
Characterization of three human sec14p-like proteins: α-Tocopherol transport activity and...
13
Research paper Characterization of three human sec14p-like proteins: a-Tocopherol transport activity and expression pattern in tissues Jean-Marc Zingg a, * ,1 , Petra Kempna a,1 , Marcel Paris a , Elke Reiter a , Luis Villacorta a , Rita Cipollone a , Adelina Munteanu b , Clara De Pascale a , Stefano Menini c , Alexandra Cueff d , Michel Arock d , Angelo Azzi a , Roberta Ricciarelli e a Institute of Biochemistry and Molecular Medicine, University of Bern, Bu ¨hlstrasse 28, CH-3012 Bern, Switzerland b Department of Physiology, Faculty of Medicine, ‘Carol Davila’ University, Bucharest, Romania c Department of Clinical Sciences, ‘‘La Sapienza’’ University, Roma, Italy d LBPA CNRS UMR 8113, Ecole Normale Supe ´rieure de Cachan 61, Ave du Pre ´sident Wilson 94235 Cachan CEDEX, France e Department of Experimental Medicine, University of Genoa, Italy Received 11 February 2008; accepted 16 July 2008 Available online 5 August 2008 Abstract Three closely related human sec14p-like proteins (hTAP1, 2, and 3, or SEC14L2, 3, and 4, respectively) have been described. These proteins may participate in intracellular lipid transport (phospholipids, squalene, tocopherol analogues and derivatives) or influence regulatory lipid- dependent events. Here, we show that the three recombinant hTAP proteins associate with the Golgi apparatus and mitochondria, and enhance the in vitro transport of radioactively labeled a-tocopherol to mitochondria in the same order of magnitude as the human a-tocopherol transfer protein (a-TTP). hTAP1 and hTAP2 are expressed in severalcell lines, whereas the expression level of hTAP3 is low. Expression of hTAP1 is induced in human umbilical cord blood-derived mast cells upon differentiation by interleukin 4. In tissues, the three hTAPs are detectable ubiquitously at low level; pronounced and localized expression is found for hTAP2 and hTAP3 in the perinuclear region in cerebellum, lung, liver and adrenal gland. hTAP3 is well expressed in the epithelial duct cells of several glands, in ovary in endothelial cells of small arteries as well as in granulosa and thecal cells, and in testis in Leydig cells. Thus, the three hTAPs may mediate lipid uptake, secretion, presentation, and sub-cellular localization in a tissue-specific manner, possibly using organelle- and enzyme-specific docking sites. Ó 2008 Elsevier Masson SAS. All rights reserved. Keywords: Vitamin E; Mitochondria; Golgi; Sec14-like proteins; Phosphatidylinositol kinase; Secretion 1. Introduction The intracellular transport of hydrophobic molecules, such as phospholipids, fatty acids, cholesterol, steroids and hydro- phobic vitamins between membranes and their enrichment at specific cellular sites are critically dependent on the binding to specific transport proteins (reviewed in Refs. [1e3]). For the intracellular transport of vitamin E (a-tocopherol), several proteins such as the microsomal triglyceride transfer protein (MTTP), the a-tocopherol transfer protein (a-TTP) and the tocopherol associated proteins (TAPs) have been identified (reviewed in Refs. [4,5]). Whereas the function of liver a-TTP in a-tocopherol transport and enrichment of VLDL is well described in vitro and in vivo [6], the role of the human tocopherol associated proteins (hTAPs) is less clear. In humans, three highly homologous TAPs (hTAP1/2/3, or SEC14L2/3/4, respectively) have been described that are related to the Saccharomyces cerevisiae SEC14p protein [7e9]. When compared to SEC14p and a-TTP, the three hTAPs carry * Corresponding author. Present address: Vascular Biology Laboratory, Office 621, JM-USDA Human Nutrition Research Center on Aging, Tufts University, 711 Washington Street, Boston, MA 02111, USA. Tel: þ1 617 556 3229; fax: þ1 617 556 3224. E-mail address: [email protected] (J.-M. Zingg). 1 These authors contributed equally to this work. 0300-9084/$ - see front matter Ó 2008 Elsevier Masson SAS. All rights reserved. doi:10.1016/j.biochi.2008.07.008 Available online at www.sciencedirect.com Biochimie 90 (2008) 1703e1715 www.elsevier.com/locate/biochi
Transcript of Characterization of three human sec14p-like proteins: α-Tocopherol transport activity and...

Available online at www.sciencedirect.com
Biochimie 90 (2008) 1703e1715www.elsevier.com/locate/biochi
Research paper
Characterization of three human sec14p-like proteins: a-Tocopheroltransport activity and expression pattern in tissues
Jean-Marc Zingg a,*,1, Petra Kempna a,1, Marcel Paris a, Elke Reiter a, Luis Villacorta a,Rita Cipollone a, Adelina Munteanu b, Clara De Pascale a, Stefano Menini c,
Alexandra Cueff d, Michel Arock d, Angelo Azzi a, Roberta Ricciarelli e
a Institute of Biochemistry and Molecular Medicine, University of Bern, Buhlstrasse 28, CH-3012 Bern, Switzerlandb Department of Physiology, Faculty of Medicine, ‘Carol Davila’ University, Bucharest, Romania
c Department of Clinical Sciences, ‘‘La Sapienza’’ University, Roma, Italyd LBPA CNRS UMR 8113, Ecole Normale Superieure de Cachan 61, Ave du President Wilson 94235 Cachan CEDEX, France
e Department of Experimental Medicine, University of Genoa, Italy
Received 11 February 2008; accepted 16 July 2008
Available online 5 August 2008
Abstract
Three closely related human sec14p-like proteins (hTAP1, 2, and 3, or SEC14L2, 3, and 4, respectively) have been described. These proteinsmay participate in intracellular lipid transport (phospholipids, squalene, tocopherol analogues and derivatives) or influence regulatory lipid-dependent events. Here, we show that the three recombinant hTAP proteins associate with the Golgi apparatus and mitochondria, and enhancethe in vitro transport of radioactively labeled a-tocopherol to mitochondria in the same order of magnitude as the human a-tocopherol transferprotein (a-TTP). hTAP1 and hTAP2 are expressed in several cell lines, whereas the expression level of hTAP3 is low. Expression of hTAP1 isinduced in human umbilical cord blood-derived mast cells upon differentiation by interleukin 4. In tissues, the three hTAPs are detectableubiquitously at low level; pronounced and localized expression is found for hTAP2 and hTAP3 in the perinuclear region in cerebellum, lung,liver and adrenal gland. hTAP3 is well expressed in the epithelial duct cells of several glands, in ovary in endothelial cells of small arteries aswell as in granulosa and thecal cells, and in testis in Leydig cells. Thus, the three hTAPs may mediate lipid uptake, secretion, presentation, andsub-cellular localization in a tissue-specific manner, possibly using organelle- and enzyme-specific docking sites.� 2008 Elsevier Masson SAS. All rights reserved.
Keywords: Vitamin E; Mitochondria; Golgi; Sec14-like proteins; Phosphatidylinositol kinase; Secretion
1. Introduction
The intracellular transport of hydrophobic molecules, suchas phospholipids, fatty acids, cholesterol, steroids and hydro-phobic vitamins between membranes and their enrichment atspecific cellular sites are critically dependent on the binding to
* Corresponding author. Present address: Vascular Biology Laboratory,
Office 621, JM-USDA Human Nutrition Research Center on Aging, Tufts
University, 711 Washington Street, Boston, MA 02111, USA. Tel: þ1 617 556
3229; fax: þ1 617 556 3224.
E-mail address: [email protected] (J.-M. Zingg).1 These authors contributed equally to this work.
0300-9084/$ - see front matter � 2008 Elsevier Masson SAS. All rights reserved.
doi:10.1016/j.biochi.2008.07.008
specific transport proteins (reviewed in Refs. [1e3]). For theintracellular transport of vitamin E (a-tocopherol), severalproteins such as the microsomal triglyceride transfer protein(MTTP), the a-tocopherol transfer protein (a-TTP) and thetocopherol associated proteins (TAPs) have been identified(reviewed in Refs. [4,5]). Whereas the function of liver a-TTPin a-tocopherol transport and enrichment of VLDL is welldescribed in vitro and in vivo [6], the role of the humantocopherol associated proteins (hTAPs) is less clear.
