique. Biochim Biophys
Acta 1098:49–60
de Wijn R, van Gorkom HJ (2001) Kinetics of electron transfer from
QA to QB in photosystem II. Biochemistry 40:11912–11922
Dekker JP, van Grondelle R (2000) Primary charge separation in
photosystem II. Photosynth Res 63:195–208
Delosme R (1967) Etude de l’induction de fluorescence des algues
vertes et des chloroplastes au debut d’une illumination intense.
Biochim Biophys Acta 143:108–128
Delosme R (1971) Photosynthese—variations du rendement de
fluorescence de la chlorophylle in vivo sous l’action d’eclairs
de forte intensite. C R Acad Sci Paris 272D:2828–2831
Delosme R, Beal D, Joliot P (1994) Photoacoustic detection of flash-
induced charge separation in photosynthetic systems. Spectral
dependence of the quantum yield. Biochim Biophys Acta
1185:56–64
Deshmukh SS, Williams JC, Allen JP, Kalman L (2011a) Light-
induced conformational changes in photosynthetic reaction
centers: dielectric relaxation in the vicinity of the dimer.
Biochemistry 50:340–348
Deshmukh SS, Williams JC, Allen JP, Kalman L (2011b) Light-
induced conformational changes in photosynthetic reaction
centers: redox regulated proton pathway near the dimer.
Biochemistry 50:3321–3331
Diner BA (1977) Dependence of the deactivation reactions of
photosystem II on the redox state of plastoquinone pool A
varied under anaerobic conditions; Equilibria on the acceptor
side of photosystem II. Biochim Biophys Acta 460:247–258
Duysens LNM, Sweers HE (1963) Mechanisms of two photochemical
reactions in algae as studied by means of fluorescence, In:
Japanese Society of Plant Physiologists (ed) Studies on micro-
algae and photosynthetic bacteria, Special Issue of Plant and Cell
Physiology. University of Tokyo Press, Tokyo, pp 353–372
Forbush B, Kok B, McGloin MP (1971) Cooperation of charges in
photosynthetic oxygen evolution. II. Damping of flash yield
oscillation, deactivation. Photochem Photobiol 14:307–321
Fowler CF, Kok B (1974) Direct observation of a light-induced
electric field in chloroplasts. Biochim Biophys Acta 357:
308–318
France LL, Geacintov NE, Breton J, Valkunas L (1992) The
dependence of the dregrees of sigmoidicities of fluorescence
induction curves in spinach chloroplasts on the duration of
actinic pulses in pump-probe experiments. Biochim Biophys
Acta 1101:105–119
Goushcha AO, Kapoustina MT, Kharkyanen VN, Holzwarth AR
(1997a) Nonlinear dynamic processes in an ensemble of
photosynthetic reaction centers; Theory and experiment. J Phys
Chem B 101:7612–7619
Goushcha AO, Kharkyanen VN, Holzwarth AR (1997b) Nonlinear
light-induced properties of photosynthetic reaction centers under
low intensity irradition. J Phys Chem B 101:259–265
Goushcha AO, Holzwarth AR, Kharkyanen VN (1999) Self-regula-
tion phenomenon of electron-conformational transitions in
biological electron transfer under nonequilibrium conditions.
Phys Rev E 59:3444–3452
Goushcha AO, Kharkyanen VN, Scott GW, Holzwarth AR (2000)
Self-regulation phenomena in bacterial reaction centers. I. Gen-
eral theory. Biophys J 79:1237–1252
Gulbinas V, Karpicz R, Garab G, Valkunas L (2006) Nonequilibrium
heating in LHCII complexes monitored by ultrafast absorbance
transients. Biochemistry 45:9559–9565
Harbinson J, Hedley CL (1993) Changes in P-700 oxidation during
the early stages of the induction of photosynthesis. Plant Physiol
103:649–660
Harbinson J, Genty B, Baker NR (1990) The relationship between
CO2 assimilation and electron transport in leaves. Photosynth
Res 25:213–224
Heber U (2002) Irrungen, Wirrungen? The Mehler reaction in relation
to cyclic electron transport in C3 plants. Photosynth Res
73:223–231
Hemelrijk PW, van Gorkom HJ (1992) No double hit involved in
fluorescence induction of photosystem II of spinach chloroplasts.
