
Chemical basis of resistance in pulses to Callosobruchus maculatus (F.) (Coleoptera: Bruchidae
11
Chemical basis of resistance in pulses to Callosobruchus maculatus (F.) (Coleoptera: Bruchidae) S. Ignacimuthu*, S. Janarthanan, B. Balachandran Entomology Research Institute, Loyola College, Chennai-600 034, India Accepted 1 July 1999 Abstract Seeds of various wild and cultivated pulses express varied degrees of resistance to damage by Callosobruchus maculatus, a major pest of stored pulses. Seeds were analysed for trypsin/chymotrypsin inhibitors (qualitatively and quantitatively) and protein profiles (SDS-polyacrylamide gel electrophoresis) to study the chemical basis of resistance to bruchid infestation. Seeds of the wild pulses Dunberia ferruginea, Neonotonia wightii, Tephrosia cararensis, Cajanus albicans and Vigna bourneae dier from other wild germplasms as well as cultivars in having characteristically higher levels of trypsin/ chymotrypsin inhibitors. SDS-PAGE analyses of seed proteins of Dolichos lablab, Lablab purpureus and Rhyncocea cana reveal the presence of high levels of non-nutrient chemicals such as vicilins, a-amylase inhibitors, and lectins in quantities that may impart resistance to C. maculatus. The importance of these protease inhibitors and other non-nutrient chemicals are discussed with reference to resistance to C. maculatus infestation. # 1999 Published by Elsevier Science Ltd. All rights reserved. Keywords: Pulses; Trypsin inhibitor; Chymotrypsin inhibitor; SDS-PAGE; Bruchid; Resistance 1. Introduction Pulses are a good source of dietary proteins and other essential nutrients. However, post- harvest insect infestation severely aects quality and storability of produce (Singh, 1978; Steele et al., 1985). Pulses are known to contain a variety of non-nutrient compounds such as lectins, proteinase inhibitors, a-amylase inhibitors and other compounds which could be part of an array of constitutive defences against attack by stored grain pests, particularly those belonging Journal of Stored Products Research 36 (2000) 89–99 0022-474X/99/$ - see front matter # 1999 Published by Elsevier Science Ltd. All rights reserved. PII: S0022-474X(99)00031-4 www.elsevier.com/locate/jspr * Corresponding author. Tel.: +91-44-826-5542; fax: +91-44-826-5544. E-mail address: [email protected] (S. Ignacimuthu)
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Chemical basis of resistance in pulses to Callosobruchus maculatus (F.) (Coleoptera: Bruchidae

Chemical basis of resistance in pulses to Callosobruchusmaculatus (F.) (Coleoptera: Bruchidae)
S. Ignacimuthu*, S. Janarthanan, B. Balachandran
Entomology Research Institute, Loyola College, Chennai-600 034, India
Accepted 1 July 1999
Abstract
Seeds of various wild and cultivated pulses express varied degrees of resistance to damage byCallosobruchus maculatus, a major pest of stored pulses. Seeds were analysed for trypsin/chymotrypsininhibitors (qualitatively and quantitatively) and protein pro®les (SDS-polyacrylamide gel electrophoresis)to study the chemical basis of resistance to bruchid infestation. Seeds of the wild pulses Dunberiaferruginea, Neonotonia wightii, Tephrosia cararensis, Cajanus albicans and Vigna bourneae di�er fromother wild germplasms as well as cultivars in having characteristically higher levels of trypsin/chymotrypsin inhibitors. SDS-PAGE analyses of seed proteins of Dolichos lablab, Lablab purpureus andRhyncocea cana reveal the presence of high levels of non-nutrient chemicals such as vicilins, a-amylaseinhibitors, and lectins in quantities that may impart resistance to C. maculatus. The importance of theseprotease inhibitors and other non-nutrient chemicals are discussed with reference to resistance to C.maculatus infestation. # 1999 Published by Elsevier Science Ltd. All rights reserved.