In humans, three highly homologous TAPs (hTAP1/2/3,or SEC14L2/3/4, respectively) have been described that arerelated to the Saccharomyces cerevisiae SEC14p protein [7e9].When compared to SEC14p and a-TTP, the three hTAPs carry

1704 J.-M. Zingg et al. / Biochimie 90 (2008) 1703e1715
an additional carboxy-terminal GOLD domain which in otherproteins (GCP60, PAP7) serves as adaptor for binding to Golgigiantin, or to the peripheral benzodiazepine receptor (PBR) (ortranslocator protein (TSPO) [10]), which is part of a complexknown as mitochondrial permeability transition pore (MPT)[9,11]. Deletion of the GOLD domain changes the intracellularlocalization of hTAPs [9], suggesting a possible role of thisdomain in targeting specific enzymes and cell compartmentssuch as Golgi or mitochondria.
hTAPs can bind the four natural tocopherol analogues (a-, b-, g-, d-tocopherol), the tocotrienols, as well as some derivativessuch as a-tocopheryl quinone, a-tocopheryl succinate and a-tocopheryl phosphate with different affinity [9,12e15], whatmay explain specific activities shown by each of these vitamin Eanalogues [16]. In support to a transport hypothesis, the cellularuptake of a-tocopherol and a-tocopheryl succinate is increasedby hTAP1 over-expression [15,17]. In addition to tocopherolanalogues and derivatives, these proteins bind in vitro severalother ligands, such as squalene, phosphatidylinositol, phos-phatidylinositol-3,4,5-phosphate, phosphatidylcholine andphosphatidylserine [7e9,12,15,17e20]. Tocopherols competewith these ligands, possibly affecting phospholipid-dependenttransport and signaling pathways [9,12,14,17].
Besides a role in direct lipid transport, the SEC14p-relatedproteins may play crucial roles in regulating exocytosis,vesicular transport and the relative composition of phospho-lipids in membranes (for reviews Refs. [1,21,22]). Intracellularlipid transport by TAPs may also influence signal transductionand gene expression, e.g. by modulating the activity of enzymesand transcription factors. In mice and humans, TAP1 interactsdirectly with phosphatidylinositol-3-kinase (PI3K) and modu-lates its activity in vitro and in vivo [9,17]. Moreover, a possibleinvolvement of mouse TAP1 in the modulation of geneexpression has been suggested [23]. Mouse TAP1 stimulates theexpression of the mast cell proteases genes mmcp-1 and mmcp-7, and the expression of TAP1 is up-regulated by activin/TGF-b treatment during differentiation of bone-marrow derived mastcells [24]. A role for TAPs in the biosynthesis of cholesterol wassuggested after the identification of TAP1/SEC14L2 as thepreviously described microsomal supernatant protein factor(SPF) [19]. hTAP1 is expressed in liver, whereas hTAP2/SEC14L3 or SPF2 is expressed in rat epithelial and respiratorytissues, including those from the olfactory bulb, trachea, lungand skin [25]. Both proteins increase squalene epoxidation,a reaction required for cholesterol synthesis, either by directlystimulating squalene transport to the enzyme or possibly byincreasing the formation or transport of vesicles carryingsqualene [19,26]. Data from TAP1/SPF-knockout mice suggestthat TAP1 maintains the steady-state plasma cholesterol levelduring fasting by compensating for decreased squalene epox-idase and HMG-CoA reductase activity [27]. Fasting is alsoassociated with reduced dietary vitamin E uptake, and elevatedde novo cholesterol synthesis may increase the requirements forvitamin E, suggesting that TAP1 up-regulation may play a dualrole in increasing cholesterol biosynthesis and in transportingand retaining sufficient vitamin E in tissues during conditions oflimited supply or increased consumption.
It is not yet clear whether the three hTAPs represent redun-dant proteins, or whether each of them performs specificfunctions, possibly as a result of their cell-type and tissue-specific distribution. In this study we have analyzed the a-tocopherol transport activity of the three recombinant hTAPs,and compared it with that of recombinant human a-TTP(hTTP). Moreover, to unravel the putative role of hTAPs indetermining the a-tocopherol concentration in peripheral cellsand tissues, the expression pattern of hTAP1, hTAP2 and hTAP3has been assessed in cell lines and human tissue samples, usingantibodies generated against each hTAP protein.
2. Materials and methods
2.1. Materials
RRR-a-tocopherol (Cognis) was dissolved in ethanol asa 50 mM stock solution and the concentration confirmed spec-trophotometrically. Radioactive D-a-S-methyl 14C-tocopherol(57 mCi/mmol) was purchased from AmershamPharmaciaBiotech and tested by Thin Layer Chromatography (TLC) onSIL G-25 TLC plates (Machery-Nagel) as previously described [9].
2.2. Cell lines and culture conditions
HMC-1 mastocytoma cells [28] and primary human umbil-ical cord blood-derived mast cells were kindly provided by Dr.M. Arock (Ecole Normale Superieure de Cachan, Cachan,CEDEX, France). HMC-1 cells were grown in Iscove’s MEM/10% FCS and 2 mM glutamine. Human umbilical cord blood-derived mast cells (CBMC) were produced by culture of CD34þ
progenitors purified from normal cord blood (CB). Briefly,CD34þ cells, purified from light-density CB cells witha specific immunoaffinity system (Miltenyi Biotech, Paris,France), were seeded at 105 cells/ml in a complete serum-freemedium (SFM) made by IMDM (Bio Whittaker, Verviers,Belgium); L-glutamine (Sigma), penicillin, streptomycin(Gibco, Cergy Pontoise, France), 1% BSA (Boehringer Man-nheim, Grenoble, France), insulin (10 mg/ml) (Sigma), trans-ferrin (5 mg/ml) (Sigma) and 100 ng/ml of recombinant humanstem cell factor (rhSCF; Amgen Inc, Thousand Oaks, CA). Atthe beginning of the culture, rhIL-3 (1 ng/ml) (Novartis, Basel,Switzerland), Flt-3 L (20 ng/ml) (R&D Systems Europe Ltd,Abingdon, UK) and thrombopoietin (2 ng/ml) (R&D SystemsEurope Ltd) were also added to induce expansion of hemato-poietic progenitors. After 7 days, cells were replaced incomplete SFM. The medium was then renewed every 7 days.After 5 weeks in culture, more than 99% of the cells wereidentified as MC by MayeGrunwaldeGiemsa staining ofcytocentrifuged samples. These pure populations of CBMCwere first primed with rhIL-4 (Novartis) at 20 ng/ml in thecomplete SFM and then suspended in the same medium sup-plemented with 2% of FBS (Dutscher, Brumath, France) beforeuse. H441 and A549 lung carcinoma cells were grown in RPMI/10% FCS and 2 mM glutamine. NCI-H295R human adreno-cortical carcinoma cells were grown in Iscove’s medium sup-plemented with 10% FCS. The remaining cell lines (Hela,

1705J.-M. Zingg et al. / Biochimie 90 (2008) 1703e1715
HepG2, LNCaP, HL-60, U937, Jurkat) were obtained fromATCC and grown as recommended.
2.3. Generation of polyclonal anti-hTAP antibodies
Affinity purified polyclonal anti-hTAP1/2/3 antibodieswere generated at Primm SRL (San Raffaele BiomedicalScience Park, Milano, Italy) and further characterized byELISA, Western Blotting and immunostaining. The specificityof the anti-hTAP antibodies was tested after expressing eachhTAP protein in E. coli BL21 Star (DE3) (Invitrogen) aspreviously described [9,13,14], and Western Blotting as indi-cated below.
2.4. Polymerase chain reaction
Reverse Transcription and Polymerase Chain Reaction (RT-PCR) for semi-quantitative detection of hTAP mRNA wasdone using primers and conditions previously described [9].
2.5. Western blotting
Immunoblots were done with 20 mg of extract per lane andseparated by 8% SDS-PAGE. The polyclonal rabbit anti-hTAPantibodies (1:300 diluted) and the monoclonal mouse anti-human b-actin (1:10,000 diluted) (Sigma) were used as primaryantibodies. Anti-mouse and anti-rabbit secondary antibodiescoupled to horseradish peroxidase (AmershamPharmaciaBiotech) were used for detection as previously described [9].
2.6. Immunohistochemistry
Human Normal Tissue Arrays (30 tissues, MaxArrays�(Zymed laboratories); 50 tissues, VastArrays�) were depar-affinizated and rehydrated as suggested by the manufacturer,blocked with 3% BSA/PBS and incubated with the anti-hTAPantibodies (1:100 diluted in 0.5% BSA/PBS) for 30 min. Thearrays were washed and stained with secondary antibodiesaccording to the Envision system kit (DAKO). The arrays werecounterstained with hematoxylin (DAKO), covered withultramount (DAKO), and analyzed for hTAP expression bymicroscopy.
2.7. Immunolocalization
Cells plated on glass slides (BD Falcon� CultureSlides)were washed with PBS and fixed with 4% paraformaldehyde/PBS, permeabilized with 0.1% saponin in PBS/0.5% BSA,blocked with 3% BSA/PBS for 30 min, and then stained witheach of the three anti-hTAP antibodies. Dilutions of antibodieswere: mouse anti-Golgi body antigen (Biogenesis) 1:200,mouse anti-CD63 (Biogenesis) 1:300, anti-hTAP1 1:50, anti-hTAP2 1:250, and mouse anti-giantin (Calbiochem) 1:200. Fordouble immunostaining, the permeabilized cells were firstincubated with organelle-specific antibodies and with thepolyclonal rabbit anti-hTAP antibodies, then with an AlexaFluor 594 labeled goat anti-mouse IgG (Hþ L) antibody
(Molecular Probes) and Alexa Fluor 488 goat anti-rabbit IgG(Hþ L) antibody (Molecular Probes). The mitochondria markerdye MitoTracker red was purchased from Molecular Probes.
