
Characterization of the cydAB-Encoded Cytochrome bd Oxidase from Mycobacterium smegmatis
12
10.1128/JB.183.24.7076-7086.2001. 2001, 183(24):7076. DOI: J. Bacteriol. Stephanie S. Dawes, Harvey Rubin and Valerie Mizrahi Bavesh D. Kana, Edward A. Weinstein, David Avarbock, Mycobacterium smegmatis Oxidase from bd Cytochrome -Encoded cydAB Characterization of the http://jb.asm.org/content/183/24/7076 Updated information and services can be found at: These include: REFERENCES http://jb.asm.org/content/183/24/7076#ref-list-1 at: This article cites 61 articles, 30 of which can be accessed free CONTENT ALERTS more» articles cite this article), Receive: RSS Feeds, eTOCs, free email alerts (when new http://journals.asm.org/site/misc/reprints.xhtml Information about commercial reprint orders: http://journals.asm.org/site/subscriptions/ To subscribe to to another ASM Journal go to: on November 17, 2014 by guest http://jb.asm.org/ Downloaded from on November 17, 2014 by guest http://jb.asm.org/ Downloaded from
Transcript of Characterization of the cydAB-Encoded Cytochrome bd Oxidase from Mycobacterium smegmatis

10.1128/JB.183.24.7076-7086.2001.
2001, 183(24):7076. DOI:J. Bacteriol. Stephanie S. Dawes, Harvey Rubin and Valerie MizrahiBavesh D. Kana, Edward A. Weinstein, David Avarbock,
Mycobacterium smegmatis Oxidase frombdCytochrome
-EncodedcydABCharacterization of the
http://jb.asm.org/content/183/24/7076Updated information and services can be found at:
These include:
REFERENCEShttp://jb.asm.org/content/183/24/7076#ref-list-1at:
This article cites 61 articles, 30 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on Novem
ber 17, 2014 by guesthttp://jb.asm
.org/D
ownloaded from
on N
ovember 17, 2014 by guest
http://jb.asm.org/
Dow
nloaded from

JOURNAL OF BACTERIOLOGY,0021-9193/01/$04.00�0 DOI: 10.1128/JB.183.24.7076–7086.2001
Dec. 2001, p. 7076–7086 Vol. 183, No. 24
Copyright © 2001, American Society for Microbiology. All Rights Reserved.
Characterization of the cydAB-Encoded Cytochrome bd Oxidasefrom Mycobacterium smegmatis
BAVESH D. KANA,1,2 EDWARD A. WEINSTEIN,3 DAVID AVARBOCK,3 STEPHANIE S. DAWES,1,2
HARVEY RUBIN,3* AND VALERIE MIZRAHI1,2*
MRC/SAIMR/WITS Molecular Mycobacteriology Research Unit, South African Institute for Medical Research,1 andDepartment of Molecular Medicine and Hematology, School of Pathology, University of the Witwatersrand,2
Johannesburg, South Africa, and Division of Infectious Diseases, University of Pennsylvania,Philadelphia, Pennsylvania3
Received 25 July 2001/Accepted 19 September 2001
The cydAB genes from Mycobacterium smegmatis have been cloned and characterized. The cydA and cydBgenes encode the two subunits of a cytochrome bd oxidase belonging to the widely distributed family of quinoloxidases found in prokaryotes. The cydD and cydC genes located immediately downstream of cydB encode aputative ATP-binding cassette-type transporter. At room temperature, reduced minus oxidized differencespectra of membranes purified from wild-type M. smegmatis displayed spectral features that are characteristicof the �-proteobacterial type cytochrome bd oxidase. Inactivation of cydA or cydB by insertion of a kanamycinresistance marker resulted in loss of d-heme absorbance at 631 nm. The d-heme could be restored bytransformation of the M. smegmatis cyd mutants with a replicating plasmid carrying the highly homologouscydABDC gene cluster from Mycobacterium tuberculosis. Inactivation of cydA had no effect on the ability of M.smegmatis to exit from stationary phase at 37 or 42°C. The growth rate of the cydA mutant was tested underoxystatic conditions. Although no discernible growth defect was observed under moderately aerobic conditions(9.2 to 37.5 � 102 Pa of pO2 or 5 to 21% air saturation), the mutant displayed a significant growth disadvantagewhen cocultured with the wild type under extreme microaerophilia (0.8 to 1.7 � 102 Pa of pO2 or 0.5 to 1% airsaturation). These observations were in accordance with the two- to threefold increase in cydAB gene expressionobserved upon reduction of the pO2 of the growth medium from 21 to 0.5% air saturation and with theconcomitant increase in d-heme absorbance in spectra of membranes isolated from wild-type M. smegmatiscultured at 1% air saturation. Finally, the cydA mutant displayed a competitive growth disadvantage in thepresence of the terminal oxidase inhibitor, cyanide, when cocultured with wild type at 21% air saturation in anoxystat. In conjunction with these findings, our results suggest that cytochrome bd is an important terminaloxidase in M. smegmatis.
The major human pathogen, Mycobacterium tuberculosis isresponsible for 3 million deaths and 8 million new cases oftuberculosis per annum (12). Most cases of tuberculosis arisefrom reactivation of an infection acquired years before theonset of symptoms, reflecting the emergence of actively repli-cating organisms from a latent or dormant state (4, 34, 55).Even after prolonged adequate therapy, M. tuberculosis DNAcan be detected in the sputum of patients, and the organismmay reactivate to cause disease following immunosuppression(15, 21). It is widely believed that oxygen limitation, amino acidstarvation, and carbon source restriction are involved in estab-lishing and maintaining dormancy in M. tuberculosis (10, 24,49). Several in vitro models for studying the physiology of the“dormant” state of M. tuberculosis have been developed on thebasis of these assumptions (42, 59). The Wayne model ofmycobacterial dormancy is based on interrupting the aerationof logarithmic-phase cultures. After cessation of growth andautolysis of most of the bacilli, a small proportion of survivors
are able to resume growth at a reduced rate (59). The evidencesuggests that mycobacteria, once thought to be obligately aer-obic, can adapt and perhaps thrive under reduced oxygen con-ditions (59–62). Significantly, Segal found evidence that thevirulence of mycobacteria, and of M. tuberculosis specifically, isrelated to the ability to grow under reduced concentrations ofO2 (48).
Aerobic and facultative aerobic bacteria are able to respondto altered levels of oxygen in the environment by utilizingalternative respiratory pathways (41). As such, many bacterialspecies possess more than one terminal oxidase, which cata-lyzes the oxidation of either cytochrome c or quinol. The cy-tochrome bd oxidase is a widely distributed prokaryotic quinoloxidase, which performs a variety of physiological functions invivo (17, 27, 33, 41). It is involved in energy-transducing res-piration in Escherichia coli (27) and Bacillus species (45). Cy-tochrome bd also plays a role in aerotolerant nitrogen fixationand protection against metal toxicity and oxidative stress inAzotobacter vinelandii (13, 28, 30). Significantly, Way et al.recently demonstrated a positive correlation between the vir-ulence of the bacterial pathogen, Shigella flexneri, and the levelof cytochrome bd expression (58), suggesting that cytochromebd may be particularly important for the growth and survival ofpathogens that encounter environments in which O2 is pro-gressively limited. Sequence analysis of the M. tuberculosis ge-
* Corresponding author. Mailing address for Harvey Rubin: Uni-versity of Pennsylvania, 225 Johnson Pavilion, Philadelphia, PA 19104.Phone: (215) 662-6475. Fax: (215) 662-7842. E-mail: [email protected].. Mailing address for Valerie Mizrahi: MRC/SAIMR/WITS Molecular Mycobacteriology Research Unit, SAIMR, P.O. Box1038, Johannesburg 2000, South Africa. Phone: 2711-4899370. Fax:2711-4899001. E-mail: [email protected].
7076
on Novem
ber 17, 2014 by guesthttp://jb.asm
.org/D
ownloaded from

nome (7) revealed the presence of a cydABDC gene cluster, thecydAB genes of which encode a putative cytochrome bd oxidasethat is highly homologous to its counterpart in E. coli. Byanalogy with the physiological function of the E. coli oxidase,we speculated that the mycobacterial bd oxidase might play arole in adaptation to conditions of reduced oxygen. To test thishypothesis, we investigated the function of the cydAB genes inMycobacterium smegmatis, which is a fast-growing nonpatho-genic saprophyte that demonstrates oxygen depletion-induceddormancy similar to that observed with M. tuberculosis (11) andis regarded as an important model for studying mycobacterialphysiology (35, 52, 66). In this study, we report the gene clon-ing, allelic disruption, spectral analysis, expression analysis,and phenotypic characterization of cyd-inactivated mutantstrains, and we demonstrate that this oxidase plays an impor-tant respiratory role in M. smegmatis.
MATERIALS AND METHODS
Materials, bacterial strains, plasmids, and growth conditions. Enzymes werefrom Boehringer Mannheim or Amersham, radiochemicals were from ICN, andall other chemicals were from Sigma. The bacterial strains and plasmids used inthis study are listed in Table 1. E. coli DH5� was used for plasmid manipulationsand JM101 was used for M13 cloning (47). For E. coli, ampicillin, kanamycin(KAN), and hygromycin (HYG) were used at 100, 50 and 200 �g/ml, respec-tively. For M. smegmatis, KAN was used at 10 (solid medium) or 25 (liquidmedium) �g/ml, and HYG was used at 50 �g/ml. E. coli strains were grown inLuria-Bertani broth (LB) or agar (LA). M. smegmatis strains were grown in LB
for targeted knockout mutagenesis or in MADC-Tw (Middlebrook 7H9 broth[Difco] supplemented with 0.085% NaCl, 0.2% glucose, 0.2% glycerol, and0.05% Tween 80) and on LA as the solid medium.
