
Characterization of blood pressure and morphological traits in cardiovascular-related organs in 13...
34
JAP-00073-2004.R1 1 FINAL ACCEPTED VERSION Characterization of blood pressure and morphologic traits in cardiovascular-related organs in 13 different inbred mouse strains Christian F. Deschepper*, Jean L. Olson + , Melissa Otis° and Nicole Gallo-Payet° *Experimental Cardiovascular Biology Research Unit, Institut de recherches cliniques de Montréal (IRCM), Montreal (QC) Canada H2W 1R7; + Department of Pathology, University of California at San Francisco, CA 94143, USA; ° Dept of Medicine, Endocrinology Service, University of Sherbrooke, Sherbrooke (QC) Canada J1H 5N4. Running head: Cardiovascular phenotypes in inbred mouse strains Corresponding author: Christian F. Deschepper IRCM 110, Pine Ave West Montréal (QC) Canada H2W 1R7 Tel: (514) 987 5759 Fax: (514) 987 5585 E-mail: [email protected] Articles in PresS. J Appl Physiol (March 26, 2004). 10.1152/japplphysiol.00073.2004 Copyright © 2004 by the American Physiological Society.
-
Upload
usherbrooke -
Category
Documents
-
view
0 -
download
0
Transcript of Characterization of blood pressure and morphological traits in cardiovascular-related organs in 13...

JAP-00073-2004.R1
1
FINAL ACCEPTED VERSION
Characterization of blood pressure and morphologic traits in cardiovascular-related
organs in 13 different inbred mouse strains
Christian F. Deschepper*, Jean L. Olson+, Melissa Otis° and Nicole Gallo-Payet°
*Experimental Cardiovascular Biology Research Unit, Institut de recherches cliniques de
Montréal (IRCM), Montreal (QC) Canada H2W 1R7; + Department of Pathology, University of
California at San Francisco, CA 94143, USA; ° Dept of Medicine, Endocrinology Service,
University of Sherbrooke, Sherbrooke (QC) Canada J1H 5N4.
Running head: Cardiovascular phenotypes in inbred mouse strains
Corresponding author: Christian F. Deschepper
IRCM
110, Pine Ave West
Montréal (QC) Canada H2W 1R7
Tel: (514) 987 5759
Fax: (514) 987 5585
E-mail: [email protected]
Articles in PresS. J Appl Physiol (March 26, 2004). 10.1152/japplphysiol.00073.2004
Copyright © 2004 by the American Physiological Society.

JAP-00073-2004.R1
2
ABSTRACT
To better understand the contributions of various genetic backgrounds to complex quantitative
phenotypes, we have measured several quantitative traits of cardiovascular interest [i.e. systolic
blood pressure, weight (corrected by body weight) of several cardiac compartments, of adrenals
and of kidneys, and histological correlates for kidneys and adrenals] in male and female mice
from 13 different inbred strains. The latter were selected in order to have representatives from
each major genealogical group and to conform to priorities set by the Mouse Phenome Database
project. Inter-strain comparisons of phenotypes made it possible to identify strains that displayed
values that belonged either to the low or the high end of the inter-strain variance for quantitative
traits such as systolic blood pressure, body weight, left ventricular weight and/or adrenocortical
structure. For instance, both male and female C3H/HeJ and A/J mice displayed either low
systolic blood pressure or low cardiac ventricular mass, respectively, and male C57Bl6/J
displayed low adrenal weights. Likewise, inter-sex comparisons made it possible to identify
phenotypic values that were sexually dimorphic for some of the same traits. For instance, female
AKR/J had relatively higher body weight and systolic blood pressure values than their male
counterparts, and may constitute an animal model of the metabolic X syndrome. These strain-
and sex-specific features will be of value either for future genetic and/or developmental studies
or to develop new animal models that will help in the generation of mechanistic hypotheses. All
data have been deposited to the Mouse Phenome Database for future integration with the Mouse
Genome Database, and can be further analyzed and compared with tools available on the site.

JAP-00073-2004.R1
3
INTRODUCTION
Cardiovascular diseases are the main cause of mortality and morbidity in industrialized societies
and remain the focus of intense research efforts. Although molecular biology has contributed
greatly to our understanding of the mechanisms of disease, all molecular data ultimately have to
be interpreted in the context of whole organs, hence the utility of inbred mouse strains. Indeed,
the genotypic and phenotypic diversity that exists between inbred strains make it possible to
generate precise linkage maps for traits of interest, and the possible roles of candidate genes can
be further explored thanks to the range of genetic manipulations that can be performed on mouse
embryos. However, it is becoming increasingly apparent that the effects of any gene
manipulation are highly dependent on the genetic background of the mouse strain that has been
used. A precise understanding of inter-strain phenotypic diversity therefore becomes an
indispensable complement of genomic databases. This need has prompted the birth of an
international consortium of partners (originating from both the academic and corporate sectors)
that is called the Mouse Phenome Database (MPD) project (http://www.jax.org/phenome). The
goals of the project are to collect reliable phenotypic data from mouse strains, and organize them
in a central, web-accessible database housed at The Jackson Laboratory (TJL), so that it can be
integrated with the Mouse Genome Database. The premise is that such phenotypic data are
essential for realizing the full utility of genomic information that will emerge from sequencing
the mouse genome.
We set out to measure several quantitative traits that include systolic blood pressure, weight
(corrected by body weight) of several cardiac compartments, of adrenals and of kidneys, and
histological correlates for kidneys and adrenals of 13 different inbred mouse strains. The latter

JAP-00073-2004.R1
4
were selected in order to have representatives from each major genealogical group (4) and to
conform to priorities set by the MPD project. In addition to providing the MPD with data of
cardiovascular interest, our hope was that such a systematic characterization might lead to the
identification of new mouse models of complex traits that could be used either to generate new
hypotheses or to perform further genetic and/or mechanistic studies.
Of note, values of systolic blood pressure and of kidney weights have been reported previously
for several inbred strains (21;22). However, these studies have been performed at a time when
viral infections (that can affect several phenotypes, including organ weights) were prevalent
among mice colonies. Likewise, blood pressures were measured in untrained mice using a
manual tail cuff apparatus. Since then, a novel device has been developed where tail blood flow
is evaluated photoelectronically, cuff inflation is automated and performed at random intervals,
and all data recorded electronically (15). That method has been used in the current study.
The complete list of data collected as a result of this project has been posted on the MPD Web
site (http://aretha.jax.org/pub-cgi/phenome/mpdcgi?rtn=projects/details&id=104), along with
built-in tools to analyze the data and visualize them in different ways. In the present manuscript,
we highlight the results of inter-strain comparisons (to identify strains that displayed values that
belonged either to the low or the high end of the inter-strain variance), as well as those of inter-
sex comparisons (to identify phenotypic values that may be sexually dimorphic), and comment
the results of these comparisons in light of their utility and in the context of other data already
present in the literature.