Keywords: Pulses; Trypsin inhibitor; Chymotrypsin inhibitor; SDS-PAGE; Bruchid; Resistance
1. Introduction
Pulses are a good source of dietary proteins and other essential nutrients. However, post-harvest insect infestation severely a�ects quality and storability of produce (Singh, 1978; Steeleet al., 1985). Pulses are known to contain a variety of non-nutrient compounds such as lectins,proteinase inhibitors, a-amylase inhibitors and other compounds which could be part of anarray of constitutive defences against attack by stored grain pests, particularly those belonging
Journal of Stored Products Research 36 (2000) 89±99
0022-474X/99/$ - see front matter # 1999 Published by Elsevier Science Ltd. All rights reserved.PII: S0022-474X(99)00031-4
www.elsevier.com/locate/jspr
* Corresponding author. Tel.: +91-44-826-5542; fax: +91-44-826-5544.E-mail address: [email protected] (S. Ignacimuthu)

to the family Bruchidae (Janzen et al., 1986; Gatehouse et al., 1990). While the exact nature ofresistance in certain pulses to Callosobruchus maculatus (F.) remains largely unknown, somerecent studies emphasize the presence of certain non-nutrient (storage) proteins in seeds ofresistant varieties as the basis of resistance (Macedo et al., 1993). In addition, trypsin andchymotrypsin inhibitors, besides their well-known function as storage proteins, are alsoimportant in imparting resistance (Liener and Kakade, 1980; Ryan, 1990). The e�cacy of anysecondary (non-nutrient) compound is indicated by biological assay against the insect pest(Agarwal et al., 1988). Several studies suggest the importance of biological assay to assess thepotential of such compounds to impart resistance (Gatehouse et al., 1989; Cardona et al., 1990;Xavier-Filho, 1991; Pusztai et al., 1993; Macedo et al., 1995; Fory et al., 1996). In view of thevariation observed in damage due to feeding by C. maculatus on various wild as well ascultivated pulses, the present study was planned to identify chemical factors that areresponsible for imparting resistance.
2. Materials and methods
2.1. Seeds
Seeds of the wild pulses were collected from the Palney Hills of the Western Ghats,Tamilnadu, India (latitude 108; longitude 788; altitude 2010 m from MSL), whereas somecultivated seeds (Cajanus cajan (L.) Millspaugh ICP14770 and ICP14990; Cicer arietinum (L.)COG29 and CO3; Vigna unguiculata (L.) Walp. CO4 and V. mungo (L.) Hepper CO5) wereobtained from the Department of Plant Breeding, Tamil Nadu Agricultural University,Coimbatore, India. The other cultivated seeds namely, V. unguiculata VCP8; V. mungo VBN1;V. radiata (L.) Wilczek K851 and UGG4 were obtained from National Pulses Research Centre,Vamban, Tamil Nadu, India.
2.2. Chemical analysis
Trypsin and chymotrypsin inhibitors were estimated following the methods described bydella Gatta et al. (1989) and Erlanger and Frances Edel Cooper (1996). They were expressed interms of units/g seed materials (a trypsin or a-chymotrypsin unit (TU or CU) is de®ned as theamount of trypsin or a-chymotrypsin that catalyses the hydrolysis of 1 mol of substrate permin. A trypsin or chymotrypsin inhibitor unit (TIU or CIU) is the reduction in activity of therespective enzymes by one unit, i.e. TU or CU respectively.
2.3. Electrophoretic studies on trypsin and chymotrypsin inhibitors
Seeds were milled to a ®ne powder (after removal of the seed coat) and protease inhibitorswere extracted by incubating overnight in Tris±HCl±CaCl2 bu�er, pH 8.1 (at a ratio of 1:10 w/v) at 48C. The homogenate (the ®ne powder with the extraction bu�er) was subsequentlycentrifuged at 12,000 rpm at 48C for 15 min. The supernatant collected was used forelectrophoretic analysis with polyacrylamide gel electrophoresis (Native PAGE) as described by
S. Ignacimuthu et al. / Journal of Stored Products Research 36 (2000) 89±9990

Kollipara et al. (1991). At the end of the run, the stacking gel was removed and the separatinggel stained for trypsin and chymotrypsin inhibitor activities based on Uriel and Bergas (1968)with some modi®cations as described by Kollipara and Hymowitz (1992).
2.4. Seed protein pro®le
To analyse proteins of both wild and cultivar pulses, SDS-polyacrylamide gel electrophoresis(SDS-PAGE) was performed according to the method of Laemmli (1970). Proteins wereextracted from powdered pulses with 0.1 M Tris±HCl bu�er, pH 8.0 (at a ratio of 1:10 w/v)using sterilised glass powder in a mortar and pestle for 15 min at 0±58C (on ice). Extracts werecentrifuged at 15,000 g for 10 min at 58C. Aliquots of supernatant were used to quantify theproteins using the Bradford method with BSA as standard (Bradford, 1976). An equal quantityof each of the protein samples (100 mg) was applied to the gels. Standard molecular markerproteins (Sigma) were simultaneously run for comparison.