Cloning and sequencing of the M. smegmatis cyd gene cluster. A cos-mid carrying the M. tuberculosis cydABDC genes was isolated from apYUB328::H37Rv cosmid library (3) by using a probe derived from the closelylinked polA gene (22). A 2.5-kb SalI fragment carrying the cydAB genes wasisolated from the cosmid and used to probe a gridded plasmid library of M.smegmatis (kindly provided by M. Everettt, Glaxo Wellcome, Stevenage, UnitedKingdom). A positive clone, p45C10, was found to contain an insert showing ahigh degree of homology to cydA and cydB from M. tuberculosis. This 1-kbBamHI-HindIII fragment was then used to probe 2- to 3-kb KpnI and 9- to 11-kbBglII size-fractionated libraries of M. smegmatis genomic DNA cloned inpGEM32F(�), from which pBK1and pBK2 were identified. The complete se-quence of the cydABDC cluster was determined by sequencing 0.5- to 1.2-kbinserts of pBK1 and pBK2 subcloned in M13mp18/19. All clones were sequencedon both strands by using a combination of universal and internal primers. DNAsequencing was carried out with an ABI Prism 377 automated DNA sequencer,and analysis was performed by using the Lasergene suite of programs (DNASTAR,Madison, Wis.) and the BLAST server located at the National Centre for Bio-technology Information (Bethesda, Md.; http://www.ncbi.nlm.nih.gov/cgi-bin/BLAST). Preliminary sequence data for M. smegmatis mc2155 was obtainedfrom The Institute for Genomic Research website (http://www.tigr.org).
Construction of cydA::aph and cydB::aph knockout mutants of M. smegmatis.The suicide plasmid, pCYDAKO, carrying a cydA::aph allele, was constructedfrom pBK1 for use in allelic exchange by cloning the Tn903 KAN resistance(Kmr) (aph) cassette from pY6002 (23) into the PstI site in cydA. The hsp60-lacZcassette from pSlacZ and the hsp60-sacB cassette from p6.11 were then sequen-tially cloned into the SalI and XbaI sites of the vector to form pCYDAKO. Theplasmid pCYDBKO was constructed by cloning the aph gene into the SphI sitelocated in the cydB gene, followed by sequential cloning of the selection markers,
TABLE 1. Strains and plasmids used in this study
M. smegmatis strainor plasmid Characteristicsa Source or
reference
M. smegmatismc2155 High-frequency transformation mutant of M. smegmatis ATCC 607 53mc2155 (pHINT) mc2155 with pHINT integrated at the attP locus; Hygr This workmc2155(pOTBCYD) mc2155 carrying M. tuberculosis cydABDC gene cluster on pOTBCYD; Hygr This workcydA::aph cydA knockout mutant of mc2155; Kmr This workcydA::aph(pOTBCYD) cydA::aph carrying M. tuberculosis cydABDC gene cluster on pOTBCYD; Kmr Hygr This workcydB::aph cydB knockout mutant of mc2155; Kmr This workcydB::aph(pOTBCYD) cydB::aph carrying M. tuberculosis cydABDC gene cluster on pOTBCYD; Kmr Hygr This workmc2155::pBK4 mc2155 carrying pBK4 integrated at the cyd locus; Kmr This work
PlasmidspGEM32F(�) E. coli cloning vector; Apr PromegapOLYG Multicopy E. coli-Mycobacterium shuttle vector; Hygr 37p6.11 pOLYG carrying hsp60-sacB cassette; Hygr S. QuanpHINT Integrative shuttle vector that integrates at the Mycobacterium attP locus; Hygr Apr 37p2NIL Suicide plasmid used for cloning homologous recombination substrates 40pOTBCYD pOLYG carrying the M. tuberculosis cydABDC genes carried on an 11kb EcoRI fragment; Hygr This workpSMT3 Derivative of pOLYG carrying M. bovis BCG hsp60 promoter; Hygr 37pSlacZ Derivative of pSMT3 carrying hsp60-lacZ cassette; Hygr This workp45C10 pBluescript SK(�) construct containing partial M. smegmatis cydAB region M. Everettp16N11 pBluescript SK(�) construct containing partial M. smegmatis cydA and upstream promoter
regionM. Everett
pBK1 pGEM32F(�) carrying 2.4-kb KpnI fragment from M. smegmatis containing part of cydA andcydB genes
This work
pBK2 pGEM32F(�) carrying 10-kb BglII fragment from M. smegmatis containing entire cydABDCgene cluster
This work
pCYDAKO pBK1 carrying cydA::aph, hsp60-sacB, and hsp60-lacZ; used for targeted knockout of M.smegmatis cydA; Kmr
This work
pCYDBKO pBK1 carrying cydB::aph, hsp60-sacB, and hsp60-lacZ; used for targeted knockout of M.smegmatis cydB; Kmr
This work
pATB12 Multicopy E. coli-Mycobacterium shuttle vector; lacZ, Hygr M. EverettpBK3 pATB12 carrying 2,634-bp PstI fragment from p16N11 containing M. smegmatis cyd promoter
region, cloned immediately upstream of lacZ; HygrThis work
pBK4 p2NIL containing the cydA�::lacZ cassette from pBK3; Kmr This work
a Apr, ampicillin resistance; Hygr, HYG resistance.
VOL. 183, 2001 M. SMEGMATIS CYTOCHROME bd OXIDASE 7077
on Novem
ber 17, 2014 by guesthttp://jb.asm
.org/D
ownloaded from

as described for pCYDAKO. These plasmids conferred sucrose sensitivity on E.coli and resulted in the development of blue color when plated on X-Gal (5-bromo-4-chloro-3-indolyl-�-D-galactopyranoside)-containing indicator medium.M. smegmatis was transformed with pCYDAKO and pCYDBKO, as previouslydescribed (6), and recombinants were selected by plating on medium containingKAN and X-Gal. Blue, Kmr colonies were picked, diluted, and plated on mediumcontaining KAN, X-Gal, and 5% sucrose. Sucrose-resistant colonies that re-mained white after patching on fresh indicator medium (3 of 200 in both cases)were selected as candidate allelic exchange mutants and were genotyped bySouthern blot analysis by previously described methods (6, 20).
Construction of cydA�::lacZ reporter strain. A 2,634-bp PstI fragment fromp16N11 (isolated from the gridded plasmid library of M. smegmatis), containingthe first 959 bp of cydA flanked by 1,675 bp of upstream sequence was used tomake the reporter construct. The 1,675-bp region consists of a 95-bp intergenicspacer immediately upstream of cydA (probably containing the cyd promoter),preceded by an open reading frame that is highly homologous to the Rv1624cgene from M. tuberculosis (7), preceded by a further 962 bp of upstream se-quence. The PstI fragment was cloned into in the ScaI site of pATB12, which islocated immediately upstream of the lacZ reporter, to form pBK3. ThecydA�::lacZ cassette was excised from pBK3 as a NotI-PstI fragment and clonedin NotI-PstI-digested p2NIL to produce pBK4. M. smegmatis was transformedwith pBK4, as previously described (6), and recombinants were selected byplating on media containing KAN and X-Gal. Southern blot analysis was used toconfirm that the blue colonies contained pBK4 integrated site specifically at thecyd locus.
Temperature sensitivity assays. To determine whether the cyd mutants weretemperature sensitive for growth and/or survival, overnight cultures of the wild-type and cydA::aph strains were diluted to an optical density at 600 nm (OD600)of 0.03 to 0.04 and grown at 37°C with shaking (100 rpm) to late stationary phase.Aliquots were removed during the late exponential and stationary phases ofgrowth, and CFU were enumerated by plating at least three serial dilutions induplicate and incubating them at 37 or 42°C.
Growth of M. smegmatis under oxystatic conditions. Growth of M. smegmatisunder defined oxygen tensions (pO2) was carried out in a Braun BIOSTAT Bfermentor in batch cultures. The water-jacketed, 3-liter fermentor vessel had a2-liter working volume and was equipped with baffles, two six-bladed impellers,a temperature probe, a ring sparger, a sampling port, and a Mettler Toledodissolved-O2 probe. The probes and water jacket were linked to a fermentorcontrol unit that monitored and adjusted all of the variables in the system, exceptfor the airflow rate. For air supply, compressed air was passed into the ringsparger through a rotometer and a 0.22-�m (pore-size) sterile filter. The roto-meter allowed for control of the air supply from 0 to 400 ml/min, and thisparameter was set manually, depending on the air saturation level required. Thetemperature was maintained by connecting the vessel water jacket to a circula-tory water-cooling unit. Stirring was accomplished by using an overhead motor toturn the impeller shaft. The fermentor control unit was also equipped with adissolved-O2 cascade control, which linked the stirring rate to the dissolved-O2
level. Agitation speeds ranged from 100 to 600 rpm. All experiments conductedin this system were carried out at the research facility of the Process Biotech-nology Division of the Council for Scientific and Industrial Research in Mod-derfontein, Johannesburg, South Africa, which is located 1,800 m above sea level,at an atmospheric pressure of 84.2 kPa.