In: Murata N (ed) Research in photosynthesis. Kluwer Aca-
demic, Dordrecht, pp 33–36
Holzwarth AR (2008a) Ultrafast primary reactions in the photosys-
tems of oxygen evolving organisms. In: Braun M, Gilch P, Zinth
W (eds) Ultrashort laser pulses in biology and medicine.
Springer, Dordrecht, pp 141–164
Holzwarth AR (2008b) Primary reactions—from isolated complexes
to intact plants. In: Allen JF, Gantt E, Golbeck JH, Osmond B
(eds) Photosynthesis. Energy from the sun. Springer, Dordrecht,
pp 77–83
Holzwarth AR, Muller MG (1996) Energetics and kinetics of radical
pairs in reaction centers from Rhodobacter sphaeroides; A
femtosecond transient absorption study. Biochemistry 35:11820–
11831
Hou J-M, Boichenko VA, Diner BA, Mauzerall D (2001) Thermo-
dynamics of electron transfer in oxygenix photosynthetic
reaction centers: volume change, enthalpy, and entropy of
electron-transfer reactions in manganese-depleted photosystem
II core complexes. Biochemistry 40:7117–7125
Joliot A (1974) Effect of low temperature (-30 to -60�C) on the
reoxidation of the photosystem II primary electron acceptor in
the presence and absence of 3(3,4-dichlorophenyl)-1,1-dimethyl-
urea. Biochim Biophys Acta 357:439–448
Joliot A, Joliot P (1964) Etude cinetique de la reaction photochimique
liberant l’oxygene au cours de la photosynthese. C R Acad Sci
Paris 258:4622–4625
Joliot P, Joliot A (1979) Comparative study of the fluorescence yield
and of the C550 absorption change at room temperature.
Biochim Biophys Acta 546:93–105
Joliot P, Joliot A (1981) A photosystem II electron acceptor which is
not a plastoquinone. FEBS Lett 134:155–158
Joliot P, Joliot A (2002) Cyclic electron transfer in plant leaf. Proc
Natl Acad Sci USA 99:10209–10214
Junge W (1977) Membrane potentials in photosynthesis. Annu Rev
Plant Physiol 28:503–536
Kautsky H, Appel W, Amann H (1960) Chlorophyllfluorescenz und
Kohlensaure-assimilation: XIII. Die Fluorescencekurve und die
Photochemie der Pflanze. Biochem Z 332:277–292
Kitajima M, Butler WL (1975) Quenching of chlorophyll fluores-
cence and primary photochemistry in chloroplasts by dibro-
mothymoquinone. Biochim Biophys Acta 376:105–115
Knox PP, Venediktov PS, Kononenko AA, Garab GI, Faludi-Daniel A
(1984) Role of electric polarization in the thermoluminescence
of chloroplasts. Photochem Photobiol 40:119–125
Kramer DM, Johnson G, Kiirats O, Edwards GE (2004) New
fluorescence parameters for the determination of QA redox state
and excitation energy fluxes. Photosynth Res 79:209–218
Krieger A, Rutherford AW (1997) Comparison of chloride-depleted
and calcium-depleted PSII: the midpoint potential of QA and
susceptibility to photodamage. Biochim Biophys Acta 1319:
91–98
Photosynth Res
123

Krieger-Liszkay A, Rutherford AW (1998) Influence of herbicide
binding on the redox potential of the quinone acceptor in
photosystem II: relevance to photodamage and phytotoxicity.