Table 1
Trypsin and chymotrypsin inhibitor contents (units/g) in wild and cultivated germplasms1
Varieties Trypsin inhibitors Chymotrypsin inhibitors
Wild pulses
Cajanus albicans 2960205.7a 412234.9a
Dunberia ferruginea 2774204.2b 425216.3a
Neonotonia wightii 2700209.0b 385213.1b
Vigna bourneae 1886207.2c 269214.8c
Tephrosia cararensis 1035204.1d 148206.5d
Lablab purpureus(brown) 890214.7e 125204.9e
Rhyncocea rufescens 523209.0f 136211.4de
Rhyncocea cana 440204.8g 62214.7f
Dolichos lablab 338206.5h 46207.3gf
Lablab purpureus (black) 300212.2i 43205.7g
Canavalia virosa 134203.2j 22207.3h
Cultivars
Cicer arietinum CO3 1238202.5a 15326.0a
Cicer arietinum COG 29 1233236.2a 17725.6b
Vigna unguiculata CO4 1075233.6b 15223.4a
Vigna unguiculata VCP8 700225.3c 6027.3c
Cajanus cajanICP 14770 575216.3d 8222.5d
Vigna radiataUGG4 418201.2e 6022.5ec
Cajanus cajan ICP 14990 408210.7e 5825.8ec
Vigna mungo VBN1 335201.8f 4824.9f
Vigna radiata K851 333218.0f 020.0Vigna mungo CO5 289209.8g 4126.5f
1 Values represent mean2SE of four replications. Data analysed using Student's t-test (P < 0.01). Values in thesame column with the same superscript letters are not signi®cantly di�erent.
S. Ignacimuthu et al. / Journal of Stored Products Research 36 (2000) 89±99 91

2.5. Bioassay
A stock culture of C. maculatus was reared on green gram (V. radiata K851 cultivar) seedsat 28218C in glass jars covered with muslin cloth. For feeding bioassays, seeds wereconditioned at 28218C and 70% r.h. for a period of 10 days before use. Seeds of eachcultivar/wild germplasm were kept in glass vials covered with muslin cloth and a single pair ofnewly emerged adults of C. maculatus from the stock culture was introduced into each of sixvials for oviposition. Six replicates were maintained for each variety. Vials were maintained at28218C and 70% r.h. After 5 days, the numbers of eggs laid on the seeds of each variety werecounted. Such seeds were observed for a maximum duration of 60 days and the numbers ofemerging adults were counted for a duration of 8 days with adults being isolated to preventfurther breeding. Developmental period, percentage adult emergence and percentage loss inweight [(initialÿ®nal)/initial seed weight � 100] were calculated to evaluate developmentalperformance of C. maculatus on cultivars and wild germplasm of pulses.
2.6. Statistical analysis
All data were subjected to Student's t-test and the level of signi®cance was tested atP< 0.01. Each value was statistically compared with other values of the same parameter.
3. Results
Seeds of wild germplasms generally were found to contain higher amounts of trypsin andchymotrypsin inhibitors than cultivated seeds (Table 1). Maximum levels of trypsin inhibitors
Fig. 1. Electrophoretogram showing trypsin inhibitor bands in the wild germplasms. Note strong reaction at the
region of Rf 0.60±0.80. Wild pulses: (a) Rhyncocea rufescens, (b) Rhyncocea cana, (c) Canavalia virosa, (d) Lablabpurpureus (black), (e) Lablab purpureus (brown), (f) Tephrosia cararensis, (g) Dolichos lablab.
S. Ignacimuthu et al. / Journal of Stored Products Research 36 (2000) 89±9992

(TI) were observed in Cajanus albicans (Wight and Arnott) Maesen with 2960 units/g followedby seeds of Dunberia ferruginea Wight and Arnott and Neonotonia wightii (Wight and Arnott)Lackey which exhibited 2774 units/g and 2700 units/g, respectively. The seeds of a brownvariety of Lablab purpureus (L.) Sweet were found to contain a signi®cantly (P< 0.01) higheramount of TI than the black variety. A signi®cant (P< 0.01) di�erence in TI content wasnoticed at intraspeci®c levels between cultivars (Table 1) except in Cicer arietinum where nosigni®cant di�erences were observed. The amount of TI was always greater than the amount ofchymotrypsin inhibitor. Among wild germplasms the amount of chymotrypsin inhibitors (CI)were higher in D. ferruginea (425 units/g), C. albicans (412 units/g) and N. wightii (385 units/g)(Table 1). Interestingly, CI was absent from the V. radiata K851 cultivar. Although the rangeof inhibitors in both the cultivated and wild accessions overlapped, overall the levels weresigni®cantly higher in the wild accessions.Electrophoretograms (Fig. 1) showed prominent bands of TI in the seeds of L. purpureus
(black) at Rf values 0.75 and 0.80, whereas L. purpureus (brown) showed two successive bandsat the same region with Rf values 0.75 and 0.77, respectively. Although the other wildgenotypes such as Rhyncocea rufescens (Willdenow) de Candolle and D. ferruginea reveal aclear TI band at Rf value 0.60, the seeds of V. bourneae Gamble, on the other hand, exhibitedthree bands at Rf values 0.50, 0.60 and 0.70. Among the various cultivars, gel electrophoresisrevealed a TI band at Rf value 0.70 in the cultivated seeds of V. radiata (K851 and UGG4)and C. arietinum (COG 29 and CO3) (Fig. 2). Interestingly, the other cultivated seeds did notreveal any inhibitor bands.
