
Characterization of Endocrine Disrupting Effects of Bisphenol ...
241
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of Characterization of Endocrine Disrupting Effects of Bisphenol ...


Characterization of Endocrine Disrupting Effects of Bisphenol A or 17α-Ethinyl Estradiol in Mouse Uterus
A dissertation submitted to the
Graduate School
of the University of Cincinnati
in partial fulfillment of the
requirements for the degree of
Doctor of Philosophy
in the Department of Pharmacology and Cell Biophysics
of the College of Medicine
by
Jessica A. Kendziorski
BA, DePauw University, 2010
Committee Chair: Scott M. Belcher, PhD

ii
ABSTRACT
Bisphenol A (BPA) and 17α-ethinyl estradiol (EE) are estrogenic endocrine disrupting
chemicals (EDCs) that adversely affect the structure and function of the uterus. Humans are
ubiquitously exposed to BPA via consumption of contaminated food and beverage from
polycarbonate packaging or food cans. Humans are exposed to EE through the use of oral
contraceptives. While human exposure to these EDCs is widespread, little is known how these
compounds alter physiological responses that lead to increased incidence of uterine pathology.
The purpose of this dissertation was to investigate and characterize uterine pathologies
and associated alterations in immune responsiveness and fibrosis. Two mouse strains were used,
the CD1 and the C57Bl/6N strains. Exposure during adulthood included mating, parturition, and
offspring rearing and mice were exposed for 12-15 weeks. Whole life exposure included
placental transfer, as well as direct oral consumption, exposing mice until postnatal day (PND)
90. In order to closely mimic human exposure, mice were orally exposed through diet to known
concentrations of control (0 ppm), BPA (0.03, 0.3, 3, 30, or 300 ppm), or EE (0.0001, 0.001,
0.01, 0.1, or 1.3 ppm), which resulted in calculated BPA doses of 0.04, 0.4, 4, 40, and 400
mg/kg/day and EE doses of 0.00002, 0.0002, and 0.001 mg/kg/day. Other exogenous estrogenic
compounds from housing, bedding, and water were eliminated.
Two distinct immune-related and fibrotic uterine pathologies were identified. Pyometra
developed in C57Bl/6N mice exposed to BPA or EE during adulthood. An equine endometrosis-
like phenotype, characterized by increased gland nests and stromal and periglandular fibrosis,
naturally occurred in control C57Bl/6N mice. In CD1 mice, this fibrotic phenotype was present
in the 30 ppm BPA group. Exposure to BPA significantly increased Col1a1 and Col3a1
expression and decreased Mmp2 and Timp2 expression. Expression and activity of matrix
metalloproteinases, MMP2 and MMP14, were significantly decreased in both the control

iii
C57Bl/6N and 30 ppm BPA-exposed CD1 mice compared to control CD1 mice. However, both
strains presented with an increased immune response, quantified as the percentage of F4/80-
positive cells. The C57Bl/6N strain had a significant increase in endometrial macrophages at a
lower dose of BPA (0.03 ppm) than the CD1 strain (30 ppm) exposed during adulthood. In both
exposure models, CD1 mice had increased macrophages in the 30 ppm BPA group.
This dissertation research was the first report of BPA altering the immune response or
collagen accumulation in the uterus, leading to the induction of pyometra or an equine
endometrosis-like phenotype. It was also the first report of strain-specific differences in
sensitivity to the actions of BPA on these endpoints of interest. Finally, this research highlighted
the potential for exposure to estrogenic compounds such as BPA and EE to impact the
development and progression of fibrotic and immune-related uterine diseases. However, based
on the differences observed between the two generations, physiological changes that occur
during mating, pregnancy, and parturition may be necessary for estrogenic compounds to elicit
fibrotic effects. These studies also emphasized the importance of understanding differences in
sensitivity to estrogenic compounds between mouse strains.

iv

v
ACKNOWLEDGEMENTS
This dissertation work is the product of the encouragement of several colleagues, family,
and friends. First and foremost, I would like to acknowledge my advisor and mentor, Dr. Scott
Belcher, for his support and assistance during my time as a graduate student. Through his
invaluable mentoring and challenges, I have grown significantly in confidence as a scientist and
a person. I am truly grateful for the guidance and support he has provided over the years.
I would like to extend gratitude for the encouragement and constructive advice given by
my dissertation committee members, Drs, Jo El Schultz, William J. Ball, and Heather Patisaul.
This dissertation would not have been completed without their attention to progress and helping
me to remain focused on the final product.
I am gratefully indebted to the past and present members of the Belcher lab, specifically
Dr. Eric Kendig, Robin Gear, Dana Buesing, Susie Christie, Dr. Vinicius Carreira, and Clifford
Cookman, for their suggestions on experimental procedures, their help in conducting
experiments, and their friendships in general during my time in graduate school. I would like to
especially acknowledge Vini for his help with identifying pathologies, which laid the foundation
for a great part of this dissertation research.
I would like to acknowledge rotating graduate students, undergraduate students, summer
students, and students from England for their contributions to this research and for their
friendship, especially Joni Ford, Rachel Steinher, Ed Durley, and Kev Ungi. Their willingness to
do whatever was asked of them was highly appreciated.
I would like to thank all of the faculty, students, and staff in the Department of
Pharmacology and Biophysics and the associated graduate program. I would like to specifically
acknowledge Nancy Thyberg for everything that she has done, by keeping me updated on

vi
deadlines, helping to organize committee meetings, and being the person I would go to for any
questions. If she didn’t have the answer, she would find someone that could answer it.
Finally, I would like to thank my family and friends for their encouragement, love, and
support. Even though they had no idea what I was doing or why I was in school for 5+ years, my
parents always asked how things were going. I would like to thank my sister Sarah for always
being able to cheer me up if I was having a rough time. I would like to thank my friends, in
particular Ian, Jamey, Natalie, Josh, Mat, Leslie, Chad, Cassie, Mason, and Kyle, for their ability
to always make sure I had fun and didn’t go too crazy from being in the lab, and my boyfriend
Carson for putting up with my complaining and for his constant reassurance and love. I would
also like to thank Megan, Chenoa, Andrea, and Kelly for their laughs. Even though we were
hundreds of miles apart, they made me smile and uplifted my spirits every time we texted or
called.

vii
TABLE OF CONTENTS Page
ABSTRACT ii
ACKNOWLEDGEMENTS v
TABLE OF CONTENTS vii
LIST OF FIGURES AND TABLES xiv
LIST OF ABBREVIATIONS xviii
Chapter 1 INTRODUCTION 1
1.1 Statement of Purpose 2
1.1.1 Objective and Rationale 2
1.1.2 Central Hypothesis and Specific Aims 2
1.2 Definition of Endocrine Disrupting Chemicals 3
1.3 Prototypical Estrogenic EDC: Diethylstilbestrol 4
1.4 17α-Ethinyl Estradiol 6
1.5 Bisphenol A 7
1.5.1 Commercial Uses of BPA and Routes of Exposure 7
1.5.2 Pharmacokinetics and Toxicologic Exposure Limits of BPA 8
1.5.3 Receptor Binding of BPA 8
1.5.4 Endocrine Disrupting Effects of BPA in the Uterus 11
1.5.5 Extracellular Matrix Alterations in Response to BPA 12
1.6 Strain Susceptibility to Effects of Estrogenic Compounds 13
1.6.1 Susceptibility to Uterine Weight Increases and Morphological Alterations 15
1.6.2 Differences in Circulating Sex Hormones and ERα Levels 15
1.6.3 Quantitative Trait Loci Controlling Phenotypic Variation 16

viii
1.6.4 Differential Sensitivity of the Ovary and Vagina to Estrogenic Compounds 17
1.6.5 Differential Sensitivity of Other Tissues to Estrogenic Compounds 18
1.7 The Endocrine System 19
1.8 Nuclear Hormone Receptors: ERα and ERβ 20
1.8.1 Structure 20
1.8.2 Nuclear Transactivation, Membrane-Initiated, 22
and Rapid Signaling Mechanisms
1.8.3 Role of ERs in Uterine Structure and Reproductive Function 26
1.8.3.1 Postnatal Uterine Development 26
1.8.3.2 Impact on Gene Expression 27
1.8.3.3 ERα in the Brain and Impact on Cyclicity 28
1.8.3.4 Role of GPR30 in the Uterus 28
1.9 The Cycling and Pregnant Uterus 29
1.9.1 Circulating Hormones and Tissue Remodeling 29
1.9.2 Role of Extracellular Matrix in Uterine Structure and Function 32
1.9.3 Role of Extracellular Matrix during Pregnancy 36
1.10 Uterine Disorders 39
1.10.1 Pyometra 39
1.10.1.1 Description, Etiology, and Treatment of Pyometra 39
1.10.1.2 Role of Bacteria and the Immune System 40
in the Development of Pyometra
1.10.1.3 Involvement of Sex Hormones and Collagen Accumulation 41
in Pyometra

ix
1.10.2 Endometriosis 41
1.10.2.1 Description, Symptoms, and Treatment of Endometriosis 41
1.10.2.2 Role of Extracellular Matrix in Pathogenesis of Endometriosis 42
1.10.3 Leiomyoma 43
1.10.3.1 Description, Symptoms, and Treatment of Leiomyoma 43
1.10.3.2 Etiology of Leiomyoma 45
1.10.3.3 Regulation of Collagen Accumulation in Leiomyoma 47
1.10.4 Equine Endometrosis 47
1.10.4.1 Description, Etiology, and Treatment of Equine Endometrosis 47
1.10.4.2 Progression of Equine Endometrosis 50
1.10.4.3 Characterization of Extracellular Matrix Alterations 51
in Equine Endometrosis
1.10.4.4 Fetal Foal Loss 52
1.11 Statement of Purpose 54
1.11.1 Objective and Rationale 54
1.11.2 Central Hypothesis and Specific Aims 54
Chapter 2 Methods 55
2.1 BPA and EE Exposure 56
2.1.1 Animal Housing 56
2.1.2 Exposure 56
2.1.3 Breeding 59
2.1.4 Vaginal Cytology 59
2.2 Assessment of Pathology 60

x
2.2.1 Necropsy and Tissue Preparation 60
2.2.2 Hematoxylin and Eosin Staining 61
2.2.3 Myometrial Wall Thickness 62
2.2.4 Histologic Assessment of Uterine Pathology 62
2.3 Assessment of Collagen Synthesis and Degradation and Immune Response 63
2.3.1 Picrosirius Red Staining and Collagen Accumulation 63
2.3.2 Immunohistochemistry 63
2.3.3 Quantitative RT-PCR 66
2.3.4 SDS-PAGE and Immunoblotting 70
2.3.5 Gelatin Zymography 72
2.4 Statistical Methods 74
Chapter 3 Strain Specific Induction of Pyometra and Differences in Immune 75
Responsiveness in Mice Exposed to 17α-Ethinyl Estradiol or the
Endocrine Disrupting Chemical Bisphenol A
3.1 Abstract 77
3.2 Introduction 78
3.3 Materials and Methods 80
3.3.1 Animal Husbandry and Dietary Exposures 80
3.3.2 Monitoring of Estrous 81
3.3.3 Necropsy and Tissue Preparation 82
3.3.4 Immunohistochemistry 82
3.3.5 Statistical Analysis 83
3.4 Results 84

xi
3.4.1 Diet Consumption 84
3.4.2 Effects on Body Weight 86
3.4.3 Fertility and Fecundity 88
3.4.4 Necropsy and Organ Weights 90
3.4.5 Uterine Pathology: C57Bl/6 Pyometra 92
3.4.6 Uterine Pathology: Effects on Uterine Morphology and 94
Macrophage Infiltration
3.5 Discussion 99
3.6 Acknowledgements 103
3.7 Supplemental Results 104
3.7.1 Uterine Morphology: Myometrial Wall Thickness 104
3.7.2 Immune Response in Uteri of CD1 and C57Bl/6N Mice 105
Chapter 4 Strain-Specific Induction of Endometrial Periglandular Fibrosis in Mice 108
Exposed During Adulthood to the Endocrine Disrupting Chemical Bisphenol A
4.1 Abstract 110
4.2 Introduction 111
4.3 Materials and Methods 116
4.3.1 Animal Husbandry and Necropsy 116
4.3.2 Histology and Immunohistochemistry 117
4.3.3 Picrosirius Red Stain and Collagen Assessment 118
4.3.4 Quantitative RT-PCR 119
4.3.5 SDS-PAGE and Immunoblotting 119
4.3.6 Gelatin Zymography 120

xii
4.3.7 Statistical Analysis 121
4.4 Results 122
4.4.1 Uterine Pathology: Gland Nests and Periglandular Fibrosis in C57Bl/6N 122
and CD-1 Mice
4.4.2 Analysis of Collagen and MMP mRNA Expression 126
4.4.3 Analysis of Protein Expression and Activity of MMPs 127
4.4.4 Immunolocalization of MMP2 and MMP14 in the Endometrium 128
4.4.5 Immunohistochemical Characterization of an Endometrosis-Like Phenotype 131
4.5 Discussion 133
4.6 Acknowledgements 139
Chapter 5 Effects of Bisphenol A and 17α-Ethinyl Estradiol in Nulligravida Uterus 140
of CD1 and C57Bl/6N Mice: Whole Life Exposure
5.1 Introduction 141
5.2 Methods 143
5.2.1 Exposure 143
5.2.2 Myometrial Wall Thickness 143
5.2.3 Uterine Pathology 144
5.2.4 Assessment of Collagen Accumulation and Immune Response 144
5.2.5 Statistical Analysis 145
5.3 Results 146
5.3.1 Uterine Morphology: Myometrial Wall Thickness 146
5.3.2 Uterine Pathology 147
5.3.3 Collagen Accumulation 148

xiii
5.3.4 Quantification of Gland Nest Density 150
5.3.5 Immune Response 151
5.4 Discussion 153
Chapter 6 SUMMARY AND CONCLUSIONS 156
6.1 Significance 157
6.2 Limitations 160
6.3 Future Directions 161
LIST OF PUBLICATIONS AND ABSTRACTS 167
BIBLIOGRAPHY 169

xiv
LIST OF FIGURES AND TABLES Page
FIGURES
Chapter 1 INTRODUCTION
Figure 1 Chemical structure, name, and CAS ID of estrogenic compounds 3
Figure 2 HPG axis and negative feedback loop 20
Figure 3 Expression of ERα and ERβ in human tissues 21
Figure 4 Schematic of domain structure of ERα and ERβ 22
Figure 5 Classical model of nuclear hormone receptor translocation 23
Figure 6 Normal uterine structure of adult CD1 mouse 30
Figure 7 Simplified representation of the human menstrual cycle 31
Figure 8 Regulation of collagen synthesis and degradation 33
Figure 9 Schematic representation of MMP structures 35
Figure 10 Schematic of decidualization and implantation in the human uterus 37
Chapter 2 METHODS
Figure 1 Exposure paradigm for sires, dams, and offspring in control and BPA or 57
EE exposure groups
Chapter 3 Pyometra and Immune Responsiveness
Figure 1 Histological and immunohistochemical assessments of C57Bl/6 uterus 93
affected by severe pyometra
Figure 2 Effects of oral exposure to BPA on uterine morphology and macrophage 95
infiltration in C57Bl/6 mice without overt pyometra
Figure 3 Effects of oral exposure to BPA on uterine morphology and macrophage 97

xv
infiltration in CD1 mice
Figure 4 Quantitative comparison of F4/80 immunoreactive cell numbers in the uterus 98
of control and BPA-exposed C57Bl/6 and CD1 mice
Figure 5 Myometrial wall thickness in mice across exposure groups and strain 105
Figure 6 Quantification of F4/80-positive cells in BPA- or EE-exposed CD1 and 107
C57Bl/6N adult mice
Chapter 4 Equine Endometrosis-Like Phenotype and Fibrosis
Figure 1 Effects of oral exposure to BPA or EE on uterine morphology and 123
collagen accumulation in C57Bl/6N mice
Figure 2 Effects of oral exposure to BPA or EE on uterine morphology and 124
collagen accumulation in CD-1 mice
Figure 3 Quantification of gland nest density in the uteri of control and 125
BPA-exposed CD-1 and C57Bl/6N mice
Figure 4 Relative expression levels and activity of mRNA and proteins involved in 126
collagen synthesis and degradation
Figure 5 Strain differences in the immunohistochemical localization of MMP2 and 130
MMP14 in the uteri of control CD-1 and C57Bl/6N mice
Figure 6 Immunohistochemical characterization of endometrosis-like phenotype 131
across BPA exposure groups or between strains
Chapter 5 Effects of BPA and EE in Uterus: Whole Life Exposure
Figure 1 Exposure paradigm for sires, dams, and offspring in control and BPA or 143
EE exposure groups

xvi
Figure 2 Myometrial wall thickness in CD1 mice across exposure groups 146
Figure 3 Effects of oral BPA or EE exposure on uterine morphology 150
and collagen accumulation in CD1 offspring
Figure 4 Quantification of gland nest density in the endometrium 151
of BPA- and EE-exposed CD1 and C57Bl/6N mice at PND90
Figure 5 Quantification of F4/80-positive cells in BPA- or EE-exposed CD1 offspring 152
TABLES
Chapter 1 INTRODUCTION
Table 1 Summary of studies examining sensitivity of rodent strains to 14
estrogenic compounds
Table 2 Characteristics of equine endometrosis phenotypes 51
Chapter 2 METHODS
Table 1 Composition of defined control and test diets 58
Table 2 Amounts of EE, BPA, and dyes incorporated into diets 58
Table 3 Primary and secondary antibodies 66
Table 4 RNA concentrations 68
Table 5 TaqMan probe sets used for qRT-PCR analysis 69
Chapter 3 Pyometra and Immune Responsiveness
Table 1 Estimated food consumption, caloric intake, and dose 85
Table 2 Female body weight 87
Table 3 Measures of fertility and fecundity 89
Table 4 Organ weights 91

xvii
Chapter 4 Equine Endometrosis-Like Phenotype and Fibrosis
Table 1 Incidence of uterine pathology in CD-1 and C57Bl/6N mice 122
at 19-23 weeks of age
Chapter 5 Effects of BPA and EE in Uterus: Whole Life Exposure
Table 1 Uterine pathology in CD1 and C57Bl/6N offspring 149
Chapter 6 SUMMARY AND CONCLUSIONS
Table 1 Comparison of relative expression and activity of collagens and MMPs 159
in mouse strains

xviii
LIST OF ABBREVIATIONS
11β-HSD2 11β-hydroxysteroid dehydrogenase
α-MG α-macroglobulin
α-SMA α-smooth muscle actin
ABC avidin-biotin complex
AF1 ligand-independent activation factor 1
AF2 ligand-dependent activation factor 2
ANOVA analysis of variance
APS ammonium persulfate
AR androgen receptor
BM basement membrane
BPA bisphenol A
BSA bovine serum albumin
CAL calbindinD9k
cAMP cyclic adenosine monophosphate
CEH cystic endometrial hyperplasia
CO2 carbon dioxide
DAB 3’3-diaminobenzidine
DBD DNA binding domain
DES diethylstilbestrol
dH2O distilled water
DMSO dimethyl sulfoxide
E2 17β-estradiol
eAD-MSC equine adipose derived-mesenchymal stem cell
ECM extracellular matrix

xix
EDC endocrine disrupting chemical
EE 17α-ethinyl estradiol
EGFR epidermal growth factor receptor
Endo endometrium
eNOS endothelial nitric oxide synthase
EPF endometrial periglandular fibrosis
ER estrogen receptor
ERα estrogen receptor α
ERβ estrogen receptor β
ERE estrogen response element
ERK extracellular signal-regulated kinase
ERR estrogen-related receptor
ERRα estrogen-related receptor α
ERRβ estrogen-related receptor β
ERRγ estrogen-related receptor γ
ERRE estrogen-related receptor response element
F0 parental generation
F4/80 glycoprotein marker on mouse macrophages
FDA US Food and Drug Administration
FSH follicle stimulating hormone
g gram
GH growth hormone
GnRH gonadotrophin releasing hormone
GPER1 G protein-coupled estrogen receptor 1
GPR30 G protein-coupled receptor 30

xx
H&E hematoxylin and eosin
H2O2 hydrogen peroxide
hCG human chorionic gonadotropin
HPG hypothalamus-pituitary-gonadal
HSP heat shock protein
IC50 half maximal inhibitory concentration
IL interleukin
IM inner myometrium
kcal kilocalorie
KD dissociation constant
kg kilogram
Kiss1 kisspeptin
LBD ligand binding domain
LE luminal epithelium
LH luteinizing hormone
LPS lipopolysaccharide
LRP low density lipoprotein receptor-related protein
µg microgram
µL microliter
µm micrometer
µM micromolar
M molar
MAPK mitogen-activated protein kinase
MED12 mediator complex subunit 12
mg milligram

xxi
mL milliliter
mm millimeter
mM millimolar
MMP matrix metalloproteinase
MR magnetic resonance
MSC mesenchymal stem cell
Myo myometrium
n sample size
NBF neutral buffered formalin
NET neutrophil extracellular trap
NHR nuclear hormone receptor
NIEHS National Institute of Environmental Health and Safety
ng nanogram
nM nanomolar
NO nitric oxide
NOAEL no observed adverse effect level
NTD N-terminal domain
OECD Organisation for Economic Co-operation and Development
OM outer myometrium
P4 progesterone
PBS phosphate buffered saline
PGC-1α peroxisome proliferator-activated receptor coactivator-1α
PGES prostaglandin E2 synthase
PGFS prostaglandin F2α synthase
PI3K phosphoinositide-3-kinase

xxii
PKA protein kinase A
PKB protein kinase B
pM picomolar
PND postnatal day
ppm parts per million
PR progesterone receptor
Prl prolactin
PTGS-2 prostaglandin-endoperoxide synthase
QTL quantitative trait loci
RBA relative binding affinity
RT room temperature
SD standard deviation
SDS sodium dodecyl sulfate
SDS-PAGE sodium dodecyl sulfate-polyacrylamide gel electrophoresis
SEM standard error of the mean
SNP single nucleotide polymorphism
T testosterone
TBS Tris-buffered saline
TBST Tris-buffered saline with Tween-20
TEMED tetramethylethylenediamine
TG2 tissue transglutaminase 2
TGF-β1 transforming growth factor-β1
THR thyroid hormone receptor
TIMP tissue inhibitor of metalloproteinase
TLR toll-like receptor

xxiii
UC uterocalin
UF uteroferrin
UG uteroglobin

1
CHAPTER 1
INTRODUCTION

2
1.1 Statement of Purpose
1.1.1 Objective and Rationale
The overall objective of this dissertation research was to characterize the endocrine
disrupting effects of BPA or EE exposure in the uterus of two mouse strains. Two exposure
paradigms were used: 1) exposure in adulthood, during which time mice mated and produced a
litter, and 2) whole life exposure without breeding. Previous studies examined incidence of
pathologies after a much longer exposure time period than is proposed in this dissertation
research. Uterine immune responsiveness and fibrosis are components of several human and
animal uterine pathologies, and the effect of BPA or EE exposure on these endpoints has not
previously been studied.
1.1.2 Central Hypothesis and Specific Aims
The central hypothesis for this dissertation was that exposure to estrogenic EDCs, such as
BPA or EE, will influence E2-regulated uterine physiology, leading to an increased incidence of
uterine pathology. A second hypothesis was that a greater sensitivity in the C57Bl/6N strain to
the estrogenic actions of EDCs also increases the incidence of uterine pathology compared to the
CD1 strain.
The specific aims of this dissertation research included 1) investigating the endocrine
disrupting actions of EDCs in both exposure models on the development of uterine pathologies,
2) examining the effects of EDCs in both exposure models on the immune responsiveness in the
uterus, 3) investigating the ECM changes associated with EDC exposure in both models, and 4)
comparing the pathological, immune, and ECM alterations between the C57Bl/6N and CD1
strains.

3
1.2 Definition of Endocrine Disrupting Chemicals
The first use of the term “endocrine disrupting chemical” (EDC) in a scientific paper was
by Colborn et al. (1993), leading to a surge of research on the deleterious effects of EDCs on
reproduction, development, and other processes and organ systems. According to the National
Institute of Environmental Health Sciences (NIEHS), EDCs were defined as “naturally occurring
compounds or man-made substances that may mimic or interfere with the function of hormones
in the body” (NIEHS 2010). The Endocrine Society defined EDCs as “an exogenous chemical,
or mixture of chemicals, that interferes with any aspect of hormone action” (Gore et al. 2014).
Given such a broad definition and the numerous hormones and hormone targets present in the
body, thousands of natural and synthetic compounds may have endocrine-disrupting properties.
Of particular interest to this dissertation research were compounds that lead to estrogenic
endocrine disrupting activities, such as diethylstilbestrol (DES), 17α-ethinyl estradiol (EE), or
bisphenol A (BPA) (Figure 1).
Figure 1. Chemical structure, name, and CAS ID of estrogenic compounds.

4
1.3 Prototypical Estrogenic EDC: Diethylstilbestrol
Diethylstilbestrol (DES) was one of the first compounds characterized as an estrogenic
EDC. Due to its known estrogenic effects, DES is often used as a positive control in EDC
studies. It was developed in 1938 as a synthetic estrogenic compound and prescribed to pregnant
women in the 1940s to early 1970s to prevent miscarriage and other pregnancy complications
(CDC 2012, NCI 1976). In the 1950s, DES treatment was found to be ineffective in reducing the
incidence of miscarriage and potentially led to premature labor (Dieckmann et al. 1953). In 1971,
it was found that using DES during pregnancy led to an increased incidence of a rare type of
cancer called vaginal clear cell adenocarcinoma in exposed female offspring as young as 15
years of age (Herbst et al. 1971). These findings led the US Food and Drug Administration
(FDA) to issue a warning to physicians to stop prescribing DES (FDA 1972).
Adverse pathophysiological effects are still being diagnosed in women exposed to DES
during pregnancy (“DES mothers”), their male and female offspring (“DES sons and
daughters”), and even in the third generation (“DES grandsons and granddaughters”). DES
mothers had an increased risk of developing breast cancer, but not other hormonal cancers such
as ovarian or endometrial cancer (Titus-Ernstoff et al. 2001, Colton et al. 1993). Animal studies
reported increased mammary tumor incidences (Greenman et al. 1987, Greenman et al. 1986), as
well as fibrosis and degeneration in ovaries (Hong et al. 2010) and thyroid neoplasias (Greenman
et al. 1990). However, these effects appeared to be strain dependent (Eastin et al. 2001).
It was already noted that DES daughters developed vaginal clear cell carcinoma, an
extremely rare cancer especially in young women. Other abnormalities were found in these
women as well, including cervical dysplasia (Robboy et al. 1984) and numerous forms of
gynecological cancers (Verloop et al. 2000). In animal studies, prenatal or neonatal DES

5
exposure caused numerous morphological and pathological alterations either in development or
adulthood, including hyperplasia, adenocarcinoma, pyometra, distended or cystic glands, and
decreased epithelial cell proliferation (Bosquiazzo et al. 2013, Yoshida et al. 2011, Yamamoto et
al. 2003, Yoshida et al. 1999, McLachlan et al. 1980), partially due to DES binding to ERα
(Couse and Korach 2004). Alterations in gene expression related to carcinogenesis, leiomyoma,
and energy metabolism in epithelial cells were reported in DES-exposed animals (Yin et al.
2012, Greathouse et al. 2008, Newbold et al. 2007a), as well as alterations in methylation
patterns (Bromer et al. 2009, Sato et al. 2009), suggesting that DES exposure also has epigenetic
effects on the offspring of DES-exposed animals and humans. In DES sons, there is much debate
as to whether in utero DES exposure increased the risk for testicular or prostate cancer due to the
inconclusive results from epidemiological studies conducted (IARC 2012). In utero DES
exposure resulted in testicular atrophy, decreased spermatogenesis, and alterations in gene
expression in animal studies (Warita et al. 2012, LaRocca et al. 2011, Li et al. 2011, Hong et al.
2010).
Children of mothers who were exposed in utero to DES are now being studied for
potential detrimental effects in the third generation. Increases in reported birth defects in the
third generation were mainly associated with skeletal/bone, heart, or male reproductive system
(Titus-Ernstoff et al. 2010). In third generation daughters, there were irregular menstrual cycles
and the potential for fertility issues (Titus-Ernstoff et al. 2006). In rodent studies, third
generation rodents had increased incidence of uterine adenocarcinomas and ovarian
cystadenocarcincomas (Newbold et al. 1998, Turusov et al. 1992, Walker 1984). Even though
direct DES exposure has not occurred in over 40 years, endocrine disrupting effects are still

6
being found in the offspring of prenatally exposed humans, supported by multigenerational
effects observed in rodent studies.
1.4 17α-Ethinyl Estradiol
17α-ethinyl estradiol (EE) is an orally bioavailable synthetic estrogen pharmaceutical
commonly used in oral contraceptive formulations. Due to its structural similarities to E2, EE
also binds to and activates the ERs (Blair et al. 2000). The major form of metabolism of EE is
oxidation by cytochrome P450 3A4 (Guengerich 1990, 1988). However, EE can also interact
with and inactivate P450 2B1 and 2B6 (Kent et al. 2006, Kent et al. 2002), preventing the
enzymes from metabolizing their target compounds and potentially leading to adverse drug-drug
interactions. Excretion occurs via the urine or feces and leads to contamination of waterways and
subsequent alterations in aquatic wildlife species (Bhandari et al. 2015).
Due to its estrogenic activity, EE is used as a positive control for estrogen-like effects
when studying oral exposures to other estrogenic EDCs. Exposure to high concentrations of EE
had detrimental effects on several parameters associated with reproduction and uterine structure
and function. In a multigenerational study, EE exposure resulted in an earlier age for vaginal
opening and increased the percentage of rats with abnormal cyclicity and cycle length (Delclos et
al. 2009). Pathology such as hyperplasia and alterations in uterine gland numbers was also
observed (Takahashi et al. 2014, Delclos et al. 2009). Uterine weight was increased by EE under
all tested conditions across species, strain, laboratory, and housing (Kanno et al. 2001). Induction
of E2-responsive gene expression in the uterus was similar between EE exposure and E2
treatment (Hyder et al. 1999); however, EE exposure decreased ERα and ERβ expression in both
the uterus and the female hypothalamus (Rebuli et al. 2014, Takahashi et al. 2014, Kang et al.
2003). Gene expression alterations in response to EE exposure included genes involved in cell

7
proliferation and apoptosis, protein metabolism and transport, regulation of transcription, water
imbibition, Wnt signaling, ECM remodeling, alternative complement activation, and estrogen-
mediated Ca2+ signaling (Fong et al. 2007, Heneweer et al. 2007, Katayama et al. 2006, Naciff et
al. 2003).
1.5 Bisphenol A
1.5.1 Commercial Uses of BPA and Routes of Exposure
Bisphenol A (BPA) is a synthetic compound that leaches from polycarbonate plastics and
epoxy resins of food cans to contaminate the contents of the food or beverage container, in
particular under heated or acidic conditions (Cooper et al. 2011, Carwile et al. 2009, Grumetto et
al. 2008, Le et al. 2008, Krishnan et al. 1993). The main route of human exposure occurs orally
through consumption of contaminated food and beverages (Hoekstra and Simoneau 2013,
Teeguarden et al. 2011, Carwile et al. 2009, Grumetto et al. 2008, Vandenberg et al. 2007a).
Humans have transdermal exposure by handling of thermal receipt paper and intravenous
exposure through tubing in medical equipment (Ehrlich et al. 2014, Calafat et al. 2009). Prenatal
exposure occurs via placental transfer to the amniotic fluid (Vom Saal et al. 2014, Edlow et al.
2012, Nishikawa et al. 2010, Ikezuki et al. 2002, Yamada et al. 2002). Greater than 93% of
humans have detectable urinary levels of BPA and metabolites (Calafat et al. 2008), highlighting
ubiquitous BPA exposure in humans. Of importance for consistency between rodent
experimental studies were controlling the potential exogenous sources of BPA and other
estrogenic compounds. Used polycarbonate caging leached BPA into drinking water
(Howdeshell et al. 2003). Diet, bedding, and drinking water also played a major factor in excess
and uncontrolled BPA exposure in rodent studies (Thigpen et al. 2013). Finally, route of
exposure had a significant impact. Because the main route of exposure in humans is orally,

8
experimental animal studies should also use the oral route of exposure in order to extrapolate
results to humans.
1.5.2 Pharmacokinetics and Toxicologic Exposure Limits of BPA
Depending on route of exposure, species, and dose, BPA concentrations reached a
maximal serum concentration between 1-6 hours and declined by 24 hours (Sieli et al. 2011,
Taylor et al. 2011). The main form of metabolism of BPA was via glucuronidation in the liver in
both humans and rodents (Vandenberg et al. 2007a, Yokota et al. 1999). However, humans
excreted BPA and its metabolites in the urine, while the main form of excretion in rodents was in
the feces (Vandenberg et al. 2007a). An acceptable oral limit of BPA exposure in humans was
set at 0.05 mg/kg body weight/day (IRIS 1988). Based on several multigenerational studies in
rodents, the no observed adverse effect level (NOAEL) for BPA was set at 5 mg/kg/day (Tyl et
al. 2008, Tyl et al. 2002). The NOAEL indicates the highest concentration of a compound that
causes no detectable alterations in the endpoint being studied and is dependent on the
experimental design.
1.5.3 Receptor Binding of BPA
Based on in vitro studies, BPA bound to several nuclear hormone receptor targets, but
had agonist or antagonist roles depending on the receptor. The main receptor targets for BPA
were estrogen receptor (ER) α and ERβ, which BPA activated at nM concentrations (Li et al.
2013, Wetherill et al. 2007, Matthews et al. 2001, Kuiper et al. 1998). The relative binding
affinity (RBA) of BPA to ERα was very low and was calculated to be between 0.008 and
0.065%, compared to 100% for E2 (Yiu et al. 2014, Blair et al. 2000). The RBA indicates the
affinity of a ligand to bind to a specific target when compared to a ligand with a known affinity

9
set to 100%. A low RBA, in the case of BPA, means that there are fewer intermolecular forces
between it and ERα compared to between E2 and ERα and that E2 will bind to ERα before BPA
binds. The IC50 is an indicator of the concentration of ligand needed to displace a known ligand
from the binding pocket of a receptor. Using an ERα competitive binding assay with [3H]-E2, E2
displaced 50% of the radioligand at 8.99 x 10-10 M and BPA had an IC50 of 1.17 x 10-5 M (Blair
et al. 2000), meaning that it took a higher concentration of BPA to displace E2 from ERα.
However, the on-rate of BPA binding to ERα was very fast compared to its RBA. The kon for
BPA was 11 x 103 M-1 s-1, compared to the kon for E2 of 1000 x 103 M-1 s-1 (Yiu et al. 2014),
suggesting that the fast on-rate of binding could produce a transient surge in downstream effects.
This transient effect could be prolonged with the recruitment of a co-activator to keep BPA
bound longer. Through mathematical modeling, it was proposed that a 10 minute surge of BPA
from pM to nM concentration in the blood could increase the fraction of BPA-activated ERα to
high enough levels to induce a physiological response (Yiu et al. 2014, Welshons et al. 2003).
The binding and activation of ERβ by BPA had significant physiological effects in the
brain, heart, and pancreas. Rapid activation of ERβ by BPA in estrogen-sensitive developing
cerebellar granule cells led to E2-like neurotoxicity and cell death (Le and Belcher 2010). In the
heart, BPA promoted sex-specific arrhythmogenesis in female rodents via rapid ERβ-mediated
alterations in Ca2+ release and reuptake (Liang et al. 2014, Gao et al. 2013, Belcher et al. 2012,
Yan et al. 2011). Similarly, BPA exposure significantly altered sex-specific Esr2 expression in
multiple areas of the brain, with potential adverse downstream effects related to brain
development. In the developing amygdala, hypothalamus, and other areas related to sociosexual
behavior, altered expression of Esr2 led to BPA-induced anxiogenic behavior (Cao et al. 2014,

10
Patisaul et al. 2012) and the potential for later-in-life defects in neuroendocrine function related
to reproduction (Rebuli et al. 2014, Cao et al. 2013, Cao et al. 2012).
Other receptor targets for BPA include the androgen receptor (AR) and thyroid hormone
receptor (THR), to which BPA bound and inhibited at µM concentrations in vitro (Teng et al.
2013, Moriyama et al. 2002, Paris et al. 2002, Sohoni and Sumpter 1998). This antagonistic
activity led to significant impairment of stromal and glandular cell function in the prostate and
could be involved in enhancing the invasiveness of prostate cancer (Ramos et al. 2001) or on
vertebrate development (Heimeier et al. 2009). The binding of BPA to AR also reduced sperm
quality and disrupted the hypothalamus-pituitary-gonadal (HPG) axis (Wisniewski et al. 2015,
Ramos et al. 2003).
Another receptor that BPA was shown to bind to in vitro is estrogen-related receptor
(ERR) γ. When examining crystal structures, BPA bound to the ligand binding domain of ERRγ
without changing internal structures of the binding pocket; in fact, BPA appeared to stabilize the
constitutively active conformation of ERRγ (Liu et al. 2014, Matsushima et al. 2007). Through
the use of a saturation binding assay, BPA had a KD of 5.5 nM for ERRγ and only one of the
phenol hydroxyl groups was essential for full binding (Okada et al. 2008). Accumulation of BPA
in the placenta was in part mediated by its binding to ERRγ, leading to increased placental
transfer to the developing fetus and significant prenatal exposure (Takeda et al. 2009). Otolith
development in zebrafish was also impaired by the interaction of BPA and ERRγ, suggesting a
potential role in deafness or hearing loss (Tohme et al. 2014)
Rapid signaling actions of BPA occurred through binding to G protein-couple receptor
(GPR) 30. In an ER-negative cell line, BPA had an IC50 of 650 nM for displacing E2 from the
transfected GPR30 (Thomas and Dong 2006). Treatment with BPA also increased

11
phosphorylation of extracellular signal-regulated kinase (ERK) in several cell types via GPR30
activation (Ge et al. 2014, Watson et al. 2014, Dong et al. 2011). Rapid signaling of BPA
binding to GPR30 affected spermatogonial cell proliferation, as well as testicular seminoma
tumor cell and breast cancer cell proliferation (Sheng et al. 2013, Chevalier et al. 2012, Pupo et
al. 2012, Sheng and Zhu 2011).
1.5.4 Endocrine Disrupting Effects of BPA in the Uterus
Exposure to BPA led to endocrine disrupting effects in the uterus, including both gross
and microscopic alterations in structure and significant impairment of uterine function. The
uterotrophic assay assessing uterine weight increases was often used to determine the estrogen-
like activity of EDCs, based on a protocol outlined by the Organisation for Economic Co-
operation and Development (OECD) (Kanno et al. 2003, Kanno et al. 2001). In rodents, BPA
exposure significantly increased uterine weights (Kendig et al. 2012, Markey et al. 2001,
Papaconstantinou et al. 2000, Ashby and Tinwell 1998), supporting that BPA has estrogen-like
activity in the uterus.
Later in life morphological alterations in the uterus from a prenatal or developmental
BPA exposure included changes in luminal and glandular epithelial and stromal thickness and
changes in the number of apoptotic or proliferative cells (Vigezzi et al. 2015, Mendoza-
Rodriguez et al. 2011, Schonfelder et al. 2004, Steinmetz et al. 1998). Increased incidence of
cystic endometrial hyperplasia, squamous metaplasia, adenomyosis, leiomyoma, and stromal
polyps due to BPA exposure were also observed (Vigezzi et al. 2015, Newbold et al. 2009,
2007b, Markey et al. 2005). While BPA altered ERα and PR expression, it is debated if
expression was increased or decreased (Vigezzi et al. 2015, Mendoza-Rodriguez et al. 2011,
Markey et al. 2005, Schonfelder et al. 2004).

12
Exposure to BPA not only impacted uterine structure, but it also significantly impaired
the function of the uterus. Epidemiological studies suggested an association between increasing
urinary or serum BPA concentrations and decreased rates of implantation success or successful
reproductive outcomes (Shen et al. 2015, Peretz et al. 2014, Ehrlich et al. 2012a, Ehrlich et al.
2012b, Mok-Lin et al. 2010). The number of implantation sites was greatly reduced in BPA-
exposed rodents due to decreased Hoxa10 expression, which led to significant impairment of
fertility (Varayoud et al. 2011). Contractility of the uterus was also decreased, which would
result in a reduced ability to deliver a full-term fetus (An et al. 2013). Expression of Hox and Wnt
genes critical for organ development was significantly decreased in the uteri of fetal rhesus
macaques (Calhoun et al. 2014), leading to impaired embryonic development. Exposure to BPA
also increased expression of c-fos, which stimulates cell proliferation, Pgr or the progesterone
receptor, and proteins involved in Ca2+ binding and the immune response (Hewitt and Korach
2011, Hong et al. 2006, Naciff et al. 2002, Steinmetz et al. 1998). In two- and three-generational
reproductive toxicity studies in rodents, only very high BPA concentrations altered fertility
parameters (Tyl et al. 2008, Tyl et al. 2002), but otherwise very few studies have examined the
effects of BPA exposure during adulthood in sexually mature rodents.
1.5.5 Extracellular Matrix Alterations in Response to BPA
Few studies have examined the effects of BPA exposure on the extracellular matrix
(ECM) in any tissue. In the fetal mammary gland, collagen localization but not density was
altered by BPA exposure (Vandenberg et al. 2007b). Whole life exposure to BPA led to sex-
specific alterations in ECM genes and proteins in the mouse heart (Belcher et al. 2015). In the
human ovarian cancer cell line OVCAR-3, BPA significantly increased MMP2 and MMP9
mRNA expression and activity of the enzymes (Ptak et al. 2014). Granulosa-lutein cells treated

13
with BPA had increased MMP9 secretion (Dominguez et al. 2008). While the impact of BPA
exposure on ECM alterations has been studied in various tissues, it still remains unknown if BPA
exposure alters ECM genes and proteins in the uterus.
1.6 Strain Susceptibility to Effects of Estrogenic Compounds
Genetic variation between rodent strains can impact physiological or pathological
responses to estrogenic compounds (Wall et al. 2014). Differences in sensitivity between strains
may lead to false conclusions and interpretations when studying the potential adverse effects of
estrogenic compounds. Therefore, the selection of strain needs to be carefully considered for all
studies that use potential or known estrogenic compounds. Table 1 summarizes several strain-
dependent endpoints with differences in response to estrogenic compound exposure.

14
Table 1. Summary of Studies Examining Sensitivity of Rodent Strains to Estrogenic Compounds
Endpoint Estrogenic Compound
Summary of Results Citation
Uterine Weight
DES ↑ in Alpk and C57Bl/6J at low and high dose, ↑ in CD1 and B6CBF1 at high dose only Ashby et al. (2003)
EE ↑ in DA/Han, Wistar, and Sprague-Dawley
Diel et al. (2004) Genistein ↑ in DA/Han, Wistar, and Sprague-Dawley at 50 and 100 mg/kg/day
BPA ↑ in Da/Han and Sprague-Dawley at 200 mg/kg/day
Uterine Morphology
Estradiol Benzoate cystic, hyperplastic glands in C57Bl/6 one week after treatment; Silberberg and Silberberg
(1951) develop two months after treatment in strain A/J
Circulating Sex Hormones
larger prolactin and P peaks in strains with small litter size; DeLeon et al. (1990)
smaller P peak in strains with large litter size
Epithelial ERα Expression
expressed in CD1 at PND3; expressed in BALB/c and C57Bl/6J at PND5
Bigsby et al. (1990), Taguchi et al. (1988)
DES advanced ERα ontogeny in C57Bl/6J Taguchi et al. (1988)
QTL
E2 Est2 and Est3control phenotypic variation in uterine weight Wall et al. (2013), Roper et
al. (1999)
E2 Est1 control phenotypic variation in eosinophil infiltration Griffith et al. (1997)
E2/DES Eutr1 (E2) and Eutr2 (DES) control phenotypic variation in pyometra development Gould et al. (2005), Pandey
et al. (2005)
Ovary
number of oocytes in neonatal development higher in CD1 than C57Bl/6;
Pepling et al. (2010) CD1 faster rate of loss of oocytes
E2 ↑ in oocytes in CD1 and C57Bl/6; greater increase in C57Bl/6
BPA ↓ follicular growth and steroidogenesis in CD1 and C57Bl/6 Peretz and Flaws (2013),
Peretz et al. (2013)
Vagina
BPA ↑ cell proliferation in F344 compared to Sprague-Dawley Long et al. (2000)
Estradiol Valerate C57Bl/6 and BALB/c susceptible and CD1 resistant to Candida albicans infection Mosci et al. (2013),
Clemons et al. (2004), Calderon et al. (2003)
Anterior Pituitary BPA hyperprolactinema in F344, no effect in Sprague-Dawley Steinmetz et al. (1997)
Mammary BPA/E2 changes in morphology in CD1 and C57Bl/6N Wadia et al. (2007)
Male Reproductive
E2 ↓ testicular weight and spermatogenesis in C57Bl/6J, CD1 resistant Spearow et al. (2001),
Spearow (1999)

15
1.6.1 Susceptibility to Uterine Weight Increases and Morphological Alterations
The uterotrophic assay is often used to determine the estrogen-like activity of a
compound. However, differences in sensitivity to estrogens between rodent strains may lead to
false results. For example, when comparing uterine weights in four DES-exposed mouse strains,
the Alpk and C57Bl/6J strains demonstrated significant increases at 1 µg/kg/day DES, while the
CD1 and B6CBF1 strains did not. All strains had significant uterine weight increases at 10
µg/kg/day DES though (Ashby et al. 2003). Several other studies using mice or rats observed
varied susceptibility to uterine weight increases in response to EE, BPA, DES, genistein, and
other E2 derivatives (Diel et al. 2004, Greenman et al. 1977, Drasher 1955).
Morphological features of the glands in the endometrium of the uterus differed between
strains in response to estradiol benzoate treatment. The C57Bl/6 strain had cystic and
hyperplastic glands one week after treatment, while it took two months before the same altered
morphology was observed in the A/J strain (Silberberg and Silberberg 1951). These findings
suggest that differences in sensitivity to estrogenic compounds impact alterations in uterine
morphology, eventually developing into pathology.
1.6.2 Differences in Circulating Sex Hormones and ERα Levels
Strain-dependent differences in circulating sex hormone levels may influence variances
in sensitivity to exogenous estrogenic compounds. Mouse strains selected for small or large litter
size had differing circulating sex hormone levels. Smaller litters coincided with larger prolactin
and P peaks, with the prolactin peak occurring in estrus. Larger litter size correlated with a
smaller P peak during proestrus (DeLeon et al. 1990). It was also possible that E2 levels differed
between strains due to alterations in circulating P levels and negative feedback mechanisms.

16
Uterine epithelial ERα expression during development also differed greatly between
strains. The CD1 strain expressed ERα as early as PND3, compared to BALB/c or C57Bl/6J
mice that expressed ERα at PND5 (Bigsby et al. 1990, Taguchi et al. 1988). Estradiol is
necessary for epithelial cell proliferation, and this finding suggests that CD1 mice have a more
advanced glandular and luminal ontogeny compared to inbred strains. However, DES exposure
advanced ERα ontogeny in epithelial cells of C57Bl/6J mice so that the uterus looked similar to
the uterus of the CD1 strain (Taguchi et al. 1988).
1.6.3 Quantitative Trait Loci Controlling Phenotypic Variation
Genetic variation between strains or even substrains influences phenotypic differences at
numerous endpoints. The two most commonly used inbred mouse strains, C57Bl/6J and
C57Bl/6N, have distinct genetic differences that led to phenotypic alterations at endpoints related
to behavior, ophthalmology, immune function, cardiovascular parameters, and metabolism
(Simon et al. 2013, Mekada et al. 2009). Specific quantitative trait loci (QTL) regulated
susceptibility to alterations in response to estrogenic compound exposure for uterine wet weight,
cellular infiltration, and the development of pyometra. Phenotypic variation in uterine wet weight
was controlled by two interacting QTL in the mouse, Est2 on chromosome 5 and Est3 on
chromosome 11. A third QTL, Est4 on chromosome 10, encoded an interacting factor that
influenced the variation of Est2 and Est3. These QTL contained several E2-responsive genes,
leading to the possibility that estrogenic EDCs may also alter uterine weight via interactions with
these genes (Wall et al. 2013, Roper et al. 1999). Eosinophil infiltration in response to E2
treatment also appeared to be genetically controlled by Est1 on chromosome 4, which interacted
with loci on chromosomes 10 and 16 (Griffith et al. 1997). Two separate but overlapping QTL
on chromosome 5 were responsible for controlling the development of pyometra in rats, Eutr1

17
for E2-induced pyometra and Eutr2 for DES-induced pyometra (Gould et al. 2005, Pandey et al.
2005). These studies highlighted that sensitivity to estrogenic compounds was in part regulated
by underlying genetic differences between strains due to specific chromosomal locations being
identified and involved in genetic control of estrogen-responsive endpoints.
1.6.4 Differential Sensitivity of the Ovary and Vagina to Estrogenic Compounds
Other organs in the female reproductive system, including the ovaries and vagina, also
showed physiological differences or alterations in susceptibility to estrogenic compounds
between rodent strains. During normal neonatal ovarian development, the number of oocytes in
the ovaries of CD1 mice was significantly higher compared to the C57Bl/6 strain. However, the
rate of loss of oocytes in the ovaries of CD1 mice was faster compared to the C57Bl/6 strain. The
same number of oocytes were present at PND4 in the CD1 and C57Bl/6 strains (Pepling et al.
2010). When comparing sensitivity to E2, both the CD1 and C57Bl/6 strain had a significantly
increased number of oocytes compared to non-treated conditions. However, the increase was
much greater in C57Bl/6 mice, suggesting that this strain is more sensitive to estrogenic
compounds (Pepling et al. 2010). Inhibited follicle growth and steroidogenesis in response to
BPA exposure in the adult ovary occurred in both CD1 and C57Bl/6 strains, suggesting that the
response of the ovary to BPA at this endpoint was not dependent on genetic variation between
mouse strains (Peretz et al. 2013, Peretz and Flaws 2013).
Another estrogen-responsive tissue, the vagina, showed differences in cell proliferation
but not BPA clearance, ERα binding, or c-fos mRNA expression levels between Fischer 344 and
Sprague-Dawley rats (Long et al. 2000). Infection with Candida albicans also differed between
C57Bl/6, BALB/c, and CD1 and derived CD10 and CD3 strains, following similar patterns of

18
susceptibility as to estrogen sensitivity (Mosci et al. 2013, Clemons et al. 2004, Calderon et al.
2003).
1.6.5 Differential Sensitivity of Other Tissues to Estrogenic Compounds
Tissues other than those in the female reproductive tract also exhibited differential
sensitivity to estrogens based on rodent strain. In the anterior pituitary, BPA exposure resulted in
hyperprolactinemia in Fischer 344 rats but did not affect the Sprague-Dawley strain (Steinmetz
et al. 1997). When examining the effects of BPA exposure on mammary gland morphology, the
CD1 strain had slightly more pronounced alterations compared to C57Bl/6N mice; however,
both strains were sensitive to the actions of E2 on mammary gland morphology (Wadia et al.
2007). The male reproductive system also displayed phenotypic variations to E2 treatment in
different mouse strains. The CD1 strain was resistant to E2 effects on inhibition of testicular
weight and spermatogenesis at doses 16 fold higher than those that led to the abolishment of
spermatogenesis in the C57Bl/6J strain (Spearow et al. 2001, Spearow 1999), demonstrating that
estrogen sensitivity is not specific to females. Genetic background also played an important role
in estrogen sensitivity in the heart (Kararigas et al. 2014), the adrenal (Westberg et al. 1957),
macrophage function, (Passwell et al. 1974), and anxiety-related behavior and associated
inflammatory responses (Benatti et al. 2011).
Important knowledge gaps still remained concerning differences in sensitive populations
and BPA exposure. In regards to uterine structure, function, and potential for the development of
pathologies, it was unknown if genetic variation at least in part controlled phenotypic
differences. It was also unclear if baseline physiological differences within the uterus between
strains had any effect on sensitivity to the endocrine disrupting actions of BPA.

19
1.7 The Endocrine System
The endocrine system is comprised of a series of interacting organs and secreted
hormones to regulate numerous biological processes and maintain an overall homeostasis in the
organism. Endocrine glands throughout the body produce and secrete hormones directly into the
bloodstream. These hormones then act on target cells, usually through binding to specific
receptors (Molina 2013). Hormones released by organs in the endocrine system affect almost any
tissue in the body; of particular interest to these studies is the uterus and response to estrogenic
compounds.
Often, hormone secretion is regulated by a feedback loop system to either increase
(positive feedback) or decrease (negative feedback) further hormone production and secretion.
An example of one such feedback loop is the hypothalamic-pituitary-gonadal (HPG) axis (Figure
2). In its simplest form, the hypothalamus produces and secretes gonadotropin-releasing
hormone (GnRH), stimulating the anterior pituitary to secrete luteinizing hormone (LH) and
follicle stimulating hormone (FSH). These hormones then stimulate the ovaries to produce
progesterone (P4) and 17β-estradiol (E2) in females or the testes to produce testosterone (T) in
males. These hormones then have regulatory roles in these reproductive organs, but they are also
secreted into circulation to play stimulatory roles in other target tissues, such as the uterus,
mammary, or prostate. Sex hormones also feed back to the hypothalamus and anterior pituitary
to work in an inhibitory fashion. High levels of E2 leads to decreased FSH secretion and
increased LH secretion, leading to the LH surge associated with ovulation. The presence of high
levels of P4 then decreases LH secretion (Molina 2013).

20
Figure 2. Simplified representation of HPG axis and negative feedback loop. As described, GnRH released by the hypothalamus stimulates the anterior pituitary to release LH and FSH. These hormones stimulate the production of E2 and P4 in the ovaries and T is the testis, which then act on target tissues. However, the sex hormones can also inhibit further release of LH and FSH from the anterior pituitary.
1.8 Nuclear Hormone Receptors: ERα and ERβ
1.8.1 Structure
Estrogens and estrogenic compounds bind to specific nuclear hormone receptors, the
estrogen receptors ERα and ERβ, and act as a ligand-activated transcription factor to modulate
responsive gene expression through recruitment of stimulatory or inhibitory complexes.
Estrogens binding to ERs can also have rapid signaling effects. The most potent endogenous
estrogen is E2. Experimental evidence in support of the idea of a receptor specific for E2 was
first reported by Jensen and Jacobson (1962), but the cDNA and amino acid sequences of the
human ER were not reported until over 20 years later (Green et al. 1986, Greene et al. 1986). In

21
1996, a second novel ER was cloned and named ERβ or ESR2 to distinguish it from the previous
ER, now named ERα or ESR1 (Kuiper et al. 1996, Mosselman et al. 1996). Using BioGPS
software, the highest mRNA expression of human ESR1 occurs in the uterus, pituitary, and
prostate. Highest mRNA expression of human ESR2 occurs in cardiomyocytes (Figure 3) (Wu et
al. 2013, Wu et al. 2009). The chromosomal location of ESR1 in humans is 6q25.1,
corresponding to a chromosomal location for Esr1 of 10 2.03 cM in mice. ESR2 in humans is
located on 14q22-24, corresponding to the location of Esr2 in mice on 12 33.52 cM.
Figure 3. Expression of ERα and β in human tissues. Shown are graphical representations of mRNA expression levels in various human tissues for ERα and ERβ. The graph for ERα is also shown zoomed in to show lower expression levels in numerous tissues. Graphs were generated at biogps.org using Dataset GeneAtlas U133A, gcrma for both receptors and Probeset 205225_at for ERα and Probeset 211120_x_at for ERβ.
The ERα and ERβ proteins are members of the nuclear hormone receptor (NHR)
superfamily. All NHRs are made up of three interacting domains, the A/B or N-terminal domain
(NTD) containing a ligand-independent activation factor (AF1), the C or DNA-binding domain
(DBD), and the D/E/F or ligand-binding domain (LBD) containing a ligand-dependent activation

22
factor (AF2) important for coregulator recruitment and receptor dimerization (Figure 4). The
DBD contains two zinc finger domains that facilitate dimerization and DNA binding of the
receptor to specific palindromic estrogen response elements (EREs) in a DNA sequence. The D
region in the LBD is a hinge region which contains a nuclear translocation signal. The LBD
contains a hydrophobic ligand binding pocket, important for ligand specificity (Planey et al.
2014, Ellmann et al. 2009).
Figure 4. Schematic of domain structure of ERα and β. Each receptor contains an A/B or N-terminal domain (NTD), a C or DNA-binding domain (DBD), and a D/E/F or ligand-binding domain (LBD). Sequence homologies between the human receptors for each domain are noted. 1.8.2 Nuclear Translocation, Membrane-Initiated, and Rapid Signaling Mechanisms
The mechanism of ER-mediated nuclear translocation begins with free E2 diffusing
through the plasma membrane and binding to an ER/heat shock protein (HSP) complex, made up
of HSP56 and 90, in the cytosol. Once E2 binds to the ER, the HSP complex dissociates and two
E2/ER molecules dimerize. The dimerized product then enters the nucleus, binds to EREs in the
DNA sequence, and recruits primary transcription factor coregulators which can then recruit
secondary coregulators to either promote or repress transcription of estrogen-responsive genes
(Figure 5) (Belcher 2008).

