
SecM-Stalled Ribosomes Adopt an Altered Geometry at the Peptidyl Transferase Center
Upload
independentCategory
view
3download
0
Glycobiology vol. 20 no. 5 pp. 617–628, 2010doi:10.1093/glycob/cwq011Advance Access publication on February 1, 2010
Characterization of a putative 3-deoxy-D-manno-2-octulosonic acid (Kdo) transferase gene fromArabidopsis thaliana
Martial Séveno2,4, Emilie Séveno-Carpentier2,5,Aline Voxeur2, Laurence Menu-Bouaouiche2,Christophe Rihouey2, Frédéric Delmas3,Christian Chevalier3, Azeddine Driouich2,and Patrice Lerouge1,2
2Laboratoire Glyco-MEV, UPRES-EA 4358, IFRMP 23, Université de Rouen,76821 Mont-Saint-Aignan, France; and 3Unité Mixte de Recherche 619Biologie du Fruit, INRA, Université de Bordeaux, Centre de Recherche INRA-Bordeaux-Aquitaine, 33883 Villenave d'Ornon cedex, France
Received on September 8, 2009; revised on January 13, 2010; accepted onJanuary 13, 2010
The structures of the pectic polysaccharide rhamnogalac-turonan II (RG-II) pectin constituent are remarkablyevolutionary conserved in all plant species. At least 12 dif-ferent glycosyl residues are present in RG-II. Among themis the seldom eight-carbon sugar 3-deoxy-D-manno-octu-losonic acid (Kdo) whose biosynthetic pathway has beenshown to be conserved between plants and Gram-negativebacteria. Kdo is formed in the cytosol by the condensationof phosphoenol pyruvate with D-arabinose-5-P and thenactivated by coupling to cytidine monophosphate (CMP)prior to its incorporation in the Golgi apparatus by aKdo transferase (KDTA) into the nascent polysaccharideRG-II. To gain new insight into RG-II biosynthesis andfunction, we isolated and characterized null mutants forthe unique putative KDTA (AtKDTA) encoded in the Ara-bidopsis genome. We provide evidence that, in contrast tomutants affecting the RG-II biosynthesis, the extinction ofthe AtKDTA gene expression does not result in any devel-opmental phenotype in the AtkdtA plants. Furthermore, thestructure of RG-II from the null mutants was not alteredand contained wild-type amount of Rha-α(1-5)Kdo sidechain. The cellular localization of AtKDTAwas investigat-ed by using laser scanning confocal imaging of the proteinfused to green fluorescent protein. In agreement with itscellular prediction, the fusion protein was demonstratedto be targeted to the mitochondria. These data, togetherwith data deduced from sequence analyses of higher plant
genomes, suggest that AtKDTA encodes a putative KDTAinvolved in the synthesis of a mitochondrial not yet identi-fied lipid A-like molecule rather than in the synthesis of thecell wall RG-II.
Keywords: Arabidopsis /Kdo/ lipid A/plant /rhamnogalacturonan II
Introduction
It was long believed that 3-deoxy-D-manno-octulosonic acid(Kdo) was synthesized only by Gram-negative bacteria as acomponent of the lipopolysaccharide (LPS) present in the outermembrane (Raetz 1990). In LPS, Kdo ensures the connectionbetween the lipid A, the hydrophobic glucosamine-based phos-pholipid anchor of LPS, and the outer core and O-antigen. Kdois crucial for bacteria since it was demonstrated that the mini-mal LPS required for the growth of Escherichia coli consists ofthe lipid A and two Kdo residues (Raetz and Whitfield 2002).Kdo was then discovered as a component of the primary cellwalls of higher plants and of cell wall polysaccharides of somegreen algae (York et al. 1985; Becker et al. 1995).The synthesis pathway of Kdo involves the intermediate for-
mation of D-arabinose, the enantiomer of L-arabinose found inthe arabinan and arabinogalactan chains of the plant cell wallpolysaccharides. The main steps of this synthesis are (i) theisomerization of D-ribulose-5-P into D-arabinose-5-P catalyzedby D-arabinose-5-P isomerase, (ii) the condensation of phos-phoenol pyruvate with D-arabinose-5-P to yield Kdo-8-P andPi catalyzed by Kdo-8-P synthase and (iii) the dephosphoryla-tion of Kdo-8-P by Kdo-8-P phosphatase. The resulting Kdo isthen activated by coupling to CMP (cytidine monophosphate;from cytidine triphosphate and free Kdo) in a reaction cata-lyzed by CMP-Kdo synthetase prior to its incorporation intothe LPS of Gram-negative bacteria mediated by Kdo transfer-ase (KDTA) (Rick 1987). In plants, the biosynthetic pathwayleading to Kdo is almost fully conserved (Figure 1). Taking ad-vantage of the complete Arabidopsis thaliana genomesequence, most of the genes have been identified (Wu et al.2004) and some were functionally characterized (Figure 1). Asingle gene (At3g54690) encodes a putative homolog of D-arab-inose-5-P isomerase (E. coli KpsF; Meredith and Woodward2006). In Arabidopsis, two genes code for a functional Kdo-8-P synthase (AtKDSA1, At1g79500 and AtKDSA2, At1g16340;Matsuura et al. 2003; Delmas et al. 2008), while a single geneoccurs in pea (Brabetz et al. 2000) and tomato (Delmas et al.2003). A single gene (AtKDSB, At1g53000) encodes the homo-