Fig. 2. Electrophoretogram showing trypsin inhibitor bands. Note the absence of reaction in cultivated germplasms.
Wild pulses: (a) Cajanus albicans, (b) Dunberia ferruginea, (c) Vigna unguiculata, (d) Neonotonia wightii. Cultivars:(e) Vigna radiata K851, (f) Vigna radiata UGG4, (g) Cicer arietinum COG29, (h) Cicer arietinum CO3.
S. Ignacimuthu et al. / Journal of Stored Products Research 36 (2000) 89±99 93

Four intense CI bands at Rf values 0.65, 0.75, 0.77 and 0.80 were observed in L. purpureus(black) while the brown variety of L. purpureus showed three intense bands with Rf values0.75, 0.77 and 0.80 (Fig. 3). The seeds of Canavalia virosa (Roxburgh) Wight and Arnott hadtwo intense bands at Rf values 0.45 and 0.75, whereas Rhyncocea rufescens had one prominentband at Rf value 0.60 and two more bands at Rf values 0.77 and 0.80. Two bands at Rf values0.77 and 0.80 were observed in the seeds of R. cana (Willdenow) de Candolle while only oneband at Rf value 0.60 was observed in the seeds of Dolichos lablab L. The electrophoretogramshowed no CI bands in the cultivated seeds.In the present investigation, trypsin and chymotrypsin inhibitors varied in both number and
migration patterns among various wild germplasms of pulses. There was also variation inintensity of these bands and some seeds showed no bands (Figs. 1±3). This could be due tolower concentration of an inhibitor protein or lower a�nity (weaker inhibition) for bovinetrypsin or a-chymotrypsin. Inhibitor contents relative to wild germplasms and cultivar pulsesare presented in Table 1. A broad variability of activities among samples was observed for alltested inhibitors. Wild varieties of pulses, D. ferruginea, N. wightii, C. albicans, R. rufescensand V. bourneae used in this study were characterised by higher values of both TI and CI(Table 1). Among them, C. albicans, D. ferruginea, R. rufescens were found to be infested bythe bruchid, C. maculatus (see results of bioassay studies, Table 2) or, in other words, thesusceptible varieties (reported above) showed higher inhibitor activity against trypsin andchymotrypsin. The other wild germplasms exhibiting lower inhibitor activity against trypsinand chymotrypsin were found to be susceptible (except the genus Lablab, in which the gelsshowed more intense TI and CI bands).SDS-PAGE seed protein pro®les showed di�erences in banding patterns among various wild
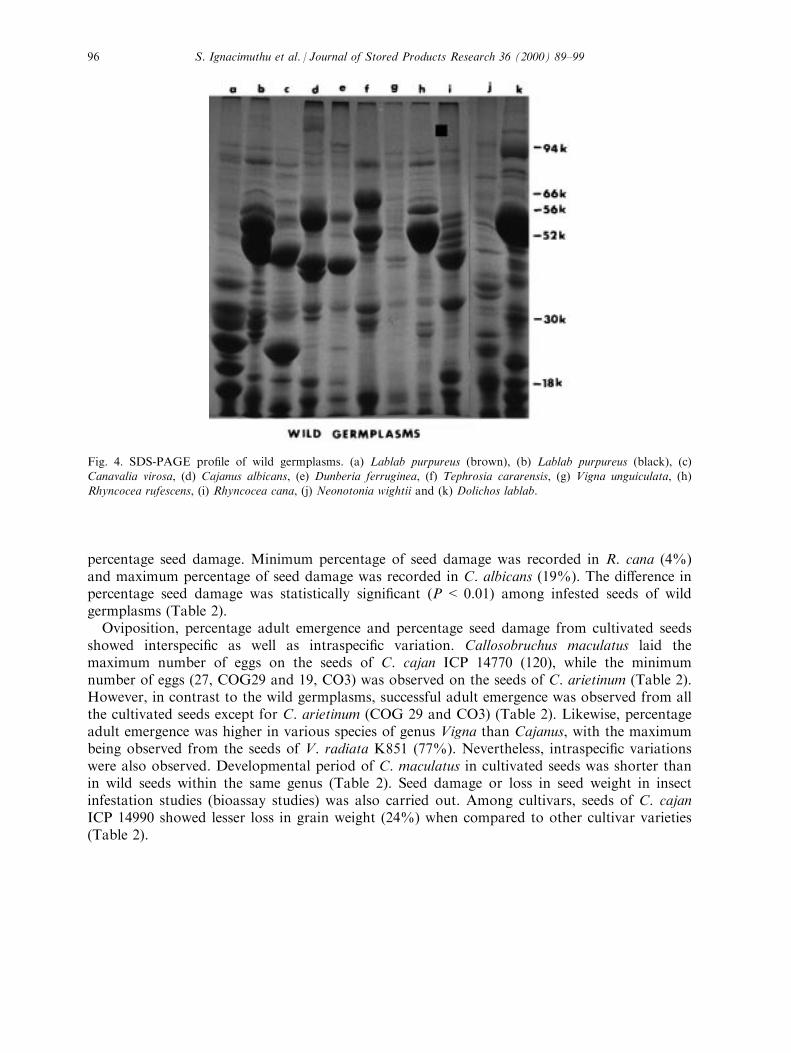
germplasm lines (Fig. 4). Major polypeptide components at 94, 56 and 52 kD regions wereidenti®ed in wild accessions of L. purpureus (brown), R. cana and D. lablab. All the other wild
Fig. 3. Electrophoretogram showing chymotrypsin inhibitor bands in the wild germplasm. Note strong reaction atthe region of Rf 0.45±0.80. Wild pulses: (a) Rhyncocea rufescens, (b) Rhyncocea cana, (c) Canavalia virosa, (d)