23
Figure 5. Classical model of nuclear hormone receptor translocation. Shown is a cartoon describing the “classical” nuclear receptor mechanism of ligand-bound estrogen receptor (ER)-mediated effects on gene transcription. Ligand binding induces a conformational change of the receptor leading to dissociation from a chaperone complex containing heat shock proteins. Activated receptors dimerize and bind at estrogen response elements (EREs) in the promoters of E2-responsive genes. Ligand/receptor complexes bound to EREs results in the recruitment of coregulatory transcription factors to the promoter which modify target gene transcription (Belcher 2008).
Rapid signaling (non-genomic) mechanisms of ERα and ERβ activation can also occur
(Banerjee et al. 2014, Levin 2009, Belcher 2008). Rapid signaling through the ERs can either be
membrane-initiated or occur intracellularly. Activation of ERα increased phosphoinositide-3-
kinase (PI3K), extracellular signal-regulated kinases (ERK) 1/2 mitogen-activated protein
kinases (MAPK), and protein kinase B (PKB)/Akt through a G protein-coupled mechanism
(Banerjee et al. 2014, Kumar et al. 2007, Revankar et al. 2005, Razandi et al. 2003b). In the
cerebellum and cerebellar granule cell neurons, E2-induced neurotoxicity was mediated by rapid

24
signaling actions of ERβ, involving a G protein- and protein kinase A (PKA)-dependent
mechanism. Treatment with E2 led to rapid activation of ERK1/2 MAPK and stimulated cell
death (Le and Belcher 2010, Belcher et al. 2005, Wong et al. 2003). In the female heart,
estrogens rapidly altered myocyte Ca2+ handling, leading to the promotion of arrythmogenesis
(Yan et al. 2011). These rapid signaling actions were mediated by an imbalance between the
stimulatory effects of ERβ and the suppressing actions of ERα (Belcher et al. 2012). The
protective effects of E2 against bone resorption may be due to rapid signaling actions as well,
based on results from studies using an E2 dendrimer conjugate incapable of inducing the nuclear
actions of ERα (Bartell et al. 2013). Another physiological action of ERα-mediated rapid
signaling is the increased production of nitric oxide (NO) within minutes of E2 treatment due to
activation of endothelial nitric oxide synthase (eNOS), which plays a crucial role in maintaining
vascular homeostatis and function (Wu et al. 2011, Wyckoff et al. 2001, Chen et al. 1999,
Lantin-Hermoso et al. 1997).
Because ERs are located intracellularly in the cytoplasm, recruitment to the membrane is
often necessary for these rapid signaling actions to occur. One such process is palmitoylation, or
the covalent attachment of fatty acids to cysteine residues (Meitzen et al. 2013). A Cys447Ala
mutation in ERα impaired localization to the plasma membrane, indicating Cys447 as the crucial
site for palmitoylation (Acconcia et al. 2005, Acconcia et al. 2004). A second palmitoylation site,
Cys451, was also implicated in the rapid signaling mechanisms of ERα (Adlanmerini et al.
2014). Recruitment of ERα to the plasma membrane also likely occurred through its interaction
with c-Src or caveolin-1 (Banerjee et al. 2014, Haynes et al. 2003, Razandi et al. 2003a).
A third membrane-associated G protein-coupled receptor that binds E2 and is expressed
in several tissue types is G protein-coupled receptor 30 (GPR30) or G protein-coupled estrogen

25
receptor 1 (GPER1) (Carmeci et al. 1997, Kvingedal and Smeland 1997, Takada et al. 1997,
Owman et al. 1996). Activation of GPR30 by E2 induced rapid signaling pathways, usually
within minutes. GPR30 and ERα both activated ERK1/2 via epidermal growth factor receptor
(EGFR) transactivation (Razandi et al. 2003b, Filardo et al. 2000). While both receptors also
activate PI3K (Revankar et al. 2005), most cell types utilized GPR30 in vivo to activate PI3K
and MAPK (Prossnitz and Maggiolini 2009, Revankar et al. 2005). Another rapid signaling
mechanism was intracellular Ca2+ mobilization via activation of GPR30 (Revankar et al. 2005).
While GPR30 is not a nuclear hormone receptor, GPR30-mediated events still indirectly led to
transcriptional activation as well (Prossnitz and Maggiolini 2009). For example, several cell
types had increased expression of c-fos and Bcl-2 through E2-mediated GPR30 activation,
leading to increased proliferation (Prossnitz and Barton 2009, Prossnitz and Maggiolini 2009). It
was suggested that GPR30 signaling played a role in tumor proliferation and invasiveness in
breast, endometrial, and ovarian cancers and indicated poor survival (Smith et al. 2009, Smith et
al. 2007, Filardo et al. 2006, Filardo et al. 2002, Filardo et al. 2000).
The estrogen-related receptors (ERRs) are members of the NHR superfamily discovered
through a cDNA screening and have a similar sequence in the DBD as the human ERα (Giguere
et al. 1988). There are currently three types of human ERRs, named ERRα (ESRRA), ERRβ
(ESRRB), and ERRγ (ESRRG). These receptors are considered orphan receptors because the
endogenous ligand is unknown. They have a low sequence homology in the LBD with ERα;
therefore, they do not bind or respond to E2 (Huss et al. 2015). However, ERRs are
constitutively active based on dimerization partners and also bind exogenous estrogenic
compounds such as BPA or DES (Liu et al. 2014, Matsushima et al. 2007, Huppunen and
Aarnisalo 2004). Similar to ERs, ERRs bind to specific DNA sequences named estrogen-related

26
receptor response elements (ERREs) as monomers, homodimers, or heterodimers (Huppunen and
Aarnisalo 2004, Gearhart et al. 2003). ERRs do not bind strongly to EREs; however, ERRs and
ERα do share some target genes and ERRs can modulate estrogen signaling by cooperation for
full gene activation or by antagonism (Zhang et al. 2006, Kraus et al. 2002, Vanacker et al. 1999,
Yang et al. 1996). Crystal structures showed that ERRs complex with a coactivator peptide from
peroxisome proliferator-activated receptor coactivator-1α (PGC-1α) and had a transcriptionally
active conformation in the absence of ligand (Kallen et al. 2004), suggesting that ligand binding
is not necessary for ERRs to bind to DNA and regulate transcription. Signaling through ERRs
was important in regulation of genes involved in oxidative phosphorylation, mitochondrial
biogenesis, fatty acid transport, and bone and cartilage formation. They were also involved in
regulating the stress response and the development of breast and prostate cancers (Misawa and
Inoue 2015, Bonnelye and Aubin 2013, Byerly et al. 2013, Huss et al. 2004, Schreiber et al.
2004).
1.8.3 Role of ERs in Uterine Structure and Reproductive Function
Estrogen receptors play a critical role in regulating numerous processes in the uterus.
However, of most importance to uterine structure and function is ERα. Studies using global ERα
knockout or uterine epithelial cell-specific ERα knockout models showed that ERα is required
for fertility because implantation and decidualization cannot occur without activation of the
receptor (Pawar et al. 2015, Hewitt and Korach 2003, Hewitt et al. 2002).
1.8.3.1 Postnatal Uterine Development
Expression and localization of ERα changed in the developing uterus after birth. Initially,
ERα was located in the stroma at low expression levels which progressively increased during
development. In rats, a gradual change in localization occurred during development, with ERα

27
initially expressed in the luminal epithelium at postnatal day (PND) 7 but then most abundant in
the glandular epithelium at PND21 (Fishman et al. 1996). These changes in localization and
increases in expression occurred until puberty and the beginning of estrous cyclicity, at which
time their localization and expression was similar to an adult rodent.
Adenogenesis, or uterine gland development, is another process that required ERα
(Lubahn et al. 1993). Glands were not present in the uterus at birth in laboratory rodents or
humans. Starting at PND5 in rodents, epithelial invaginations in the uterus were observed that
formed glandular epithelium (Cooke et al. 2012a, Cooke et al. 2012b). Glands were not present
in the uterus until PND7 in mice and PND9 in rats (Cooke et al. 2013, Branham et al. 1985). On
PND15 in mice, a general organization of the adult uterus was established (Hu et al. 2004). In
humans, glandular epithelium differentiated from luminal epithelium to form tubular glands
throughout the stroma (Cooke et al. 2013).
1.8.3.2 Impact on Gene Expression
Expression of several genes in the uterus is dependent on ERα signaling. In the absence
of ligand, ERα still bound to chromatin structures, but ERα binding was greatly increased by the
presence of E2. Often, RNA polymerase II was located in similar binding sites, suggesting co-
localization in preparation for transcriptional activation (Hewitt et al. 2012). Over 1800 genes
were found to be regulated by E2 in the uterus (Leitman et al. 2012). In the cycling uterus,
clusters of genes involved in extracellular matrix (ECM), cell adhesion, proliferation, immune
response, coagulation, metabolism, and Wnt and hedgehog signaling were differentially
expressed in stages of the reproductive cycle, suggesting that their expression is dependent in
part on E2 activation of ERs (Yip et al. 2013).

28
1.8.3.3 ERα in the Brain and Impact on Cyclicity
In the brain, ERα also influences uterine function through responses to circulating E2
levels and negative feedback mechanisms. ERα-expressing neurons in the arcuate nucleus were
involved in this negative feedback and normal cyclicity (Yeo and Herbison 2014). Inhibition of
GnRH release from GnRH neurons occurred through E2 binding to ERs (Radovick et al. 2012,
Sanchez-Criado et al. 2006). Using in situ hybridization, expression of kisspeptin (Kiss1) and
ERα in the hypothalamus was highly overlapping. Kiss1 is a protein that can directly regulate
GnRH release from GnRH neurons due to binding to and activating GPR54 located on GnRH
neurons (Navarro et al. 2004). Regulation of GnRH neurons by Kiss1 significantly impacted
estrous cyclicity and puberty by regulating LH and FSH expression (Witham et al. 2013, Navarro
et al. 2004). Other studies supported that regulation of GnRH release was mediated by E2
binding to ERα in Kiss1 neurons (Radovick et al. 2012, Cao and Patisaul 2011, Vida et al. 2010).
Because ERα expression and signaling in the brain was highly involved in regulating estrous
cyclicity, alterations in this signaling significantly impaired uterine function. Administration of
exogenous estrogenic compounds during neonatal development significantly decreased Kiss1
neuron density and signaling in the hypothalamus, leading to impaired reproductive function
(Patisaul et al. 2009, Bateman and Patisaul 2008).
1.8.3.4 Role of GPR30 in the Uterus
GPR30 is expressed in both the endometrium and myometrium of the uterus, with
expression levels varying during the reproductive cycle (Maiti et al. 2011, Kolkova et al. 2010,
Otto et al. 2009). However, GPR30 had little effect on uterine structure and fertility based on
knockout studies (Otto et al. 2009). In the endometrium, GPR30 activation negatively regulated
rapid signaling of ERα by inhibiting ERK1/2 phosphorylation and reducing uterine growth,

29
suggesting that these signaling events are relevant for balancing ERα-mediated effects (Gao et al.
2011). Within the myometrium, GPR30 activation increased smooth muscle contraction in
response to oxytocin, implying an important role in labor (Maiti et al. 2011).
Progression of several uterine diseases is partially mediated by GPR30. In leiomyoma
samples, GPR30 expression was significantly increased compared to normal myometrium (Tian
et al. 2013). Proliferation and invasion of endometrial carcinoma cells occurred via activation of
GPR30 by E2 or hydroxytamoxifen and downstream signaling of the ERK/MAPK pathway (Du
et al. 2012, He et al. 2009, Vivacqua et al. 2006).
1.9 The Cycling and Pregnant Uterus
1.9.1 Circulating Hormones and Tissue Remodeling
The main function of the uterus is to protect and provide nourishment for a growing fetus.
The myometrium and endometrium are two distinct morphological and functional tissue layers.
The myometrium is divided into two layers composed of smooth muscle and vasculature and
surrounds the endometrium to provide structural support and protection. It can greatly expand
during pregnancy and contracts during labor (Thompson 2015). The endometrium consists of
lumen and gland structures lined by epithelial cells, which are connected by the lamina propria or
stroma and ECM (Figure 6). These structures are crucial for preparing the uterus for implantation
and fetal development and differentiate based on circulating E2 and P levels, also known as the
endometrial or menstrual cycle in humans and the estrous cycle in rodents (Buy and Ghossain
2013, Molina 2013).

30
Figure 6. Normal uterine structure in adult CD1 mouse. Uterus is divided into myometrium (Myo) and endometrium (Endo). Myo consists of inner myometrium (IM) and the outer myometrium (OM). Endo is made up of luminal and glandular structures and connected by stroma. Similar uterine structures are found in normal human tissue.
In humans, the entire reproductive cycle lasts on average 28 days, beginning with the
endometrium shedding in a process called menstruation. As described in Molina (2013),
menstruation occurs in the menstrual phase of the uterus if fertilization and implantation does not
occur. The menstrual phase is considered day 1 of the cycle. Menstruation also indicates day 1 of
the follicular phase, or maturation of the developing egg, in the ovary. During the beginning of
the follicular phase, sex hormone levels are low. Starting at day 3 of the follicular phase, E2
levels begin to gradually increase, peaking at day 13. The proliferative phase in the uterus occurs
between days 4 to 13 and is characterized by thickening of the endometrium in preparation for
implantation. Around day 14, ovulation occurs due to a rapid increase in LH and FSH and a peak
in E2 levels, signifying the end of the follicular phase. Following ovulation, the ovary enters the
luteal phase, in which the follicle that released the egg gradually matures to form a corpus
luteum. The luteal phase lasts from day 14 to day 26. A mature corpus luteum secretes P4,

31
leading to the increased P4 levels observed after ovulation. Levels of LH, FSH, and E2 rapidly
decrease; however, a second, smaller peak in E2 is also observed during high P4 levels around
day 20. In the uterus, the secretory phase occurs from day 13 to day 24 and is characterized by an
inhibition of endometrial thickening by high P4 levels (Molina 2013, Hinson et al. 2010).
Morphological changes prepare the uterus for implantation, a process called decidualization
(Gardner and Shoback 2007). If implantation has not occurred at day 28, the uterus reenters the
menstrual phase and the thickened endometrium is shed (Molina 2013, Hinson et al. 2010).
Graphical representation of the human menstrual cycle is found in Figure 7.
Figure 7. Simplified representation of the human menstrual cycle. The menstrual cycle has an ovarian and endometrial component. The ovarian cycle is divided into the follicular and luteal phases. The endometrial cycle is divided into the menstrual (Mens), proliferative and secretory phases. The relative sex hormone levels are also graphically shown from Molina (2013).
A similar process occurs in rodents, but on a shorter time scale. While the menstrual
cycle in humans is roughly 28 days, the estrous cycle is only 4 days in rodents (Cora et al. 2015,
Goldman et al. 2007). The proestrus and estrus stages are similar to the proliferative phase in
humans in that the endometrium is thickening in preparation for implantation. Ovulation occurs

32
at the end of the estrus stage. However, peaks in E2, LH, and FSH do not occur at the same time.
Instead, all three hormones peak between the proestrus and estrus stages, with FSH having a
second, smaller peak at the end of the estrus stage (McLean et al. 2012). Following ovulation, the
rodent enters metestrus if fertilization has not occurred. This stage is characterized by
endometrial degeneration and is similar to the luteal phase in humans (Hedrich and Bullock
2004). However, instead of menstruation like in humans, rodent tissue degenerates and
eventually regenerates. A second small peak in E2 occurs in metestrus, as well as gradual
increases in P4 that peaks in diestrus. The diestrus stage is characterized by a quiescent state in
the endometrium (McLean et al. 2012).
1.9.2 Role of Extracellular Matrix in Uterine Structure and Function
The extracellular matrix (ECM) is a group of interacting proteins and polysaccharides
that are secreted by surrounding cells to provide structural support and initiate biochemical and
biomechanical processes (Frantz et al. 2010). The ECM plays a dynamic and tightly regulated
role in maintaining structure of the cycling uterus. Regulation of collagen synthesis and
degradation is summarized in Figure 8. In the cycling uterus, expression of ECM proteins
involves several levels of regulation, almost all of which are differentially expressed in response
to circulating levels of E2 or P, highlighting that sex hormones are important for numerous
aspects of tissue remodeling during the menstrual or estrous cycles.
Accumulation of collagens involves a balance between hormonal control of collagen
gene expression and matrix metalloproteinase (MMP) expression and activity (Gaide
Chevronnay et al. 2012, Henriet et al. 2012). The collagen types I and III are primary collagens
expressed in mammalian uterus, with collagen types IV, V, and VI expressed variably during the
cycle, mainly in the stroma, basement membranes of glandular epithelium, and myometrium

33
(Augsburger and Henzi 2008, Boos 2000, Tanaka et al. 1998, Stenback 1989, Aplin et al. 1988,
Karkavelas et al. 1988). In response to E2, synthesis of mRNA and protein for collagen subtypes
increased (Frankel et al. 1988, Dyer et al. 1980, Salvador and Tsai 1973).
Figure 8. Regulation of collagen synthesis and degradation. Shown is a simplified schematic of the major levels of control of transcriptional and posttranslational regulation of collagens and MMPs. In stromal and epithelial cells in the uterus, E2 binds to ERα, dimerizes, and translocates to the nucleus. Binding to EREs and recruitment of co-activators leads to transcription of collagen type 1 (pink) and type 3 (green) and proMMP2 (blue) and proMMP14 (purple). Once collagen mRNA is translated, it is released into the extracellular compartment. ProMMP14 is activated and localized to the plasma membrane, where it forms a complex with TIMP2 (orange). This complex interacts with proMMP2 to activate the enzyme via a cysteine-switch mechanism, which is then released into the extracellular compartment. MMP2 can be inhibited by TIMPs and α-macroglobulin.
Matrix metalloproteinases are a group of Ca2+-dependent collagen degrading enzymes
whose expression and activity are also tightly regulated by E2 levels during the menstrual or
estrous cycle (Gaide Chevronnay et al. 2012, Russo et al. 2009, Helvering et al. 2005, Rodgers et
al. 1994). They are divided into different subgroups based on their structure (Figure 9). For

34
example, collagenases (MMP1, 8, and 13) differ from gelatinases (MMP2, 9) due to the lack of
fibronectin type II domains located within the catalytic domain. Substrates for each MMP also
differ. However, most collagens can be degraded by several different MMPs. For example,
collagen type I is a substrate for and degraded by MMP1, 2, 7, 8, 13, and 14 (Curry and Osteen
2003). The MMP9 and MMP2 proteins are key gelatinases present in both latent and active
forms in endometrial tissue of the uterus throughout the reproductive cycle (Gaide Chevronnay
et al. 2012). Other MMPs in the uterus include MMP1, 3, 7, 10, 11 and various membrane-
associated MMPs, which also have differential expression during the estrous or menstrual cycle
(Goffin et al. 2003, Goldman and Shalev 2003, Curry and Osteen 2001, Hulboy et al. 1997).
Initially, MMPs are transcribed and translated as quiescent proMMPs and activated in the
extracellular space by proteolytic cleavage. Regulation of MMP2 activity during the reproductive
cycle is in part controlled by MMP14, which activates proMMP2 by proteolytic cleavage via a
“cysteine switch” mechanism of action. A cysteine residue in the propeptide domain interacts
with Zn2+ in the catalytic domain, preventing cleavage of the propeptide domain and activation
of the MMP. Disruption of this interaction leads to opening of the cleavage site in the propeptide
domain and the active site in the catalytic domain and activation of the MMP (Gaide Chevronnay
et al. 2012, Murphy et al. 1999, Strongin et al. 1995, Sato et al. 1994, Springman et al. 1990, Van
Wart and Birkedal-Hansen 1990).

35
Figure 9. Schematic representation of MMP structures. The common name and associated number for MMPs are shown. The MMPs contain several domains, including a signal peptide, propeptide, catalytic domain containing a Zn2+ binding site, hinge region, and hemopexin-like domain. The MMP is activated by cleavage of the signal peptide and propeptide. The gelatinases contain several fibronectin type II domains within the catalytic domain. A furin site that allows for intracellular activation can be found in MMP11 and the transmembrane MMPs. The transmembrane MMPs also contain a transmembrane linker and a cytoplasmic domain. The activity of MMPs is also tightly controlled and is inhibited by P4 or specific
inhibitory proteins. Attenuation of MMP14 expression by P4 led to decreased MMP2 activation
(Zhang et al. 2000). Tissue inhibitors of metalloproteinases (TIMPs) and α-macroglobulin (α-
MG) also inhibit MMPs in active form (Dong et al. 2002). Based on crystal structures, TIMPs
have an overall “wedge-like” shape that fits into the active site of MMPs, inhibiting ECM
components from being degraded (Nagase et al. 2006, Fernandez-Catalan et al. 1998, Gomis-

36
Ruth et al. 1997). Inhibition by α-MG occurred by entrapping the MMP between the four
subunits of one α-MG molecule. The MMP/α-MG complex was then cleared by endocytosis
mediated by low density lipoprotein receptor-related protein (LRP) (Nagase et al. 2006,
Strickland et al. 1990). TIMPs-1, -2, and -3 were differentially expressed during the estrous or
menstrual cycle in response to cycling levels of sex hormones, increasing and decreasing at
similar time points to MMPs (Gaide Chevronnay et al. 2012, Goffin et al. 2003, Zhang and
Salamonsen 1997, Hampton and Salamonsen 1994, Sato et al. 1991). The fluctuating expression
levels prevented excess degradation of collagens and other ECM components by high levels of
MMPs. However, TIMP-2 also aided in activation of proMMP2 to MMP2, binding to proMMP2
and sequestering the quiescent protein near MMP14 (Butler et al. 1998). Overall, several levels
of regulation exist that control collagen synthesis and degradation in the cycling uterus.
1.9.3 Role of Extracellular Matrix during Pregnancy
Components of the ECM are highly involved in the various processes of a successful
pregnancy, including decidualization, implantation, placentation, and involution (Figure 10).
Serious complications or pregnancy failure may arise from aberrant ECM expression or activity
(Plaks et al. 2013, Singh et al. 2013, Pereza et al. 2012). During decidualization, endometrial
fibroblasts differentiated into decidual cells and denoted potential implantation sites
(Abrahamsohn and Zorn 1993). Surrounding these sites were collagen fibrils organized into a
basket-like structure comprised of collagens I, III, V, and VI (Carbone et al. 2006, Dziadek et al.
1995). Collagen IV and laminin were also expressed in the pericellular region of stromal cells in
the decidua (Iwahashi et al. 1996). Collagen diameter and thickness were greatly increased in the
decidua, while the collagen fibrils in the normal endometrium were unaffected (Spiess et al.
2007, Spiess and Zorn 2007, Teodoro et al. 2003, Iwahashi et al. 1996, Zorn et al. 1986).

37
The overall amount of collagen in the decidua was increased, particularly collagen V,
which was expressed exclusively in the decidua. In humans, the ratio of collagen III/collagen I
was decreased and the ratio of collagen V/collagen I was significantly increased in the decidua
compared to normal endometrial tissue. Collagen V played a vital role in successful
decidualization and implantation, since a low collagen V/collagen I ratio was found in decidual
tissues in which a spontaneous abortion occurred (Iwahashi and Nakano 1998). It was
hypothesized that the ECM greatly increases in thickness and amount in order to reinforce the
decidual cells as they support the implanting and developing embryos, as well as forming a
barrier to prevent the invasion of immune cells and subsequent rejection of the embryo (Carbone
et al. 2006). The process of placentation and trophoblast invasion also involved alterations in
collagen accumulation, mainly collagens I, III, and IV (Kitaoka et al. 1996, Kitaoka et al. 1994).
During involution, the total amount of collagen rapidly decreased, reaching nulliparous levels by
day 5 in rats and 1-2 weeks in humans (Morrione and Seifter 1962, Harkness and Moralee 1956).
Figure 10. Schematic of decidualization and implantation in the human uterus. The embryo attaches to the luminal epithelium (LE) and begins pushing through the basement membrane (BM) to invade the underlying stroma. Simultaneously, stromal cells are differentiating into decidualized cells. Part of the trophoblast eventually attaches as the placenta. This figure is adapted from Sharkey and Macklon (2013) with the addition of labels denoting stages of embryo implantation and invasion.

38
During decidualization, endometrial decidual cells exhibited an upregulation of
expression of TIMPs (Nuttall and Kennedy 1999), potentially to inhibit degradation of collagen
surrounding the implantation sites by the increased expression of MMP2 (Tsai et al. 2009).
Implantation and placentation occurred due to a balance between expression and activity of
MMPs and TIMPs. In order to enable invasion of trophoblast cells through the decidua, MMP2
and 9 degraded collagen IV in the basement membrane (Staun-Ram and Shalev 2005).
Expression of MMP2 and 9 and TIMPs was also found at implantation sites in the uterus and in
pre-implantation embryos (Fontana et al. 2012, Wang et al. 2003, Das et al. 1997). Co-
localization of MMP2, MMP14, and collagen IV mRNA was found in invading trophoblast cells,
as well as co-localization of MMP2 and MMP14 mRNA in decidual cells (Bjorn et al. 1997).
Human chorionic gonadotropin (hCG), a hormone indicative of pregnancy released by the
trophoblast, increased MMP2 and 9 secretion from trophoblast cells and decreased TIMP-1, -2,
and -3 expression in endometrial stromal cells to aid in trophoblast invasion and placentation
(Fluhr et al. 2008). During involution in the human uterus, the overall collagen content
significantly decreased (Morrione and Seifter 1962). Using a rodent model, mRNA expression of
Mmp2 and Mmp14 and activity of MMP2 were highly increased until five days postpartum. At
this time, Timp2 mRNA expression increased to inhibit MMP2 expression and activity (Manase
et al. 2006), which aid in explaining the significant decrease in collagen in the involuting human
uterus postpartum.
Preeclampsia, a serious and potentially life-threatening pregnancy complication, may be
promoted by aberrant expression of MMP2 and 9 to alter the ECM components of the
decidualized areas of the endometrium (Plaks et al. 2013, Lockwood et al. 2008). Increased
MMP9 and MMP2/TIMP-2 complex expression in serum or genetic polymorphisms in MMP2 or

39
MMP9 gene promoters were correlated with spontaneous abortions (Nissi et al. 2013, Pereza et
al. 2012). Because of the associated increase in implantation failure, spontaneous abortions, or
pregnancy complications due to aberrant MMP expression, collagen and MMPs play distinct and
important roles in preparation of pregnancy, during pregnancy, and postpartum.
1.10 Uterine Disorders
1.10.1 Leiomyoma
1.10.1.1 Description, Symptoms, and Treatment of Leiomyoma
Leiomyoma, or uterine fibroids, are benign fibrotic growths in the myometrium that arise
from smooth muscle cells and are one of the most common benign pathologies to affect women
of reproductive age (Baird et al. 2003, Kempson and Hendrickson 2000). Some of the more
common symptoms in women with uterine fibroids are heavy and prolonged menstrual bleeding,
dysmenorrhea, pelvic pain or pressure, anemia, urinary incontinence, constipation, lower back
pain, compression of other pelvic organs, sexual dysfunction, infertility and recurrent pregnancy
loss (Commandeur et al. 2015). Treatment usually involves administration of GnRH agonists,
progestins, selective PR modulators, or oral contraceptives, implantation of levonorgestrel
intrauterine devices, or interventions such as myomectomy (removal of fibroids while preserving
the uterus), uterine artery embolization, and magnetic resonance (MR)-guided ultrasound
treatments. However, these treatments are often variable in effectiveness or ineffective all
together (Commandeur et al. 2015, Vilos et al. 2015, Gorny et al. 2014). In most instances,
hysterectomy is the only effective treatment for fibroids and is the most common cause for
hysterectomy in the United States (Commandeur et al. 2015, Vilos et al. 2015).

40
1.10.1.2 Etiology of Leiomyoma
Hormonal and genetic factors play a role in the development of uterine fibroids. For
example, preadolescent and early adolescent and postmenopausal females have a low risk due to
the low circulating levels of sex hormones. Women taking oral contraceptives or that have had
multiple pregnancies also have a lower risk compared to normally cycling women (Commandeur
et al. 2015, Ross et al. 1986). Epidemiological studies have correlated having relatives with
leiomyoma as a risk factor for a woman to develop leiomyoma (Sato et al. 2002). Ethnicity also
plays a role, with African American woman developing fibroids more often and at a younger age
than Caucasian women (Baird et al. 2003).
Fibroids are clonal in origin and all cells in the mass arise from one cell, suggesting that
the parental cell has multipotent stem cell properties (Commandeur et al. 2015, Holdsworth-
Carson et al. 2014, Cai et al. 2007, Rogalla et al. 1995). Conditional knockout of β-catenin in the
embryo led to adipose gradually replacing smooth muscle cells in the Müllerian duct
mesenchyme (Arango et al. 2005). Cells in the uterus were immunopositive for α-smooth muscle
actin (α-SMA), ERα, and β-catenin and were located near cells positive for stem cell markers
and negative for several hematopoietic markers (Szotek et al. 2007). These studies provided
strong evidence that there exists a population of myometrial stem or progenitor cells, leading to
the theory that these stem/progenitor cells become dysregulated by a genetic mutation and
differentiate into actively dividing fibroid cells instead of smooth muscle cells (Bulun 2013). The
mediator complex subunit 12 (MED12) gene was mutated in over 70% of uterine fibroid samples
(Makinen et al. 2011), suggesting that mutations in MED12 may play a role in abnormal
differentiation of stem/progenitor cells.

41
1.10.1.3 Regulation of Collagen Accumulation in Leiomyoma
The connection between fibrosis and alterations in collagen accumulation in uterine
fibroids is well established. During the proliferative phase, mRNA for collagen I and III was
overexpressed in uterine fibroids compared to surrounding tissue, suggesting that ECM
components are under hormonal control within the fibroid (Stewart et al. 1994, Brandon et al.
1993). This idea was supported by in vitro studies that treated normal myometrial and
leiomyoma cells with low levels of E2 and observed significant increases in collagen
biosynthesis and in vivo studies reporting that estrogen treatment promoted proliferation of
fibroids through the activation of fibroblasts (Luo et al. 2014, Zbucka et al. 2007). The
orientation and alignment of collagen fibrils in the fibroid were also altered, suggesting
differences in the amount of proteoglycan between the fibrils or alterations in the number of
cross links (Leppert et al. 2004). Expression of MMP2 and MMP14 and activity of MMP2 were
also increased in leiomyoma samples (Tsigkou et al. 2015, Wang et al. 2015, Bogusiewicz et al.
2007), potentially in an attempt to regulate the increased amount of collagen present in the
fibroid.
1.10.2 Endometriosis
1.10.2.1 Description, Symptoms, and Treatment of Endometriosis
Endometriosis is a benign estrogen-dependent gynecological condition characterized by
the presence and growth of ectopic endometrial glands and stroma, often associated with
inflammation and increased fibrosis. Ectopic tissue is most often found in the pelvic region or
peritoneum, but it is also found in the bowel, diaphragm, or pleural cavity. Prevalence of
endometriosis in the general population ranges from 6 to 10%; however, in women with
infertility or pain, prevalence increases to 35-50% (Giudice and Kao 2004). The most common

42
symptom reported is chronic pelvic pain in 58% of women. Between 30 and 50% of women with
endometriosis are also infertile; however, the mechanism and causation are not well understood
(Hickey et al. 2014). Laparoscopy is the only tool to definitively diagnose a woman with
endometriosis due to the lack of biomarkers (Fassbender et al. 2015, Macer and Taylor 2012,
Kennedy et al. 2005). Treatments for endometriosis center mainly on pain management or relief.
Several accepted therapies include the use of GnRH analogues or levonorgestrel intrauterine
devices. Laparoscopic surgery is often used to ablate and excise the tissue or transect nerves.
These interventions significantly reduce pain and lead to increased pregnancy and live birth rates
(Brown and Farquhar 2015, Kennedy et al. 2005).
1.10.2.2 Role of Extracellular Matrix in Pathogenesis of Endometriosis
Retrograde menstruation, in which endometrial tissue is disseminated and implanted into
the peritoneal cavity via the fallopian tubes, is the most accepted theory for the pathogenesis of
endometriosis (Ahn et al. 2015, Sampson 1927). Between 75-90% of women undergo retrograde
menstruation; however, women that develop endometriosis are likely to have other genetic and
biochemical factors leading to development of the disease (Ahn et al. 2015). It is not clearly
understood how the endometrial tissue attaches to the peritoneum, whether it is directly to the
peritoneal mesothelium or to the underlying ECM (Adachi et al. 2011, Lucidi et al. 2005, Witz et
al. 2003, Dunselman et al. 2001, Nisolle et al. 2000, Sillem et al. 1999). Differences in
expression of adhesion molecules that are part of the ECM and ECM signaling, such as integrins
and cadherins, appear to play a role in the development of endometriosis (reviewed in Young et
al. (2013)). Numerous integrin subtypes were present in human peritoneal mesothelium and in
endometriotic lesions in a nude mouse model (Hull et al. 2008, Witz et al. 2000). Endometriotic
samples also had impaired expression and localization of several integrin receptors, suggesting a

43
role in enhanced invasion (Giannelli et al. 2007, Klemmt et al. 2007, Sillem et al. 1999). E-
cadherin was expressed in healthy peritoneal and normal endometrial tissue (Jimenez-Heffernan
et al. 2004, Groothuis et al. 1999). An increased number of endometriotic lesions appeared to be
E-cadherin negative though (Gaetje et al. 1997). Increased expression of P-cadherin in
endometriotic tissue compared to normal endometrial tissue suggested that it also played a role in
increased cell adhesion (Chen et al. 2002).
Endometriotic tissue invasion was also facilitated by MMPs (Sharpe-Timms and Cox
2002). Expression of numerous MMPs, including MMP3, 9, 10, 12, and 13, was increased in
human samples and animal models. The most common MMP to increase was MMP9.
Interestingly, eutopic endometrial tissue in women with endometriosis also had increased
expression and activity of MMP9 compared to normal eutopic endometrial tissue. Expression of
TIMP-1 appeared to vary though (Pelch et al. 2010, Pino et al. 2009, Collette et al. 2006, Collette
et al. 2004). Increased MMP9 expression and decreased TIMP-1 expression was also found in
peritoneal fluid (Szamatowicz et al. 2002). These findings suggest that MMP9 may aid in
degrading the ECM in the peritoneum, allowing ectopic endometrial tissue to invade and implant
into the peritoneal cavity.
1.10.3 Pyometra
1.10.3.1 Description, Etiology, and Treatment of Pyometra
Pyometra is an inflammatory condition characterized by the accumulation of intraluminal
purulent material in the uterus and can be divided into open- and closed-cervix distinctions.
Open-cervix pyometra means that the purulent fluid can drain, while closed-cervix pyometra
means that the fluid remains trapped in the uterine cavity. Closed-cervix pyometra is an
emergency requiring rapid intervention to prevent systemic infection and death (Smith 2006). In

44
humans, pyometra is extremely rare yet life threatening, with a 0.04% incidence rate in the
general population. The development of pyometra has been reported in children as young as four
months of age. In the elderly, this rate jumps to >13%, showing an age-dependent increase in
development (Barry 2015, Geggie and Walton 2006, Yildizhan et al. 2006). In humans, pyometra
often occurs due to impairment in natural drainage subsequent to damage from a malignant
disease such as cervical cancer and radiotherapy treatment (for example Vyas et al. (2009),
Yildizhan et al. (2006)). In veterinary medicine, pyometra is most common in intact canines,
with nearly 25% presenting with the disease before 10 years of age (Smith 2006, Egenvall et al.
2001), but pyometra is also found in small and captive large felids, livestock, mares, and even
rhesus monkeys, rhinoceros, and orca whales (Knudsen et al. 2015, Patisaul 2012, McCain et al.
2009, McCarthy et al. 1989, Ridgway 1979).
Treatment for pyometra varies with the severity of the condition. Complete
ovariohysterectomy is preferred in older animals, those with systemic inflammation, or animals
with closed-cervix pyometra (Smith 2006). Otherwise, the standard protocol is treatment with
antibiotics, fluids, and either prostaglandins or antiprogestins in an attempt to reverse progression
and save breeding capabilities (Fieni et al. 2014, Smith 2006).
A combination of hormonal, genetic, and inflammatory or infectious factors play a role in
the etiology of pyometra. However, not all rodent strains treated with estrogenic compounds will
develop pyometra. Treatment with estrogenic or progestogenic compounds, including
phytoestrogens, E2, DES, and contraceptives, in various species led to the development of the
disease (Asa et al. 2014, Patisaul 2012, Brossia et al. 2009, Gould et al. 2005, Pandey et al. 2005,
McCarthy et al. 1989, McLachlan et al. 1980, Burrows 1936). Brown Norway, but not Sprague
Dawley, ACI, or Fischer 344, rats developed pyometra after E2 treatment or DES exposure

45
(Brossia et al. 2009, Gould et al. 2005, Pandey et al. 2005). Using quantitative trait loci (QTL)
mapping, key genetic variations between the Brown Norway, ACI, and Fischer 344 strains were
found in Eutr1 for E2-induced pyometra and Eutr2 for DES-induced inflammation and
pyometra, supporting that genetic factors play a role in the etiology of the disease (Gould et al.
2005, Pandey et al. 2005). Strain susceptibility can also be used to identify other endpoints that
are estrogen-responsive as well.
1.10.3.2 Role of Bacteria and the Immune System in the Development of Pyometra
During metestrus in the reproductive cycle, the uterus is primed for embryonic
implantation and the maternal immune system is decreased in order to allow the embryo,
considered a foreign body, to implant. However, bacterial infection in the uterus from the normal
flora in the vaginal or digestive tract is also highly likely to occur during this stage due to the
decreased immune response. In roughly 70% of cases in canines in which bacteria are present in
the uterine cavity, Escherichia coli was cultured with a high percentage displaying virulence
factors (Agostinho et al. 2014, Hagman 2012, Coggan et al. 2008, Hagman and Kuhn 2002).
Canines inoculated with E. coli at different estrous cycle stages only developed a severe
pyometric phenotype when inoculated in early metestrus (Tsumagari et al. 2005). In the bursa
surrounding the ovaries in canines, E. coli was present but there was also bacterial isolates of
strains of Staphylococcus and Streptococcus, among others (Rubio et al. 2014), suggesting that
infection from several different bacterial types can lead to the development of pyometra.
Antibiotics are often used to combat the bacterial infection; however, 50% of canines with
pyometra had multidrug resistance (Agostinho et al. 2014), suggesting that treatment with
antibiotics may not be useful and could lead to further antibiotic resistance.

46
The presence of bacteria leads to an immune or inflammatory response in the uterus.
Toll-like receptors (TLRs) are important in regulating the innate and adaptive immune responses
in the host tissue in response to bacterial lipopolysaccharide (LPS) binding (Takeda et al. 2003,
Janeway and Medzhitov 2002, Hoshino et al. 1999). In canines with pyometra, mRNA and
protein expression of TLR2 and TLR4 were significantly increased in all cell types examined
(Chotimanukul and Sirivaidyapong 2012, 2011, Silva et al. 2010), potentially due to the
increased bacterial content in the uterus. This increased expression could also represent a change
associated with the development of pyometra, rather than be playing a causative role. Other
immune cells types (ie. granulocytes, macrophages, and leucocytes) were decreased in cows with
pyometra, while lymphocytes were increased (Brodzki et al. 2014). Expression of mRNA and
protein for pro-inflammatory interleukins (ILs), including IL-1α, IL-1β, and IL-6, were increased
in pyometric samples from affected animals (Brodzki et al. 2015, Voorwald et al. 2015,
Bukowska et al. 2014). Anti-inflammatory ILs, such as IL-4 and IL-10, were also increased in
animals with pyometra (Maciel et al. 2014), suggesting an attempt by the animal to combat the
effects of the present bacteria and developing disease. Alterations in cytokine production resulted
in differences in prostaglandin synthesis. Prostaglandin F2α synthase (PGFS) and prostaglandin
E2 synthase (PGES) mRNA levels were significantly increased in animals with pyometra,
leading to increases in prostaglandin E2 and F2α levels (Silva et al. 2010). It has been proposed
that several of the altered proteins could be potential predictive biological markers to detect
animals with pyometra or animals which could develop pyometra without invasive procedures
(Dabrowski et al. 2015, Voorwald et al. 2015, Dabrowski et al. 2013, Karlsson et al. 2012).

47
1.10.3.3 Involvement of Sex Hormones and Collagen Accumulation in Pyometra
Alterations in sex hormone receptor levels and collagen accumulation were observed in
canines and felids with pyometra. Serum P levels were unaffected, but serum E2 levels were
greatly increased in both halves of diestrus and early anestrus (Ververidis et al. 2004). While
progesterone receptor (PR) expression was decreased in surface epithelium only, ERα expression
was decreased in all areas of tissue, including surface epithelium, stromal cells, normal glands,
cystic glands, and smooth muscle (Kunkitti et al. 2011, Misirlioglu et al. 2006, De Bosschere et
al. 2002). Within the cervix of canines with pyometra, the closed-cervix phenotype had a
significantly decreased percentage of collagen compared to open-cervix, while both had a
decreased percentage of collagen compared to normal cervical tissue (Linharattanaruksa et al.
2014). These decreases in collagen may be due to the substantial upregulation in MMPs and
associated activity (Voorwald et al. 2015, Hagman 2012, Chu et al. 2002).
1.10.4 Equine Endometrosis
1.10.4.1 Description, Etiology, and Treatment of Equine Endometrosis
Equine endometrosis is distinct from human endometriosis and is defined as an age-
related and irreversible degenerative uterine disease characterized by gland nest structures
surrounded by increased endometrial stromal and periglandular fibrosis (EPF). The disease can
be classified into Kenney categories I, IIA, IIB, and III based on disease severity, progression,
and amount of fibrosis and inflammation (Buczkowska et al. 2014, Hanada et al. 2014, Snider et
al. 2011, Evans et al. 1998, Ferreira-Dias et al. 1994, Ricketts and Alonso 1991, Kenney 1978).
When classifying equine endometrosis, several potential pathological changes are examined,
including the presence of inflammatory infiltrates, infection, fibrosis and gland nesting,
hypertrophy associated with fibrosis, lymphatic lacunae, cystic glands or nonglandular cysts,

48
atrophy/hypoplasia, and glandular hyperplasia. Kenney category I is characterized by no
pathologic changes or changes that are slight and scattered. Category I mares do not have an
impaired ability to support a fetus to term. Kenney category II is characterized by more extensive
or severe inflammatory changes, increase in fibrosis of individual gland branches or gland nests,
presence of atrophy, and presence of lacunae but are not large enough to palpate. Mares in
category II have impairment in the ability to carry a foal to term. The severity of the fibrotic
changes distinguishes categories IIA and IIB, with category IIA having 10-35% of glands
affected and category IIB having 35-60% of glands affected. Kenney category III is
characterized by widespread fibrosis, severe and widespread inflammation, and severe lymphatic
lacunae. These pathologic changes greatly reduce fertility, with only about 10% of mares having
the ability to carry a foal to term (Buczkowska et al. 2014, Kenney 1978). The severity, or higher
category, of EPF is inversely correlated to successful conception and gestation and is associated
with increased rates of embryonic or fetal foal loss and increased susceptibility to infection
(Buczkowska et al. 2014, Kenney 1978). An endometrosis-like phenotype has also been reported
in canine biopsy samples (Christensen et al. 2012).
While it is unclear if genetic or hormonal factors are involved in the etiology of equine
endometrosis, the involvement of increased immune cell infiltration has been reported. Increased
neutrophil infiltration possibly occurred in response to bacterial challenge or non-infectious
stimuli, such as semen (Rebordao et al. 2014). This increase in inflammation led to the formation
of neutrophil extracellular traps (NETs) containing pro-inflammatory proteins that damaged
nearby tissue and contributed to the development of fibrosis (Rebordao et al. 2014).
No effective treatment for equine endometrosis has been developed. Intrauterine
infusions with 30% dimethyl sulfoxide (DMSO) in 0.9% saline decreased inflammatory cells and

49
periglandular fibrosis in barren mares but fertility rates remained low (Ley et al. 1989).
However, follow-up studies to determine if the severity of endometrosis returned to that
observed pre-treatment were lacking. Recently, mesenchymal stem cell (MSC) therapy has been
proposed as a potential treatment for endometrosis due to their ability to give rise to several cell
types and their trophic mediation, including in inflammatory and fibrotic processes. The
differentiated cells then secrete growth factors, cytokines, and MMPs to degrade the fibrotic
components and inhibit the inflammatory response (van Poll et al. 2008, Caplan and Dennis
2006, Aggarwal and Pittenger 2005, Caplan 1991). A simple delivery method, similar to that
used for artificial insemination, using equine adipose-derived MSCs (eAD-MSCs) has been
developed and eAD-MSCs delivered through this technique are incorporated into and distributed
throughout uterine tissue (Mambelli et al. 2013). Mares with endometrosis treated with eAD-
MSCs exhibited decreases in vimentin, laminin, cytokeratin 18, and α-SMA staining as early as
Day 7 after treatment, as well as improved morphological characteristics. An increase in Ki-67
staining was present in control and experimental groups at Days 7, 21, and 60 after treatment, but
the experimental group exhibited larger increases at all three time points. However, a large
sample size was lacking, with only four mares treated with eAD-MSCs, and one mare with
advanced degeneration exhibiting no changes in protein expression with treatment (Mambelli et
al. 2014). While decreases in phenotypic markers were present, it was unknown if symptoms
were decreased or if fertility was improved. This method is limited by the local nature of
treatment, preventing progression or reoccurrence of endometrosis, and will require a suitable
systemic treatment method to be developed as well in order to fully abolish endometrosis from
the endometrium. Also, the treatment may not be useful on mares with highly advanced disease.

50
Several questions still exist about etiology, progression, and treatment of equine endometrosis
without an experimental model that can be used for rapid and high powered experimentation.
1.10.4.2 Progression of Equine Endometrosis
Early stages of endometrosis were accompanied by periglandular-associated stromal cells
that actively produce collagen fibers (Buczkowska et al. 2014). A progression of the disease was
proposed based on alterations in proteins involved in proliferation and ECM and sex hormone
protein levels (Hoffmann et al. 2009b). After the early stages of fibrosis were underway, either
an active or inactive non-destructive phenotype developed (Table 2); however, the factors
regulating if the tissue becomes an active or inactive phenotype were unknown (Hoffmann et al.
2009b) . An active phenotype was characterized by the presence of metabolically active fibrotic
stromal cells and myofibroblasts, while an inactive phenotype was characterized by the
arrangement and contractibility of myofibroblasts (Hoffmann et al. 2009b). However, the
presence of myofibroblasts in general was indicative of advanced endometrosis (Walter et al.
2001).
To distinguish between the active and inactive phenotypes, immunohistochemical
examinations was performed, with the inactive phenotype having a mild decrease in Ki-67
antigen in fibrotic stromal cells compared to normal tissue while the active phenotype was not
changed. In glandular epithelia, the inactive phenotype had decreases in ERα and PR compared
to normal tissue while the active phenotype had increases in both receptors (Hoffmann et al.
2009b). Eventually, endometrosis progressed to an active or inactive destructive form. An active
destructive form resulted from an increase in myofibroblasts and ECM proteins and the inactive
destructive phenotype arose from mechanical stress (Hoffmann et al. 2009b). Based on an
immunohistochemical assessment, inactive destructive endometrosis again had a decrease in Ki-

51
67 antigen in fibrotic stromal cells compared to normal tissue while the active destructive type
did not. Active destructive endometrosis had 50-80% of cells staining for fibronectin or
proteoglycane, while the inactive destructive phenotype had 20-40% of cells staining for
fibronectin and 10-20% of cells staining for proteoglycane. Finally, the amount of Ki-67 antigen
in the glandular epithelia was decreased in the active destructive phenotype compared to normal
tissue, while the inactive destructive phenotype had no change in the amount of Ki-67 antigen. In
the glandular epithelia, the active destructive phenotype displayed decreases in ER while the
active non-destructive phenotype displayed increases in ER (Hoffmann et al. 2009b). However,
this proposed progression was limited due to the potential bias when estimating the percentage of
positive cells and the staining intensity. Some of the proposed changes appeared too small to
quantify, so the distinction between active and inactive phenotypes might not be meaningful.
Table 2. Characteristics of Equine Endometrosis Phenotypes
Non-destructive Destructive Active Inactive Active Inactive
Fibrotic Stromal Ki-67 − ↓ − ↓
Glandular Epithelium Ki-67 − − ↓ −
PR ↑ ↓ ↓ ↓ ERα ↑ ↓ ↓ ↓
ECM Fibronectin 10-20% 20-40% 50-80% 20-40%
Proteoglycane 10-20% 10-20% 50-80% 10-20%
1.10.4.3 Characterization of Extracellular Matrix Alterations in Equine Endometrosis
Because fibrosis is highly involved in the development of endometrosis, varying
expression patterns of ECM proteins between the defined categories or phenotypes were
characterized. Periglandular-associated fibroblasts produced increased levels of collagen IV,
laminin, and fibronectin. These fibroblasts appeared to have also differentiated into

52
myofibroblasts, based on their expression of α-SMA, tropomyosin, transforming growth factor-
β1 (TGF-β1), and 11β-hydroxysteroid dehydrogenase (11β-HSD2) (Ganjam and Evans 2006,
Walter et al. 2001). While synthesis of ECM proteins was increased in endometrosis, MMP2 and
tissue transglutaminase 2 (TG2), an enzyme involved in ECM protein cross linking, expression
was also increased (Walter et al. 2005). Because TG2 cross links are resistant to proteolytic
cleavage, the rate and level of collagen deposition was increased (Walter et al. 2005, Skill et al.
2004, Greenberg et al. 1991). While TG2 was degraded by an MMP2/MMP14 complex, it also
interacted with an intermediate form of MMP2 and decreased the rate of MMP2 maturation
(Walter et al. 2005, Belkin et al. 2004). This imbalance in collagen and MMP2 homeostasis may
lead to the increased fibrosis observed in affected, degenerating regions of the uterus.
1.10.4.4 Fetal Foal Loss
Fetal foal loss in mares with equine endometrosis may be due to alterations in the
endometrial environment, specifically expression patterns of interleukins and prostaglandins that
are involved in estrous cycle regulation and pregnancy in several farm animals (Franczak et al.
2010, Weems et al. 2006, Tanikawa et al. 2005). Myofibroblasts, which are present in advanced
endometrosis, increased cytokine production (Zhang et al. 1996). When comparing Kenney
categories II and III to category I in each stage of the estrous cycle, significant alterations in
prostaglandin-endoperoxide synthase (PTGS-2), PGFS, and PGES mRNA levels were found,
which correlated with alterations in prostaglandin F2α and prostaglandin E2 concentrations
(Szostek et al. 2012). Comparing the same categories within the stages of the estrous cycle,
significant differences in IL mRNA transcription were observed as well, which translated into
differences in protein expression. Specific IL proteins examined include IL-1α, IL-1β and the IL-
1 receptor subunits (IL-1RI and IL-1RII), and IL-6 and the IL-6 receptor (IL-6Rα) (Szostek et al.

53
2013). Endometrial samples from each category were treated with either IL-1α, IL-1β, or IL-6 to
determine the influence of ILs on prostaglandin secretion and synthase mRNA transcription.
Each category exhibited distinctive alterations in patterns of secretion and transcription levels
(Szostek et al. 2013). The increased differentiation of fibroblasts into myofibroblasts in
categories II and III of endometrosis may be leading to altered expression in ILs and subsequent
altered prostaglandin synthase expression and prostaglandin concentrations. The abnormal
endometrial environment then resulted in decreased fertility and fetal loss.
Several proteins are normally secreted from endometrial glands during the estrous cycle
in fertile humans and animals. Aberrant expression of these proteins may indicate a reduction in
fertility in general and may be used to determine the differences between fertile and barren
mares. Uteroglobin (UG), uterocalin (UC), uteroferrin (UF), and calbindinD9k (CAL) are
important secreted proteins involved in a successful pregnancy. UG has anti-inflammatory
properties and prevents the maternal immune system from attacking trophoblastic cells during
implantation and placental growth (Mukherjee et al. 1994, Miele et al. 1990). UC is a carrier
protein that transports hydrophobic proteins and molecules to the conceptus (Suire et al. 2001,
Crossett et al. 1998). UF is primarily an iron transporter to the conceptus, but also is a
hematopoietic growth factor (Bazer et al. 1991, Ducsay et al. 1984, McDowell et al. 1982). CAL
is a Ca2+-binding transporter in the uterus (Wooding et al. 1996, Inpanbutr et al. 1994). In both
non-destructive and destructive endometrosis, expressions of UG, UC, and CAL were largely
decreased compared to unaffected tissue, while UF expression was increased (Lehmann et al.
2011, Hoffmann et al. 2009a). Decreases in secreted proteins essential for viable growth of a
fetus in endometrosis also played a significant factor in decreased fertility and fetal loss.

54
1.11 Statement of Purpose
1.11.1 Objective and Rationale
To repeat, the overall objective of this dissertation research was to characterize the
endocrine disrupting effects of BPA or EE exposure in the uterus of two mouse strains. Two
exposure paradigms were used: 1) exposure in adulthood, during which time mice mated and
produced a litter, and 2) whole life exposure without breeding. Previous studies examined
incidence of pathologies after a much longer exposure time period than is proposed in this
dissertation research. Uterine immune responsiveness and fibrosis are components of several
human and animal uterine pathologies, and the effect of BPA or EE exposure on these endpoints
has not been studied.
1.11.2 Central Hypothesis and Specific Aims
The central hypothesis for this dissertation was that exposure to estrogenic EDCs, such as
BPA or EE, will influence E2-regulated uterine physiology, leading to an increased incidence of
uterine pathology. A second hypothesis was that a greater sensitivity in the C57Bl/6N strain to
the estrogenic actions of EDCs also increases the incidence of uterine pathology compared to the
CD1 strain.
The specific aims of this dissertation research included 1) investigating the endocrine
disrupting actions of EDCs in both exposure models on the development of uterine pathologies,
2) examining the effects of EDCs in both exposure models on the immune responsiveness in the
uterus, 3) investigating the ECM changes associated with EDC exposure in both models, and 4)
comparing the pathological, immune, and ECM alterations between the C57Bl/6N and CD1
strains.

55
CHAPTER 2
METHODS

56
2.1 BPA and EE Exposure
2.1.1 Animal Housing
All animal procedures were performed in accordance with protocols approved by the
University of Cincinnati Institutional Animal Care and Use Committee and followed
recommendations of the Panel on Euthanasia of the American Veterinary Medical Association.
C57Bl/6NHsd and Hsd:ICR (CD1) mice (Harland; Indianapolis, IN) aged six to seven weeks
were evenly distributed between the nine exposure groups, with five of a single sex occupying a
cage for an acclimation period of two weeks. During this period, general health and vaginal
cytology were monitored daily and body weight and food consumption were monitored weekly.
Animals were kept on a polycarbonate-free housing system with single-use polyethylene cages
and water bottles certified BPA-free (Innovive; San Diego, CA). Sanichip bedding (Irradiated
Aspen Sani-chip; PJ Murphy Forest Products Corp.; Montville, NJ) was used to prevent
contamination from mycoestrogens in corncob bedding (Thigpen et al. 2013). Sterile drinking
water containing <1% of oxidizable organics was produced from a water purification system
(Millipore Rios 16 with ELIX UV/Progard 2; Billerica, MA). A 14-hour on, 10-hour off light
cycle was maintained throughout the study. Diet and water were provided ad libitum.
2.1.2 Exposure
C57Bl/6N and CD1 mice were randomly assigned and divided equally between a control
group (0 ppm), five BPA groups (0.03, 0.3, 3, 30, or 300 ppm), and three EE groups (0.0001,
0.001, 0.01) as a positive control for estrogenic activity. In a pilot study, higher EE doses were
used (0.1 and 1.3 ppm) but were discontinued in later phases. Mice remained on diet from arrival
to necropsy. Offspring were exposed to BPA or EE from conception to necropsy (Figure 1).
Food consumption was recorded weekly. The amount of food eaten per cage was divided by the

57
number of animals per cage and the number of days between food weight collections to estimate
the amount of food consumed per animal. Actual doses of BPA or EE were calculated based on
estimated food consumption. Animals were fed a defined casein-based phytoestrogen-free diet
(Product # D1010501, Research Diets, Inc.; New Brunswick, NJ) unsupplemented (control or 0
ppm) or supplemented with either BPA (2,2-bis(4-hydroxyphenyl)propane; CAS No. 80-05-7;
Lot 11909; USEPA/NIEHS standard) or EE (1,3,5(10)-estratrien-17α-ethinyl-3,17β-diol; CAS
No. 57-63-6; Batch No. H923; Steraloids Inc.; Newport, RI) that was incorporated
homogenously in the diet at each desired concentration. Final BPA concentrations (0.03, 0.3, 3,
30, and 300 ppm) resulted in doses of 0.004, 0.04, 0.4, 4, and 40 mg/kg/day. Concentrations of
EE (0.0001, 0.001, and 0.01 ppm) resulted in doses of 0.00002, 0.0002, and 0.001 mg/kg/day
(Kendziorski and Belcher 2015, Kendig et al. 2012, Kendziorski et al. 2012). Details of diet
composition are found in Table 1 and amounts of BPA or EE and dyes incorporated into the diets
are found in Table 2.
Figure 1. Exposure paradigm for sires, dams, and offspring in control and BPA or EE exposure groups.