© The Author 2010. Published by Oxford University Press. All rights reserved. For permissions, please e-mail: [email protected] 617
1To whom correspondence should be addressed: Tel: +33-2-35-146394; Fax:+33-2-35-146615; e-mail: [email protected] address: CNRS UMR 5203, Plate-forme de ProtéomiqueFonctionnelle, Institut de Génomique Fonctionnelle, IFR3, 141 rue de laCardonille, 34094 Montpellier cedex 05, France5Present address: UMR 5004 BPMP, Campus INRA, SupAgro - IBIP,Bâtiment 7, 2 place Pierre Viala, 34060 Montpellier cedex 2, France
by guest on February 25, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from

log of CMP-Kdo synthetase that was previously characterized inmaize (Royo et al. 2000). The only exception is the Kdo-8-Pphosphatase for which homologous genes are not clearly pre-dicted in Arabidopsis. Interestingly, all these sequences sharestrong similarities with their bacterial homologues, thus sug-gesting that they may have appeared in plants followingsymbiosis with bacteria. In addition to genes encoding enzymesinvolved in the Kdo synthesis or activation, Wu et al. (2004)mentioned the occurrence of a unique putative KDTA in Arabi-dopsis (AtKDTA encoded by At5g03770). However, the inplanta function of this putative AtKDTA has not been elucidatedso far.
Kdo is one of the remarkable monomers of rhamnogalactur-onan II (RG-II). This pectic polysaccharide of cell walls iscomposed of an α-1,4-linked homogalacturonan backbone thatis substituted with four structurally different oligosaccharideside chains, namely A to D side chains (O'Neill et al. 2004).At least 12 different glycosyl residues are present in RG-II, in-cluding, in addition to Kdo, the rare aceric acid, apiose and 3-deoxy-D-lyxo-heptulosonic acid (Dha) (O'Neill et al. 1996; Pérezet al. 2000). As a result of its complex structure, the biosynthe-sis of RG-II is likely to require at least 22 glycosyltransferaseswhich is estimated to represent half of the glycosyltransferasesneeded for the synthesis of pectin (O'Neill et al. 2004). Despiteits highly complex structure, RG-II is evolutionarily conservedin the plant kingdom as it is present in the primary cell wall ofall higher plants predominantly in the form of a dimer that iscross-linked by a borate di-ester between two apiosyl residues(O'Neill et al. 1996; Kobayashi et al. 1996). This suggests thatproteins involved in its synthesis appeared early in land plantevolution and that RG-II has fundamental functions in the pri-mary wall organization. RG-II is believed to be involved in theregulation of cell wall properties and plant growth due to theboron-mediated cross-linking of RG-II that generates a cova-lently cross-linked pectic network (Fleisher et al. 1999; Ishii
et al. 1999; Ishii et al. 2001). So far, all mutations that affectthe RG-II structure deeply alter the plant development (O'Neillet al. 2001; Ahn et al. 2006). Furthermore, the inactivation ofthe Kdo synthesis resulted in nonviable null mutants. Pollentubes from mutants of the cytosolic D-arabinose-5-P isomeraseor Kdo-8-P synthase were demonstrated to be unable to elon-gate properly and perform fertilization (Johnson et al. 2004;Delmas et al. 2008). Thus, although the precise function ofRG-II remains to be established, RG-II is believed to play akey function in primary cell wall formation and its in murofunctionality requires the presence of Kdo.
In the present study, we report on the characterization of nullmutants for a putative KDTA in A. thaliana to elucidate its po-tential involvement in the transfer of Kdo into the cell wallRG-II. Our data suggest that AtKDTA encodes a putativeKDTA involved in the synthesis of a mitochondrial not yetidentified lipid A-like molecule rather than in the biosynthesisof the cell wall pectic polysaccharide RG-II.