Biochemistry 37:17339–17344
Kurreck J, Schodel R, Renger G (2000) Investigation of the
plastoquinone pool size and fluorescence quenching in thylakoid
membranes and photosystem II (PS II) membrane fragments.
Photosynth Res 63:171–182
Lambrev PH, Schmitt F-J, Kussin S, Schoengen M, Varkonyi Z,
Eichler HJ, Garab G, Renger G (2011) Functional domain size in
aggregates of light-harvesting complex II and thylakoid mem-
branes. Biochim Biophys Acta 1807:1022–1031
Lavergne J (1982) Two types of primary acceptors in chloroplasts
photosystem II. I. Different recombination properties. Photobio-
chem Photobiophys 3:257–271
Lazar D (2003) Chlorophyll a fluorescence rise induced by high light
illumination of dark-adapted plant tissue studied by means of a
model of photosystem II and considering photosystem II
heterogeneity. J Theor Biol 220:469–503
Lazar D (2009) Modelling of light-induced chlorophyll a fluorescence
rise (O-J-I-P transient) and changes in 820 nm-transmittance
signal of photosynthesis. Photosynthetica 47:483–498
Lazar D, Pospısil P (1999) Mathematical simulation of chlorophyll
a fluorescence rise measured with 3-(30,40-dichlorophenyl)-1,1-
dimethylurea-treated barley leaves at room and high tempera-
tures. Eur Biophys J 28:468–477
Lazar D, Schansker G (2009) Models of Chlorophyll a fluorescence
transients. In: Laisk A, Nedbal L, Govindjee (eds) Photosynthe-
sis in silico: understanding complexity from molecules to
ecosystems, vol 29., Advances in photosynthesis and respiration.
Springer, Dordrecht, pp 85–123
Loach PA (1976) Oxidation-reduction potentials, absorbance bands
and molar absorbance of compounds used in biochemical
studies. In: Fasman GD (ed) Handbook of biochemistry and
molecular biology, 3rd edn, vol I., Physical Chemical Data. CRC
Press, Cleveland, pp 122–130
Mano J, Hideg E, Asada K (2004) Ascorbate in thylakoid lumen
functions as an alternative electron donor to photosystem II and
photosystem I. Arch Biochem Biophys 429:71–80
Martinez-Junza V, Szczepaniak M, Braslavsky SE, Sander J, Now-
aczyk M, Rogner M, Holzwarth AR (2008) A photoprotection
mechanism involving the D2 branch in photosystem II cores with
closed reaction centers. Photochem Photobiol Sci 7:1337–1343
Maxwell K, Johnson GN (2000) Chlorophyll fluorescence— a
practical guide. J Exp Bot 51:659–668
McCauley SW, Melis A, Tang GM-S, Arnon DI (1987) Protono-
phores induce plastoquinol oxidation and quench chloroplast
fluorescence: evidence for a cyclic, proton-conducting pathway
in oxygenic photosynthesis. Proc Natl Acad Sci USA
84:8424–8428
McMahon BH, Muller JD, Wraight CA, Nienhaus GU (1998)
Electron transfer and protein dynamics in the photosynthetic
reaction center. Biophys J 74:2567–2587
Melis A (1985) Functional properties of photosystem IIß in spinach
chloroplasts. Biochim Biophys Acta 808:334–342
Melis A, Homann PH (1975) Kinetic analysis of the fluorescence
induction in 3-(3,4- dichlorophenyl)-1,1-dimethylurea poisoned
chloroplasts. Photochem Photobiol 21:431–437
Melis A, Homann PH (1976) Heterogeneity of the photochemical
centers in system II of chloroplasts. Photochem Photobiol
23:343–350
Moise N, Moya I (2004a) Correlation between lifetime heterogeneity
and kinetics heterogeneity during chlorophyll fluorescence
induction in leaves: 1. Mono-frequency phase and modulation
analysis reveals a conformational change of a PSII pigment
complex during the IP thermal phase. Biochim Biophys Acta
1657:33–46
Moise N, Moya I (2004b) Correlation between lifetime heterogeneity
and kinetics heterogeneity during chlorophyll fluorescence
induction in leaves: 2. Multi-frequency phase and modulation
analysis evidences a loosely connected PSII pigment-protein
complex. Biochim Biophys Acta 1657:47–60
Morin P (1964) Etudes des cinetiques de fluorescence de la
chlorophylle in vivo, dans les premiers instants qui suivent le
debut de l’illumination. J Chim Phys 61:674–680
Muller MG, Dorra D, Holzwarth AR, Gad’on N, Drews G (1995)
Time-dependent radical pair relaxation in chromatophores of an
antenna-free mutant from Rhodobacter capsulatus. In: Mathis P
(ed) Photosynthesis: from light to biosphere, vol 1. Kluwer
Academic, Dordrecht, pp 595–598
Munday JCM, Govindjee (1969) Light-induced changes in the
fluorescence yield of chlorophyll a in vivo. III. The dip and
the peak in the fluorescence transient of Chlorella pyrenoidosa.