Lablab purpureus (black), (e) Lablab purpureus (brown), (f) Tephrosia cararensis, (g) Dolichos lablab and (h) Cajanusalbicans.
S. Ignacimuthu et al. / Journal of Stored Products Research 36 (2000) 89±9994

germplasms showed variations in polypeptides of low molecular weights in the range of 20±45kD. All cultivar accessions show similar banding patterns except Cicer arietinum (Fig. 5). Ingeneral, polypeptide components of 94, 66, 56, 52 and 30 kD were identi®ed in all accessionsof cultivars together with some minor bands.Developmental performance of C. maculatus on various varieties of wild germplasms has
been summarized in Table 2. Maximum numbers of eggs (96) were laid on the seeds of C.albicans, whereas eggs were not laid on the seeds of Tephrosia cararensis de Candolle.Variation in percentage adult emergence was observed in both cultivated and wild germplasmswith observed higher values in wild germplasms. Percentage adult emergence was found to be41, 45, 57 and 20 in the wild seeds of Dunberia ferruginea, C. albicans, R. rufescens and R.cana, respectively (Table 2). Adult emergence was not observed from other wild germplasms.Accordingly, they supported lower levels of infestation by the bruchid as is re¯ected by
Table 2Biological performance of Callosobruchus maculatus on di�erent species of wild and cultivated pulses1
Varieties Fecundity(No. of eggs/female)
Adult emergence(%)
Developmental period(days)
Seed damage(%)
Wild pulses
Cajanus albicans 9626.3a 4523.6a 3721.6a 1921.7a
Dolichos lablab 7625.4b Nil2 ± ±Vigna bourneae 7125.6bc Nil2 ± ±
Lablab purpureus (brown) 7020.7cd Nil2 ± ±Canavalia virosa 6824.0cde Nil2 ± ±Lablab purpureus (black) 5929.0e Nil2 ± ±Dunberia ferruginea 3428.3f 4125.8a 3221.4b 1422.8b
Rhyncocea cana 2726.4fg 2025.2b 3423.0ab 421.4c
Rhyncocea rufescens 2524.6g 5727.2c 3820.7a 1523.4a
Neonotonia wightii 1422.3h Nil2 ± ±
Tephrosia cararensis ± ± ± ±
Cultivars
Cajanus cajan ICP14770 12028.8a 3921.4a 2722.2a 3922.5a
Cajanus cajan ICP14990 11329.5a 4722.6b 2521.6a 2423.1b
Cicer arietinum COG29 2722.1b Nil2 ± ±Cicer arietinum CO3 1923.6c Nil2 ± ±Vigna unguiculata CO4 8525.8d 6925.9c 2824.1a 4323.5ac
Vigna unguiculata VCP8 9724.6e 6324.2cd 2723.5a 3721.6a
Vigna mungo CO5 5826.2f 7526.8ce 2421.5a 3821.4a
Vigna mungo VBN1 4923.4g 7025.3cf 2421.3a 3722.0a
Vigna radiata K851 7623.5h 7721.5e 2522.5a 4424.5ac
Vigna radiata UGG4 8826.5id 7224.2ef 2620.5a 4221.7c
1 Values represent mean2SE of six replications. Data analysed by Student's t-test (P < 0.01); ±, not tested.
Values in the same column with the same superscript letters are not signi®cantly di�erent.2 No adult emergence was observed.