58
Table 1. Composition of Defined Control and Test Diets
Macronutrients Source gm% kcal% Ingredients g kcal
Protein Casein 18.1 20 Casein 200 800
Carbohydrate D-glucose 60.3 65 L-Cysteine 3 12
Corn Starch Maltodextrin
Corn Starch 346 1384
Fat Soy Oil 6.2 15 Maltodextrin 10 45 180
Fiber Cellulose 11.7 Dextrose 250 1000
Inulin 2.3 Cellulose, BW 200 125 0
Total ME (kcal/g)* 3.63 100 Inulin 25 25
Soybean Oil 70 630
Mineral Mix S10026 10 0
Dicalcium Phosphate 13 0
Calcium Carbonate 5.5 0
Potassium Citrate, 1
H2O 16.5 0
Vitamin Mix V10001 10 40
Choline Bitartrate 2 0
Total 1121.05 4071
*Metabolizable energy estimate based on the Atwater factors: protein: 4 kcal/g; fat: 9 kcal/g; carbohydrate: 4 kcal/g
Table 2. Amounts of EE, BPA, and Dyes Incorporated into Diets Control BPA EE 0 0.03 0.3 3 30 300 0.0001 0.001 0.01 0.1 1.3
17α-Ethinyl Estradiol (µg)
0 0 0 0 0 0 0.12 1.2 12 120 1470
Bisphenol A (µg) 0 33.7 337 3370 33700 337000 0 0 0 0 0
Red Dye #40,
FD&C (g) 0 0.04 0 0.04 0.025 0 0 0.05 0.025 0 0.05
Blue Dye #1, FD&C (g)
0 0.01 0.04 0 0 0.025 0.05 0 0.025 0.05 0
Yellow Dye #5, FD&C (g)
0.05 0 0.01 0.01 0.025 0.025 0 0 0 0 0

59
2.1.3 Breeding
After the acclimation period, males and females within an exposure group were paired
single-housed for breeding and observed daily within the first three hours of the light-on cycle
for a copulation plug. Once a plug was observed, vaginal cytological assessment ceased and the
breeding pair was separated. If a copulation plug was not observed after two weeks, female body
weight gain was used as an indicator of pregnancy and the breeding pair was separated. Females
were observed twice daily for general health, signs of pregnancy, and parturition (designated as
PND0). Offspring were separated at PND22 and group housed ≤5 per sex per cage. The dams
were housed 5 per cage after separation from their offspring.
2.1.4 Vaginal Cytology
In order to examine the stage of reproductive cycle and to ensure female mice were
analyzed and sacrifice occurred during the estrus phase of the estrous cycle, vaginal cytology
was performed. Vaginal lavage was performed daily during the first three hours of the light-on
cycle using a 0.09% sterile saline solution and a fire polished glass pipette. Samples were
collected and a microscopic cytological assessment was completed. Each stage of the cycle had
different morphological features to the vaginal cells in the collected sample. Estrus is indicated
by the presence of cornified, non-nucleated epithelial cells. The presence of infiltrating immune
cells such as leukocytes and few epithelial cells with nuclear degeneration is indicative of
diestrus. Proestrus is indicated by the presence of round, nucleated epithelial cells (McLean et al.
2012).
Estrous stage identification determined by vaginal cytology was confirmed by
morphological assessment of the hematoxylin & eosin (H&E)-stained uteri for C57Bl/6N and
CD1 dams and offspring. Staining of uteri is described in Section 2.2.2. Mice in estrus and

60
proestrus have a distended lumen, with the lumen being thicker in estrus, indicative of tissue
breakdown. Mice in diestrus have a thin lumen in the process of regeneration (Hedrich and
Bullock 2004). Mice that were classified as being in estrus or diestrus were used for all studies.
2.2 Assessment of Pathology
2.2.1 Necropsy and Tissue Preparation
Dams were sacrificed between 19 and 23 weeks of age, and offspring were sacrificed
~PND90. All animals were sacrificed in estrus, based on cytological assessment of vaginal cells.
Animals were evenly and randomly divided into a group in which all tissue would be frozen and
a group in which all tissue would be perfused. Randomization was based on the identification
number of the animal, with the first half being frozen and the second half being perfused. All
tissues were collected as part of a larger study (Kendig et al. 2012). Half of the study mice were
euthanized by rapid cervical dislocation to collect frozen tissue or CO2 asphyxiation to collect
perfused tissue. Tissues were quickly excised and excess fat and connective tissue were
removed. Tissues were weighed and placed in a 2 mL free standing microcentrifuge tube with
screw cap (Fisher Scientific; Waltham, MA) for freezing in a 100% ethanol/dry ice mixture.
Frozen tissue samples were stored at -80oC until used. For the other half of the study animals, the
chest cavity was opened and a cardiac puncture of the left ventricle was used in order to flush the
animal with heparinized saline and then perfuse with 10% neutral-buffered formalin (NBF;
Polysciences, Inc.; Warrington, PA). Tissues were collected, trimmed of excess fat, weighed, and
placed in properly sized cassettes and washed in multiple changes of 10% NBF and then washed
in six washes of 70% ethanol before tissue processing.
Tissues were processed using a Tissue Tek VIP 3000 Tissue Processor (Sakura/Miles
Scientific; Torrance, CA) using the following program: one wash in 70% ethanol for 20 minutes,

61
two washes in 80% ethanol for 20 minutes each, two washes in 95% ethanol for 20 minutes each,
one wash in 100% ethanol for 20 minutes, two washes in 100% ethanol for 30 minutes each, two
washes in xylene for 30 minutes each, and three washes in heated paraffin under vacuum seal for
30 minutes each (58oC; Type 3 paraffin, Richard Allen Scientific; Kalamazoo, MI). After
processing, tissues were embedded in paraffin (Histoplast IM; Richard Allen Scientific;
Kalamazoo, MI) using a Shandon Histocentre 3 Embedding Station (Thermo Shandon;
Kalamazoo, MI). To embed uteri, a section of one uterine horn was cut and embedded
transversely to assess myometrial wall thickness. The rest of each uterus was placed
longitudinally in the same paraffin block.
2.2.2 Hematoxylin and Eosin Staining
Paraffin-embedded uteri from C57Bl/6N and CD1 dams and offspring were sectioned at a
5 µm thickness on an HM 325 Rotary Microtome (Thermo Shandon; Kalamazoo, MI) and placed
on SuperFrost Plus slides (Fisher Scientific; Waltham, MA). Slides were incubated at 60oC for
>20 minutes in a hybridization incubator (Fisher Scientific; Waltham, MA). Once cooled to
room temperature (RT), an H&E stain was performed in order to visualize the nuclei of cells
(hematoxylin) and the basic compartments or cytoplasm (eosin). Sections were deparaffinized in
three washes of xylene for 5 minutes each, rehydrated through a graded series of ethanol (two
washes in 100% ethanol for 3 minutes each, two washes in 95% ethanol for 1 minute each, and
one wash in 70% ethanol for 1 minute), and rinsed in running tap water for 5 minutes. Slides
were rinsed in dH2O, stained in Hematoxylin 2 (Cat# 7231; Richard Allen Scientific;
Kalamazoo, MI) for 30 seconds, and rinsed in running tap water. Slides were placed in Clarifier
2 (Cat# 7442; Richard Allen Scientific; Kalamazoo, MI) for 30 seconds and rinsed in running tap
water for 30 seconds. Slides were placed in Bluing Reagent (Cat# 7341; Richard Allen

62
Scientific; Kalamazoo, MI) for 1 minute and rinsed in running tap water for 1 minute. Slides
were placed in 95% ethanol for 1 minute and counterstained with Eosin-Y (Cat# 7111; Richard
Allen Scientific; Kalamazoo, MI) for 2 minutes. Excess eosin was blotted with paper towel and
slides were dehydrated through three washes of 100% ethanol for 15 dips each. Slides were
placed in three washes of xylene for 15 dips each and coverslipped using Permount (Fisher
Scientific; Waltham, MA), which were allowed to dry overnight.
2.2.3 Myometrial Wall Thickness
Transverse sections of the H&E-stained uterine tube of C57Bl/6N and CD1 dams from
the preliminary study were measured in order to quantify the thickness of the inner and outer
myometrial layers. Using a Nikon Eclipse 55i microscope using a DS-Fi1 CCD camera
controlled with DigitalSight software, five random areas at 20x magnification of the outer edge
of the transverse section were imaged by an observer blinded to strain and exposure group.
Within each image, five measurements from the beginning of the inner myometrium to the outer
edge of the outer myometrium were collected using DigitalSight software. Using a stage
micrometer, the ratio of 60 µm/100 pixels was measured. Wall thickness measurements were
converted from pixels to micrometers (µm) using the following formula:
X = 60 µm (pixel measurement) 100 pixels
2.2.4 Histologic Assessment of Uterine Pathology
The H&E-stained uteri of C57Bl/6N and CD1 dams and offspring (PND21, PND49, and
~PND90) were assessed for distended glands, gland nests, cysts, and pyometra. Distended glands
are characterized by a thinning of the epithelial cells that make up the gland. Gland nests are
groups of glands that appear clumped together in an oval or circle and are surrounded by fibrotic

63
stromal cells instead of being dispersed throughout the endometrium. Cysts appear as circular
holes in the endometrium surrounded by a thin layer of epithelial cells. The presence of the
pathology and, when applicable, the number of incidences were noted for each uterus. The
observer was blinded to strain, exposure, and generation.
2.3 Assessment of Collagen Synthesis and Degradation and Immune Response
2.3.1 Picrosirius Red Staining and Collagen Accumulation
Paraffin-embedded uterine sections for C57Bl/6N and CD1 dams and offspring were cut
at a 5 µm thickness on an HM 325 Rotary Microtome and put on SuperFrost Plus slides. Slides
were incubated at 60oC for >20 minutes in a hybridization incubator. Slides were deparaffinized
in three washes of xylene for 5 minutes each, rehydrated in graded series of ethanol (two washes
in 100% ethanol for 2 minutes each, two washes in 95% ethanol for 2 minutes each, and one
wash in 70% ethanol for 2 minutes), and rinsed in dH2O. Slides were stained with Weigert’s
hematoxylin (American MasterTech; Lodi, CA) for 8 minutes and rinsed with running tap water
for 5 minutes. Sections were placed in a 0.2% phosphomolybdic acid hydrate in water for 2
minutes, rinsed with dH2O for 30 seconds, and placed in a Picrosirius Red F3BA (1.4% 2,4,6-
trinitrophenol, 0.4% Direct Red 80) solution in water for 60 minutes. Slides were transferred to a
0.1 N hydrochloric acid solution for 2 minutes and placed in 70% ethanol for 45 seconds. Slides
were dehydrated (one wash of 95% ethanol for 15 dips and three washes of xylene for 15 dips
each) and washed in xylene three times for 15 dips each. Sections were coverslipped using
Permount.
2.3.2 Immunohistochemistry
Uteri sections were cut at a 5 µm thickness on an HM 325 Rotary Microtome and put on
SuperFrost Plus slides. Slides were incubated at 60oC for >20 minutes in a hybridization

64
incubator. Slides were deparaffinized in three washes of xylene for 5 minutes each, rehydrated in
graded series of ethanol (two washes of 100% ethanol for 3 minutes each, two washes of 95%
ethanol for 1 minute each, and one wash of 70% ethanol for 1 minute), and rinsed in running tap
water for 5 minutes. Slides were immersed in 0.01 M citrate buffer (anhydrous citric acid, pH
6.0; Fisher Scientific; Waltham, MA) containing 0.05% Tween-20 at 100oC for 10 minutes and
allowed to cool for 30 minutes. Slides were washed twice in their respective wash solutions, with
20 dips per wash. The wash solution for MMP2, F4/80, α-SMA, and ERα was phosphate-
buffered saline (PBS, pH 7.6; 3.8 mM NaH2PO4 (Fisher Scientific; Waltham, MA), 16 mM
Na2HPO4 (Fisher Scientific; Waltham, MA), and 0.15 M NaCl (Fisher Scientific; Waltham,
MA)) and the wash solution for MMP14 was Tris-buffered saline with Tween-20 (TBST, pH
7.6; 20 mM Tris base (Fisher Scientific; Waltham, MA), 137 mM NaCl, 0.1% Tween-20 (Fisher
Scientific; Waltham, MA)). Endogenous peroxidase activity was blocked by immersing slides in
PBS containing 3% H2O2 (Fisher Scientific; Waltham, MA) for 10 minutes and rinsing in
running tap water for 5 minutes. Non-specific binding sites were blocked by 3% normal goat
serum (Vector Laboratories; Burlingame, CA) in PBS for one hour at RT in a humidified
incubator tray (Ciba Corning Diagnostics; Medfield, MA). Sections were then incubated
overnight at 4oC in a humidified incubator tray with rabbit polyclonal anti-MMP2 (3.33 µg/mL
(1:300); NB200-193, Novus Biologicals; Littleton, CO), rabbit monoclonal anti-MMP14 (0.5
µg/mL (1:250); ab51074, Abcam; Cambridge, MA), rat monoclonal anti-F4/80 (C1:A3-1; 4
µg/mL (1:250); ab6640, Abcam; Cambridge, MA), rabbit polyclonal anti-α-SMA (1 µg/mL
(1:200); ab5694, Abcam; Cambridge, MA), or rabbit polyclonal anti-ERα (MC-20; 1 µg/mL
(1:200); sc-542, Santa Cruz Biotechnology; Dallas, TX) Additional information regarding these
antibodies can be found in Table 3. Coverslips were removed in PBS and slides were washed

65
three times in wash solution for 5 minutes each. Sections incubated in biotinylated goat anti-rat
(mouse adsorbed) (10 µg/mL (1:50); Vector Laboratories; Burlingame, CA) or biotinylated goat
anti-rabbit (30 µg/mL (1:50); Vector Laboratories; Burlingame, CA) diluted in 3% normal goat
serum in PBS for 1 hour at RT in humidified incubator trays. Additional information regarding
these antibodies can be found in Table 3. Slides were washed in wash solution three times for 5
minutes per wash. The avidin-biotin complex (ABC; Vector Laboratories; Burlingame, CA) was
prepared 30 minutes prior to use by adding 2 drops of avidin DH (reagent A) and two drops of
biotinylated horseradish peroxidase H (reagent B) to 5 mL of PBS. Sections were incubated in
ABC solution for 1 hour at RT in humidified incubator trays. Slides were washed in wash
solution three times for 5 minutes per wash. Sections were visualized using 0.05% 3’3-
diaminobenzidine (DAB; Vector Laboratories; Burlingame, CA). DAB was prepared
immediately before use by adding 2 drops of buffer (pH 7.5), 4 drops of DAB, and 2 drops of
H2O2 to 5 mL of dH2O. Visualization was stopped by placing the slides in PBS. After all of the
sections developed, slides were washed in tap water three times for 5 minutes per wash. Slides
were rinsed in dH2O for 15 dips, counterstained in Hematoxylin 2 for 30 seconds, and rinsed in
running tap water. Slides were placed in Clarifier 2 for 30 seconds and rinsed in running tap
water for 30 seconds. Slides were placed in Bluing Reagent for 1 minute and rinsed in running
tap water for 1 minute. Slides were dehydrated through one wash of 95% ethanol for 15 dips and
three washes of 100% ethanol for 15 dips each. Slides were placed in three washes of xylene for
15 dips each and coverslipped using Permount. Mouse spleen was used as positive control tissue
for F4/80 staining, mouse lung was used as positive control tissue for MMP2 and MMP14
staining, mouse heart was used as positive control tissue for α-SMA staining, and rat mammary
tumor was used as positive control tissue for ERα staining.

66
The number of F4/80-positive cells in the uterus was quantified. Five random images at
40x magnification of myometrium and five random images at 40x magnification of endometrium
in which mostly stroma was visible were collected. For each image of endometrium and
myometrium, the total number of positively stained cells and the total number of stromal or
smooth muscle cells were counted using Photoshop CC and ImagePro Plus v4.5 (Media
Cybernetics; Rockville, MD). The number of positively stained cells was then divided by the
total number of cells and multiplied by 100 to generate the percent of F4/80-positive cells in each
mouse.
Table 3. Primary and Secondary Antibodies
Antigen Source Catalog # Conc IHC
(µg/mL)
Dilution IHC
Conc WB
(µg/mL)
Dilution WB
Species/Type Ref for
Specificity
MMP2 Novus NB200-193 3.33 1:300 0.25 1:4000 Rabbit
polyclonal
MMP14 Abcam ab51074 0.5 1:250 0.066 1:2000 Rabbit
monoclonal
Farahani et al.
(2012)
β-actin Cell
Signaling 8457 0.02 1:2000
Rabbit monoclonal
F4/80 Abcam ab6640 4 1:250 Rat
monoclonal
α-SMA Abcam ab5694 1 1:200 Rabbit
polyclonal
Farahani et al.
(2012)
ERα Santa Cruz
sc-542 1 1:200 Rabbit
polyclonal
Garcia-Falgueras
et al. (2011)
goat
anti-rabbit Vector BA-1000 30 1:50 Goat IgG
goat anti-rat
Vector BA-9401 10 1:50 Goat IgG
2.3.3 Quantitative RT-PCR
Frozen tissue of uterine horns ≤30 mg from CD1 mice exposed to 0 ppm and 30 ppm
BPA were homogenized for RNA isolation using an RNeasy Fibrous Tissue Mini Kit (Qiagen;

67
Valencia, CA). Tissue was homogenized in 300 µL of Buffer RLT (RNA lysis buffer containing
guanidine thiocyanate). To the homogenized tissue, 590 µL of RNase-free water (Teknova;
Hollister, CA) and 10 µL of proteinase K were added, mixed, and incubated at 55oC for 10
minutes. Samples were then centrifuged at 10,000 x g for 3 minutes. The supernatant was
transferred to a new tube, 450 µL of 100% ethanol was added, and the tube was mixed. Of the
total amount of supernatant, 700 µL was transferred to an RNeasy Mini column and the column
was centrifuged for 15 seconds at 8,000 x g. The flow through was discarded and the step was
repeated until all of the supernatant was centrifuged. Next, 350 µL of Buffer RW1 (RNA wash
buffer 1 containing ethanol) was added to the RNeasy column, centrifuged for 15 seconds at
8,000 x g, and the flow through was discarded. In the column, 10 µL of DNase stock solution
was mixed with 70 µL of Buffer RDD (DNase wash buffer) and incubated for 15 minutes at RT.
After the incubation, 350 µL of Buffer RW1 was added to the column and centrifuged for 15
seconds at 8,000 x g. The flow through was discarded and 500 µL of Buffer RPE (wash buffer)
was added to the column. The column was centrifuged for 15 seconds at 8,000 x g and the flow
through was discarded. A second 500 µL of Buffer RPE was added to the column and
centrifuged for 2 minutes at 8,000 x g. The column was placed in a new 1.5 mL microcentrifuge
tube and 30-50 µL of RNase-free water was added to the column. The column was centrifuged
for 1 minute at 8,000 x g and repeated with the RNA eluate. RNA concentrations and potential
contamination were determined using a Nanodrop 2000 spectrophotometer (Thermo Scientific;
Waltham, MA). RNase-free water was used as a standard. Concentrations of RNA are given as
ng/µL. The 260/280 ratio can indicate protein contamination and should be around 2.0
(ThermoScientific 2011). The 260/230 ratio indicates salt, carbohydrate, phenol, EDTA, or
guanidine HCl contamination and should also be around 2.0-2.2 (ThermoScientific 2011). RNA
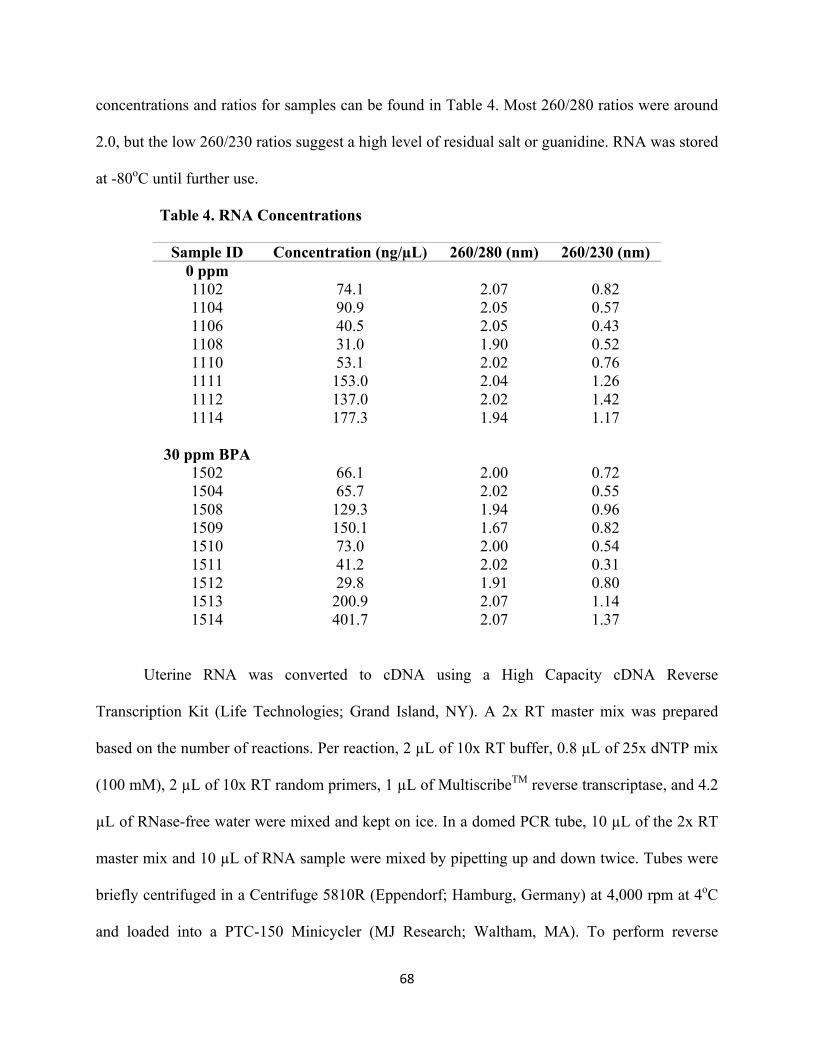
68
concentrations and ratios for samples can be found in Table 4. Most 260/280 ratios were around
2.0, but the low 260/230 ratios suggest a high level of residual salt or guanidine. RNA was stored
at -80oC until further use.
Table 4. RNA Concentrations
Sample ID Concentration (ng/µL) 260/280 (nm) 260/230 (nm) 0 ppm 1102 74.1 2.07 0.82 1104 90.9 2.05 0.57 1106 40.5 2.05 0.43 1108 31.0 1.90 0.52 1110 53.1 2.02 0.76 1111 153.0 2.04 1.26 1112 137.0 2.02 1.42 1114 177.3 1.94 1.17
30 ppm BPA
1502 66.1 2.00 0.72 1504 65.7 2.02 0.55 1508 129.3 1.94 0.96 1509 150.1 1.67 0.82 1510 73.0 2.00 0.54 1511 41.2 2.02 0.31 1512 29.8 1.91 0.80 1513 200.9 2.07 1.14 1514 401.7 2.07 1.37
Uterine RNA was converted to cDNA using a High Capacity cDNA Reverse
Transcription Kit (Life Technologies; Grand Island, NY). A 2x RT master mix was prepared
based on the number of reactions. Per reaction, 2 µL of 10x RT buffer, 0.8 µL of 25x dNTP mix
(100 mM), 2 µL of 10x RT random primers, 1 µL of MultiscribeTM reverse transcriptase, and 4.2
µL of RNase-free water were mixed and kept on ice. In a domed PCR tube, 10 µL of the 2x RT
master mix and 10 µL of RNA sample were mixed by pipetting up and down twice. Tubes were
briefly centrifuged in a Centrifuge 5810R (Eppendorf; Hamburg, Germany) at 4,000 rpm at 4oC
and loaded into a PTC-150 Minicycler (MJ Research; Waltham, MA). To perform reverse

69
transcription, the following conditions were used: 10 minutes at 25oC, 120 minutes at 37oC, 5
minutes at 85oC, and ≤24 hours at 4oC. The resulting cDNA was stored at -20oC until further use.
Predesigned TaqMan RT-PCR assays (Applied Biosystems; Grand Island, NY) were
used for the genes found in Table 5. PCR amplification was performed in a total volume of 20
µL containing 20x TaqMan expression assay primers (1 µL/reaction), TaqMan Universal Master
Mix (10 µL/reaction; Applied Biosystems; Grand Island, NY), RNase-free water (8 µL/reaction),
and 10 ng of cDNA (1 µL/reaction). Amplification was performed in triplicates on a Step One
Plus Real-Time PCR System (Applied Biosystems; Grand Island, NY) under the following fast
conditions: 20 seconds at 95oC for one holding stage, then 1 second at 95oC and 20 seconds at
60oC for the cycling stage (60 cycles). Relative expression was quantified using the ΔΔCt
method, using 18s (Mm03928990_g1) as the endogenous control for normalization.
Table 5. TaqMan Probe Sets Used for qRT-PCR Analysis
Gene Name Gene
Symbol ABI Assay
Entrez Gene
ID Unigene # NCBI Location Chromosome
collagen, type I, alpha 1
Col1a1 Mm00801632_gH 12842 Mm.277735 Chr.11: 94936270 - 94951867
collagen, type III, alpha 1
Col3a1 Mm00802296_g1 12825 Mm.249555 Chr.1: 45311538 - 45349706
matrix metallopeptidase 2
Mmp2 Mm00439498_m1 17390 Mm.29564 Chr.8: 92827292-92853420
tissue inhibitor of metalloproteinase 1
Timp1 Mm00441818_m1 21857 Mm.8245 Chr.X: 20870166 - 20874737
tissue inhibitor of metalloproteinase 2
Timp2 Mm00441825_m1 21858 Mm.206505 Chr.11: 118301061 - 118355411
tissue inhibitor of metalloproteinase 3
Timp3 Mm00441826_m1 21859 Mm.4871 Chr.10: 86300412 - 86349505
18s ribosomal RNA 18s Mm03928990_g1 19791

70
2.3.4 SDS-PAGE and Immunoblotting
Frozen uterine tissue was homogenized in lysis buffer (20 mM Tris-HCl (Fisher
Scientific; Waltham, MA), 150 mM NaCl, 1 mM EDTA (Fisher Scientific; Waltham, MA), 1
mM EGTA (Fisher Scientific; Waltham, MA), 1% Triton X-100 (Bio-Rad; Hercules, CA),
cOmplete Protease Inhibitor Cocktail (EDTA-free; Roche; Basel, Switzerland), pH 7.5) and
centrifuged at 4,000 rpm for 10 minutes at 4oC on a Centrifuge 5810R (Eppendorf; Hamburg,
Germany). The supernatant was collected and stored at -20oC until further use.
A colorimetric DC protein assay (Bio-Rad; Hercules, CA) was used in order to quantify
the protein concentration of the uterine homogenates. A standard protein curve using bovine
serum albumin (Frac V) (BSA; Fisher Scientific; Waltham, MA) was generated by diluting 50
mg of BSA in 1 mL of lysis buffer. Serial dilutions were made for 1.5 mg/mL, 1.0 mg/mL, 0.5
mg/mL, and 0.25 mg/mL from the 50 mg/mL solution. Lysis buffer was used for 0.0 mg/mL. A
working reagent was prepared by combining 20 µL of reagent S with each mL of reagent A
needed to make reagent A’. Reagent A’ is needed to solubilize detergent in the lysis buffer. If
necessary, uterine lysates were diluted in lysis buffer. In a 1.5 mL microcentrifuge tube, 20 µL of
standard or lysate was combined with 100 µL of reagent A’ and 800 µL of reagent B. After a 15
minute incubation at RT, 230 µL of the reaction was transferred to a flat bottom 96-well plate
(Denville Scientific; South Plainfield, NJ) in triplicate. Absorbance at 750 nm was determined
using a SpectraFluor Plus spectrofluorometer with Magellan 3 software (Tecan; Männedorf,
Switzerland). Absorbance readings were converted to protein concentrations in Microsoft Excel
(Microsoft; Redmond, WA) by first averaging the absorbance readings for the 0 mg/mL standard
and subtracting this value from all readings. A standard curve was generated by averaging the

71
values for each protein standard and linear regression analysis was used to calculate protein
concentrations for the uterine lysates.
Once protein concentrations were calculated, total protein (30 µg) was mixed 3:1 with
sample buffer (0.25 M Tris-HCl, 40% glycerol (Fisher Scientific; Waltham, MA), 8% SDS (Bio-
Rad; Hercules, CA), 0.01% bromophenol blue (Fisher Scientific; Waltham, MA), pH 6.8)
containing 0.04% β-mercaptoethanol (Fisher Scientific; Waltham, MA) and incubated at 96oC
for 5 minutes. Prestained SDS-PAGE protein standards of known molecular weights (Broad
Range; Bio-Rad; Hercules, CA) were used to determine relative molecular weights based on
mobility. Standards and protein lysates were loaded onto and resolved by electrophoresis on a
10% polyacrylamide gel (10% acrylamide (Fisher Scientific; Waltham, MA), 0.27%
bisacrylamide (Bio-Rad; Hercules, CA), 375 mM Tris base, 0.1% SDS, 0.025% ammonium
persulfate (APS; Bio-Rad; Hercules, CA), and 0.067% tetramethylethylenediamine (TEMED;
Bio-Rad; Hercules, CA), pH 8.8) with a 4% stacking gel (3.9% acrylamide, 0.1% bisacrylamide,
125 mM Tris base, 0.1% SDS, 0.0375% APS, and 0.1% TEMED, pH 6.8).
After electrophoresis, gels and Odyssey® nitrocellulose membranes (Li-Cor Biosciences;
Lincoln, NE) were placed in transfer buffer (48 mM Tris base, 39 mM glycine (Fisher Scientific;
Waltham, MA), 0.0375% SDS, 20% methanol (Fisher Scientific; Waltham, MA)) for 30 minutes
at 4oC. A wet transfer was then set up in the following order: black side of cassette, sponge, filter
paper, gel, membrane, filter paper, sponge, and white side of cassette. In between each layer, a
glass rod was used to expel of any bubbles. The black side of the cassette was placed facing the
black side of the Mini Trans-Blot Cell (Bio-Rad; Hercules, CA) for wet transfer. Transfer
occurred for 16 hours at 4oC at a constant amperage of 70 mA. Membranes were stained with
0.1% Ponceau S (Fisher Scientific; Waltham, MA) in 1% acetic acid (Acros Organics; Waltham,

72
MA) to ensure protein transfer. Membranes were blocked with 5% non-fat dry milk (Blotting
Grade Blocker, Bio-Rad; Hercules, CA) in TBST for 1 hour at RT and then incubated overnight
at 4oC with either rabbit polyclonal anti-MMP2 (0.25 µg/mL (1:4000); NB200-193, Novus
Biologicals; Littleton, CO) or rabbit monoclonal anti-MMP14 (0.066 µg/mL (1:2000); ab51074,
Abcam; Cambridge, MA). A rabbit monoclonal anti-β-actin antibody (0.02 µg/mL (1:2000);
8457, Cell Signaling; Danvers, MA) was used as a loading control. Additional information
regarding these antibodies can be found in Table 3. Membranes were washed three times in
TBST for 5 minutes per wash and incubated with IRDye® 800CW goat anti-rabbit IgG (1:5000;
Li-Cor Biosciences; Lincoln, NE) for 1 hour at RT. Membranes were washed in TBST three
times for 5 minutes per wash and placed in TBS. Membranes were imaged and analyzed using an
Odyssey® CLx Infrared Imaging System (Li-Cor Biosciences; Lincoln, NE) and Image Studio
Software (Li-Cor Biosciences; Lincoln, NE).
2.3.5 Gelatin Zymography
Zymography is a molecular technique that uses electrophoretic methodologies to quantify
the activity of certain enzymes by adding a substrate to the gel. In this case, activity of MMP2
was quantified by using gelatin as a substrate. Methods for gelatin zymography were adapted
from previously described protocols (Toth et al. 2012, Belcher et al. 2005).
Frozen uterine tissue was homogenized in lysis buffer (25 mM Tris-HCl, 0.1 M NaCl,
1% Nonidet P-40 (Shell Chemicals; The Hague, Netherlands), cOmpleteTM Protease Inhibitor
Cocktail (EDTA-free), pH 7.5; Roche; Basel, Switzerland) and centrifuged at 4,000 rpm for 10
minutes at 4oC on a Centrifuge 5810R. The supernatant was collected and stored at -20oC until
further use.

73
Protein concentrations of the uterine lysates were calculated using the colorimetric DC
protein assay, similar to the procedure used to quantify protein concentration for Western
blotting. The BSA standards were generated and lysates were diluted using lysis buffer specific
for zymography.
Once protein concentrations were calculated, 30 µg of total protein was mixed 3:1 with
sample buffer (0.25 M Tris-HCl, 40% glycerol, 8% SDS, 0.01% bromophenol blue, pH 6.8)
without the presence of a reducing agent. The lysate:sample buffer mixture was incubated at RT
for 15 minutes. Samples were loaded onto a 10% polyacrylamide gelatin-based zymography gel
(10% acrylamide, 0.27% bisacrylamide, 375 mM Tris base , 0.1% SDS, 0.025% APS, 0.067%
TEMED, 0.1% gelatin (Bio-Rad; Hercules, CA), pH 8.8) with a 4% stacking gel (3.9%
acrylamide, 0.1% bisacrylamide, 125 mM Tris base, 0.1% SDS, 0.0375% APS, and 0.1%
TEMED, pH 6.8). Following electrophoresis, zymograms were incubated in 2.5% Triton X-100
for 30 minutes at RT with gentle agitation in order to remove SDS from the samples and renature
the proteins. After renaturation, zymograms were rinsed in MilliQ H2O and incubated in
developing buffer (50 mM Tris-HCl, 0.2 M NaCl, 5 mM CaCl2 (Fisher Scientific; Waltham,
MA), pH 7.8) for 30 minutes at RT with gentle agitation. The developing buffer was decanted
and zymograms incubated in fresh developing buffer for 16 hours at 37oC in an isotemp
incubator (Fisher Scientific; Waltham, MA). Zymograms were stained with Coomassie Brilliant
Blue R-250 (Bio-Rad; Hercules, CA) dissolved in a 5% methanol/10% acetic acid solution for 3
hours at RT with gentle agitation. Zymograms were destained in a 5% methanol/10% acetic acid
solution for 15 minutes, 30 minutes, and 1 hour at RT with gentle agitation. Clear bands denoted
areas of proteolytic activity. Conditioned media from the human brain tumor cell line DAOY
(ATCC HTB-186) was used as a positive control for MMP activity (Belcher et al. 2009). In

74
control experiments, the addition of EDTA (1 mM final concentration) to lysates or developing
buffer resulted in loss of proteolytic activity at the expected molecular weights. Gels were
imaged using an Alpha Innotech Multiimage II Gel Documentation System (Alpha Innotech
Corp.; San Leandro, CA) and quantified using Image Studio Lite Software.
2.4 Statistical Methods
When applicable, the observer was blinded to strain, exposure group, and generation. The
litter was the statistical unit used; one animal was selected from each litter as representative of
that litter. Pathology was assessed with statistical differences analyzed using Fisher’s exact test
or χ2 test for trend when assessing across PND. Myometrial wall thickness, gland nest density,
and percentage of F4/80-positive cell counts were analyzed using a two-way ANOVA and
Bonferroni post hoc test in dams and a one-way ANOVA and Dunnett’s multiple comparison test
in offspring. A Student’s t-test was used for analysis of mRNA and protein expression and
protein activity. C57Bl/6N offspring endpoints were not statistically analyzed due to a low
sample size for all groups. Outliers were removed based on Grubb’s test. Significance between
differences in values was defined as p<0.05. All data was analyzed using GraphPad Prism®
v5/v6 software (GraphPad; La Jolla, California).

75
CHAPTER 3
Strain Specific Induction of Pyometra and Differences in Immune Responsiveness in Mice
Exposed to 17α-Ethinyl Estradiol or the Endocrine Disrupting Chemical Bisphenol A
Kendziorski JA, EL Kendig, RB Gear, and SM Belcher. 2012. Strain specific induction of pyometra and differences in immune responsiveness in mice exposed to 17α-ethinyl estradiol or the endocrine disrupting chemical bisphenol A. Reprod Toxicol. 34(1): 22-30. doi: 10.1016/j.reprotox.2012.03.001. PMID: 22429997

76
Strain Specific Induction of Pyometra and Differences in Immune Responsiveness in Mice Exposed to 17α-Ethinyl Estradiol or the Endocrine Disrupting Chemical Bisphenol A
Jessica A. Kendziorski, Eric L. Kendig, Robin L. Gear and Scott M. Belcher*
Department of Pharmacology and Cell Biophysics University of Cincinnati College of Medicine
Cincinnati, OH, 45267-0575
*Corresponding Author: Scott M. Belcher Department of Pharmacology and Cell Biophysics University of Cincinnati College of Medicine 231 Albert Sabin Way; PO Box 670575 Cincinnati, OH, 45267-0575 Telephone: 513-558-1721 Fax: 513-558-4329 Email: [email protected]

77
3.1 Abstract
Pyometra is an inflammatory disease of the uterus that can be caused by chronic exposure to
estrogens. It is unknown whether weakly estrogenic endocrine disruptors can cause pyometra.
We investigated whether dietary exposures to the estrogenic endocrine disruptor bisphenol A
(BPA) induced pyometra. Pyometra did not occur in CD1 mice exposed to different dietary
doses of BPA ranging from 4.1 to >4000 µg/kg/day or 17α-ethinyl estradiol (EE; 1.2 to >150
µg/kg/day). In the C57Bl/6 strain, pyometra occurred in the 15 µg/kg/day EE and 33 µg/kg/day
BPA treatment groups. At the effective concentration of BPA, histological analysis revealed
pathological alterations of uterine morphology associated with a >5.3-fold increase in
macrophage numbers in non-pyometra uteri of C57Bl/6 mice exposed to BPA. These results
suggest that BPA enhances immune responsiveness of the uterus and that heightened
responsiveness in C57Bl/6 females is related to increased susceptibility to pyometra.

78
3.2 Introduction
Inflammatory conditions of the uterus occur in specific strains of rats and mice receiving
extended exposures to estrogens (Burrows 1935). Pyometra is literally defined as “puss in the
uterus.” This potentially life-threatening infectious disease is characterized by inflammation and
intraluminal accumulation of purulent material in the uterus. Pyometra is clinically most well-
known to veterinary medical science, where it is a common occurrence in unspayed female dogs;
nearly 25% of bitches will be affected by pyometra before the age of 10 years (Smith 2006,
Egenvall et al. 2001). In addition to dogs, pyometra, while less common, is also a significant
disease in domestic cats and captive large felids, some livestock species, and laboratory animals
such as rabbits and some strains of mice and rats. In humans, pyometra is a relatively uncommon
condition with rare but significant mortality resulting from spontaneous uterine perforation or
rupture leading to septicemia. In the general population, it is estimated to account for about
0.04% of gynecological admissions; however, the incidence becomes increased to >13% in the
elderly (Yildizhan et al. 2006).
Hormonal factors, particularly the actions of estrogen on the uterus, play a central role in
the etiology of pyometra. Along with hormonal status and age, genetic factors related to cellular
infiltration of leukocytes into the uterus play a major role in regulating sensitivity to estrogen-
induced uterine inflammation and pyometra (Hagman et al. 2011, Gould et al. 2005, Roper et al.
1999). In sensitive strains of laboratory rats and mice, it is well established that chronic exposure
to estradiol or the highly efficacious non-steroidal estrogen, diethylstilbestrol (DES), can induce
pyometra (Gould et al. 2005, Pandey et al. 2005, McLachlan et al. 1980, Stone et al. 1979,
Gardner and Allen 1937, Zondek 1936, Burrows 1935). These findings suggest the possibility
that the highly prevalent environmental estrogenic endocrine disrupting chemical, bisphenol A

79
(BPA), may have a similar effect. Bisphenol A is a synthetic monomer used in the production of
epoxy resin-based sealants and polycarbonate plastics. Measurable levels of BPA have been
found in >90% of humans tested at levels that affect fecundity, embryonic development and
carcinogenesis in experimental models (Ye et al. 2011, Richter et al. 2007, Calafat et al. 2005) .
The ability of endocrine disrupting chemicals such as BPA, that are considered weak estrogens
based on potency in a uterotrophic bioassay or binding affinity at the nuclear estrogen receptors,
to cause pyometra is essentially unknown. However, BPA can cause morphological changes in
the CD1 mouse uterus at doses considered to be safe in humans, suggesting mechanisms other
than those assessed by the rodent uterotrophic bioassay are involved with some actions of BPA
in the uterus (Markey et al. 2001). As part of a larger effort to determine the physiological
consequences of dietary exposure to the endocrine disrupting chemical BPA, we investigated
whether BPA might act in an estrogen-like fashion to induce pyometra in mice exposed to BPA
or 17α-ethinyl estradiol (EE) as a positive control for estrogenic activity in their diet.

80
3.3 Materials and Methods
3.3.1 Animal husbandry and dietary exposures
All animal procedures were performed in accordance with protocols approved by the
University of Cincinnati Institutional Animal Care and Use Committee and followed
recommendations of the Panel on Euthanasia of the American Veterinary Medical Association.
Animals were maintained on a 14 h light, 10 h dark cycle and provided food and water ad
libitum in single-use polyethylene caging certified BPA-free (Innovive, San Diego California).
Sani-chip bedding (Irradiated Aspen Sani-chip; P.J. Murphy Forest Products Corp. Montville,
NJ) was used to avoid corncob-derived mycoestrogen exposure. Sterile drinking water was
generated by a dedicated water purification system (Millipore Rios 16 with ELIX UV/Progard 2)
that reduces organic contaminants to less than 1% of source levels. Drinking water was
dispensed from BPA-free polyethylene water bottles (Innovive, San Diego California).
A modified open standard diet (supplemental data table 1; Product #10010504 Research
Diets) was used to eliminate possible dietary phytoestrogen contamination. Test compounds
were directly incorporated into the extruded pellet diets that were unsupplemented (control) or
supplemented with 17α-ethinyl estradiol (EE; 1,3,5(10)-estratrien-17α-ethinyl-3,17β-diol, CAS
No. 57-63-6, Batch No. H923; Steraloids Inc., Newport, RI) or bisphenol A (2,2-bis(4-
hydroxyphenyl)propane; BPA; CAS 80-05-7; Lot 11909; USEPA/NIEHS standard) at desired
final concentrations (supplemental data table 1). For BPA, all three doses used were designed to
fall below the NOAEL (50 mg/kg/da). The high dose (30 ppm) was between the NOAEL and the
reference dose (50 μg/kg/da), the middle dose (0.3 ppm) approximated the reference dose, and
the low dose (0.03 ppm) fell below the reference dose (1982). Diets containing EE were

81
estimated to be at or below an effective dose previously reported to decrease fecundity in mice
(1.3 ppm, 0.1 ppm and 0.01 ppm) (Thayer et al. 2001) .
Seventy (35 of each sex) C57Bl/6NHsd and seventy Hsd:ICR (CD1 Swiss) mice aged six
to seven weeks were received from Harlan Laboratories, Inc. (Indianapolis, IN). Males and
females from each strain were group housed five per cage with the same sex and evenly divided
among control or one of the six dietary treatments and maintained on those treatments from
arrival until necropsy. Mice were allowed to acclimate to housing and treatment for two weeks.
Body weight and food consumption were monitored weekly, while general health and vaginal
cytology were monitored daily. Following the acclimation period, males and females in the same
group were randomly paired in individual housing and observed daily within the first 3 hr of the
light cycle for copulation plug. Upon observation of a copulation plug, assessment of vaginal
cytology ceased, the male was removed, and females were individually housed and observed
twice daily. Dams were supplied with nestlets, monitored for general health, and observed for
parturition. Body weights of both males and females were monitored weekly until necropsy.
Dams and their offspring were housed together until postnatal day 21, at which time the
offspring were separated and dams were group housed with F0 females from the same treatment
group.
3.3.2 Monitoring of estrous
Progression of estrous cycles was monitored in F0 females by morphological analysis of
daily vaginal lavage prior to copulation. Briefly, approximately 200 µl of sterile NaCl solution
(0.09%) was used to flush the vulva using a fire polished pipette. Stage of the estrous cycle was
determined by microscopic assessment of the vaginal cell morphology in each lavage sample.
Samples with epithelial cells in various stages of nuclear degeneration and cytoplasmic shrinkage

82
and polymorphonuclear leukocytes indicated diestrus; samples with nucleated endothelial cells
were indicative of proestrus; and samples with cornified, the presence of non-nucleated cellular
debris was characteristic of estrus.
3.3.3 Necropsy and tissue preparation
Female F0 mice were euthanized by CO2 asphyxiation and complete necropsy was
performed between the ages of 19 and 23 weeks. Digital images of animals, uteri, and ovaries
were captured as part of gross examination during necropsy (Nikon D3000 with 50 mM Nikkor
macro lens). Tissues were excised and cleaned of excess connective tissue and fat, blotted of
excess fluids, and weighed. Isolated tissues were fixed with several changes of 5%
paraformaldehyde or 10% neutral buffered formalin. Tissues were blocked by sectioning with a
sterile razor blade and washed several times in 70% ethanol prior to automated tissue processing
and embedding in paraffin (Histocenter 3; Thermo-Shandon Kalamazoo, MI). Microtome
sections were cut at 5 µm thickness from blocks at 4˚C and placed on positively charged slides
for hematoxylin and eosin (H&E) staining and immunohistochemistry. Standard H&E staining
was performed in order to examine tissue structure and morphology.
3.3.4 Immunohistochemistry
Paraffin sections were dewaxed and rehydrated through graded series of ethanol
concentrations into phosphate buffered saline (PBS, pH 7.4) and endogenous peroxidase blocked
by treatment with 3% H2O2 in PBS. After washing, heat-mediated epitope retrieval was
performed at 100oC in 10 mM citric acid with 0.05% Tween-20 (pH 6.0). Sections were
incubated at room temperature for >30 min with 3% normal goat serum in PBS, washed, and
then incubated overnight at 4˚C with 4 µg/mL of a macrophage specific F4/80 rat monoclonal
antibody (Cl:A3-1; Abcam Cambridge, Mass). Immunoreactivity was visualized with 0.05%

83
3’3-diaminobenzidine by the avidin-biotin peroxidase complex method (Vector Laboratories).
Stained sections were lightly counterstained in hematoxylin for 30 seconds (Richard Allan;
Kalamazoo, MI) and mounted in Permount (Fisher Scientific, Fair Lawn NJ). Internal controls
for specificity of immunostaining included replacement of primary or secondary antibodies with
non-specific serum. Sections prepared from spleen served as positive controls. Microscopic
examination of immunostained material was carried out using a Nikon Eclipse 55i microscope
using a DS-Fi1 CCD camera controlled with Digital Sight software (Nikon Instruments, Inc.,
Melville, NY). For quantitative assessments of immunopositive cell numbers, five random 40x
fields of the myometrium and five random 40x fields of the endometrium were collected from a
single section for each animal. The number of immunopositive cells per field was manually
counted by an investigator blinded to treatment and strain. To account for possible region
specific differences in infiltration, the mean number of cells in each of five 40x fields for the
myometrium and the endometrium was determined for one section per individual. The sum of
those values was taken to be representative of the number of F4/80 positive cells for that
individual. Final photomicrograph graphics were generated and labeled using Adobe Photoshop
CS3 (Adobe Systems, Inc., San Jose, CA).
3.3.5 Statistical analysis
Analysis of weight and cell count data was performed using one-way ANOVA followed
by Tukey’s Multiple Comparison post-hoc test with Cramer’s correction or Dunnett’s multiple
comparison test using GraphPad Prism v5 software (GraphPad Prism®, GraphPad Software,
Inc., La Jolla, CA). Significance between differences in values was defined by a p-value of
<0.05.

84
3.4 Results
3.4.1 Diet consumption
The amount of chow consumed by each female animal was determined immediately after
mating. From this consumption data, estimations of caloric intake and oral dose of EE and BPA
were calculated (Table 1).
For C57Bl/6 animals, the presence of 0.1 and 1.3 ppm EE in diet resulted in a significant
decrease in food consumption and the intake of significantly fewer calories per day as compared
to control diet. While EE dose dependently decreased consumption and caloric intake in CD1
animals, the difference in values for any one treatment group were not different from control. No
differences in consumption or caloric intake were detected in additional studies comparing
control diet and diet supplemented with 0.001 and 0.0001 ppm EE (data not shown). The
presence of BPA was not associated with any change in consumption or caloric intake for either
strain.
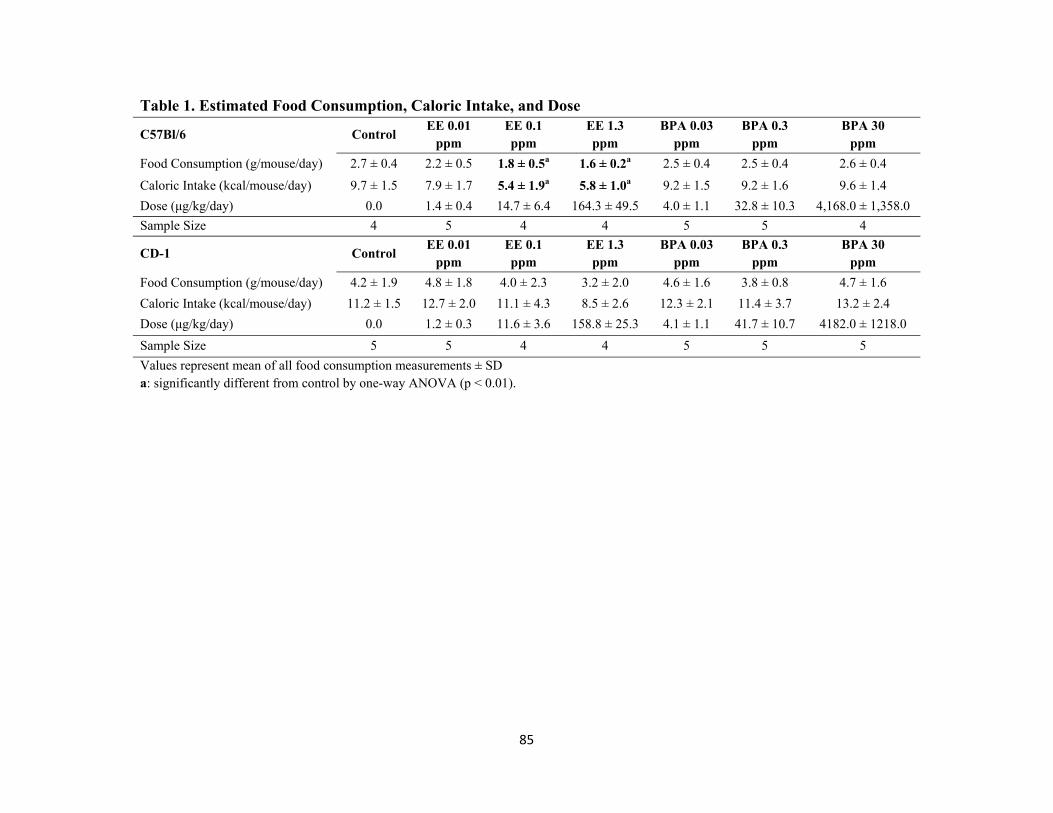
85
Table 1. Estimated Food Consumption, Caloric Intake, and Dose
C57Bl/6 Control EE 0.01
ppm EE 0.1 ppm
EE 1.3 ppm
BPA 0.03 ppm
BPA 0.3 ppm
BPA 30 ppm
Food Consumption (g/mouse/day) 2.7 ± 0.4 2.2 ± 0.5 1.8 ± 0.5a 1.6 ± 0.2a 2.5 ± 0.4 2.5 ± 0.4 2.6 ± 0.4
Caloric Intake (kcal/mouse/day) 9.7 ± 1.5 7.9 ± 1.7 5.4 ± 1.9a 5.8 ± 1.0a 9.2 ± 1.5 9.2 ± 1.6 9.6 ± 1.4
Dose (μg/kg/day) 0.0 1.4 ± 0.4 14.7 ± 6.4 164.3 ± 49.5 4.0 ± 1.1 32.8 ± 10.3 4,168.0 ± 1,358.0
Sample Size 4 5 4 4 5 5 4
CD-1 Control EE 0.01
ppm EE 0.1 ppm
EE 1.3 ppm
BPA 0.03 ppm
BPA 0.3 ppm
BPA 30 ppm
Food Consumption (g/mouse/day) 4.2 ± 1.9 4.8 ± 1.8 4.0 ± 2.3 3.2 ± 2.0 4.6 ± 1.6 3.8 ± 0.8 4.7 ± 1.6
Caloric Intake (kcal/mouse/day) 11.2 ± 1.5 12.7 ± 2.0 11.1 ± 4.3 8.5 ± 2.6 12.3 ± 2.1 11.4 ± 3.7 13.2 ± 2.4
Dose (μg/kg/day) 0.0 1.2 ± 0.3 11.6 ± 3.6 158.8 ± 25.3 4.1 ± 1.1 41.7 ± 10.7 4182.0 ± 1218.0
Sample Size 5 5 4 4 5 5 5
Values represent mean of all food consumption measurements ± SD a: significantly different from control by one-way ANOVA (p < 0.01).

86
3.4.2 Effects on body weight
Body weights were monitored from the onset of study until study completion when
animals were between 19 and 23 weeks of age. Not surprisingly, EE treatment dose dependently
impacted the body weight (Table 2). In the 1.3 ppm EE dose group, body weight was
significantly decreased at the end of the first study week. Lower concentrations of EE similarly
decreased body weight following an inverse correlation with dose (Table 2). There was also a
dose responsiveness observed in the onset of detectable differences in body weight of control and
EE dose groups. After the second week on treatment, both 0.1 and 1.3 ppm EE groups weighed
significantly less than controls. Those trends continued through the end of study. No difference
in either the amount or the rate of weight gained was observed in females of any of the BPA
groups (Table 1; Table 2). It thus appears that there is a dose dependent aversion to consuming
diet containing EE and that the sensitivity to EE diet was strain dependent.
After mating, body weight measurements for mated gravida and nulligravida females
were analyzed separately to avoid confounders on body weight related to metabolic
consequences of pregnancy, parturition, and lactation status. While impacts on body weight
related to treatment and dose remain evident during those later time points, the resulting
reduction in sample size prevented meaningful statistical analysis post copulation.