Biophys J 9:1–21
Nagy L, Maroti P, Terazima M (2008) Spectrally silent light induced
conformation change in photosynthetic reaction centers. FEBS
Lett 582:3657–3662
Neubauer C, Schreiber U (1987) The polyphasic rise of chlorophyll
fluorescence upon onset of strong continuous illumination.
I. Saturation characteristics and partial control by the photosys-
tem II acceptor side. Z Naturforsch 42c:1246–1254
Oberhuber W, Dai Z-Y, Edwards GE (1993) Light dependence of
quantum yields of photosystem II and CO2 fixation in C3 and C4
plants. Photosynth Res 35:265–274
Osvath S, Meszena G, Barzda V, Garab G (1994) Trapping
magnetically oriented chloroplast thylakoid membranes in gels
for electric measurements. J Photochem Photobiol, B 26:
287–292
Paillotin G (1976) Movement of excitations in photosynthetic
domains of photosystem II. J Theor Biol 58:237–252
Papageorgiou GC, Govindjee (2004) Chlorophyll a fluorescence: a
signature of photosynthesis, vol 19., Advances in photosynthesis
and respiration. Springer, Dordrecht 818 pp
Petrouleas V, Crofts AR (2005) The iron-quinone acceptor complex.
In: Wydrzynski TJ, Satoh K (eds) Photosystem II: the light-
driven water: plastoquinone oxidoreductase, vol 22., Advances
in photosynthesis and respiration. Springer, Dordrecht,
pp 177–206
Pospısil P, Dau H (2002) Valinomycin sensitivity proves that light-
induced thylakoid voltages result in millisecond phase of
chlorophyll fluorescence transients. Biochim Biophys Acta
1554:94–100
Prasil O, Kolber Z, Berry JA, Falkowski PG (1996) Cyclic electron
flow around Photosystem II in vivo. Photosynth Res 48:395–410
Prince RC, Linkletter SJG, Dutton PL (1981) The thermodynamic
properties of some commonly used oxidation-reduction media-
tors, inhibitors and dyes, as determined by polarography.