S. Ignacimuthu et al. / Journal of Stored Products Research 36 (2000) 89±99 95
percentage seed damage. Minimum percentage of seed damage was recorded in R. cana (4%)and maximum percentage of seed damage was recorded in C. albicans (19%). The di�erence inpercentage seed damage was statistically signi®cant (P< 0.01) among infested seeds of wildgermplasms (Table 2).Oviposition, percentage adult emergence and percentage seed damage from cultivated seeds
showed interspeci®c as well as intraspeci®c variation. Callosobruchus maculatus laid themaximum number of eggs on the seeds of C. cajan ICP 14770 (120), while the minimumnumber of eggs (27, COG29 and 19, CO3) was observed on the seeds of C. arietinum (Table 2).However, in contrast to the wild germplasms, successful adult emergence was observed from allthe cultivated seeds except for C. arietinum (COG 29 and CO3) (Table 2). Likewise, percentageadult emergence was higher in various species of genus Vigna than Cajanus, with the maximumbeing observed from the seeds of V. radiata K851 (77%). Nevertheless, intraspeci®c variationswere also observed. Developmental period of C. maculatus in cultivated seeds was shorter thanin wild seeds within the same genus (Table 2). Seed damage or loss in seed weight in insectinfestation studies (bioassay studies) was also carried out. Among cultivars, seeds of C. cajanICP 14990 showed lesser loss in grain weight (24%) when compared to other cultivar varieties(Table 2).
Fig. 4. SDS-PAGE pro®le of wild germplasms. (a) Lablab purpureus (brown), (b) Lablab purpureus (black), (c)Canavalia virosa, (d) Cajanus albicans, (e) Dunberia ferruginea, (f) Tephrosia cararensis, (g) Vigna unguiculata, (h)Rhyncocea rufescens, (i) Rhyncocea cana, (j) Neonotonia wightii and (k) Dolichos lablab.
S. Ignacimuthu et al. / Journal of Stored Products Research 36 (2000) 89±9996

4. Discussion
Identi®cation and analysis of sources of resistance in seeds provide us with opportunities toidentify and breed pulse varieties that can naturally resist bruchid attacks. The production ofgrain legumes is devastated by C. maculatus and, with current limitations on chemical controlof stored grain pests, the probability of identifying the chemical basis of resistance in pulses,speci®cally the enzyme inhibitors and some deterrent proteins, prompted us to pursue thisstudy.Protease inhibitors are proteins or polypeptides which bind proteolytic enzymes and, in
plants which produce them, are thought to provide a form of natural defence againstherbivorous insects (Green and Ryan, 1972; Ryan, 1979; Broadway et al., 1986; Xavier-Filhoand Campos, 1989; Burgass et al., 1994). In the present investigation in cultivars, bothquantitative and qualitative analyses on TI and CI showed lower activities when compared tothose from their wild counterparts, except C. arietinum, and all were susceptible to C.maculatus. It has been explained that, since a threshold value to discriminate between resistantand susceptible lines cannot be established for any tested inhibitors, evaluating only one or twoantimetabolites cannot provide an e�ective way of checking the degree of resistance towardsstorage pests (Piergiovanni et al., 1994). This conclusion agreed with previous studies whereseveral antimetabolites in a larger number of germplasms of pulses had been analysed (Bakeret al., 1989; Xavier-Filho et al., 1989; Cardona et al., 1990; Piergiovanni et al., 1991; Fory et
Fig. 5. SDS-PAGE pro®le of cultivated seeds. (a) Vigna unguiculata VCP8, (b) Vigna unguiculata CO4, (c) Cicerarietinum COG29, (d) Cicer arietinum CO3, (e) Vigna mungo CO5, (f) Vigna mungo VBN1, (g) Vigna radiata K851,(h) Vigna radiata UGG4, (i) Vigna radiata CO3, (j) Vigna radiata CO4, (k) Cajanus cajan ICP 14770 and (l) Cajanus
cajanICP 14990.
S. Ignacimuthu et al. / Journal of Stored Products Research 36 (2000) 89±99 97

al., 1996). It seems more likely, in the light of these observations, that more antimetabolitescould be involved in the mechanism of bruchid beetle resistance in the case of the present pulsevarieties. Furthermore, antinutrients, ``like the vicilins (55 kD polypeptides), lectins (30 kDpolypeptides), and a-amylase inhibitors (13±17 kD polypeptides) which are reported to have arole in imparting resistance to bruchids'' (Gatehouse et al., 1989; Pratt et al., 1990; Pereira DeSales et al., 1992; Macedo et al., 1993, 1995; Fory et al., 1996), may also be involved inimparting resistance in the present investigation. The present work on protein pro®lesenumerates the presence of bands of 56 and 52 kD in wild accessions of Dolichos lablab, R.cana and L. purpureus under denaturing conditions. In addition, our results displayed severalminor bands of higher mobilities on SDS-PAGE gels which match well with the protein anti-metabolites reported in earlier ®ndings (Liener and Kakade, 1980; Ryan, 1990; Macedo et al.,1993, 1995).