ResultsArabidopsis contains a single-copy gene for a putative KDTAChromosome V of A. thaliana harbors a genomic sequence(At5g03770) that could encode a KDTA protein referred toas AtKDTA, as revealed by basic local alignment search tool(BLAST) searches using the E. coli KDTA (Clementz andRaetz 1991) as a query sequence. Full-length sequences forplant KDTA proteins were also identified in Oryza sativa(EEE55697.1),Medicago truncatula (ABE80128.2), Vitis vinif-era (CAO15996.1),Populus trichocarpa (EEF04661.1),Ricinuscommunis (EEF33745.1) and Zea mays (ACN31913.1) as wellas in the moss Physcomitrella patens (PHYPADRAFT109233). The predicted translation products of these plantKDTA genes display significant amino acid identity percentagewith various KDTA sequences of prokaryotic organisms(CAZy GT30, http://www.cazy.org; Cantarel et al. 2009) whichall belong to the Gram-negative bacterial family. Despite thesesequence similarities with the bacterial enzymes, the plant pro-teins represent a distinct lineage, which could be clearlyseparated from the Gram-negative bacterial proteins as seenin the phylogenetic tree (Figure 2). The amino acid sequencealignment of plant putative KDTA (Figure 3) indicates thatAtKDTA shares from 48 to 67% of identity with other plantKDTA sequences. Sequence identity with the Gram-negativebacterial KDTA varies from 15 to 35%. The best rates were ob-served for rhizobial specieswith31–35%of identityand50–55%of similarity between plant and bacterial KDTA sequences sup-porting a direct homology between plant and rhizobialsequences (Figure 2). The A. thaliana putative KDTA is com-posed of 447 amino acids (Mr = 49871, pI = 9.96) and displaysthe two identified Pfam domains, Glyco_transf_N (PF04413)from residue 36 to 221 and Glyco_transf_1 (PF00534) from res-idue 242 to 415, of KDTA protein (http://pfam.sanger.ac.uk/;Finn et al. 2008). The prediction of the 3D structure of AtKDTA(Q8VZA5entry ofModBase, http://modbase.compbio.ucsf.edu;Pieper et al. 2009) indicated that the protein is composed of twoRossman fold protein domains as reported for GT-B fold of gly-cosyltransferases (Lairson et al. 2008). Sequence alignmentsrevealed the presence of highly conserved protein motifs withinboth plant and bacterial KDTA (Figure 3). Although no data are
618
Fig. 1. Biosynthesis and activation of Kdo in plant cells. Arabidopsis locusnumbers and corresponding gene names (when functionally characterized) areindicated in the diagram.
M Séveno et al.
by guest on February 25, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from

available on amino acids involved in substrate recognition andCMP-Kdo recognition in bacterial KDTA, these motifs may becrucial for transferase activities in both prokaryotic and eukary-otic proteins. Among these, the two conserved sequences W58–G66 andG345–E350 couldbe involved in the bindingofCMP-Kdoonto the KDTAs since these motifs were shown to be implicatedin the CMP-Kdo binding site in CMP-Kdo synthases (Jelakovicand Schulz 2002). Furthermore, the asparagine residue (R279 inthe Arabidopis sequence) of the conserved PR sequence wasdemonstrated to be required for the transfer of Kdo onto lipid Ain bacteria (Figure 3) (Ekpo and Nano 2006).
Gene expression analysis of the Kdo biosynthetic genes inArabidopsis plantsWe analyzed the expression of the genes involved in the Kdobiosynthetic pathway in Arabidopsis: AtKDSA1 and AtKDSA2encoding the two isoforms of Kdo-8-P synthase (Matsuura etal. 2003), AtKBSB encoding the CMP-Kdo synthetase andAtKDTA encoding the putative KDTA. The expression analysiswas performed using total RNAs extracted from mature flow-ers, stems, immature and mature leaves from wild-type Col0plants. Results of transcript copy number estimations with realtime quantitative reverse transcriptase-polymerase chain reac-
619
Fig. 2. Phylogenetic tree of KDTA based on the maximum likelihood method. The scale bar (0.5) represents the number of amino acid residue substitutions per site.Plant and rhizobial KDTA are in bold and underlined, respectively. Accession numbers for the different protein sequences are indicated in the Materials and methodssection.
Characterization of Arabidopsis Kdo transferase
by guest on February 25, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from

tion (RT-PCR) are presented in Figure 4. All four genes werepreferentially expressed in flowers and in immature leaves, i.e.in plant organs displaying cell division activities. This expres-
sion profile is in agreement with our previous work showingthat the AtKDSA2 promoter is strongly activated in shoot apicalmeristem, first rosette leaves and flowers (Delmas et al. 2008),
620
Fig. 3. Protein sequence multiple alignments of KDTA from plants and from Gram-negative bacteria. The gray line above the alignments indicates the twoconserved sequences likely involved in the binding of CMP-Kdo. The arrow pointed out to the conserved asparagine residue required for the transfer of Kdo ontolipid A in bacteria. Identical amino acids are shaded in black, and similar amino acids are in bold. Abbreviations and accession numbers are indicated in Materialsand methods section.