JAP-00073-2004.R1
5
MATERIAL AND METHODS
Mouse models: Thirteen strains of inbred mice originated from The Jackson Laboratories
(TJL) and were shipped at 8 weeks of age. For each strain, we received 10 male and 10 female
individuals. Strains were selected in order to have representatives from each major genealogical
group (see Tables 1 and 2) and according to priorities set by the MPD project. After arrival, the
mice were housed in the animal facility at the “Institut de recherches cliniques de Montréal”
(IRCM), and maintained on the same chow as that used at TJL (LabDiet 5K54 for DBA/2J,
LabDiet 5K52 for all other strains).
Systolic blood pressure measurements: After 7 days of acclimation in the IRCM animal
facilities, systolic blood pressure (SBP) was measured daily in conscious mice (between 10 AM
and 1 PM) for 10 continuous days using the Visitech BP-2000 system (Apex, NC), as described
previously (15). The first 6 days of measurements were mostly for training purposes. Data
collected during these days were not used for calculations, but were used to check that reliable
flow waveforms could be obtained in each mouse. During the following 2-3 days of recording,
sets of 10 individual measurements were recorded. The final systolic value was computed as the
average of the 20-30 measurements obtained during the last 2-3 days.
Tissue collection: Whole body weight (BW) was measured the day of arrival and the day of
sacrifice. All mice were sacrificed at 10 weeks of age (16 days after their arrival). Organs
collected were the thymus, the heart, and both kidneys and adrenals. The hearts were dissected
into 4 parts: free wall of the right ventricle (RV), left ventricle (LV; comprising the

JAP-00073-2004.R1
6
interventricular septal wall), left and right atrium (LA and RA). Each tissue sample was weighed
individually, and divided by the value of whole body weight of the corresponding animal to
generate a weight index value (RV/BW, LV/BW, RA/BW, LA/BW, respectively). Likewise, the
weight of the right kidney (RK) and of the combined left and right adrenals (bi-adr) were
determined and corrected by BW to generated corresponding weight index values (RK/BW and
bi-adr/BW, respectively).
Histology: Adrenals and kidneys were collected, and kidneys were sectioned either
longitudinally or coronally. Both types of tissues were fixed by overnight immersion in a
solution of 4% paraformaldehyde in phosphate buffered saline, and then processed for paraffin
embedding. For kidneys, sections were cut at 5 µm, then were stained with either hematoxylin
and eosin (H&E) or periodic acid-Schiffs (PAS) and examined. The slides were assessed for
changes in the appearance of either glomeruli, tubules, interstitium or vessels. Glomeruli were
evaluated for changes in overall size, in the volume occupied by the mesangium, and in the
appearance of the juxtaglomerular apparatus (JGA) was also noted. For adrenals, sections were
cut at 3 µm, stained with H&E, and mounted with non-aqueous VectaMount mounting media,
(Vector Laboratories, Burlingame, CA, USA). Images were observed using a Nikon Eclipse 300
microscope equipped with a CoolSnap color digital camera. Acquired images were processed
and analyzed using Adobe Photoshop 4.0.
Distribution of values: For each phenotypic measurement, corresponding values from all
13 strains were averaged to calculate mean and standard deviation (SD). Assuming that the 13
strains represented a random and normally distributed sample of all values found among all

JAP-00073-2004.R1
7
laboratory strains, we then transformed each value obtained for a given strain (µ) into the Z
normal deviate, where Z = (mean - µ)/SD. We then estimated to which percentile each value
belonged on the basis of the normal distribution of Z normal deviates.
RESULTS
Systolic blood pressure:
SBP values ranged from 103.4 to 139 mm Hg in males, and from 102.6 to 134.7 mm Hg in
females (Table 1 & 2). In male and female C3H/HeJ mice, SBP values corresponded to the
lower 5.2% and 3.4 % percentile, respectively. For both sexes, high SBP values were observed
for wild derived-strains. In males, SBP corresponded to the higher 91% and 98.4 % percentile of
the normal distribution for PWK/Ph and CAST/Ei, respectively. In females, SBP was high only
in PWK/Ph, where it corresponded to the high 94 % percentile of the normal distribution. Inter-
sex comparisons for all strains revealed that for 9 out of the 13 strains, there was a significant (p
< 0.001) linear correlation (r2 = 0.92) between values from males and females (Fig. 2). Four
other strains had SBP values that were outside of the 99% confidence interval of the correlation
between the values of males and females (Fig. 2), thus identifying strains where SBP values were
sexually dimorphic. These strains corresponded to 129S1/SvlmJ and CAST/Ei mice (where SBP
was higher in males than in females), and to AKR/J and C57Bl/6J mice (were SBP values were
higher in females than in males).
Gross morphology:

JAP-00073-2004.R1
8
The BW of the wild-derived strains were much lower than that of the other laboratory strains, as
BW of CAST corresponded to the lower 1% and 3.2 % percentile of the normal distribution of
values among in males and females, respectively, whereas the BW of PWK/Ph corresponded to
the lower 2.6% and 7 % percentile of the normal distribution of values among males and
females, respectively (Tables 1 & 2). Inter-sex comparison of the BW values from all 13 strains
demonstrated a highly significant (r2 = 0.79, P < 0.01) linear correlation. However, it appeared
that there was sexual dimorphism for the AKR/J strain, since BW from AKR/J females
corresponded to the higher 98.8% percentile of the normal distribution of values among female
strains, and the corresponding point was above the 99% higher confidence interval of the linear
regression of male vs. female values. When the AKR/J strain was excluded from the male vs.
female comparison, a better correlation score (r2 = 0.95, P < 0.001) was obtained (Fig. 2).
For hearts, the lowest LV/BW values for both males and females were those found in the A/J
strain, where they corresponded to the lower 7.6 % and 13.1 % percentile of the normal
distribution in males and females, respectively. Conversely, the highest values were found in
PWK, were they corresponded to the higher 97.2 % and 98.4 % percentile of the normal
distribution in males and females, respectively. Since SBP may influence LV/BW within any
particular strain, we tested whether there was a correlation between both values among all strains
in either male or female mice. For males, there was a significant correlation (r2 = 0.38, P < 0.05)
between both variables, although the values of 2 strains (A/J and FVB/NJ) were below the
boundaries of the 99% confidence interval (Fig 3a). When values from these 2 strains were
excluded from the linear regression, a better correlation score (r2 = 0.49, P < 0.05) was obtained.
In contrast, we found no significant correlation between LV/BW and SBP in female strains.

JAP-00073-2004.R1
9
When we compared the values of LV/BW between males and females, there was a highly
significant linear correlation (r2 = 0.76, P < 0.01) between the values of all strains (Fig. 3b).
Only one strain (AKR/J) had a LV/BW ratio that was below the 99% lower confidence interval
of the regression. Of note, this was likely due to the fact that body weight was also sexually
dimorphic in the AKR/J (Fig. 2). When the value from the latter strain was excluded from the
linear regression, the inter-sex correlation score did improve (r2 = 0.88, P < 0.01).
Macroscopically, the RV of males and female DBA/2J mice were found to contain calcifications,
which probably explained why RV/BW was higher than the 96.6% and 94.5 % for male and
female DBA/2J mice, respectively (Tables 1 and 2). However, RV/BW was also found to be
high for PWK mice, as it was higher than the 97 % and 94 % percentile in males and females,
respectively, in the absence of any other obvious anomaly.
For kidneys, there was generally no significant differences between the weight of left and right
kidney, with the exception of male C57Bl/6J mice where the corrected weight of the left kidney
was significantly lower than that of the right one (3.45 ± 0.29 vs. 5.93 ± 0.76, mean ± S.D.).
Consequently, inter-strain comparisons were performed only for the corrected weight of RK. In
males, RK/BW was also low (as compared to other strains) in C57Bl/6 mice, as it corresponded
to the lower 3 % percentile of the normal distribution of the RKW/BW values across all strains.
In contrast, RK/BW was high in male C3H/HeJ mice, since it corresponded to the higher 93 %
percentile of the normal distribution of the RK/BW values across all strains. In females, the
most divergent values were found only in the wild-derived strains, as the values of RK/BW
corresponded to the higher 95 % and 98 % percentiles for PWK and CAS mice, respectively.
These high kidney weights in female wild-derived mice reflected a sexual dimorphism, since the