Acknowledgements
The authors gratefully acknowledge the ®nancial assistance and support provided by a grantto Fr S. Ignacimuthu from the Department of Biotechnology, Government of India, NewDelhi. We also thank Dr Alok Sen, Entomology Research Institute, Loyola College, Chennai,for his fruitful and critical suggestions.
References
Agarwal, A., Sone, Lal, Gupta, K.C., 1988. Natural products as protectants of pulses against pulse beetles. Bulletinof Grain Technology 26, 154±164.
Baker, T.A., Nielsen, S., Shade, R.E., Singh, B.B., 1989. Physical and chemical attributes of cowpea lines resistantand susceptible to Callosobruchus maculatus (F.) (Coleoptera: Bruchidae). Journal of Stored Products Research25, 1±8.
Bradford, M.M., 1976. A rapid and sensitive method for the quanti®cation of microgram quantities of protein utilis-ing the principle of dye binding. Analytical Biochemistry 72, 248±254.
Broadway, R.M., Du�ey, S.S., Pearce, G., Ryan, C.A., 1986. Plant proteinase inhibitors: a defence against herbivor-ous insects? Entomologia Experimentalis et Applicata 41, 33±38.
Burgass, E.P.J., Main, C.A., Stevens, P.S., Christeller, J.T., Gatehouse, A.M.R., Laing, W.A., 1994. E�ects of pro-tease inhibitor concentration and combinations on the survival, growth and gut enzyme activities of the black®eld cricket, Teleogryllus commodus. Journal of Insect Physiology 40, 803±811.
Cardona, C., Kornegay, J., Posso, C.E., Morales, F., Ramirez, H., 1990. Comparative value of four arcelin variantsin the development of dry bean lines resistant to the Mexican bean weevil. Entomologia Experimentalis etApplicata 56, 197±206.
della Gatta, C., Piergiovanni, A.R., Ng, N.Q., Carnovale, E., Perrino, P., 1989. Trypsin inhibitor levels in raw andcooked cowpea (Vigna unguiculata ) seeds. Lebensmittel-Wissenschaft und Technologie 22, 78±80.
Erlanger, B.F., Frances Edel Cooper, A.G., 1996. The action of chymotrypsin on two new chromogenic substrates.Archives of Biochemistry and Biophysics 115, 206.
Fory, L.F., Finhardi-Filho, F., Quintero, C.M., Osborn, T.C., Cardona, C., Chrispeels, M.J., Mayer, J., 1996.Alpha amylase inhibitors in resistance to common beans to the Mexican bean weevil and the bean weevil(Coleoptera: Bruchidae). Journal of Economic Entomology 89, 204±210.
Gatehouse, A.M.R., Dewey, F.M., Dove, J., Fenton, K.A., Pusztai, A., 1989. E�ect of seed lectins from Phaseolusvulgaris on the development of larvae of Callosobruchus maculatus. Mechanism of toxicity. Journal of the Scienceof Food and Agriculture 35, 373±380.
S. Ignacimuthu et al. / Journal of Stored Products Research 36 (2000) 89±9998

Gatehouse, A.M.R., Minney, H., Dobie, P., Hilder, V.A., 1990. Biochemical resistance to bruchid attack in legumes;
investigation and exploitation. In: Fujii, K., Gatehouse, A.M.R., Johnson, C.D., Mitchell, R., Yoshida, T. (Eds.),
Bruchids and Legumes: Economics, Ecology and Coevolution. Kluwer Academic Publishers, Dordrecht, pp. 241±
256.
Green, T.R., Ryan, C.A., 1972. Wound induced proteinase inhibitors in plant leaves: a possible defence mechanism
against insects. Science 175, 776±777.
Janzen, D.H., Ryan, C.A., Liener, I.E., Pearse, G., 1986. Potentially defensive proteins in mature seeds of 59 species
of tropical Leguminosae. Journal of Chemical Ecology 12, 1469±1480.
Kollipara, K.P., Domagalsjki, J.M., Hymowitz, I.A., 1991. Quick method of resolving soybean Kunitz trypsin in-
hibitors using polyacrylamide gel electrophoresis. Soybean Genetics Newsletter 18, 234±236.