2.8. Isolation of mitochondria from Hela cells
Isolation of mitochondria from cells in culture was per-formed essentially as described [29]. Briefly, 3� 108 Helacells were harvested by trypsinisation and washed two timeswith DMEM/10% FCS. Cells were then washed two timeswith isolation buffer (320 mM sucrose, 1 mM potassiumeEDTA, 10 mM TriseHCl, pH 7.4), and homogenized witha tight-fitting glass-Teflon homogenizer using 20e30 up anddown strokes at about 550 rpm. The samples were centrifugedat 1500�g at 4 �C for 10 min, the supernatant S1 was removedand homogenization and centrifugation was repeated with thepellet generating the S2 supernatant. S1 and S2 were thencombined and centrifuged at 17,000�g at 4 �C for 10 min.The pellet was dissolved in 1 ml of isolation buffer and usedimmediately. The identity of mitochondria was confirmed byWestern Blotting with anti-cytochrome c and anti-cytochromeoxidase subunit IV (COX4) antibodies (ApoAlert� CellFractionation kit, Clonetech).
2.9. Isolation of mitochondria from yeast
Overnight cultures (Saccharomyces cerevisiae strain CTY-MATa, ura3-52, Dhis 3-200, lys2-801) were centrifuged at1500�g for 5 min, washed with 10 mM EDTA, 1% b-mer-captoethanol, centrifuged again and resuspended in SE buffer(1 M sorbitol, 1 mM EDTA, pH 7.5e8). Lyticase was added ata concentration of 0.2 mg/g cells; suspension was incubatedfor 1 h at 30 �C and collected by centrifugation at 1500�g for10 min. The pellet (protoplasts) was resuspended in buffer SE,homogenized with a Dounce homogenizer and centrifuged at1500�g at 4 �C for 10 min, to remove nuclei and debris. Thepost nuclear fraction was centrifuged at 12,000�g at 4 �C, thesupernatant was removed and the pellet (mitochondria) wasresuspended again in SE buffer. Both centrifugation steps wererepeated. Mitochondria were resuspended in reaction buffer(320 mM sucrose, 1 mM potassiumeEDTA, 10 mM TriseHCl, pH 7.4) at a concentration of 0.1 mg of mitochondria/mland used directly for tocopherol transfer assays or stored at�80 �C.
2.10. Purification of recombinant hTAPs andhTTP from Escherichia coli
Recombinant hTAPs and hTTP were expressed and purifiedas previously described [9,13,14].
2.11. Mitochondrial tocopherol transfer assay
Isolated mitochondria (2 mg of wet weight) were incubatedat 37 �C with recombinant hTAPs or hTTP (4 mg/tube) in100 ml isolation buffer containing phosphatidylcholine lipo-somes, 20 mM cold a-tocopherol and 0.3 ml D-a-S-methyl

1706 J.-M. Zingg et al. / Biochimie 90 (2008) 1703e1715
14C-tocopherol (10 nmol/ml, 1.196 mCi) for 1 h with gentleshaking. Control reactions were carried out in the absence ofrecombinant proteins. The samples were centrifuged at16,000�g, washed twice with reaction buffer, and the distri-bution of radioactive tocopherol in pellet and supernatant wasquantified by scintillation counting. Enrichment of mito-chondria with 14C-a-tocopherol was corrected for background,expressed by spontaneous uptake of a-tocopherol in theabsence of recombinant proteins.
2.12. Mitochondrial binding assay
50 ml phosphatidylcholine (Sigma, dissolved in chloroformat 5 mg/ml) and 20 mM cold a-tocopherol were combined,dried under nitrogen, taken up in 200 ml buffer (50 mM TriseHCl, 50 mM KCl and 50 mM DTT, pH 7.5) and bath sonicatedfor 60 s. 20 ml of purified mitochondria, purified recombinanthTAPs, and 10 ml of phospholipid vesicles were assembled in100 ml of isolation buffer and incubated with shaking at 37 �Cfor 1 h. The samples were centrifuged at 16,000�g for 10 min,taken up in 100 ml of isolation buffer, and centrifuged again.Aliquots of pellet and supernatant were separated by 8% SDS-PAGE. Western Blotting was done as described above.
3. Results
3.1. Expression levels of hTAPs in HMC-1 mastocytomacells and H295R adrenocortical cells
In a previous survey, the three hTAP mRNAs were shownto be expressed at low level in several cancer cells [9]. Toidentify cell lines with higher levels of hTAP mRNAs, we
HMC-1
1 3
hTAP
H29
5R
HM
C-1
H295R
GAPDH
1hTAP
anti-hTAP1
anti-hTAP2
anti-hTAP3
Coomassie
A B
2
Fig. 1. Expression of the hTAPs mRNA and proteins in H295R and HMC-1 cells. (
PCR. The expression of the GAPDH gene was used as internal control. (B) Sp
expressing recombinant hTAP1, hTAP2 or hTAP3 or empty control were prepared,
as indicated in Section 2. Bottom: Coomassie stained gel run in parallel. (C) Imm
vesicular staining in both cell lines.
screened a number of further cancer cell lines; again, most ofthem expressed relatively low levels of hTAP mRNAs (datanot shown). However, two cell lines appeared to expresshigher levels of hTAP1 mRNA, the HMC-1 mastocytoma andthe H295R human adrenocortical carcinoma cells. In line withthe results of Ikeda et al. [30], HMC-1 expressed mainlyhTAP1 (Fig. 1A). H295R expressed about equal amounts ofhTAP1 and hTAP2 mRNAs, whereas hTAP3 was hardlydetectable (Fig. 1A). The relatively high expression of hTAP1mRNA in HMC-1 and H295R cells suggests the involvementof hTAP1 in the secretory aspects of these cells, possiblyrelated to the formation, transport and sub-cellular localizationof specific lipids involved in the vesicular storage and releaseof mediators (histamine, cytokines or proteases such as chy-mase or tryptase) in mast cells, or of steroid hormones inadrenocortical cells [31]. This would represent a conservedfunction of the TAP proteins similar to the one of sec14p inmaintaining and regulating Golgi-derived vesicle flow inSaccharomyces cerevisiae (reviewed in Refs. [1,22,32]).
To further investigate the expression of hTAPs in mast andadrenal cells, specific antibodies were generated againsthTAP1, hTAP2 and hTAP3. Their specificity was confirmedusing the recombinant hTAP proteins and Western Blotting(Fig. 1B). H295R cells expressed relatively high amounts ofhTAP1, as assessed by Western Blotting (Table 1). HMC-1cells expressed robust amounts of hTAP1 compared to theother tested cell lines; hTAP2 was expressed at various levels,and hTAP3 was not detectable in any of the tested cell lines(Table 1). Immunofluorescence indicated that hTAP1 islocated to cytoplasmic granules in both HMC-1 and H295Rcells (Fig. 1C), further supporting a role in the secretoryfunctions of these cells.
H295R
HMC-1
3 -2
C
A) Expression of the three hTAP mRNAs as assessed by semi-quantitative RT-
ecificity of polyclonal rabbit anti-hTAP antibodies. Escherichia coli extracts
separated and probed with anti-hTAP1, anti-hTAP2 and anti-hTAP3 antibodies
unostaining of HMC-1 and H295R cells with anti-hTAP1 antibodies shows

Table 1
Expression level of the three hTAP proteins in different cell lines as analyzed
by Western blots
Cell line TAP1 TAP2 TAP3
A549 (lung carcinoma) þ þþ �HepG2 (liver carcinoma) þþ þ/� �Hela (cervix carcinoma) þ þþ �HMC-1 (mastocytoma) þþ þþ �LNCaP (prostate carcinoma) þ � �H295R (adrenocortical cells) þþ þþ �Primary mast cells (differentiated by
interleukin 4(IL-4))
þþ þþ �
HL-60 (monocyte-like) � þ �HL-60/ cis-Retinoic acid (differentiated
to macrophage-like cells)
þ þþ �
U937 (monocyte-like) þ þ/� �� no expression detectable, þ/� weak expression, þþ clearly detectable
band.
1707J.-M. Zingg et al. / Biochimie 90 (2008) 1703e1715
3.2. hTAP1 is induced by IL-4 during differentiation ofprimary human mast cells
To investigate whether the relatively high expression ofhTAP1 in HMC-1 mastocytoma cells reflects their state ofexpression in normal mast cells, primary umbilical cord blood-derived mast cells were used to analyze the expression of thethree hTAP proteins. To generate fully differentiated mast cells,primary human mast cells were treated with interleukin 4 (IL-4). This cytokine is known to repress c-kit, tryptase and Fc3RIaexpression [33,34], to increase the percentage of chymase andtryptase double-positive mast cells and their morphologicalmaturation [35], to induce apoptosis [36], and to stimulate mastcell mediator release [37,38]. Interestingly, whereas primarymast cells expressed relatively little hTAP1, the expression wasincreased up to 6 fold upon treatment with IL-4 (Fig. 2). Co-treatment with a-tocopherol did not change the expression levelof hTAP1 (Fig. 2). hTAP2 expression was very low and hTAP3
Fig. 2. Expression of hTAP1 during primary human mast cells differentiation.