The fermentor vessel was first sterilized with 1.5 liter of Middlebrook 7H9 plus3 ml of glycerol. As the vessel cooled down, Tween 80, NaCl, and glucose wereadded, and the final volume was made up to 1.6 liter. The vessel was allowed toequilibrate overnight. Thereafter, the dissolved-O2 probe was calibrated to 0%air saturation by sparging the medium with 99.99% nitrogen and to 100% airsaturation by sparging with 100% compressed air (21% O2; 176.8 Pa). Precul-tures were prepared by inoculating 100 ml of MADC plus Tw medium with 4 mlof a freezer stock of the culture and growing the mixture for 16 h on a rotaryshaker (100 rpm), at which time the OD600 of the culture had reached 2 (ca. 108
CFU/ml). For competitive growth experiments with the mc2155::pHINT (wildtype) and cydA::aph (mutant) strains, the precultures were mixed and added tothe 1.6 liter of medium contained in the vessel to yield a starting culture con-taining a total of ca. 107 CFU/ml. An 800-�l aliquot of sterile antifoam (Durapol;AECI Bioproducts) was added to the culture, the stirrer was set to 100 rpm, airinflow was shut off, and the bacteria were allowed to consume the oxygen presentin the medium. Once the air saturation had dropped to the desired level, therotometer was set at a flow rate of 60 ml/min (for 21% air saturation), 40 ml/min(for 5% air saturation), or 20 ml/min (for 1 and 0.5% air saturation), and thestirrer was set to cascade control. Under these conditions, the pO2 could bemaintained within �10% of the desired level over the duration of the experiment(24 h). Samples were withdrawn at 3-h intervals for the first 12 h of the exper-
iment, and a final sample was taken 12 h later. Growth was monitored by platingon LA containing either KAN (to select for cydA::aph) or HYG (to select formc2155::pHINT).
Oxystatic growth of the mc2155::pBK4 reporter strain was carried out byinoculating 0.6 liter of culture medium (MADC plus Tw containing KAN [10�g/ml]) with 200 ml of a preculture to give a starting cell density of ca. 7 � 107
CFU/ml. After the addition of 400 �l of antifoam, the culture was grown for 6 h,with air saturation being controlled as described above. Duplicate samples (1 to2 ml) were withdrawn at 2-h intervals, and the experiment was carried out intriplicate at each level of air saturation (0.5, 1, 5, and 21%).
Inhibition by cyanide under oxystatic conditions. A New Brunswick ScientificBioflow 3000 fermentor system (University of Pennsylvania) was used for thecyanide inhibition studies. The mechanical organization of this system, the pO2
probe calibration, vessel preparation, and other relevant methods were similar tothose of the Braun fermentor system decribed above. The strains used in thisstudy were mc2155 and cydA::aph, which were grown in MADC medium, butwithout Tween and antifoam additives. Instead, clumps were disrupted by vor-texing samples with 1/8-in. glass beads (3 ml of beads in 4 ml of culture, vortexedfor 90 s in a 22-by-70-mm vial). For the competitive growth experiments, the finalworking volume was 3 liters, to which was added an inoculum of 5 ml of a frozenpreculture of each strain (ca. 106 CFU of each/ml) to yield a starting coculturecontaining ca. 102 CFU of each strain/ml. KCN (1 mM) was added 30 min afterinoculation, and CFU were enumerated by plating in duplicate on 7H9 agar inthe presence (to score mutant CFU) or absence (to score total CFU) of KAN.
�-Galactosidase assays. Assays were performed on cell extracts, as previouslydescribed (18), with the following modifications. Cells were lysed in Z-buffer ina Savant BIO101 Fastprep cell disruptor according to the manufacturer’s instruc-tions. Assays were carried out in Z-buffer (without �-mercaptoethanol), with 50to 100 �l of soluble cell extract, in a final volume of 1 ml by using 2-nitrophenyl-�-D-galactopyranoside (ONPG) at 0.8 mg/ml as the substrate. Protein concen-trations in the cell extracts were determined by using a Bradford-based Bio-Radkit and following the manufacturer’s instructions. For a given experiment, thespecific activities determined for the four duplicate samples taken over the 6-htime course were averaged to obtain the mean for a given air saturation value.Overall means and standard deviations were obtained from the data generatedfrom three independent growth experiments.
Spectral analysis of membrane fractions. Unless otherwise indicated, culturesof M. smegmatis were grown aerobically in MADC plus Tw for 3 days at 37°C,with shaking at 100 rpm. Cultures were then transferred to 1-liter Schott bottles,the headspaces were purged with argon, and the bottles were sealed and main-tained at 37°C for a further 5 to 6 days. During this period, the bacteria settledslowly to the bottom of the vessel. The settled bacteria were then harvested bycentrifugation. Cells (8 g [wet weight]) were washed with 5 ml of Tris-cholate(TC) buffer (10 mM Tris-HCl [pH 7.4], 16 mM cholate) and resuspended in thesame buffer. The bacteria were lysed by passage through a French press at 14,000kPa, and the lysate was centrifuged at 12,000 � g for 15 min to remove cellulardebris. The supernatant was further clarified by centrifugation at 50,000 � g for50 min. The ultracentrifuged supernatant was used as the membrane-containingfraction for spectral studies. Further centrifugation at 150,000 � g for 90 min toisolate membrane particles did not improve the quality of the spectra. Proteinconcentrations were determined by bicinchoninic acid assay (Pierce) by usingbovine serum albumin as the standard. Reduced minus oxidized spectra wererecorded at room temperature on a Cary 4E dual-beam spectrophotometer, byusing a few grains of fresh solid sodium dithionite as the reductant and 100 �Mpotassium ferricyanide as the oxidant.
Quantitation of cytochrome bd oxidase. The amount of cytochrome bd oxidasewas determined as described by Junemann and Wrigglesworth (26) by using anextinction coefficient of 27 mM�1cm�1 for the 629- to 650-nm wavelength pair.
RESULTS
Identification and characterization of the M. smegmatis cydgene cluster. The high degree of homology between the cydABgenes of M. tuberculosis and their counterparts from M. smeg-matis allowed the entire M. smegmatis cyd gene cluster to becloned by using a cydAB probe derived from M. tuberculosis(Fig. 1A). The cydABDC organization in M. smegmatis paral-lels that observed in M. tuberculosis (7), B. subtilis (63), andStreptomyces coelicolor. The spacing of 13 bp between cydA andcydB suggests that these genes, like their counterparts from E.coli and B. subtilis, are polycistronic. However, the intergenic
7078 KANA ET AL. J. BACTERIOL.
on Novem
ber 17, 2014 by guesthttp://jb.asm
.org/D
ownloaded from

spacing of 90 bp between cydB and cydD suggests that thecydDC genes may be expressed independently from cydAB. Asexpected, the M. smegmatis cyd genes encode polypeptidesshowing the greatest homology to their counterparts from M.tuberculosis (78, 67, 65, and 64% identity for CydA, CydB,CydC, and CydD, respectively). In the -proteobacterial CydAprototype from E. coli, His19, His186, and Met393 providethree of the four axial ligands to the irons of the hemes in thecytochrome bd complex (14, 29). These three residues areconserved in the CydA polypeptides from B. subtilis, M. smeg-matis, and M. tuberculosis. The counterparts of the Glu99,Glu107 residues of E. coli CydA, which have been proposed toform part of a proton channel leading from the cytoplasmicside of the membrane to the oxygen reactive site (38) are alsoconserved in M. smegmatis and M. tuberculosis CydA. Further-more, the GRQPW motif, which is implicated in the binding ofthe quinol or heme-d in E. coli CydA (38), is also conserved inthe mycobacterial CydA homologues, but the C-terminal halfof the Q-loop of E. coli CydA is deleted. The mycobacterialcytochrome bd oxidases therefore belong to the more widelydistributed and evolutionarily older class of cytochrome bdoxidases found in gram-positive, cyanobacterial and archaealorganisms (46).
Construction and characterization of cyd knockout mutantsof M. smegmatis. To investigate the in vivo function of thecydAB-encoded cytochrome bd oxidase in M. smegmatis, weconstructed cydA::aph and cydB::aph mutants by homologous
recombination. The mutant alleles were delivered on suicidevectors carrying lacZ (39) and sacB markers (42). Southernblot analysis confirmed that the white, sucrose-resistant clonesrecovered from the mutagenesis procedure with pCYDAKOand pCYDBKO were site-specific, allelic exchange mutants(Fig. 1B). Inactivation of cydA and/or cydB genes had no dis-cernible effect on the rate of growth of M. smegmatis during theexponential phase, when the organism was grown in shakingflasks (data not shown). To investigate whether cyd gene inac-tivation conferred a stationary-phase exit defect analogous tothat observed in E. coli cyd mutants (19, 51), the M. smegmatiscydA::aph mutant was tested for temperature sensitivity forgrowth and survival. The mutant displayed no defect duringthe exponential phase of growth when cultured at 42°C. More-over, comparison of the plating efficiencies at 37 and 42°C ofcultures sampled 6 to 14 h after the cessation of growth re-vealed that the wild-type and mutant strains were indistin-guishable (data not shown).