JAP-00073-2004.R1
10
RK/BW values were significantly higher than in their male counterparts for these 2 strains.
There was no correlation between the values of RK/BW and SBP in either males or females.
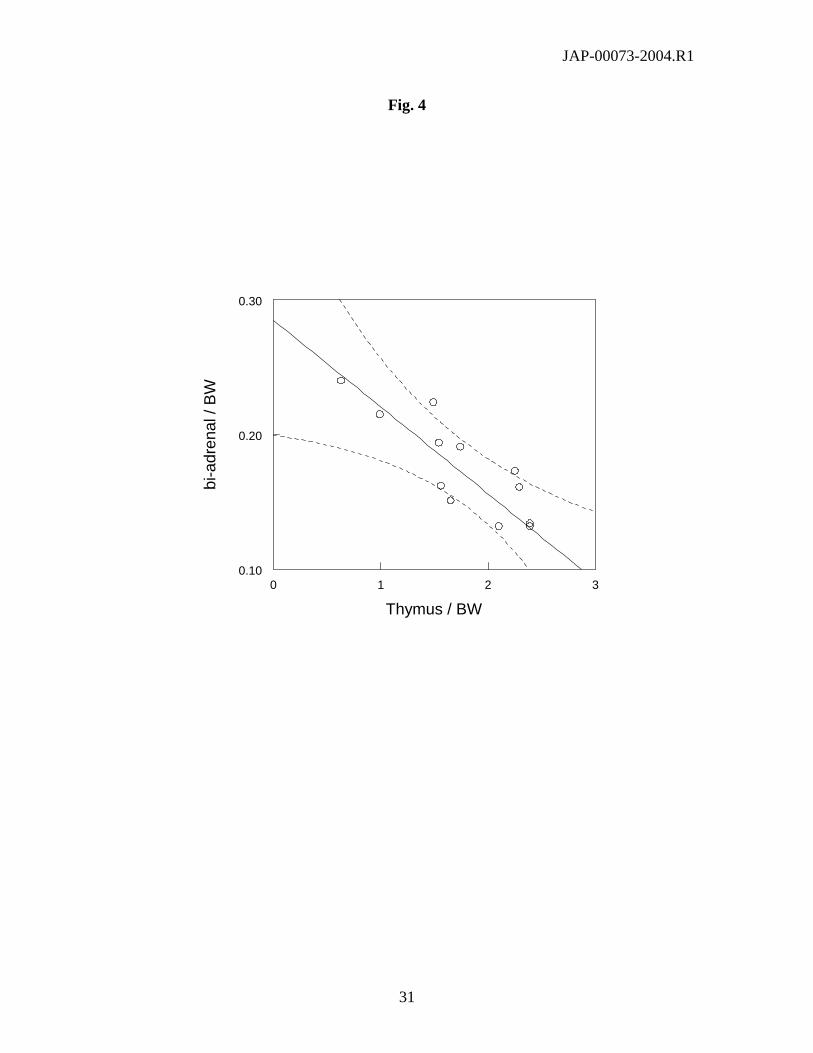
In males, the bi-adr/BW values spanned a 3-fold range, and belonged to the lower 2% percentile
or higher 95% percentile of the normal distribution of values among male strains for either
C57Bl/6J or PWK/Ph mice, respectively. There was a significant linear correlation (r2 = 0.70, P
< 0.01) between the values of bi-adr/BW and those of corrected thymus weight (Fig. 4).
Adrenals from females were macroscopically different than that of males, since female adrenals
were generally larger and had a whitish appearance.
Histology:
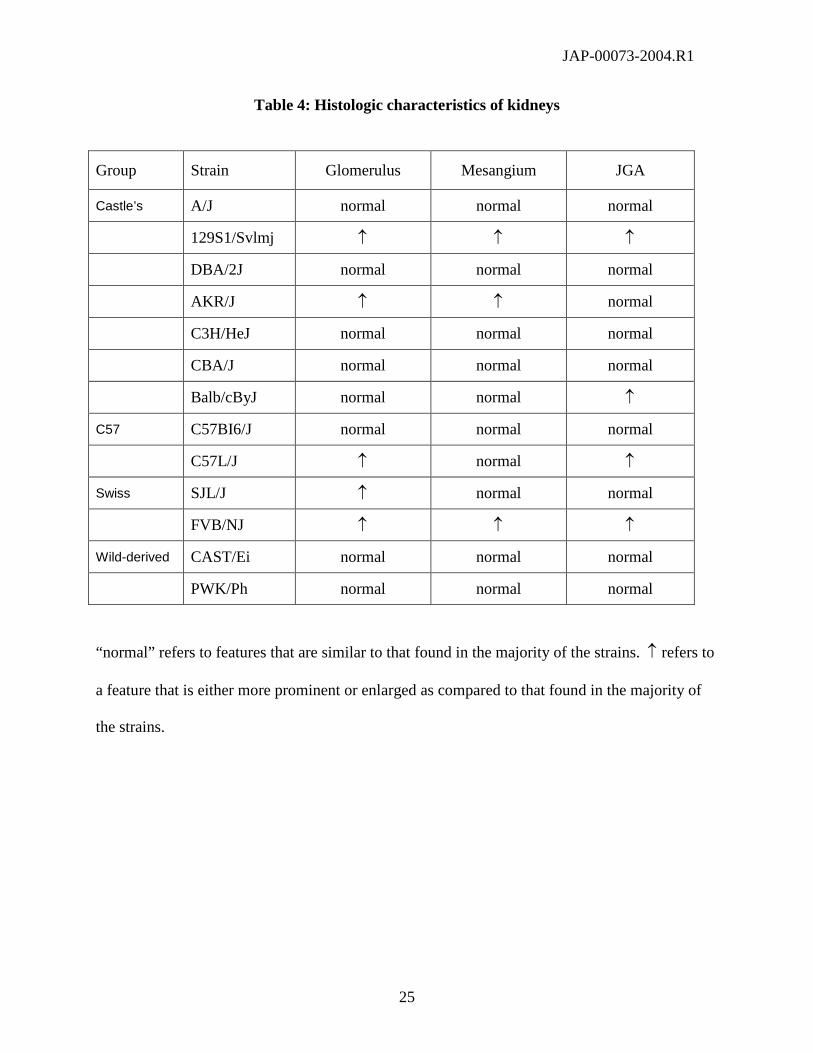
For kidneys, there was no particular histological feature that could account for differences in
gross kidney weight between strains. In particular, tubules, interstitium and vessels were
unremarkable in all groups. However, some differences were observed in kidney cortices, as
some strains displayed either larger glomeruli, an increase of the mesangial compartment (with
increased mesangial cellularity and mesangial matrix), or enlarged juxtaglomerular appariti
(Table 4). With the exception of glomerular size (which was increased in both strains from the
Swiss genealogical group), there was no particular pattern within groups.
For adrenals, apart from a well-known difference in size between males and females, adrenal
cortices from all strains all clearly exhibit the 3 zones usually found in all vertebrates, i.e. the
zona glomerulosa (ZG), zona fasciculata (ZF) and zona reticularis (ZR) (Fig. 5). As in the rat,
there was a clear demarcation in all strains between ZG and ZF due to 1) specific arrangements