Kollipara, K.P., Hymowitz, T., 1992. Characterization of trypsin and chymotrypsin inhibitors in the wild perennial
Glycine species. Journal of Agriculture and Food Chemistry 40, 2356±2383.
Laemmli, U.K., 1970. Cleavage of structural proteins during assembly of the head of bacteriophage T4. Nature 227,
680±685.
Liener, I.E., Kakade, M.L., 1980. Protease inhibitors. In: Liener, I.E. (Ed.), Toxic Constituents of Plant Foodstu�s.
Academic Press, New York.
Macedo, M.L.R., Andrade, L.B.S., Morales, R.A., Xavier-Filho, J., 1993. Vicilin variants and the resistance of cow-
pea (Vigna unguiculata ) seeds to the cowpea weevil (Callosobruchus maculatus ). Comparative Biochemistry and
Physiology 105C, 89±94.
Macedo, M.L.R., Fernandes, K.V.S., Sales, M.P., Xavier-Filho, J., 1995. Puri®cation and properties of storage pro-
teins (vicilins) from cowpea (Vigna unguiculata ) seed which are susceptible or resistant to the bruchid beetle
Callosobruchus maculatus. Brazilian Journal of Medical Biology Research 28, 183±190.
Pereira De Sales, M., Macedo, M.L.R., Xavier-Filho, J., 1992. Digestibility of cowpea (Vigna unguiculata ) vicilins
by pepsin, papain and bruchid (insect) midgut proteinases. Comparative Biochemistry and Physiology 1033, 945±
950.
Piergiovanni, A.R., Sonnante, G., della Gatta, C., Perrino, P., 1991. Digestive enzyme inhibitors and storage pest re-
sistance in cowpea (Vigna unguiculata ) seeds. Euphytica 54, 191±194.
Piergiovanni, A.R., della Gatta, C., Sergio, L., Perrino, P., 1994. High antinutrient levels and bruchid resistance of
cowpea (Vigna unguiculata ) seeds. Euphytica 80, 59±62.
Pratt, R.C., Singh, N.K., Shade, R.E., Murdock, L.L., Bressan, R.A., 1990. Isolation and partial characterization of
a seed lectin from tepary bean that delays bruchid beetle development. Plant Physiology 93, 1453±1459.
Pusztai, A., Even, S.W.B., Grant, G., Brown, D.S., Stewart, J.C., Peumans, W.J., Van Damme, E.J.M., Bardocz, S.,
1993. Antinutritive e�ects of wheat germ agglutinin and other N-acetylglucosamine speci®c lectins. Brazilian
Journal of Nutrition 70, 313±321.
Ryan, C.A., 1979. Proteinase inhibitors. In: Rosenthal, G.A., Janzen, D.H. (Eds.), Herbivores: Their Interaction
with Secondary Plant Metabolites. Academic Press, New York, pp. 599±618.
Ryan, C.A., 1990. Protease inhibitors: genes for improving defenses against insects and pathogens. Annual Review
of Phytopathology 28, 425±449.
Singh, S.R., 1978. Pests of grain legumes and their control in Nigeria. In: Singh, S.R., Van Emden, H.F., Taylor,
T.A. (Eds.), Resistance to Pests of Cowpeas in Nigeria. Academic Press, New York, pp. 267±297.
Steele, W.M., Allen, D.J., Summer®eld, R.J., 1985. Cowpea (Vigna unguiculata ) (L.) (Walp.). In: Summer®eld, R.J.,
Roberts, E.H. (Eds.), Grain Legume Crops. Collins, London, pp. 537±544.
Uriel, J., Bergas, J., 1968. Characterization of natural inhibitors of trypsin and chymotrypsin by electrophoresis in
acrylamide agarose gels. Nature 218, 578±580.
Xavier-Filho, J., Campos, F.A.P., 1989. Proteinase inhibitors. In: Cheeke, P.R. (Ed.), Toxicants of Plant Origin,
vol. III. CRC Press.
Xavier-Filho, J., Campos, F.A.P., Ary, M.B., Silva, C.P., Carvalho, M.M.M., Macedo, M.L.R., Lemos, F.J.A.,
Grant, G., 1989. Poor correlation between the levels of proteinase inhibitors found in seeds of di�erent cultivars
of cowpea (Vigna unguiculata ) and the resistance/susceptibility to predation by Callosobruchus maculatus. Journal
of Agriculture and Food Chemistry 37, 1139±1143.
Xavier-Filho, J., 1991. The resistance of seeds of cowpea (Vigna unguiculata ) to the cowpea weevil (Callosobruchus
maculatus ). Memoris Institute of Oswaldo 86, 75±77.
S. Ignacimuthu et al. / Journal of Stored Products Research 36 (2000) 89±99 99




