(A) Primary human mast cells were differentiated with interleukin (IL-4,
20 ng/ml) for 24 h, without treatment (c), in the presence of 0.1% ethanol
(solvent control, E), or 50 mM a-tocopherol (aT). The results show up to 6-
fold induction of hTAP1 protein expression by IL-4, and ethanol or a-
tocopherol had no effect. Shown is a representative blot of two experiments.
not detectable on Western blots in both undifferentiated and IL-4 differentiated mast cells (data not shown).
3.3. Association of recombinant hTAP with Golgi andmitochondria
A previous study reported the localization of hemagglutinin-tagged hTAPs to the Golgi, the endoplasmic reticulum (ER) andthe plasma membrane [9]. In order to transport ligands betweendifferent membranes, TAPs are presumed to be able to transientlybind to different membranes, and it may depend on the cell type,activation state and stage within the cell cycle, to which or fromwhich membrane system a given ligand is transported. Specificorganelles may have docking sites/receptors that attract TAPsenabling the more efficient transport of the TAP-ligands. Here,double-immunostaining with anti-hTAP1 and organelle-specificantibodies/markers shows that the localization of endogenoushTAP1 partially overlaps with Golgi giantin in HMC-1 cells,with Golgi marker in A549 lung adenocarcinoma cells and alsowith mitotracker red in Hela and H295R cells (Fig. 3). Onlypartial overlap was seen with the tetraspanin protein CD63 inHMC-1 cells (Fig. 3), indicating a certain selectivity for mem-brane association of hTAP1. Whereas the location of hTAP1 tothe Golgi apparatus and vesicles in mast cells may be similar tothe role of Saccharomyces cerevisae sec14p in vesicle flow andsecretion from the Golgi, the association with mitochondria mayreflect a specific function in maintaining transport of specificproducts (e.g. vitamin E analogues, squalene, phospholipids,steroids) to and from mitochondria in these cells.
3.4. Binding of recombinant hTAP1 to isolatedmitochondria
To assess the ability of hTAP1 to interact with mitochondria,recombinant hTAP1 was incubated with mitochondria isolatedfrom Hela cells, Jurkat cells or Saccharomyces cerevisiae. Aftercentrifugation, the amount of mitochondria-bound hTAP1 in thepellet was determined by Western Blotting on extracts madefrom supernatant (S) and pellet (P). hTAP1 was best bound bymitochondria isolated from Hela cells, whereas mitochondriafrom Jurkat and Saccharomyces cerevisiae showed less bindingcapacity (Fig. 4A). Control reactions in the absence of mito-chondria showed that the binding assay conditions did not leadto precipitation of recombinant hTAP1 (Fig. 4B). Lowerbinding of hTAP1 to Jurkat or yeast mitochondria may reflectthe low/mutant or absent expression of PBR in these cells [39].PBR is part of the mitochondrial permeability transition pore(MPT) involved in the transport of molecules into and out ofmitochondria. In mast cells, PBR plays a role in releasingseveral mediators [40,41], and the hTAPs may be targeted to itthrough their GOLD domains [9,11,39].
3.5. Transport of a-tocopherol to mitochondria isstimulated by the recombinant hTAP and hTTP
To test the possible involvement of the three hTAPs in thetransport of a-tocopherol to mitochondria, we performed an in

-
-
Fig. 3. Localization of hTAP1 in cell lines. Double-immunostaining of HMC-1 mastocytoma cells, A549 lung cells, H295R adrenocortical and Hela cells. Left
panels: green staining with anti-hTAP1 antibody; middle panels: staining with anti-giantin antibodies, anti-Golgi antibodies, anti-CD63 antibodies, or with
Mitotracker red (as indicated); right panels: overlays are shown in yellow. (For interpretation of the references to colour in figure legends, the reader is refered to
the web version of this article)
1708 J.-M. Zingg et al. / Biochimie 90 (2008) 1703e1715
vitro a-tocopherol transfer assay [42,43]. Mitochondria isolatedfrom Hela cells and Saccharomyces cerevisiae were incubatedfor 1 h with 14C-a-tocopherol, in the presence of recombinanthTAPs or human a-TTP (hTTP). The mitochondrial uptake of
radioactive a-tocopherol, quantified by scintillation counting,was highest with hTTP; hTAP1 and hTAP2 also increased theuptake, whereas hTAP3 gave only a weak increase (Fig. 4C).These results suggest that hTAP1 and hTAP2 may be involved
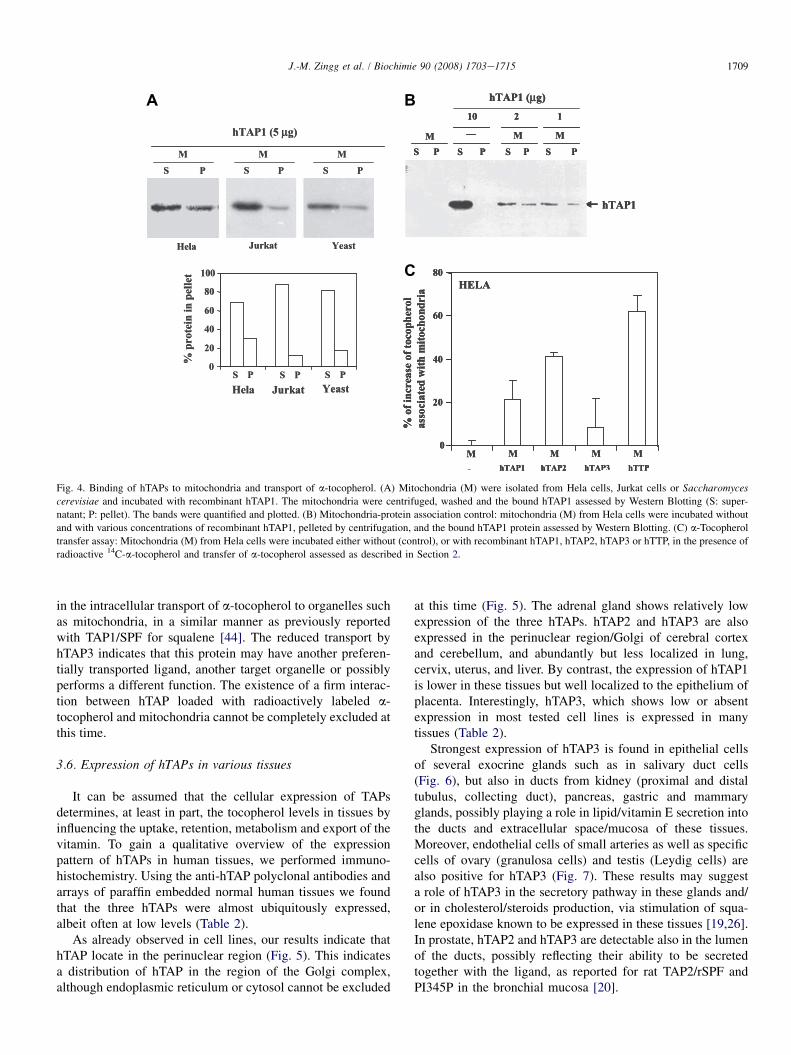
A B
C
Fig. 4. Binding of hTAPs to mitochondria and transport of a-tocopherol. (A) Mitochondria (M) were isolated from Hela cells, Jurkat cells or Saccharomyces
cerevisiae and incubated with recombinant hTAP1. The mitochondria were centrifuged, washed and the bound hTAP1 assessed by Western Blotting (S: super-
natant; P: pellet). The bands were quantified and plotted. (B) Mitochondria-protein association control: mitochondria (M) from Hela cells were incubated without
and with various concentrations of recombinant hTAP1, pelleted by centrifugation, and the bound hTAP1 protein assessed by Western Blotting. (C) a-Tocopherol
transfer assay: Mitochondria (M) from Hela cells were incubated either without (control), or with recombinant hTAP1, hTAP2, hTAP3 or hTTP, in the presence of
radioactive 14C-a-tocopherol and transfer of a-tocopherol assessed as described in Section 2.
1709J.-M. Zingg et al. / Biochimie 90 (2008) 1703e1715
in the intracellular transport of a-tocopherol to organelles suchas mitochondria, in a similar manner as previously reportedwith TAP1/SPF for squalene [44]. The reduced transport byhTAP3 indicates that this protein may have another preferen-tially transported ligand, another target organelle or possiblyperforms a different function. The existence of a firm interac-tion between hTAP loaded with radioactively labeled a-tocopherol and mitochondria cannot be completely excluded atthis time.