87
Table 2. Female Body Weight C57Bl/6
Wks Control EE 0.01 ppm EE 0.1 ppm EE 1.3 ppm BPA 0.03 ppm BPA 0.3 ppm BPA 30 ppm
0 14.4 ± 0.6 14.1 ± 0.4 14.4 ± 0.4 14.7 ± 0.6 14.3 ± 0.8 13.7 ± 0.9 14.2 ± 0.9
1 15.5 ± 0.6 15.4 ± 0.9 14.0 ± 0.4 13.1 ± 0.4a 16.1 ± 0.8 15.8 ± 0.6 16.4 ± 1.8
2 16.5 ± 0.6 15.7 ± 0.9 14.7 ± 0.9a 13.2 ± 1.0a 16.6 ± 0.7 16.1 ± 0.7 16.4 ± 0.6
3 17.5 ± 1.0 16.3 ± 1.3 14.6 ± 0.7a 13.3 ± 0.7a 17.8 ± 0.9 17.3 ± 0.4 17.2 ± 0.3
Status gravida nulli
gravida gravida
nulli gravida
gravida nulli
gravida gravida
nulli gravida
gravida nulli
gravida gravida
nulli gravida
gravida nulli
gravida
4 17.9 ± 0.9 17.1 ± 0.4 15.2 16.3 ± 1.1 NA 14.7 ± 0.8 NA 12.8 ± 0.9 18.2 ± 0.6 17.0 ± 0.5 17.7 ± 0.4 17.2 ± 0.6 17.6 ± 0.3 16.4
5 19.8 ± 1.1 19.0 ± 0.8 15.9 17.3 ± 1.5 NA 15.4 ± 0.4 NA 13.1 ± 1.5 19.9 ± 0.8 18.1 ± 1.2 19.7 ± 0.7 18.8 ± 0.0 19.7 ± 0.6 17.6
6 26.4 ± 1.7 20.4 ± 0.2 17.1 19.2 ± 0.6 NA 15.4 ± 1.1 NA 13.1 ± 0.6 25.5 ± 0.2 19.3 ± 0.1 26.6 ± 2.6 20.1 ± 0.1 26.7 ± 3.0 18.9
7 24.3 ± 0.8 21.2 ± 1.1 18.2 20.2 ± 2.0 NA 16.2 ± 0.6 NA 13.5 ± 1.3 32.3 ± 8.5 21.4 ± 0.2 27.8 ± 6.1 21.7 ± 1.2 26.3 ± 4.7 19.6
CD-1
Wks Control EE 0.01 ppm EE 0.1 ppm EE 1.3 ppm BPA 0.03 ppm BPA 0.3 ppm BPA 30 ppm
0 20.1 ± 1.4 20.8 ± 0.6 19.9 ± 0.7 20.2 ± 1.7 20.3 ± 0.9 19.6 ± 1.3 19.3 ± 1.4
1 22.5 ± 1.2 24.0 ± 1.2 20.8 ± 1.7 18.6 ± 1.4a 23.3 ± 1.0 23.4 ± 2.7 23.7 ± 1.6
2 24.6 ± 1.5 26.6 ± 1.8 21.8 ± 1.7 18.7 ± 1.3a 25.4 ± 2.0 26.4 ± 4.2 27.0 ± 3.4
3 23.6 ± 1.0 25.0 ± 1.6 21.3 ± 1.3 18.3 ± 1.1a 24.0 ± 1.8 25.5 ± 3.2 25.6 ± 2.3
Status gravida nulli
gravida gravida
nulli gravida
gravida nulli
gravida gravida
nulli gravida
gravida nulli
gravida gravida
nulli gravida
gravida nulli
gravida
4 26.0 ± 1.3 NA 27.9 ± 1.5 26.4 ± 2.0 NA 22.5 ± 1.2 NA 19.0 ± 1.4 25.2 ± 2.3 27.3 ± 2.6 27.4 ± 3.0 25.6 ± 2.8 26.8 ± 0.5 29.7
5 33.6 ± 2.4 NA 35.7 ± 6.2 31.2 ± 0.5 NA 23.0 ± 1.3 NA 19.3 ± 1.3 32.3 ± 5.1 29.0 ± 3.2 32.1 ± 1.3 26.5 ± 2.3 34.3 ± 1.9 29.6
6 53.4 ± 5.2 NA 33.7 ± 3.4 32.9 ± 3.5 NA 24.1 ± 2.4 NA 19.4 ± 1.4 47.6 ± 13.3 31.8 ± 4.2 45.6 ± 8.2 27.6 ± 3.2 33.7 ± 1.6 32.7
7 35.5 ± 2.7 NA 36.0 ± 0.8 32.2 NA 29.1 ± 1.9 NA 19.9 37.1 ± 1.6 34.8 ± 4.5 38.5 ± 1.0 38.5 ± 3.9 36.5 ± 2.8 37.9
Values represent group mean ± SD; a Significantly different from Control by one-way ANOVA (p < 0.05). Weeks 4-7 of treatment are separated by those F0 females that produced a live litter (Gravida) and those that did not produce a live litter (Nulligravida).

88
3.4.3 Fertility and fecundity
In both strains, diets containing 0.1 and 1.3 ppm EE resulted in no productive mating
(Table 3). This is considered likely to have resulted from insufficient nutritional intake, as is the
marked delay in mating and decreased percentage of mating that was also observed. In the
C57Bl/6 mice in the low dose EE group, a single productive mating occurred that resulted in a
small litter with 4 pups. In the low dose EE CD1 group, a modest reduction in fertility and
fecundity was also observed (Table 3). In contrast, no effect on fertility or fecundity in the
C57Bl/6 strain for any dose of BPA was observed (Table 3). In the CD1 strain 0.03 ppm (4
µg/kg/da) and 0.3 ppm (41 µg/kg/da) BPA treatment groups, an increase in latency of mating
and a decrease in productive mating was noted.

89
Table 3. Measures of Fertility and Fecundity C57Bl/6
Control EE 0.01 EE 0.1 EE 1.3 BPA 0.03 BPA 0.3 BPA 30 No. Breeding Pairs 5 5 5 5 5 5 5 No. litters born 3 1 0 0 3 3 4 Mating index (%) 100 100 20 100 100 100 100 Fertility index (%) 60 20 0 0 60 60 80 Precoital time (da) 2.0 ± 0.8 3.0 ± 0.0 3.0 ± 0.0 3.8 ± 3.0 2.2 ± 0.8 2.2 ± 1.5 1.8 ± 0.5 Gestation (da) 19.0 ± 0.0 N/A N/A N/A 19.0 ± 0.0 18.7 ± 0.6 19.0 ± 1.0 Pups per litter 7.7 ± 1.2 4.0 0.0 0 8.0 ± 0.0 7.0 ± 0.0 6.8 ± 3.2 % Males 50.0 ± 10.1 50 N/A N/A 50.0 ± 17.68 40.0 ± 18.7 37.2 ± 10.2
CD1 Control EE 0.01 EE 0.1 EE 1.3 BPA 0.03 BPA 0.3 BPA 30
No. Breeding Pairs 5 5 5 5 5 5 5 No. litters born 5 3 0 0 3 3 4 Mating index (%) 100 80 60 40 80 80 100 Fertility index (%) 100 75 0 0 75 75 80 Precoital time (da) 2.6 ± 0.5 2.3 ± 0.5 7.2 ± 3.3 7.3 ± 6.5 4.2 ± 3.3 4.4 ± 3.1 2.2 ± 0.8 Gestation (da) 18.6 ± 0.5 18.5 ± 0.7 N/A N/A 19.0 ± 0.0 19.0 ± 0.0 19.0 ± 0.8 Pups per litter 8.4 ± 3.5 10.0 ± 3.0 N/A N/A 10.0 ± 4.0 10.0 ± 1.7 8.0 ± 4.8 % Males 34.9 ± 12.7 44.0 ± 18.7 N/A N/A 50.5 ± 29.3 45.4 ± 30.0 31.7 ± 19.9 Values represent mean ± SD where relevant; Mating index (%) = No. copulation plug positive females / No. females paired X 100; Fertility index (%) = No. females with live litters / No. copulation plug positive females X 100; Precoital time = days from introduction of mating pairs to observation of copulation plug.

90
3.4.4 Necropsy and organ weights
Because of morbidity, the C57Bl/6 mice in the EE treatment groups were euthanized, and
necropsy was performed one to two weeks prior to necropsy of the control or BPA treated
groups. Similarly, one C57Bl/6 female was sacrificed prematurely (~wk 4) due to an undefined
neurological pathology; pyometra was not observed in that individual. Due to the small sample
size, variability of some endpoints, and pathology associated with differences in body weights,
statistical assessments of differences in these endpoints compared to controls are of limited value
in determining impact of treatments on gross organ weights. However, these values clearly
illustrate the dramatic strain specific differences observed in the uterine weights in response to
treatment with 0.1 ppm EE and 0.3 ppm BPA (Table 4). An average 23.5 fold increase in uterine
weight compared to controls was observed in the C57Bl/6 0.1 ppm EE treatment group. In
contrast, the uterine weights for the CD1 0.1 ppm and 1.3 ppm EE treated groups were not
different than the control groups. Spleen weights for the C57Bl/6 0.1 ppm EE treatment group
were significantly increased compared to controls, a result consistent with a substantial immune
response in females with affected uteri (p<0.05). With the exception of uterus and spleen weights
in each of the affected C57Bl/6 treatment groups, there were no additional changes in organ
weights.
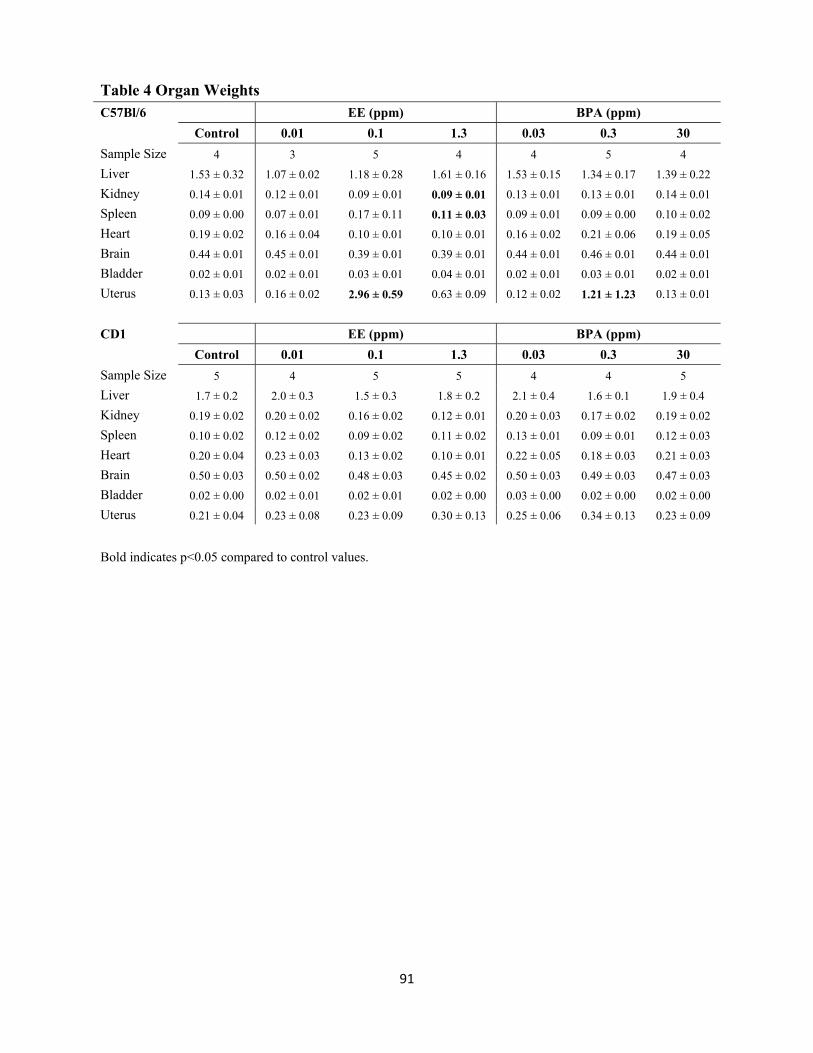
91
Table 4 Organ Weights C57Bl/6 EE (ppm) BPA (ppm)
Control 0.01 0.1 1.3 0.03 0.3 30
Sample Size 4 3 5 4 4 5 4
Liver 1.53 ± 0.32 1.07 ± 0.02 1.18 ± 0.28 1.61 ± 0.16 1.53 ± 0.15 1.34 ± 0.17 1.39 ± 0.22
Kidney 0.14 ± 0.01 0.12 ± 0.01 0.09 ± 0.01 0.09 ± 0.01 0.13 ± 0.01 0.13 ± 0.01 0.14 ± 0.01
Spleen 0.09 ± 0.00 0.07 ± 0.01 0.17 ± 0.11 0.11 ± 0.03 0.09 ± 0.01 0.09 ± 0.00 0.10 ± 0.02
Heart 0.19 ± 0.02 0.16 ± 0.04 0.10 ± 0.01 0.10 ± 0.01 0.16 ± 0.02 0.21 ± 0.06 0.19 ± 0.05
Brain 0.44 ± 0.01 0.45 ± 0.01 0.39 ± 0.01 0.39 ± 0.01 0.44 ± 0.01 0.46 ± 0.01 0.44 ± 0.01
Bladder 0.02 ± 0.01 0.02 ± 0.01 0.03 ± 0.01 0.04 ± 0.01 0.02 ± 0.01 0.03 ± 0.01 0.02 ± 0.01
Uterus 0.13 ± 0.03 0.16 ± 0.02 2.96 ± 0.59 0.63 ± 0.09 0.12 ± 0.02 1.21 ± 1.23 0.13 ± 0.01
CD1 EE (ppm) BPA (ppm)
Control 0.01 0.1 1.3 0.03 0.3 30
Sample Size 5 4 5 5 4 4 5
Liver 1.7 ± 0.2 2.0 ± 0.3 1.5 ± 0.3 1.8 ± 0.2 2.1 ± 0.4 1.6 ± 0.1 1.9 ± 0.4
Kidney 0.19 ± 0.02 0.20 ± 0.02 0.16 ± 0.02 0.12 ± 0.01 0.20 ± 0.03 0.17 ± 0.02 0.19 ± 0.02
Spleen 0.10 ± 0.02 0.12 ± 0.02 0.09 ± 0.02 0.11 ± 0.02 0.13 ± 0.01 0.09 ± 0.01 0.12 ± 0.03
Heart 0.20 ± 0.04 0.23 ± 0.03 0.13 ± 0.02 0.10 ± 0.01 0.22 ± 0.05 0.18 ± 0.03 0.21 ± 0.03
Brain 0.50 ± 0.03 0.50 ± 0.02 0.48 ± 0.03 0.45 ± 0.02 0.50 ± 0.03 0.49 ± 0.03 0.47 ± 0.03
Bladder 0.02 ± 0.00 0.02 ± 0.01 0.02 ± 0.01 0.02 ± 0.00 0.03 ± 0.00 0.02 ± 0.00 0.02 ± 0.00
Uterus 0.21 ± 0.04 0.23 ± 0.08 0.23 ± 0.09 0.30 ± 0.13 0.25 ± 0.06 0.34 ± 0.13 0.23 ± 0.09
Bold indicates p<0.05 compared to control values.

92
3.4.5 Uterine Pathology: C57Bl/6 pyometra
During the course of the study, it was observed that four of the C57Bl/6 females in the
0.1 ppm EE group and one in the 0.3 ppm BPA group (none of which produced a live litter)
developed a moribund phenotype characterized by lethargy and highly distended abdomens.
Upon necropsy, the uterus of each of the affected animals was found to be grossly enlarged and
filled with purulent fluid, indicative of severe pyometra (Fig. 1A). No pyometra was observed in
any CD1 mice from any treatment group, indicating an increased sensitivity of estrogen-induced
pyometra for C57Bl/6 mice.
Histological examination of the pyometic uteri revealed thin remnants of the
myometrium and endometrium. The remaining cells of the endometrium were in a disorganized
and ulcerated layer that was highly necrotized (Fig. 1B-G). Immunostaining for the macrophage
specific antigen F4/80 revealed a high degree of macrophage infiltration associated with the
outer layer of myometrium and localized to the inner border of the endometrial-like cell layer
remnant (Fig. 1C; 1E; 1G).
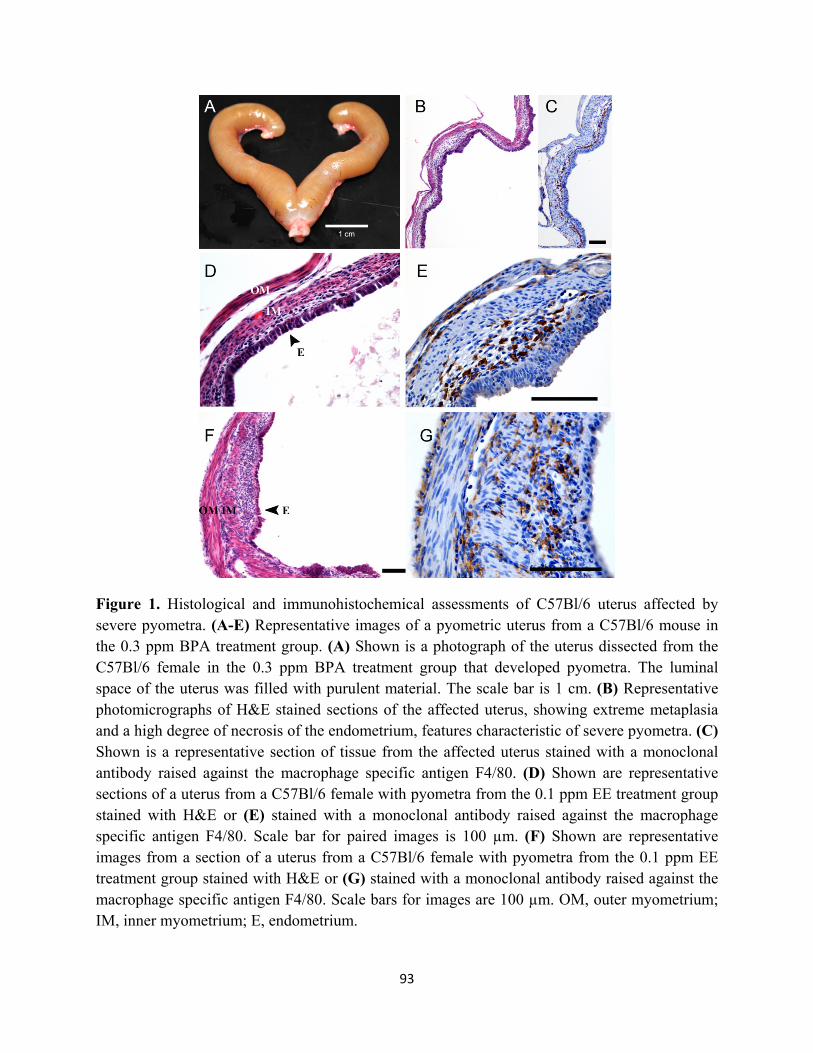
93
Figure 1. Histological and immunohistochemical assessments of C57Bl/6 uterus affected by severe pyometra. (A-E) Representative images of a pyometric uterus from a C57Bl/6 mouse in the 0.3 ppm BPA treatment group. (A) Shown is a photograph of the uterus dissected from the C57Bl/6 female in the 0.3 ppm BPA treatment group that developed pyometra. The luminal space of the uterus was filled with purulent material. The scale bar is 1 cm. (B) Representative photomicrographs of H&E stained sections of the affected uterus, showing extreme metaplasia and a high degree of necrosis of the endometrium, features characteristic of severe pyometra. (C) Shown is a representative section of tissue from the affected uterus stained with a monoclonal antibody raised against the macrophage specific antigen F4/80. (D) Shown are representative sections of a uterus from a C57Bl/6 female with pyometra from the 0.1 ppm EE treatment group stained with H&E or (E) stained with a monoclonal antibody raised against the macrophage specific antigen F4/80. Scale bar for paired images is 100 µm. (F) Shown are representative images from a section of a uterus from a C57Bl/6 female with pyometra from the 0.1 ppm EE treatment group stained with H&E or (G) stained with a monoclonal antibody raised against the macrophage specific antigen F4/80. Scale bars for images are 100 µm. OM, outer myometrium; IM, inner myometrium; E, endometrium.

94
3.4.6 Uterine Pathology: effects on uterine morphology and macrophage infiltration
To investigate whether exposures to EE or BPA were associated with uterine pathology
and increased macrophage infiltration, even in the absence of severe overt pyometra, histological
comparison of uteri from C57Bl/6 females in the control group with those from the 0.3 ppm BPA
group without pyometra was performed. In the BPA treatment group, the general structural
composition of the uterus including the myometrium, luminal and glandular epithelium, and
endometrial stroma was intact. However, disorganization in these uterine structures was evident
(compare Fig. 2A with Fig. 2B; 2C). The myometrium contained two distinctive smooth muscle
layers each of proper thickness, and the endometrium contained groups of glands surrounding the
luminal epithelium. However, hypertrophic luminal epithelial cells were evident. In some cases,
cells of the stroma were tightly packed and disorganized (Fig. 2C). Cystic endometrial
hyperplasia (CEH) was evident in the uterus of one of the C57Bl/6 female in the 0.3 ppm BPA
group (Fig. 2C1). Compared to control (Fig. 2A2), a marked increase in F4/80 positive
immunostaining was observed in all uteri of the 0.3 ppm BPA treatment group (Fig. 2 B2; C2).
Quantitative assessment of F4/80 immunopositive cell numbers revealed a statistically
significant 5.3-fold increase in the number of macrophages present in the uterus of C57Bl/6 mice
from the 0.3 ppm BPA treatment group (Fig. 4). There were no detectable differences in the
density of immunopositive cells localized to the endometrium or the myometrium in either
control or 0.3 ppm BPA groups. Comparison of the numbers of F4/80 immunopositive cells in
uteri from primagravida and nulliparous F0 C57Bl/6 females without pyometra, revealed no
differences within a given treatment group.

95
Figure 2. Effects of oral exposure to BPA on uterine morphology and macrophage infiltration in C57Bl/6 mice without overt pyometra. Shown are representative photomicrographs of uterine sections from a control female C57Bl/6 mouse (Panels A1-A4) and from two different animals in the 0.3 ppm BPA treatment group without symptomatic pyometra (Panels B1-B4; C1-C4). The uteri of BPA treated animals displayed metaplasia (C1, inset) throughout the various regions of the uterus with glandular disorganization and apparent luminal epithelial hyperplasia (C1, arrowhead). Cystic endometrial hyperplasia was also observed in one uterus (C1; denoted by *). Comparison of the density of F4/80 immunopositive cells present in sections from controls (Panels A2; A4) with those from the BPA treatment groups (Panels B2; B4 and C2; C4) demonstrates increased macrophage infiltration. Scale bars for each high magnification and low magnification photomicrograph is 100 µm.

96
To assess whether heightened immune responsiveness with BPA and EE exposure was
specific to the uteri from C57Bl/6 animals that developed pyometra, we also assessed uterine
morphology and the numbers of macrophages present in uterine tissue from CD1 mice (Fig. 3A-
H). In contrast to uteri from C57Bl/6 animals, the overall morphology of the uterus was
generally normal for the CD1 females in the 0.3 ppm BPA (Fig. 3B; E) and 0.1 ppm EE (Fig. 3F-
H) treatment groups. However, CEH was detected in one of four CD1 females in the 0.3 ppm
BPA group, and three of four uteri in the 0.1 ppm EE group. Compared to the C57Bl/6 control
group, increased numbers of F4/80 immunopositive cells were evident in uterine tissues from
CD1 controls (Fig. 3A; D; Fig 4). Similar increases in macrophages were apparent for the 0.3
ppm BPA (Fig 3B; E) and 0.1 ppm EE treated females (Fig. 3F-H). Quantitative comparison of
the numbers of macrophages present in the uteri showed 2.7-fold higher F4/80 positive cells in
CD1 controls compared to C57Bl/6 control females (Fig. 4). Compared to CD1 controls, a
significant increase in the number of F4/80 positive cells was not observed in the uteri of CD1
mice in the 0.3 ppm BPA treatment group (Fig. 4).

97
Figure 3. Effects of oral exposure to BPA on uterine morphology and macrophage infiltration in CD1 mice, which did not display pyometra. Shown are representative low and high magnification photomicrographs of uterine sections stained for F4/80 positive macrophages from female CD1 mouse in the control group (A; D), the 0.3 ppm BPA treatment group (B; E) or the 0.1 ppm EE treatment group (F-H). Control for specificity of immunostaining included replacement of primary antibodies with non-specific serum (C1). Immunostaining of sections prepared from spleen served as positive controls (C2). Scale bars represent 100 µm.
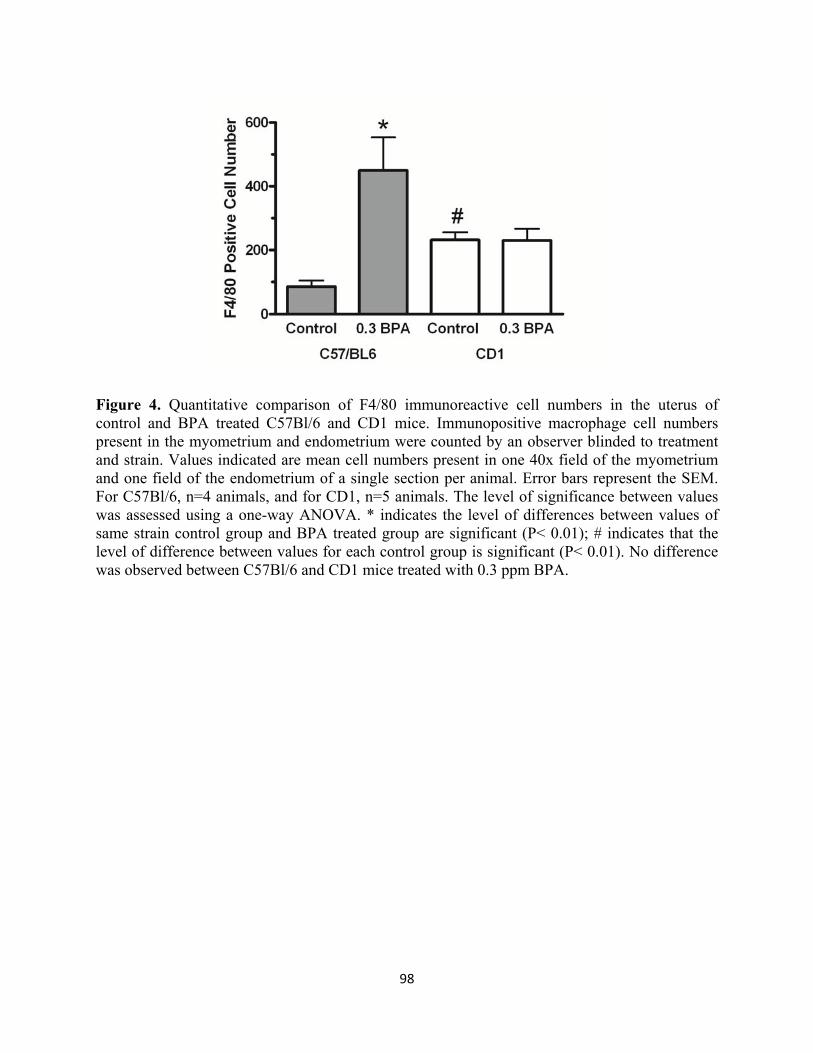
98
Figure 4. Quantitative comparison of F4/80 immunoreactive cell numbers in the uterus of control and BPA treated C57Bl/6 and CD1 mice. Immunopositive macrophage cell numbers present in the myometrium and endometrium were counted by an observer blinded to treatment and strain. Values indicated are mean cell numbers present in one 40x field of the myometrium and one field of the endometrium of a single section per animal. Error bars represent the SEM. For C57Bl/6, n=4 animals, and for CD1, n=5 animals. The level of significance between values was assessed using a one-way ANOVA. * indicates the level of differences between values of same strain control group and BPA treated group are significant (P< 0.01); # indicates that the level of difference between values for each control group is significant (P< 0.01). No difference was observed between C57Bl/6 and CD1 mice treated with 0.3 ppm BPA.

99
3.5 Discussion
It should first be pointed out that the intent of this study was to compare appropriate
concentrations of dietary estrogenic chemicals that would allow reproduction and not to
determine whether or not BPA had specific effects upon reproductive or other endpoints. As a
result, the observed changes in those endpoints (e.g. fertility and fecundity, organ weights, etc.)
must not be over interpreted. However, in the case where food consumption is markedly altered
and reproduction was completely blocked, some interesting and valuable insight to the impact of
these dietary estrogens is possible. A clear and significant differential sensitivity to estrogens
between the CD1 and C57Bl/6 strains was evident. This difference in sensitivity was dose
dependent and extended to an apparent aversion for consuming EE. That effect was dramatic and
statistically significant for two related, though independent, endpoint assessments (body weight
and food consumption), even in a study with limited statistical power. Additionally, the
development of pyometra, a well characterized uterine pathology related to estrogen exposure,
was found to occur in the C57Bl/6 strain but not in the CD1 strain. The relatively increased
sensitivity of the C57Bl/6 strain to impacts of estrogens on endpoints as varied as decreased food
consumption and a uterine immunopathology could be interpreted to suggest that the differential
sensitivity of these strains to estrogens is related to a common fundamental aspect of estrogen
signaling rather than isolated tissue-specific differences between strains. However, the apparent
trend toward decreased fecundity observed in the BPA exposed CD1 mice suggests there could
be increased reproductive sensitivities to some actions of BPA. The phenomenon of strain
differences in endpoint- and tissue-specific responses to estrogens in vivo has been extensively
supported in the EDC literature (Pepling et al. 2010, Brossia et al. 2009, Long et al. 2000,
Spearow 1999, Morozova 1991, Greenman et al. 1977). The effects observed here suggest about

100
a 10-fold increase in estrogen sensitivity of the C57Bl/6 mice and support the notion of
differential sensitivity of these experimental animal models. However, statements regarding
strain specific estrogen sensitivity should take into account the specific endpoints analyzed.
In contrast to the relatively well-documented cases in veterinary medicine, estrogen-
induced pyometra has received relatively little attention from researchers investigating the
actions of endogenous estrogens and estrogenic endocrine disrupting chemicals. The first
reported observation of pyometra occurring in response to long periods of estrogen exposure in
laboratory rodents was made by Burrows (Burrows 1935). A similar study indicated that some
strains of rat remained resistant to follicular hormone-induced pyometra; a phenomenon not seen
in other species tested (Zondek 1936). Later studies demonstrated that estrogenic hormones
altered bacterial content of the murine uterus, suggesting that there could be a link between the
ability of estrogens to modify the structure of the uterus and the observed changes in the uterine
microbiome (Weinstein et al. 1943). Studies investigating the actions of the non-steroidal
estrogen, DES, clearly demonstrated strain specific differences in the susceptibility to developing
pyometra. Those strain-related differences may result from direct, estrogen receptor-mediated,
activation of the innate immune response (Calippe et al. 2010, Calippe et al. 2008, Hendry et al.
1997, McLachlan et al. 1980). During long-term DES exposure, Sprague-Dawley rats, but not
ACI rats, developed DES-induced pyometra (Stone et al. 1979). Subsequently, the Brown
Norway rat was identified as sensitive to developing pyometra, which allowed demonstration,
via quantitative trait loci mapping, that the strain specific differences in susceptibility to
developing estrogen-induced pyometra were, in part, genetically controlled (Gould et al. 2005).
The results of this study clearly demonstrate strain differences in susceptibility as dietary
exposure to approximately 15 µg/kg/day EE results in essentially quantitative induction of

101
pyometra in C57Bl/6 mice, while the CD1 mice did not develop pyometra at that dose or even at
a 10-fold greater dose. Further, one of the five female mice that consumed diet containing 0.3
ppm BPA, resulting in exposure of about 33 µg/kg/day, developed pyometra. Again, no
pyometra was observed in any CD1 animals or in C57Bl/6 mice maintained on control diet
lacking estrogens. The development of pyometra in a BPA-treated animal is important in that it
is the first report of a weakly-estrogenic xenoestrogen inducing pyometra; this demonstrates that,
in regard to its pathological actions on the uterus, BPA is presumably acting as an estrogen
mimetic. This may be of concern in humans (especially sensitive subpopulations) given that
>90% of Americans tested have measurable levels of BPA in their urine (Ye et al. 2011, Calafat
et al. 2005). It is also notable that the dose of BPA that induced pyometra in this study was
below the dose considered safe for human exposure (50 μg/kg/day), which was based on a
NOAEL of 50 mg/kg/day from rodent dietary studies with a BPA concentration of 1000 ppm
(1982).
To assess the role of the immune response in the differential strain susceptibility to
develop pyometra with exposure to estrogens, we investigated the degree of uterine macrophage
infiltration in each strain. Regardless of treatment, we found high levels of macrophages in the
remnants of the necrotic myometrium and the endometrial-like cells of the pyometric uteri.
Based on those findings we hypothesized that BPA and EE were increasing the basal
inflammation in the uteri and reasoned that BPA-exposed C57Bl/6 mice without overt pyometra
would have an increased immune response, as indicated by increased presence of F4/80 positive
macrophages. Comparison of the numbers of macrophages present in the uteri of control CD1
and C57Bl/6 mice revealed a higher number of F4/80-staining macrophages in CD1 samples,
suggesting that there was an elevated basal level compared to C57Bl/6. However, there was a

102
significant increase in macrophage counts in C57Bl/6 mice treated with 0.3 ppm BPA compared
to control with no increase seen in 0.3 ppm BPA treated CD1 mice. Along with increased
numbers of macrophages in the mice that did not develop pyometra, significant structural
differences were observed in the C57Bl/6, but not the CD1 strain. The C57Bl/6 mice exposed to
0.3 ppm BPA displayed stromal and glandular metaplasia, with no structural differences
observed in the corresponding CD1 treatment group. Those differences in uterine phenotype
further support the differential sensitivity to estrogens between the C57Bl/6 and CD1. Thus, it
seems likely that these immunologic and structural differences between the strains indicate a
potential key difference in susceptibility to developing overt pyometra which is related to the
immune response. While the basal (control) number of macrophages in the CD1 uterus was
significantly higher than that in the C57Bl/6 mice, the increase with BPA exposure was very
robust in the C57Bl/6 mice. It would appear that the magnitude of the uterine immune response
represents a possible reason for increased susceptibility in C57Bl/6 mice, and may also explain
the difference in structural response to BPA treatment in C57Bl/6 mice. Therefore, the
heightened responsiveness of the C57Bl/6 animals is considered a probable contributing factor
for the sensitivity of this strain to developing estrogen-induced pyometra. Previous studies in
“sensitive” strains of rat have identified quantitative trait loci associated with estrogen-induced
pyometra; however, there is no understanding of the mechanisms involved with sensitivity or
resistance to pyometra. This pathology may be related to interactions between the uterine
immune response and the uterine microbiome. The results here suggest that pyometra in the
C57Bl/6 strain might serve as a sensitive endpoint for understanding the mechanisms responsible
for the impact of estrogen and estrogenic endocrine disrupting chemicals on immunity. Further,

103
the C57Bl/6 strain might serve as a useful model of sensitive subpopulations at risk for
developing immunological disorders related to exposures to estrogenic EDCs.
3.6 Acknowledgements
This work was supported by National Institute of Health grants R01 ES015145, RC2 ES018765,
T32 ES016646, and the University of Cincinnati Center for Environmental Genetics (P30-
ES06096).

104
3.7 Supplemental Results
3.7.1 Uterine morphology: myometrial wall thickness
To characterize the effects of BPA or EE exposure on the myometrium, the thickness of
both outer and inner layers was measured in both strains and generations of mice. In adult CD1
mice, BPA exposure did not alter wall thickness (Figure 5A). However, EE exposure dose
dependently decreased wall thickness, with a significant decrease at 1.3 ppm EE (p<0.05; Figure
5B). In adult C57Bl/6N mice, the 0.03 ppm BPA exposure group exhibited a decreased wall
thickness that was not statistically significant, while increasing BPA exposure led to a dose
dependent increase in wall thickness (Figure 5A). A trend toward dose dependent decreases in
wall thickness in EE-exposed mice was also observed (Figure 5B). No C57Bl/6N mice in the 1.3
ppm EE exposure group were included due to the high incidence of pyometra in this group.
Strain had a significant main effect on wall thickness in the BPA-exposed (p=0.0008) and the
EE-exposed (p=0.0098) groups, with the 0.03 ppm BPA exposure groups in each strain being
significantly different (p<0.01).

105
Figure 5. Myometrial wall thickness in mice across exposure groups and strain. Myometrial wall thickness for adult CD1 (A) and C57Bl/6N (B) mice are shown. Wall thickness measurements were collected as described in section 2.2.4 of the Methods chapter. Values indicated are the mean wall thickness (µm) per group and error bars represent the SEM. For adult CD1, n ≥ 4 animals. For adult C57Bl/6N, n ≥ 3 animals. Wall thickness for BPA- and EE-exposed groups was compared separately to the control group. The level of statistical significance between values in BPA-exposed adult mice was assessed using a two-way ANOVA, with exposure and strain as the main effects, and Bonferroni post hoc test. The 1.3 ppm EE exposure group was not included in two-way ANOVA analysis due to no C57Bl/6N mice being measured in this group. One-way ANOVA and Dunnett’s multiple comparison post hoc test were used to determine the statistical significance in the EE-exposed adult CD1 mice. * indicates a level of difference between values is significant (p<0.05); ** indicates a levels of differences between values is very significant (p<0.01).
3.7.2 Immune response in uteri of CD1 and C57Bl/6N mice
The percent of F4/80-positive cells was quantified in the endometrium and myometrium
of BPA- or EE-exposed adult CD1 and C57Bl/6N mice (Figure 6). In BPA-exposed adult mice,
endometrial F4/80-positive cells exhibited significant increases in both strains (Figure 6A), with
a significant main effect of exposure (p=0.0004) and a significant interaction between exposure
and strain (p=0.0308) based on two-way ANOVA. In the CD1 strain, a significant increase in
F4/80-positive cells occurred at 30 ppm BPA (p<0.001) compared to control. In the C57Bl/6N
strain, significant increases in F4/80-positive cells occurred at 0.03 ppm BPA (p<0.01), 3 ppm
BPA (p<0.01), and 30 ppm BPA (p<0.01) compared to control. No significant differences in

106
endometrial F4/80 counts were observed in EE-exposed adult CD1 or C57Bl/6N mice, but all
groups appear increased compared to control in the C57Bl/6N strain (Figure 6B). However,
exposure had a significant main effect (p=0.0419). When quantifying myometrial F4/80-positive
cells in BPA-exposed mice (Figure 6C), a significant main effect of exposure (p=0.0199) and a
significant interaction between exposure and strain (p=0.0094) occurred. Adult CD1 mice again
had a significant increase at 30 ppm BPA (p<0.05) compared to control, similar to that observed
in endometrial cell counts. In the C57Bl/6N strain, significant increases occurred at 0.03 ppm
BPA (p<0.05), 3 ppm BPA (p<0.01), and 300 ppm BPA (p<0.05) compared to control.
Differences in percent of F4/80-positive cells between strains were observed in the control
(p<0.05) and 30 ppm BPA (p<0.01) groups. A similar difference between strains in the control
group was also observed when quantifying myometrial F4/80-positive cells in EE-exposed adult
mice (p<0.05; Figure 6D). Adult C57Bl/6N mice in the 0.01 ppm EE exposure group had a
significant increase in myometrial F4/80-positive cells compared to control (p<0.05; Figure 6D).
When quantifying F4/80-positive cells in EE-exposed adult mice, strain had a significant main
effect (p=0.0011).

107
Figure 6. Quantification of F4/80-positive cells in BPA- or EE-exposed CD1 and C57Bl/6N adult mice. The percent of F4/80-positive cells was counted in the endometrium (A-B) or myometrium (C-D) of BPA- (A, C) or EE-exposed (B, D) adult CD1 and C57Bl/6N mice. Endometrial and myometrial F4/80-positive cells were quantified separately and normalized to either stromal or smooth muscle cells respectively. F4/80-positive and total cells were counted by an observer blinded to strain and exposure group. Values indicated are the percent of F4/80-positive cells per animal and error bars represent the SEM. For adult CD1, n≥7 animals except n=2 animals in the 3 ppm BPA and 0.001 ppm EE groups. For adult C57Bl/6N, n≥3 animals. The level of statistical significance between values was assessed using a two-way ANOVA, in which strain and exposure group were used as main effects, and Bonferroni post hoc test. * indicates a level of difference between values is significant (p<0.05); ** indicates a level of difference between values is very significant (p<0.01). *** indicates a level of difference between values is extremely significant (p<0.001).

108
CHAPTER 4
Strain-Specific Induction of Endometrial Periglandular Fibrosis in Mice Exposed During
Adulthood to the Endocrine Disrupting Chemical Bisphenol A
Kendziorski JA and SM Belcher. 2015. Strain-specific induction of endometrial periglandular fibrosis in mice exposed during adulthood to the endocrine disrupting chemical bisphenol A. Reprod Toxicol, 58:119-130. doi: 10.1016/j.reprotox.2015.08.001. PMID: 26307436

109
Strain-Specific Induction of Endometrial Periglandular Fibrosis in Mice Exposed During Adulthood to the Endocrine Disrupting Chemical Bisphenol A
Jessica A. Kendziorski and Scott M. Belcher*
Department of Pharmacology and Cell Biophysics University of Cincinnati College of Medicine
Cincinnati, OH, 45267-0575 *Corresponding Author: Scott M. Belcher Department of Pharmacology and Cell Biophysics University of Cincinnati College of Medicine 231 Albert Sabin Way; PO Box 670575 Cincinnati, OH, 45267-0575 Telephone: 513-558-1721 Email: [email protected]

110
4.1 Abstract
The aim of this study was to compare effects of bisphenol A (BPA) on collagen
accumulation in uteri of two mouse strains. Adult C57Bl/6N and CD-1 mice were exposed to
dietary BPA (0.004-40 mg/kg/day) or 17α-ethinyl estradiol (0.00002-0.001 mg/kg/day) as effect
control. An equine endometrosis-like phenotype with increased gland nesting and periglandular
collagen accumulation was characteristic of unexposed C57Bl/6N, but not CD-1, endometrium.
BPA non-monotonically increased gland nest density and periglandular collagen accumulation in
both strains. Increased collagen I and III expression, decreased matrix metalloproteinase 2
(MMP2) and MMP14 expression, and increased immune response were associated with the
endometrosis phenotype in the C57Bl/6N strain and the 30 ppm BPA CD-1 group. The
association between the pro-collagen shift in increased collagen expression and decreased
MMP2 expression and activity implies that strain differences and BPA exposure alter regulation
of endometrial remodeling and contribute to increased fibrosis, a component of several human
uterine diseases.

111
4.2 Introduction
Bisphenol A (BPA) is an endocrine disrupting chemical (EDC) released from
polycarbonate plastics and linings of food and beverage containers that contaminates the
consumed contents of the container (Cooper et al. 2011, Carwile et al. 2009, Krishnan et al.
1993). Consumption of BPA-contaminated foods and beverages is the major route of exposure in
humans, with additional exposure occurring through handling of thermal receipt paper,
application of dental sealants, and transfer from medical equipment (Ehrlich et al. 2014, Calafat
et al. 2009, Pulgar et al. 2000). Supporting widespread and prolonged human exposure to BPA,
measurable levels of BPA metabolites were found in over 93% of human urine samples in the
2003-2004 National Health and Nutrition Examination Survey (Calafat et al. 2005). Because of
the known estrogenic and androgenic endocrine disrupting activity of BPA, results from
numerous in vitro and in vivo studies have demonstrated that development and function of
reproductive tissues, including the uterus, were sensitive to BPA exposures (Caserta et al. 2014,
Peretz et al. 2014, Bredhult et al. 2007, Singleton et al. 2006, Schonfelder et al. 2004, Markey et
al. 2001, Papaconstantinou et al. 2000, Ashby and Tinwell 1998). Estrogenic activity of BPA in
the uterus has been established with the rodent uterotrophic assay, for example (Kendig et al.
2012, Laws et al. 2000), and numerous examples of changes in BPA-responsive gene expression
have been reported from studies in a variety of animal models (Calhoun et al. 2014, Hewitt and
Korach 2011, Hong et al. 2006, Naciff et al. 2002, Diel et al. 2000). Endocrine disrupting actions
of BPA have also been associated with pathological changes in the uteri of BPA-exposed rats
and mice (Kendziorski et al. 2012, Signorile et al. 2010, Newbold et al. 2009, 2007b).
Outside of the reproductive axis, we recently demonstrated that lifelong exposure to as
little as 0.004 mg/kg/day of BPA resulted in functional alterations of the collagen extracellular

112
matrix (ECM) in the hearts of male and female CD-1 mice (Belcher et al. 2015). In the uterus,
the collagen ECM plays a dynamic structural and functional role in tissue remodeling of the
cycling uterus and during pregnancy (Gaide Chevronnay et al. 2012), whereas dysregulation of
ECM function and collagen accumulation contributes to endometrial adenocarcinoma, fibroids
(leiomyomas), and endometriosis (Kim et al. 2013, Klemmt et al. 2007, Jussila et al. 2004, Kao
et al. 2003, Stewart et al. 1994). In addition to being associated with pathology of the human
uterus, dysregulation of collagen accumulation and excessive fibrosis is a hallmark of equine
endometrosis, one of the most important causes of infertility in mares (Allen 1993) . Equine
endometrosis is an age-related and irreversible degenerative disease of the uterus characterized
by markedly increased endometrial stromal and periglandular fibrosis (EPF) associated with
“gland nest” structures and is unrelated to human endometriosis (Hanada et al. 2014, Snider et al.
2011, Evans et al. 1998, Ferreira-Dias et al. 1994, Kenney 1978). The severity of EPF is
inversely correlated to successful conception and gestation and is associated with increased rates
of embryonic or fetal foal loss and increased susceptibility to infection (Kenney 1978). The
etiology of EPF is unknown, in part due to the lack of suitable laboratory animal models to
facilitate experimental studies. The presence of EPF has also been noted in canines (Christensen
et al. 2012); however, it is unknown whether a similar pathology characterized by excessive
periglandular collagen accumulation and gland nest formation occurs in humans or laboratory
rodent models.
Accumulation of collagen in the uterus during the female reproductive cycle is a dynamic
and tightly regulated process involving hormonal control of collagen gene expression and matrix
metalloproteinase (MMP) expression and activity (Gaide Chevronnay et al. 2012). The types I
and III are primary collagens expressed in mammalian uterus, with lower levels of collagen types

113
IV, V, and VI expressed variably during the cycle (Augsburger and Henzi 2008, Boos 2000,
Aplin et al. 1988, Karkavelas et al. 1988). 17β-Estradiol (E2) can increase mRNA and protein
synthesis of many collagen subtypes in the uterus (Frankel et al. 1988, Dyer et al. 1980, Salvador
and Tsai 1973). Estradiol also plays a critical role in the cyclic remodeling of the endometrium
through dynamic modulation of collagen degradation by tightly regulating MMP expression and
activity (Gaide Chevronnay et al. 2012, Russo et al. 2009, Helvering et al. 2005). The MMP9
and MMP2 proteins are key collagen degrading enzymes present in both latent and active forms
in endometrial tissue of the uterus throughout the reproductive cycle (Gaide Chevronnay et al.
2012). Regulation of MMP2 activity during the estrous cycle of rodents and the menstrual cycle
in humans is in part controlled by MMP14, which activates latent MMP2 (proMMP2) by
proteolytic cleavage (Gaide Chevronnay et al. 2012, Strongin et al. 1995, Sato et al. 1994).
Vascular smooth muscle cells respond to increasing levels of estrogen by induction of MMP14
protein expression and a corresponding increase in MMP2 activity (Grandas et al. 2009). Control
of collagen accumulation also involves inhibition of MMP activity through serum and tissue
inhibitors of metalloproteinases (TIMPs) which, in response to varying levels of E2 and
progesterone, are also differentially expressed during the phases of the estrous and menstrual
cycles (Gaide Chevronnay et al. 2012, Goffin et al. 2003, Zhang and Salamonsen 1997, Hampton
and Salamonsen 1994, Sato et al. 1991). TIMP-1, -2, and -3 are all expressed at high levels in the
uterus throughout the cycle, with TIMP-3 exhibiting the most dynamic spatio-temporal
expression profile (Gaide Chevronnay et al. 2012). While the effects of BPA on reproductive
tissues are well-studied, the impacts of exposures to BPA and other EDCs on collagens, MMPs
and the ECM of most tissues, including the uterus, are poorly understood. Evidence supporting
the possibility that BPA may alter MMP activity leading to dysregulation of collagen dynamics

114
in the uterus is limited. However studies in developing mammary gland have identified an
exposure related change in ECM collagen content, and BPA can increase MMP2 and MMP9
expression in ovarian granulosa cells and ovarian cancer cell lines (Ptak et al. 2014, Dominguez
et al. 2008, Vandenberg et al. 2007b).
There are well known differences in the sensitivity of rodent strains to pathologic effects
of estrogenic compounds, including E2, diethylstilbestrol (DES) and BPA. Differences in
sensitivity of a variety of hormonally regulated responses, among them male reproductive
development, prolactin release, oocyte development, and expression of estrogen receptors (ERs)
in uterine epithelium, have been characterized (Pepling et al. 2010, Long et al. 2000, Spearow
1999, Steinmetz et al. 1997, Bigsby et al. 1990, Greenman et al. 1977). Strain-specific responses
to estrogen-like EDCs vary and are dependent on the tissue of interest and the phenotypic or
physiological response being examined. Even substrains derived from the C57Bl/6 line have
been shown to have important genetic and phenotypic differences (Simon et al. 2013, Mekada et
al. 2009). Numerous strain-specific differences in sensitivity of uterine and female reproductive
responses to estrogenic compounds have been reported in regards to uterine weight alterations,
development of pyometra, immune cell infiltration, and onset of puberty, including findings from
our previous work which demonstrated that C57Bl/6N mice were more sensitive to the actions of
17α-ethinyl estradiol (EE) and BPA than CD-1 mice (Kendziorski et al. 2012, Roper et al. 1999,
Griffith et al. 1997, Greenman et al. 1977, Drasher 1955). Based on that differential sensitivity
we hypothesized that the sensitivity to developing collagen-related pathology of the uterus would
likewise differ between the CD-1 and C57Bl/6N strains and that dietary BPA exposure would
alter the regulation of collagen. To examine whether BPA differentially altered collagen
accumulation in the adult uterus, uterine pathology was analyzed in young adult female

115
C57Bl/6N and CD-1 mice that were exposed for 12-15 weeks to dietary BPA at doses extending
below the accepted safe exposure level in humans (0.05 mg/kg/day) to the defined no observed
adverse effect level (NOAEL) of 50 mg/kg/day (Kendig et al. 2012, Kendziorski et al. 2012,
FAO/WHO 2011, IRIS 1988). To characterize the differential impacts of these BPA exposures,
histological examination was performed to examine collagen-related uterine pathology, including
presence of EPF, gland nest formation, and collagen accumulation, with phenotypes compared
across exposure groups and between strains. Immunohistochemistry was also used to compare
differences in the distribution of MMP2, MMP14, F4/80, α-smooth muscle actin (α-SMA), and
ERα in the uterus. MMP expression and activity were also assessed to examine whether changes
in collagen degradation were contributing to strain- and exposure-related differences in collagen
accumulation.

116
4.3 Materials and Methods
4.3.1 Animal Husbandry and Necropsy
All animal procedures were performed in accordance with protocols approved by the
University of Cincinnati Institutional Animal Care and Use Committee and followed
recommendations of the Panel on Euthanasia of the American Veterinary Medical Association.
Six to seven week old C57Bl/6NHsd and Hsd:ICR (CD-1 Swiss) mice were received from
Harlan Laboratories, Inc. (Indianapolis, IN), group housed five per cage and randomly assigned
to control or dietary exposure groups. Animals were housed in a polycarbonate-free housing
system with single-use BPA-free polyethylene cages and water bottles (Innovive, San Diego,
CA). Sanichip bedding (Irradiated Aspen Sani-chip; PJ Murphy Forest Products Corp, Montville,
NJ) was used to prevent contamination from mycoestrogens in corncob bedding. Sterile drinking
water containing <1% of oxidizable organics was produced from a water purification system
designed to eliminate organic chemical contamination (Millipore Rios 16 with ELIX
UV/Progard 2, Billerica, MA). Animals were fed a defined casein-based phytoestrogen-free diet
(Product # D1010501, Research Diets, Inc.; New Brunswick, NJ) unsupplemented (control or 0
ppm) or supplemented with either BPA (2,2-bis(4-hydroxyphenyl)propane; CAS No. 80-05-7;
Lot 11909; USEPA/NIEHS standard) or EE (1,3,5(10)-estratrien-17α-ethinyl-3,17β-diol; CAS
No. 57-63-6; Batch No. H923; Steraloids Inc.; Newport, RI) that was homogenously
incorporated in the diet at desired concentrations. Final dietary BPA concentrations (0.03, 0.3, 3,
30, and 300 ppm) resulted in doses of 0.004, 0.04, 0.4, 4, and 40 mg/kg/day, and concentrations
of EE (0.0001, 0.001, and 0.01 ppm) resulted in doses of 0.00002, 0.0002, and 0.001 mg/kg/day
(Kendig et al. 2012, Kendziorski et al. 2012). Following a 2 week acclimation to the BPA-free
system, study animals were maintained on assigned control or test diet ad libitum until necropsy
at 19 to 23 weeks of age, for a total exposure period of 12 to 15 weeks. During this time, animals

117
were bred and those that produced a litter were included in this study. Progression of estrous
cycles was monitored by morphological analysis of vaginal lavage and necropsy was performed
while in estrus. Estrous stage was confirmed by histological assessment of uterine tissues.
4.3.2 Histology and Immunohistochemistry
Tissue isolation, fixation and preparation were described in detail previously (Kendig et
al. 2012, Kendziorski et al. 2012). Formalin fixed tissues were blocked for sectioning with a
sterile razor blade and washed several times in 70% ethanol prior to automated tissue processing
and embedding in paraffin (Histoplast IM; Richard Allen, Kalamazoo, MI). Uteri were placed in
the block longitudinally to examine the entire length of the uterine horn, as well as the dorsal and
lateral sides of the uterus. Microtome sections were cut at 5 µm thickness from blocks at 4˚C and
placed on positively charged slides for hematoxylin and eosin (H&E) staining and
immunohistochemistry. Standard H&E staining was performed in order to examine tissue
structure and morphology and to conduct histopathological analysis. For immunohistochemistry,
sections were dewaxed in xylene and rehydrated through graded series of ethanol concentrations
into phosphate buffered saline (PBS) for MMP2, F4/80, α-SMA, and ERα immunostaining or
Tris-buffered saline with 0.1% Tween 20 (TBST) for MMP14. Heat-mediated antigen retrieval
was performed at 100oC in 0.01M citrate buffer (pH 6.0). Endogenous peroxidase was blocked
with 3% H2O2 in PBS. Sections were incubated at room temperature (RT) in 3% normal goat
serum in PBS for 1 hour and then incubated overnight at 4oC with either rabbit polyclonal anti-
MMP2 (3.33 µg/mL (1:300); NB200-193, Novus Biologicals; Littleton, CO), rabbit monoclonal
anti-MMP14 (0.5 µg/mL (1:250); ab51074, Abcam; Cambridge, MA), rat monoclonal anti-F4/80
(C1:A3-1; 4 µg/mL (1:250); ab6640, Abcam; Cambridge, MA), rabbit polyclonal anti-α-SMA (1
µg/mL (1:200); ab5694, Abcam; Cambridge, MA), or rabbit polyclonal anti-ERα (MC-20; 1

118
µg/mL (1:200); sc-542, Santa Cruz Biotechnology; Dallas, TX). Immunoreactivity was
visualized with 0.05% 3’3-diaminobenzidine by the avidin-biotin peroxidase complex method
(Vector Laboratories; Burlingame, CA). Sections were counterstained in Hematoxylin 2 (Richard
Allen; Kalamazoo, MI) for 30 seconds, dehydrated, and coverslipped using Permount (Fisher
Scientific; Fair Lawn, NJ). Mouse lung tissue was used as a positive control for MMP2 and
MMP14, mouse spleen tissue was used as a positive control for F4/80, mouse heart tissue was
used as a positive control for α-SMA, and rat mammary tumor was used as a positive control for
ERα. Negative staining controls included replacement of primary antibodies with non-specific
serum. Stained sections were examined on a Nikon Eclipse 55i microscope using a DS-Fi1 CCD
camera controlled with Digital Sight Software (Nikon; Melville, NY). Final figures were
generated using Adobe Photoshop (San Jose, CA).
4.3.3 Picrosirius Red Stain and Collagen Assessment
Paraffin-embedded uterine sections were stained with Picrosirius Red (Polysciences;
Warrington, PA) to visualize total collagen (red) with bright field illumination and by
polarization birefringence to assess the thickness and packing density of collagen fibers (Dayan
et al. 1989, Junqueira et al. 1979, Junqueira et al. 1978). Briefly, tissue sections were
deparaffinized, rehydrated, and stained for 8 minutes with Weigert’s hematoxylin (American
MasterTech; Lodi, CA). Stained sections were rinsed with tap water for 5 minutes, incubated for
2 minutes in 0.2% phosphomolybdic acid hydrate, and rinsed in deionized H2O for 30 seconds.
Slides were then stained for 1 hour in Picrosirius Red F3BA solution (1.3% 2,4,6-trinitrophenol,
0.4% Direct Red 80), transferred to 0.1 N hydrochloric acid solution for 2 minutes, washed in
70% ethanol for 45 seconds, dehydrated and then coverslipped. Stained sections were examined
on a Nikon Eclipse 80i microscope equipped with a polarized light attachment and a DS-Fi1

119
CCD camera controlled with Digital Sight Software (Nikon; Melville, NY). Final figures were
generated using Adobe Photoshop.
4.3.4 Quantitative RT-PCR
Frozen uterine tissue ≤30 mg from CD-1 control and 30 ppm BPA exposure groups were
homogenized for RNA isolation using an RNeasy Fibrous Tissue Mini Kit (Qiagen; Valencia,
CA). Uterine RNA was converted to cDNA using a High Capacity cDNA Reverse Transcription
Kit (Life Technologies; Grand Island, NY). Predesigned TaqMan RT-PCR assays (Applied
Biosystems; Grand Island, NY) were used for the following genes: Col1a1 (Mm00801632_gH),
Col3a1 (Mm00802296_g1), Mmp2 (Mm00439498_m1), Timp1 (Mm00441818_m1), Timp2
(Mm00441825_m1), and Timp3 (Mm00441826_m1). PCR amplification was performed in a
final volume of 20 µL containing 1x TaqMan expression assay primers, Universal Master Mix
(Applied Biosystems; Grand Island, NY), and 10 ng of cDNA. Amplification was performed in
triplicates on a Step One Plus Real-Time PCR System (Applied Biosystems; Grand Island, NY)
under the following fast conditions: 20 seconds at 95oC for one holding stage, then 1 second at
95oC and 20 seconds at 60oC for the cycling stage (60 cycles). Relative expression was
quantified using the ΔΔCt method, using 18s as the endogenous control for normalization.
4.3.5 SDS-PAGE and Immunoblotting
Uterine lysates from frozen tissue were generated by homogenization in 20 mM Tris-HCl
(pH 7.5) with 150 mM NaCl, 1 mM EDTA, 1 mM EGTA, and 1% Triton containing protease
(Roche; Nutley, NJ) and phosphatase (Sigma-Aldrich; St. Louis, MO) inhibitors. Protein
concentrations of cleared lysates were determined using the Bio-Rad Dc protein assay (Bio-Rad;
Hercules, CA). Protein lysate samples (30 µg) were mixed 3:1 with sample buffer (0.25 M Tris-
HCl, 40% glycerol, 8% SDS, 0.01% bromophenol blue, pH 6.8) containing 0.04% β-

120
mercaptoethanol and incubated at 96oC for 5 minutes. Protein was loaded onto 10%
polyacrylamide gel and resolved by electrophoresis. Proteins were electrotransferred to an
Odyssey® nitrocellulose membrane (Li-Cor Biosciences; Lincoln, NE) for 16 hours at 4oC.
Membranes were stained with 0.1% Ponceau S in 1% acetic acid to confirm homogenous protein
transfer, destained in water and then blocked with 5% non-fat dry milk (Blotting Grade Blocker,
Bio-Rad; Hercules, CA) in TBST for 1 hour at RT. Membranes were incubated overnight at 4oC
with either rabbit polyclonal anti-MMP2 (0.25 µg/mL (1:4000); NB200-193, Novus Biologicals;
Littleton, CO) or rabbit monoclonal anti-MMP14 (0.066 µg/mL (1:2000); ab51074, Abcam;
Cambridge, MA). Blots were coincubated with a rabbit monoclonal anti-β-actin antibody (0.02
µg/mL (1:2000); 8457, Cell Signaling; Danvers, MA) for normalization of loading. Membranes
were washed and incubated with IRDye® 800CW goat anti-rabbit IgG (0.2 µg/mL (1:5000); Li-
Cor Biosciences; Lincoln, NE) for 1 hour at RT. Membranes were imaged using an Odyssey®
CLx Infrared Imaging System (Li-Cor Biosciences; Lincoln, NE) and immunoreactive bands
were visualized and analyzed with Image Studio Lite Software (Li-Cor Biosciences; Lincoln,
NE).
4.3.6 Gelatin Zymography
Methods for gelatin zymography were adapted from previously described protocols (Toth
et al. 2012, Belcher et al. 2005). Frozen uterine tissue was homogenized in 0.025 M Tris-HCl
(pH 7.5), 0.1 M NaCl, 1% Nonidet P-40 containing EDTA-free Protease Inhibitor Cocktail
(Roche; Nutley, NJ) and cleared by centrifugation at 4oC. Protein concentration was determined
using the Bio-Rad Dc protein assay (Bio-Rad, Hercules, CA). Equal amounts of total protein (30
µg) were mixed 3:1 with sample buffer (0.25 M Tris-HCl, 40% glycerol, 8% SDS, 0.01%
bromophenol blue, pH 6.8) and incubated at RT for 15 minutes. Protein was resolved onto native

121
10% polyacrylamide gel containing 0.1% gelatin. Following electrophoresis, gels were washed
with gentle agitation in 2.5% Triton X-100 at RT for 30 minutes, rinsed in dH2O, and then
washed for an additional 30 minutes at RT with gentle agitation in developing buffer (0.05 M
Tris-HCl (pH 7.8), 0.2 M NaCl, 0.005 M CaCl2). Washed gels were then incubated in fresh
developing buffer at 37oC for 16 hours, stained with 0.5% Coomassie Brilliant Blue R-250 in 5%
methanol and 10% glacial acetic acid at RT for 3 hours, and then destained at RT for consecutive
washes of 15 minutes, 30 minutes, and 1 hour with 5% methanol and 10% acetic acid.
Conditioned media from the human brain tumor cell line DAOY (ATCC HTB-186) was used as
a positive control for MMP activity. In control experiments, the addition of EDTA (1 mM final
concentration) to lysates or developing buffer resulted in loss of proteolytic activity at the
expected molecular weights. Gels were imaged using an Alpha Innotech Multiimage II (Alpha
Innotech; San Leandro, CA) and optical density of cleared bands denoting areas of proteolytic
activity were quantified using Image Studio Lite Software.
4.3.7 Statistical Analysis
Pathology was assessed by an observer blinded to exposure and strain with statistical
differences analyzed using Fisher’s exact test. Gland nest density was analyzed using a two-way
ANOVA. Differences between control and exposure groups were analyzed using Dunnett’s
multiple comparison test and a Student’s t-test was used for analysis of mRNA and protein
expression and protein activity. Significance between differences in values was defined as
p<0.05. All data was analyzed using GraphPad Prism® v5/v6 software (GraphPad; La Jolla,
California).