M Séveno et al.
by guest on February 25, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from

and that the tomato kdsA gene expression and relevant Kdo-8-Psynthase activity are preferentially associated with dividingcells in a cell cycle-dependent manner (Delmas et al. 2003).To a lesser extent, the expression of AtKDSA1, AtKDSA2,AtKBSB and AtKDTA was also detected in stems and matureleaves.
Isolation of AtkdtA null mutantsWe obtained two AtKDTAT-DNA insertion lines from the Ara-bidopsis Biological Resource Center (SALK_035981 and
GABI_140B12) designated as AtkdtA1 and AtkdtA2, respec-tively. The T-DNA insertion site of AtkdtA1 is illustrated inFigure 5A; it disrupts the AtKDTA gene within intron 7. Thiswas confirmed by PCR analyses using genomic DNA as com-pared to control wild-type Col0 plants (not shown). RT-PCRanalyses of T4 homozygous AtkdtA1 plants confirmed thatAtkdtA1 is a null allele (Figure 5B). Identical results were ob-tained with AtkdtA2 mutant which is disrupted in exon 10(Figure 5B). The general development and growth phenotypeof AtkdtA null mutants appear to be similar to those of the wild-type Col0 plants (not shown). Thus under standard growthconditions and in contrast to mutants altered in the cytosolicbiosynthesis of Kdo (Johnson et al. 2004; Delmas et al.2008), the knock-out mutation of AtKDTA does not induceany apparent modifications of the plant phenotype.
Biochemical characterization of RG-II isolated from theputative AtkdtA null mutantsStudying the biological function of the Arabidopsis putativeKDTA is hardly achievable through an enzymatic assay sinceCMP-Kdo required for the bioassay is an unstable compound(Belunis et al. 1995). As a consequence, to determine whetherthis putative transferase is involved in the incorporation of Kdointo RG-II, we investigated the RG-II structure from the homo-zygous AtkdtA1 null mutant. Pectins were extracted from leafmaterial by treatment of an alcohol-insoluble fraction with en-dopolygalacturonase (EPG). RG-II was separated from RG-Iand oligogalacturonides by size exclusion chromatography(SEC), and its sugar composition was determined by gas liquidchromatography. As shown in Table I, the sugar compositionof the AtkdtA1 RG-II fraction was similar to that of RG-II fromwild-type Arabidopsis plants, including the Kdo content which
Fig. 4. Expression analysis of Kdo biosynthetic genes in Arabidopsis plants.Total RNAs were prepared from flowers, stems, immature and mature leaves ofwild-type Col0 plants. The copy number of Kdo biosynthetic pathway genetranscripts estimated from quantitative RT-PCR experiments in real time isrepresented by bar graphs. Copy number estimates were conducted usingcDNA synthesized from 8ng of total RNAs. Error bars represent experimentalvariation observed in four independent experiments using different wild-typeplants.
Fig. 5. Characterization of the AtkdtA null mutants. (A) Schematic representation of the T-DNA insertions in the AtkdtA gene. The coding region of the geneconsists of 11 exons, represented as black boxes. White boxes represent the untranslated 5′ and 3′ regions of the corresponding mRNA. The location of primers usedin RT-PCR experiments to characterize the AtkdtA null mutant is given in the diagram. (B) Molecular characterization of the AtkdtA null mutants by RT-PCRanalysis of Arabidopsis AtKDTA mRNA levels in AtkdtA1 and AtkdtA2 mutants.