3.6. Expression of hTAPs in various tissues
It can be assumed that the cellular expression of TAPsdetermines, at least in part, the tocopherol levels in tissues byinfluencing the uptake, retention, metabolism and export of thevitamin. To gain a qualitative overview of the expressionpattern of hTAPs in human tissues, we performed immuno-histochemistry. Using the anti-hTAP polyclonal antibodies andarrays of paraffin embedded normal human tissues we foundthat the three hTAPs were almost ubiquitously expressed,albeit often at low levels (Table 2).
As already observed in cell lines, our results indicate thathTAP locate in the perinuclear region (Fig. 5). This indicatesa distribution of hTAP in the region of the Golgi complex,although endoplasmic reticulum or cytosol cannot be excluded
at this time (Fig. 5). The adrenal gland shows relatively lowexpression of the three hTAPs. hTAP2 and hTAP3 are alsoexpressed in the perinuclear region/Golgi of cerebral cortexand cerebellum, and abundantly but less localized in lung,cervix, uterus, and liver. By contrast, the expression of hTAP1is lower in these tissues but well localized to the epithelium ofplacenta. Interestingly, hTAP3, which shows low or absentexpression in most tested cell lines is expressed in manytissues (Table 2).
Strongest expression of hTAP3 is found in epithelial cellsof several exocrine glands such as in salivary duct cells(Fig. 6), but also in ducts from kidney (proximal and distaltubulus, collecting duct), pancreas, gastric and mammaryglands, possibly playing a role in lipid/vitamin E secretion intothe ducts and extracellular space/mucosa of these tissues.Moreover, endothelial cells of small arteries as well as specificcells of ovary (granulosa cells) and testis (Leydig cells) arealso positive for hTAP3 (Fig. 7). These results may suggesta role of hTAP3 in the secretory pathway in these glands and/or in cholesterol/steroids production, via stimulation of squa-lene epoxidase known to be expressed in these tissues [19,26].In prostate, hTAP2 and hTAP3 are detectable also in the lumenof the ducts, possibly reflecting their ability to be secretedtogether with the ligand, as reported for rat TAP2/rSPF andPI345P in the bronchial mucosa [20].

Table 2
Expression level of the three TAP proteins in human tissues as measured by
immunohistochemistry
Tissue TAP1 TAP2 TAP3
Adrenal gland þþ þþ þþþBone marrow þþ þþ þþBreast þþ þþ þþþCerebellum þ þþ þþCerebral cortex þ þþ þCervix þ þ þþColon þ þþ þþþEsophagus þþ þ þþþHeart muscle þ þþþ þþKidney þ þþ þþLiver þ þþþ þþþLung þ þþ þþþOmentum � þ þOvary þ þþ þþþPancreas þ þþ þþþParathyroid gland þ þ þþþPeripheral nerve þ þ þþPituitary gland þ þþþ þþþPlacenta þþ þþ þþProstate gland þþ þþ þþSalivary gland þ þþ þþþSkeletal muscle þþ þþ þþþSkin þ þþþ þþþSmall intestine þþ þ þþþSpleen þ þ þStomach � þþ þþTestis þþ þþ þThymus gland þ þ �Thyroid gland þ þ þTonsil þ þþ þþUterus � þ þþExpression level: � without, þweak, þþ middle, þþþ high.
1710 J.-M. Zingg et al. / Biochimie 90 (2008) 1703e1715
4. Discussion and conclusion
Whereas recent research has clarified the uptake andtransport of a-tocopherol from the diet, the enrichment inplasma by a-TTP, as well as the transport across the bloodbrain barrier (reviewed in Ref. [5]), less is known about thetransport within peripheral tissues and in particular within oracross cells. In neurons, a-tocopherol is concentrated at thesomiteeneurite junction suggesting specific intracellularmechanisms of enrichment [45]. Since the tocopherolconcentration is highest in mitochondria, Golgi, endoplasmicreticulum and lysosomes [46], intracellular transport mecha-nisms to enrich a-tocopherol in the membranes of theseorganelles must exist. In mitochondria, a-tocopherol concen-trations are higher in the inner membrane than in the outermembrane [47], and tocopherol transport proteins may main-tain an optimal level of mitochondrial a-tocopherol and a-tocopheryl quinone required for the optimal function of thisorganelle [48e51].
Using an in vitro a-tocopherol transfer assay, we find thathTAPs increase the localization of a-tocopherol to mito-chondria. Transport proteins that are able to enrich tocopherolin organelles could thus favor their optimal functional activity,in particular in organelles able to generate reactive oxygen
species (ROS) such as mitochondria. This function cannot befulfilled in all tissues by a-TTP since this protein is expressedmainly in the liver, whereas hTAPs are expressed in manyperipheral tissues. Increased mitochondria-mediated apoptosisinduced by a-tocopheryl succinate in hTAP1-overexpressingmesothelioma cells suggests that hTAP1 plays a role inintracellular transport of vitamin E analogues and derivativesto specific sites at mitochondria, such as Bcl-xL/Bcl-2 orsuccinate oxidase [15,17,52e54]. A relatively high level ofhTAP1 in cytoplasmic granuli of mast cells may indicate theincreased requirements of mast cells for vitamin E transport orsuggests a role of hTAP1 in the endocytic/secretory pathwayor in degranulation. Moreover, since TAP1 binds to andregulates phosphatidylinositol-3-kinase (PI3K) [9,17], a keyprotein required for IgE-driven activation of mast cells, hTAP1may influence signal transduction in these cells, possibly ina tocopherol analogue specific manner (reviewed in Ref. [55]).TAPs-dependent transport of different lipids and their mutualexchange may facilitate secretory processes by potentiatingPI4K and subsequent vesicle formation [56]; thus, the trans-port and presentation of lipids by TAPs to phosphatidylinositolkinases may link lipid-dependent signal transduction with theformation of vesicles and secretion. Modulation of PI3K byTAPs may also affect gene expression in a tocopherol-specificmanner [23,24], e.g. by affecting the PI3K/PKB/PPARg signaltransduction pathway [57]; however, in preliminary experi-ments (data not shown), over-expression of hTAP1 did notaffect PKB phosphorylation or a PPARg-responsive reportergene [58] in a tocopherol-specific manner.
It can be assumed that the expression level of intracellulara-tocopherol transport proteins may determine in part thetocopherol abundance in peripheral tissues, e.g. by determiningthe cellular uptake, retention, metabolism and export oftocopherols. Among the tested human tissues, the three hTAPsare expressed at different levels and also in different cell typeswithin the same tissue. Most striking was the high expression ofhTAP3 in the epithelial duct cells of different glands. Thissuggests that hTAP3 may play a role in maintaining or regu-lating the (possibly polarized) lipid/vitamin E and vesicle flowwithin or across these secretory cells, a function related to theSaccharomyces cerevisiae sec14p protein, which maintains thesecretory vesicle flow from the late Golgi compartment [59].The expression of hTAP3 in epithelial duct cells of mammary orsalivary gland may increase the level of tocopherols in breastmilk sustaining the newborn (reviewed in Ref. [60]), and insaliva possibly preventing ulceration and oral cancers [61,62],respectively. The high expression of hTAP3 in endothelial cellsof small arteries seen in ovary may reflect the higher demand fora-tocopherol in this tissue [63], as a protective agent e.g. againstethanol-induced changes in reproductive tissues [64]. Thepresence of hTAP3 in granulosa cells of ovary and Leydig cellsof testis may be important for the function of a-tocopherol infertility; moreover, since these cells express also squaleneepoxidase, hTAP3 may exert a similar role in cholesterol andsubsequent steroid hormone synthesis as described for hTAP1in liver tissues [19]. The absence of symptoms in mouse TAP1knockout mice suggests that this protein may exert its main

Fig. 5. Comparison of the expression pattern of the three hTAP proteins in human tissue arrays. Immunohistochemistry of tissues-sections show clear expression
differences between the three hTAP proteins. The nuclei are stained in blue with hematoxylin, arrows indicate localized expression of the hTAP proteins (For
interpretation of the references to colour in figure legends, the reader is refered to the web version of this article).
1711J.-M. Zingg et al. / Biochimie 90 (2008) 1703e1715
function under certain physiological conditions such as fooddeprivation [27], supported by the relatively low staining seen inmany tissues. On the other hand, the expression of hTAP3 intissues known to be strongest affected by vitamin E deficiency
in animals, such as brain, ovary, testes, liver, kidney and skeletalmuscle [65], may indicate an important function of hTAP3 intissue distribution of vitamin E and/or in mediating its biolog-ical effects.