Spectral analysis of cytochromes in membranes of wild-typeand cyd mutant strains of M. smegmatis. Reduced minus oxi-dized difference spectroscopy at room temperature was used toanalyze the cytochrome content of M. smegmatis (Fig. 2A).Major absorbance peaks observed at 600, 563, and 552 nmwere characteristic of a-, b-, and c-type cytochromes, respec-tively (25). A discrete peak at 631 nm denoted the presence ofheme-d, which is characteristic of the cytochrome bd oxidase inE. coli (36) and B. subtilis (63). The spectra of the cydA::aph
FIG. 1. Construction and genotypic characterization of M. smegmatis cyd mutants. (A) Restriction map of the cyd locus of M. smegmatis showingthe insertion sites of the aph cassette in the cyd knockout mutants. The 2.4-kb KpnI fragment from pBK1, used as the probe for Southern blotanalysis (see panel B), is denoted by a dashed, double-headed arrow. Restriction sites: Bg, BglII; H, HindIII; K, KpnI; P, PstI; S, SphI. The sequenceextending from the PstI site upstream of cydA to a SalI site downstream of cydC has been deposited in GenBank under accession no. AF196488.A hypothetical open reading frame that showed high homology to Rv1624c from M. tuberculosis was found upstream of cydA, confirming that thegenetic organization at this locus is conserved between M. smegmatis and M. tuberculosis. (B) Southern blot analysis of cyd mutants. Genomic DNAfrom the various strains was digested with the indicated enzyme(s) and probed with the 2.4-kb KpnI fragment from pBK1. In all cases, the sizesof the hybridizing fragments derived from the mutant strains were consistent with site-specific, allelic exchange.
VOL. 183, 2001 M. SMEGMATIS CYTOCHROME bd OXIDASE 7079
on Novem
ber 17, 2014 by guesthttp://jb.asm
.org/D
ownloaded from
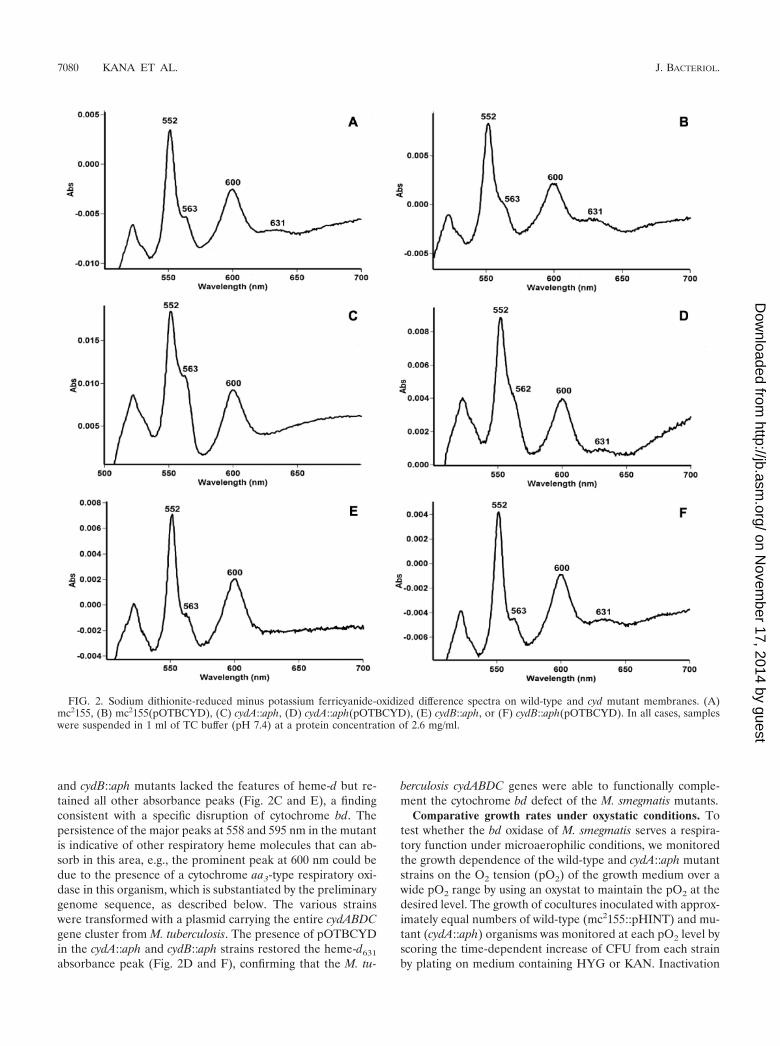
and cydB::aph mutants lacked the features of heme-d but re-tained all other absorbance peaks (Fig. 2C and E), a findingconsistent with a specific disruption of cytochrome bd. Thepersistence of the major peaks at 558 and 595 nm in the mutantis indicative of other respiratory heme molecules that can ab-sorb in this area, e.g., the prominent peak at 600 nm could bedue to the presence of a cytochrome aa3-type respiratory oxi-dase in this organism, which is substantiated by the preliminarygenome sequence, as described below. The various strainswere transformed with a plasmid carrying the entire cydABDCgene cluster from M. tuberculosis. The presence of pOTBCYDin the cydA::aph and cydB::aph strains restored the heme-d631
absorbance peak (Fig. 2D and F), confirming that the M. tu-
berculosis cydABDC genes were able to functionally comple-ment the cytochrome bd defect of the M. smegmatis mutants.
Comparative growth rates under oxystatic conditions. Totest whether the bd oxidase of M. smegmatis serves a respira-tory function under microaerophilic conditions, we monitoredthe growth dependence of the wild-type and cydA::aph mutantstrains on the O2 tension (pO2) of the growth medium over awide pO2 range by using an oxystat to maintain the pO2 at thedesired level. The growth of cocultures inoculated with approx-imately equal numbers of wild-type (mc2155::pHINT) and mu-tant (cydA::aph) organisms was monitored at each pO2 level byscoring the time-dependent increase of CFU from each strainby plating on medium containing HYG or KAN. Inactivation
FIG. 2. Sodium dithionite-reduced minus potassium ferricyanide-oxidized difference spectra on wild-type and cyd mutant membranes. (A)mc2155, (B) mc2155(pOTBCYD), (C) cydA::aph, (D) cydA::aph(pOTBCYD), (E) cydB::aph, or (F) cydB::aph(pOTBCYD). In all cases, sampleswere suspended in 1 ml of TC buffer (pH 7.4) at a protein concentration of 2.6 mg/ml.
7080 KANA ET AL. J. BACTERIOL.
on Novem
ber 17, 2014 by guesthttp://jb.asm
.org/D
ownloaded from
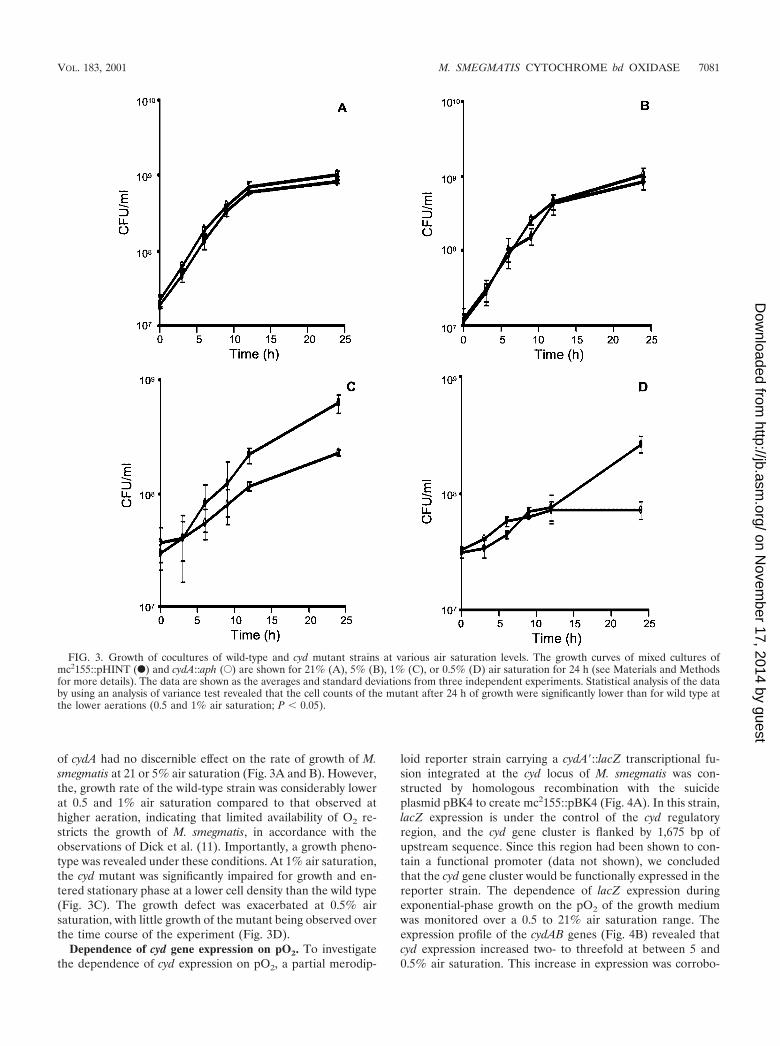
of cydA had no discernible effect on the rate of growth of M.smegmatis at 21 or 5% air saturation (Fig. 3A and B). However,the, growth rate of the wild-type strain was considerably lowerat 0.5 and 1% air saturation compared to that observed athigher aeration, indicating that limited availability of O2 re-stricts the growth of M. smegmatis, in accordance with theobservations of Dick et al. (11). Importantly, a growth pheno-type was revealed under these conditions. At 1% air saturation,the cyd mutant was significantly impaired for growth and en-tered stationary phase at a lower cell density than the wild type(Fig. 3C). The growth defect was exacerbated at 0.5% airsaturation, with little growth of the mutant being observed overthe time course of the experiment (Fig. 3D).