122
4.4 Results
4.4.1 Uterine Pathology: Gland Nests and Periglandular Fibrosis in C57Bl/6N and CD-1 Mice
A pathological assessment of a longitudinal section of the uterine horn between the
oviduct and the cervix was conducted. The number of animals presenting with each pathology is
summarized in Table 1. The uteri of control C57Bl/6N and CD-1 mice were characterized by a
low incidence of uterine cysts. Uterine cysts were defined as hyperplastic glands surrounded by a
thin layer of cuboidal epithelial cells. Distension of the glands and increased stromal
proliferation was commonly observed (Table 1).
Table 1. Incidence of Uterine Pathology in CD-1 and C57Bl/6N Mice at 19-23 Weeks of Age
BPA (ppm) EE (ppm)
CD-1 0 0.03 0.3 3 30 300 0.0001 0.001 0.01
n= 12 9 10 10 8 10 10 10 10
Gland Nests 1 (8%) 0 2 (20%) 0 6 (75%)a 3 (30%) 4 (40%) 0 0
Distended Glands 7 (58%) 7 (78%) 6 (60%) 6 (60%) 6 (75%) 8 (80%) 8 (80%) 8 (80%) 8 (80%)
Cysts 1 (8%) 1 (11%) 2 (20%) 2 (20%) 1 (13%) 0 0 0 0
C57Bl/6N
n= 7 7 7 4 6 3 4 3 6
Gland Nests 5 (71%) 6 (86%) 5 (71%) 4 (100%) 5 (83%) 3 (100%) 3 (75%) 2 (67%) 4 (67%)
Distended Glands 6 (86%) 5 (71%) 4 (57%) 3 (75%) 4 (67%) 2 (67%) 1 (25%) 2 (67%) 2 (33%)
Cysts 0 2 (29%) 3 (43%) 2 (50%) 0 1 (33%) 1 (25%) 1 (33%) 1 (17%)
a p = 0.0044 by two-tailed Fisher's exact test
Gland nests, characterized by multiple uterine glands surrounded by collagen producing
stromal cells with resulting periglandular fibrosis and marked collagen accumulation (Fig. 1
arrows), were frequently observed in C57Bl/6N control and in each of the C57Bl/6N BPA and
EE exposure groups (Table 1; Fig. 1). Compared to control, an increased intensity of red/yellow
and decreased green polarization birefringence, characteristic of increased collagen fiber
thickness and density, was associated with periglandular regions of stromal cells in the 30 ppm
BPA and 0.01 ppm EE exposure groups in C57Bl/6N mice (Fig. 1B; D; F).

123
Figure 1. Effects of oral exposure to BPA or EE on uterine morphology and collagen accumulation in C57Bl/6N mice. Shown are representative photomicrographs of H&E- and Picrosirius Red-stained uterine sections from a control C57Bl/6N mouse (A-B) and from a mouse exposed to 30 ppm BPA (C-D) or 0.01 ppm EE (E-F). Similar uterine locations are highlighted in each panel. Arrows designate gland nests. Scale bar represents 50 µm.
In contrast to C57Bl/6N controls, only a single gland nest-like structure was observed in
one uterus from the CD-1 control group (Table 1). A significant increase in gland nests (Table 1;
Fig. 2C, arrows) and extensive periglandular fibrosis was observed in the CD-1 30 ppm BPA
exposure group (Fig. 2D) compared to control (Fig. 2A). In contrast to controls, intense

124
red/yellow polarization birefringence, forming net-like patterns in areas between gland nest
structures, was characteristic of the CD-1 30 ppm BPA exposure group (Fig 2D). Gland nests
were not present in the phenotypically estrogenized uteri of CD-1 mice exposed to 0.01 ppm EE
(Fig. 2E-F), and notably lower levels of red/yellow and higher levels of green polarization
birefringence were observed, suggesting a less densely packed collagen (Fig. 2F).
Figure 2. Effects of oral exposure to BPA or EE on uterine morphology and collagen accumulation in CD-1 mice. Shown are representative photomicrographs of H&E- and Picrosirius Red-stained uterine sections from a control CD-1 mouse (A-B) and from a mouse exposed to 30 ppm BPA (C-D) or 0.01 ppm EE (E-F). Similar uterine locations are highlighted in each panel. Arrows designate gland nests. Scale bar represents 50 µm.

125
Figure 3. Quantification of gland nest density in the uteri of control and BPA-exposed CD1 and C57Bl/6N mice. Comparison of gland nest density between the control groups of CD1 and C57Bl/6N mice (A), control and BPA-exposed CD1 mice (B), and control and BPA-exposed C57Bl/6N mice (C). The number of gland nests was counted by an observer blinded to strain and exposure group. Values indicated are the number of gland nests per mm2 per animal. Error bars represent the SEM. For C57Bl/6N, n≥3 animals and for CD1, n≥8 animals. The level of statistical significance between values was assessed using a two-way ANOVA, in which exposure and strain were the main effects. * indicates a level of differences between values is significant (p<0.05); ** indicates a level of differences between values is very significant (p<0.01).
The density of gland nests in the endometrium for each strain and exposure group was
calculated (Figure 3). Two-way ANOVA of gland nest density indicated significant main effects
of both strain [F(1,77) = 26.99, (p<0.0001)] and exposure [F(5,77) = 4.411, (p=0.0014)] with no
significant interaction between factors [F(5, 77) = 1.493, (p=0.2019)]. The mean density of gland
nest in the endometrium of control C57Bl/6N uteri (M = 0.895 ± SD 0.927) was 9 times greater
than the density observed in the CD-1 strain (M = 0.101± SD 0.349; Fig. 3A). Gland nest density
of neither C57Bl/6N [F (3, 15) = 1.568, (p=0.2385)] nor CD-1 uteri [F (3, 38) = 0.6947,
(p=0.5610)] was affected by EE exposure. Exposure to 30 ppm BPA significantly increased
endometrial gland nest density in the CD-1 strain (Fig. 3B) and significant increases in density
were detected in 0.03 ppm BPA group and in the two highest BPA groups in the C57Bl/6N strain
(Fig. 3C). The increased density detected in the lowest exposure group and the dose dependent
increase in gland nest density observed across the 0.3 to 300 ppm dose range is suggestive of a
non-monotonic dose response relationship. The significant increase in gland nest density in the

126
30 ppm BPA CD-1 mice correlated with the increased number of animals observed to have gland
nests in Table 1. In C57Bl/6N mice, however, gland nest density also increased in response to
BPA whereas the number of animals with gland nests was not changed (Table 1).
Figure 4. Relative expression levels and activity of mRNA and proteins involved in collagen synthesis and degradation. Quantitative RT-PCR analysis between control (0) and BPA-exposed (30) CD-1 mice for Col1a1, Col3a1, Mmp2, Timp1, Timp2, and Timp3 (A). The relative differences in gene expression between control and exposed groups were determined by the ΔΔ cycle threshold method with specific TaqMan assays. Representative Western blots for proMMP2 and MMP2 (B) and proMMP14 and MMP14 (C) are shown for control CD-1 and C57Bl/6N mice and 30 ppm BPA-exposed CD-1 mice. Relative proMMP2 and MMP2 protein expression levels (D) and activity (E) and protein expression levels of proMMP14 and MMP14 (F) are shown. Protein expression levels are based on Western blot analysis and activity is based on gelatin zymography as described in Methods. Results are expressed as mean of group ± SEM. Expression and activity of BPA-exposed CD-1 mice and control C57Bl/6N mice is graphed in relation to control CD-1 results for all experiments. Control CD-1 mice were compared to BPA-exposed CD-1 mice and control C57Bl/6N mice separately. For qRT-PCR, n≥4 animals. For protein expression and activity, n≥4 animals for CD-1 mice and n≥3 animals for C57Bl/6N mice. The level of statistical significance for differences between mean values of control and BPA-exposed groups was determined by Student’s t test for all experiments and is indicated by * (p<0.05).
4.4.2 Analysis of Collagen and MMP mRNA Expression
The extensive periglandular fibrosis and increased levels of collagen observed in the CD-
1 30 ppm BPA exposure group were suggestive of increased collagen expression and/or

127
alterations in regulation of collagen fibril accumulation. The relative expression levels for
Col1a1, Col3a1, Mmp2, Timp1, Timp2, and Timp3 mRNA in uterine tissues from CD-1 mice in
the control (0) and 30 ppm BPA (30) exposure groups were quantified using qRT-PCR (Fig.
4A). Compared to control, significant increases in Col1a1 (p=0.0379) and Col3a1 (p=0.0161)
and a significant decrease (p=0.0162) in Mmp2 mRNA expression levels were detected in uteri
from CD-1 mice in the 30 ppm BPA exposure group. While Timp1 was not altered by BPA
exposure, Timp2 was significantly decreased (p = 0.0013) in the 30 ppm BPA exposure group
compared to control. Because of its broad inhibitory spectrum and more variable regulation in
the uterus (Gaide Chevronnay et al. 2012), Timp3 expression was also compared in the uterus of
control and 30 ppm BPA exposure groups and was found to be unchanged by BPA exposure.
4.4.3 Analysis of Protein Expression and Activity of MMPs
Western blot analysis was used to determine if protein expression of MMP2 (Fig. 4B)
and MMP14 (Fig. 4C) in uteri from the C57Bl/6N control group and CD-1 30 ppm BPA
exposure group were decreased compared to unexposed CD-1 controls, because these groups
displayed significant differences in gland nest formation and collagen accumulation compared to
CD-1 control animals. Relative to unexposed CD-1 controls, decreased proMMP2 and MMP2
protein levels were detected in both CD-1 30 ppm BPA and unexposed C57Bl/6N groups (Fig.
4B). Quantification found the decrease in expression of proMMP2 (t(9) = 3.014, p=0.0073) and
MMP2 (t(8) = 2.397, p=0.0073) in CD-1 30 ppm BPA and proMMP2 (t(7) = 3.404, p=0.0057)
and MMP2 (t(6) = 1.971, p=0.0481) in C57Bl/6N controls was significant (Fig. 4D). The
decreased expression of MMP2 was reflected by relative decreases in MMP2 gelatinase activities
of the CD-1 30 ppm BPA exposure group and C57Bl/6N control group as compared to
unexposed CD-1 controls (Fig 4E). The relative decreases in proMMP2 (t(10) = 5.486,

128
p=0.0003) and MMP2 (t(6) = 4.610, p=0.0037) activity in the CD-1 30 ppm BPA exposure
group and proMMP2 (t(9) = 5.789, p=0.0003) and MMP2 (t(5) = 6.357, p=0.0007) activity in
C57Bl/6N controls were significant (Fig. 4E). As expected (Gaide Chevronnay et al. 2012), the
relative activity of MMP9 in C57Bl/6N and CD-1 uterus were not significantly different (t(4) =
0.0906, p=0.9322). Decreased levels of proMMP14 and MMP14 protein levels were also
detected in both CD-1 30 ppm BPA and unexposed C57Bl/6N groups (Fig. 4C). Expression of
proMMP14 in CD-1 30 ppm BPA (t(9) = 2.056, p=0.0350) and C57Bl/6N control (t(7) = 4.367,
p=0.0016) groups was significantly decreased. While it appeared that relative expression of
MMP14 was also decreased in the C57Bl/6N control group (t(7) = 1.481, p=0.0910), results
were not statistically significant due to the high levels of variance associated with quantifying the
more diffusely migrating MMP14 immunoreactive band observed in the CD-1 controls (Fig. 4C;
F).
4.4.4 Immunolocalization of MMP2 and MMP14 in the Endometrium
In the endometrium of the CD-1 control group, intense positive staining for MMP2 was
found throughout the stroma, as well as in the cytoplasmic compartment of luminal and
glandular epithelial cells (Fig. 5A). Intense staining of stromal cells was more variable with
moderate to high intracellular staining and more diffuse staining in extracellular regions (Fig.
5A). Expression did not appear altered in stroma closer to the glandular or luminal structures.
In C57Bl/6N controls, the cell specific staining pattern in the endometrium was similar,
but comparatively less intense (Fig. 5B). Immunoreactivity for MMP2 was also weaker in the
cytoplasm of glandular and luminal epithelial cells, reflecting the overall decreased expression
levels of MMP2 in the C57Bl/6N uterus. Intracellular MMP2 staining of stromal cells was

129
generally weak and further reduced to background levels in the periglandular stroma surrounding
the gland nest (Fig. 5B; arrows).
Immunoreactivity for MMP14 was weak to moderate in the intracellular compartment of
stromal cells in endometrium of both CD-1 and C57Bl/6N controls. In the CD-1 endometrium,
mainly scattered stromal cells surrounding glands were MMP14 immunopositive, and glandular
and luminal epithelial cells were immunonegative (Fig. 5F). In the C57Bl/6N endometrium,
intracellular immunostaining was weak to moderate and more diffusely granular in appearance.
Notably less intense periglandular staining was typical (Fig. 5G; arrow). Weak to moderate
immunostaining was detectable in the borders of luminal epithelial cells.

130
Figure 5. Strain differences in the immunohistochemical localization of MMP2 and MMP14 in the uteri of control CD1 and C57Bl/6N mice. Shown are representative photomicrographs of uterine sections stained for MMP2 in CD1 (A) and C57Bl/6N (B) mice and for MMP14 in CD1 (F) and C57BL/6N (G) mice. Arrows in B and G point to periglandular areas with decreased staining. Control for specificity of immunostaining included the replacement of primary antibody with non-specific goat serum for MMP2 (C) and MMP14 (H). Mouse lung tissue was used as a positive control for immunostaining for MMP2 (D) and MMP14 (I), with the replacement of primary antibody with non-specific goat serum for MMP2 (E) and MMP14 (J) used to control for specificity of staining. Scale bar represents 50 µm.

131
Figure 6. Immunohistochemical characterization of endometrosis-like phenotype between BPA exposure groups or strain. Shown are representative photomicrographs of uterine sections from 0 ppm CD-1 mice (A-C), 30 ppm BPA CD-1 mice (D-F), and 0 ppm C57Bl/6N mice (G-I) stained for F4/80 (A, D, G), α-SMA (B, E, H), or ERα (C, F, I). Control for specificity of immunostaining included the replacement of primary antibody with non-specific goat serum for F4/80 (A, inset), α-SMA (B, inset), and ERα (C, inset). As a positive control for immunostaining, mouse spleen tissue was used for F4/80 (D, inset), mouse heart tissue was used for α-SMA (E, inset) and rat mammary tumor was used for ERα (F, inset). Scale bar represents 50 µm.
4.4.5 Immunohistochemical Characterization of an Endometrosis-Like Phenotype
Immunohistochemistry using antibodies against F4/80, α-SMA, and ERα was performed
on uterine sections from the 0 ppm CD-1, 30 ppm BPA CD-1, and 0 ppm C57Bl/6N groups to
further characterize the effects of BPA exposure or strain on the observed endometrosis-like

132
phenotype (Fig. 6). An increase in F4/80 positive macrophages, indicative of a generally
increased immune response, surrounding gland nests and throughout the stroma was found in the
30 ppm BPA CD-1 exposure group (Fig. 6D) compared to the 0 ppm CD-1 group (Fig. 6A). In
the 0 ppm C57Bl/6N group, higher levels of F4/80 positive macrophages were observed in the
stroma surrounding gland nests (Fig. 6G) as compared to the stroma surrounding glands in the 0
ppm CD-1 group. No staining for α-SMA was observed in periglandular stromal cells
surrounding gland nests in the 30 ppm BPA CD-1 exposure group (Fig. 6E) or the 0 ppm
C57Bl/6N group (Fig. 6H). Endothelial cells surrounding blood vessels in the stroma were α-
SMA immunopositive as expected. Staining for ERα was localized to the nuclear regions in the
glandular epithelium or stromal cells and relative levels did not appear altered by BPA exposure
(Fig. 6F) or strain (Fig. 6I) compared to 0 ppm CD-1 (Fig. 6C).

133
4.5 Discussion
Bisphenol A is an EDC with known pathophysiologic activity in the uterus; little
however is known about the relationship between BPA exposure and fibrosis, a common feature
of several uterine pathologies. In this study, the presence of periglandular-associated stromal
cells and fibrosis surrounding clusters or “nests” of endometrial glands, a hallmark of equine
endometrosis, was found to commonly occur in the uterus of C57Bl/6N mice. To our knowledge,
this is the first report of an endometrosis-like phenotype in the murine uterus. The histological
features of the observed EPF and gland nest phenotype suggests that the C57Bl/6N strain may be
a useful “off the shelf” experimental model for understanding the etiology and pathogenesis of
progressive increases in periglandular fibrosis and chronic endometrial degeneration in horses.
The decreased reproductive efficiencies and loss of fertility caused by equine endometrosis in
mares suggests that the endometrosis-like phenotype may play a role in the decreased average
litter size characteristic of C57Bl/6N (6.5 pups) compared to CD-1 mice (11 pups) (Harlan
2009a, 2009b, Evans et al. 1998). Whereas age is the most important factor related to onset of
equine endometrosis in mares (Hanada et al. 2014, Snider et al. 2011), the observed increases in
density and incidence of gland nests, as well as increased periglandular fibrosis resulting from
BPA exposure in both strains of mice suggest that genetic and endocrine disruptive effects
contribute to alterations in stromal collagen accumulation and the development of endometrosis.
The horse and the mouse have distinct similarities and differences in regards to uterine
biology that need to be taken into account when considering the mouse as a model to study a
disease mainly found in equine populations. Adenogenesis in both species continues postnatally
and can be inhibited by progesterone treatment (Cooke et al. 2012b, Filant et al. 2012,
Gerstenberg and Allen 2000). In both species, adenogenesis results in simple tubular gland

134
structures (Gray et al. 2001); however, glands eventually branch in mares (Gerstenberg and
Allen 2000), whereas they remain simple tubular structures in mice. Regarding estrous cyclicity,
the two species display distinct patterns. While mice are polyestrous throughout the year,
cyclicity in mares is seasonal, occurring from late April to August. Outside of those months,
mares remain in a state of anestrus (Aiello 2014, Sendel 2010). In order for parturition to occur
in both species, glands and their secretions are crucial for blastocyst implantation,
decidualization, and fetal survival as implantation does not occur in mares until day 35
postovulation (Filant and Spencer 2014, Gray et al. 2001), with implantation occurring on day
4.5 in mice (Wang and Dey 2006). Postpartum, the uterus involutes and returns to normal
cyclicity within ten days for both species (Stewart et al. 2011, Gray et al. 2001, Gomez-Cuetara
et al. 1995). Differences aside, the important similarities in uterine biology between the two
species suggest that the mouse may be a useful model to study equine disease.
In response to BPA exposure, gland nest density in both mouse strains examined here
showed a non-monotonic dose response relationship commonly observed for EDCs. These
complex dose-response relationships can be a result of several separate, yet simultaneously
acting, mechanisms of action (Vandenberg et al. 2012). Estrogens and BPA are known to act
with numerous receptor mediated pathways, either through classical nuclear receptor or rapid
signaling mechanisms (Cookman and Belcher 2014, Belcher 2008). While BPA has estrogen-
like activity at ERα and ERβ at nM concentrations (Wetherill et al. 2007), it can also bind to and
inhibit the androgen and thyroid hormone receptors at µM concentrations (Teng et al. 2013,
Moriyama et al. 2002, Paris et al. 2002, Sohoni and Sumpter 1998) to cause oppositional effects
at low and high dose ranges. BPA can also elicit multiple responses through binding of a single
receptor, as shown in developing neurons and rat cardiomyocytes (Liang et al. 2014, Belcher et

135
al. 2005) and partial agonist-like activity of BPA at ERα in the uterus is well established. For
example BPA can increase progesterone receptor (PR) expression in the immature rat uterus, but
also antagonizes E2-induced stimulation in PR expression (Gould et al. 1998). Further study is
required to elucidate the nature of the multiple regulatory mechanisms responsible for the
differential effects of BPA on gland nest density that were observed at low and high BPA doses,
and the nature of the baseline differences observed between the CD-1 and C57Bl/6N strains of
mice.
Excessive collagen accumulation and fibrosis in endometrial tissue is associated with the
progression of numerous diseases of the human uterus, most notably leiomyomas. Leiomyomas,
or uterine fibroids, are one of the most common benign pathologies to affect women of
reproductive age (Baird et al. 2003). The connection between fibrosis and alterations in ECM
signaling in uterine fibroids is well established (Leppert et al. 2004). During the proliferative
phase, mRNA for collagen I and III is overexpressed in uterine fibroids compared to surrounding
tissue, suggesting that ECM components are under hormonal control within the fibroid (Stewart
et al. 1994, Brandon et al. 1993). Exposure to BPA and the development of leiomyomas has also
been experimentally established, since exposure led to the presence of leiomyomas in murine
uteri at 18 months of age (Newbold et al. 2007b), and in vitro studies in human UL uterine
fibroid cells found that BPA increased proliferation and increased mRNA and protein expression
of ERα, IGF-1 and VEGF (Shen et al. 2014). The marked increase in collagen accumulation
induced by BPA in our study further supports the possibility that BPA exposure may contribute
to fibrotic uterine diseases and defined dysregulation of collagen accumulation dynamics is a
plausible mechanism of action. It is notable that the doses of BPA that led to alterations in
collagen accumulation observed here are below the defined NOAEL for BPA and that increases

136
in gland nest density in the C57Bl/6N strain occurred at exposures below what is considered safe
for humans (1982).
Proteins involved in collagen synthesis and degradation are transcriptionally controlled
throughout the estrous or menstrual cycle (Gaide Chevronnay et al. 2012, Dyer et al. 1980). To
assess the relationship between BPA exposure and changes in collagen synthesis and
degradation, we first quantified mRNA expression of representative genes involved in these
processes at an exposure level (30 ppm) in CD-1 mice that led to clear increases in gland nest
formation and collagen accumulation. While it is known that E2 can increase mRNA levels of
collagens and MMPs in the uterus (Helvering et al. 2005, Frankel et al. 1988), exposure to BPA
resulted in increased Col1a1 and Col3a1 expression and decreased Mmp2 expression. In relation
to collagen synthesis, BPA acted similarly to E2 (Frankel et al. 1988). However, BPA had an
effect opposite of E2 on genes involved in collagen degradation, a finding in support of BPA
having antagonist-like actions that are interfering with the normal function of E2 in the uterus.
These findings point toward an imbalance between collagen synthesis and degradation leading to
an overall fibrotic phenotype.
Protein expression and activity of MMPs were quantified in order to assess the effect of
exposure and strain. Exposure to BPA significantly decreased the expression and activity of
latent and active forms of MMP2 and expression of the latent form of MMP14. The C57Bl/6N
strain also had significantly decreased latent and active MMP2 expression and activity and latent
MMP14 expression compared to the CD-1 strain. These results coincide with the mRNA
expression results, supporting that BPA decreased Mmp2 mRNA expression leading to a
decrease in protein expression. Decreased expression of proMMP14 and a trend toward
decreased expression of the active form add another layer of complexity to the dysregulation of

137
collagen degradation in the BPA-exposed CD-1 uteri or the control C57Bl/6N uteri. A lower
level of MMP14 may also lead to a decrease in activation of MMP2, which would contribute to
an overall increase in collagen accumulation in the uterus.
The diffuse MMP2 immunostaining in the stromal compartment is similar to previous
observations in equine endometrosis (Walter et al. 2005). One difference between equine
endometrosis and the pathology observed in the C57Bl/6N and CD-1 strains was a lack of
MMP2 localization to the glandular and luminal epithelial cells in the mouse endometrium. Also
similar to equine endometrosis (Rebordao et al. 2014), an increased immune response associated
with gland nests was observed, which may have a role in the development of the phenotype. In
equine endometrosis, the periglandular stromal cells can differentiate into myofibroblasts that
express α-SMA (Walter et al. 2001). Differences in ERα staining and localization may also
indicate an active or inactive phenotype (Hoffmann et al. 2009b). We did not observe α-SMA
staining in the periglandular stromal cells or differences in ERα staining and localization, further
supporting the interpretation that gland nests present in these young adult mice represent an early
stage of pathology prior to loss of fertility. Thus, the C57Bl/6N mouse may serve as a small
animal model for examining the etiology and progression of equine endometrosis and also may
be useful in determining the role of EDCs in this disease.
We previously found that life-long BPA exposure resulted in sex-specific pathologic
changes in the ECM of the CD-1 mouse heart (Belcher et al. 2015). Along with those sex-
specific responses, differential responses to BPA exposure based on strain or genetic background
have also been reported (Kendziorski et al. 2012, Drasher 1955). In the uterus, BPA or EE can
significantly increase immune responses in C57Bl/6N mice resulting in development of
pyometra, yet this pathology was not observed in CD-1 mice (Kendziorski et al. 2012). Related

138
strain-specific responses to BPA have also been reported in other estrogen-responsive tissues
(Wadia et al. 2007, Long et al. 2000, Steinmetz et al. 1997). In this study, a distinct strain
difference in the response to BPA was observed, as well as differences between the control
groups of mice in each strain. Strain differences may help to explain the etiology of
endometrosis. C57Bl/6N control mice already exhibit a high level of gland nests and
periglandular fibrosis, signifying a potentially unrecognized underlying genetic component to
progression of this phenotype, whereas BPA exposure in the CD-1 strain was required to induce
gland nest development through mechanisms presumably involving dysregulation of collagen
accumulation. Potential differences in baseline levels of ovarian hormones, ER and PR
expression, or cellular responsiveness to E2 or progesterone could be playing a role in the
differential development of an endometrotic-like phenotype observed in the CD-1 and C57Bl/6N
strains (Pepling et al. 2010, Roper et al. 1999, Griffith et al. 1997, Bigsby et al. 1990).
In conclusion, BPA may act as a partial ER agonist to inhibit actions of E2 and result in
collagen dysregulation and potentially fibrosis in the uterus. Up until now, little was known
about the impact of BPA exposure on the regulation of collagen in the uterus. This study informs
that BPA exposure increases gland nest formation and stromal and periglandular collagen
accumulation in both CD-1 and C57Bl/6N mouse strains. In relation to collagen synthesis, BPA
acts similarly to E2 by increasing mRNA expression levels of Col1a1 and Col3a1. However,
BPA exposure has the opposite effect on collagen degradation as would be expected from an
exposure to E2. Mmp2 mRNA expression, protein expression levels of MMP2 and MMP14 and
activity of MMP2 were decreased in BPA-exposed CD-1 mice and control C57Bl/6N mice
compared to control CD-1 mice. These decreases in MMP expression and activity may result in
decreased collagen degradation and lead to an accumulation of collagen in the stromal and

139
periglandular compartments of the endometrium. Over time, this accumulation may lead to
fibrosis, a component of several uterine diseases that affect humans and animals, suggesting that
exposure to EDCs may play a role in the development or progression of those diseases. This
study is the first report of an endometrosis-like phenotype in the murine uterus, as well as the
first report of an EDC leading to the presence of this phenotype in any animal model. Based on
these results, the C57Bl/6N strain may serve as an experimental animal model for studying the
etiology and progression of equine endometrosis, as well as determining the effects of exposures
to EDCs on this disease and other uterine pathologies.
4.6 Acknowledgements
This work was supported by research grants R03ES023098 and RC2ES018765 from the National
Institute of Environmental Health Sciences (NIEHS) and a predoctoral fellowship awarded to
JAK from the Albert J. Ryan Foundation. The authors would like to acknowledge and thank Dr.
Vinicius Carreira for his assistance with pathological assessment, Ms. Robin Gear for her
guidance on qRT-PCR, Dr. Eric Kendig, Ms., Susie Christie, and Ms. Dana Buesing for their
planning and execution of the oral BPA/EE exposure study, and Dr. Burns Blaxall for his
generous gift of the α-SMA antiserum.

140
CHAPTER 5
Effects of Bisphenol A and 17α-Ethinyl Estradiol in Nulligravida Uterus
of CD1 and C57Bl/6N Mice: Whole Life Exposure

141
5.1 Introduction
The aims of this study were to examine the uteri of CD1 and C57Bl/6N mice with whole
life exposure to BPA or EE for increases in pathology, collagen dysregulation, and immune
responsiveness. Based on studies in adult mice exposed to BPA or EE (Kendziorski and Belcher
2015, Kendziorski et al. 2012), it was hypothesized that these endpoints would be significantly
and adversely altered in both strains by BPA or EE exposure. Myometrial wall thickness in CD1
mice was not affected by BPA or EE exposure though. Significant age- and exposure-related
increases in distended glands and gland nests were observed in CD1 mice. It appeared that BPA
and EE slightly increased collagen accumulation in the endometrium of CD1 mice; however,
gland nest density was not increased in BPA or EE exposure groups. Significant increases in
macrophage accumulation in the CD1 uterus were observed in BPA and EE exposure groups.
In humans, BPA exposure is ubiquitous and begins in utero. The main route of BPA
exposure in humans occurs orally by ingesting contaminated food or beverages contained in
polycarbonate plastics or food cans with epoxy resin linings, with additional exposure occurring
through handling of thermal receipt paper, application of dental sealants, and transfer from
medical equipment (Ehrlich et al. 2014, Calafat et al. 2009, Carwile et al. 2009, Pulgar et al.
2000, Krishnan et al. 1993). Fetuses and newborns have indirect exposure via placental transfer
to amniotic fluid (Edlow et al. 2012, Nishikawa et al. 2010, Ikezuki et al. 2002, Yamada et al.
2002). Greater than 93% of humans have detectable urinary levels of BPA and metabolites
(Calafat et al. 2008).
Uterine weights for CD1 mice in this study were significantly increased in all EE
exposure groups, and CD1 mice in the 30 ppm BPA group had an increased uterine weight as
well. Vaginal opening (VO) and days to cornification were not altered by BPA or EE exposure
(Kendig et al. 2012). A prenatal or neonatal exposure to BPA led to morphological alterations in

142
epithelial cell proliferation or apoptosis and expression of the ERs and PR later in life (Vigezzi et
al. 2015, Mendoza-Rodriguez et al. 2011, Markey et al. 2005). Exposure to BPA and E2
increased myometrial thickness (Papaconstantinou et al. 2000), suggesting that proliferation and
growth of the myometrium are under estrogenic control. Increased incidence of cystic
endometrial hyperplasia, adenomyosis, and leiomyoma were found in CD1 mice prenatally
exposed to BPA (Newbold et al. 2009, 2007b). In the uterus of adult BPA-exposed mice, mice
display differences in the immune response between strains (Kendziorski et al. 2012), prompting
an examination of the effects of whole life exposure on the immune response in the uterus. Due
to the observed increase in leiomyoma and an equine endometrosis-like phenotype in BPA-
exposed mice, as well as significant alterations in collagen synthesis and degradation in the adult
uterus (Kendziorski and Belcher 2015, Newbold et al. 2007b), potential alterations in collagen
accumulation were also studied.

143
5.2 Methods
5.2.1 Exposure
As described in section 2.1.1 of Methods, mice were housed in a polycarbonate-free
caging system with excess estrogenic compounds controlled. Adult CD1 and C57Bl/6N mice
were divided equally between control, BPA, and EE groups and exposed through diet to known
concentrations of BPA (0.03, 0.3, 3, 30, and 300 ppm) or EE (0.0001, 0.001, and 0.01 ppm).
Offspring were exposed from gestation day 0 under either PND21, 49, or 90 either indirectly via
placental transfer or directly via diet (Figure 1). Females were sacrificed in estrus when possible
and tissues were collected for to examine morphology and potential pathologies.
Figure 1. Exposure paradigm for sires, dams, and offspring in control and BPA or EE exposure groups.
5.2.2 Myometrial Wall Thickness
Tissue preparation and H&E staining were described previously in sections 2.2.1 and
2.2.2 of Methods. Transverse sections of the H&E-stained uterine tube from CD1 offspring were
measured for myometrial wall thickness. Five random areas at 20x magnification of the outer
edge of the transverse section were imaged by an observer blinded to exposure group. Within
each image, five measurements from the beginning of the inner myometrium to the outer edge of
the outer myometrium were collected. Using a stage micrometer, the ratio of 60 µm/100 pixels

144
was measured. Wall thickness measurements were converted from pixels to micrometers (µm)
using the following formula:
X = 60 µm (pixel measurement) 100 pixels
5.2.3 Uterine Pathology
The H&E-stained uteri of C57Bl/6N and CD1 offspring (PND21, PND49, and ~PND90)
were assessed for distended glands, gland nests, and cysts. Distended glands were characterized
by a thinning of the glandular epithelial cells. Gland nests were characterized as groups of glands
that appear clumped together in an oval or circle surrounded by fibrotic stromal cells. Cysts
appeared as circular holes in the endometrium surrounded by a thin layer of epithelial cells. The
presence of the pathology and, when applicable, the number of incidences were noted for each
uterus. The observer was blinded to strain and exposure group.
5.2.4 Assessment of Collagen Accumulation and Immune Response
Picrosirious Red staining was used to visualize collagen accumulation in the uterus of
CD1 offspring, as described in section 2.3.1 of Methods. Immunohistochemistry using a rat
monoclonal anti-F4/80 (C1:A3-1; 4 µg/mL (1:250); ab6640, Abcam; Cambridge, MA) antibody
was performed to visualize and quantify the presence of macrophages in the uterus, as described
in section 2.3.2 of Methods. To quantify the number of F4/80-positive cells in the uterus, five
random images at 40x magnification of myometrium and five random images at 40x
magnification of endometrium in which mostly stroma was visible were collected. For each
image of endometrium and myometrium, the total number of positively stained cells and the total
number of stromal or smooth muscle cells were counted using Photoshop CC and ImagePro Plus
v4.5 (Media Cybernetics; Rockville, MD). The number of positively stained cells was then

145
divided by the total number of cells and multiplied by 100 to generate the percent of F4/80-
positive cells in each mouse.
5.2.5 Statistical Analysis
When applicable, the observer was blinded to strain and exposure group. The litter was
the statistical unit used; one animal was selected from each litter as representative of that litter.
Pathology was assessed with statistical differences analyzed using Fisher’s exact test or χ2 test
for trend when assessing across PND. Significance of differences in percentage of F4/80-positive
cells was determined after arc sin transformation. Myometrial wall thickness, gland nest density,
and percentage of F4/80-positive cell counts were analyzed using a one-way ANOVA and
Dunnett’s multiple comparison test. C57Bl/6N offspring endpoints were not statistically
analyzed due to a low sample size for all groups. Outliers were removed based on Grubb’s test.
Significance between differences in values was defined as p < 0.05. All data was analyzed using
GraphPad Prism® v5/v6 software.

146
5.3 Results
5.3.1 Uterine Morphology: Myometrial Wall Thickness
The myometrium is made up of two smooth muscle layers involved in protecting the
endometrium and supporting pregnancy and labor by inducing uterine contractions (Thompson
2015). Alterations in myometrial wall thickness may adversely impact these functions. To
determine if BPA or EE exposure affected myometrial thickness, the thickness of both outer and
inner layers was measured in CD1 mice. No differences in wall thickness were observed in BPA-
or EE-exposed CD1 offspring with a whole life exposure (Figure 2). The lack of differences in
myometrial thickness suggests that alterations in uterine morphology by BPA or EE begin at the
cellular level and gross differences are not observed until later in life.
Figure 2. Myometrial wall thickness in CD1 mice across exposure groups. Myometrial wall thickness for CD1 offspring is shown. Wall thickness measurements were collected as described in section 5.2.2. Values indicated are the mean wall thickness (µm) per group and error bars represent the SEM. n ≥ 3 animals. Wall thickness for BPA- and EE-exposed groups was compared separately to the control group. One-way ANOVA and Dunnett’s multiple comparison post hoc test was used to determine the level of statistical significance for BPA-exposed offspring and Student’s t test was used to determine the level of statistical significance in EE-exposed offspring. Statistical significance was set at p < 0.05.

147
5.3.2 Uterine Pathology
The presence of distended glands, cysts, and gland nests were noted in the uteri of CD1
mice at PND21, 49, and 90 (Table 1). The presence of gland nests in the uteri of C57Bl/6N
offspring is also summarized in Table 1. At PND21, no animals presented with cysts or gland
nests. Distended glands were absent from control animals and were observed at low frequency in
the 0.03, 30, and 300 ppm BPA groups and the 0.0001 and 0.01 ppm EE groups. At PND49, a
low frequency of mice with distended glands was observed in control and most BPA and EE
exposure groups. In the BPA groups, the number of mice with distended glands followed a non-
monotonic response curve, with the highest incidence at 3 ppm BPA (63%). Cysts were observed
in one CD1 mouse in the 0.3 ppm BPA group and the 0.001 ppm EE group. The frequency of
mice with gland nests in the 0.03 ppm BPA exposure group (75%) significantly increased
compared to the control group (22%; p = 0.0445). All other BPA exposure groups and the 0.0001
and 0.001 ppm EE exposure groups appeared to have a higher frequency of mice with gland
nests compared to the control group as well.
Figure 3 shows representative images of uterine morphology for control (Figure 3A), 30
ppm BPA (Figure 3D), and 0.01 ppm EE (Figure 3G). At PND90, the incidence of mice with
distended glands was high in control (70%) and all BPA (50-85%) and EE (44-75%) groups. At
least one mouse in each BPA or EE group developed a cyst, except for 300 ppm BPA and 0.001
ppm EE (frequency <20%). Gland nests were present in control (20%) and all BPA and EE
groups, with the number of mice at or above 50% in the 0.3 and 300 ppm BPA exposure groups.
In the C57Bl/6N strain at PND90, gland nests were present in almost every animal. These results
provide insight into experiments related to collagen accumulation and normal uterine aging.

148
When examining trends across PND in the CD1 strain, the number of mice with
distended glands significantly increased over time in the control (p = 0.0046), 0.03 ppm BPA (p
= 0.0287), 3 ppm BPA (p = 0.0208), 30 ppm BPA (p = 0.0287), 300 ppm BPA (p = 0.0025),
0.0001 ppm EE (p = 0.0273), and 0.001 ppm EE (p = 0.0068) groups. The time-dependent
observed increase in distended glands serves as an internal control due to the increase being well
known (Chapin et al. 2008). The presence of dilated or distended glands also regularly fluctuates
throughout the menstrual or estrous cycle (Rubin and Strayer 2012). No trends were found for
cyst formation over time. The number of mice presenting with gland nests clearly increased over
time, especially between PND21 and PND49. However, between PND49 and PND90, the
number of mice with gland nests did not increase further. Some exposure groups even had a
decrease in the number of mice with gland nests, the most drastic change being in the 0.03 ppm
BPA group (75% at PND49 vs 27% at PND90). In the 30 ppm BPA group, trends for increasing
the number of mice with gland nests over time approached significance (p = 0.0566).
5.3.3 Collagen Accumulation
A slightly increased intensity and dispersal of red/yellow birefringence, indicative of
increased collagen fiber thickness and density, was associated with stromal cells in the BPA-
exposed CD1 offspring (Figure 3F) compared to control (Figure 3C). This increase in stromal
cell-associated red/yellow birefringence was also observed in EE-exposed CD1 offspring, as well
as a slight increase in periglandular-associated green birefringence, suggestive of less densely
packed periglandular collagen (Figure 3I). The observed increase was much lower than that
observed in the adulthood exposure model though (Kendziorski and Belcher 2015).

149
Table 1. Uterine Pathology in CD1 and C57Bl/6N Offspring BPA (ppm) EE (ppm)
PND21 CD1 0 0.03 0.3 3 30 300 0.0001 0.001 0.01
n= 5 6 5 7 7 5 5 6 5
Distended Glands 0 (0%) 1 (17%) 0 (0%) 0 (0%) 1 (14%) 1 (20%) 1 (20%) 0 (0%) 2 (40%)
Cysts 0 (0%) 0 (0%) 0 (0%) 0 (0%) 0 (0%) 0 (0%) 0 (0%) 0 (0%) 0 (0%)
Gland Nests 0 (0%) 0 (0%) 0 (0%) 0 (0%) 0 (0%) 0 (0%) 0 (0%) 0 (0%) 0 (0%)
PND49 CD1
n= 9 8 11 8 5 10 10 5 7
Distended Glands 2 (22%) 1 (13%) 4 (36%) 5 (63%) 2 (40%) 2 (20%) 2 (20%) 1 (20%) 2 (29%)
Cysts 0 (0%) 0 (0%) 1 (9%) 0 (0%) 0 (0%) 0 (0%) 0 (0%) 1 (20%) 0 (0%)
Gland Nests 2 (22%) 6 (75%)a 6 (55%) 5 (63%) 2 (40%) 6 (60%) 5 (50%) 2 (40%) 1 (14%)
PND90 CD1
n= 10 11 12 10 14 13 8 11 9 Distended Glands 7 (70%)b 7 (64%)b 6 (50%) 6 (60%)b 9 (64%)b 11 (85%)b 6 (75%)b 7 (64%)b 4 (44%)
Cysts 0 (0%) 2 (18%) 1 (8%) 1 (10%) 2 (14%) 0 (0%) 1 (13%) 0 (0%) 1 (11%)
Gland Nests 2 (20%) 3 (27%) 6 (50%) 3 (30%) 6 (43%)c 7 (54%) 3 (38%) 4 (36%) 3 (33%)
PND90 C57Bl/6N
n= 2 2 3 2 2 1 0 1 1
Gland Nests 1 (50%) 2 (100%) 3 (100%) 1 (50%) 2 (100%) 1 (100%) 0 0 (0%) 1 (100%) a p=0.0445 by one-tailed Fisher's exact test b p<0.05 by χ2 test for trend across all PNDs for increases in distended glands c p=0.0566 χ2 test for trend across all PNDs for increases in gland nests

150
Figure 3. Effects of oral BPA or EE exposure on uterine morphology and collagen accumulation in CD1 offspring at PND90. Shown are representative photomicrographs of H&E- and Picrosirius Red-stained uterine sections from a control CD1 mouse (A-C) and from a mouse exposed to 30 ppm BPA (D-F) or 0.01 ppm EE (G-I). Similar uterine locations are highlighted in each panel. Arrows indicate gland nest structures. Scale bars represent 50 µm.
5.3.4 Quantification of Gland Nest Density
Quantification of gland nest density in the endometrium of both strains at PND90 was
performed (Figure 4). In the C57Bl/6N strain, the mean density of the control group (M = 1.138
± 1.138) was ~37 fold greater than the mean density of the control group in the CD1 strain (M =
0.031 ± 0.020). However, due to the low sample number in the C57Bl/6N strain, the results
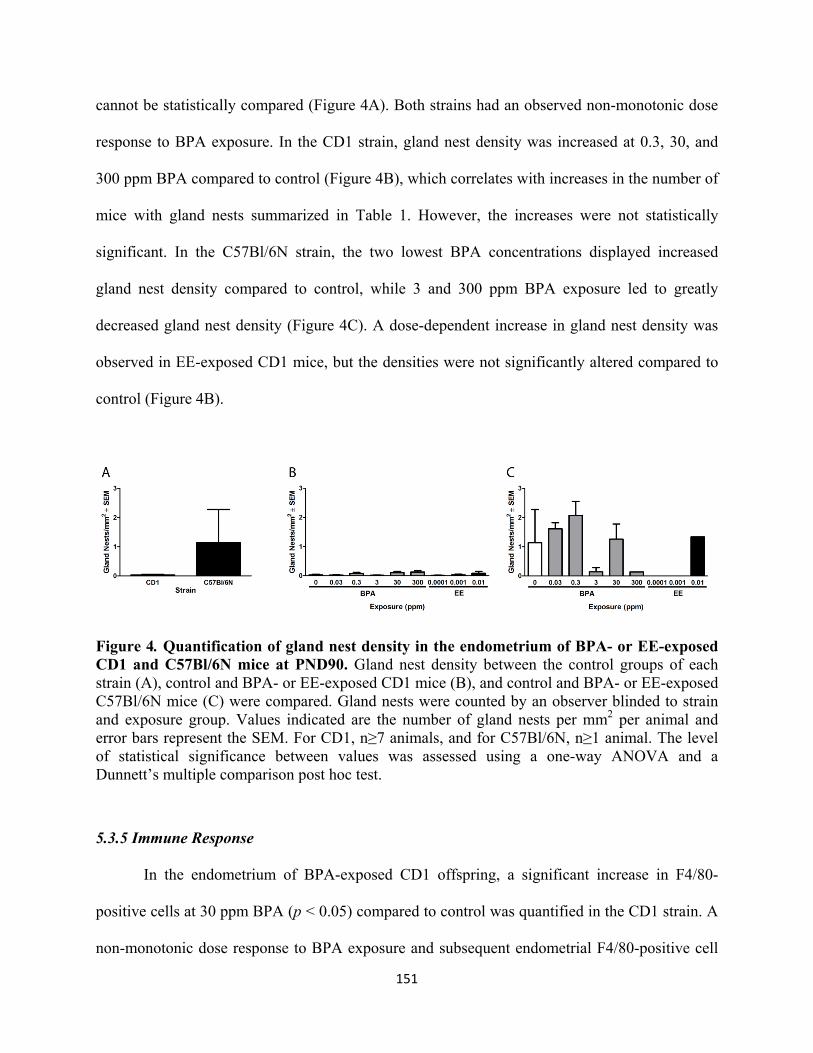
151
cannot be statistically compared (Figure 4A). Both strains had an observed non-monotonic dose
response to BPA exposure. In the CD1 strain, gland nest density was increased at 0.3, 30, and
300 ppm BPA compared to control (Figure 4B), which correlates with increases in the number of
mice with gland nests summarized in Table 1. However, the increases were not statistically
significant. In the C57Bl/6N strain, the two lowest BPA concentrations displayed increased
gland nest density compared to control, while 3 and 300 ppm BPA exposure led to greatly
decreased gland nest density (Figure 4C). A dose-dependent increase in gland nest density was
observed in EE-exposed CD1 mice, but the densities were not significantly altered compared to
control (Figure 4B).
Figure 4. Quantification of gland nest density in the endometrium of BPA- or EE-exposed CD1 and C57Bl/6N mice at PND90. Gland nest density between the control groups of each strain (A), control and BPA- or EE-exposed CD1 mice (B), and control and BPA- or EE-exposed C57Bl/6N mice (C) were compared. Gland nests were counted by an observer blinded to strain and exposure group. Values indicated are the number of gland nests per mm2 per animal and error bars represent the SEM. For CD1, n≥7 animals, and for C57Bl/6N, n≥1 animal. The level of statistical significance between values was assessed using a one-way ANOVA and a Dunnett’s multiple comparison post hoc test.
5.3.5 Immune Response
In the endometrium of BPA-exposed CD1 offspring, a significant increase in F4/80-
positive cells at 30 ppm BPA (p < 0.05) compared to control was quantified in the CD1 strain. A
non-monotonic dose response to BPA exposure and subsequent endometrial F4/80-positive cell

152
counts was also present (Figure 5A). No significant differences were observed in the
myometrium of BPA-exposed offspring (Figure 5B). When comparing the endometrial and
myometrial F4/80-positive cell counts, the 30 ppm BPA exposure group was significantly
different between the two tissue types (p < 0.05; Figure 5). In the endometrium of EE-exposed
offspring, the 0.0001 ppm EE group had a significant increase in F4/80-positive cells (p < 0.013)
compared to control in CD1 offspring (Figure 5A). Similar patterns of F4/80-positive cell counts
in the endometrium of EE-exposed offspring were observed in the myometrium of EE-exposed
offspring (Figure 5B). The 0.0001 ppm EE exposure group had a significant increase in F4/80-
positive cells (p < 0.01) compared to control in CD1 offspring (Figure 5B). The 0.001 and 0.01
ppm EE-exposed CD1 offspring appeared to have decreases in the percent of F4/80-positive cells
compared to the control group.
Figure 5. Quantification of F4/80-positive cells in BPA- or EE-exposed CD1 offspring. In CD1 offspring, endometrial (A) or myometrial (B) F4/80-positive cells were counted in BPA- (gray bars) or EE-exposed (black bars) mice. Endometrial and myometrial F4/80-positive cells were quantified separately and normalized to either stromal or smooth muscle cells respectively. F4/80-positive and total cells were counted by an observer blinded to exposure group. Values indicated are the percent of F4/80-positive cells per animal and error bars represent the SEM. n ≥ 4 animals. The level of statistical significance between values was assessed using a one-way ANOVA and Dunnett’s post hoc test. * indicates a level of difference between values is significant (p<0.05); ** indicates a level of difference between values is very significant (p<0.01).

153
5.4 Discussion
Bisphenol A is a compound with known endocrine disrupting activity in the uterus.
Through studies on adult CD1 and C57Bl/6N mice, differences in responses between mouse
strains to BPA exposure in regards to development of pyometra and an equine endometrosis-like
phenotype, collagen dysregulation, and immune response were reported (Kendziorski and
Belcher 2015, Kendziorski et al. 2012). In this study, the development of various pathologies
including an equine endometrosis-like phenotype, collagen accumulation, and immune response
to whole life BPA exposure in the offspring of CD1 mice were examined. While C57Bl/6N
offspring were examined for pathology and gland nest density, a small sample size prevented
analysis.
The myometrium is made of distinct outer and inner smooth muscle layers that provide
support for the uterus during pregnancy and parturition. During development, the layers can be
seen as early as PND10 in mice, with the adult configuration observed around PND15. By
PND42, the entire uterus has grown in size and complexity but retains its configuration
(Mehasseb et al. 2009). Treatment or exposure to estrogenic compounds has varying effects on
alterations in myometrial development and thickness. In young adult rats, myometrial thickness
is increased 24 hours after one dose of E2 (Kang et al. 1975), similar to results from E2-treated
or BPA-exposed ovariectomized adult mice (Papaconstantinou et al. 2000). However, prenatal
BPA exposure decreases the volume of myometrium (Markey et al. 2005) or perinatal estradiol
benzoate treatment speeds up uterine development without structural alterations (Mehasseb et al.
2009). These studies indicate that the developmental time point at which exposure occurs is
important for the end result. In this study, whole life BPA or EE exposure had no effect on
myometrial thickness. While morphology and pathology of the endometrium were examined at

154
earlier time points (PND21 and 42), normal myometrial structure was not investigated. It is
possible that myometrial development was sped up, as was observed in Mehasseb et al. (2009).
However, even earlier time points than PND21 are necessary to examine to determine if
development did occur at an earlier time point.
Previous studies examining pathology in the rodent uterus prenatally or neonatally
exposed to BPA assess the tissue at 16-18 months of age (Newbold et al. 2009, 2007b). This
study assessed pathology at much younger ages, PND21, 49, and 90. As expected, few
pathological alterations were found at PND21 because endometrial development does not finish
until ~PND15 (Cooke et al. 2013). However, at PND49, at least one CD1 mouse presented with
distended glands or gland nests in control and all BPA or EE exposure groups. The frequency of
CD1 mice with distended glands further increased at PND90, suggesting that this pathological
alteration is a natural occurrence of development, aging, and cyclicity (Rubin and Strayer 2012).
The frequency of CD1 mice with gland nests remained fairly constant between PND49 and 90
and did not further increase with BPA or EE exposure, potentially indicative of gland nest
formation also being a natural occurrence of development. However, when gland nest density
was quantified, the occurrence of gland nests was almost negligible in the CD1 strain in all
exposure groups. The increased frequency of mice with gland nests was more than likely not
physiologically relevant. Offspring in the C57Bl/6N strain had a higher gland nest density than
the CD1 strain, similar to what was observed in the adult exposure model (Kendziorski and
Belcher 2015). When assessing collagen accumulation in the CD1 strain, BPA or EE exposure
did appear to shift stromal collagen to a more sheet-like pattern and a thicker fibril, while
periglandular collagen was largely unchanged. Because transcription of collagen types is
estrogenically regulated (Frankel et al. 1988, Dyer et al. 1980), BPA or EE may be increasing

155
transcription and leading to increased collagen accumulation without a high incidence of gland
nests.
In both the whole life and adulthood exposures, the number of F4/80-positive cells
significantly increased in the 30 ppm BPA exposure group of CD1 mice (Figure 6 in Chapter 3),
suggesting that time of exposure does not matter in regards to alterations in immune
responsiveness in the uterus. However, other endpoints examined, including collagen
accumulation, gland nest density, and pathology, were not significantly altered in the whole life
exposure model as they were in the adulthood exposure model. Other than time of exposure, the
main difference is the mating, pregnancy, and parturition that occurred in the adult exposure
model. This difference between exposure models highlights the lack of understanding of the
relationship between estrogenic EDC exposure and potential alterations in the response to drastic
changes in the uterine environment that occur during mating and pregnancy. The presence of
semen or sperm induces an inflammatory response in the uterus (Bromfield 2014, Robertson et
al. 2013, Schuberth et al. 2008, Sharkey et al. 2007); however, it is unknown if the presence of
semen alters collagen regulation. Regulation of collagen and other ECM proteins is significantly
altered during pregnancy though (Tsai et al. 2009, Carbone et al. 2006, Bjorn et al. 1997,
Iwahashi et al. 1996), so the combination of these events may be important for creating an
environment sensitive to potential long-term endocrine disrupting effects.