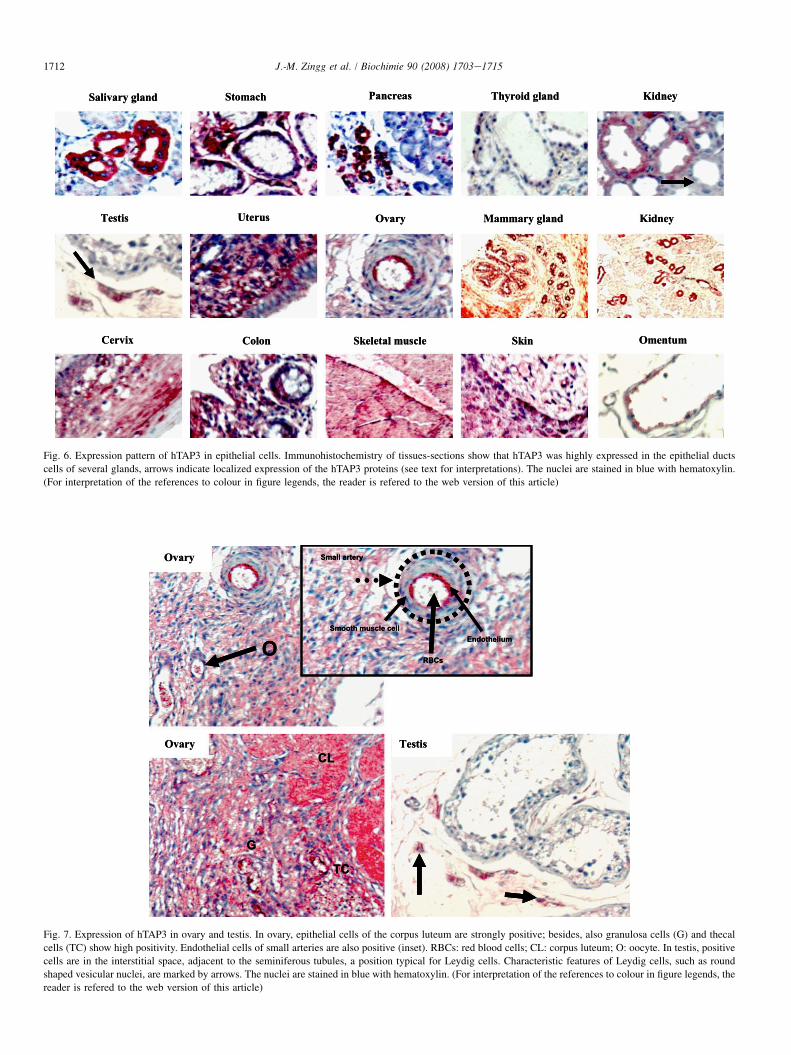
Fig. 6. Expression pattern of hTAP3 in epithelial cells. Immunohistochemistry of tissues-sections show that hTAP3 was highly expressed in the epithelial ducts
cells of several glands, arrows indicate localized expression of the hTAP3 proteins (see text for interpretations). The nuclei are stained in blue with hematoxylin.
(For interpretation of the references to colour in figure legends, the reader is refered to the web version of this article)
Fig. 7. Expression of hTAP3 in ovary and testis. In ovary, epithelial cells of the corpus luteum are strongly positive; besides, also granulosa cells (G) and thecal
cells (TC) show high positivity. Endothelial cells of small arteries are also positive (inset). RBCs: red blood cells; CL: corpus luteum; O: oocyte. In testis, positive
cells are in the interstitial space, adjacent to the seminiferous tubules, a position typical for Leydig cells. Characteristic features of Leydig cells, such as round
shaped vesicular nuclei, are marked by arrows. The nuclei are stained in blue with hematoxylin. (For interpretation of the references to colour in figure legends, the
reader is refered to the web version of this article)
1712 J.-M. Zingg et al. / Biochimie 90 (2008) 1703e1715

1713J.-M. Zingg et al. / Biochimie 90 (2008) 1703e1715
The expression of hTAP2 and hTAP3 in lung, cervix anduterus may reflect their possible involvement in the secretionof a-tocopherol with mucus, as reported for rat SPF2/hTAP2[20]. Expression of hTAPs in brain (cerebellum and cortex)may play a role in the intracellular enrichment of vitamin E atspecific sub-cellular sites [45], what may in turn explain itsessential function for the nervous system or mediate itspreventive effects against several neuro-degenerative diseases.Alternatively, since the brain synthesizes almost all of itscholesterol in situ [66], hTAP could play an important role inbrain cholesterol as well as neurosteroids homeostasis viamodulating squalene epoxidase [19,26]. Interestingly, in someorgans such as the kidney, not all the ducts express hTAP3(Fig. 6), indicating that hTAP3 does not determine theepithelial duct structures, but rather modulates the functionalaspect of these cells. Moreover, as reported for rat TAP2/rSPF2 in the bronchial mucosa [20], the presence of hTAPproteins in the lumen of the prostate may reflect their ability tobe secreted together with the ligand (e.g. in exosomes or asprostasomes) which may then protect against oxidation ormodulate the function of spermatozoa and influence fertility.
The possible reasons for the absence of hTAP3 expressionin most cell lines also require further analysis. As far one cantell for the tested cancer cell lines, none of them was directlyderived from epithelia. The absence of hTAP3 expression incell lines could be explained by the need for local cues onlypresent in intact tissues, or that hTAP3 somehow interfereswith proliferation required for cell culture expansion and issilenced by DNA hyper-methylation. In fact, results suggestthat hTAP3 mRNA expression can be increased by treatmentwith the demethylating agent 5-azacytidine (data not shown).Accordingly, our hypothesis is consistent with the observationthat proliferation of prostate cancer cells is associated withreduced expression of hTAP1 [17,67], and it remains to bedetermined whether similar changes occur with the threehTAP proteins and in other epithelial-derived tumors.
The affinity constants measured for hTAP1 indicate that it isable to bind several other ligands, and the results shown herewith a-tocopherol may reflect one of the possible functions ofhTAPs [12,68]. In particular, the transport of squalene to/fromsub-cellular sites such as mitochondria may play a similarprotective function as postulated for vitamin E [44,69]. In vivo,the three hTAPs could represent versatile lipid shuttle proteinsor ‘‘regulatory lipid chaperones’’ that play multiple roles in cell-type specific intracellular lipid localization, dynamics andhomeostasis by transporting hydrophobic ligands across thehydrophilic cytosol to enzymes, transcription factors, receptors,organelles, to lipid domains at the plasma membrane, or even toextracellular sites such as exosomes.
The findings that the hTAP proteins can enrich cells andmitochondria with a-tocopherol could be relevant for thepreventive effects of vitamin E described against severaldegenerative diseases, ranging from cardiomyopathy to Alz-heimer’s and Parkinson’s disease, and several ataxias withunknown etiologies. These disorders share common signs ofmitochondrial impairment, a condition that may become worsewhen the vitamin E levels are reduced [70e73]. The hTAP
proteins may play a role in these diseases by affecting vitaminE transport to sub-cellular sites in a tissue-specific manner.
Acknowledgments
This study was supported by the Swiss National ScienceFoundation and the Foundation for Nutrition Research inSwitzerland. We thank M. Feher for excellent technicalassistance.
References
[1] C.J. Mousley, K.R. Tyeryar, P. Vincent-Pope, V.A. Bankaitis, The Sec14-
superfamily and the regulatory interface between phospholipid metabo-
lism and membrane trafficking, Biochim. Biophys. Acta 1771 (2007)
727e736.
[2] S. Cockcroft, Trafficking of phosphatidylinositol by phosphatidylinositol
transfer proteins, Biochem. Soc. Symp. (2007) 259e271.
[3] M.A. De Matteis, A. Di Campli, G. D’Angelo, Lipid-transfer proteins in
membrane trafficking at the golgi complex, Biochim. Biophys. Acta 1771
(2007) 761e768.
[4] D.E. Kaempf-Rotzoll, M.G. Traber, H. Arai, Vitamin E and transfer
proteins, Curr. Opin. Lipidol. 14 (2003) 249e254.
[5] A. Rigotti, Absorption, transport, and tissue delivery of vitamin E, Mol.
Aspect. Med. 28 (2007) 423e436.
[6] A. Hentati, H.X. Deng, W.Y. Hung, M. Nayer, M.S. Ahmed, X. He,
R. Tim, D.A. Stumpf, T. Siddique, Ahmed, Human alpha-tocopherol
transfer protein: gene structure and mutations in familial vitamin E
deficiency, Ann. Neurol. 39 (1996) 295e300.
[7] M.I. Merkulova, S.G. Andreeva, T.M. Shuvaeva, S.V. Novoselov,
I.V. Peshenko, M.F. Bystrova, V.I. Novoselov, E.E. Fesenko,
V.M. Lipkin, A novel 45 kDa secretory protein from rat olfactory
epithelium: primary structure and localisation, FEBS Lett. 450 (1999)
126e130.
[8] S. Zimmer, A. Stocker, M.N. Sarbolouki, S.E. Spycher, J. Sassoon,
A. Azzi, A novel human tocopherol-associated protein: cloning, in vitro
expression, and characterization, J. Biol. Chem. 275 (2000) 25672e
25680.
[9] P. Kempna, J.M. Zingg, R. Ricciarelli, M. Hierl, S. Saxena, A. Azzi,
Cloning of novel human SEC14p-like proteins: cellular localization,
ligand binding and functional properties, Free Radic. Biol. Med. 34
(2003) 1458e1472.