JAP-00073-2004.R1
11
of ZG as glomeruli or loops, and of ZF cells in fascicular columns, 2) the presence of larger lipid
droplets in ZF than in ZG, and 3) the presence in some strains, as reported previously (17), of an
intermediate zone between ZG and ZF. In addition, a so-called X zone of variable size could be
distinguished in adrenals from male and female of some strains (Figs. 6 and 7) (Table 3).
Finally, one additional zone (that we call “lipoid zone”) was seen only in female representatives
from some of the strains (Fig. 5 and 7) (Table 3). The relative importance of each zone varied
across strains, as summarized in Table 3.
DISCUSSION:
When we compared the distribution of phenotypic values among 13 different inbred strains, we
typically found that for each variable, there was one or 2 strains that displayed values that were
at one extremity of the normal distribution. By proceeding in this fashion, it was possible to
identify inbred strains that had phenotypic characteristics that were very different from that of
most other strains. Comparisons of values between males and females from each strain also
made it possible to identify sexually dimorphic traits. All data are summarized in Tables 1 and
2, and have been posted MPD Web site (http://aretha.jax.org/pub-
cgi/phenome/mpdcgi?rtn=projects/details&id=104). The same site comprises tools to visualize
“outliers” (which makes it possible to identify strains that display values of given traits that are
particularly high or low as compared to other strains), and to compare data as a function of
chosen criteria (strain, sex, etc.).
Body weight was much lower in the wild-derived strains (CAST/Ei and PKW/Ph) than in any of
the other laboratory mouse strains. At the other end of the range of phenotypic values, body
weight was highest in male AKR/J mice. There was also sexual dimorphism, as the average BW

JAP-00073-2004.R1
12
value of AKR/J mice deviated from the inter-strain average to a greater extent in females than in
males. Of note, it has been reported that the male AKR/J mice were the most sensitive to the
development of dietary obesity (when compared to males from 8 other strains of inbred mice)
(32). The sexual dimorphism detected in the present study suggests that the female hormonal
environment interacts in a particular way with the obesity-prone background of AKR/J mice.
Moreover, since female AKR/J mice have higher values of both body weight and systolic blood
pressure than their male counterparts, it is possible that female AKR/J mice constitute a so far
unrecognized animal model of the so-called metabolic X-syndrome, which is comprised of a
combination of symptoms that include obesity, hypertension and insulin-resistance (10). Of
note, it has usually been considered in other animal models of the metabolic syndrome that the
phenotype was less severe in females than in males (30), which is contrary to what we have
observed with AKR/J mice. However, it has also been found that one of the loci linked to
obesity in AKR mice was located on chromosome X (33). The current finding that the metabolic
phenotype is more severe in female than in male AKR/J suggests that the male chromosome Y
contains a locus whose effect counteracts that of the locus on chromosome X. This possibility
had indeed been considered previously (33).
In male mice, the lowest and highest values for SBP were observed with the C3H/HeJ and the
wild-derived strains (respectively). In female mice, the lowest and highest values for SBP were
observed with the C3H/HeJ and only one of the wild-derived strains (i.e. PKW/Ph). Of note, a
distribution of systolic blood pressure values has been reported previously for 19 strains of
inbred mice (23). However, the values from that previous report cannot been readily compared
to those reported in the current study, because values from the previous report had been obtained

JAP-00073-2004.R1
13
with aging retired breeders using a different experimental device to measure blood pressure. The
correlations we have observed for the SBP values between males and females from most strains
or for the values of LV/BW and SBP in male strains provide independent validations for the
reliability of our measurements. Of note, others had reported that the SBP values (measured
with an apparatus that was identical to the one used by ourselves) of 8-week old male Balb/cByJ
and CBA/J mice were 104 and 96 mm Hg, respectively(25), whereas we detected no difference
between 10 week-old mice from both strains. We do not know whether such differences are due
to differences in the age of the animals or in experimental and/or housing conditions.
Cardiac ventricular weight is a variable of particular interest, because it has been shown in
human epidemiological studies that cardiac ventricular mass is one of the most important
independent predictor of cardiovascular mortality and morbidity (7;9;16). Although there is a
paucity of information about cardiac mass in mouse strains, others have reported that the left
ventricular weight index of male C57Bl/6 mice was 27 % greater than that of male A/J mice
(12). Although the absolute values of LV/BW were different between that study and our own
(presumably because of differences in the way of separating the left and right ventricles), we
report the same 27 % difference between the values of LV/BW of male mice from both strains.
Likewise, others had reported (like us) that cardiac mass was greater in CBA/J than in Balb/cByJ
male mice (25). However, direct comparisons between both studies are not possible because
values in the latter study were not corrected for body weight (25). Concerning the RV, male and
female DBA/2J mice had the particular feature of displaying readily visible calcifications on the
surface of the RV. This finding has been described previously (20), and is considered to
constitute the sequel of spontaneous eosinophil-mediated myocarditis in this particular strain
(11).

JAP-00073-2004.R1
14
Since blood pressure is one factor that may influence the value of LV/BW within any given
strain, we first sought to determine whether we could detect any correlation between both
variables across strains. Among males from all strains, we detected a weak but significant
correlation between both variables. However, there was no correlation between the two
variables when inter-strain comparisons were performed for female individuals. In contrast,
there was a very tight correlation between the values of LV/BW from males and females, which
indicates that genetic background accounts for a greater part of the variance of LV/BW among
inbred strains than systolic blood pressure. These findings are compatible with those obtained by
others with 23 inbred strains of rats (29). In the latter report, no correlation had been found
between cardiac mass and SBP (when the latter was smaller than 140 mm Hg), but is has been
calculated that the degree of genetic determination for cardiac mass was between 65 and 75 %
(29).
Considerable inter-strain variability was also found for adrenal weight among male mice. Of
note, it has been reported that within a given strain of male rats, the trophic effects of ACTH on
the adrenal fasciculata were the strongest determinant of changes in adrenal weight (2).
Conversely, corticosteroids have well-documented thymolytic effects, so that thymus weight has
been reported to be inversely related to circulating levels of corticosteroids within a given rat
strain (1). The present data show that similar correlations can be found between the weight of
thymuses and adrenals of male mice belonging to several inbred strains. It is therefore likely that
differences in either adrenal or thymus weight reflect primarily different average levels of
activation of the hypothalamo-pituitary-adrenal axis. Interestingly, others who had measured
biologic responses to stress in different mouse strains have referred to C57Bl/6J as stress-
resistant and to BalB/cByJ as stress-reactive mice (3;24); these 2 particular strains were the ones