621
Characterization of Arabidopsis Kdo transferase
by guest on February 25, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from
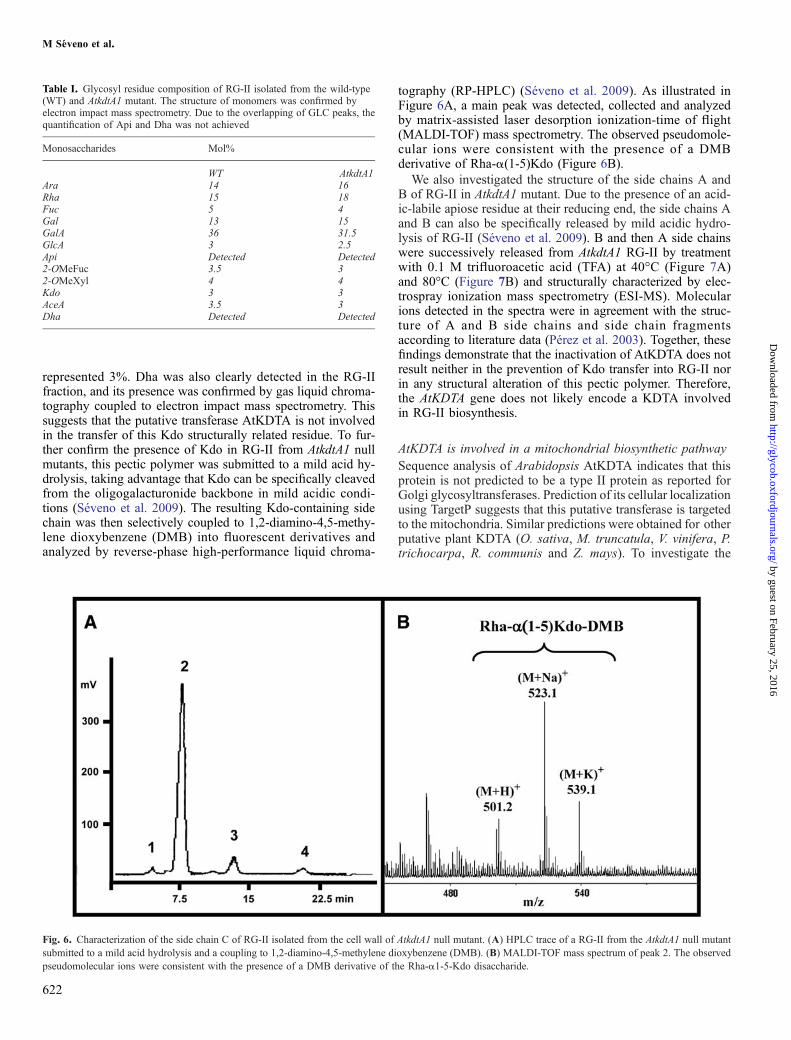
represented 3%. Dha was also clearly detected in the RG-IIfraction, and its presence was confirmed by gas liquid chroma-tography coupled to electron impact mass spectrometry. Thissuggests that the putative transferase AtKDTA is not involvedin the transfer of this Kdo structurally related residue. To fur-ther confirm the presence of Kdo in RG-II from AtkdtA1 nullmutants, this pectic polymer was submitted to a mild acid hy-drolysis, taking advantage that Kdo can be specifically cleavedfrom the oligogalacturonide backbone in mild acidic condi-tions (Séveno et al. 2009). The resulting Kdo-containing sidechain was then selectively coupled to 1,2-diamino-4,5-methy-lene dioxybenzene (DMB) into fluorescent derivatives andanalyzed by reverse-phase high-performance liquid chroma-
tography (RP-HPLC) (Séveno et al. 2009). As illustrated inFigure 6A, a main peak was detected, collected and analyzedby matrix-assisted laser desorption ionization-time of flight(MALDI-TOF) mass spectrometry. The observed pseudomole-cular ions were consistent with the presence of a DMBderivative of Rha-α(1-5)Kdo (Figure 6B).
We also investigated the structure of the side chains A andB of RG-II in AtkdtA1 mutant. Due to the presence of an acid-ic-labile apiose residue at their reducing end, the side chains Aand B can also be specifically released by mild acidic hydro-lysis of RG-II (Séveno et al. 2009). B and then A side chainswere successively released from AtkdtA1 RG-II by treatmentwith 0.1 M trifluoroacetic acid (TFA) at 40°C (Figure 7A)and 80°C (Figure 7B) and structurally characterized by elec-trospray ionization mass spectrometry (ESI-MS). Molecularions detected in the spectra were in agreement with the struc-ture of A and B side chains and side chain fragmentsaccording to literature data (Pérez et al. 2003). Together, thesefindings demonstrate that the inactivation of AtKDTA does notresult neither in the prevention of Kdo transfer into RG-II norin any structural alteration of this pectic polymer. Therefore,the AtKDTA gene does not likely encode a KDTA involvedin RG-II biosynthesis.
AtKDTA is involved in a mitochondrial biosynthetic pathwaySequence analysis of Arabidopsis AtKDTA indicates that thisprotein is not predicted to be a type II protein as reported forGolgi glycosyltransferases. Prediction of its cellular localizationusing TargetP suggests that this putative transferase is targetedto the mitochondria. Similar predictions were obtained for otherputative plant KDTA (O. sativa, M. truncatula, V. vinifera, P.trichocarpa, R. communis and Z. mays). To investigate the
Table I. Glycosyl residue composition of RG-II isolated from the wild-type(WT) and AtkdtA1 mutant. The structure of monomers was confirmed byelectron impact mass spectrometry. Due to the overlapping of GLC peaks, thequantification of Api and Dha was not achieved
Monosaccharides Mol%
WT AtkdtA1Ara 14 16Rha 15 18Fuc 5 4Gal 13 15GalA 36 31.5GlcA 3 2.5Api Detected Detected2-OMeFuc 3.5 32-OMeXyl 4 4Kdo 3 3AceA 3.5 3Dha Detected Detected
Fig. 6. Characterization of the side chain C of RG-II isolated from the cell wall of AtkdtA1 null mutant. (A) HPLC trace of a RG-II from the AtkdtA1 null mutantsubmitted to a mild acid hydrolysis and a coupling to 1,2-diamino-4,5-methylene dioxybenzene (DMB). (B) MALDI-TOF mass spectrum of peak 2. The observedpseudomolecular ions were consistent with the presence of a DMB derivative of the Rha-α1-5-Kdo disaccharide.