156
CHAPTER 6
SUMMARY AND CONCLUSIONS

157
6.1 Significance
This dissertation research highlighted the impact of BPA and EE exposure on alterations
in immune responsiveness and fibrosis, along with the development of uterine pathologies such
as pyometra and an equine endometrosis-like phenotype (Kendziorski and Belcher 2015,
Kendziorski et al. 2012). Changes in immune responsiveness and ECM regulation are also
components of human uterine pathologies, including leiomyoma and endometriosis, and
important factors in infertility, suggesting that exposure to estrogenic EDCs is a contributing
factor to the development of uterine diseases and reproductive dysfunction.
Pyometra is a uterine disorder caused in part by hormonal factors. Exposure to estrogenic
compounds such as E2 and DES led to the development of this disease (Brossia et al. 2009,
Gould et al. 2005, Pandey et al. 2005). This dissertation research was the first report of BPA
exposure leading to the development of pyometra in any animal model (Kendziorski et al. 2012),
supporting that BPA acts as an estrogenic EDC in the uterus. In this study, the oral exposure in
intact female mice can be translated to humans due to the relevant route of exposure and dosing.
A second aspect to the studies presented in this dissertation was related to susceptibility to
pyometra development. Only BPA- and EE-exposed C57Bl/6N mice developed the pathology,
suggesting that genetic variation in humans could also impact sensitivity to estrogenic EDCs and
subsequent development of uterine pathologies. Genome regions involved in susceptibility to E2-
or DES-induced pyometra were known (Gould et al. 2005, Pandey et al. 2005), suggesting a
QTL involved in BPA-induced pyometra may be regulating susceptibility as well.
The observed alteration in immune responsiveness in both the adulthood and whole life
exposure model suggests that BPA exposure may have a detrimental effect on the immune-
related functions in fertility. Successful implantation depends greatly on the contributions of the

158
varying immune cells present in the uterus (Gnainsky et al. 2014). Aberrant recruitment and
presence of immune cells in the uterus that occurs with BPA exposure may significantly reduce
the probability of successful implantation. Increased immune cell infiltration and macrophage
activation are also highly involved in the growth of endometriotic lesions (Ahn et al. 2015),
suggesting that BPA exposure can play a role in the progression and severity of uterine disorders
based on altering the immune environment in reproductive tissue.
The results presented in this dissertation highlighted that BPA exposure in adulthood led
to increased collagen accumulation in the uterus of mice due to dysregulation of collagen
synthesis and degradation. In adult CD1 mice, BPA directly increased collagen I and III
expression via transcriptional activation and directly and indirectly decreased MMP2, MMP14,
and TIMP2 expression and MMP2 activity. Exposure groups with observed increases in stromal
and periglandular collagen also had increases in gland nest density, another hallmark of equine
endometrosis (Kendziorski and Belcher 2015). These findings suggest that BPA exposure factors
in the progression of fibrotic human and veterinary uterine diseases by altering collagen
synthesis and degradation, leading to the observed fibrotic phenotype. Estrogens increased the
transcription of collagens and MMPs during the physiological cycling of sex hormones (Gaide
Chevronnay et al. 2012, Frankel et al. 1988). Exposure to BPA led to further transcription of
collagens, increasing the overall levels of collagen. However, BPA exposure did not increase
MMP expression levels, as would be expected with E2 treatment, suggesting that BPA acted
through a different mechanism than E2 to alter collagen degradation. The combination of
increased collagen synthesis and decreased collagen degradation led to fibrosis in the uterus,
potentially increasing the severity of present uterine diseases as well.

159
Although collagen synthesis and degradation in the uterus are under estrogenic control
(Gaide Chevronnay et al. 2012, Henriet et al. 2012), no previous studies had been conducted
comparing and contrasting the differences in collagen accumulation between different mouse
strains. Due to the prevalence of gland nest structures and periglandular fibrosis in the C57Bl/6N
uterus, part of this dissertation research examined the differences in collagen accumulation
between the C57Bl/6N and CD1 strains (Kendziorski and Belcher 2015). Differences in
expression of proteins involved in regulating collagen accumulation between the strains are
summarized in Table 1. There was a general decrease in MMPs in the C57Bl/6N strain, leading
to an overall increase in collagen accumulation.
Table 1. Comparison of Relative Expression and Activity of Collagens and MMPs in Mouse Strains
Expression Activity Collagen proMMP2 MMP2 proMMP14 MMP14 proMMP2 MMP2
CD1 ‐ − − − − − −
C57Bl/6N ↑ ↓ ↓ ↓ ↓ ↓ ↓
Collagen dysregulation combined with increases in gland nest density are characteristics
of equine endometrosis (Evans et al. 1998, Kenney 1978). Further characterization of the stromal
fibroblasts surrounding gland nests in the equine endometrosis-like phenotype resulted in an
increase in F4/80 staining, but ERα and α-SMA staining in the stroma and glandular epithelium
were unaffected (Kendziorski and Belcher 2015, Hoffmann et al. 2009b, Walter et al. 2001). The
endometrosis-like phenotype observed in the C57Bl/6N strain appeared to be an early stage of
the progressive pathology, suggesting that aging and repeated mating cycles may lead to more of
the destructive degenerative phenotypes observed in equine populations. Under control
conditions, the C57Bl/6N mouse may be useful as an experimental model to study the etiology,
progression, and treatment of equine endometrosis.

160
When comparing the two exposure models, mice with a whole life exposure to BPA or
EE did not develop significant alterations in collagen accumulation, increased gland nest density,
or pathology. Other than exposure, distinguishing characteristics were mating, gravid status, and
parturition, with mice in the adulthood exposure having delivered one litter and mice in the
whole life exposure model never mating and were nulligravid. This difference between exposure
models highlights the lack of understanding of the relationship between estrogenic EDC
exposure and potential alterations in the uterine environment that occur during mating and
pregnancy.
6.2 Limitations
While each technique is not without its limitations, the quantification of F4/80-positive
cells could have been performed differently using flow cytometry or more specific antibodies for
in situ hybridization. These techniques would have allowed for distinguishing between M1 and
M2 macrophages, which are differentially activated by various stimuli and have varying roles in
the periphery and tissues (Martinez and Gordon 2014). The techniques could also have been used
to characterize the presence and abundance of other immune cells, such as leukocytes,
neutrophils, and B and T cells. This knowledge would allow for a better understanding of the
specific types of immune cells regulated by BPA or EE exposure and potentially identify early
stages of immune alterations before the induction of pyometra. However, the choice to use a
general macrophage marker (F4/80) was made to examine an initial general immune response.
Further characterization of the specific immune cells involved in this response could be
performed using more sophisticated techniques, such as flow cytometry.
Loss of tissue integrity was another limitation to these studies that needs to be
considered. Depth in the uterine tissue was an important variable that needed to be controlled and

161
known. When tissue sectioning, the depth was recorded and sections were collected at a similar
level in the uterus for all animals. However, it was possible that there was still variability
between samples, which could play a role in differences observed between samples. Loss of
tissue integrity was also an important factor in RNA isolation and protein homogenization. While
temperature of the tissue was kept as cold as possible, activity of degrading enzymes could still
have affected integrity of the tissue, leading to potential decreases in the amount of RNA or
protein isolated.
While the endometrium, specifically stromal cells surrounding glandular epithelium, was
the main localized site of interest in the uterus, both the myometrium and endometrium were
contained in protein homogenates used to quantify protein expression and activity. Laser capture
microdissection would be an appropriate technique to isolate the desired cell types of interest.
However, the significant decreases in MMP protein expression and activity observed in BPA-
exposed CD1 mice or control C57Bl/6N mice compared to control CD1 mice suggested that the
decreases in these proteins was so robust between the groups that it was unnecessary to isolate
specific cell types. Also, immunolocalization of MMP2 and MMP14 supported that expression
of these proteins is decreased in periglandular stromal cells.
6.3 Future Directions
With these novel findings regarding immune responsiveness and the development of
immune-related uterine pathology though come new questions regarding the impact of BPA
exposure and genetic variation in these processes. While it was known that estrogenic
compounds, now including BPA, can lead to the induction of pyometra, specific genes and
proteins involved in its etiology are still partially unknown. As described, mapping of
overlapping QTLs named Eutr1 and Eutr2, which include large chromosomal areas containing

162
several genes, has been performed for E2- and DES-induced pyometra in rat strains (Gould et al.
2005, Pandey et al. 2005). A third QTL in mice involved in eosinophil infiltration, Est1 (Griffith
et al. 1997), also overlaps with Eutr1 and Eutr2. This overlapping region between three QTL
may contain genes involved in the development of pyometra. There are four genes in the
overlapping region, those encoding IL11Rα, Ccl19, Ccl21a, and Ccl21c, all of which are
involved in the immune system. The impact of exposure to EDCs on expression of these genes
and translated proteins need to be characterized to determine the role of these genes of interest in
immune responsiveness.
Of these four genes, only IL11Rα has been extensively studied in the uterus. In human
endometrium, IL11Rα was found in both the stroma and the apical surface of epithelial cells
(Karpovich et al. 2003) and was expressed in decidual tissue in the rat uterus peri-implantation
(Li et al. 2001) and glandular epithelial cells in canines during pregnancy (Guo et al. 2009). It
was important for proper decidualization and placentation in humans and animals (Menkhorst et
al. 2009, Paiva et al. 2009, Karpovich et al. 2005, Ain et al. 2004, Dimitriadis et al. 2003,
Bilinski et al. 1998, Robb et al. 1998). In women with infertility or recurrent miscarriages, there
were significant decreases in IL-11, the ligand for IL11Rα (Dimitriadis et al. 2007, Linjawi et al.
2004), while women with endometriosis and associated infertility had an absence of IL11Rα
staining in the glandular epithelium (Dimitriadis et al. 2006). Increased IL11Rα staining was also
found in human endometrial carcinoma samples, suggesting that aberrant signaling may play a
role in progression of the disease (Yap et al. 2010). It is currently unknown if IL11Rα is involved
in the induction of pyometra though or whether exposures to estrogens or estrogenic compounds
such as BPA directly or indirectly interact with the receptor. Located within the promoter are
three EREs to which an activated ERα dimer could bind, leading to potential activation or

163
repression of the receptor or its downstream targets (SABiosciences 2012). To determine how
estrogenic EDCs, including BPA and EE, affect Il11ra expression, targeted ChIP assays could be
used. Also, sequencing of the Il11ra promoter can identify single nucleotide polymorphisms
(SNPs) that can impact differences in gene expression between the CD1 and C57Bl/6N strains
and aids in explaining the sensitivity between strains to estrogenic compounds. The sensitivity
between strains can be correlated to differences in sensitivities between humans to estrogenic
EDCs. Identification of SNPs can also aid in defining the etiology of pyometra and lead to the
development of a treatment for the disease.
Research for this dissertation determined that BPA exposure altered collagen
accumulation by increasing collagen synthesis and decreasing collagen degradation, leading to
an overall fibrotic phenotype. However, the mechanism of action of BPA that led to the
alterations in collagen synthesis and degradation remains unknown. Under physiological
conditions, E2 activated ERα and led to transcriptional upregulation of collagens and MMPs
(Gaide Chevronnay et al. 2012, Frankel et al. 1988). Exposure to BPA also increased
transcription of collagens, suggesting that BPA was also activating ERα to upregulate collagen
synthesis. However, the effects of BPA and E2 on MMPs and proteins involved in collagen
degradation were opposite, suggesting a different mechanism of action of BPA compared to E2.
One possibility is that BPA may be acting as a partial or inverse agonist on ERα or be
antagonizing the effects of E2. A partial agonist is a ligand that binds to a receptor at the active
site but produces a submaximal response (Golan et al. 2012). An inverse agonist is a ligand that
binds to the same receptor target as an agonist but the induced pharmacological response is in the
opposite direction (Golan et al. 2012). It is possible that BPA was still binding to ERα but had a
decreased efficacy compared to E2, leading to decreased expression and activity of MMPs.

164
Partial agonist-like and antagonist activity of BPA at ERα in the uterus has been reported. For
example, BPA increased PR expression in the immature rat uterus, but also antagonized E2-
induced stimulation in PR expression (Gould et al. 1998).
A second possibility is that BPA was binding to another receptor target altogether. At nM
concentrations, BPA bound to and activated ERα and ERβ (Li et al. 2013, Wetherill et al. 2007,
Matthews et al. 2001, Kuiper et al. 1998). However, BPA also had strong binding to ERRγ and
GPR30 at similar concentrations (Okada et al. 2008, Thomas and Dong 2006). Other receptor
targets for BPA included AR and THR, at which BPA bound and inhibited at µM concentrations
(Teng et al. 2013, Moriyama et al. 2002, Paris et al. 2002, Sohoni and Sumpter 1998). Binding of
BPA to any of these receptors could potentially elicit the observed alterations in collagen
degradation and decreased expression and activity of MMPs. To determine this, a luciferase
assay containing the MMP2 promoter could be used. Treatment with BPA or E2 would help
answer which receptor BPA acts through to alter collagen degradation. ChIP experiments can
also be performed to determine if ERα or other receptor targets of BPA are bound to the
promoters of Mmp genes. These experiments would help to narrow down through which receptor
BPA is acting to induce the observed alterations in collagen degradation. However, translocation
and signaling of nuclear receptors also relies on the recruitment of coactivators or corepressors.
Experiments designed to identify the specific coregulators involved in Mmp transcription would
greatly expand knowledge on the differential activation of these genes by BPA.
The mechanisms behind gland nest formation and progression of gland nest severity is an
aspect of equine endometrosis that needs to be further explored. Because equine endometrosis is
irreversible, understanding the etiology and progression could help to develop treatments to
either prevent or halt progression. The mare would then have a better chance of carrying a fetus

165
to term. To the best of knowledge, it is unknown if gland nest formation occurs in humans too.
However, gland nest formation and associated periglandular fibrosis could also play a role in
infertility in humans. Again, understanding the etiology of periglandular fibrosis may lead to
treatments for fibrosis in the human uterus and increased fertility. Because the C57Bl/6N strain
already exhibits increased incidence of gland nests and stromal and periglandular fibrosis, this
strain could potentially be a useful small animal experimental model to study the etiology and
progression of equine endometrosis.
A third future direction for this dissertation research would be to determine the effects of
alterations in collagen accumulation on uterine physiology. In particular, it would be important
to examine fertility, including decidualization, implantation, uterine stretch, and parturition.
While collagen fibrils are involved in decidualization and MMPs degrade these fibrils during
implantation (Carbone et al. 2006, Staun-Ram and Shalev 2005, Dziadek et al. 1995,
Abrahamsohn and Zorn 1993), the increased expression of collagens and the decreased
expression and activity of MMPs observed in this research could significantly decrease fertility
due to the inability of the uterus to undergo decidualization and implantation. Alterations in
collagen accumulation could have a significant detrimental impact on fertility and may help to
explain the increases in infertility in women with fibrotic uterine diseases.
When comparing CD1 mice in the whole life and adult exposure models, there was a
clear lack of effect on the development of fibrotic uterine diseases in the whole life exposure
model, as well as only a slight increase in collagen accumulation. However, the 30 ppm BPA-
exposed CD1 group in both models was shown to have increases in immune responsiveness
(Kendziorski and Belcher 2015). These findings suggest that physiological changes in the uterine

166
environment during mating, pregnancy and parturition are important for the development of
fibrotic uterine pathologies and should be further investigated.

167
LIST OF PUBLICATIONS AND ABSTRACTS
Publications
JA Kendziorski and SM Belcher. 2015. Effects of whole life exposure to bisphenol A or 17α- ethinyl estradiol in uterus of nulligravida CD1 mice. Data in Brief.
JA Kendziorski and SM Belcher. 2015. Strain-specific induction of endometrial periglandular fibrosis in mice exposed during adulthood to the endocrine disrupting chemical bisphenol A. Reprod Toxicol, 58: 119-130. doi: 10.1016/j.reprotox.2015.08.001.
EL Kendig, DR Buesing, SM Christie, CJ Cookman, RB Gear, ER Hugo, JA Kendziorski, KR Ungi, K Williams, SN Kasper, and SM Belcher. 2012. Estrogen-like disruptive effects of dietary exposure to bisphenol A or 17α-ethinyl estradiol in CD1 mice. Int J Toxicol, 31(6): 537-50. doi: 10.1177/1091581812463254
JA Kendziorski, EL Kendig, RB Gear, and SM Belcher. 2012. Strain specific induction of pyometra and differences in immune responsiveness in mice exposed to 17α-ethinyl estradiol or the endocrine disrupting chemical bisphenol A. Reprod Toxicol, 34(1): 22-30, doi: 10.1016/j.reprotox.2012.03.001
Abstracts
J Kendziorski and S Belcher. Strain-Specific Induction of Endometrial Periglandular Fibrosis in Mice Exposed During Adulthood to the Endocrine Disrupting Chemical Bisphenol A. Oral Presentation. 49th Annual Albert J. Ryan Fellowship Symposium. North Conway, NH. May 15, 2015.
J Kendziorski and S Belcher. Exposure to Bisphenol A Increases Gland Nest Formation and Fibrosis in the Murine Uterus. College of Medicine Graduate Student Poster Forum. University of Cincinnati, Cincinnati, OH. March 6, 2015.
J Kendziorski and S Belcher. Exposure to Bisphenol A Increases Gland Nest Formation and Fibrosis in the Murine Uterus. Graduate Student Expo and Poster Forum. University of Cincinnati, Cincinnati, OH. March 6, 2015.
J Kendziorski and S Belcher. Sensitivity to estrogenic actions of bisphenol A in the uterus of two mouse strains. Oral presentation. 48th Annual Albert J. Ryan Fellowship Symposium. North Conway, NH. May 17, 2014.
J Kendziorski and S Belcher. Bisphenol A blocks the adverse growth effects of in utero arsenic exposure. Poster. University of Cincinnati Graduate Student Poster Forum. Cincinnati, OH. February 28, 2014.

168
J Kendziorski, E Kendig, R Gear, and S Belcher. Dietary bisphenol A increases the uterine immune response and induces pyometra in mice. Poster. NIEHS BPA Grantee Research Meeting. NIEHS Research Triangle Park, NC. January 28-29, 2013.
J Kendziorski, E Kendig, R Gear, and S Belcher. Dietary bisphenol A increases the uterine immune response and pyometra in mice. Oral presentation. Society of Toxicology 2012 Annual Meeting. San Francisco, CA. March 11-15, 2012.
E Kendig, C Lo, C Cookman, S Christie, D Buesing, R Gear, J Kendziorski, K Ungi, and S Belcher. The effects of oral Bisphenol A exposure in the developing CD-1 mouse. Poster. ENDO 2011: The Endocrine Society’s 93rd Annual Meeting and Expo. Boston, MA. June 4- 7, 2011.

169
BIBLIOGRAPHY
1982. "Carcinogenesis Bioassay of Bisphenol A (CAS No. 80-05-7) in F344 Rats and B6C3F1 Mice (Feed Study)." Natl Toxicol Program Tech Rep Ser 215:1-116.
Abrahamsohn, PA, and TM Zorn. 1993. "Implantation and decidualization in rodents." J Exp Zool 266 (6):603-628.
Acconcia, F, P Ascenzi, A Bocedi, E Spisni, V Tomasi, A Trentalance, P Visca, and M Marino. 2005. "Palmitoylation-dependent estrogen receptor alpha membrane localization: regulation by 17beta-estradiol." Mol Biol Cell 16 (1):231-7. doi: 10.1091/mbc.E04-07-0547.
Acconcia, F, P Ascenzi, G Fabozzi, P Visca, and M Marino. 2004. "S-palmitoylation modulates human estrogen receptor-alpha functions." Biochem Biophys Res Commun 316 (3):878-83. doi: 10.1016/j.bbrc.2004.02.129.
Adachi, M, K Nasu, A Tsuno, A Yuge, Y Kawano, and H Narahara. 2011. "Attachment to extracellular matrices is enhanced in human endometriotic stromal cells: a possible mechanism underlying the pathogenesis of endometriosis." Eur J Obstet Gynecol Reprod Biol 155 (1):85-8. doi: 10.1016/j.ejogrb.2010.10.026.
Adlanmerini, M, R Solinhac, A Abot, A Fabre, I Raymond-Letron, AL Guihot, F Boudou, L Sautier, E Vessieres, SH Kim, P Liere, C Fontaine, A Krust, P Chambon, JA Katzenellenbogen, P Gourdy, PW Shaul, D Henrion, JF Arnal, and F Lenfant. 2014. "Mutation of the palmitoylation site of estrogen receptor alpha in vivo reveals tissue-specific roles for membrane versus nuclear actions." Proc Natl Acad Sci U S A 111 (2):E283-90. doi: 10.1073/pnas.1322057111.
Aggarwal, S, and MF Pittenger. 2005. "Human mesenchymal stem cells modulate allogeneic immune cell responses." Blood 105:1815-1822. doi: 10.1182/blood-2004-041559.
Agostinho, JM, A de Souza, RP Schocken-Iturrino, LG Beraldo, CA Borges, FA Avila, and JM Marin. 2014. "Escherichia coli Strains Isolated from the Uteri Horn, Mouth, and Rectum of Bitches Suffering from Pyometra: Virulence Factors, Antimicrobial Susceptibilities, and Clonal Relationships among Strains." Int J Microbiol 2014:979584. doi: 10.1155/2014/979584.
Ahn, SH, SP Monsanto, C Miller, SS Singh, R Thomas, and C Tayade. 2015. "Pathophysiology and Immune Dysfunction in Endometriosis." Biomed Res Int 2015:795976. doi: 10.1155/2015/795976.
Aiello, SE, ed. 2014. The Merck Veterinary Manual. Edited by R Tremblay, Management and Nutrition.

170
Ain, R, ML Trinh, and MJ Soares. 2004. "Interleukin-11 signaling is required for the differentiation of natural killer cells at the maternal-fetal interface." Dev Dyn 231 (4):700-8. doi: 10.1002/dvdy.20183.
Allen, WR. 1993. "Proceedings of the John P. Hughes International Workshop on Equine Endometritis. Davis, California, August 1992." Equine Vet J 25 (3):184-193.
An, BS, HJ Ahn, HS Kang, EM Jung, H Yang, EJ Hong, and EB Jeung. 2013. "Effects of estrogen and estrogenic compounds, 4-tert-octylphenol, and bisphenol A on the uterine contraction and contraction-associated proteins in rats." Mol Cell Endocrinol 375 (1-2):27-34. doi: 10.1016/j.mce.2013.04.025.
Aplin, JD, AK Charlton, and S Ayad. 1988. "An immunohistochemical study of human endometrial extracellular matrix during the menstrual cycle and first trimester of pregnancy." Cell Tissue Res 253:231-240.
Arango, NA, PP Szotek, TF Manganaro, E Oliva, PK Donahoe, and J Teixeira. 2005. "Conditional deletion of beta-catenin in the mesenchyme of the developing mouse uterus results in a switch to adipogenesis in the myometrium." Dev Biol 288 (1):276-83. doi: 10.1016/j.ydbio.2005.09.045.
Asa, CS, KL Bauman, S Devery, M Zordan, GR Camilo, S Boutelle, and A Moresco. 2014. "Factors associated with uterine endometrial hyperplasia and pyometra in wild canids: implications for fertility." Zoo Biol 33 (1):8-19. doi: 10.1002/zoo.21069.
Ashby, J, W Owens, J Odum, and H Tinwell. 2003. "The Intact Immature Rodent Uterotrophic Bioassay: Possible Effects on Assay Sensitivity of Vomeronasal Signals from Male Rodents and Strain Differences." Environmental Health Perspectives 111 (12):1568-1570. doi: 10.1289/ehp.5981.
Ashby, J, and H Tinwell. 1998. "Uterotrophic Activity of Bisphenol A in the Immature Rat." Environ Health Perspect 106:719-720.
Augsburger, HR, and D Henzi. 2008. "Immunohistochemical expression of collagen types I, III, IV and alpha-actin in the uterine horns of nulliparous and multiparous beagles." Theriogenology 69 (9):1070-6. doi: 10.1016/j.theriogenology.2008.01.019.
Baird, DD, DB Dunson, MC Hill, D Cousins, and JM Schectman. 2003. "High cumulative incidence of uterine leiomyoma in black and white women: Ultrasound evidence." American Journal of Obstetrics and Gynecology 188 (1):100-107. doi: 10.1067/mob.2003.99.
Banerjee, S, KL Chambliss, C Mineo, and PW Shaul. 2014. "Recent insights into non-nuclear actions of estrogen receptor alpha." Steroids 81:64-9. doi: 10.1016/j.steroids.2013.11.002.

171
Barry, C. 2015. "Pyometra in childhood." Obstet Gynecol 125 (5):1035-8. doi: 10.1097/AOG.0000000000000664.
Bartell, SM, L Han, HN Kim, SH Kim, JA Katzenellenbogen, BS Katzenellenbogen, KL Chambliss, PW Shaul, PK Roberson, RS Weinstein, RL Jilka, M Almeida, and SC Manolagas. 2013. "Non-nuclear-initiated actions of the estrogen receptor protect cortical bone mass." Mol Endocrinol 27 (4):649-56. doi: 10.1210/me.2012-1368.
Bateman, HL, and HB Patisaul. 2008. "Disrupted female reproductive physiology following neonatal exposure to phytoestrogens or estrogen specific ligands is associated with decreased GnRH activation and kisspeptin fiber density in the hypothalamus." Neurotoxicology 29 (6):988-97. doi: 10.1016/j.neuro.2008.06.008.
Bazer, FW, D Worthington-White, MFV Fliss, and S Gross. 1991. "Uteroferrin: A Progesterone-induced Hematopoietic Growth Factor of Uterine Origin." Exp Hematol 19:910-915.
Belcher, SM. 2008. "Rapid signaling mechanisms of estrogens in the developing cerebellum." Brain Res Rev 57 (2):481-92. doi: 10.1016/j.brainresrev.2007.07.020.
Belcher, SM, Y Chen, S Yan, and HS Wang. 2012. "Rapid estrogen receptor-mediated mechanisms determine the sexually dimorphic sensitivity of ventricular myocytes to 17beta-estradiol and the environmental endocrine disruptor bisphenol A." Endocrinology 153 (2):712-20. doi: 10.1210/en.2011-1772.
Belcher, SM, R Gear, and EL Kendig. 2015. "Bisphenol A Alters Autonomic Tone, Extracellular Matrix Structure and Induces Sex-Specific Effects on Cardiovascular Function in Male and Female CD-1 Mice." Endocrinology.
Belcher, SM, HH Le, L Spurling, and JK Wong. 2005. "Rapid estrogenic regulation of extracellular signal- regulated kinase 1/2 signaling in cerebellar granule cells involves a G protein- and protein kinase A-dependent mechanism and intracellular activation of protein phosphatase 2A." Endocrinology 146 (12):5397-406. doi: 10.1210/en.2005-0564.
Belcher, SM, X Ma, and HH Le. 2009. "Blockade of estrogen receptor signaling inhibits growth and migration of medulloblastoma." Endocrinology 150 (3):1112-21. doi: 10.1210/en.2008-1363.
Belkin, AM, EA Zemskov, J Hang, SS Akimov, S Sikora, and AY Strongin. 2004. "Cell-surface-associated tissue transglutaminase is a target of MMP-2 proteolysis." Biochemistry 43 (37):11760-9. doi: 10.1021/bi049266z.
Benatti, C, S Alboni, C Montanari, F Caggia, F Tascedda, N Brunello, and JM Blom. 2011. "Central effects of a local inflammation in three commonly used mouse strains with a different anxious phenotype." Behav Brain Res 224 (1):23-34. doi: 10.1016/j.bbr.2011.05.011.

172
Bhandari, RK, SL Deem, DK Holliday, CM Jandegian, CD Kassotis, SC Nagel, DE Tillitt, FS Vom Saal, and CS Rosenfeld. 2015. "Effects of the environmental estrogenic contaminants bisphenol A and 17alpha-ethinyl estradiol on sexual development and adult behaviors in aquatic wildlife species." Gen Comp Endocrinol 214:195-219. doi: 10.1016/j.ygcen.2014.09.014.
Bigsby, RM, L Aixin, K Luo, and GR Cunha. 1990. "Strains Differences in the Ontogeny of Estrogen Receptors in Murine Uterine Epithelium." Endocrinology 126:2592-2596.
Bilinski, P, D Roopenian, and A Gossler. 1998. "Maternal IL-11Ralpha function is required for normal decidua and fetoplacental development in mice." Genes Dev 12:2234-2243.
Bjorn, SF, N Hastrup, LR Lund, K Dano, JF Larsen, and C Pyke. 1997. "Co-ordinated expression of MMP-2 and its putative activator, MT1-MMP, in human placentation." Mol Hum Reprod 3 (8):713-723.
Blair, RM, H Fang, WS Branham, BS Hass, SL Dial, CL Moland, W Tong, L Shi, R Perkins, and DM Sheehan. 2000. "The Estrogen Receptor Relative Binding Affinities of 188 Natural and Xenochemicals: Structural Diversity of Ligands." Toxicol Sci 54:138-153.
Bogusiewicz, M, M Stryjecka-Zimmer, K Postawski, AJ Jakimiuk, and T Rechberger. 2007. "Activity of matrix metalloproteinase-2 and -9 and contents of their tissue inhibitors in uterine leiomyoma and corresponding myometrium." Gynecol Endocrinol 23 (9):541-6. doi: 10.1080/09513590701557416.
Bonnelye, E, and JE Aubin. 2013. "An energetic orphan in an endocrine tissue: a revised perspective of the function of estrogen receptor-related receptor alpha in bone and cartilage." J Bone Miner Res 28 (2):225-33. doi: 10.1002/jbmr.1836.
Boos, A. 2000. "Immunohistochemical Assessment of Collagen Types I, III, IV and VI in Biopsy Samples of the Bovine Uterine Wall Collected during the Oestrous Cycle." Cells Tissue Organs 167:225-238.
Bosquiazzo, VL, L Vigezzi, M Munoz-de-Toro, and EH Luque. 2013. "Perinatal exposure to diethylstilbestrol alters the functional differentiation of the adult rat uterus." J Steroid Biochem Mol Biol 138:1-9. doi: 10.1016/j.jsbmb.2013.02.011.
Brandon, DD, CL Bethea, EY Strawn, MJ Novy, KA Burry, MS Harrington, TE Erickson, C Warner, EJ Keenan, and GM Clinton. 1993. "Progesterone receptor messenger ribonucleic acid and protein are overexpressed in human uterine leiomyomas." Am J Obstet Gynecol 169:78-85.
Branham, WS, DM Sheehan, DR Zehr, E Ridlon, and CJ Nelson. 1985. "The Postnatal Ontogeny of Rat Uterine Glands and Age-Related Effects of 17beta-Estradiol." Endocrinology 117:2229-2237.

173
Bredhult, C, BM Backlin, and M Olovsson. 2007. "Effects of some endocrine disruptors on the proliferation and viability of human endometrial endothelial cells in vitro." Reprod Toxicol 23 (4):550-9. doi: 10.1016/j.reprotox.2007.03.006.
Brodzki, P, K Kostro, A Brodzki, K Niemczuk, and U Lisiecka. 2014. "Cytometric analysis of surface molecules of leucocytes and phagocytic activity of granulocytes and monocytes/macrophages in cows with pyometra." Reprod Domest Anim 49 (5):858-64. doi: 10.1111/rda.12381.
Brodzki, P, K Kostro, A Brodzki, and J Zietek. 2015. "The concentrations of inflammatory cytokines and acute-phase proteins in the peripheral blood and uterine washings in cows with pyometra." Reprod Domest Anim 50 (3):417-22. doi: 10.1111/rda.12507.
Bromer, JG, J Wu, Y Zhou, and HS Taylor. 2009. "Hypermethylation of homeobox A10 by in utero diethylstilbestrol exposure: an epigenetic mechanism for altered developmental programming." Endocrinology 150 (7):3376-82. doi: 10.1210/en.2009-0071.
Bromfield, JJ. 2014. "Seminal fluid and reproduction: much more than previously thought." J Assist Reprod Genet 31 (6):627-36. doi: 10.1007/s10815-014-0243-y.
Brossia, LJ, CS Roberts, JT Lopez, RM Bigsby, and JR Dynlacht. 2009. "Interstrain Differences in the Development of Pyometra after Estrogen Treatment of Rats." J Am Assoc Lab Anim Sci 48 (5):517-520.
Brown, J, and C Farquhar. 2015. "An Overview of Treatments for Endometriosis." JAMA 313 (3):296-297.
Buczkowska, J, R Kozdrowski, M Nowak, A Raś, and J Mrowiec. 2014. "Endometrosis – significance for horse reproduction, pathogenesis, diagnosis, and proposed therapeutic methods." Polish Journal of Veterinary Sciences 17 (3). doi: 10.2478/pjvs-2014-0083.
Bukowska, D, B Kempisty, P Zawierucha, K Jopek, H Piotrowska, P Antosik, S Ciesiolka, M Wozna, KP Brussow, and JM Jaskowski. 2014. "Microarray analysis of inflammatory response-related gene expression in the uteri of dogs with pyometra." J Biol Regul Homeost Agents 28 (4):637-648.
Bulun, SE. 2013. "Uterine fibroids." N Engl J Med 369 (14):1344-55. doi: 10.1056/NEJMra1209993.
Burrows, H. 1935. "Leucocytic invasion as an accompaniment of epithelial metaplasia." J Pathol and Bacteriol 41 (1):43-49.
Burrows, H. 1936. "Pyelitis of Pregnancy in the Light of Conditions Found in Mice after the Prolonged Administration of Oestrogenic Compounds." Proc R Soc Med 29 (4):404-406.
Butler, GS, MJ Butler, SJ Atkinson, H Will, T Tamura, SS van Westrum, T Crabbe, J Clements, MP d'Ortho, and G Murphy. 1998. "The TIMP2 Membrane Type 1 Metalloproteinase

174
"Receptor" Regulates the Concentration and Efficient Activation of Progelatinase A." J Biol Chem 273 (2):871-880.
Buy, JN, and M Ghossain. 2013. "Anatomy, Histology, and Normal Imaging of the Endometrium."543-548. doi: 10.1007/978-3-642-31012-6_21.
Byerly, MS, RD Swanson, GW Wong, and S Blackshaw. 2013. "Estrogen-related receptor beta deficiency alters body composition and response to restraint stress." BMC Physiol 13:10. doi: 10.1186/1472-6793-13-10.
Cai, YR, XL Diao, SF Wang, W Zhang, HT Zhang, and Q Su. 2007. "X-chromosomal inactivation analysis of uterine leiomyomas reveals a common clonal origin of different tumor nodules in some multiple leiomyomas." Int J Oncol 31:1379-1389.
Calafat, AM, J Weuve, X Ye, LT Jia, H Hu, S Ringer, K Huttner, and R Hauser. 2009. "Exposure to bisphenol A and other phenols in neonatal intensive care unit premature infants." Environ Health Perspect 117 (4):639-44. doi: 10.1289/ehp.0800265.
Calafat, AM, X Ye, LY Wong, JA Reidy, and LL Needham. 2008. "Exposure of the U.S. population to bisphenol A and 4-tertiary-octylphenol: 2003-2004." Environ Health Perspect 116 (1):39-44. doi: 10.1289/ehp.10753.
Calafat, Antonia M., Zsuzsanna Kuklenyik, John A. Reidy, Samuel P. Caudill, John Ekong, and Larry L. Needham. 2005. "Urinary Concentrations of Bisphenol A and 4-Nonylphenol in a Human Reference Population." Environmental Health Perspectives 113 (4):391-395. doi: 10.1289/ehp.7534.
Calderon, L, R Williams, M Martinez, KV Clemons, and DA Stevens. 2003. "Genetic susceptibility to vaginal candidiasis." Med Mycol 41:143-147.
Calhoun, KC, E Padilla-Banks, WN Jefferson, L Liu, KE Gerrish, SL Young, CE Wood, PA Hunt, CA Vandevoort, and CJ Williams. 2014. "Bisphenol a exposure alters developmental gene expression in the fetal rhesus macaque uterus." PLoS One 9 (1):e85894. doi: 10.1371/journal.pone.0085894.
Calippe, B, V Douin-Echinard, L Delpy, M Laffargue, K Lelu, A Krust, B Pipy, F Bayard, JF Arnal, JC Guery, and P Gourdy. 2010. "17Beta-estradiol promotes TLR4-triggered proinflammatory mediator production through direct estrogen receptor alpha signaling in macrophages in vivo." J Immunol 185 (2):1169-76. doi: 10.4049/jimmunol.0902383.
Calippe, B, V Douin-Echinard, M Laffargue, H Laurell, V Rana-Poussine, B Pipy, JC Guery, F Bayard, JF Arnal, and P Gourdy. 2008. "Chronic Estradiol Administration In Vivo Promotes the Proinflammatory Response of Macrophages to TLR4 Activation: Involvement of the Phosphatidylinositol 3-Kinase Pathway." The Journal of Immunology 180 (12):7980-7988. doi: 10.4049/jimmunol.180.12.7980.

175
Cao, J, L Joyner, JA Mickens, SM Leyrer, and HB Patisaul. 2014. "Sex-specific Esr2 mRNA expression in the rat hypothalamus and amygdala is altered by neonatal bisphenol A exposure." Reproduction 147 (4):537-54. doi: 10.1530/REP-13-0501.
Cao, J, JA Mickens, KA McCaffrey, SM Leyrer, and HB Patisaul. 2012. "Neonatal Bisphenol A exposure alters sexually dimorphic gene expression in the postnatal rat hypothalamus." Neurotoxicology 33 (1):23-36. doi: 10.1016/j.neuro.2011.11.002.
Cao, J, and HB Patisaul. 2011. "Sexually dimorphic expression of hypothalamic estrogen receptors alpha and beta and Kiss1 in neonatal male and female rats." J Comp Neurol 519 (15):2954-77. doi: 10.1002/cne.22648.
Cao, J, ME Rebuli, J Rogers, KL Todd, SM Leyrer, SA Ferguson, and HB Patisaul. 2013. "Prenatal bisphenol A exposure alters sex-specific estrogen receptor expression in the neonatal rat hypothalamus and amygdala." Toxicol Sci 133 (1):157-73. doi: 10.1093/toxsci/kft035.
Caplan, AI. 1991. "Mesenchymal Stem Cells." J Orthop Res 9 (5):641-650.
Caplan, AI, and JE Dennis. 2006. "Mesenchymal stem cells as trophic mediators." J Cell Biochem 98 (5):1076-84. doi: 10.1002/jcb.20886.
Carbone, K, NM Pinto, PA Abrahamsohn, and TM Zorn. 2006. "Arrangement and fine structure of collagen fibrils in the decidualized mouse endometrium." Microsc Res Tech 69 (1):36-45. doi: 10.1002/jemt.20265.
Carmeci, C, DA Thompson, HZ Ring, U Francke, and RJ Weigel. 1997. "Identification of a Gene (GPR30) with Homology to the G-Protein-Coupled Receptor Superfamily Associated with Estrogen Receptor Expression in Breast Cancer." Genomics 45:607-617.
Carwile, JL, HT Luu, LS Bassett, DA Driscoll, C Yuan, JY Chang, X Ye, AM Calafat, and KB Michels. 2009. "Polycarbonate bottle use and urinary bisphenol A concentrations." Environ Health Perspect 117 (9):1368-72. doi: 10.1289/ehp.0900604.
Caserta, D, N Di Segni, M Mallozzi, V Giovanale, A Mantovani, R Marci, and M Moscarini. 2014. "Bisphenol a and the female reproductive tract: an overview of recent laboratory evidence and epidemiological studies." Reprod Biol Endocrinol 12:37. doi: 10.1186/1477-7827-12-37.
CDC. 2012. DES: Yesterday, Today, Tomorrow. Center for Disease Control and Prevention.
Chapin, RE, J Adams, K Boekelheide, LE Gray, SW Hayward, PS Lees, BS McIntyre, KM Portier, TM Schnorr, SG Selevan, JG Vandenbergh, and SR Woskie. 2008. "NTP-CERHR expert panel report on the reproductive and developmental toxicity of bisphenol A." Birth Defects Res B Dev Reprod Toxicol 83 (3):157-395. doi: 10.1002/bdrb.20147.

176
Chen, GT, CT Tai, LS Yeh, TC Yang, and HD Tsai. 2002. "Identification of the cadherin subtypes present in the human peritoneum and endometriotic lesions: potential role for P-cadherin in the development of endometriosis." Mol Reprod Dev 62 (3):289-94. doi: 10.1002/mrd.10121.
Chen, Z, IS Yuhanna, Z Galcheva-Gargova, RH Karas, ME Mendelsohn, and PW Shaul. 1999. "Estrogen receptor alpha mediates the nongenomic activation of endothelial nitric oxide synthase by estrogen." J Clin Invest 103:401-406.
Chevalier, N, A Bouskine, and P Fenichel. 2012. "Bisphenol A promotes testicular seminoma cell proliferation through GPER/GPR30." Int J Cancer 130 (1):241-2. doi: 10.1002/ijc.25972.
Chotimanukul, S, and S Sirivaidyapong. 2011. "Differential expression of Toll-like receptor 4 (TLR4) in healthy and infected canine endometrium." Theriogenology 76 (6):1152-61. doi: 10.1016/j.theriogenology.2011.05.024.
Chotimanukul, S, and S Sirivaidyapong. 2012. "The localization of Toll-like receptor 2 (TLR2) in the endometrium and the cervix of dogs at different stages of the oestrous cycle and with pyometra." Reprod Domest Anim 47 Suppl 6:351-5. doi: 10.1111/rda.12104.
Christensen, BW, DH Schlafer, DW Agnew, C Wang, C Kozlowski, and CS Asa. 2012. "Diagnostic value of transcervical endometrial biopsies in domestic dogs compared with full-thickness uterine sections." Reprod Domest Anim 47 Suppl 6:342-6. doi: 10.1111/rda.12109.
Chu, P, LA Salamonsen, CS Lee, and PJ Wright. 2002. "Matrix metalloproteinases (MMPs) in the endometrium of bitches." Reproduction 123:467-477.
Clemons, KV, JL Spearow, R Parmar, M Espiritu, and DA Stevens. 2004. "Genetic susceptibility of mice to Candida albicans vaginitis correlates with host estrogen sensitivity." Infect Immun 72 (8):4878-80. doi: 10.1128/IAI.72.8.4878-4880.2004.
Coggan, JA, PA Melville, CM de Oliveria, M Faustino, AM Moreno, and NR Benites. 2008. "Microbiological and Histopathological Aspects of Canine Pyometra." Braz J Microbiol 39:477-483.
Colborn, T, FS vom Saal, and AM Soto. 1993. "Developmental Effects of Endocrine-Disrupting Chemicals in Wildlife and Humans." Environ Health Perspect 101:378-384.
Collette, T, C Bellehumeur, R Kats, R Maheux, J Mailloux, M Villeneuve, and A Akoum. 2004. "Evidence for an increased release of proteolytic activity by the eutopic endometrial tissue in women with endometriosis and for involvement of matrix metalloproteinase-9." Human Reprod 19 (6):1257-1264.

177
Collette, T, R Maheux, J Mailloux, and A Akoum. 2006. "Increased expression of matrix metalloproteinase-9 in the eutopic endometrial tissue of women with endometriosis." Hum Reprod 21 (12):3059-67. doi: 10.1093/humrep/del297.
Colton, T, ER Greenberg, K Noller, L Resseguie, C Van Bennekom, T Heeren, and Y Zhang. 1993. "Breast Cancer in Mothers Prescribed Diethylstilbestrol in Pregnancy." JAMA 269:2096-2100.
Commandeur, AE, AK Styer, and JM Teixeira. 2015. "Epidemiological and genetic clues for molecular mechanisms involved in uterine leiomyoma development and growth." Hum Reprod Update 21 (5):593-615. doi: 10.1093/humupd/dmv030.
Cooke, PS, DC Borsdorf, GC Ekman, KF Doty, SG Clark, PJ Dziuk, and FF Bartol. 2012a. "Uterine gland development begins postnatally and is accompanied by estrogen and progesterone receptor expression in the dog." Theriogenology 78 (8):1787-95. doi: 10.1016/j.theriogenology.2012.05.028.
Cooke, PS, GC Ekman, J Kaur, J Davila, IC Bagchi, SG Clark, PJ Dziuk, K Hayashi, and FF Bartol. 2012b. "Brief exposure to progesterone during a critical neonatal window prevents uterine gland formation in mice." Biol Reprod 86 (3):63. doi: 10.1095/biolreprod.111.097188.
Cooke, PS, TE Spencer, FF Bartol, and K Hayashi. 2013. "Uterine glands: development, function and experimental model systems." Mol Hum Reprod 19 (9):547-58. doi: 10.1093/molehr/gat031.
Cookman, CJ, and SM Belcher. 2014. "Classical nuclear hormone receptor activity as a mediator of complex concentration response relationships for endocrine active compounds." Curr Opin Pharmacol 19C:112-119. doi: 10.1016/j.coph.2014.09.013.
Cooper, JE, EL Kendig, and SM Belcher. 2011. "Assessment of bisphenol A released from reusable plastic, aluminium and stainless steel water bottles." Chemosphere 85 (6):943-7. doi: 10.1016/j.chemosphere.2011.06.060.
Cora, MC, L Kooistra, and G Travlos. 2015. "Vaginal Cytology of the Laboratory Rat and Mouse: Review and Criteria for the Staging of the Estrous Cycle Using Stained Vaginal Smears." Toxicol Pathol 43 (6):776-93. doi: 10.1177/0192623315570339.
Couse, JF, and KS Korach. 2004. "Estrogen receptor-alpha mediates the detrimental effects of neonatal diethylstilbestrol (DES) exposure in the murine reproductive tract." Toxicology 205 (1-2):55-63. doi: 10.1016/j.tox.2004.06.046.
Crossett, B, S Suire, A Herrler, WR Allen, and F Stewart. 1998. "Transfer of a Uterine Lipocalin from the Endometrium of the Mare to the Developing Equine Conceptus." Biol Reprod 59:483-490.

178
Curry, TE, and KG Osteen. 2001. "Cyclic Changes in the Matrix Metalloproteinase System in the Ovary and Uterus." Biol Reprod 64:1285-1296.
Curry, TE, and KG Osteen. 2003. "The matrix metalloproteinase system: changes, regulation, and impact throughout the ovarian and uterine reproductive cycle." Endocr Rev 24 (4):428-65. doi: 10.1210/er.2002-0005.
Dabrowski, R, K Kostro, and M Szczubial. 2013. "Concentrations of C-reactive protein, serum amyloid A, and haptoglobin in uterine arterial and peripheral blood in bitches with pyometra." Theriogenology 80 (5):494-7. doi: 10.1016/j.theriogenology.2013.05.012.
Dabrowski, R, M Szczubial, K Kostro, W Wawron, JJ Ceron, and A Tvarijonaviciute. 2015. "Serum insulin-like growth factor-1 and C-reactive protein concentrations before and after ovariohysterectomy in bitches with pyometra." Theriogenology 83 (4):474-7. doi: 10.1016/j.theriogenology.2014.09.024.
Das, SK, S Yano, J Wang, DR Edwards, H Nagase, and SK Dey. 1997. "Expression of Matrix Metalloproteinases and Tissue Inhibitors of Metalloproteinases in the Mouse Uterus During the Peri-Implantation Period." Dev Genet 21:44-54.
Dayan, D, Y Hiss, A Hirshberg, JJ Bubis, and M Wolman. 1989. "Are the polarization colors of picrosirius red-stained collagen determined only by the diameter of the fibers?" Histochemistry 93 (1):27-29.
De Bosschere, H, R Ducatelle, H Vermeirsch, P Simoens, and M Coryn. 2002. "Estrogen-alpha and progesterone receptor expression in cystic endometrial hyperplasia and pyometra in the bitch." Anim Reprod Sci 70:251-259.
Delclos, KB, CC Weis, TJ Bucci, G Olson, P Mellick, N Sadovova, JR Latendresse, B Thorn, and RR Newbold. 2009. "Overlapping but distinct effects of genistein and ethinyl estradiol (EE(2)) in female Sprague-Dawley rats in multigenerational reproductive and chronic toxicity studies." Reprod Toxicol 27 (2):117-32. doi: 10.1016/j.reprotox.2008.12.005.
DeLeon, DD, MB Zelinski-Wooten, and MS Barkley. 1990. "Hormonal basis of variation in oestrous cyclicity in selected strains of mice." J Reprod Fert 89:117-126.
Dieckmann, WJ, ME Davis, LM Rynkiewicz, and RE Pottinger. 1953. "Does the Administration of Diethylstilbestrol During Pregnancy Have Therapeutic Value?" Am J Obstet Gynecol 66 (5):1062-1081.
Diel, P, S Schmidt, G Vollmer, P Janning, A Upmeier, H Michna, HM Bolt, and GH Degen. 2004. "Comparative responses of three rat strains (DA/Han, Sprague-Dawley and Wistar) to treatment with environmental estrogens." Arch Toxicol 78 (4):183-93. doi: 10.1007/s00204-003-0535-y.

179
Diel, P, T Schulz, K Smolnikar, E Strunck, G Vollmer, and H Michna. 2000. "Ability of xeno- and phytoestrogens to modulate expression of estrogen-sensitive genes in rat uterus: estrogenicity profiles and uterotropic activity." J Steroid Biochem Mol Biol 73:1-10.
Dimitriadis, E, L Robb, Y Liu, AC Enders, H Martin, C Stoikos, E Wallace, and LA Salamonsen. 2003. "IL-11 and IL-11Ralpha immunolocalisation at primate implantation sites supports a role for IL-11 in placentation and fetal development." Reprod Biol Endocrinol 1:34.
Dimitriadis, E, AM Sharkey, YL Tan, LA Salamonsen, and JR Sherwin. 2007. "Immunolocalisation of phosphorylated STAT3, interleukin 11 and leukaemia inhibitory factor in endometrium of women with unexplained infertility during the implantation window." Reprod Biol Endocrinol 5:44. doi: 10.1186/1477-7827-5-44.
Dimitriadis, E, C Stoikos, M Stafford-Bell, I Clark, P Paiva, G Kovacs, and LA Salamonsen. 2006. "Interleukin-11, IL-11 receptoralpha and leukemia inhibitory factor are dysregulated in endometrium of infertile women with endometriosis during the implantation window." J Reprod Immunol 69 (1):53-64. doi: 10.1016/j.jri.2005.07.004.
Dominguez, MA, MA Petre, MS Neal, and WG Foster. 2008. "Bisphenol A concentration-dependently increases human granulosa-lutein cell matrix metalloproteinase-9 (MMP-9) enzyme output." Reprod Toxicol 25 (4):420-5. doi: 10.1016/j.reprotox.2008.05.059.
Dong, JC, H Dong, A Campana, and P Bischof. 2002. "Matrix metalloproteinases and their specific tissue inhibitors in menstruation." Reproduction 123:621-631.
Dong, S, S Terasaka, and R Kiyama. 2011. "Bisphenol A induces a rapid activation of Erk1/2 through GPR30 in human breast cancer cells." Environ Pollut 159 (1):212-8. doi: 10.1016/j.envpol.2010.09.004.
Drasher, ML. 1955. "Strain Differences in the Response of the Mouse Uterus to Estrogens." J Hered 46:190-192.
Du, GQ, L Zhou, XY Chen, XP Wan, and YY He. 2012. "The G protein-coupled receptor GPR30 mediates the proliferative and invasive effects induced by hydroxytamoxifen in endometrial cancer cells." Biochem Biophys Res Commun 420 (2):343-9. doi: 10.1016/j.bbrc.2012.02.161.
Ducsay, CA, WC Buhi, FW Bazer, RM Roberts, and GE Combs. 1984. "Role of Uteroferrin in Placental Iron Transport: Effect of Maternal Iron Treatment on Fetal Iron and Uteroferrin Content and Neonatal Hemoglobin." J Animal Sci 59 (5):1303-1308.
Dunselman, GAJ, PG Groothuis, AFPM de Goeij, and JLH Evers. 2001. "The Mesothelium, Teflon or Velcro? Mesothelium in endometriosis pathogenesis." Hum Reprod 16 (4):605-607.

180
Dyer, RF, J Sodek, and JNM Heersche. 1980. "The Effect of 17beta-Estradiol on Collagen and Noncollagenous Protein Synthesis in the Uterus and Some Periodontal Tissues." Endocrinology 107:1014-1021.
Dziadek, M, P Darling, RZ Zhang, TC Pan, E Tillet, R Timpl, and ML Chu. 1995. "Expression of Collagen alpha1(VI), alpha2(VI), and alpha 3(VI) Chains in the Pregnant Mouse Uterus." Biol Reprod 52:885-894.
Eastin, WC, JH Mennear, RW Tennant, RE Stoll, DG Branstetter, JR Bucher, B McCullough, RL Binder, JW Spalding, and JF Mahler. 2001. "Tg.AC Genetically Altered Mouse: Assay Working Group Overview of Available Data." Toxicol Pathol 29 (Suppl):60-80.
Edlow, AG, M Chen, NA Smith, C Lu, and TF McElrath. 2012. "Fetal bisphenol A exposure: concentration of conjugated and unconjugated bisphenol A in amniotic fluid in the second and third trimesters." Reprod Toxicol 34 (1):1-7. doi: 10.1016/j.reprotox.2012.03.009.
Egenvall, A, R Hagman, BN Bonnett, A Hedhammar, P Olson, and AS Lagerstedt. 2001. "Breed risk of pyometra in insured dogs in Sweden." J Vet Intern Med 15 (6):530-538.
Ehrlich, S, AM Calafat, O Humblet, T Smith, and R Hauser. 2014. "Handling of thermal receipts as a source of exposure to bisphenol A." JAMA 311 (8):859-860.
Ehrlich, S, PL Williams, SA Missmer, JA Flaws, KF Berry, AM Calafat, X Ye, JC Petrozza, D Wright, and R Hauser. 2012a. "Urinary bisphenol A concentrations and implantation failure among women undergoing in vitro fertilization." Environ Health Perspect 120 (7):978-83. doi: 10.1289/ehp.1104307.
Ehrlich, S, PL Williams, SA Missmer, JA Flaws, X Ye, AM Calafat, JC Petrozza, D Wright, and R Hauser. 2012b. "Urinary bisphenol A concentrations and early reproductive health outcomes among women undergoing IVF." Hum Reprod 27 (12):3583-92. doi: 10.1093/humrep/des328.
Ellmann, S, H Sticht, F Thiel, MW Beckmann, R Strick, and PL Strissel. 2009. "Estrogen and progesterone receptors: from molecular structures to clinical targets." Cell Mol Life Sci 66 (15):2405-26. doi: 10.1007/s00018-009-0017-3.
Evans, TJ, MA Miller, VK Ganjam, KD Niswender, MR Ellersieck, WJ Krause, and RS Youngquist. 1998. "Morphometric analysis of endometrial periglandular fibrosis in mares." Am J Vet Res 59:1209-1214.
FAO/WHO. 2011. Toxicological and Health Aspects of Bisphenol A: Report of Joint FAO/WHO Expert Meeting and Report of Stakeholder Meeting on Bisphenol A. World Health Organization.
Farahani, RM, B Sarrafpour, M Simonian, Q Li, and N Hunter. 2012. "Directed glia-assisted angiogenesis in a mature neurosensory structure: pericytes mediate an adaptive response

181
in human dental pulp that maintains blood-barrier function." J Comp Neurol 520 (17):3803-26. doi: 10.1002/cne.23162.
Fassbender, A, RO Burney, DF O, T D'Hooghe, and L Giudice. 2015. "Update on Biomarkers for the Detection of Endometriosis." Biomed Res Int 2015:130854. doi: 10.1155/2015/130854.
FDA. 1972. "Diethylstilbestrol Contraindicated in Pregnancy: Drug's Use Linked to Adenocarcinoma in the Offspring." Western Journal of Medicine 116 (2):85-86.
Fernandez-Catalan, C, W Bode, R Huber, D Turk, JJ Calvete, A Lichte, H Tschesche, and K Maskos. 1998. "Crystal structure of the complex formed by the membrane type 1-matrix metalloproteinase with the tissue inhibitor of metalloproteinase-2, the soluble progelatinase A receptor." EMBO J 17 (17):5238-5248.
Ferreira-Dias, G, LG Nequin, and SS King. 1994. "Morphologic characteristics of equine endometrium classified as Kenney categories I, II, and III, using light and scanning electron microscopy." Am J Vet Res 55 (8):1060-1065.
Fieni, F, E Topie, and A Gogny. 2014. "Medical treatment for pyometra in dogs." Reprod Domest Anim 49 Suppl 2:28-32. doi: 10.1111/rda.12302.
Filant, J, and TE Spencer. 2014. "Uterine glands: biological roles in conceptus implantation, uterine receptivity and decidualization." Int J Dev Biol 58 (2-4):107-16. doi: 10.1387/ijdb.130344ts.
Filant, J, H Zhou, and TE Spencer. 2012. "Progesterone inhibits uterine gland development in the neonatal mouse uterus." Biol Reprod 86 (5):146, 1-9. doi: 10.1095/biolreprod.111.097089.
Filardo, EJ, CT Graeber, JA Quinn, MB Resnick, D Giri, RA DeLellis, MM Steinhoff, and E Sabo. 2006. "Distribution of GPR30, a seven membrane-spanning estrogen receptor, in primary breast cancer and its association with clinicopathologic determinants of tumor progression." Clin Cancer Res 12 (21):6359-66. doi: 10.1158/1078-0432.CCR-06-0860.
Filardo, EJ, JA Quinn, KI Bland, and AR Frackelton. 2000. "Estrogen-Induced Activation of Erk-1 and Erk-2 Requires the G Protein-Coupled Receptor Homolog, GPR30, and Occurs via Trans-Activation of the Epidermal Growth Factor Receptor through Release of HB-EGF." Mol Endocrinol 14:1649-1660.
Filardo, EJ, JA Quinn, AR Frackelton, and KI Bland. 2002. "Estrogen Action Via the G Protein-Coupled Receptor, GPR30: Stimulation of Adenylyl Cyclase and cAMP-Mediated Attenuation of the Epidermal Growth Factor Receptor-to-MAPK Signaling Axis." Mol Endocrinol 16:70-84.