JAP-00073-2004.R1
15
we found to be at the lower and higher end of the distribution of adrenal weight values,
respectively. Likewise, others had reported strain-related differences among inbred mouse
strains for the values of corrected thymus weight, with this variable being highest in C57Bl/6 and
AKR/J mice. This report is compatible with our findings, since we found that male mice from
these 2 strains both had corrected adrenal weights at the lower end of the distribution of
corresponding values. It also has been reported that thymus size was under genetic control, as it
could be linked to one quantitative trait locus (designated Tsz-1) on chromosome 5 in
recombinant inbred strains derived from C57Bl/6 and A/J mice (19). It is therefore possible that
this locus relates in fact to responsiveness of the hypothalamo-pituitary-adrenal axis.
It also has been reported that adrenocortical structure may vary across mouse strains (18;26).
Strains whose general adrenocortical architecture corresponded most closely to the general
histological features described in the literature for male and female mice were those belonging to
the Cast/Ei, C57Bl/6J and Balb/cByJ strains (17). We also found that the relative importance of
the zona reticularis varied according to the sex and the strain of each mice. In addition to the 3
traditional adrenocortical zones, one feature that is found in the adrenals of adult mice is the so-
called “X-zone”, as first reported by Howard (13). Although the X-zone is not found in the
adrenals of other adult vertebrates, it has been recently pointed out that it resembles the human
adrenal fetal zone, and hypothesized that the murine X-zone might in fact be homologous to the
“fetal cortex” of human adrenals (31). In male mice, the X-zone can be seen in immature male
mice, but it typically disappears at puberty at ~6 weeks of age (8). In females, the X-zone
remains usually visible and distinct from the ZR in virgin animals, but it regresses after
pregnancy (8). These dynamic changes in the X-zone appear to be controlled by hypophyseal
gonadotropic hormones, since all degenerative changes are abolished after gonadectomy in both

JAP-00073-2004.R1
16
sexes (8). However, we found that the presence or absence of the X zone is not uniform in adult
mice from different strains, since it was still detectable in males from 3 strains (Balb/cByJ and
the 2 wild-derived strains), and it could no longer be seen in virgin females from 2 other strains
(A/J and C3H/HeJ)(Table 3). Of note, we have looked only at 10 week-old mice. Since the X
zone is a dynamic structure that evolves according to age and/or endocrine status, a
reinvestigation of the same strains at different ages may therefore reveal differences in the age-
related changes of the X-zone. The interest of these findings may reside in the fact that
experiments with inbred strains or genetic crosses have shown that X-zone development is under
the control of genes within specific loci (14;27;28). Some of these genes include SF-1, DAX-1
and ACD (5). The strains identified in Table 3 may therefore be useful for genetic studies aimed
at studying the roles of such genes in X zone development.
In addition to the X-zone, some female mice displayed a zone (that we called the lipoid zone)
that was not present in male mice. This zone has not been described previously, but has been
shown in a figure of a recent paper showing a section from the adrenal of a female DBA/2J
mouse (6). One possibility is that this zone derives from the degenerescence of other
adrenocortical cells, but further investigations are needed to determine whether this is the case.
Altogether, the data commented in the present manuscript extend the body of data already
present in the MPD by identifying mouse strains with phenotypic characteristics of particular
interest. It is expected that such strains will proved useful in further studies aimed at identifying
loci linked to this particular phenotypes or at studying the roles of certain genes in the course of
development, or to generate new animal models of disease that will help to formulate novel
mechanistic hypotheses.

JAP-00073-2004.R1
17
Acknowledgements:
Animals used in this study have been donated by The Jackson Laboratories and the Mouse
Phenome Database Project. Generous funds from AstraZeneca were used to defray the cost of
mice through the Mouse Phenome Project Collaboration Program. This work has been supported
by the NIH/NHLBI grant HL69122 (to CFD), by a Group Grant of the Canadian Institutes for
Health Research (CIHR) to the IRCM Multidisciplinary Research Group in Hypertension, and by
CIHR grant MPO-10998 (to NGP). NGP is a recipient of a Canada Research Chair in
Endocrinology of the Adrenal Gland.
References:
1. Akana, S., C. S. Cascio, J. Shinsako, and M. F. Dallman. Corticosterone: narrow range
required for normal body and thymus weight and ACTH. Am.J.Physiol. 249: R527-R532,
1985.
2. Akana, S. F., J. Shinsako, and M. F. Dallman. Relationships among adrenal weight,
corticosterone, and stimulated adrenocorticotropin levels in rats. Endocrinology 113: 2226-
2231, 1983.
3. Anisman, H., S. Lacosta, P. Kent, D. C. McIntyre, and Z. Merali. Stressor-induced
corticotropin-releasing hormone, bombesin, ACTH and corticosterone variations in strains
of mice differentially responsive to stressors. Stress 2: 209-220, 1998.
4. Beck, J. A., S. Lloyd, M. Hafezparast, M. Lennon-Pierce, J. T. Eppig, M. F. Festing, and E.
M. Fisher. Genealogies of mouse inbred strains. Nat.Genet. 24: 23-25, 2000.

JAP-00073-2004.R1
18
5. Beuschlein, F., C. E. Keegan, D. L. Bavers, C. Mutch, J. E. Hutz, S. Shah, Y. M. Ulrich-
Lai, W. C. Engeland, B. Jeffs, J. L. Jameson, and G. D. Hammer. SF-1, DAX-1, and ACD:
molecular determinants of adrenocortical growth and steroidogenesis. Endocr.Res. 28: 597-
607, 2002.
6. Bielinska, M., H. Parviainen, S. B. Porter-Tinge, S. Kiiveri, E. Genova, N. Rahman, I. T.
Huhtaniemi, L. J. Muglia, M. Heikinheimo, and D. B. Wilson. Mouse strain susceptibility
to gonadectomy-induced adrenocortical tumor formation correlates with the expression of
GATA-4 and luteinizing hormone receptor. Endo 144: 4123-4133, 2003.
7. de Simone, G., P. Verdecchia, S. Pede, M. Gorini, and A. P. Maggioni. Prognosis of
inappropriate left ventricular mass in hypertension. The MAVI study. Hypertension 40:
470-476, 2002.
8. Deacon, C. F., W. Mosley, and I. C. Jones. The X zone of the mouse adrenal cortex of the
swiss albino strain. Gen.Comp.Endocrinol. 61: 87-99, 1986.
9. Devereux, R. B., G. de Simone, A. Ganau, and M. J. Roman. Left ventricular hypertrophy
and geometric remodeling in hypertension: stimuli, functional consequences and prognostic
implications. J.Hypertens. 12 (suppl. 10): S117-S127, 1994.
10. Grundy, S. M., H. B. Jr. Brewer, J. I. Cleeman, S. C. Jr. Smith, and C. Lenfant. Definition
of the metabolic syndrome. Circulation 109: 433-438, 2004.
11. Hirasawa, M., Y. Kitaura, H. Deguchi, A. Ukimura, and K. Kawamura. Spontaneous
myocarditis in DBA/2 mice. Virchows Arch. 432: 461-468, 1998.
12. Hoit, B. D., S. Kiatchoosakun, J. Restivo, D. Kirkpatrick, K. Olszens, H. Shao, Y.-H. Pao,
and J. H. Nadeau. Naturally occurring variation in cardiovascular traits among inbred
strains of mice. Genomics 79: 679-685, 2003.