622
M Séveno et al.
by guest on February 25, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from

cellular localization of AtKDTA, this putative transferase wastransiently expressed in tobacco leaves as a fusion protein togreen fluorescent protein (GFP) and visualized using confocalmicroscopy. As shown in Figure 8A (green channel), 48 h after
infiltration, the AtKDTA-GFP fusion protein was mainly locat-ed in mobile 1-μm dots. To identify the nature of these dots, wecoexpressed AtKDTA-GFP with a Golgi marker, the ERD2-cy-an fluorescent protein (CFP) fusion protein (Saint-Jore et al.2002) (Figure 8A). While the ERD2-CFP fusion protein accu-mulated in both endoplasmic reticulum and Golgi apparatus, theAtKDTA-GFP fusion protein appeared in dots that are clearlydistinct from Golgi bodies. To determine whether the AtKD-TA-GFP-labeled structures are related to mitochondria, weused the MitoTracker® orange to specifically stain these orga-nelles. MitoTracker® orange is a mitochondrion-selective dyethat enters the cells and is sequestered in the mitochondria whereit reacts with thiols on proteins to form a fluorescent conjugate.Figure 8B shows a cell expressing the AtKDTA-GFP constructwhose mitochondria are labeled with MitoTracker®. Mitochon-dria appear as dot structures of 1-μm diameter that are highlymobile. AtKDTA-GFP accumulated in the same dots as Mito-Tracker® and showed a ring-like labeling surrounding themitochondria (arrow in Figure 8B). Additional localization ex-periments were carried out on tobacco mesophyll cells thatcontain chloroplasts. No colocalization of AtKDTA-GFP withchloroplasts was observed (not shown). Together these datademonstrate that, under our labeling conditions, the fusion pro-tein AtKDTA-GFP is localized in the mitochondria.
Discussion
Altering RG-II composition and structure is of an indisputableinterest to understand its function in cell wall and its relation-ship with growth. Hence the isolation of plant mutants isneeded to offer such a means of investigation. In this study,we aimed at performing a functional analysis of a candidategene putatively encoding a KDTA referred to as AtKDTA.AtKDTA was identified in the Arabidopsis genome, based onamino acid sequence alignments with Gram-negative bacterialKDTAs (Figures 2 and 3). The best sequence homologies wereobserved for Rhizobial species with 31–35% of identity and50–55% of similarity between plant and bacterial KDTA se-quences. Sequence alignments revealed the presence ofhighly conserved protein motifs within both plant and bacterialKDTA which could be implicated in catalytic activity.Two biochemical lines of evidence strongly suggest that
AtKDTA is not involved in RG-II biosynthesis in A. thaliana.First, Kdo was detected in the glycosyl residue composition ofAtKDTA1 null mutant in a ratio that is consistent with a presenceof one Kdo residue for one RG-II monomer. Second, the Rha-α1-5-Kdo side chain of RG-II was structurally characterized inthis mutant after release by mild acidic hydrolysis and couplingto DMB (Figure 7). Furthermore, this putative transferase is notpredicted to be a membrane protein as required for its localiza-tion in the Golgi apparatus, and the AtKDTA-GFP fusionprotein does not accumulate in Golgi structures where RG-IIis synthesized. As a consequence, we postulate that AtKDTA isnot responsible for the incorporation of Kdo in RG-II. Other pro-teins such as sialic acid transferase-like enzymes (At3g48820and At1g08660) have to be considered. Sialic acids are not syn-thesized in plants and therefore these putative transferases arenot involved in the transfer of this residue onto glycopolymers(Séveno et al. 2004; Daskalova et al. 2009). Arabidopsis sialicacid transferase-like proteins were demonstrated to be located
623
Fig. 7. Characterization of the side chains A and B of RG-II isolated from theAtkdtA1 null mutant. Structure of A side chain (A) and B side chain (B) ofArabidopsis RG-II. ESI-MS of oligosaccharide fragments released by themild acid hydrolysis in TFA 0.1M at 40°C (C) and 80°C (D) of RG-II isolatedfrom the cell wall of AtkdtA1 mutant. *: potassium adduct. •: A side chainmethyl esters. Api, D-apiose; AceA, L-aceric acid; 2-OMeFuc, 2-O-methylL-fucose; 2-OMeXyl, 2-O-methyl D-xylose. B - Rha and A - 2OMeXyl referto B side chain lacking L-rhamnose residue and A side chain lacking 2-O-methyl D-xylose, respectively. *: potassium adduct.