Biochim Biophys Acta 635:132–148
Raszewski G, Renger T (2008) Light harvesting in photosystem II
core complexes is limited by the transfer to the trap: can the core
complex turn into a photoprotective mode? J Am Chem Soc
130:4431–4446
Robinson HH, Crofts AR (1984) Kinetics of proton uptake and the
oxidation-reduction reactions of the quinone acceptor complex
of PS II from pea chloroplasts. In: Sybesma C (ed) Advances in
photosynthesis research, vol I. Martinus Nijhoff/Dr W Junk, Den
Haag, pp 477–480
Rubin A, Riznichenko G (2009) Modeling of the primary processes in
a photosynthetic membrane. In: Laisk A, Nedbal L, Govindjee
(eds) Photosynthesis in silico: understanding complexity from
Photosynth Res
123

molecules to ecosystems, vol 29., Advances in photosynthesis
and respiration. Springer, Dordrecht, pp 151–176
Samson G, Bruce D (1996) Origins of the low yield of chlorophyll
a fluorescence induced by a single turnover flash in spinach
thylakoids. Biochim Biophys Acta 1276:147–153
Samson G, Prasil O, Yaakoubd B (1999) Photochemical and thermal
phases of chlorophyll a fluorescence. Photosynthetica 37:163–182
Satoh K (1981) Fluorescence induction and activity of ferredoxin-
NADP? reductase in Bryopsis chloroplasts. Biochim Biophys
Acta 638:327–333
Schansker G, Strasser RJ (2005) Quantification of non-QB-reducing
centers in leaves using a far-red pre-illumination. Photosynth
Res 84:145–151
Schansker G, van Rensen JJS (1999) Performance of active photo-
system II centers in photoinhibited pea leaves. Photosynth Res
62:175–184
Schansker G, Toth SZ, Strasser RJ (2005) Methylviologen and
dibromothymoquinone treatments of pea leaves reveal the role of
photosystem I in the Chl a fluorescence rise OJIP. Biochim
Biophys Acta 1706:250–261
Schansker G, Toth SZ, Strasser RJ (2006) Dark-recovery of the Chl
a fluorescence transient (OJIP) after light adaptation: the qT-
component of non-photochemical quenching is related to an
activated photosystem I acceptor side. Biochim Biophys Acta
1757:787–797
Schansker G, Yuan Y, Strasser RJ (2008) Chl a fluorescence and
820 nm transmission changes occurring during a dark-to-light
transition in pine needles and pea leaves: a comparison. In: Allen
JF, Osmond B, Golbeck JH, Gantt E (eds) Energy from the sun.
Springer, Dordrecht, pp 951–955
Schansker G, Toth SZ, Kovacs L, Holzwarth AR, Garab G (2011)
Evidence for a fluorescence yield change driven by a light-
induced conformational change within photosystem II during the
fast chlorophyll a fluorescence rise. Biochim Biophys Acta
1807:1032–1043
Schlodder E (2008) Temperature dependence of the reduction kinetics
of P680? in oxygen-evolving PSII complexes throughout the
range from 320 to 80 K. In: Allen JF, Osmond B, Golbeck JH,
Gantt E (eds) Energy from the sun. Springer, Dordrecht,
pp 187–190
Schreiber U (1986) Detection of rapid induction kinetics with a new
type of high-frequency modulated chlorophyll fluorometer.
Photosynth Res 9:261–272
Schreiber U (2002) Assessment of maximal fluorescence yield:
donor-side dependent quenching and QB-quenching. In: Van
Kooten O, Snel JFH (eds) Plant spectrofluorometry: applications
and basic research. Rozenberg, Amsterdam, pp 23–47
Schreiber U (2004) Pulse-amplitude-modulation (PAM) fluorometry
and saturation pulse method: An overview. In: Papageorgiou
GC, Govindjee (eds) Chlorophyll a fluorescence: a signature of
photosynthesis, vol 19. Springer, Dordrecht, pp 279–319
Schreiber U, Neubauer C (1990) O2-dependent electron flow,
membrane energization and the mechanism of non-photochem-
ical quenching of chlorophyll fluorescence. Photosynth Res
25:279–293
Schreiber U, Neubauer C, Klughammer C (1989) Devices and
methods for room-temperature fluorescence analysis. Philos
Trans R Soc Lond B 323:241–251
Schreiber U, Hormann H, Neubauer C, Klughammer C (1995)
Assessment of photosystem II photochemical quantum yield by
chlorophyll fluorescence quenching analysis. Aust J Plant
Physiol 22:209–220
Shinkarev V (2005) Flash-Induced oxygen evolution in photosynthe-
sis: simple solution for the extended S-state model that includes
misses, double-hits, inactivation, and backward-transitions. Bio-
phys J 88:412–421
Sokolove PM, Marsho TV (1979) The effect of valinomycin on
electron transport in intact spinach chloroplasts. FEBS Lett
100:179–184
Steffen R, Christen G, Renger G (2001) Time-resolved monitoring of
flash-induced changes of fluorescence quantum yield and decay
of delayed light emission in oxygen-evolving photosynthetic
organisms. Biochemistry 40:173–180
Stirbet A, Govindjee (2012) Chlorophyll a fluorescence induction: a
personal perspective of the thermal phase, the J-I-P rise.