JAP-00073-2004.R1
19
13. Howard, E. The representation of the adrenal X-zone in rats, in light of the observations on
the X-zone variability in mice. J.Anat.(Lond.) 62: 351-373, 1938.
14. Janat, M. F. and J. G. M. Shire. The adrenal X-zone of mice: genetic analysis of its
development with recombinant-inbred strains. Exp.Biology 46: 217-221, 1987.
15. Krege, J. H., J. B. Hodgin, J. R. Hagaman, and O. Smithies. A noninvasive computerized
tail-cuff system for measuring blood presure in mice. Hypertension 25: 1111-1115, 1995.
16. Levy, D., R. J. Garrison, D. D. Savage, W. B. Kannel, and W. P. Castelli. Prognostic
implications of echocardiographically determined left ventricular mass in the Framingham
heart study. N.Engl.J.Med. 322: 1561-1566, 1990.
17. Nussdorfer, G. Cytophysiology of the adrenal cortex: the adrenal cortex of normal adult
vertebrates. Int.Rev.Cytol. 98: 1-71, 1986.
18. Pawlus, M. Genetic differences in mouse adrenocortical structure. Folia
Histochem.Cytochem. 21: 239-251, 1983.
19. Peleg, L. and M. N. Nesbitt. Genetic control of thymus size in inbred mice. J.Heredity 75:
126-130, 1984.
20. Rings, R. W. and J. E. Wagner. Incidence of cardiac and other soft tissue mineralized
lesions in DBA/2 mice. Lab.Animal Sci. 344-352, 1971.
21. Schlager, G. Systolic blood pressure in eight inbred strains of mice. Nature 212: 519-520,
1966.
22. Schlager, G. Kidney weight in mice: strain differences and genetic determination.
J.Heredity 59: 171-174, 1968.

JAP-00073-2004.R1
20
23. Schlager, G. and R. S. Weibust. Genetic control of blood pressure in mice. Genetics 55:
497-506, 1967.
24. Shanks, N., J. Griffiths, S. Zalcman, R. M. Zacharko, and H. Anisman. Mouse strain
differences in plasma corticosterone following uncontrollable footshock.
Pharmacol.Biochem.Behavior 36: 515-519, 1990.
25. Sugiyama, F., G. A. Churchill, R. Li, L. J. M. Libby, T. Carver, K.-I. Yagami, S. W. M.
John, and B. Paigen. QTL associated with blood pressure, heart rate, and heart weight in
CBA/CaJ and BalB/cJ mice. Physiol.Genomics 10: 5-12, 2002.
26. Tanaka, S. and A. Matsuzawa. Comparison of adrenocortical zonation in C57BL/6J and
DDD mice. Exp.Anim. 44: 285-291, 1995.
27. Tanaka, S., M. Nishimura, J. Kitoh, and A. Matsuzawa. Strain difference of the adrenal
cortex between A/J and SM/J mioce, progenitors of SMxA recombinant inbred group.
Exp.Anim. 44: 127-130, 1995.
28. Tanaka, S., M. Nishimura, and A. Matsuzawa. Genetic association between Agouti locus
and adrenal X-zone morphology in SM/J mice. Acta Anat. 149: 170-173, 1994.
29. Tanase, H., Y. Yamori, C. T. Hansen, and W. Lovenberg. Heart size in inbred strains of
rats. Part 1. Genetic determination of the development of cardiovascular enlargement in
rats. Hypertension 4: 864-872, 1982.
30. Tofovic, S. P. and E. K. Jackson. Rat models of the metabolic syndrome. In Goligorsky, M.
S., ed. Methods in Molecular Medicine, vol. 86: Renal Disease: Techniques and Protocols.
Totowa,NJ, Humana Press. 2002, 29-46.
31. Vinson, G. P. Adrenocortical zonation and ACTH. Microsc.Res.Tech. 61: 227-239, 2003.

JAP-00073-2004.R1
21
32. West, D. B., C. N. Boozer, D. L. Moody, and R. L. Atkinson. Dietary obesity in nine
inbred mouse strains. Am.J.Physiol. 262: R1025-R1032, 1992.
33. York, B., K. Lei, and D. B. West. Inherited non-autosomal efects on body fat in F2 mice
derived from AKR/J x SWR/J cross. Mamm.Genome 8: 726-730, 1997.
JAP-00073-2004.R1
22
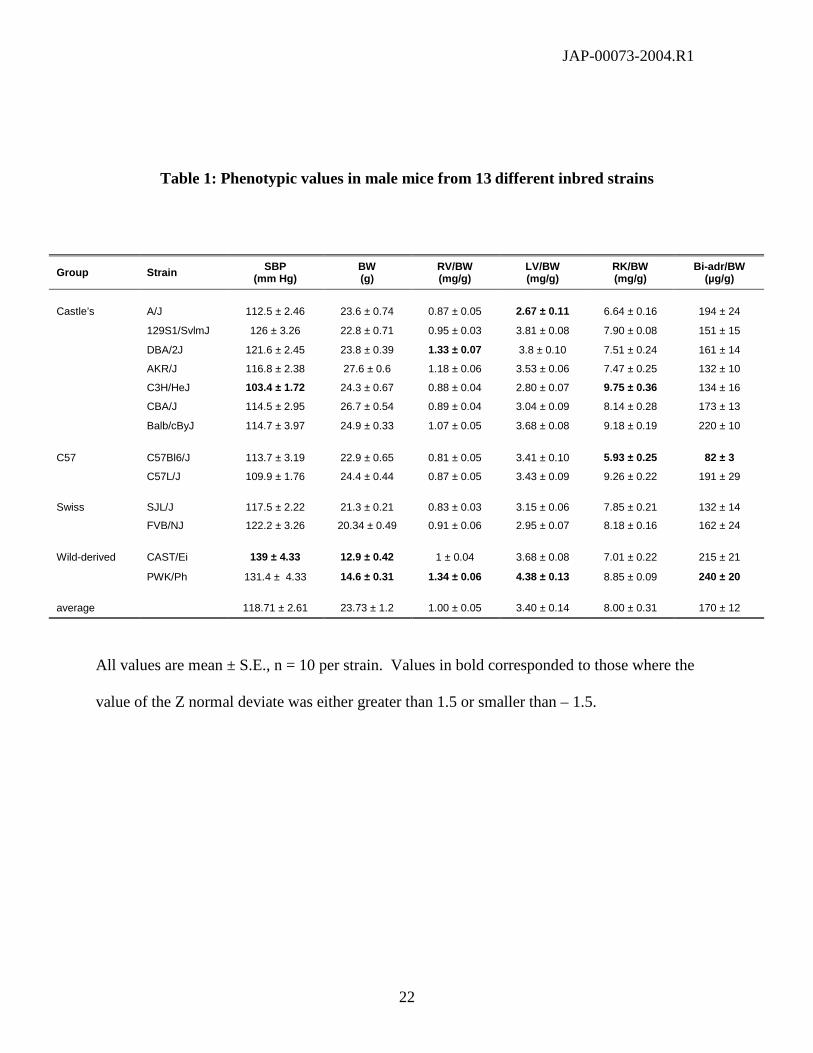
Table 1: Phenotypic values in male mice from 13 different inbred strains
All values are mean ± S.E., n = 10 per strain. Values in bold corresponded to those where the
value of the Z normal deviate was either greater than 1.5 or smaller than – 1.5.
Group Strain SBP(mm Hg)
BW(g)
RV/BW(mg/g)
LV/BW(mg/g)
RK/BW(mg/g)
Bi-adr/BW(µg/g)
Castle’s A/J 112.5 ± 2.46 23.6 ± 0.74 0.87 ± 0.05 2.67 ± 0.11 6.64 ± 0.16 194 ± 24
129S1/SvlmJ 126 ± 3.26 22.8 ± 0.71 0.95 ± 0.03 3.81 ± 0.08 7.90 ± 0.08 151 ± 15
DBA/2J 121.6 ± 2.45 23.8 ± 0.39 1.33 ± 0.07 3.8 ± 0.10 7.51 ± 0.24 161 ± 14
AKR/J 116.8 ± 2.38 27.6 ± 0.6 1.18 ± 0.06 3.53 ± 0.06 7.47 ± 0.25 132 ± 10
C3H/HeJ 103.4 ± 1.72 24.3 ± 0.67 0.88 ± 0.04 2.80 ± 0.07 9.75 ± 0.36 134 ± 16
CBA/J 114.5 ± 2.95 26.7 ± 0.54 0.89 ± 0.04 3.04 ± 0.09 8.14 ± 0.28 173 ± 13
Balb/cByJ 114.7 ± 3.97 24.9 ± 0.33 1.07 ± 0.05 3.68 ± 0.08 9.18 ± 0.19 220 ± 10
C57 C57Bl6/J 113.7 ± 3.19 22.9 ± 0.65 0.81 ± 0.05 3.41 ± 0.10 5.93 ± 0.25 82 ± 3
C57L/J 109.9 ± 1.76 24.4 ± 0.44 0.87 ± 0.05 3.43 ± 0.09 9.26 ± 0.22 191 ± 29
Swiss SJL/J 117.5 ± 2.22 21.3 ± 0.21 0.83 ± 0.03 3.15 ± 0.06 7.85 ± 0.21 132 ± 14
FVB/NJ 122.2 ± 3.26 20.34 ± 0.49 0.91 ± 0.06 2.95 ± 0.07 8.18 ± 0.16 162 ± 24
Wild-derived CAST/Ei 139 ± 4.33 12.9 ± 0.42 1 ± 0.04 3.68 ± 0.08 7.01 ± 0.22 215 ± 21
PWK/Ph 131.4 ± 4.33 14.6 ± 0.31 1.34 ± 0.06 4.38 ± 0.13 8.85 ± 0.09 240 ± 20
average 118.71 ± 2.61 23.73 ± 1.2 1.00 ± 0.05 3.40 ± 0.14 8.00 ± 0.31 170 ± 12