[10] V. Papadopoulos, M. Baraldi, T.R. Guilarte, T.B. Knudsen, J.J. Lacapere,
P. Lindemann, M.D. Norenberg, D. Nutt, A. Weizman, M.R. Zhang,
M. Gavish, Translocator protein (18 kDa): new nomenclature for the
peripheral-type benzodiazepine receptor based on its structure and
molecular function, Trends Pharmacol. Sci. 27 (2006) 402e409.
[11] V. Anantharaman, L. Aravind, The GOLD domain, a novel protein
module involved in golgi function and secretion, Genome Biol. 3 (2002)
research0023.
[12] C. Panagabko, S. Morley, M. Hernandez, P. Cassolato, H. Gordon,
R. Parsons, D. Manor, J. Atkinson, Ligand specificity in the CRAL-TRIO
protein family, Biochemistry 42 (2003) 6467e6474.
[13] P. Kempna, R. Cipollone, L. Villacorta, R. Ricciarelli, J.M. Zingg,
Isoelectric point mobility shift assay for rapid screening of charged and
uncharged ligands bound to proteins, IUBMB Life 55 (2003) 103e107.
[14] D. Habermehl, P. Kempna, A. Azzi, J.M. Zingg, Recombinant SEC14-
like proteins (TAP) possess GTPase activity, Biochem. Biophys. Res.
Commun. 326 (2005) 254e259.
[15] J. Neuzil, L.F. Dong, X.F. Wang, J.M. Zingg, Tocopherol-associated
protein-1 accelerates apoptosis induced by alpha-tocopheryl succinate in
mesothelioma cells, Biochem. Biophys. Res. Commun. 343 (2006)
1113e1117.
[16] A. Azzi, R. Ricciarelli, J.M. Zingg, Non-antioxidant molecular functions
of alpha-tocopherol (vitamin E), FEBS Lett. 519 (2002) 8e10.

1714 J.-M. Zingg et al. / Biochimie 90 (2008) 1703e1715
[17] J. Ni, X. Wen, J. Yao, H.C. Chang, Y. Yin, M. Zhang, S. Xie, M. Chen,
B. Simons, P. Chang, A. di Sant’agnese, E.M. Messing, S. Yeh,
Tocopherol-associated protein suppresses prostate cancer cell growth by
inhibition of the phosphoinositide 3-kinase pathway, Cancer. Res. 65
(2005) 9807e9816.
[18] I.W. Caras, E.J. Friedlander, K. Bloch, Interactions of supernatant protein
factor with components of the microsomal squalene epoxidase system.
Binding of supernatant protein factor to anionic phospholipids, J. Biol.
Chem. 255 (1980) 3575e3580.
[19] N. Shibata, M. Arita, Y. Misaki, N. Dohmae, K. Takio, T. Ono, K. Inoue,
H. Arai, Supernatant protein factor, which stimulates the conversion of
squalene to lanosterol, is a cytosolic squalene transfer protein and
enhances cholesterol biosynthesis, Proc. Natl. Acad. Sci. U.S.A. 98
(2001) 2244e2249.
[20] M. Merkulova, H. Huynh, V. Radchenko, K. Saito, V. Lipkin,
T. Shuvaeva, T. Mustelin, Secretion of the mammalian Sec14p-like
phosphoinositide-binding p45 protein, FEBS J. 272 (2005) 5595e5605.
[21] S.E. Phillips, K.E. Ile, M. Boukhelifa, R.P. Huijbregts, V.A. Bankaitis,
Specific and nonspecific membrane-binding determinants cooperate in
targeting phosphatidylinositol transfer protein beta-isoform to the
mammalian trans-golgi network, Mol. Biol. Cell. 17 (2006) 2498e2512.
[22] K. Saito, L. Tautz, T. Mustelin, The lipid-binding SEC14 domain, Bio-
chim. Biophys. Acta 1771 (2007) 719e726.
[23] J. Yamauchi, T. Iwamoto, S. Kida, S. Masushige, K. Yamada, T. Esashi,
Tocopherol-associated protein is a ligand-dependent transcriptional
activator, Biochem. Biophys. Res. Commun. 285 (2001) 295e299.
[24] M. Funaba, M. Murakami, T. Ikeda, K. Ogawa, K. Tsuchida, H. Sugino,
Identification of tocopherol-associated protein as an activin/TGF-
beta-inducible gene in mast cells, Biochim. Biophys. Acta 1763 (2006)
900e906.
[25] V. Mokashi, D.K. Singh, T.D. Porter, Rat supernatant protein factor-like
protein stimulates squalene monooxygenase and is activated by protein
kinase A, Biochem. Biophys. Res. Commun. 316 (2004) 688e692.
[26] V. Mokashi, D.K. Singh, T.D. Porter, Supernatant protein factor stimu-
lates HMG-CoA reductase in cell culture and in vitro, Arch. Biochem.
Biophys. 433 (2005) 474e480.
[27] N. Shibata, K. Jishage, M. Arita, M. Watanabe, Y. Kawase,
K. Nishikawa, Y. Natori, H. Inoue, H. Shimano, N. Yamada,
M. Tsujimoto, H. Arai, Regulation of hepatic cholesterol synthesis by
a novel protein (SPF) that accelerates cholesterol biosynthesis, FASEB J.
20 (2006) 2642e2644.
[28] J.H. Butterfield, D. Weiler, G. Dewald, G.J. Gleich, Establishment of an
immature mast cell line from a patient with mast cell leukemia, Leuk.
Res. 12 (1988) 345e355.
[29] A. Almeida, J.M. Medina, A rapid method for the isolation of meta-
bolically active mitochondria from rat neurons and astrocytes in primary
culture, Brain. Res. Brain. Res. Protoc. 2 (1998) 209e214.
[30] T. Ikeda, M. Murakami, M. Funaba, Expression of tocopherol-associated
protein in mast cells, Clin. Diagn. Lab. Immunol. 11 (2004) 1189e1191.
[31] M. Hecker, J.L. Newsted, M.B. Murphy, E.B. Higley, P.D. Jones, R. Wu,
J.P. Giesy, Human adrenocarcinoma (H295R) cells for rapid in vitro
determination of effects on steroidogenesis: hormone production, Tox-
icol. Appl. Pharmacol. 217 (2006) 114e124.
[32] C.J. Mousley, K.R. Tyeryar, M.M. Ryan, V.A. Bankaitis, Sec14p-like
proteins regulate phosphoinositide homoeostasis and intracellular protein
and lipid trafficking in yeast, Biochem. Soc. Trans. 34 (2006) 346e350.
[33] C. Sillaber, H. Strobl, D. Bevec, L.K. Ashman, J.H. Butterfield,
K. Lechner, D. Maurer, P. Bettelheim, P. Valent, IL-4 regulates c-kit
proto-oncogene product expression in human mast and myeloid
progenitor cells, J. Immunol. 147 (1991) 4224e4228.
[34] F. Thienemann, B.M. Henz, M. Babina, Regulation of mast cell char-
acteristics by cytokines: divergent effects of interleukin-4 on immature
mast cell lines versus mature human skin mast cells, Arch. Dermatol.
Res. 296 (2004) 134e138.
[35] H. Toru, M. Eguchi, R. Matsumoto, M. Yanagida, J. Yata, T. Nakahata,
Interleukin-4 promotes the development of tryptase and chymase double-
positive human mast cells accompanied by cell maturation, Blood 91
(1998) 187e195.
[36] C.A. Oskeritzian, W. Zhao, A.L. Pozez, N.M. Cohen, M. Grimes,
L.B. Schwartz, Neutralizing endogenous IL-6 renders mast cells of the
MCT type from lung, but not the MCTC type from skin and lung,
susceptible to human recombinant IL-4-induced apoptosis, J. Immunol.
172 (2004) 593e600.
[37] J.W. Coleman, M.R. Holliday, I. Kimber, K.M. Zsebo, S.J. Galli,
Regulation of mouse peritoneal mast cell secretory function by stem cell
factor, IL-3 or IL-4, J. Immunol. 150 (1993) 556e562.
[38] K. Matsuda, A.M. Piliponsky, M. Iikura, S. Nakae, E.W. Wang,
S.M. Dutta, T. Kawakami, M. Tsai, S.J. Galli, Monomeric IgE enhances
human mast cell chemokine production: IL-4 augments and dexameth-
asone suppresses the response, J. Allergy. Clin. Immunol. 116 (2005)
1357e1363.
[39] B. Costa, A. Salvetti, L. Rossi, F. Spinetti, A. Lena, B. Chelli,
M. Rechichi, E. Da Pozzo, V. Gremigni, C. Martini, Peripheral benzo-
diazepine receptor: characterization in human T-lymphoma Jurkat cells,
Mol. Pharmacol. 69 (2006) 37e44.
[40] M. Bidri, B. Royer, G. Averlant, G. Bismuth, J.J. Guillosson, M. Arock,
Inhibition of mouse mast cell proliferation and proinflammatory mediator
release by benzodiazepines, Immunopharmacology 43 (1999) 75e86.