Characterization of Arabidopsis Kdo transferase
by guest on February 25, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from

in the Golgi apparatus by a proteomic approach (Dunkley et al.2006) and by the analysis of the localization of thecorresponding GFP fusion proteins (Daskalova et al. 2009)but their biological functions in this compartment have notbeen elucidated so far. Since Kdo and sialic acid transferasesshare common features such as the use of CMP-activated su-gars as cosubstrates (Münster et al. 1998), these Golgitransferases may be considered as putative candidates for thetransfer of Kdo and/or Dha into RG-II.
A relationship between AtKDTA and RG-II biosynthesis isalso highly questioned due to the absence of phenotypical alter-ation in the AtkdtA mutant lines when compared to wild-typeplants. Indeed all mutations so far identified that target theRG-II composition and thereby structure deeply alter plant de-velopment (O'Neill et al. 2001; Ahn et al. 2006). Furthermore,the inactivation of the D-arabinose-5-P isomerase or Kdo-8-Psynthase that is involved in the Kdo synthesis in the cytosolresulted in nonviable null mutants (Johnson et al. 2004; Delmaset al. 2008). It would be expected that preventing Kdo incorpo-ration into the RG-II would also result in the inability of pollenfrom the null mutant to perform fertilization. In addition, theKdo structurally related monosaccharide Dha was also clearlyidentified in the RG-II fraction, thus suggesting that AtKDTAis not involved in the transfer of this monomer in the Ara-β(1-5)Dha side chain of RG-II.
The cellular localization of AtKDTA was investigatedthrough confocal imaging of the protein fused to GFP. The fu-sion protein was observed in 1-μm size mobile dots.Coexpression with Golgi marker clearly showed that these dotsare not Golgi stacks as expected for glycosyltransferases in-volved in the RG-II biosynthesis. In contrast, AtKDTA-GFPaccumulated in the same dots as MitoTracker®, a mitochondrialmarker, and showed a ring-like labeling surrounding the mito-chondria. As a consequence, in agreement with its predictedcellular localization, we conclude that AtKDTA protein is tar-geted to the mitochondria.
Recently, BLASTsearch for candidate genes in the Arabidop-sis genome revealed the presence of a set of genes potentiallyencoding homologues of LpxA, LpxB, LpxC, LpxD and LpxKinvolved in the biosynthesis of lipid A, the hydrophobic glucos-amine-based anchor of LPS in bacteria (Wu et al. 2004). Wesearched in other sequenced plant genomes and found thatLpx genes are conserved in the plant kingdom. Furthermore,most of these genes encode proteins that are predicted to be tar-geted to mitochondria. Although the presence of lipid A-likemolecules is still highly speculative, this observation suggeststhat the putative Arabidopsis AtKDTA may be involved in thebiosynthesis of a not yet identified Kdo-lipid A mitochondrialmembrane component in plants and not in the transfer of Kdointo RG-II. The presence of lipid A-like molecules in plants is
624
Fig. 8. Subcellular localization of AtKDTA. (A) Tobacco leaf epidermis expressing both ERD2-CFP (red) and AtKDTA-GFP (green) constructs at 48 h afterinfiltration. The two images were merged to show that AtKDTA-GFP fusion protein accumulated in mobile dots was distinct from the Golgi stacks (arrows). Bars =5 μm. (B) Tobacco leaf epidermis expressing the AtKDTA-GFP construct (green) (48 h) was infiltrated with the MitoTracker® dye (red). The fluorescence wasobserved 4 h after MitoTracker® orange treatment. Note that AtKDTA-GFP dots and MitoTracker®-stained mitochondria colocalize (arrows). Bars = 5 μm.
M Séveno et al.
by guest on February 25, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from

debatable. Recent reports on this hypothetic bacteria-like glyco-lipid suggested its presence onto the chloroplast membrane(Raetz and Whitfield 2002; Armstrong et al. 2006). Lipid Amay have appeared in plants following symbiosis with bacte-ria which would explain the high homology of all proteins ofits biosynthetic pathway with bacterial sequences. This bio-chemical process led to the accumulation in the plant cytosolof Kdo that might have then been channeled to the synthesisof the plant cell wall RG-II during the early land plant evolu-tion. Efforts are currently made in our laboratory to providebiochemical evidence for the presence of a lipid A-like on plantmitochondria and to unravel its putative function in higherplants. Since numerous genes encoding homologues of theToll-like receptors are also found in the Arabidopsis genome,this plant lipid A may be involved in some yet unknown signal-ing mechanisms.