JAP-00073-2004.R1
23
Table 2: Phenotypic values in female mice from 13 different inbred strains
All values are mean ± S.E., n = 10 per strain. Values in bold corresponded to those where the
value of the Z normal deviate was either greater than 1.5 or smaller than – 1.5.
Group Strain SBP(mm Hg)
BW(g)
RV/BW(mg/g)
LV/BW(mg/g)
RK/BW(mg/g)
bi-adr/BW(µg/g)
Castle’s A/J 109.6 ± 2 17.6 ± 0.3 0.92 ± 0.04 3.05 ± 0.04 6.60 ± 0.12 301 ± 15
129S1/SvlmJ 115.9 ± 3.7 17.8 ± 0.4 0.90 ± 0.03 3.61 ± 0.05 6.70 ± 0.11 234 ± 21
DBA/2J 129.3 ± 1.74 19.2 ± 0.5 1.38 ± 0.06 3.96 ± 0.08 6.79 ± 0.22 240 ± 11
AKR/J 129.8 ± 3.33 27.9 ± 0.8 1.01 ± 0.04 3.12 ± 0.15 5.91 ± 0.11 250± 10
C3H/HeJ 102.6 ± 1.56 19.3 ± 0.4 0.85 ± 0.05 3.18 ± 0.10 7.45 ± 0.11 356 ± 25
CBA/J 115.8 ± 2.34 21.0 ± 0.4 0.77 ± 0.04 3.41 ± 0.06 7.06 ± 0.24 373 ± 55
Balb/cByJ 119.7 ± 3.3 19.6 ± 0.2 1.27 ± 0.07 3.60 ± 0.08 7.41 ± 0.09 336 ± 27
C57 C57Bl6/J 128.9 ± 2.0 18.6 ± 0.3 0.84 ± 0.04 3.39 ± 0.05 6.54 ± 0.15 270 ± 30
C57L/J 111 ± 4.6 20.8 ± 0.5 1.01 ± 0.05 3.73 ± 0.09 8.23 ± 0.25 332 ± 13
Swiss SJL/J 122.3 ± 1.93 18.7 ± 0.5 0.95 ± 0.06 3.32 ± 0.05 7.66 ± 0.12 287± 24
FVB/NJ 125.1 ± 3.2 25.5 ± XX 0.79 ± 0.07 3.06 ± 0.08 6.28 ± 0.14 410 ± 18
Wild-derived CAST/Ei 114.2 ± 3.12 11.8 ± 0.2 1.12 ± 0.05 3.86 ± 0.15 9.59 ± 0.56 392 ± 58
PWK/Ph 134.7 ± 3.5 13.3 ± 0.2 1.44 ± 0.04 4.49 ± 0.19 9.17 ± 0.24 313 ± 25
average 119.9 ± 2.6 18.9 ± 1 1.02 ± 0.06 3.52 ± 0.11 7.35 ± 0.3 315 ± 16

JAP-00073-2004.R1
24
Table 3: Morphologic characteristics of adrenals
Male FemaleGroup Strain
X-zone ZR X zone ZR Lipoid
zone
Castle’s A/J - + - ++ +++
129S1/SvlmJ - + ++ + -
DBA/2J - + ± + +
AKR/J - + + + ++
C3H/HeJ - ++ - + -
CBA/J - + + ++ +
Balb/cByJ + + + ++ ++
C57 C57Bl6/J - + + ++ -
C57L/J - + ++ -
Swiss SJL/J - + + + -
FVB/NJ - + ++ +
Wild-derived
CAST/Ei + + + + -
PWK/Ph + ++ + ++ -
Characteristics of the X zone, of zona reticularis (ZR) and the lipoid zone in male and female mice from all strains. - , Absent; +, poorly developed; ++ , well developed; +++ , very well developed.
JAP-00073-2004.R1
25
Table 4: Histologic characteristics of kidneys
Group Strain Glomerulus Mesangium JGA
Castle’s A/J normal normal normal
129S1/Svlmj ↑ ↑ ↑
DBA/2J normal normal normal
AKR/J ↑ ↑ normal
C3H/HeJ normal normal normal
CBA/J normal normal normal
Balb/cByJ normal normal ↑
C57 C57BI6/J normal normal normal
C57L/J ↑ normal ↑
Swiss SJL/J ↑ normal normal
FVB/NJ ↑ ↑ ↑
Wild-derived CAST/Ei normal normal normal
PWK/Ph normal normal normal
“normal” refers to features that are similar to that found in the majority of the strains. ↑ refers to
a feature that is either more prominent or enlarged as compared to that found in the majority of
the strains.