[41] L. Veenman, M. Gavish, The peripheral-type benzodiazepine receptor
and the cardiovascular system. Implications for drug development,
Pharmacol. Ther. 110 (2006) 503e524.
[42] A.K. Dutta-Roy, D.J. Leishman, M.J. Gordon, F.M. Campbell,
G.G. Duthie, Identification of a low molecular mass (14.2 kDa) alpha-
tocopherol-binding protein in the cytosol of rat liver and heart, Biochem.
Biophys. Res. Commun. 196 (1993) 1108e1112.
[43] M. Arita, Y. Sato, A. Miyata, T. Tanabe, E. Takahashi, H.J. Kayden,
H. Arai, K. Inoue, Human alpha-tocopherol transfer protein: cDNA
cloning, expression and chromosomal localization, Biochem. J. 306
(1995) 437e443.
[44] D. Fuks-Holmberg, K. Bloch, Intermembrane transfer of squalene
promoted by supernatant protein factor, J. Lipid. Res. 24 (1983)
402e408.
[45] T. Cecchini, S. Ciaroni, P. Ferri, P. Ambrogini, R. Cuppini, S. Santi,
P. Del Grande, Alpha-tocopherol, an exogenous factor of adult hippo-
campal neurogenesis regulation, J. Neurosci. Res. 73 (2003) 447e455.
[46] C.A. Drevon, Absorption, transport and metabolism of vitamin E, Free
Radic. Res. Commun. 14 (1991) 229e246.
[47] A.J. Ham, D.C. Liebler, Vitamin E oxidation in rat liver mitochondria,
Biochemistry 34 (1995) 5754e5761.
[48] F. Flamigni, C. Guarnieri, R. Toni, C.M. Caldarera, Effect of oxygen
radicals on heart mitochondrial function in alpha-tocopherol deficient
rabbits, Int. J. Vitam. Nutr. Res. 52 (1982) 402e406.
[49] J. Neuzil, P.K. Witting, R. Stocker, Alpha-tocopheryl hydroquinone is an
efficient multifunctional inhibitor of radical-initiated oxidation of low
density lipoprotein lipids, Proc. Natl. Acad. Sci. U.S.A. 94 (1997) 7885e
7890.
[50] W. Gregor, K. Staniek, H. Nohl, L. Gille, Distribution of tocopheryl
quinone in mitochondrial membranes and interference with ubiquinone-
mediated electron transfer, Biochem. Pharmacol. 71 (2006) 1589e1601.
[51] S.L. Cuddihy, S.S. Ali, E.S. Musiek, J. Lucero, S.J. Kopp, J.D. Morrow,
L.L. Dugan, Prolonged alpha-tocopherol deficiency decreases oxidative
stress and unmasks alpha-tocopherol-dependent regulation of mito-
chondrial function in the brain, J. Biol. Chem. 283 (2008) 6915e6924.
[52] C.W. Shiau, J.W. Huang, D.S. Wang, J.R. Weng, C.C. Yang, C.H. Lin,
C. Li, C.S. Chen, alpha-Tocopheryl succinate induces apoptosis in
prostate cancer cells in part through inhibition of BCL-XL/BCL-2
function, J. Biol. Chem. 281 (2006) 11819e11825.
[53] J. Neuzil, J.C. Dyason, R. Freeman, L.F. Dong, L. Prochazka, X.F. Wang,
I. Scheffler, S.J. Ralph, Mitocans as anti-cancer agents targeting mito-
chondria: lessons from studies with vitamin E analogues, inhibitors of
complex II, J. Bioenerg. Biomembr. 39 (2007) 65e72.
[54] L.F. Dong, P. Low, J.C. Dyason, X.F. Wang, L. Prochazka, P.K. Witting,
R. Freeman, E. Swettenham, K. Valis, J. Liu, R. Zobalova, J. Turanek,
D.R. Spitz, F.E. Domann, I.E. Scheffler, S.J. Ralph, J. Neuzil, Alpha-
tocopheryl succinate induces apoptosis by targeting ubiquinone-binding
sites in mitochondrial respiratory complex II, Oncogene (2008).

1715J.-M. Zingg et al. / Biochimie 90 (2008) 1703e1715
[55] J.M. Zingg, Vitamin E and Mast Cells, vol. 76, Academic Press, 2007,
pp. 391e416.
[56] G. Schaaf, E.A. Ortlund, K.R. Tyeryar, C.J. Mousley, K.E. Ile,
T.A. Garrett, J. Ren, M.J. Woolls, C.R. Raetz, M.R. Redinbo,
V.A. Bankaitis, Functional anatomy of phospholipid binding and regu-
lation of phosphoinositide homeostasis by proteins of the sec14 super-
family, Mol. Cell. 29 (2008) 191e206.
[57] A. Munteanu, M. Taddei, I. Tamburini, E. Bergamini, A. Azzi,
J.M. Zingg, Antagonistic effects of oxidized low density lipoprotein and
{alpha}-tocopherol on cd36 scavenger receptor expression in monocytes:
involvement of protein kinase b and peroxisome proliferator-activated
receptor-{gamma}, J. Biol. Chem. 281 (2006) 6489e6497.
[58] M.C. De Pascale, A.M. Bassi, V. Patrone, L. Villacorta, A. Azzi,
J.M. Zingg, Increased expression of transglutaminase-1 and PPAR-
gamma after vitamin E treatment in human keratinocytes, Arch. Bio-
chem. Biophys. 447 (2006) 97e106.
[59] J.C. Hay, T.F. Martin, Phosphatidylinositol transfer protein required for
ATP-dependent priming of Ca(2þ)-activated secretion, Nature 366
(1993) 572e575.
[60] C. Debier, Vitamin E during pre- and postnatal periods, Vitam. Horm. 76
(2007) 357e373.
[61] Y. Saral, B.K. Coskun, P. Ozturk, F. Karatas, A. Ayar, Assessment of
salivary and serum antioxidant vitamins and lipid peroxidation in patients
with recurrent aphthous ulceration, Tohoku J. Exp. Med. 206 (2005)
305e312.
[62] B. Rai, S. Kharb, R. Jain, S.C. Anand, Salivary vitamins E and C in oral
cancer, Redox. Rep. 12 (2007) 163e164.
[63] R.F. Aten, T.R. Kolodecik, H.R. Behrman, Ovarian vitamin E accumulation:
evidence for a role of lipoproteins, Endocrinology 135 (1994) 533e539.
[64] Q. Zhu, M.A. Emanuele, N. Lapaglia, E.J. Kovacs, N.V. Emanuele,
Vitamin E prevents ethanol-induced inflammatory, hormonal, and cyto-
toxic changes in reproductive tissues, Endocrine 32 (2007) 59e68.
[65] R.H. Wasserman, A.N. Taylor, Metabolic roles of fat-soluble vitamins D,
E, and K, Annu. Rev. Biochem. 41 (1972) 179e202.
[66] J.M. Dietschy, S.D. Turley, Cholesterol metabolism in the brain, Curr.
Opin. Lipidol. 12 (2001) 105e112.
[67] X.Q. Wen, X.J. Li, Z.L. Su, Y. Liu, X.F. Zhou, Y.B. Cai, W.T. Huang,
X. Gao, Reduced expression of alpha-tocopherol-associated protein is
associated with tumor cell proliferation and the increased risk of prostate
cancer recurrence, Asian J. Androl. 9 (2007) 206e212.
[68] J.M. Zingg, A. Azzi, Modulation of cellular signalling and gene
expression by vitamin E, in: O.H. Bendrick (Ed.), New Topics in Vitamin
E Research, NOVA Publishers, 2006.
[69] S. Buddhan, R. Sivakumar, N. Dhandapani, B. Ganesan, R. Anandan,
Protective effect of dietary squalene supplementation on mitochondrial
function in liver of aged rats, Prostaglandins. Leukot. Essent. Fatty.
Acids. 76 (2007) 349e355.
[70] K. Hirai, G. Aliev, A. Nunomura, H. Fujioka, R.L. Russell, C.S. Atwood,
A.B. Johnson, Y. Kress, H.V. Vinters, M. Tabaton, S. Shimohama,
A.D. Cash, S.L. Siedlak, P.L. Harris, P.K. Jones, R.B. Petersen, G. Perry,
M.A. Smith, Mitochondrial abnormalities in Alzheimer’s disease, J.
Neurosci. 21 (2001) 3017e3023.
[71] M.W. Fariss, J.G. Zhang, Vitamin E therapy in Parkinson’s disease,
Toxicology 189 (2003) 129e146.
[72] B. Marriage, M.T. Cladinin, D.M. Glerum, Nutritional cofactor treatment
in mitochondrial disorders, J. Am. Diet. Assoc. 103 (2003) 1029e1038.
[73] J. Kaplan, Spinocerebellar ataxias due to mitochondrial defects, Neuro-
chem. Int. 40 (2002) 553e557.




