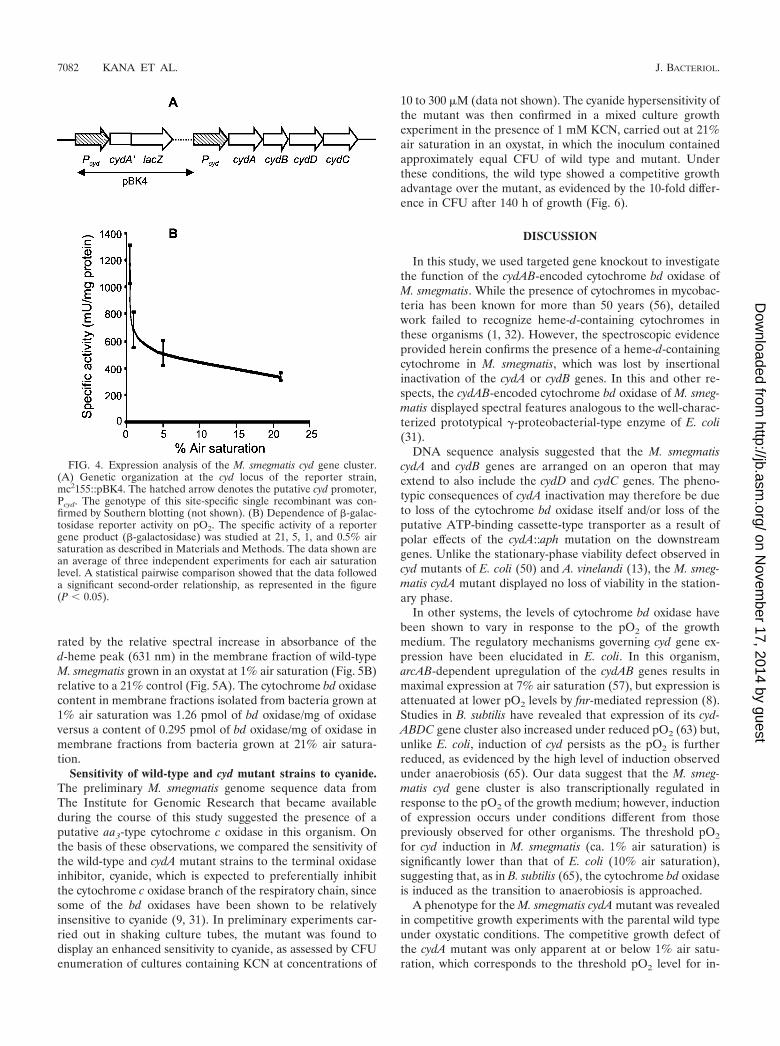
Dependence of cyd gene expression on pO2. To investigatethe dependence of cyd expression on pO2, a partial merodip-
loid reporter strain carrying a cydA�::lacZ transcriptional fu-sion integrated at the cyd locus of M. smegmatis was con-structed by homologous recombination with the suicideplasmid pBK4 to create mc2155::pBK4 (Fig. 4A). In this strain,lacZ expression is under the control of the cyd regulatoryregion, and the cyd gene cluster is flanked by 1,675 bp ofupstream sequence. Since this region had been shown to con-tain a functional promoter (data not shown), we concludedthat the cyd gene cluster would be functionally expressed in thereporter strain. The dependence of lacZ expression duringexponential-phase growth on the pO2 of the growth mediumwas monitored over a 0.5 to 21% air saturation range. Theexpression profile of the cydAB genes (Fig. 4B) revealed thatcyd expression increased two- to threefold at between 5 and0.5% air saturation. This increase in expression was corrobo-
FIG. 3. Growth of cocultures of wild-type and cyd mutant strains at various air saturation levels. The growth curves of mixed cultures ofmc2155::pHINT (F) and cydA::aph (E) are shown for 21% (A), 5% (B), 1% (C), or 0.5% (D) air saturation for 24 h (see Materials and Methodsfor more details). The data are shown as the averages and standard deviations from three independent experiments. Statistical analysis of the databy using an analysis of variance test revealed that the cell counts of the mutant after 24 h of growth were significantly lower than for wild type atthe lower aerations (0.5 and 1% air saturation; P 0.05).
VOL. 183, 2001 M. SMEGMATIS CYTOCHROME bd OXIDASE 7081
on Novem
ber 17, 2014 by guesthttp://jb.asm
.org/D
ownloaded from
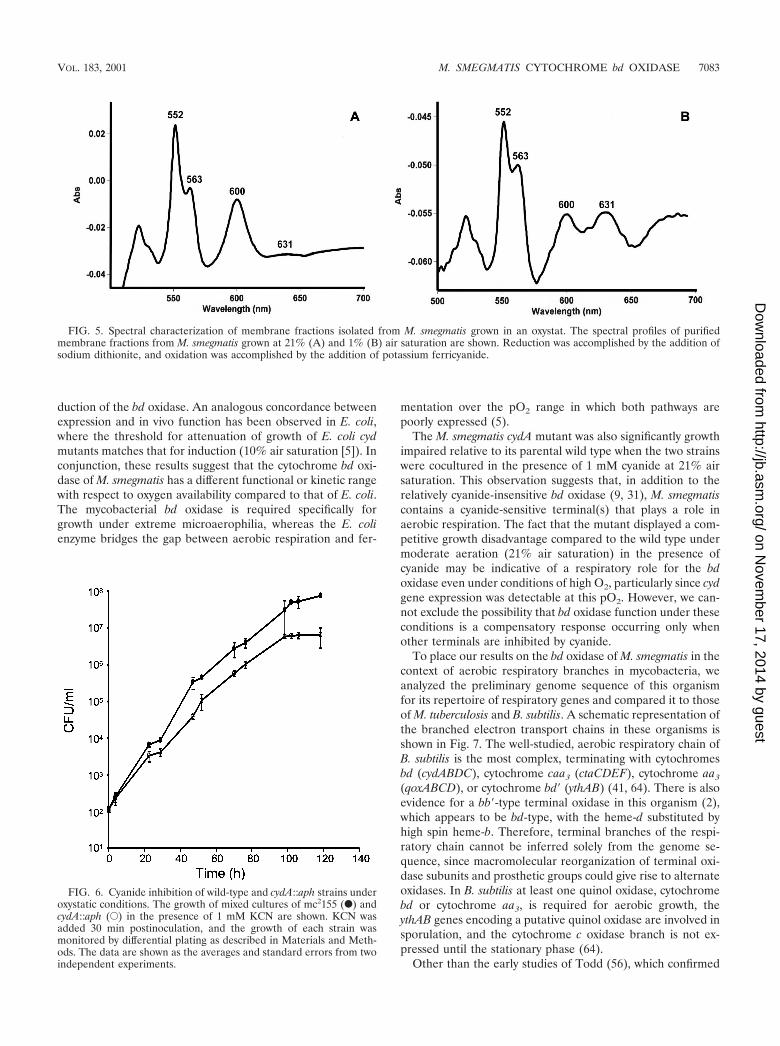
rated by the relative spectral increase in absorbance of thed-heme peak (631 nm) in the membrane fraction of wild-typeM. smegmatis grown in an oxystat at 1% air saturation (Fig. 5B)relative to a 21% control (Fig. 5A). The cytochrome bd oxidasecontent in membrane fractions isolated from bacteria grown at1% air saturation was 1.26 pmol of bd oxidase/mg of oxidaseversus a content of 0.295 pmol of bd oxidase/mg of oxidase inmembrane fractions from bacteria grown at 21% air satura-tion.
Sensitivity of wild-type and cyd mutant strains to cyanide.The preliminary M. smegmatis genome sequence data fromThe Institute for Genomic Research that became availableduring the course of this study suggested the presence of aputative aa3-type cytochrome c oxidase in this organism. Onthe basis of these observations, we compared the sensitivity ofthe wild-type and cydA mutant strains to the terminal oxidaseinhibitor, cyanide, which is expected to preferentially inhibitthe cytochrome c oxidase branch of the respiratory chain, sincesome of the bd oxidases have been shown to be relativelyinsensitive to cyanide (9, 31). In preliminary experiments car-ried out in shaking culture tubes, the mutant was found todisplay an enhanced sensitivity to cyanide, as assessed by CFUenumeration of cultures containing KCN at concentrations of
10 to 300 �M (data not shown). The cyanide hypersensitivity ofthe mutant was then confirmed in a mixed culture growthexperiment in the presence of 1 mM KCN, carried out at 21%air saturation in an oxystat, in which the inoculum containedapproximately equal CFU of wild type and mutant. Underthese conditions, the wild type showed a competitive growthadvantage over the mutant, as evidenced by the 10-fold differ-ence in CFU after 140 h of growth (Fig. 6).
DISCUSSION
In this study, we used targeted gene knockout to investigatethe function of the cydAB-encoded cytochrome bd oxidase ofM. smegmatis. While the presence of cytochromes in mycobac-teria has been known for more than 50 years (56), detailedwork failed to recognize heme-d-containing cytochromes inthese organisms (1, 32). However, the spectroscopic evidenceprovided herein confirms the presence of a heme-d-containingcytochrome in M. smegmatis, which was lost by insertionalinactivation of the cydA or cydB genes. In this and other re-spects, the cydAB-encoded cytochrome bd oxidase of M. smeg-matis displayed spectral features analogous to the well-charac-terized prototypical -proteobacterial-type enzyme of E. coli(31).