JAP-00073-2004.R1
26
FIGURE LEGENDS:
Fig. 1: Comparison of SBP (expressed in mm Hg) between male and female mice from 13
different strains. The filled circles correspond to strains whose SBP values were outside of the
boundaries defined by the 99% confidence interval of the linear correlation.
Fig. 2: Comparison of BW (expressed in g) between male and female mice from 13 different
strains. The filled circle corresponded to the AKR/J mice where BW values were outside of the
boundaries defined by the 99% confidence interval of the linear correlation.
Fig. 3: Left panel: relationship between SBP and LV/BW in male mice from 13 different strains.
Right panel: comparison of LV/BW between between male and female mice from 13 different
strains. The filled circles correspond to strains whose values were outside of the boundaries
defined by the 99% confidence interval of the linear correlation.
Fig.4: Relationship between the values of thymus / BW and that of bi-adr / BW male mice from
13 different strains.
Fig. 5. H&E stained sections of adrenals from 10-week-old C57BL/6J (A), Balb/cByJ (B) and AJ
(C) female mice. In all strains, the zona glomerulosa (ZG) is characterized by three to five layers
of cells of small cells, the zona fasciculata (ZF) by cells arranged as centripetal columns
separated by sinusoids, and the zona reticularis (ZR) by smaller cells with an irregular
disposition. In females Balb/cByJ (B) and in AJ (C), a well-developed lipoid zone (occupying

JAP-00073-2004.R1
27
uo to 50% of the adrenal volume) is present. Such a zone is absent in C57BL/6J (A). Scale bars:
100 µm.
Fig. 6. Representations of the inner adrenal cortex from 10-week-old Balb/cByJ (A), Cast/Ei (B)
and C57BL/6J male mice (H&E stained sections). Cells from the zonal fasciculata (ZF)
progressively changed into the inner zona fasciculata (IZF), then into the zona reticularis (ZR),
where cells are small, flattened and arranged in irregular groups and cords. In strains such as
Balb/cByJ (A) and Cast/Ei (B), a well-developed X zone (XZ) is still clearly present, whereas it
is much less demarcated or visible in C57BL/6J (C). Scale bars, 25 µm.
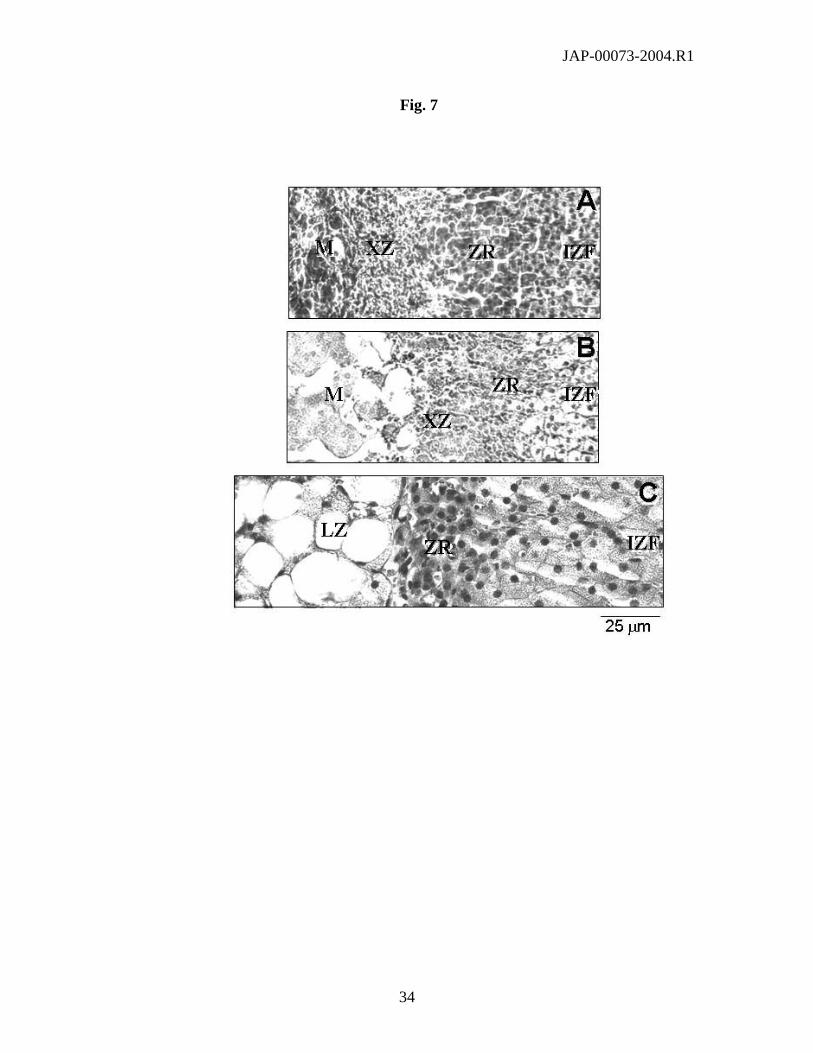
Fig. 7. Representations of the inner adrenal cortex from 10-week-old 129S1/SvlmJ (A), CBA/J
(B), and A/J (C) female mice (H&E stained sections). Fasciculata cells from CBA/J (B) and A/J
(C) adrenals contain much more lipid droplets than fasciculata cells from 129S1/SvlmJ (A)
adrenal glands. In all strains, fasciculata cells progressively changed into the zona reticularis
(ZR), where cells are small, flattened with a nucleus small and dark. The X zone (XZ), consisting
of small clumps of dispersed cells distributed in a more or less loose connective tissue, is clearly
observed in the 129S1/SvlmJ strain (A), but less evident in CBA/J (B) and is absent in the A/J
strain (C). In the A/J strain, the cortex is clearly separated from the medulla by a large lipoid
zone (LZ), where the nucleus is compressed against the cell membrane by a large lipid droplet.
Scale bars: 25 µm.

JAP-00073-2004.R1
28
Fig. 1
100 110 120 130 140
Males SBP (mm Hg)
100
110
120
130
140
Fem
ales
SB
P (
mm
Hg)
AKR/J
C57Bl6/J
CAST/Ei
129S1/SvlmJ

JAP-00073-2004.R1
29
Fig. 2
10 15 20 25 30
body weight (males)
10
15
20
25
30
body
wei
ght (
fem
ales
)
AKR/J

JAP-00073-2004.R1
30
Fig. 3
100 110 120 130 140
SBP (mm Hg)
2.00
2.50
3.00
3.50
4.00
4.50
5.00
LV/B
W
Males
2.50 3.00 3.50 4.00 4.50
Male LV/BW
2.50
3.00
3.50
4.00
4.50
5.00
Fem
ale
LV/B
W
Males vs. females
AKR/JA/J
FVB/NJ
JAP-00073-2004.R1
31
Fig. 4
0 1 2 3
Thymus / BW
0.10
0.20
0.30
bi-a
dren
al /
BW

JAP-00073-2004.R1
32
Fig. 5

JAP-00073-2004.R1
33
Fig. 6
JAP-00073-2004.R1
34
Fig. 7




