182
Fishman, RB, WS Branham, RD Streck, and DM Sheehan. 1996. "Ontogeny of Estrogen Receptor Messenger Ribonucleic Acid Expression in the Postnatal Rat Uterus." Biol Reprod 55:1221-1230.
Fluhr, H, D Bischof-Islami, S Krenzer, P Licht, P Bischof, and M Zygmunt. 2008. "Human chorionic gonadotropin stimulates matrix metalloproteinases-2 and -9 in cytotrophoblastic cells and decreases tissue inhibitor of metalloproteinases-1, -2, and -3 in decidualized endometrial stromal cells." Fertil Steril 90 (4 Suppl):1390-5. doi: 10.1016/j.fertnstert.2007.08.023.
Fong, CJ, LD Burgoon, KJ Williams, AL Forgacs, and TR Zacharewski. 2007. "Comparative temporal and dose-dependent morphological and transcriptional uterine effects elicited by tamoxifen and ethynylestradiol in immature, ovariectomized mice." BMC Genomics 8:151. doi: 10.1186/1471-2164-8-151.
Fontana, V, TA Coll, CM Sobarzo, LP Tito, JC Calvo, and E Cebral. 2012. "Matrix metalloproteinase expression and activity in trophoblast-decidual tissues at organogenesis in CF-1 mouse." J Mol Histol 43 (5):487-96. doi: 10.1007/s10735-012-9429-8.
Franczak, A, A Zmijewska, B Kurowicka, B Wojciechowicz, and G Kotwica. 2010. "Interleukin 1beta-Induced Synthesis and Secretion of Prostaglandin E2 in the Porcine Uterus During Various Periods of Pregnancy and the Estrous Cycle." J Physiol and Pharmacol 61 (6):733-742.
Frankel, FR, CJ Hsu, JC Myers, E Lin, CR Lyttle, B Komm, and K Mohn. 1988. "Regulation of Alpha2(I), Alpha1(III), and Alpha2(V) Collagen mRNAs by Estradiol in the Immature Rat Uterus." DNA 7 (5):347-354.
Frantz, C, KM Stewart, and VM Weaver. 2010. "The extracellular matrix at a glance." J Cell Sci 123:4195-4200.
Gaetje, R, S Kotzian, G Herrmann, R Baumann, and A Starzinski-Powitz. 1997. "Nonmalignant Epithelial Cells, Potentially Invasive in Human Endometriosis, Lack the Tumor Suppressor Molecular E-Cadherin." Am J Pathol 150:461-467.
Gaide Chevronnay, HP, C Selvais, H Emonard, C Galant, E Marbaix, and P Henriet. 2012. "Regulation of matrix metalloproteinases activity studied in human endometrium as a paradigm of cyclic tissue breakdown and regeneration." Biochim Biophys Acta 1824 (1):146-56. doi: 10.1016/j.bbapap.2011.09.003.
Ganjam, VK, and TJ Evans. 2006. "Equine endometrial fibrosis correlates with 11beta-HSD2, TGF-beta1 and ACE activities." Mol Cell Endocrinol 248 (1-2):104-8. doi: 10.1016/j.mce.2005.12.008.
Gao, F, X Ma, AB Ostmann, and SK Das. 2011. "GPR30 activation opposes estrogen-dependent uterine growth via inhibition of stromal ERK1/2 and estrogen receptor alpha (ERalpha) phosphorylation signals." Endocrinology 152 (4):1434-47. doi: 10.1210/en.2010-1368.

183
Gao, X, Q Liang, Y Chen, and HS Wang. 2013. "Molecular mechanisms underlying the rapid arrhythmogenic action of bisphenol A in female rat hearts." Endocrinology 154 (12):4607-17. doi: 10.1210/en.2013-1737.
Garcia-Falgueras, A, L Ligtenberg, FP Kruijver, and DF Swaab. 2011. "Galanin neurons in the intermediate nucleus (InM) of the human hypothalamus in relation to sex, age, and gender identity." J Comp Neurol 519 (15):3061-84. doi: 10.1002/cne.22666.
Gardner, DG, and D Shoback, eds. 2007. Greenspan's Basic and Clinical Endocrinology. 8 ed. United States: McGraw-Hill.
Gardner, WU, and E Allen. 1937. "Some Effects of Estrogens on the Uterus of the Mouse." Endocrinology 21:727-730.
Ge, LC, ZJ Chen, HY Liu, KS Zhang, H Liu, HB Huang, G Zhang, CK Wong, JP Giesy, J Du, and HS Wang. 2014. "Involvement of activating ERK1/2 through G protein coupled receptor 30 and estrogen receptor alpha/beta in low doses of bisphenol A promoting growth of Sertoli TM4 cells." Toxicol Lett 226 (1):81-9. doi: 10.1016/j.toxlet.2014.01.035.
Gearhart, MD, SMA Holmbeck, RM Evans, HJ Dyson, and PE Wright. 2003. "Monomeric Complex of Human Orphan Estrogen Related Receptor-2 with DNA: A Pseudo-dimer Interface Mediates Extended Half-site Recognition." Journal of Molecular Biology 327 (4):819-832. doi: 10.1016/s0022-2836(03)00183-9.
Geggie, D, and L Walton. 2006. "An unusual acute abdomen in a 4-month-old infant." Emerg Med J 23 (11):e61. doi: 10.1136/emj.2006.040196.
Gerstenberg, C, and WR Allen. 2000. "Development of equine endometrial glands from fetal life to ovarian cyclicity." J Reprod Fert Supplement 56:317-326.
Giannelli, G, C Sgarra, E Di Naro, C Lavopa, U Angelotti, M Tartagni, O Simone, P Trerotoli, S Antonaci, and G Loverro. 2007. "Endometriosis is characterized by an impaired localization of laminin-5 and alpha3beta1 integrin receptor." Int J Gynecol Cancer 17 (1):242-7. doi: 10.1111/j.1525-1438.2006.00750.x.
Giguere, V, N Yang, P Segui, and RM Evans. 1988. "Identification of a new class of steroid hormone receptors." Nature 331 (91-94).
Giudice, LC, and LC Kao. 2004. "Endometriosis." The Lancet 364 (9447):1789-1799. doi: 10.1016/s0140-6736(04)17403-5.
Gnainsky, Y, N Dekel, and I Granot. 2014. "Implantation: mutual activity of sex steroid hormones and the immune system guarantee the maternal-embryo interaction." Semin Reprod Med 32 (5):337-45. doi: 10.1055/s-0034-1376353.

184
Goffin, F, C Munaut, F Frankenne, S Perrier D'Hauterive, A Beliard, V Fridman, P Nervo, A Colige, and JM Foidart. 2003. "Expression pattern of metalloproteinases and tissue inhibitors of matrix-metalloproteinases in cycling human endometrium." Biol Reprod 69 (3):976-84. doi: 10.1095/biolreprod.103.015933.
Golan, DE, AH Tashjian, EJ Armstrong, and AW Armstring. 2012. Principles of Pharmacology: The Pathophysiologic Basis of Drug Therapy. 3rd ed. Philadelphia, PA: Lippincott, Williams, & Wilkins.
Goldman, M, AS Murr, and RL Cooper. 2007. "The rodent estrous cycle: characterization of vaginal cytology and its utility in toxicological studies." Birth Defects Res B Dev Reprod Toxicol 80 (2):84-97. doi: 10.1002/bdrb.20106.
Goldman, S, and E Shalev. 2003. "The role of the matrix metalloproteinases in human endometrial and ovarian cycles." European Journal of Obstetrics & Gynecology and Reproductive Biology 111 (2):109-121. doi: 10.1016/s0301-2115(03)00341-5.
Gomez-Cuetara, C, JM Flores, J Sanchez, A Rodriguez, and MA Sanchez. 1995. "Histological Changes in the Uterus during Postpartum in the Mare." Anat Histol Embryol 24:19-23.
Gomis-Ruth, FX, K Maskos, M Betz, A Bergner, R Huber, K Suzuki, N Yoshida, H Nagase, K Brew, GP Bourenkov, H Bartunik, and W Bode. 1997. "Mechanism of inhibition of the human matrix metalloproteinase stromelysin-1 by TIMP-1." Nature 389:77-81.
Gore, AC, D Crews, LL Doan, M La Merrill, H Patisaul, and A Zota. 2014. Introduction to Endocrine Disrupting Chemicals (EDCs): A Guide for Public Interest Organizations and Policy-Makers. edited by The Endocrine Society and IPEN.
Gorny, KR, BJ Borah, DL Brown, DA Woodrum, EA Stewart, and GK Hesley. 2014. "Incidence of additional treatments in women treated with MR-guided focused US for symptomatic uterine fibroids: review of 138 patients with an average follow-up of 2.8 years." J Vasc Interv Radiol 25 (10):1506-12. doi: 10.1016/j.jvir.2014.05.012.
Gould, JC, LS Leonard, SC Maness, BL Wagner, K Conner, T Zacharewski, S Safe, DP McDonnell, and KW Gaido. 1998. "Bisphenol a interacts with the estrogen receptor a in a distinct manner from estradiol." Mol Cell Endocrinol 142:203-214.
Gould, KA, J Pandey, CM Lachel, CR Murrin, LA Flood, KL Pennington, BS Schaffer, M Tochacek, RD McComb, JL Meza, DL Wendell, and JD Shull. 2005. "Genetic mapping of Eutr1, a locus controlling E2-induced pyometritis in the Brown Norway rat, to RNO5." Mamm Genome 16 (11):854-64. doi: 10.1007/s00335-005-0070-7.
Grandas, OH, DH Mountain, SS Kirkpatrick, DC Cassada, SL Stevens, MB Freeman, and MH Goldman. 2009. "Regulation of vascular smooth muscle cell expression and function of matrix metalloproteinases is mediated by estrogen and progesterone exposure." J Vasc Surg 49 (1):185-91. doi: 10.1016/j.jvs.2008.07.080.

185
Gray, CA, FF Bartol, BJ Tarleton, AA Wiley, GA Johnson, FW Bazer, and TE Spencer. 2001. "Developmental Biology of Uterine Glands." Biol Reprod 65:1311-1323.
Greathouse, KL, JD Cook, K Lin, BJ Davis, TD Berry, TG Bredfeldt, and CL Walker. 2008. "Identification of uterine leiomyoma genes developmentally reprogrammed by neonatal exposure to diethylstilbestrol." Reprod Sci 15 (8):765-78. doi: 10.1177/1933719108322440.
Green, S, P Walter, G Greene, A Krust, C Goffin, E Jensen, G Scrace, M Waterfield, and P Chambon. 1986. "Cloning of the human oestrogen receptor cDNA." J Steroid Biochem 24 (1):77-83.
Greenberg, CS, PJ Birckbichler, and RH Rice. 1991. "Transglutaminases: multifunctional cross-linking enzymes that stabilize tissues." FASEB J 5:3071-3077.
Greene, GL, P Gilna, M Waterfield, A Baker, Y Hort, and J Shine. 1986. "Sequence and Expression of Human Estrogen Receptor Complementary DNA." Science 231:1150-1154.
Greenman, DL, K Dooley, CR Breeden, and GH Gass. 1977. "Strains differences in the response of the mouse to diethylstilbestrol." J Toxicol Environ Health 3 (3):589-597.
Greenman, DL, B Highman, J Chen, W Sheldon, and G Gass. 1990. "Estrogen-Induced Thyroid Follicular Cell Adenomas in C57Bl/6 Mice." J Toxicol Environ Health 29:269-278.
Greenman, DL, B Highman, JJ Chen, GJ Schieferstein, and MJ Norvell. 1986. "Influence of Age on Induction of Mammary Tumors by Diethylstilbestrol in C3H/HeN Mice with Low Murine Mammary Tumor Virus Titer." J Natl Cancer Inst 77:891-898.
Greenman, DL, RL Kodell, B Highman, GJ Schieferstein, and MJ Norvell. 1987. "Mammary Tumorigenesis in C3H/HEN-MTV+ Mice Treated with Diethylstilbestrol for Varying Periods." Fd Chem Toxic 25 (3):229-232.
Griffith, JS, SM Jensen, JK Lunceford, MW Kahn, Y Zheng, EAO Falase, CR Lyttle, and C Teuscher. 1997. "Evidence for the Genetic Control of Estradiol-Regulated Responses. Implications for Variation in Normal and Pathological Hormone-Dependent Phenotypes." Am J Pathol 150:2223-2230.
Groothuis, PG, CAM Koks, AFPM de Goeij, GAJ Dunselman, JW Arends, and JLH Evers. 1999. "Adhesion of human endometrial fragments to peritoneum in vitro." Fertil Steril 71 (6):1119-1124.
Grumetto, L, D Montesano, S Seccia, S Albrizio, and F Barbato. 2008. "Determination of bisphenol a and bisphenol B residues in canned peeled tomatoes by reversed-phase liquid chromatography." J Agric Food Chem 56 (22):10633-7. doi: 10.1021/jf802297z.

186
Guengerich, FP. 1988. "Oxidation of 17alpha-Ethynylestradiol by Human Liver Cytochrome P-450." Mol Pharmacol 33:500-508.
Guengerich, FP. 1990. "Metabolism of 17alpha-ethynylestradiol in humans." Life Sci 47:1981-1988.
Guo, B, BC Han, Z Tian, TY Zhao, L Zeng, ZM Yang, and ZP Yue. 2009. "Expression and hormonal regulation of IL-11Ralpha in canine uterus during early pregnancy." Reprod Domest Anim 44 (5):779-82. doi: 10.1111/j.1439-0531.2008.01072.x.
Hagman, R. 2012. "Clinical and molecular characteristics of pyometra in female dogs." Reprod Domest Anim 47 Suppl 6:323-5. doi: 10.1111/rda.12031.
Hagman, R, and I Kuhn. 2002. "Escherichia coli strains isolated from the uterus and urinary bladder of bitches suffering from pyometra: comparison by restriction enzyme digestion and pulsed-field gel electrophoresis." Vet Microbiol 84:143-153.
Hagman, R, AS Lagerstedt, A Hedhammar, and A Egenvall. 2011. "A breed-matched case-control study of potential risk-factors for canine pyometra." Theriogenology 75 (7):1251-7. doi: 10.1016/j.theriogenology.2010.11.038.
Hampton, AL, and LA Salamonsen. 1994. "Expression of Messenger Ribonucleic Acid Encoding Matrix Metalloproteinases and Their Tissue Inhibitors is Related to Menstruation." J Endocrinol 141:R1-R3.
Hanada, M, Y Maeda, and M Oikawa. 2014. "Histopathological Characteristics of Endometrosis in Thoroughbred Mares in Japan: Results from 50 Necropsy Cases." J Equine Sci 25 (2):45-52.
Harkness, RD, and BE Moralee. 1956. "The time-course and route of loss of collagen from the rat's uterus during post-partum involution." J Physiol 132:502-508.
Harlan. 2009a. "RMS0809-US-EN-01-PS-73 C57Bl/6NHsd."
Harlan. 2009b. "RMS1209-US-EN-01-PS-78 Hsd:ICR (CD-1)."
Haynes, MP, L Li, D Sinha, KS Russell, K Hisamoto, R Baron, M Collinge, WC Sessa, and JR Bender. 2003. "Src kinase mediates phosphatidylinositol 3-kinase/Akt-dependent rapid endothelial nitric-oxide synthase activation by estrogen." J Biol Chem 278 (4):2118-23. doi: 10.1074/jbc.M210828200.
He, YY, B Cai, YX Yang, XL Liu, and XP Wan. 2009. "Estrogenic G protein-coupled receptor 30 signaling is involved in regulation of endometrial carcinoma by promoting proliferation, invasion potential, and interleukin-6 secretion via the MEK/ERK mitogen-activated protein kinase pathway." Cancer Sci 100 (6):1051-61. doi: 10.1111/j.1349-7006.2009.01148.x.

187
Hedrich, HJ, and G Bullock, eds. 2004. The Laboratory Mouse. Edited by P Petrudz, The Handbook of Experimental Animals. London, UK: Elsevier Academic Press.
Heimeier, RA, B Das, DR Buchholz, and YB Shi. 2009. "The xenoestrogen bisphenol A inhibits postembryonic vertebrate development by antagonizing gene regulation by thyroid hormone." Endocrinology 150 (6):2964-73. doi: 10.1210/en.2008-1503.
Helvering, LM, MD Adrian, AG Geiser, ST Estrem, T Wei, S Huang, P Chen, ER Dow, JN Calley, JA Dodge, TA Grese, SA Jones, DL Halladay, RR Miles, JE Onyia, YL Ma, M Sato, and HU Bryant. 2005. "Differential effects of estrogen and raloxifene on messenger RNA and matrix metalloproteinase 2 activity in the rat uterus." Biol Reprod 72 (4):830-41. doi: 10.1095/biolreprod.104.034595.
Hendry, WJ, X Zheng, WW Leavitt, WS Branham, and DM Sheehan. 1997. "Endometrial Hyperplasia and Apoptosis following Neonatal Diethylstilbestrol Exposure and Subsequent Estrogen Stimulation in both Host and Transplanted Hamster Uteri." Cancer Res 57:1903-1908.
Heneweer, M, R Houtman, J Poortman, M Groot, C Maliepaard, and A Peijnenburg. 2007. "Estrogenic effects in the immature rat uterus after dietary exposure to ethinylestradiol and zearalenone using a systems biology approach." Toxicol Sci 99 (1):303-14. doi: 10.1093/toxsci/kfm151.
Henriet, P, HP Gaide Chevronnay, and E Marbaix. 2012. "The endocrine and paracrine control of menstruation." Mol Cell Endocrinol 358 (2):197-207. doi: 10.1016/j.mce.2011.07.042.
Herbst, AL, H Ulfelder, and DC Poskanzer. 1971. "Adenocarcinoma of the Vagina: Association of Maternal Stilbestrol Therapy with Tumor Appearance in Young Women." NEJM 284 (16):878-881.
Hewitt, SC, EH Goulding, EM Eddy, and KS Korach. 2002. "Studies Using the Estrogen Receptor alpha Knockout Uterus Demonstrate That Implantation but Not Decidualization-Associated Signaling is Estrogen Dependent." Biol Reprod 67:1268-1277. doi: 10.1095/biolreprod.101.002436.
Hewitt, SC, and KS Korach. 2003. "Oestrogen receptor knockout mice: roles for oestrogen receptors A and B in reproductive tissues." Reproduction 125:143-149.
Hewitt, SC, and KS Korach. 2011. "Estrogenic activity of bisphenol A and 2,2-bis(p-hydroxyphenyl)-1,1,1-trichloroethane (HPTE) demonstrated in mouse uterine gene profiles." Environ Health Perspect 119 (1):63-70. doi: 10.1289/ehp.1002347.
Hewitt, SC, L Li, SA Grimm, Y Chen, L Liu, Y Li, PR Bushel, D Fargo, and KS Korach. 2012. "Research resource: whole-genome estrogen receptor alpha binding in mouse uterine tissue revealed by ChIP-seq." Mol Endocrinol 26 (5):887-98. doi: 10.1210/me.2011-1311.

188
Hickey, M, K Ballard, and C Farquhar. 2014. "Endometriosis." BMJ 348:g1752. doi: 10.1136/bmj.g1752.
Hinson, J, P Raven, and S Chew, eds. 2010. The Endocrine System. 2 ed, Systems of the Body. China: Elsevier.
Hoekstra, EJ, and C Simoneau. 2013. "Release of bisphenol A from polycarbonate: a review." Crit Rev Food Sci Nutr 53 (4):386-402. doi: 10.1080/10408398.2010.536919.
Hoffmann, C, FW Bazer, J Klug, H Aupperle, C Ellenberger, and HA Schoon. 2009a. "Immunohistochemical and histochemical identification of proteins and carbohydrates in the equine endometrium Expression patterns for mares suffering from endometrosis." Theriogenology 71 (2):264-74. doi: 10.1016/j.theriogenology.2008.07.008.
Hoffmann, C, C Ellenberger, RC Mattos, H Aupperle, S Dhein, B Stief, and HA Schoon. 2009b. "The equine endometrosis: new insights into the pathogenesis." Anim Reprod Sci 111 (2-4):261-78. doi: 10.1016/j.anireprosci.2008.03.019.
Holdsworth-Carson, SJ, M Zaitseva, BJ Vollenhoven, and PA Rogers. 2014. "Clonality of smooth muscle and fibroblast cell populations isolated from human fibroid and myometrial tissues." Mol Hum Reprod 20 (3):250-9. doi: 10.1093/molehr/gat083.
Hong, EJ, SH Park, KC Choi, PC Leung, and EB Jeung. 2006. "Identification of estrogen-regulated genes by microarray analysis of the uterus of immature rats exposed to endocrine disrupting chemicals." Reprod Biol Endocrinol 4:49. doi: 10.1186/1477-7827-4-49.
Hong, Y, J Wang, P Zhang, S Yang, K Song, F Yu, and W Liu. 2010. "Histopathological and gene expression analysis of mice exposed to diethylstilbestrol." Toxicol Mech Methods 20 (3):105-11. doi: 10.3109/15376510903572631.
Hoshino, K, O Takeuchi, T Kawai, H Sanjo, T Ogawa, Y Takeda, K Takeda, and S Akira. 1999. "Toll-Like Receptor 4 (TLR4)-Deficient Mice are Hyporesponsive to Lipopolysaccharide: Evidence for TLR4 as the Lps Gene Product." J Immunol 162:3749-3752.
Howdeshell, KL, PH Peterman, BM Judy, JA Taylor, CE Orazio, RL Ruhlen, FS vom Saal, and WV Welshons. 2003. "Bisphenol A Is Released from Used Polycarbonate Animal Cages into Water at Room Temperature." Environmental Health Perspectives 111 (9):1180-1187. doi: 10.1289/ehp.5993.
Hu, J, CA Gray, and TE Spencer. 2004. "Gene expression profiling of neonatal mouse uterine development." Biol Reprod 70 (6):1870-6. doi: 10.1095/biolreprod.103.026336.
Hulboy, DL, LA Rudolph, and LM Matrisian. 1997. "Matrix metalloproteinases as mediators of reproductive function." Mol Hum Reprod 3 (1):27-45.

189
Hull, ML, CR Escareno, JM Godsland, JR Doig, CM Johnson, SC Phillips, SK Smith, S Tavare, CG Print, and DS Charnock-Jones. 2008. "Endometrial-peritoneal interactions during endometriotic lesion establishment." Am J Pathol 173 (3):700-15. doi: 10.2353/ajpath.2008.071128.
Huppunen, J, and P Aarnisalo. 2004. "Dimerization modulates the activity of the orphan nuclear receptor ERRγ." Biochemical and Biophysical Research Communications 314 (4):964-970. doi: 10.1016/j.bbrc.2003.12.194.
Huss, JM, WG Garbacz, and W Xie. 2015. "Constitutive activities of estrogen-related receptors: Transcriptional regulation of metabolism by the ERR pathways in health and disease." Biochim Biophys Acta 1852 (9):1912-27. doi: 10.1016/j.bbadis.2015.06.016.
Huss, JM, IP Torra, B Staels, V Giguere, and DP Kelly. 2004. "Estrogen-related receptor alpha directs peroxisome proliferator-activated receptor alpha signaling in the transcriptional control of energy metabolism in cardiac and skeletal muscle." Mol Cell Biol 24 (20):9079-91. doi: 10.1128/MCB.24.20.9079-9091.2004.
Hyder, SM, C Chiappetta, and GM Stancel. 1999. "Synthetic Estrogen 17{alpha}-Ethinyl Estradiol Induces Pattern of Uterine Gene Expression Similar to Endogenous Estrogen 17{beta}-Estradiol." J Pharmacol Exp Ther 290:740-747.
IARC. 2012. "Diethylstilbestrol." In IARC Monographs on the Evaluation of Carcinogenic Risks to Humans, edited by International Agency for Research on Cancer World Health Organization, 175-218.
Ikezuki, Y, O Tsutsumi, Y Takai, Y Kamei, and Y Taketani. 2002. "Determination of bisphenol A concentrations in human biological fluids reveals significant early prenatal exposure." Hum Reprod 17 (11):2839-2841.
Inpanbutr, N, EK Miller, BK Petroff, and AM Iacopino. 1994. "CaBP9K Levels during the Luteal and Follicular Phases of the Estrous Cycle in the Bovine Uterus." Biol Reprod 50:561-571.
IRIS, US EPA. 1988. "Bisphenol A. (CASRN 80-05-7) ", Last Modified October 31, 2014. http://www.epa.gov/iris/subst/0356.htm.
Iwahashi, M, Y Muragaki, A Ooshima, M Yamoto, and R Nakano. 1996. "Alterations in distribution and composition of the extracellular matrix during decidualization of the human endometrium." J Reprod Fert 108:147-155.
Iwahashi, M, and R Nakano. 1998. "Decreased type V collagen expression in human decidual tissues of spontaneous abortion during early pregnancy." J Clin Pathol 51:44-46.
Janeway, CA, and R Medzhitov. 2002. "Innate immune recognition." Annu Rev Immunol 20:197-216. doi: 10.1146/annurev.immunol.20.083001.084359.

190
Jensen, EV, and HI Jacobson. 1962. "Basic guides to the mechanism of estrogen action." Recent Prog Horm Res 18:387-414.
Jimenez-Heffernan, JA, A Aguilera, LS Aroeira, E Lara-Pezzi, MA Bajo, G del Peso, M Ramirez, C Gamallo, JA Sanchez-Tomero, V Alvarez, M Lopez-Cabrera, and R Selgas. 2004. "Immunohistochemical characterization of fibroblast subpopulations in normal peritoneal tissue and in peritoneal dialysis-induced fibrosis." Virchows Arch 444 (3):247-56. doi: 10.1007/s00428-003-0963-3.
Junqueira, LC, G Bignolas, and RR Brentani. 1979. "Picrosirius staining plus polarization microscopy, a specific method for collagen detection in tissue sections." Histochem J 11 (4):447-455.
Junqueira, LC, W Cossermelli, and R Brentani. 1978. "Differential staining of collagens type I, II and III by Sirius Red and polarization microscopy." Arch Histol Jpn 41 (3):267-274.
Jussila, T, S Kauppila, M Bode, J Tapanainen, J Risteli, L Risteli, A Kauppila, and F Stenbäck. 2004. "Synthesis and maturation of type I and type III collagens in endometrial adenocarcinoma." European Journal of Obstetrics & Gynecology and Reproductive Biology 115 (1):66-74. doi: 10.1016/s0301-2115(02)00406-2.
Kallen, J, JM Schlaeppi, F Bitsch, I Filipuzzi, A Schilb, V Riou, A Graham, A Strauss, M Geiser, and B Fournier. 2004. "Evidence for ligand-independent transcriptional activation of the human estrogen-related receptor alpha (ERRalpha): crystal structure of ERRalpha ligand binding domain in complex with peroxisome proliferator-activated receptor coactivator-1alpha." J Biol Chem 279 (47):49330-7. doi: 10.1074/jbc.M407999200.
Kang, JS, BJ Lee, B Ahn, DJ Kim, SY Nam, YW Yun, KT Nam, M Choi, HS Kim, DD Jang, YS Lee, and K Yang. 2003. "Expression of Estrogen Receptor {alpha} and {beta} in the Uterus and Vagina of Immature Rats Treated with 17-Ethinyl Estradiol." J Vet Med Sci 65 (12):1293-1297.
Kang, YH, WA Anderson, and ER DeSombre. 1975. "Modulation of Uterine Morphology and Growth by Estradiol-17beta and an Estrogen Antagonist." J Cell Bio 64:682-691.
Kanno, J, L Onyon, J Haseman, P Fenner-Crisp, J Ashby, and W Owens. 2001. "The OECD Program to Validate the Rat Uterotrophic Bioassay to Screen Compounds for in Vivo Estrogenic Responses: Phase 1." Environ Health Perspect 109:785-794.
Kanno, J, L Onyon, S Peddada, J Ashby, E Jacob, and W Owens. 2003. "The OECD Program to Validate the Rat Uterotrophic Bioassay. Phase 2: Coded Single-Dose Studies." Environmental Health Perspectives 111 (12):1550-1558. doi: 10.1289/ehp.5870.
Kao, LC, A Germeyer, S Tulac, S Lobo, JP Yang, RN Taylor, K Osteen, BA Lessey, and LC Giudice. 2003. "Expression profiling of endometrium from women with endometriosis

191
reveals candidate genes for disease-based implantation failure and infertility." Endocrinology 144 (7):2870-81. doi: 10.1210/en.2003-0043.
Kararigas, G, BT Nguyen, LC Zelarayan, M Hassenpflug, K Toischer, H Sanchez-Ruderisch, G Hasenfuss, MW Bergmann, H Jarry, and V Regitz-Zagrosek. 2014. "Genetic background defines the regulation of postnatal cardiac growth by 17beta-estradiol through a beta-catenin mechanism." Endocrinology 155 (7):2667-76. doi: 10.1210/en.2013-2180.
Karkavelas, G, NA Kefalides, PS Amenta, and A Martinez-Hernandez. 1988. "Comparative Ultrastructural Localization of Collagen Types III, IV, VI and Laminin in Rat Uterus and Kidney." J Ultrastruct Mol Struct Res. 100:137-155.
Karlsson, I, R Hagman, A Johannisson, L Wang, E Karlstam, and S Wernersson. 2012. "Cytokines as immunological markers for systemic inflammation in dogs with pyometra." Reprod Domest Anim 47 Suppl 6:337-41. doi: 10.1111/rda.12034.
Karpovich, N, K Chobotova, J Carver, JK Heath, DH Barlow, and HJ Mardon. 2003. "Expression and function of interleukin-11 and its receptor alpha in the human endometrium." Mol Hum Reprod 9 (2):75-80.
Karpovich, N, P Klemmt, JH Hwang, JE McVeigh, JK Heath, DH Barlow, and HJ Mardon. 2005. "The production of interleukin-11 and decidualization are compromised in endometrial stromal cells derived from patients with infertility." J Clin Endocrinol Metab 90 (3):1607-12. doi: 10.1210/jc.2004-0868.
Katayama, S, K Ashizawa, T Fukuhara, M Hiroyasu, Y Tsuzuki, H Tatemoto, T Nakada, and K Nagai. 2006. "Differential expression patterns of Wnt and beta-catenin/TCF target genes in the uterus of immature female rats exposed to 17alpha-ethynyl estradiol." Toxicol Sci 91 (2):419-30. doi: 10.1093/toxsci/kfj167.
Kempson, RL, and MR Hendrickson. 2000. "Smooth Muscle, Endometrial Stromal, and Mixed Mullerian Tumors of the Uterus." Mod Pathol 13 (3):328-342.
Kendig, EL, DR Buesing, SM Christie, CJ Cookman, RB Gear, ER Hugo, SN Kasper, JA Kendziorski, KR Ungi, K Williams, and SM Belcher. 2012. "Estrogen-like disruptive effects of dietary exposure to bisphenol A or 17alpha-ethinyl estradiol in CD1 mice." Int J Toxicol 31 (6):537-50. doi: 10.1177/1091581812463254.
Kendziorski, JA, and SM Belcher. 2015. "Strain-Specific Induction of Endometrial Periglandular Fibrosis in Mice Exposed During Adulthood to the Endocrine Disrupting Chemical Bisphenol A." Reprod Toxicol 58:119-130. doi: 10.1016/j.reprotox.2015.08.001.
Kendziorski, JA, EL Kendig, RB Gear, and SM Belcher. 2012. "Strain specific induction of pyometra and differences in immune responsiveness in mice exposed to 17alpha-ethinyl estradiol or the endocrine disrupting chemical bisphenol A." Reprod Toxicol 34 (1):22-30. doi: 10.1016/j.reprotox.2012.03.001.

192
Kennedy, S, A Bergqvist, C Chapron, T D'Hooghe, G Dunselman, R Greb, L Hummelshoj, A Prentice, and E Saridogan. 2005. "ESHRE guideline for the diagnosis and treatment of endometriosis." Hum Reprod 20 (10):2698-704. doi: 10.1093/humrep/dei135.
Kenney, RM. 1978. "Cyclic and Pathologic Changes of the Mare Endometrium as Detected by Biopsy, with a Note on Early Embryonic Death." J Am Vet Med Assoc 172 (3):241-262.
Kent, UM, H Lin, DE Mills, KA Regal, and PF Hollenberg. 2006. "Identification of 17-alpha-Ethynylestradiol-Modified Active Site Peptides and Glutathione Conjugates Formed during Metabolism and Inactivation of P450s 2B1 and 2B6." Chem Res Toxicol 19:279-287.
Kent, UM, DE Mills, RV Rajnarayanan, WL Alworth, and PF Hollenberg. 2002. "Effect of 17-alpha-Ethynylestradiol on Activities of Cytochrome P450 2B (P450 2B) Enzymes: Characterization of Inactivation of P450s 2B1 and 2B6 and Identification of Metabolites." J Pharmacol Exp Ther 300:549-558.
Kim, JJ, T Kurita, and SE Bulun. 2013. "Progesterone action in endometrial cancer, endometriosis, uterine fibroids, and breast cancer." Endocr Rev 34 (1):130-62. doi: 10.1210/er.2012-1043.
Kitaoka, M, K Iyama, H Yoshioka, M Monda, and G Usuku. 1994. "Immunohistochemical localization of procollagen types I and III during placentation in pregnant rats by type-specific procollagen antibodies." Journal of Histochemistry & Cytochemistry 42 (11):1453-1461. doi: 10.1177/42.11.7930527.
Kitaoka, M, KI Iyama, T Ushijima, C Mimata, H Hori, N Abe, and H Yoshioka. 1996. "Differential Expressions of Collagen Types IV, III, and I During the Development of Invasive Trophoblasts in Rats." Dev Dyn 207:219-331.
Klemmt, PA, JG Carver, P Koninckx, EJ McVeigh, and HJ Mardon. 2007. "Endometrial cells from women with endometriosis have increased adhesion and proliferative capacity in response to extracellular matrix components: towards a mechanistic model for endometriosis progression." Hum Reprod 22 (12):3139-47. doi: 10.1093/humrep/dem262.
Knudsen, LR, CC Karstrup, HG Pedersen, JS Agerholm, TK Jensen, and K Klitgaard. 2015. "Revisiting bovine pyometra--new insights into the disease using a culture-independent deep sequencing approach." Vet Microbiol 175 (2-4):319-24. doi: 10.1016/j.vetmic.2014.12.006.
Kolkova, Z, V Noskova, A Ehinger, S Hansson, and B Casslen. 2010. "G protein-coupled estrogen receptor 1 (GPER, GPR 30) in normal human endometrium and early pregnancy decidua." Mol Hum Reprod 16 (10):743-51. doi: 10.1093/molehr/gaq043.

193
Kraus, RJ, EA Ariazi, ML Farrell, and JE Mertz. 2002. "Estrogen-related receptor alpha 1 actively antagonizes estrogen receptor-regulated transcription in MCF-7 mammary cells." J Biol Chem 277 (27):24826-34. doi: 10.1074/jbc.M202952200.
Krishnan, AV, P Stathis, SF Permuth, L Tokes, and D Feldman. 1993. "Bisphenol-A an estrogenic substance is released from polycarbonate flasks during autoclaving." Endocrinology 132:2279-2286.
Kuiper, GGJM, E Enmark, M Pelto-Huikko, S Nilsson, and J Gustafsson. 1996. "Cloning of a novel estrogen receptor expressed in rat prostate and ovary." Proc Natl Acad Sci U S A 93:5925-5930.
Kuiper, GGJM, JG Lemmen, B Carlsson, JC Corton, SH Safe, PT van der Saag, B van der Burg, and J Gustafsson. 1998. "Interaction of estrogenic chemicals and phytoestrogens with estrogen receptor beta." Endocrinology 139:4252-4263.
Kumar, P, Q Wu, KL Chambliss, IS Yuhanna, SM Mumby, C Mineo, GG Tall, and PW Shaul. 2007. "Direct interactions with G alpha i and G betagamma mediate nongenomic signaling by estrogen receptor alpha." Mol Endocrinol 21 (6):1370-80. doi: 10.1210/me.2006-0360.
Kunkitti, P, S Srisuwatanasagul, and K Chatdarong. 2011. "Distribution of estrogen receptor alpha and progesterone receptor, and leukocyte infiltration in the cervix of cyclic bitches and those with pyometra." Theriogenology 75 (6):979-87. doi: 10.1016/j.theriogenology.2010.11.004.
Kvingedal, AM, and EB Smeland. 1997. "A novel putative G-protein-coupled receptor expressed in lung, heart and lymphoid tissue." FEBS Letters 407 (1):59-62. doi: 10.1016/s0014-5793(97)00278-0.
Lantin-Hermoso, RL, CR Rosenfeld, IS Yuhanna, Z German, Z Chen, and PW Shaul. 1997. "Estrogen acutely stimulates nitric oxide synthase activity in fetal pulmonary artery endothelium." Am J Physiol Lung Cell Mol Physiol 273:L119-L126.
LaRocca, J, A Boyajian, C Brown, SD Smith, and M Hixon. 2011. "Effects of in utero exposure to Bisphenol A or diethylstilbestrol on the adult male reproductive system." Birth Defects Res B Dev Reprod Toxicol 92 (6):526-33. doi: 10.1002/bdrb.20336.
Laws, SC, SA Carey, JM Ferrell, GJ Bodman, and RL Cooper. 2000. "Estrogenic Activity of Octylphenol, Nonylphenol, Bisphenol A and Methoxylchlor in Rats." Toxicol Sci 54:154-167.
Le, HH, and SM Belcher. 2010. "Rapid signaling actions of environmental estrogens in developing granule cell neurons are mediated by estrogen receptor ss." Endocrinology 151 (12):5689-99. doi: 10.1210/en.2010-0710.

194
Le, HH, EM Carlson, JP Chua, and SM Belcher. 2008. "Bisphenol A is released from polycarbonate drinking bottles and mimics the neurotoxic actions of estrogen in developing cerebellar neurons." Toxicol Lett 176 (2):149-56. doi: 10.1016/j.toxlet.2007.11.001.
Lehmann, J, C Ellenberger, C Hoffmann, FW Bazer, J Klug, WR Allen, H Sieme, and HA Schoon. 2011. "Morpho-functional studies regarding the fertility prognosis of mares suffering from equine endometrosis." Theriogenology 76 (7):1326-36. doi: 10.1016/j.theriogenology.2011.06.001.
Leitman, DC, S Paruthiyil, C Yuan, CB Herber, M Olshansky, M Tagliaferri, I Cohen, and TP Speed. 2012. "Tissue-specific regulation of genes by estrogen receptors." Semin Reprod Med 30 (1):14-22. doi: 10.1055/s-0031-1299593.
Leppert, PC, T Baginski, C Prupas, WH Catherino, S Pletcher, and JH Segars. 2004. "Comparative ultrastructure of collagen fibrils in uterine leiomyomas and normal myometrium." Fertil Steril 82 Suppl 3:1182-7. doi: 10.1016/j.fertnstert.2004.04.030.
Levin, ER. 2009. "Plasma membrane estrogen receptors." Trends Endocrinol Metab 20 (10):477-82. doi: 10.1016/j.tem.2009.06.009.
Ley, WB, JM Bowen, DP Sponenberg, and PN Lessard. 1989. "Dimethyl Sulfoxide Intrauterine Therapy in the Mare: Effects upon Endometrial Histological Features and Biopsy Classification." Theriogenology 32 (2):263-276.
Li, R, L Hartley, and L Robb. 2001. "Cloning of rat interleukin 11 and interleukin 11 receptor alpha chain and analysis of their expression in rat uterus in the peri-implantation period." Reproduction 122 (593-600).
Li, Y, CJ Luh, KA Burns, Y Arao, Z Jiang, CT Teng, RR Tice, and KS Korach. 2013. "Endocrine-Disrupting Chemicals (EDCs): In Vitro Mechanism of Estrogenic Activation and Differential Effects on ER Target Genes." Environ Health Perspect 121 (4):459-66. doi: 10.1289/ehp.1205951.
Li, Y, K Okumura, S Nomura, N Maeda, T Miyasho, and H Yokota. 2011. "Oxidatively damaged proteins in the early stage of testicular toxicities in male rats by orally administered with a synthetic oestrogen, diethylstilbestrol." Reprod Toxicol 31 (1):26-34. doi: 10.1016/j.reprotox.2010.09.007.
Liang, Q, X Gao, Y Chen, K Hong, and HS Wang. 2014. "Cellular mechanism of the nonmonotonic dose response of bisphenol A in rat cardiac myocytes." Environ Health Perspect 122 (6):601-8. doi: 10.1289/ehp.1307491.
Linharattanaruksa, P, S Srisuwatanasagul, S Ponglowhapan, M Khalid, and K Chatdarong. 2014. "Collagen and Glycosaminoglycan Profiles in the Canine Cervix during Different Stages of the Estrous Cycle and in Open- and Closed-Cervix Pyometra." Journal of Veterinary Medical Science 76 (2):197-203. doi: 10.1292/jvms.13-0114.

195
Linjawi, S, TC Li, EM Tuckerman, AI Blakemore, and SM Laird. 2004. "Expression of interleukin-11 receptor alpha and interleukin-11 protein in the endometrium of normal fertile women and women with recurrent miscarriage." J Reprod Immunol 64 (1-2):145-55. doi: 10.1016/j.jri.2004.08.007.
Liu, X, A Matsushima, M Shimohigashi, and Y Shimohigashi. 2014. "A characteristic back support structure in the bisphenol A-binding pocket in the human nuclear receptor ERRgamma." PLoS One 9 (6):e101252. doi: 10.1371/journal.pone.0101252.
Lockwood, CJ, C Oner, YH Uz, UA Kayisli, SJ Huang, LF Buchwalder, W Murk, EF Funai, and F Schatz. 2008. "Matrix metalloproteinase 9 (MMP9) expression in preeclamptic decidua and MMP9 induction by tumor necrosis factor alpha and interleukin 1 beta in human first trimester decidual cells." Biol Reprod 78 (6):1064-72. doi: 10.1095/biolreprod.107.063743.
Long, X, R Steinmetz, N Ben-Jonathan, A Caperell-Grant, PCM Young, KP Nephew, and RM Bigsby. 2000. "Strain Differences in Vaginal Responses to the Xenoestrogen Bisphenol A." Environ Health Perspect 108:243-247.
Lubahn, DB, JS Moyer, TS Golding, JF Couse, KS Korach, and O Smithies. 1993. "Alteration of reproductive function but not prenatal sexual development after insertional disruption of the mouse estrogen receptor gene." Proc Natl Acad Sci U S A 90:11162-11166.
Lucidi, RS, CA Witz, M Chrisco, PA Binkley, SA Shain, and RS Schenken. 2005. "A novel in vitro model of the early endometriotic lesion demonstrates that attachment of endometrial cells to mesothelial cells is dependent on the source of endometrial cells." Fertil Steril 84 (1):16-21. doi: 10.1016/j.fertnstert.2004.10.058.
Luo, N, Q Guan, L Zheng, X Qu, H Dai, and Z Cheng. 2014. "Estrogen-mediated activation of fibroblasts and its effects on the fibroid cell proliferation." Transl Res 163 (3):232-41. doi: 10.1016/j.trsl.2013.11.008.
Macer, ML, and HS Taylor. 2012. "Endometriosis and infertility: a review of the pathogenesis and treatment of endometriosis-associated infertility." Obstet Gynecol Clin North Am 39 (4):535-49. doi: 10.1016/j.ogc.2012.10.002.
Maciel, GS, RR Uscategui, VT de Almeida, ME Oliveira, MA Feliciano, and WR Vicente. 2014. "Quantity of IL-2, IL-4, IL-10, INF-gamma, TNF-alpha and KC-like cytokines in serum of bitches with pyometra in different stages of oestrous cycle and pregnancy." Reprod Domest Anim 49 (4):701-4. doi: 10.1111/rda.12360.
Maiti, K, JW Paul, M Read, EC Chan, SC Riley, P Nahar, and R Smith. 2011. "G-1-activated membrane estrogen receptors mediate increased contractility of the human myometrium." Endocrinology 152 (6):2448-55. doi: 10.1210/en.2010-0979.

196
Makinen, N, M Mehine, J Tolvanen, E Kaasinen, Y Li, HJ Lehtonen, M Gentile, J Yan, M Enge, M Taipale, M Aavikko, R Katainen, E Virolainen, T Bohling, TA Koski, V Launonen, J Sjoberg, J Taipale, P Vahteristo, and LA Aaltonen. 2011. "MED12, the Mediator Complex Subunit 12 Gene, Is Mutated at High Frequency in Uterine Leiomyomas." Science 334:252-255.
Mambelli, LI, RC Mattos, GH Winter, DS Madeiro, BP Morais, E Malschitzky, MA Miglino, A Kerkis, and I Kerkis. 2014. "Changes in expression pattern of selected endometrial proteins following mesenchymal stem cells infusion in mares with endometrosis." PLoS One 9 (6):e97889. doi: 10.1371/journal.pone.0097889.
Mambelli, LI, GH Winter, A Kerkis, E Malschitzky, RC Mattos, and I Kerkis. 2013. "A novel strategy of mesenchymal stem cells delivery in the uterus of mares with endometrosis." Theriogenology 79 (5):744-50. doi: 10.1016/j.theriogenology.2012.11.030.
Manase, K, T Endo, M Chida, K Nagasawa, H Honnma, K Yamazaki, Y Kitajima, T Goto, M Kanaya, T Hayashi, T Mitaka, and T Saito. 2006. "Coordinated elevation of membrane type 1-matrix metalloproteinase and matrix metalloproteinase-2 expression in rat uterus during postpartum involution." Reprod Biol Endocrinol 4:32. doi: 10.1186/1477-7827-4-32.
Markey, CM, CL Michaelson, EC Veson, C Sonnenschein, and AM Soto. 2001. "The Mouse Uterotrophic Assay: A Reevaluation of its Validity in Assessing the Estrogenicity of Bisphenol A." Environ Health Perspect 109:55-60.
Markey, CM, PR Wadia, BS Rubin, C Sonnenschein, and AM Soto. 2005. "Long-term effects of fetal exposure to low doses of the xenoestrogen bisphenol-A in the female mouse genital tract." Biol Reprod 72 (6):1344-51. doi: 10.1095/biolreprod.104.036301.
Martinez, FO, and S Gordon. 2014. "The M1 and M2 paradigm of macrophage activation: time for reassessment." F1000Prime Rep 6:13. doi: 10.12703/P6-13.
Matsushima, A, Y Kakuta, T Teramoto, T Koshiba, X Liu, H Okada, T Tokunaga, S Kawabata, M Kimura, and Y Shimohigashi. 2007. "Structural evidence for endocrine disruptor bisphenol A binding to human nuclear receptor ERR gamma." J Biochem 142 (4):517-24. doi: 10.1093/jb/mvm158.
Matthews, JB, K Twomey, and TR Zacharewski. 2001. "In Vitro and in Vivo Interactions of Bisphenol A and Its Metabolite, Bisphenol A Glucuronide, with Estrogen Receptors {alpha} and {beta}." Chem Res Toxicol 14 (2):149-157.
McCain, S, E Ramsay, MC Allender, C Souza, and J Schumacher. 2009. "Pyometra in captive large felids: a review of eleven cases." J Zoo Wildl Med 40 (1):147-51. doi: 10.1638/2008-0008.1.

197
McCarthy, TJ, FZ Beluhan, WA Bardawil, and BT Bennett. 1989. "Pyometra in a Rhesus Monkey Secondary to Prolonged Therapy with Medroxyprogesterone Acetate." Lab Animal Sci 39 (1):71-72.
McDowell, KJ, DC Sharp, A Fazleabas, RM Roberts, and FW Bazer. 1982. "Partial characterization of the equine uteroferrin-like protein." J Reprod Fert Supplement 32:329-334.
McLachlan, JA, RR Newbold, and BC Bullock. 1980. "Long-Term Effects on the Female Mouse Genital Tract Associated with Prenatal Exposure to Diethylstilbestrol." Cancer Res 40:3988-3999.
McLean, AC, N Valenzuela, S Fai, and SA Bennett. 2012. "Performing vaginal lavage, crystal violet staining, and vaginal cytological evaluation for mouse estrous cycle staging identification." J Vis Exp (67):e4389. doi: 10.3791/4389.
Mehasseb, MK, SC Bell, and MA Habiba. 2009. "The effects of tamoxifen and estradiol on myometrial differentiation and organization during early uterine development in the CD1 mouse." Reproduction 138 (2):341-50. doi: 10.1530/REP-09-0054.
Meitzen, J, JI Luoma, MI Boulware, VL Hedges, BM Peterson, K Tuomela, KA Britson, and PG Mermelstein. 2013. "Palmitoylation of estrogen receptors is essential for neuronal membrane signaling." Endocrinology 154 (11):4293-304. doi: 10.1210/en.2013-1172.
Mekada, K, K Abe, A Murakami, S Nakamura, H Nakata, K Moriwaki, Y Obata, and A Yoshiki. 2009. "Genetic Differences among C57Bl/6 Substrains." Exp Anim 58 (2):141-149.
Mendoza-Rodriguez, CA, M Garcia-Guzman, N Baranda-Avila, S Morimoto, M Perrot-Applanat, and M Cerbon. 2011. "Administration of bisphenol A to dams during perinatal period modifies molecular and morphological reproductive parameters of the offspring." Reprod Toxicol 31 (2):177-83. doi: 10.1016/j.reprotox.2010.10.013.
Menkhorst, E, L Salamonsen, L Robb, and E Dimitriadis. 2009. "IL11 antagonist inhibits uterine stromal differentiation, causing pregnancy failure in mice." Biol Reprod 80 (5):920-7. doi: 10.1095/biolreprod.108.073601.
Miele, L, E Cordella-Miele, A Facchiano, and AB Mukherjee. 1990. "Inhibition of Phospholipase A2 by Uteroglobin and Antiflammin Peptides." Adv Exp Med Biol 279:137-160.
Misawa, A, and S Inoue. 2015. "Estrogen-Related Receptors in Breast Cancer and Prostate Cancer." Front Endocrinol (Lausanne) 6:83. doi: 10.3389/fendo.2015.00083.
Misirlioglu, D, D Nak, A Sevimli, Y Nak, MO Ozyigit, A Akkoc, and IT Cangul. 2006. "Steroid Receptor Expression and HER-2/neu (c-erbB-2) Oncoprotein in the Uterus of Cats with Cystic Endometrial Hyperplasia-Pyometra Complex." J Vet Med A 53:225-229.

198
Mok-Lin, E, S Ehrlich, PL Williams, J Petrozza, DL Wright, AM Calafat, X Ye, and R Hauser. 2010. "Urinary bisphenol A concentrations and ovarian response among women undergoing IVF." Int J Androl 33 (2):385-93. doi: 10.1111/j.1365-2605.2009.01014.x.
Molina, PE. 2013. "General Principles of Endocrine Physiology." In Endocrine Physiology, ed PE Molina. New York, NY: McGraw-Hill.
Moriyama, K, T Tagami, T Akamizu, T Usui, M Saijo, N Kanamoto, Y Hataya, A Shimatsu, H Kuzuya, and K Nakao. 2002. "Thyroid hormone action is disrupted by bisphenol A as an antagonist." J Clin Endocrinol Metab 87 (11):5185-90. doi: 10.1210/jc.2002-020209.
Morozova, OV. 1991. "Effects of estrogen on the uterus of mice of different strains." Biull Eksp Biol Med 112 (12):631-633.
Morrione, TG, and S Seifter. 1962. "Alteration in the Collagen Content of the Human Uterus during Pregnancy and Post Partum Involution." J Exp Med 115:357-365.
Mosci, P, D Pietrella, G Ricci, N Pandey, C Monari, E Pericolini, E Gabrielli, S Perito, F Bistoni, and A Vecchiarelli. 2013. "Mouse strain-dependent differences in estrogen sensitivity during vaginal candidiasis." Mycopathologia 175 (1-2):1-11. doi: 10.1007/s11046-012-9589-9.
Mosselman, S, J Polman, and R Dijkema. 1996. "ERbeta: identification and characterization of a novel human estrogen receptor." FEBS Letters 392:49-53.
Mukherjee, AB, L Miele, and N Pattabiraman. 1994. "Phospholipase A2 Enzymes: Regulation and Physiological Role." Biochem Pharmacol 48 (1):1-10.
Murphy, G, H Stanton, S Cowell, G Butler, V Knauper, S Atkinson, and J Gavrilovic. 1999. "Mechanisms of pro matrix metalloproteinase activation." APMIS 107:38-44.
Naciff, JM, ML Jump, SM Torontali, GJ Carr, JP Tiesman, GJ Overmann, and GP Daston. 2002. "Gene Expression Profile Induced by 17alpha-Ethynyl Estradiol, Bisphenol A, and Genistein in the Developing Female Reproductive System of the Rat." Toxicol Sci 68:184-199.
Naciff, JM, GJ Overmann, SM Torontali, GJ Carr, JP Tiesman, BD Richardson, and GP Daston. 2003. "Gene expression profile induced by 17 alpha-ethynyl estradiol in the prepubertal female reproductive system of the rat." Toxicol Sci 72 (2):314-30. doi: 10.1093/toxsci/kfg037.
Nagase, H, R Visse, and G Murphy. 2006. "Structure and function of matrix metalloproteinases and TIMPs." Cardiovasc Res 69 (3):562-73. doi: 10.1016/j.cardiores.2005.12.002.
Navarro, VM, JM Castellano, R Fernandez-Fernandez, ML Barreiro, J Roa, JE Sanchez-Criado, E Aguilar, C Dieguez, L Pinilla, and M Tena-Sempere. 2004. "Developmental and hormonally regulated messenger ribonucleic acid expression of KiSS-1 and its putative

199
receptor, GPR54, in rat hypothalamus and potent luteinizing hormone-releasing activity of KiSS-1 peptide." Endocrinology 145 (10):4565-74. doi: 10.1210/en.2004-0413.
NCI. 1976. "Exposure In Utero to Diethylstilbestrol and Related Synthetic Hormones: Association with Vaginal and Cervical Cancers and Other Abnormalities." JAMA 236 (10):1107-1109.
Newbold, RR, RB Hanson, WN Jefferson, BC Bullock, J Haseman, and JA McLachlan. 1998. "Increased tumors but uncompromised fertility in the female descendents of mice exposed developmentally to diethylstilbestrol." Carcinogenesis 19 (9):1655-1663.
Newbold, RR, WN Jefferson, SF Grissom, E Padilla-Banks, RJ Snyder, and EK Lobenhofer. 2007a. "Developmental Exposure to Diethylstilbestrol Alters Uterine Gene Expression That May Be Associated with Uterine Neoplasia Later in Life." Mol Carcinog 46 (9):783-796.
Newbold, RR, WN Jefferson, and E Padilla-Banks. 2007b. "Long-term adverse effects of neonatal exposure to bisphenol A on the murine female reproductive tract." Reprod Toxicol 24 (2):253-8. doi: 10.1016/j.reprotox.2007.07.006.
Newbold, RR, WN Jefferson, and E Padilla-Banks. 2009. "Prenatal Exposure to Bisphenol A at Environmentally Relevant Doses Adversely Affects the Murine Female Reproductive Tract Later in Life." Environ Health Perspect 117:879-885. doi: 10.1289/.
NIEHS. 2010. "Endocrine Disruptors Fact Sheet."
Nishikawa, M, H Iwano, R Yanagisawa, N Koike, H Inoue, and H Yokota. 2010. "Placental transfer of conjugated bisphenol A and subsequent reactivation in the rat fetus." Environ Health Perspect 118 (9):1196-203. doi: 10.1289/ehp.0901575.
Nisolle, M, F Casanas-Roux, and J Donnez. 2000. "Early-stage endometriosis: adhesion and growth of human menstrual endometrium in nude mide." Fertil Steril 74 (2):306-312.
Nissi, R, A Talvensaari-Mattila, V Kotila, M Niinimaki, I Jarvela, and T Turpeenniemi-Hujanen. 2013. "Circulating matrix metalloproteinase MMP-9 and MMP-2/TIMP-2 complex are associated with spontaneous early pregnancy failure." Reprod Biol Endocrinol 11:2. doi: 10.1186/1477-7827-11-2.
Nuttall, RK, and TG Kennedy. 1999. "Gelatinases A and B and Tissue Inhibitors of Metalloproteinases 1, 2, and 3 during In Vivo and In Vitro Decidualization of Rat Endometrial Stromal Cells." Biol Reprod 60:471-478.
Okada, H, T Tokunaga, X Liu, S Takayanagi, A Matsushima, and Y Shimohigashi. 2008. "Direct evidence revealing structural elements essential for the high binding ability of bisphenol A to human estrogen-related receptor-gamma." Environ Health Perspect 116 (1):32-8. doi: 10.1289/ehp.10587.