DNA sequence analysis suggested that the M. smegmatiscydA and cydB genes are arranged on an operon that mayextend to also include the cydD and cydC genes. The pheno-typic consequences of cydA inactivation may therefore be dueto loss of the cytochrome bd oxidase itself and/or loss of theputative ATP-binding cassette-type transporter as a result ofpolar effects of the cydA::aph mutation on the downstreamgenes. Unlike the stationary-phase viability defect observed incyd mutants of E. coli (50) and A. vinelandi (13), the M. smeg-matis cydA mutant displayed no loss of viability in the station-ary phase.
In other systems, the levels of cytochrome bd oxidase havebeen shown to vary in response to the pO2 of the growthmedium. The regulatory mechanisms governing cyd gene ex-pression have been elucidated in E. coli. In this organism,arcAB-dependent upregulation of the cydAB genes results inmaximal expression at 7% air saturation (57), but expression isattenuated at lower pO2 levels by fnr-mediated repression (8).Studies in B. subtilis have revealed that expression of its cyd-ABDC gene cluster also increased under reduced pO2 (63) but,unlike E. coli, induction of cyd persists as the pO2 is furtherreduced, as evidenced by the high level of induction observedunder anaerobiosis (65). Our data suggest that the M. smeg-matis cyd gene cluster is also transcriptionally regulated inresponse to the pO2 of the growth medium; however, inductionof expression occurs under conditions different from thosepreviously observed for other organisms. The threshold pO2
for cyd induction in M. smegmatis (ca. 1% air saturation) issignificantly lower than that of E. coli (10% air saturation),suggesting that, as in B. subtilis (65), the cytochrome bd oxidaseis induced as the transition to anaerobiosis is approached.
A phenotype for the M. smegmatis cydA mutant was revealedin competitive growth experiments with the parental wild typeunder oxystatic conditions. The competitive growth defect ofthe cydA mutant was only apparent at or below 1% air satu-ration, which corresponds to the threshold pO2 level for in-
FIG. 4. Expression analysis of the M. smegmatis cyd gene cluster.(A) Genetic organization at the cyd locus of the reporter strain,mc2155::pBK4. The hatched arrow denotes the putative cyd promoter,Pcyd. The genotype of this site-specific single recombinant was con-firmed by Southern blotting (not shown). (B) Dependence of �-galac-tosidase reporter activity on pO2. The specific activity of a reportergene product (�-galactosidase) was studied at 21, 5, 1, and 0.5% airsaturation as described in Materials and Methods. The data shown arean average of three independent experiments for each air saturationlevel. A statistical pairwise comparison showed that the data followeda significant second-order relationship, as represented in the figure(P 0.05).
7082 KANA ET AL. J. BACTERIOL.
on Novem
ber 17, 2014 by guesthttp://jb.asm
.org/D
ownloaded from
duction of the bd oxidase. An analogous concordance betweenexpression and in vivo function has been observed in E. coli,where the threshold for attenuation of growth of E. coli cydmutants matches that for induction (10% air saturation [5]). Inconjunction, these results suggest that the cytochrome bd oxi-dase of M. smegmatis has a different functional or kinetic rangewith respect to oxygen availability compared to that of E. coli.The mycobacterial bd oxidase is required specifically forgrowth under extreme microaerophilia, whereas the E. colienzyme bridges the gap between aerobic respiration and fer-
mentation over the pO2 range in which both pathways arepoorly expressed (5).
The M. smegmatis cydA mutant was also significantly growthimpaired relative to its parental wild type when the two strainswere cocultured in the presence of 1 mM cyanide at 21% airsaturation. This observation suggests that, in addition to therelatively cyanide-insensitive bd oxidase (9, 31), M. smegmatiscontains a cyanide-sensitive terminal(s) that plays a role inaerobic respiration. The fact that the mutant displayed a com-petitive growth disadvantage compared to the wild type undermoderate aeration (21% air saturation) in the presence ofcyanide may be indicative of a respiratory role for the bdoxidase even under conditions of high O2, particularly since cydgene expression was detectable at this pO2. However, we can-not exclude the possibility that bd oxidase function under theseconditions is a compensatory response occurring only whenother terminals are inhibited by cyanide.
To place our results on the bd oxidase of M. smegmatis in thecontext of aerobic respiratory branches in mycobacteria, weanalyzed the preliminary genome sequence of this organismfor its repertoire of respiratory genes and compared it to thoseof M. tuberculosis and B. subtilis. A schematic representation ofthe branched electron transport chains in these organisms isshown in Fig. 7. The well-studied, aerobic respiratory chain ofB. subtilis is the most complex, terminating with cytochromesbd (cydABDC), cytochrome caa3 (ctaCDEF), cytochrome aa3
(qoxABCD), or cytochrome bd� (ythAB) (41, 64). There is alsoevidence for a bb�-type terminal oxidase in this organism (2),which appears to be bd-type, with the heme-d substituted byhigh spin heme-b. Therefore, terminal branches of the respi-ratory chain cannot be inferred solely from the genome se-quence, since macromolecular reorganization of terminal oxi-dase subunits and prosthetic groups could give rise to alternateoxidases. In B. subtilis at least one quinol oxidase, cytochromebd or cytochrome aa3, is required for aerobic growth, theythAB genes encoding a putative quinol oxidase are involved insporulation, and the cytochrome c oxidase branch is not ex-pressed until the stationary phase (64).
Other than the early studies of Todd (56), which confirmed
FIG. 5. Spectral characterization of membrane fractions isolated from M. smegmatis grown in an oxystat. The spectral profiles of purifiedmembrane fractions from M. smegmatis grown at 21% (A) and 1% (B) air saturation are shown. Reduction was accomplished by the addition ofsodium dithionite, and oxidation was accomplished by the addition of potassium ferricyanide.
FIG. 6. Cyanide inhibition of wild-type and cydA::aph strains underoxystatic conditions. The growth of mixed cultures of mc2155 (F) andcydA::aph (E) in the presence of 1 mM KCN are shown. KCN wasadded 30 min postinoculation, and the growth of each strain wasmonitored by differential plating as described in Materials and Meth-ods. The data are shown as the averages and standard errors from twoindependent experiments.
VOL. 183, 2001 M. SMEGMATIS CYTOCHROME bd OXIDASE 7083
on Novem
ber 17, 2014 by guesthttp://jb.asm
.org/D
ownloaded from

that mycobacteria contained cytochromes and the subsequentdemonstration of a complex containing cytochromes aa3 and oin Mycobacterium phlei (43), the respiratory chains of myco-bacteria are poorly understood. The aerobic respiratory chainof M. tuberculosis deduced from its genome sequence (7) is farsimpler than that of B. subtilis, terminating in only two mainbranches, namely, cytochrome bd and an aa3-type cytochromec oxidase belonging to the heme-copper respiratory oxidasesuperfamily (Fig. 7). Assignment of this aa3-type oxidase as acytochrome c oxidase rather than a quinol oxidase is based onthe presence in subunit II (CtaC) of a CuA binding site (16).The M. smegmatis respiratory chain is of intermediate com-plexity, possessing the same two branches as M. tuberculosisand a third possible branch terminating in the YthAB quinoloxidase. The other notable difference is the presence of twodistinct ctaD alleles in M. smegmatis versus one in M. tubercu-losis. We therefore speculate that alternate isoforms of cyto-chrome c oxidase may exist in M. smegmatis, as has been
proposed for Paracoccus denitrificans, which also containsctaDI and ctaDII alleles (44).
The cytochrome c branch of mycobacteria has some unusualfeatures that are shared by other high-G�C-content gram-positive bacteria (54). The only identifiable c-type cytochromein M. smegmatis and M. tuberculosis is the di-heme c-typecytochrome QcrC, which, like its counterpart from Corynebac-terium glutamicum, is a tandem product of two class I cyto-chromes c (54). This cytochrome has been proposed to acceptelectrons from the Rieske Fe-S cluster and to donate them toa terminal oxidase (54), suggesting that the quinol cytochromec reductase (QcrCAB) may form a complex with cytochrome coxidase (CtaCDE) in mycobacteria and related organisms (Fig.7). Interestingly, the qcrCAB operon in M. tuberculosis and M.smegmatis is located immediately downstream of ctaE, suggest-ing that these genes may be coordinately regulated.
In summary, we have shown that the cydAB-encoded quinoloxidase plays a particularly important respiratory role in M.smegmatis under conditions of extreme microaerophilia. Al-though the bd oxidase might also operate under moderatelyaerobic conditions, we propose that the major oxidase usedunder these conditions is the cytochrome c oxidase. Experi-ments aimed at testing this hypothesis and further elucidatingthe respiratory chains of M. smegmatis are currently under wayin our laboratories. In addition, the confirmed respiratory rolefor the M. smegmatis bd oxidase in microaerophilic growth hasobvious implications for studies of respiration in M. tuberculo-sis. In this respect, in vivo phenotyping of recently constructedcyd mutant strains of M. tuberculosis (S. S. Dawes and V.Mizrahi, unpublished data) should reveal whether loss of thebd oxidase affects the virulence of this organism.