Photosynth Res 113:15–61
Strasser RJ (1978) The grouping model of plant photosynthesis. In:
Akoyunoglou G, Argyroudi-Akoyunoglou JH (eds) Chloroplast
development. Elsevier/North-Holland Biomedical Press,
Amsterdam, pp 513–524
Strasser RJ, Govindjee (1991) The F0 and the O-J-I-P fluorescence
rise in higher plants and algae. In: Argyroudi-Akoyunoglou JH
(ed) Regulation of chloroplast biogenesis. Plenum Press, New
York, pp 423–426
Strasser BJ, Strasser RJ (1995) Measuring fast fluorescence transients
to address environmental questions: The JIP test. In: Mathis P
(ed) Photosynthesis: from light to biosphere, vol V. Kluwer
Academic, Dordrecht, pp 977–980
Strasser RJ, Tsimilli-Michael M, Srivastava A (2004) Analysis of the
chlorophyll a fluorescence transient. In: Papageorgiou GC,
Govindjee (eds) Chlorophyll a fluorescence: a signature of
photosynthesis, vol 19., Advances in photosynthesis and respi-
ration. Springer, Dordrecht, pp 321–362
Szczepaniak M, Sugiura M, Holzwarth AR (2008) The role of TyrD
in the electron transfer kinetics in photosystem II. Biochim
Biophys Acta 1777:1510–1517
Szczepaniak M, Sander J, Nowaczyk M, Muller MG, Rogner M,
Holzwarth AR (2009) Charge separation, stabilization, and
protein relaxation in photosystem II core particles with closed
reaction center. Biophys J 96:621–631
Tamura N, Inoue H, Inoue Y (1990) Inactivation of the water-
oxidizing complex by exogenous reductants in PSII membranes
depleted of extrinsic proteins. Plant Cell Physiol 31:469–477
Toth SZ, Schansker G, Strasser RJ (2005a) In intact leaves, the
maximum fluorescence level (FM) is independent of the redox
state of the plastoquinone pool: a DCMU-inhibition study.
Biochim Biophys Acta 1708:275–282
Toth SZ, Schansker G, Strasser RJ (2005b) Biophysical studies of
photosystem II-related recovery processes after a heat pulse in
barley seedling (Hordeum vulgare L.). J Plant Physiol
162:181–194
Toth SZ, Schansker G, Garab G, Strasser RJ (2007a) Photosynthetic
electron transport activity in heat-treated barley leaves: the role
of internal alternative electron donors to photosystem II.
Biochim Biophys Acta 1767:295–305
Toth SZ, Schansker G, Strasser RJ (2007b) A non-invasive assay of
the plastoquinone pool redox state based on the OJIP-transient.
Photosynth Res 93:193–203
Toth SZ, Puthur JT, Nagy V, Garab G (2009) Experimental evidence
for ascorbate-dependent electron transport in leaves with inac-
tive oxygen-evolving complexes. Plant Physiol 149:1568–1578
Trissl H-W, Lavergne J (1994) Fluorescence induction from photo-
system II: analytical equations for the yields of photochemistry
and fluorescence derived from analysis of a model including
exciton-radical pair equilibrium and restricted energy transfer
between photosynthetic units. Aust J Plant Physiol 22:183–193
van Kooten O, Snel JFH (1990) The use of chlorophyll fluorescence
nomenclature in plant stress physiology. Photosynth Res
25:147–150
van Wijk KJ, Krause GH (1991) O2-dependence of photoinhibition at
low temperature in intact protoplasts of Locusta valerianella L.