200
Otto, C, I Fuchs, G Kauselmann, H Kern, B Zevnik, P Andreasen, G Schwarz, H Altmann, M Klewer, M Schoor, R Vonk, and KH Fritzemeier. 2009. "GPR30 does not mediate estrogenic responses in reproductive organs in mice." Biol Reprod 80 (1):34-41. doi: 10.1095/biolreprod.108.071175.
Owman, C, P Blay, C Nilsson, and SJ Lolait. 1996. "Cloning of Human cDNA Encoding a Novel Heptahelix Receptor Expressed in Burkitt's Lymphoma and Widely Distributed in Brain and Peripheral Tissues." Biochem Biophys Res Commun 228:285-292.
Paiva, P, LA Salamonsen, U Manuelpillai, and E Dimitriadis. 2009. "Interleukin 11 inhibits human trophoblast invasion indicating a likely role in the decidual restraint of trophoblast invasion during placentation." Biol Reprod 80 (2):302-10. doi: 10.1095/biolreprod.108.071415.
Pandey, J, KA Gould, RD McComb, JD Shull, and DL Wendell. 2005. "Localization of Eutr2, a locus controlling susceptibility to DES-induced uterine inflammation and pyometritis, to RNO5 using a congenic rat strain." Mamm Genome 16 (11):865-72. doi: 10.1007/s00335-005-0071-6.
Papaconstantinou, AD, TH Umbreit, BR Fisher, PL Goering, NT Lappas, and KM Brown. 2000. "Bisphenol A-Induced Increase in Uterine Weight and Alterations in Uterine Morphology in Ovariectomized B6C3F1 Mice: Role of the Estrogen Receptor." Toxicol Sci 56:332-339.
Paris, F, P Balaguer, B Terouanne, N Servant, C Lacoste, JP Cravedi, JC Nicolas, and C Sultan. 2002. "Phenylphenols, biphenols, bisphenol-A and 4-tert-octylphenol exhibit alpha and beta estrogen activities and antiandrogen activity in reporter cell lines." Mol Cell Endocrinol 193:43-49.
Passwell, JH, MW Steward, and JF Soothill. 1974. "Inter-Mouse Strain Differences in Macrophage Function and its Relationship to Antibody Responses." Clin Exp Immunol 17:159-167.
Patisaul, HB. 2012. "Infertility in the Southern White Rhino: is diet the source of the problem?" Endocrinology 153 (4):1568-71. doi: 10.1210/en.2012-1008.
Patisaul, HB, AW Sullivan, ME Radford, DM Walker, HB Adewale, B Winnik, JL Coughlin, B Buckley, and AC Gore. 2012. "Anxiogenic effects of developmental bisphenol A exposure are associated with gene expression changes in the juvenile rat amygdala and mitigated by soy." PLoS One 7 (9):e43890. doi: 10.1371/journal.pone.0043890.
Patisaul, HB, KL Todd, JA Mickens, and HB Adewale. 2009. "Impact of neonatal exposure to the ERalpha agonist PPT, bisphenol-A or phytoestrogens on hypothalamic kisspeptin fiber density in male and female rats." Neurotoxicology 30 (3):350-7. doi: 10.1016/j.neuro.2009.02.010.

201
Pawar, S, MJ Laws, IC Bagchi, and MK Bagchi. 2015. "Uterine epithelial estrogen receptor alpha controls decidualization via a paracrine mechanism." Mol Endocrinol:me20151142. doi: 10.1210/me.2015-1142.
Pelch, KE, AL Schroder, PA Kimball, KL Sharpe-Timms, JW Davis, and SC Nagel. 2010. "Aberrant gene expression profile in a mouse model of endometriosis mirrors that observed in women." Fertil Steril 93 (5):1615-1627 e18. doi: 10.1016/j.fertnstert.2009.03.086.
Pepling, ME, EA Sundman, NL Patterson, GW Gephardt, L Medico, and KI Wilson. 2010. "Differences in oocyte development and estradiol sensitivity among mouse strains." Reproduction 139 (2):349-57. doi: 10.1530/REP-09-0392.
Peretz, J, and JA Flaws. 2013. "Bisphenol A down-regulates rate-limiting Cyp11a1 to acutely inhibit steroidogenesis in cultured mouse antral follicles." Toxicol Appl Pharmacol 271 (2):249-56. doi: 10.1016/j.taap.2013.04.028.
Peretz, J, SL Neese, and JA Flaws. 2013. "Mouse strain does not influence the overall effects of bisphenol a-induced toxicity in adult antral follicles." Biol Reprod 89 (5):108. doi: 10.1095/biolreprod.113.111864.
Peretz, J, L Vrooman, WA Ricke, PA Hunt, S Ehrlich, R Hauser, V Padmanabhan, HS Taylor, SH Swan, CA VandeVoort, and JA Flaws. 2014. "Bisphenol A and Reproductive Health: Update of Experimental and Human Evidence, 2007-2013." Environ Health Perspect 122:775-786. doi: 10.1289/ehp.1307728.
Pereza, N, S Ostojic, M Volk, M Kapovic, and B Peterlin. 2012. "Matrix metalloproteinases 1, 2, 3 and 9 functional single-nucleotide polymorphisms in idiopathic recurrent spontaneous abortion." Reprod Biomed Online 24 (5):567-75. doi: 10.1016/j.rbmo.2012.01.008.
Pino, M, C Galleguillos, M Torres, H Sovino, A Fuentes, MA Boric, and MC Johnson. 2009. "Association between MMP1 and MMP9 activities and ICAM1 cleavage induced by tumor necrosis factor in stromal cell cultures from eutopic endometria of women with endometriosis." Reproduction 138 (5):837-47. doi: 10.1530/REP-09-0196.
Plaks, V, J Rinkenberger, J Dai, M Flannery, M Sund, K Kanasaki, W Ni, R Kalluri, and Z Werb. 2013. "Matrix metalloproteinase-9 deficiency phenocopies features of preeclampsia and intrauterine growth restriction." Proc Natl Acad Sci U S A 110 (27):11109-14. doi: 10.1073/pnas.1309561110.
Planey, SL, R Kumar, and JA Arnott. 2014. "Estrogen receptors (ERalpha versus ERbeta): friends or foes in human biology?" J Recept Signal Transduct Res 34 (1):1-5. doi: 10.3109/10799893.2013.853188.
Prossnitz, ER, and M Barton. 2009. "Signaling, physiological functions and clinical relevance of the G protein-coupled estrogen receptor GPER." Prostaglandins Other Lipid Mediat 89 (3-4):89-97. doi: 10.1016/j.prostaglandins.2009.05.001.

202
Prossnitz, ER, and M Maggiolini. 2009. "Mechanisms of estrogen signaling and gene expression via GPR30." Mol Cell Endocrinol 308 (1-2):32-8. doi: 10.1016/j.mce.2009.03.026.
Ptak, A, M Hoffmann, I Gruca, and J Barc. 2014. "Bisphenol A induce ovarian cancer cell migration via the MAPK and PI3K/Akt signalling pathways." Toxicol Lett 229 (2):357-365. doi: 10.1016/j.toxlet.2014.07.001.
Pulgar, R, MF Olea-Serrano, A Novillo-Fertrell, A Rivas, P Pazos, V Pedraza, J Navajas, and N Olea. 2000. "Determination of Bisphenol A and Related Aromatic Compounds Released from Bis-GMA-Based Composites and Sealants by High Performance Liquid Chromatography." Environ Health Perspect 108:21-27.
Pupo, M, A Pisano, R Lappano, MF Santolla, EM De Francesco, S Abonante, C Rosano, and M Maggiolini. 2012. "Bisphenol A induces gene expression changes and proliferative effects through GPER in breast cancer cells and cancer-associated fibroblasts." Environ Health Perspect 120 (8):1177-82. doi: 10.1289/ehp.1104526.
Radovick, S, JE Levine, and A Wolfe. 2012. "Estrogenic regulation of the GnRH neuron." Front Endocrinol (Lausanne) 3:52. doi: 10.3389/fendo.2012.00052.
Ramos, JG, J Varayoud, L Kass, H Rodriguez, L Costabel, M Munoz-De-Toro, and EH Luque. 2003. "Bisphenol a induces both transient and permanent histofunctional alterations of the hypothalamic-pituitary-gonadal axis in prenatally exposed male rats." Endocrinology 144 (7):3206-15. doi: 10.1210/en.2002-0198.
Ramos, JG, J Varayoud, C Sonnenschein, AM Soto, M Munoz De Toro, and EH Luque. 2001. "Prenatal Exposure to Low Doses of Bisphenol A Alters the Periductal Stroma and Glandular Cell Function in the Rat Ventral Prostate." Biol Reprod 65:1271-1277.
Razandi, M, G Alton, A Pedram, S Ghonshani, P Webb, and ER Levin. 2003a. "Identification of a Structural Determinant Necessary for the Localization and Function of Estrogen Receptor at the Plasma Membrane." Molecular and Cellular Biology 23 (5):1633-1646. doi: 10.1128/mcb.23.5.1633-1646.2003.
Razandi, M, A Pedram, ST Park, and ER Levin. 2003b. "Proximal events in signaling by plasma membrane estrogen receptors." J Biol Chem 278 (4):2701-12. doi: 10.1074/jbc.M205692200.
Rebordao, MR, A Galvao, A Szostek, A Amaral, L Mateus, DJ Skarzynski, and G Ferreira-Dias. 2014. "Physiopathologic mechanisms involved in mare endometrosis." Reprod Domest Anim 49 Suppl 4:82-7. doi: 10.1111/rda.12397.
Rebuli, ME, J Cao, E Sluzas, KB Delclos, L Camacho, SM Lewis, MM Vanlandingham, and HB Patisaul. 2014. "Investigation of the Effects of Subchronic Low Dose Oral Exposure to Bisphenol A (BPA) and Ethinyl Estradiol (EE) on Estrogen Receptor Expression in the

203
Juvenile and Adult Female Rat Hypothalamus." Toxicol Sci 140 (1):190-203. doi: 10.1093/toxsci/kfu074.
Revankar, CM, DF Cimino, LA Sklar, JB Arterburn, and ER Prossnitz. 2005. "A Transmembrane Intracellular Estrogen Receptor Mediates Rapid Cell Signaling." Science 307:1625-1630.
Richter, CA, LS Birnbaum, F Farabollini, RR Newbold, BS Rubin, CE Talsness, JG Vandenbergh, DR Walser-Kuntz, and FS vom Saal. 2007. "In vivo effects of bisphenol A in laboratory rodent studies." Reprod Toxicol 24 (2):199-224. doi: 10.1016/j.reprotox.2007.06.004.
Ricketts, SW, and S Alonso. 1991. "The effect of age and parity on the development of equine chronic endometrial disease." Equine Vet J 23 (3):189-192.
Ridgway, SH. 1979. "Reported causes of death of captive killer whales (Orcinus orca)." J Wildl Dis 15 (1):99-104.
Robb, L, R Li, L Hartley, HH Nandurkar, F Koentgen, and CG Begley. 1998. "Infertility in female mice lacking the receptor for interleukin 11 is due to a defective uterine response to implantation." Nat Med 4 (3):303-308.
Robboy, SJ, KL Noller, P O'Brien, RH Kaufman, D Townsend, AB Barnes, J Gundersen, WD Lawrence, E Bergstrahl, S McGorray, BC Tilley, J Anton, and G Chazen. 1984. "Increased Incidence of Cervical and Vaginal Dysplasia in 3,980 Diethylstilbestrol-Exposed Young Women." JAMA 252:2979-2983.
Robertson, SA, JR Prins, DJ Sharkey, and LM Moldenhauer. 2013. "Seminal fluid and the generation of regulatory T cells for embryo implantation." Am J Reprod Immunol 69 (4):315-30. doi: 10.1111/aji.12107.
Rodgers, WH, LM Matrisian, LC Giudice, B Dsupin, P Cannon, C Svitek, F Gorstein, and KG Osteen. 1994. "Patterns of matrix metalloproteinase expression in cycling endometrium imply differential functions and regulation by steroid hormones." J Clin Invest 94 (3):946-53. doi: 10.1172/JCI117461.
Rogalla, P, C Rohen, Y Hennig, U Deichert, U Bonk, and J Bullerdiek. 1995. "Telomere repeat fragment sizes do not limit the growth potential of uterine leiomyomas." Biochem Biophys Res Commun 211 (1):175-182.
Roper, RJ, JS Griffith, CR Lyttle, RW Doerge, AW McNabb, RE Broadbent, and C Teuscher. 1999. "Interacting Quantitative Trait Loci Control Phenotypic Variation in Murine Estradiol-Regulated Responses." Endocrinology 140:556-561.
Ross, RK, MC Pike, MP Vessey, D Bull, D Yeates, and JT Casagrande. 1986. "Risk factors for uterine fibroids: reduced risk associated with oral contraceptives." Br Med J 293 (6543):359-362.

204
Rubin, R, and DS Strayer. 2012. Rubin's Pathology: Clinicopathologic Foundations of Medicine. 6th ed. Baltimore, MD: Lippincott, Williams, and Wilkins.
Rubio, A, F Boyen, O Tas, A Kitshoff, I Polis, B Van Goethem, and H de Rooster. 2014. "Bacterial colonization of the ovarian bursa in dogs with clinically suspected pyometra and in controls." Theriogenology 82 (7):966-71. doi: 10.1016/j.theriogenology.2014.07.016.
Russo, LA, BJ Peano, SP Trivedi, TD Cavalcanto, BA Olenchock, JA Caruso, AR Smolock, O Vishnevsky, and RM Gardner. 2009. "Regulated expression of matrix metalloproteinases, inflammatory mediators, and endometrial matrix remodeling by 17beta-estradiol in the immature rat uterus." Reprod Biol Endocrinol 7:124. doi: 10.1186/1477-7827-7-124.
SABiosciences. 2012. "EpiTect ChIP qPCR Primers." http://www.sabiosciences.com/chipqpcrsearch.php?gene=il11ra&species_id=0&factor=ER-alpha&ninfo=n&ngene=n&nfactor=n.
Salvador, RA, and I Tsai. 1973. "Accumulation of Collagen in the Uterus of the Immature Rat Administered Estradiol-17beta." Biochem Pharmacol 22:37-46.
Sampson, JA. 1927. "Peritoneal endometriosis due to the menstrual dissemination of endometrial tissue into the peritoneal cavity." Am J Obstet Gynecol 14:422-469.
Sanchez-Criado, JE, JM de Las Mulas, C Bellido, VM Navarro, R Aguilar, JC Garrido-Gracia, MM Malagon, M Tena-Sempere, and A Blanco. 2006. "Gonadotropin-secreting cells in ovariectomized rats treated with different oestrogen receptor ligands: a modulatory role for ERbeta in the gonadotrope?" J Endocrinol 188 (2):167-77. doi: 10.1677/joe.1.06377.
Sato, F, M Mori, M Nishi, R Kudo, and H Miyake. 2002. "Familial Aggregation of Uterine Myomas in Japanese Women." J Epidemiol 12:249-253.
Sato, H, T Takino, Y Okada, J Cao, A Shinagawa, E Yamamoto, and M Seiki. 1994. "A matrix metalloproteinase expressed on the surface of invasive tumour cells." Nature 370:61-65.
Sato, K, H Fukata, Y Kogo, J Ohgane, K Shiota, and C Mori. 2009. "Neonatal Exposure to Diethylstilbestrol Alters Expression of DNA Methyltransferases and Methylation of Genomic DNA in the Mouse Uterus." Endocr J 56 (1):131-139.
Sato, T, A Ito, Y Mori, K Yamashita, T Hayakawa, and H Nagase. 1991. "Hormonal regulation of collagenolysis in uterine cervical fibroblasts." Biochem J 275:645-650.
Schonfelder, G, K Friedrich, M Paul, and I Chahoud. 2004. "Developmental effects of prenatal exposure to bisphenol a on the uterus of rat offspring." Neoplasia 6 (5):584-94. doi: 10.1593/neo.04217.

205
Schreiber, SN, R Emter, MB Hock, D Knutti, J Cardenas, M Podvinec, EJ Oakeley, and A Kralli. 2004. "The estrogen-related receptor alpha (ERRalpha) functions in PPARgamma coactivator 1alpha (PGC-1alpha)-induced mitochondrial biogenesis." Proc Natl Acad Sci U S A 101 (17):6472-7. doi: 10.1073/pnas.0308686101.
Schuberth, HJ, U Taylor, H Zerbe, D Waberski, R Hunter, and D Rath. 2008. "Immunological responses to semen in the female genital tract." Theriogenology 70 (8):1174-81. doi: 10.1016/j.theriogenology.2008.07.020.
Sendel, T. 2010. Anatomy, Physiology and Reproduction in the Mare. edited by Food and Rural Affairs Ministry of Agriculture. Ontario, Canada.
Sharkey, AM, and NS Macklon. 2013. "The science of implantation emerges blinking into the light." Reprod Biomed Online 27 (5):453-60. doi: 10.1016/j.rbmo.2013.08.005.
Sharkey, DJ, AM Macpherson, KP Tremellen, and SA Robertson. 2007. "Seminal plasma differentially regulates inflammatory cytokine gene expression in human cervical and vaginal epithelial cells." Mol Hum Reprod 13 (7):491-501. doi: 10.1093/molehr/gam028.
Sharpe-Timms, KL, and KE Cox. 2002. "Paracrine Regulation of Matrix Metalloproteinase Expression in Endometriosis." Ann N Y Acad Sci 955:147-158.
Shen, Y, ML Ren, X Feng, YL Cai, YX Gao, and Q Xu. 2014. "An evidence in vitro for the influence of bisphenol A on uterine leiomyoma." Eur J Obstet Gynecol Reprod Biol 178:80-3. doi: 10.1016/j.ejogrb.2014.03.052.
Shen, Y, Y Zheng, J Jiang, Y Liu, X Luo, Z Shen, X Chen, Y Wang, Y Dai, J Zhao, H Liang, A Chen, and W Yuan. 2015. "Higher urinary bisphenol A concentration is associated with unexplained recurrent miscarriage risk: evidence from a case-control study in eastern China." PLoS One 10 (5):e0127886. doi: 10.1371/journal.pone.0127886.
Sheng, ZG, W Huang, YX Liu, and BZ Zhu. 2013. "Bisphenol A at a low concentration boosts mouse spermatogonial cell proliferation by inducing the G protein-coupled receptor 30 expression." Toxicol Appl Pharmacol 267 (1):88-94. doi: 10.1016/j.taap.2012.12.014.
Sheng, ZG, and BZ Zhu. 2011. "Low concentrations of bisphenol A induce mouse spermatogonial cell proliferation by G protein-coupled receptor 30 and estrogen receptor-alpha." Environ Health Perspect 119 (12):1775-80. doi: 10.1289/ehp.1103781.
Sieli, PT, E Jasarevic, DA Warzak, J Mao, MR Ellersieck, C Liao, K Kannan, SH Collet, PL Toutain, FS Vom Saal, and CS Rosenfeld. 2011. "Comparison of serum bisphenol A concentrations in mice exposed to bisphenol A through the diet versus oral bolus exposure." Environ Health Perspect 119 (9):1260-5. doi: 10.1289/ehp.1003385.
Signorile, PG, EP Spugnini, L Mita, P Mellone, A D'Avino, M Bianco, N Diano, L Caputo, F Rea, R Viceconte, M Portaccio, E Viggiano, G Citro, R Pierantoni, V Sica, B Vincenzi, DG Mita, F Baldi, and A Baldi. 2010. "Pre-natal exposure of mice to bisphenol A elicits

206
an endometriosis-like phenotype in female offspring." Gen Comp Endocrinol 168 (3):318-25. doi: 10.1016/j.ygcen.2010.03.030.
Silberberg, M, and R Silberberg. 1951. "Susceptibility to Estrogen of Breast, Vagina, and Endometrium of Various Strains of Mice." Proc Soc Exp Biol Med 76 (1):161-164.
Sillem, M, S Prifti, B Monga, T Arslic, and B Runnebaum. 1999. "Integrin-mediated adhesion of uterine endometrial cells from endometriosis patients to extracellular matrix proteins is enhanced by tumor necrosis factor alpha (TNFa) and interleukin-1 (IL-1)." Eur J Obstet Gynecol Reprod Biol 87:123-127.
Silva, E, S Leitao, S Henriques, MP Kowalewski, B Hoffmann, G Ferreira-Dias, LL da Costa, and L Mateus. 2010. "Gene transcription of TLR2, TLR4, LPS ligands and prostaglandin synthesis enzymes are up-regulated in canine uteri with cystic endometrial hyperplasia-pyometra complex." J Reprod Immunol 84 (1):66-74. doi: 10.1016/j.jri.2009.10.004.
Simon, MM, S Greenaway, JK White, H Fuchs, V Gailus-Durner, S Wells, T Sorg, K Wong, E Bedu, EJ Cartwright, R Dacquin, S Djebali, J Estabel, J Graw, NJ Ingham, IJ Jackson, A Lengeling, S Mandillo, J Marvel, H Meziane, F Preitner, O Puk, M Roux, DJ Adams, S Atkins, A Ayadi, L Becker, A Blake, D Brooker, H Cater, MF Champy, R Combe, P Danecek, A di Fenza, H Gates, AK Gerdin, E Golini, JM Hancock, W Hans, SM Holter, T Hough, P Jurdic, TM Keane, H Morgan, W Muller, F Neff, G Nicholson, B Pasche, LA Roberson, J Rozman, M Sanderson, L Santos, M Selloum, C Shannon, A Southwell, GP Tocchini-Valentini, VE Vancollie, H Westerberg, W Wurst, M Zi, B Yalcin, R Ramirez-Solis, KP Steel, AM Mallon, MH de Angelis, Y Herault, and SD Brown. 2013. "A comparative phenotypic and genomic analysis of C57BL/6J and C57BL/6N mouse strains." Genome Biol 14 (7):R82. doi: 10.1186/gb-2013-14-7-r82.
Singh, AK, R Chattopadhyay, B Chakravarty, and K Chaudhury. 2013. "Altered circulating levels of matrix metalloproteinases 2 and 9 and their inhibitors and effect of progesterone supplementation in women with endometriosis undergoing in vitro fertilization." Fertil Steril 100 (1):127-34 e1. doi: 10.1016/j.fertnstert.2013.03.006.
Singleton, DW, Y Feng, J Yang, A Puga, AV Lee, and SA Khan. 2006. "Gene expression profiling reveals novel regulation by bisphenol-A in estrogen receptor-alpha-positive human cells." Environ Res 100 (1):86-92. doi: 10.1016/j.envres.2005.05.004.
Skill, NJ, TS Johnson, IG Coutts, RE Saint, M Fisher, L Huang, AM El Nahas, RJ Collighan, and M Griffin. 2004. "Inhibition of transglutaminase activity reduces extracellular matrix accumulation induced by high glucose levels in proximal tubular epithelial cells." J Biol Chem 279 (46):47754-62. doi: 10.1074/jbc.M402698200.
Smith, FO. 2006. "Canine pyometra." Theriogenology 66 (3):610-2. doi: 10.1016/j.theriogenology.2006.04.023.

207
Smith, HO, H Arias-Pulido, DY Kuo, T Howard, CR Qualls, SJ Lee, CF Verschraegen, HJ Hathaway, NE Joste, and ER Prossnitz. 2009. "GPR30 predicts poor survival for ovarian cancer." Gynecol Oncol 114 (3):465-71. doi: 10.1016/j.ygyno.2009.05.015.
Smith, HO, KK Leslie, M Singh, CR Qualls, CM Revankar, NE Joste, and ER Prossnitz. 2007. "GPR30: a novel indicator of poor survival for endometrial carcinoma." Am J Obstet Gynecol 196 (4):386 e1-9; discussion 386 e9-11. doi: 10.1016/j.ajog.2007.01.004.
Snider, TA, C Sepoy, and GR Holyoak. 2011. "Equine endometrial biopsy reviewed: observation, interpretation, and application of histopathologic data." Theriogenology 75 (9):1567-81. doi: 10.1016/j.theriogenology.2010.12.013.
Sohoni, P, and JP Sumpter. 1998. "Several environmental oestrogens are also anti-androgens." J Endocrinol 158:327-339.
Spearow, JL. 1999. "Genetic Variation in Susceptibility to Endocrine Disruption by Estrogen in Mice." Science 285 (5431):1259-1261. doi: 10.1126/science.285.5431.1259.
Spearow, JL, P O'Henley, P Doemeny, R Sera, R Leffler, T Sofos, and M Barkley. 2001. "Genetic variation in physiological sensitivity to estrogen in mice." APMIS 109:356-364.
Spiess, K, WR Teodoro, and TM Zorn. 2007. "Distribution of collagen types I, III, and V in pregnant mouse endometrium." Connect Tissue Res 48 (2):99-108. doi: 10.1080/03008200601166194.
Spiess, K, and TM Zorn. 2007. "Collagen types I, III, and V constitute the thick collagen fibrils of the mouse decidua." Microsc Res Tech 70 (1):18-25. doi: 10.1002/jemt.20381.
Springman, EB, EL Angleton, H Birkedal-Hansen, and HE Van Wart. 1990. "Multiple modes of activation of latent human fibroblast collagenase: Evidence for the role of a Cys73 active-site zinc complex in latency and a "cysteine switch" mechanism of activation." Proc Natl Acad Sci U S A 87:364-368.
Staun-Ram, E, and E Shalev. 2005. "Human trophoblast function during the implantation process." Reprod Biol Endocrinol 3:56. doi: 10.1186/1477-7827-3-56.
Steinmetz, R, NG Brown, DL Allen, RM Bigsby, and N Ben-Jonathan. 1997. "The Environmental Estrogen Bisphenol A Stimulates Prolactin Release in Vitro and in Vivo." Endocrinology 138:1780-1786.
Steinmetz, R, NA Mitchner, A Grant, DL Allen, RM Bigsby, and N Ben-Jonathan. 1998. "The Xenoestrogen Bisphenol A Induces Growth, Differentiation, and c-fos Gene Expression in the Female Reproductive Tract." Endocrinology 139:2741-2747.
Stenback, F. 1989. "Collagen Type III Formation and Distribution in the Uterus: Effects of Hormones and Neoplasm Development." Oncology 46:326-334.

208
Stewart, CA, SJ Fisher, Y Wang, MD Stewart, SC Hewitt, KF Rodriguez, KS Korach, and RR Behringer. 2011. "Uterine gland formation in mice is a continuous process, requiring the ovary after puberty, but not after parturition." Biol Reprod 85 (5):954-64. doi: 10.1095/biolreprod.111.091470.
Stewart, EA, AJ Friedman, K Peck, and RA Nowak. 1994. "Relative Overexpression of Collagen Type I and Collagen Type III Messenger Ribonucleic Acids by Uterine Leiomyomas during the Proliferative Phase of the Menstrual Cycle." J Clin Endocrinol Metab 79:900-906.
Stone, JP, S Holtzman, and CJ Shellabarger. 1979. "Neoplastic Responses and Correlated Plasma Prolactin Levels in Diethylstilbestrol-treated ACI and Sprague-Dawley Rats." Cancer Res 39:773-778.
Strickland, DK, JD Ashcom, S Williams, WH Burgess, M Migliorini, and WS Argraves. 1990. "Sequence Identity between the alpha2-Macroglobulin Receptor and Low Density Lipoprotein Receptor-related Protein Suggests That This Molecule is a Multifunctional Receptor." J Biol Chem 265 (29):17401-17404.
Strongin, AY, I Collier, G Bannikov, BL Marmer, GA Grant, and GI Goldberg. 1995. "Mechanism of Cell Surface Activation of 72-kDa Type IV Collagenase." J Biol Chem 270:5331-5338.
Suire, S, F Stewart, J Beauchamp, and MW Kennedy. 2001. "Uterocalin, a lipocalin provisioning the preattachment equine conceptus: fatty acid and retinol binding properties and structural characterization." Biochem J 356:369-376.
Szamatowicz, J, P Laudanski, and I Tomaszewska. 2002. "Matrix metaloproteinase-9 and tissue inhibitor of matrix metalloproteinase-1: a possible role in the pathogenesis of endometriosis." Human Reprod 17 (2):284-288.
Szostek, AZ, K Lukasik, AM Galvao, GM Ferreira-Dias, and DJ Skarzynski. 2013. "Impairment of the interleukin system in equine endometrium during the course of endometrosis." Biol Reprod 89 (4):79. doi: 10.1095/biolreprod.113.109447.
Szostek, AZ, MJ Siemieniuch, K Lukasik, AM Galvao, GM Ferreira-Dias, and DJ Skarzynski. 2012. "mRNA transcription of prostaglandin synthases and their products in the equine endometrium in the course of fibrosis." Theriogenology 78 (4):768-76. doi: 10.1016/j.theriogenology.2012.03.024.
Szotek, PP, HL Chang, L Zhang, F Preffer, D Dombkowski, PK Donahoe, and J Teixeira. 2007. "Adult mouse myometrial label-retaining cells divide in response to gonadotropin stimulation." Stem Cells 25 (5):1317-25. doi: 10.1634/stemcells.2006-0204.
Taguchi, O, RM Bigsby, and GR Cunha. 1988. "Estrogen Responsiveness and the Estrogen Receptor during Development of the Murine Female Reproductive Tract." Develop Growth & Differ 30 (3):301-313.

209
Takada, Y, C Kato, S Kondo, R Korenaga, and J Ando. 1997. "Cloning of cDNAs Encoding G Protein-Coupled Receptor Expressed in Human Endothelial Cells Exposed to Fluid Shear Stress." Biochem Biophys Res Commun 240:737-741.
Takahashi, M, K Inoue, T Mokikawa, S Matsuo, S Hayashi, K Tamura, G Watanabe, K Taya, and M Yoshida. 2014. "Early indicators of delayed adverse effects in female reproductive organs in rats receiving neonatal exposure to 17alpha-ethynylestradiol." J Toxicol Sciences 39 (5):775-784.
Takeda, K, T Kaisho, and S Akira. 2003. "Toll-like receptors." Annu Rev Immunol 21:335-76. doi: 10.1146/annurev.immunol.21.120601.141126.
Takeda, Y, X Liu, M Sumiyoshi, A Matsushima, M Shimohigashi, and Y Shimohigashi. 2009. "Placenta expressing the greatest quantity of bisphenol A receptor ERR{gamma} among the human reproductive tissues: Predominant expression of type-1 ERRgamma isoform." J Biochem 146 (1):113-22. doi: 10.1093/jb/mvp049.
Tanaka, T, C Wang, and N Umesaki. 1998. "Autocrine/paracrine regulation of human endometrial stromal remodeling by laminin and type IV collagen." International Journal of Molecular Medicine. doi: 10.3892/ijmm_00000059.
Tanikawa, M, TJ Acosta, T Fukui, S Murakami, A Korzekwa, DJ Skarzynski, KK Piotrowska, CK Park, and K Okuda. 2005. "Regulation of prostaglandin synthesis by interleukin-1alpha in bovine endometrium during the estrous cycle." Prostaglandins Other Lipid Mediat 78 (1-4):279-90. doi: 10.1016/j.prostaglandins.2005.09.003.
Taylor, JA, FS Vom Saal, WV Welshons, B Drury, G Rottinghaus, PA Hunt, PL Toutain, CM Laffont, and CA VandeVoort. 2011. "Similarity of bisphenol A pharmacokinetics in rhesus monkeys and mice: relevance for human exposure." Environ Health Perspect 119 (4):422-30. doi: 10.1289/ehp.1002514.
Teeguarden, JG, AM Calafat, X Ye, DR Doerge, MI Churchwell, R Gunawan, and MK Graham. 2011. "Twenty-four hour human urine and serum profiles of bisphenol a during high-dietary exposure." Toxicol Sci 123 (1):48-57. doi: 10.1093/toxsci/kfr160.
Teng, C, B Goodwin, K Shockley, M Xia, R Huang, J Norris, BA Merrick, AM Jetten, CP Austin, and RR Tice. 2013. "Bisphenol A affects androgen receptor function via multiple mechanisms." Chem Biol Interact 203 (3):556-64. doi: 10.1016/j.cbi.2013.03.013.
Teodoro, WR, SS Witzel, APP Velosa, M Shimokomaki, PA Abrahamsohn, and TMT Zorn. 2003. "Increase of Interstitial Collagen in the Mouse Endometrium During Decidualization." Connective Tissue Research 44 (2):96-103. doi: 10.1080/03008200390200238.
Thayer, KA, RL Ruhlen, KL Howdeshell, DL Buchanan, PS Cooke, D Preziosi, WV Welshons, J Haseman, and FS vom Saal. 2001. "Altered prostate growth and daily sperm production

210
in male mice exposed prenatally to subclinical doses of 17alpha-ethinyl oestradiol." Human Reprod 16 (5):988-996.
ThermoScientific. 2011. Assessment of Nucleic Acid Purity.
Thigpen, JE, KDR Setchell, GE Kissling, J Locklear, GF Caviness, T Whiteside, SM Belcher, NM Brown, BJ Collins, FB Lih, KB Tomer, E Padilla-Banks, L Camacho, FG Adsit, and M Grant. 2013. "The Estrogenic Content of Rodent Diets, Bedding, Cages, and Water Bottles and Its Effect on Bisphenol A Studies." J Am Assoc Lab Anim Sci 52 (2):130-141.
Thomas, P, and J Dong. 2006. "Binding and activation of the seven-transmembrane estrogen receptor GPR30 by environmental estrogens: a potential novel mechanism of endocrine disruption." J Steroid Biochem Mol Biol 102 (1-5):175-9. doi: 10.1016/j.jsbmb.2006.09.017.
Thompson, GS. 2015. Understanding Anatomy and Physiology: A Visual, Auditory, Interactive Approach. 2nd ed. Philadelphia, PA: FA Davis Company.
Tian, R, Z Wang, Z Shi, D Li, Y Wang, Y Zhu, W Lin, Y Gui, and XL Zheng. 2013. "Differential expression of G-protein-coupled estrogen receptor-30 in human myometrial and uterine leiomyoma smooth muscle." Fertil Steril 99 (1):256-63. doi: 10.1016/j.fertnstert.2012.09.011.
Titus-Ernstoff, L, EE Hatch, RN Hoover, J Palmer, ER Greenberg, W Ricker, R Kaufman, K Noller, AL Herbst, T Colton, and P Hartge. 2001. "Long-term cancer risk is women given diethylstilbestrol (DES) during pregnancy." Br J Cancer 84 (1):126-133. doi: 10.1054/.
Titus-Ernstoff, L, R Troisi, EE Hatch, JR Palmer, M Hyer, R Kaufman, E Adam, K Noller, and RN Hoover. 2010. "Birth defects in the sons and daughters of women who were exposed in utero to diethylstilbestrol (DES)." Int J Androl 33 (2):377-84. doi: 10.1111/j.1365-2605.2009.01010.x.
Titus-Ernstoff, L, R Troisi, EE Hatch, LA Wise, J Palmer, M Hyer, R Kaufman, E Adam, W Strohsnitter, K Noller, AL Herbst, J Gibson-Chambers, P Hartge, and RN Hoover. 2006. "Menstrual and reproductive characteristics of women whose mothers were exposed in utero to diethylstilbestrol (DES)." Int J Epidemiol 35 (4):862-8. doi: 10.1093/ije/dyl106.
Tohme, M, SM Prud'homme, A Boulahtouf, E Samarut, F Brunet, L Bernard, W Bourguet, Y Gibert, P Balaguer, and V Laudet. 2014. "Estrogen-related receptor gamma is an in vivo receptor of bisphenol A." FASEB J 28 (7):3124-33. doi: 10.1096/fj.13-240465.
Toth, M, A Sohail, and R Fridman. 2012. "Assessment of gelatinases (MMP-2 and MMP-9) by gelatin zymography." Methods Mol Biol 878:121-35. doi: 10.1007/978-1-61779-854-2_8.

211
Tsai, JH, JM Hwang, TH Ying, JC Shyu, CC Tsai, YS Hsieh, YW Wang, JY Liu, and SH Kao. 2009. "The activation of matrix metalloproteinase-2 induced by protein kinase C alpha in decidualization." J Cell Biochem 108 (3):547-54. doi: 10.1002/jcb.22285.
Tsigkou, A, FM Reis, P Ciarmela, MH Lee, B Jiang, C Tosti, FR Shen, Z Shi, YG Chen, and F Petraglia. 2015. "Expression Levels of Myostatin and Matrix Metalloproteinase 14 mRNAs in Uterine Leiomyoma are Correlated With Dysmenorrhea." Reprod Sci. doi: 10.1177/1933719115592710.
Tsumagari, S, T Ishinazaka, H Kamata, S Ohba, S Tanaka, M Ishii, and MA Memon. 2005. "Induction of canine pyometra by inoculation of Escherichia coli into the uterus and its relationship to reproductive features." Anim Reprod Sci 87 (3-4):301-8. doi: 10.1016/j.anireprosci.2004.11.006.
Turusov, VS, LS Trukhanova, YD Parfenov, and L Tomatis. 1992. "Occurrence of Tumours in the Descendants of CBA Male Mice Prenatally Treated with Diethylstilbestrol." Int J Cancer 50:131-135.
Tyl, RW, CB Myers, MC Marr, CS Sloan, NP Castillo, MM Veselica, JC Seely, SS Dimond, JP Van Miller, RN Shiotsuka, D Beyer, SG Hentges, and JM Waechter. 2008. "Two-generation reproductive toxicity study of dietary bisphenol A in CD-1 (Swiss) mice." Toxicol Sci 104 (2):362-84. doi: 10.1093/toxsci/kfn084.
Tyl, RW, CB Myers, MC MArr, BF Thomas, AR Keimowitz, DR Brine, MM Veselica, PA Fail, TY Chang, JC Seely, RL Joiner, JH Butala, SS Dimond, SZ Cagen, RN Shiotsuka, GD Stropp, and JM Waechter. 2002. "Three-Generation Reproductive Toxicity Study of Dietary Bisphenol A in CD Sprague-Dawley Rats." Toxicol Sci 68:121-146.
van Poll, D, B Parekkadan, IHM Borel Rinkes, AW Tilles, and ML Yarmush. 2008. "Mesenchymal Stem Cell Therapy for Protection and Repair of Injured Vital Organs." Cellular and Molecular Bioengineering 1 (1):42-50. doi: 10.1007/s12195-008-0001-2.
Van Wart, HE, and H Birkedal-Hansen. 1990. "The cysteine switch: A principle of regulation of metalloproteinase activity with potential applicability to the entire matrix metalloproteinase gene family." Proc Natl Acad Sci U S A 87:5578-5582.
Vanacker, JM, K Pettersson, JA Gustafsson, and V Laudet. 1999. "Transcriptional targets shared by estrogen receptor-related receptors (ERRs) and estrogen receptor (ER) alpha, but not ERbeta." EMBO J 18 (15):4270-4279.
Vandenberg, LN, T Colborn, TB Hayes, JJ Heindel, DR Jacobs, DH Lee, T Shioda, AM Soto, FS vom Saal, WV Welshons, RT Zoeller, and JP Myers. 2012. "Hormones and endocrine-disrupting chemicals: low-dose effects and nonmonotonic dose responses." Endocr Rev 33 (3):378-455. doi: 10.1210/er.2011-1050.

212
Vandenberg, LN, R Hauser, M Marcus, N Olea, and WV Welshons. 2007a. "Human exposure to bisphenol A (BPA)." Reprod Toxicol 24 (2):139-77. doi: 10.1016/j.reprotox.2007.07.010.
Vandenberg, LN, MV Maffini, PR Wadia, C Sonnenschein, BS Rubin, and AM Soto. 2007b. "Exposure to environmentally relevant doses of the xenoestrogen bisphenol-A alters development of the fetal mouse mammary gland." Endocrinology 148 (1):116-27. doi: 10.1210/en.2006-0561.
Varayoud, J, JG Ramos, VL Bosquiazzo, M Lower, M Munoz-de-Toro, and EH Luque. 2011. "Neonatal exposure to bisphenol A alters rat uterine implantation-associated gene expression and reduces the number of implantation sites." Endocrinology 152 (3):1101-11. doi: 10.1210/en.2009-1037.
Verloop, J, MA Rookus, and FE van Leeuwen. 2000. "Prevalence of Gynecologic Cancer in Women Exposed to Diethylstilbestrol in Utero." NEJM 342:1838-1839.
Ververidis, HN, CM Boscos, A Stefanakis, P Saratsis, AI Stamou, and E Krambovitis. 2004. "Serum estradiol-17 beta, progesterone and respective uterine cytosol receptor concentrations in bitches with spontaneous pyometra." Theriogenology 62 (3-4):614-23. doi: 10.1016/j.theriogenology.2003.11.011.
Vida, B, L Deli, E Hrabovszky, T Kalamatianos, A Caraty, CW Coen, Z Liposits, and I Kallo. 2010. "Evidence for suprachiasmatic vasopressin neurones innervating kisspeptin neurones in the rostral periventricular area of the mouse brain: regulation by oestrogen." J Neuroendocrinol 22 (9):1032-9. doi: 10.1111/j.1365-2826.2010.02045.x.
Vigezzi, L, VL Bosquiazzo, L Kass, JG Ramos, M Munoz-de-Toro, and EH Luque. 2015. "Developmental exposure to bisphenol A alters the differentiation and functional response of the adult rat uterus to estrogen treatment." Reprod Toxicol 52:83-92. doi: 10.1016/j.reprotox.2015.01.011.
Vilos, GA, C Allaire, PY Laberge, and N Leyland. 2015. "The Management of Uterine Leiomyomas." J Obstet Gynaecol Can 37 (2):157-178.
Vivacqua, A, D Bonofiglio, AG Recchia, AM Musti, D Picard, S Ando, and M Maggiolini. 2006. "The G protein-coupled receptor GPR30 mediates the proliferative effects induced by 17beta-estradiol and hydroxytamoxifen in endometrial cancer cells." Mol Endocrinol 20 (3):631-46. doi: 10.1210/me.2005-0280.
Vom Saal, FS, CA VandeVoort, JA Taylor, WV Welshons, PL Toutain, and PA Hunt. 2014. "Bisphenol A (BPA) pharmacokinetics with daily oral bolus or continuous exposure via silastic capsules in pregnant rhesus monkeys: Relevance for human exposures." Reprod Toxicol 45:105-16. doi: 10.1016/j.reprotox.2014.01.007.
Voorwald, FA, FA Marchi, RA Villacis, CE Alves, GH Toniollo, RL Amorim, SA Drigo, and SR Rogatto. 2015. "Molecular Expression Profile Reveals Potential Biomarkers and

213
Therapeutic Targets in Canine Endometrial Lesions." PLoS One 10 (7):e0133894. doi: 10.1371/journal.pone.0133894.
Vyas, S, A Kumar, M Prakash, R Kapoor, P Kumar, and N Khandelwal. 2009. "Spontaneous perforation of pyometra in a cervical cancer patient: a case report and literature review." Cancer Imaging 9:12-4. doi: 10.1102/1470-7330.2009.0002.
Wadia, PR, LN Vandenberg, CM Schaeberle, BS Rubin, C Sonnenschein, and AM Soto. 2007. "Perinatal bisphenol A exposure increases estrogen sensitivity of the mammary gland in diverse mouse strains." Environ Health Perspect 115 (4):592-8. doi: 10.1289/ehp.9640.
Walker, BE. 1984. "Tumors of Female Offspring of Mice Exposed Prenatally to Diethylstilbestrol." JNCI Journal of the National Cancer Institute 73:133-140.
Wall, EH, SC Hewitt, LK Case, CY Lin, KS Korach, and C Teuscher. 2014. "The role of genetics in estrogen responses: a critical piece of an intricate puzzle." FASEB J 28 (12):5042-54. doi: 10.1096/fj.14-260307.
Wall, EH, SC Hewitt, L Liu, R del Rio, LK Case, CY Lin, KS Korach, and C Teuscher. 2013. "Genetic control of estrogen-regulated transcriptional and cellular responses in mouse uterus." FASEB J 27 (5):1874-86. doi: 10.1096/fj.12-213462.
Walter, I, J Handler, I Miller, and C Aurich. 2005. "Matrix metalloproteinase 2 (MMP-2) and tissue transglutaminase (TG 2) are expressed in periglandular fibrosis in horse mares with endometrosis." Histol Histopathol 20:1105-1113.
Walter, I, J Handler, M Reifinger, and C Aurich. 2001. "Association of endometrosis in horses with differentiation of periglandular myofibroblasts and changes of extracellular matrix proteins." Reproduction 121:581-586.
Wang, H, and SK Dey. 2006. "Roadmap to embryo implantation: clues from mouse models." Nat Rev Genet 7 (3):185-99. doi: 10.1038/nrg1808.
Wang, H, Y Wen, S Mooney, H Li, B Behr, and ML Polan. 2003. "Matrix metalloproteinase and tissue inhibitor of matrix metalloproteinase expression in human preimplantation embryos." Fertility and Sterility 80:736-742. doi: 10.1016/s0015-0282(03)00782-9.
Wang, Y, G Feng, J Wang, Y Zhou, Y Liu, Y Shi, Y Zhu, W Lin, Y Xu, and Z Li. 2015. "Differential effects of tumor necrosis factor-alpha on matrix metalloproteinase-2 expression in human myometrial and uterine leiomyoma smooth muscle cells." Hum Reprod 30 (1):61-70. doi: 10.1093/humrep/deu300.
Warita, K, T Mitsuhashi, Y Tabuchi, K Ohta, S Suzuki, N Hoshi, T Miki, and Y Takeuchi. 2012. "Microarray and gene ontology analyses reveal downregulation of DNA repair and apoptotic pathways in diethylstilbestrol-exposed testicular Leydig cells." J Toxicol Sciences 37 (2):287-295.

214
Watson, CS, G Hu, and AA Paulucci-Holthauzen. 2014. "Rapid actions of xenoestrogens disrupt normal estrogenic signaling." Steroids 81:36-42. doi: 10.1016/j.steroids.2013.11.006.
Weems, CW, YS Weems, and RD Randel. 2006. "Prostaglandins and reproduction in female farm animals." Vet J 171 (2):206-28. doi: 10.1016/j.tvjl.2004.11.014.
Weinstein, L, WU Gardner, and E Allen. 1943. "The Effect of Estrogenic Hormones on the Bacterial Content of the Uterus." Yale J Biol Med 16 (1):43-51.
Welshons, WV, KA Thayer, BM Judy, JA Taylor, EM Curran, and FS vom Saal. 2003. "Large Effects from Small Exposures. I. Mechanisms for Endocrine-Disrupting Chemicals with Estrogenic Activity." Environmental Health Perspectives 111 (8):994-1006. doi: 10.1289/ehp.5494.
Westberg, JA, HA Bern, and EB Barnawell. 1957. "Strain Differences in the Response of the Mouse Adrenal to Oestrogen." Acta Endocrinol 25:70-82.
Wetherill, YB, BT Akingbemi, J Kanno, JA McLachlan, A Nadal, C Sonnenschein, CS Watson, RT Zoeller, and SM Belcher. 2007. "In vitro molecular mechanisms of bisphenol A action." Reprod Toxicol 24 (2):178-98. doi: 10.1016/j.reprotox.2007.05.010.
Wisniewski, P, RM Romano, MM Kizys, KC Oliveira, T Kasamatsu, G Giannocco, MI Chiamolera, MR Dias-da-Silva, and MA Romano. 2015. "Adult exposure to bisphenol A (BPA) in Wistar rats reduces sperm quality with disruption of the hypothalamic-pituitary-testicular axis." Toxicology 329:1-9. doi: 10.1016/j.tox.2015.01.002.
Witham, EA, JD Meadows, HM Hoffmann, S Shojaei, D Coss, AS Kauffman, and PL Mellon. 2013. "Kisspeptin regulates gonadotropin genes via immediate early gene induction in pituitary gonadotropes." Mol Endocrinol 27 (8):1283-94. doi: 10.1210/me.2012-1405.
Witz, CA, S Cho, VE Centonze, IA Montoya-Rodriguez, and RS Schenken. 2003. "Time series analysis of transmesothelial invasion by endometrial stromal and epithelial cells using three-dimensional confocal microscopy." Fertility and Sterility 79:770-778. doi: 10.1016/s0015-0282(02)04834-3.
Witz, CA, A Takahashi, IA Montoya-Rodriguez, S Cho, and RS Schenken. 2000. "Expression of the alpha2beta1 and alpha3beta1 integrins at the surface of mesothelial cells: a potential attachment site of endometrial cells." Fertil Steril 73 (3):579-584.
Wong, JK, HH Le, A Zsarnovszky, and SM Belcher. 2003. "Estrogens and ICI182,780 (Faslodex) Modulate Mitosis and Cell Death in Immature Cerebellar Neurons via Rapid Activation of p44/p42 Mitogen-Activated Protein Kinase." J Neurosci 23 (12):4984-4995.
Wooding, FBP, G Morgan, GV Jones, and AD Care. 1996. "Calcium transport and the localisation of calbindin-D9k in the ruminant placenta during the second half of pregnancy." Cell Tissue Res 285:477-489.

215
Wu, C, I Macleod, and AI Su. 2013. "BioGPS and MyGene.info: organizing online, gene-centric information." Nucleic Acids Res 41 (Database issue):D561-5. doi: 10.1093/nar/gks1114.
Wu, C, C Orozco, J Boyer, M Leglise, J Goodale, S Batalov, CL Hodge, J Haase, J Janes, JW Huss, and AI Su. 2009. "BioGPS: an extensible and customizable portal for querying and organizing gene annotation resources." Genome Biol 10 (11):R130. doi: 10.1186/gb-2009-10-11-r130.
Wu, Q, K Chambliss, M Umetani, C Mineo, and PW Shaul. 2011. "Non-nuclear estrogen receptor signaling in the endothelium." J Biol Chem 286 (17):14737-43. doi: 10.1074/jbc.R110.191791.
Wyckoff, MH, KL Chambliss, C Mineo, IS Yuhanna, ME Mendelsohn, SM Mumby, and PW Shaul. 2001. "Plasma membrane estrogen receptors are coupled to endothelial nitric-oxide synthase through Galpha(i)." J Biol Chem 276 (29):27071-6. doi: 10.1074/jbc.M100312200.
Yamada, H, I Furuta, EH Kato, S Kataoka, Y Usuki, G Kobashi, F Sata, R Kishi, and S Fujimoto. 2002. "Maternal serum and amniotic fluid bisphenol A concentrations in the early second trimester." Reprod Toxicol 16:735-739.
Yamamoto, M, M Shirai, K Sugita, N Nagai, Y Miura, R Mogi, K Yamamoto, A Tamura, and K Arishima. 2003. "Effects of Maternal Exposure to Diethylstilbestrol on the Development of the Reproductive System and Thyroid Function in Male and Female Rat Offspring." J Toxicol Sciences 28 (5):385-394.
Yan, S, Y Chen, M Dong, W Song, SM Belcher, and HS Wang. 2011. "Bisphenol A and 17beta-estradiol promote arrhythmia in the female heart via alteration of calcium handling." PLoS One 6 (9):e25455. doi: 10.1371/journal.pone.0025455.
Yang, N, H Shigeta, H Shi, and CT Teng. 1996. "Estrogen-related Receptor, hERR1, Modulates Estrogen Receptor-mediated Response of Human Lactoferrin Gene Promoter." J Biol Chem 271 (10):5795-5804.
Yap, J, LA Salamonsen, T Jobling, PK Nicholls, and E Dimitriadis. 2010. "Interleukin 11 is upregulated in uterine lavage and endometrial cancer cells in women with endometrial carcinoma." Reprod Biol Endocrinol 8:63. doi: 10.1186/1477-7827-8-63.
Ye, X, LY Wong, AM Bishop, and AM Calafat. 2011. "Variability of urinary concentrations of bisphenol A in spot samples, first morning voids, and 24-hour collections." Environ Health Perspect 119 (7):983-8. doi: 10.1289/ehp.1002701.
Yeo, SH, and AE Herbison. 2014. "Estrogen-negative feedback and estrous cyclicity are critically dependent upon estrogen receptor-alpha expression in the arcuate nucleus of adult female mice." Endocrinology 155 (8):2986-95. doi: 10.1210/en.2014-1128.

216
Yildizhan, B, E Uyar, A Sismanoglu, G Gulluoglu, and ZN Kavak. 2006. "Spontaneous perforation of pyometra." Infect Dis Obstet Gynecol 2006:26786. doi: 10.1155/IDOG/2006/26786.
Yin, Y, C Lin, GM Veith, H Chen, M Dhandha, and L Ma. 2012. "Neonatal diethylstilbestrol exposure alters the metabolic profile of uterine epithelial cells." Dis Model Mech 5 (6):870-80. doi: 10.1242/dmm.009076.
Yip, KS, A Suvorov, J Connerney, NJ Lodato, and DJ Waxman. 2013. "Changes in mouse uterine transcriptome in estrus and proestrus." Biol Reprod 89 (1):13. doi: 10.1095/biolreprod.112.107334.
Yiu, KW, CK Lee, KC Kwok, and NH Cheung. 2014. "Measuring the kinetics of the binding of xenoestrogens and estrogen receptor alpha by fluorescence polarization." Environ Sci Technol 48 (19):11591-9. doi: 10.1021/es503801c.
Yokota, H, H Iwano, M Endo, T Kobayashi, H Inoue, S Ikushiro, and A Yuasa. 1999. "Glucuronidation of the environmental oestrogen bisphenol A by an isoform of UDP-glucuronosyltransferase, UGT2B1, in the rat liver." Biochem J 340:405-409.
Yoshida, A, RR Newbold, and D Dixon. 1999. "Effects of Neonatal Diethylstilbestrol (DES) Exposure on Morphology and Growth Patterns of Endometrial Epithelial Cells in CD-1 Mice." Toxicologic Pathology 27 (3):325-333. doi: 10.1177/019262339902700308.
Yoshida, M, M Takahashi, K Inoue, S Hayashi, A Maekawa, and A Nishikawa. 2011. "Delayed adverse effects of neonatal exposure to diethylstilbestrol and their dose dependency in female rats." Toxicol Pathol 39 (5):823-34. doi: 10.1177/0192623311413785.
Young, VJ, JK Brown, PT Saunders, and AW Horne. 2013. "The role of the peritoneum in the pathogenesis of endometriosis." Hum Reprod Update 19 (5):558-69. doi: 10.1093/humupd/dmt024.
Zbucka, M, W Miltyk, T Bielawski, A Surazynski, J Palka, and S Wolczynski. 2007. "Mechanism of collagen biosynthesis up-regulation in cultured leiomyoma cells." Folia Histochemica et Cytobiologica 45 (Suppl 1):181-185.
Zhang, J, AL Hampton, G Nie, and LA Salamonsen. 2000. "Progesterone Inhibits Activation of Latent Matrix Metalloproteinase (MMP)-2 by Membrane-Type 1 MMP: Enzymes Coordinately Expressed in Human Endometrium." Biol Reprod 62:85-94.
Zhang, J, and LA Salamonsen. 1997. "Tissue inhibitor of metalloproteinases (TIMP)-1, -2, and -3 in human endometrium during the menstrual cycle." Mol Hum Reprod 3:735-741.
Zhang, S, PH Howarth, and WR Roche. 1996. "Cytokine Production by Cell Cultures from Bronchial Subepithelial Myofibroblasts." J Pathol 180:95-101.

217
Zhang, Z, K Chen, JC Shih, and CT Teng. 2006. "Estrogen-related receptors-stimulated monoamine oxidase B promoter activity is down-regulated by estrogen receptors." Mol Endocrinol 20 (7):1547-61. doi: 10.1210/me.2005-0252.
Zondek, B. 1936. "The Effect of Prolonged Application of Large Doses of Follicular Hormone on the Uterus of Rabbits." J Exp Med 63 (6):789-794.
Zorn, TMT, EMAF Bevilacqua, and PA Abrahamsohn. 1986. "Collagen remodeling during decidualization in the mouse." Cell Tissue Res 244:443-448.




