ACKNOWLEDGMENTS
This work was supported by grant AI-43420 (to H.R.) from theNational Institutes of Health and by grants from the Medical ResearchCouncil of South Africa, the DACST-MRC Lead Programme for Tu-berculosis, the National Research Foundation of South Africa, and theSouth African Institute for Medical Research (to V.M.). V.M was alsosupported by an International Scholar’s grant from the HowardHughes Medical Institute, and B.K. was supported by an NRF Presti-gious bursary.
We thank the Council for Scientific and Industrial Research for theuse of the Braun fermentor and are indebted to Paul Axelsen forallowing us the use of his Cary spectrophotometer. We also thankBrian James, as well as Bhavna Gordhan and other members of theMizrahi lab, for advice and assistance; Selwyn Quan for providingp6.11; Peadar O’Gaora for pOLYG and pSMT3; and Martin Everettfor providing the gridded plasmid library and for recovering clones.We thank Piet Becker of the South African MRC Biostatistics Unit forassistance with the statistical analysis of our data. Preliminary se-quence data was obtained from The Institute for Genomic Researchwebsite at http://www.tigr.org.
REFERENCES
1. Asano, A., and A. F. Brodie. 1965. Oxidative phosphorylation in fractionatedbacterial systems. 18. Phosphorylation coupled to different segments of therespiratory chains of Mycobacterium phlei. J. Biol. Chem. 240:4002–4010.
2. Azarkina, N., S. Siletsky, V. Borisov, C. Von Wachenfeldt, L. Hederstedt, andA. Konstantinov. 1999. A cytochrome bb�-type quinol oxidase in Bacillussubtilis strain 168. J. Biol. Chem. 274:32810–32817.
3. Balasubramanian, V., M. S. Pavelka, Jr., S. S. Bardarov, J. Martin, T. R.Weisbrod, R. A. McAdam, B. R. Bloom, and W. R. Jacobs, Jr. 1996. Allelicexchange in Mycobacterium tuberculosis with long linear recombination sub-strates. J. Bacteriol. 178:273–279.
4. Barnes, P. F., H. el-Hajj, S. Preston-Martin, M. D. Cave, B. E. Jones, M.Otaya, J. Pogoda, and K. D. Eisenach. 1996. Transmission of tuberculosis
FIG. 7. Proposed aerobic respiratory pathways in M. smegmatis andrelated organisms. The pathways in B. subtilis were adapted fromreferences 41 and 64. The M. tuberculosis pathways were deduced fromits genome sequence (7), and those of M. smegmatis are based on thisstudy and analysis of the preliminary genome sequence data obtainedfrom The Institute for Genomic Research website (http://www.tigr.org). Solid arrows denote confirmed pathways, and dashed arrowsdenote putative pathways. The mycobacterial cytochrome reductasecomplex (QcrCAB) is denoted as “bcc” in accordance with the desig-nation of QcrC as cytochrome cc (54).
7084 KANA ET AL. J. BACTERIOL.
on Novem
ber 17, 2014 by guesthttp://jb.asm
.org/D
ownloaded from

among the urban homeless. JAMA 275:305–307.5. Becker, S., D. Vlad, S. Schuster, P. Pfeiffer, and G. Unden. 1997. Regulatory
O2 tensions for the synthesis of fermentation products in Escherichia coli andrelation to aerobic respiration. Arch. Microbiol. 168:290–296.
6. Boshoff, H. I. M., and V. Mizrahi. 1998. Purification, gene cloning, targetedknockout, overexpression, and biochemical characterization of the majorpyrazinamidase from Mycobacterium smegmatis. J. Bacteriol. 180:5809–5814.
7. Cole, S. T., R. Brosch, J. Parkhill, T. Garnier, C. Churcher, D. Harris, S. V.Gordon, K. Eiglmeier, S. Gas, C. E. Barry III, F. Tekaia, K. Badcock, D.Basham, D. Brown, T. Chillingworth, R. Connor, R. Davies, K. Devlin, T.Feltwell, S. Gentles, N. Hamlin, S. S. Holroyd, T. Hornsby, K. Jagels, A.Krogh, J. McLean, S. Moule, L. Murphy, K. Oliver, J. Osborne, M. A. Quail,M. A. Rajandream, J. Rogers, S. Rutter, K. Seeger, J. Skelton, S. Squares,J. E. Sulston, K. Taylor, S. Whitehead, and B. G. Barrell. 1998. Decipheringthe biology of Mycobacterium tuberculosis from the complete genome se-quence. Nature 393:537–544.
8. Cotter, P. A., S. B. Melville, J. A. Albrecht, and R. P. Gunsalus. 1997.Aerobic regulation of cytochrome d oxidase (cydAB) operon expression inEscherichia coli: roles of Fnr and ArcA in repression and activation. Mol.Microbiol. 25:605–615.
9. Cunningham, L., M. Pitt, and H. D. Williams. 1997. The cioAB genes fromPseudomonas aeruginosa code for a novel cyanide-insensitive terminal oxi-dase related to the cytochrome bd quinol oxidases. Mol. Microbiol. 24:579–591.
10. Cunningham, A. F., and C. L. Spreadbury. 1998. Mycobacterial stationaryphase induced by low oxygen tension: cell wall thickening and localization ofthe 16-kilodalton alpha-crystallin homolog. J. Bacteriol. 180:801–808.
11. Dick, T., B. H. Lee, and B. Murugasu-Oei. 1998. Oxygen depletion induceddormancy in Mycobacterium smegmatis. FEMS Microbiol. Lett. 163:159–164.
12. Dye, C., S. Scheele, P. Dolin, V. Pathania, and M. C. Raviglione. 1999.Consensus statement. Global burden of tuberculosis: estimated incidence,prevalence, and mortality by country. W.H.O. Global Surveillance and Mon-itoring Project. JAMA 282:677–686.
13. Edwards, S. E., C. S. Loder, G. Wu, H. Corker, B. W. Bainbridge, S. Hill, andR. K. Poole. 2000. Mutation of cytochrome bd quinol oxidase results inreduced stationary phase survival, iron deprivation, metal toxicity and oxi-dative stress in Azotobacter vinelandii. FEMS Microbiol. Lett. 185:71–77.
14. Fang, H., R. J. Lin, and R. B. Gennis. 1989. Location of heme axial ligandsin the cytochrome d terminal oxidase complex of Escherichia coli determinedby site-directed mutagenesis. J. Biol. Chem. 264:8026–8032.
15. Gangadharam, P. R. J. 1995. Mycobacterial dormancy. Tuber. Lung Dis.76:477–479.
16. Garcıa-Horsman, J. A., B. Barquera, J. Rumbley, J. Ma, and R. B. Gennis.1994. The superfamily of heme-copper respiratory oxidases. J. Bacteriol.176:5587–5600.
17. Gennis, R. B., and V. Stewart. 1996. Respiration, p. 217–261. In F. C.Neidhardt, R. Curtiss III, J. L. Ingraham, E. C. C. Lin, K. B. Low, B.Magasanik, W. S. Reznikoff, M. Riley, M. Schaechter, and H. E. Umbarger(ed.), Escherichia coli and Salmonella: cellular and molecular biology, 2nded. ASM Press, Washington, D.C.
18. Ghanekar, K., A. McBride, O. Dellagostin, S. Thorne, R. Mooney, and J.McFadden. 1999. Stimulation of transposition of the Mycobacterium tuber-culosis insertion sequence IS6110 by exposure to microaerobic environment.Mol. Microbiol. 33:982–993.
19. Goldman, B. S., K. K. Gabbert, and R. G. Kranz. 1996. The temperature-sensitive growth and survival phenotypes of Escherichia coli cydDC andcydAB strains are due to deficiencies in cytochrome bd and are corrected byexogenous catalase and reducing agents. J. Bacteriol. 178:6348–6351.
20. Gordhan, B. G., S. J. Andersen, A. R. De Meyer, and V. Mizrahi. 1996.Construction by homologous recombination and phenotypic characterizationof a polA mutant of Mycobacterium smegmatis. Gene 178:125–130.
21. Grange, J. M. 1992. The mystery of the mycobacterial persistor. Tuberc.Lung Dis. 73:249–251.
22. Huberts, P., and V. Mizrahi. 1995. Cloning and sequence analysis of the geneencoding the DNA polymerase I from Mycobacterium tuberculosis. Gene164:133–136.
23. Husson, R. N., B. E. James, and R. A. Young. 1990. Gene replacement andexpression of foreign DNA in mycobacteria. J. Bacteriol. 172:519–524.
24. Imboden, P., and G. K. Schoolnik. 1998. Construction and characterizationof a partial Mycobacterium tuberculosis cDNA library of genes expressed atreduced oxygen tension. Gene 213:107–117.
25. Jones, C. W., and R. K. Poole. 1985. The analysis of cytochromes, p. 285–328.In G. Gottschalk (ed.), Methods in microbiology. Academy Press, London,England.
26. Junemann, S., and J. M. Wrigglesworth. 1995. Cytochrome bd oxidase fromAzotobacter vinelandii. J. Biol. Chem. 270:16213–16220.
27. Junemann, S. 1997. Cytochrome bd terminal oxidase. Biochim. Biophys.Acta 1321:107–127.
28. Juty, N. S., F. Moshiri, M. Merrick, C. Anthony, and S. Hill. 1997. TheKlebsiella pneumoniae cytochrome bd terminal oxidase complex and its rolein microaerobic nitrogen fixation. Microbiology 143:2673–2683.