Planta 186:135–142
Photosynth Res
123

Vasil’ev S, Bruce D (1998) Nonphotochemical quenching of excita-
tion energy in photosystem II. A picoseconds time-resolved
study of the low yield of chlorophyll a fluorescence induced by
single-turnover flash in isolated spinach thylakoids. Biochemis-
try 37:11046–11054
Vernotte C, Etienne AL, Briantais J-M (1979) Quenching of the
system II chlorophyll fluorescence by the plastoquinone pool.
Biochim Biophys Acta 545:519–527
Vredenberg WJ (2000) A three-state model for energy trapping and
chlorophyll fluorescence in photosystem II incorporating radical
pair recombination. Biophys J 79:26–38
Vredenberg WJ (2004) System analysis and photoelectrochemical
control of chlorophyll fluorescence in terms of trapping models
of photosystem II: A challenging view. In: Papageorgiou GC,
Govindjee (eds) Chlorophyll a fluorescence: a signature of
photosynthesis, vol 19., Advances in photosynthesis and respi-
ration. Springer, Dordrecht, pp 133–172
Vredenberg WJ (2008) Analysis of initial chlorophyll fluorescence
induction kinetics in chloroplasts in terms of rate constants of
donor side quenching release and electron trapping in photosys-
tem II. Photosynth Res 96:83–97
Vredenberg WJ (2011) Kinetic analysis and mathematical modeling
of primary photochemical and photoelectrochemical processes in
plant photosystems. Biosystems 103:138–151
Vredenberg WJ, Bulychev AA (2002) Photo-electrochemical control
of photosystem II chlorophyll fluorescence in vivo. Bioelectro-
chemistry 57:123–128
Vredenberg WJ, Bulychev A (2003) Photoelectric effects on chloro-
phyll fluorescence of photosystem II in vivo. Kinetics in the
absence and presence of valinomycin. Bioelectrochemistry
60:87–95
Vredenberg W, Kasalicky V, Durchan M, Prasil O (2006) The
chlorophyll a fluorescence induction pattern in chloroplasts upon
repetitive single turnover excitations: accumulation and function
of QB-nonreducing centers. Biochim Biophys Acta 1757:173–181
Vredenberg W, Durchan M, Prasil O (2007) On the chlorophyll a
fluorescence yield in chloroplasts upon excitation with twin
turnover flashes (TTF) and high frequency flash trains. Photo-
synth Res 93:183–192
Vredenberg W, Durchan M, Prasil O (2012) The analysis of PS II
photochemical activity using single and multi-turnover excita-
tions. J Photochem Photobiol, B 107:45–54
Wientjes E, Croce R (2012) PMS: photosystem I electron donor or
fluorescence quencher. Photosynth Res 111:185–191
Yaakoubd B, Andersen R, Desjardins Y, Samson G (2002) Contri-
butions of the free oxidized and QB-bound plastoquinone
molecules to the thermal phase of chlorophyll-a fluorescence.
Photosynth Res 74:251–257
Zhu X-G, Govindjee, Baker NR, deSturler E, Ort DR, Long SP (2005)
Chlorophyll a fluorescence induction kinetics in leaves predicted
from a model describing each discrete step of excitation energy
and electron transfer associated with Photosystem II. Planta
223:114–133
Zimanyi L, Garab G (1989) Configuration of the electric field and
distribution of ions in energy transducing biological membranes:
model calculations in a vesicle containing discrete charges.
J Theor Biol 138:59–76
Photosynth Res
123
Copyright © 2022 FDOKUMEN




