29. Kaysser, T. M, J. B. Ghaim, C. Georgiou, and R. B. Gennis. 1995. Methio-
nine-393 is an axial ligand of the heme b558 component of the cytochromebd ubiquinol oxidase from Escherichia coli. Biochemistry 34:13491–13501.
30. Kelly, M. J. S., R. K. Poole, M. G. Yates, and C. Kennedy. 1990. Cloning andmutagenesis of genes encoding the cytochrome bd terminal oxidase complexin Azotobacter vinelandii: mutants deficient in the cytochrome d complex areunable to fix nitrogen in air. J. Bacteriol. 172:6010–6019.
31. Kita, K., K. Konishi, and Y. Anraku. 1986. Purification and properties of twoterminal oxidase complexes of Escherichia coli aerobic respiratory chain.Methods Enzymol. 126:94–113.
32. Kusaka, T., R. Sato, and K. Shoji. 1964. Comparison of cytochromes inmycobacteria grown in vitro and in vivo. J. Bacteriol. 87:1383–1388.
33. Kusumoto, K., M. Sakiyama, J. Sakamoto, S. Noguchi, and N. Sone. 2000.Menaquinol oxidase activity and primary structure of cytochrome bd fromthe amino-acid fermenting bacterium Corynebacterium glutamicum. Arch.Microbiol. 173:390–397.
34. Le, H. Q., and P. T. Davidson. 1996. Reactivation and exogenous infection:their relative roles in the pathogenesis of tuberculosis. Curr. Clin. Top.Infect. Dis. 16:260–276.
35. Lee, B. H., B. Murugasu-Oei, and T. Dick. 1998. Upregulation of a histone-like protein in dormant Mycobacterium smegmatis. Mol. Gen. Genet. 260:475–479.
36. Lorence, R. M., J. G. Koland, and R. B. Gennis. 1986. Coulometric andspectroscopic analysis of the purified cytochrome d complex of Escherichiacoli: evidence for the identification of “cytochrome a1” as cytochrome b595.Biochemistry 25:2314–2321.
37. O’Gaora, P., S. Barnini, C. Hayward, E. Filley, G. Rook, D. Young, andJ. Thole. 1997. Mycobacteria as immunogens: development of expressionvectors for use in multiple mycobacterial species. Med. Princ. Pract. 6:91–96.
38. Osborne, J. P., and R. B. Gennis. 1999. Sequence analysis of cytochrome bdoxidase suggests a revised topology for subunit I. Biochim. Biophys. Acta1410:32–50.
39. Parish, T. A., B. G. Gordhan, R. A. McAdam, K. Duncan, V. Mizrahi, andN. G. Stoker. 1999. Production of mutants in amino acid biosynthesis genesof Mycobacterium tuberculosis by homologous recombination. Microbiology145:3497–3503.
40. Parish, T., and N. G. Stoker. 2000. Use of a flexible cassette method togenerate a double unmarked Mycobacterium tuberculosis tlyA plcABC mutantby gene replacement. Microbiology 146:1969–1975.
41. Poole, R. K., and G. Cook. 2000. Redundancy of aerobic respiratory chainsin Bacteria? Routes, reasons, and regulation. Adv. Microb. Physiol. 43:165–224.
42. Primm, T. P., S. J. Andersen, V. Mizrahi, D. Avarbock, H. Rubin, and C. E.Barry III. 2000. The stringent response of Mycobacterium tuberculosis isrequired for long-term survival. J. Bacteriol. 182:4889–4898.
43. Revsin, B., E. D. Marquez, and A. F. Brodie. 1970. Cytochromes fromMycobacterium phlei: isolation and spectral properties of a mixture of cyto-chromes (a � a3) (o). Arch. Biochem. Biophys. 139:114–120.
44. Raitio, M., J. M. Pispa, T. Metso, and M. Saraste. 1990. Are there isoen-zymes of cytochrome c oxidase in Paracoccus denitrificans? FEBS Lett. 261:431–435.
45. Sakamoto, J., A. Matsumoto, K. Oobuchi, and N. Sone. 1996. Cytochromebd-type quinol oxidase in a mutant of Bacillus stearothermophilus deficient incaa3-type cytochrome c oxidase. FEMS Microbiol. Lett. 143:151–158.
46. Sakamoto, J., E. Koga, T. Mizuta, C. Sato, S. Noguchi, and N. Sone. 1999.Gene structure and quinol oxidase activity of a cytochrome bd-type oxidasefrom Bacillus stearothermophilus. Biochim. Biophys. Acta 1411:147–158.
47. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning: alaboratory manual, 2nd ed. Cold Spring Harbor Laboratory, Cold SpringHarbor, N.Y.
48. Segal, W. 1984. Growth dynamics of in vivo and in vitro grown mycobacterialpathogens, p. 547–573. In G. P. Kubica and L. G. Wayne (ed.), The Myco-bacteria—a sourcebook. Marcel Dekker, New York, N.Y.
49. Sherman, D. R., M. Voskuil, D. Schnappinger, R. Liao, M. I. Harrell, andG. K. Schoolnik. 2001. Regulation of the Mycobacterium tuberculosis hypoxicresponse gene encoding �-crystallin. Proc. Natl. Acad. Sci. USA 98:7534–7539.
50. Siegele, D. A., and R. Kolter. 1993. Isolation and characterization of anEscherichia coli mutant defective in resuming growth after starvation. GenesDev. 7:2629–2640.
51. Siegele, D. A., K. R. C. Imlay, and J. A. Imlay. 1996. The stationary-phase-exit defect of cydC (surB) mutants is due to the lack of a functional terminalcytochrome oxidase. J. Bacteriol. 178:6091–6096.
52. Smeulders, M. J., J. Keer, R. A. Speight, and H. D. Williams. 1999. Adap-tation of Mycobacterium smegmatis to stationary phase. J. Bacteriol. 181:270–283.
53. Snapper, S. B., R. E. Melton, S. Mustafa, T. Keiser, and W. R. Jacobs, Jr.1990. Isolation and characterization of efficient plasmid transformation mu-tants of Mycobacterium smegmatis. Mol. Microbiol. 4:1911–1919.
54. Sone, N., K. Nagata, H. Kojima, J. Tajima, Y. Kodera, T. Kanamaru, S.Noguchi, and J. Sakamoto. 2001. A novel hydrophobic diheme c-type cyto-chrome: purification from Corynebacterium glutamicum and analysis of theqcrCBA operon encoding three subunit proteins of a putative cytochrome
VOL. 183, 2001 M. SMEGMATIS CYTOCHROME bd OXIDASE 7085
on Novem
ber 17, 2014 by guesthttp://jb.asm
.org/D
ownloaded from

reductase complex. Biochim. Biophys. Acta 1503:279–290.55. Starke, J. R. 1995. Tuberculosis in children. Curr. Opin. Pediatr. 7:268–277.56. Todd, C. M. 1949. Occurrence of cytochrome and coproporphyrin in myco-
bacteria. Biochem. J. 45:386–390.57. Tseng, C. P., J. Albrecht, and R. P. Gunsalus. 1996. Effect of microaerophilic
cell growth conditions on expression of the aerobic (cyoABCDE and cydAB)and anaerobic (narGHJI, frdABCD, and dmsABC) respiratory pathway genesin Escherichia coli. J. Bacteriol. 178:1094–1098.
58. Way, S. S., S. Sallustio, R. S. Magliozzo, and M. B. Goldberg. 1999. Impactof either elevated or decreased levels of cytochrome bd expression on Shi-gella flexneri virulence. J. Bacteriol. 181:1229–1237.
59. Wayne, L. G., and G. A. Diaz. 1967. Autolysis and secondary growth ofMycobacterium tuberculosis in submerged culture. J. Bacteriol. 93:1374–1381.
60. Wayne, L. G., and K. Y. Lin. 1982. Glyoxylate metabolism and adaptation ofMycobacterium tuberculosis to survival under anaerobic conditions. Infect.Immun. 37:1042–1049.
61. Wayne, L. G. 1994. Dormancy of Mycobacterium tuberculosis and latency of
disease. Eur. J. Clin. Microbiol. Infect. Dis. 13:908–914.62. Wayne, L. G., and L. G. Hayes. 1996. An in vitro model for sequential study
of shiftdown of Mycobacterium tuberculosis through two stages of nonrepli-cating persistence. Infect. Immun. 64:2062–2069.
63. Winstedt, L., K. Yoshida, Y. Fujita, and C. von Wachenfeldt. 1998. Cyto-chrome bd biosynthesis in Bacillus subtilis: characterization of the cydABCDoperon. J. Bacteriol. 180:6571–6580.
64. Winstedt, L., and C. von Wachenfeldt. 2000. Terminal oxidases of Bacillussubtilis strain 168: one quinol oxidase, cytochrome aa3 or cytochrome bd, isrequired for aerobic growth. J. Bacteriol. 182:6557–6564.
65. Ye, R. W., W. Tao, L. Bedzyk, T. Young, M. Chen, and L. Li. 2000. Globalgene expression profiles of Bacillus subtilis grown under anaerobic condi-tions. J. Bacteriol. 182:4458–4465.
66. Young, D. B., and K. Duncan. 1995. Prospects for new interventions in thetreatment and prevention of mycobacterial disease. Annu. Rev. Microbiol.49:641–673.
7086 KANA ET AL. J. BACTERIOL.
on Novem
ber 17, 2014 by guesthttp://jb.asm
.org/D
ownloaded from




