Materials and methodsPlant materialsWild-type and AtkdtA mutant plants of A. thaliana are in theColumbia (Col0) ecotype background. For germination, seedswere surface sterilized for 15 min in 12.5% (v/v) sodium hy-pochlorite and 0.02% (v/v) Triton X-100, rinsed at least fivetimes and plated in Petri plates containing Murashige andSkoog's growth medium (Murashige and Skoog 1962). Aftercold treatment at 4°C for 2 days in the dark, the plates wereincubated in a growth chamber at 22°C in a cycle of 16-h light/8-h darkness. After 10 days of growth the plantlets weretransferred in soil in a growth chamber in the same conditions.
Amino acid sequence analysisThe phylogenetic analysis and drawing of the KDTA multiplealignments were performed on the phylogeny.fr server (http://www.phylogeny.fr; Dereeper et al. 2008) with the PhyML pro-gram (Guindon and Gascuel 2003; Anisimova and Gascuel2006) after blocks selection using Gblocks (Castresana 2000).The phylogenetic tree was drawn with TreeDyn (Chevenet etal. 2006). The multiple KDTA sequences alignment used forphylogenetic analysis was generated by the multAlin program(http://bioinfo.genotoul.fr/multalin; Corpet 1988). Accessionnumbers and abbreviations for the different protein sequencesare as follows: Aquifex aeolicus, AaKDTA, aq_326; Acidobac-teria bacterium, AbKDTA, Acid345_4718; Anaeromyxobacterdehalogenans, AdKDTA, Adeh_2615; Arabidopsis thaliana,AtKDTA, Q8VZA5; Agrobacterium tumefaciens, AtuKDTA,Atu0695; Brucella melitensis, BmKDTA, BMEII1029; Bacter-oides thetaiotaomicron, BtKDTA, BT_2747; Caulobactercrescentus, CcKDTA, CC_0302; Chlorobaculum tepidum,CteKDTA, CT0593; Chlamydia trachomatis, CtrKDTA,CT208; Escherichia coli, EcKDTA, b3633; Fusobacterium nu-cleatum, FnKDTA, FN1606; Geobacter sulfurreducens,GsKDTA, GSU2259; Haemophilus influenzae, HiKDTA,HI0652; Helicobacter pylori, HpKDTA, HP0957; Leptospirainterrogans, LiKDTA, LA1477; Legionella pneumophila Paris,LpKDTA, lpp2288; Mesorhizobium loti, MlKDTA, mlr8269;Medicago truncatula, MtKDTA, ABE80128.2; Neisseria me-ningitidis, NmKDTA, NMB0014; Oryza sativa, OsKDTA,EEE55697.1; Opitutus terrae, OtKDTA, Oter_2533; Pseudo-monas aeruginosa, PaKDTA, PA4988; Physcomitrella patens,
PpKDTA, PHYPADRAFT_109233; Populus trichocarpa,PtKDTA, EEF04661.1; Rhodopirellula baltica, RbKDTA,RB11688; Ricinus communis, RcKDTA, EEF33745.1; Rhizobi-um leguminosarum, RlKDTA, RL0902; Rickettsia prowazekii,RprKDTA, RP089; Rhodopseudomonas palustris, RpaKDTA,RPA1158; Ralstonia solanacearum, RsoKDTA, RSc0693; Rho-dobacter sphaeroides, RspKDTA, RSP_2394; Sinorhizobiummeliloti, SmKDTA, SMc00894; Vibrio cholerae, VcKDTA,VC0233; Vitis vinifera, VvKDTA, CAO15996.1; Xanthomonascampestris, XcKDTA, XCC3337; Yersinia pestis, YpKDTA,YPO0055; Zea mays, ZmKDTA, ACN31913.1. An extract ofthe multiple alignments presented in Figure 3 was rendered bythe ESPript/ENDscript program (http://espript.ibcp.fr; Gouet etal. 2003).Putative cellular localization of the proteins was analyzed us-
ing TargetP program (http://www.cbs.dtu.dk/services/TargetP;Emanuelsson et al. 2007).
Isolation of AtkdtA null mutantsTo isolate the homozygous AtkdtA1 null mutant, Arabidopsisgenomic DNA was PCR amplified with the primers Kdo-Transf5′ (5′-TGTTGTCTGCTTTTTGTTCCA-3′) andKdoTransf3′ (5′-TTCGTCTTCCTTGGTGTAAGC-3′). PCRreactions were performed using the LBa1 primer (5′-TGGTTCACGTAGTGGGCCATCG-3′), located within theLB sequence of the T-DNA, and KdoTransf5′ or KdoTransf3′primer in order to amplify the T-DNA junction. The use of pri-mers KdoTransf5′ and KdoTransf3′ generates a 606 bpfragment of the AtKDTA genomic sequence. PCR reactionswere performed with genomic DNA extracted as describedby Edwards et al. (1991). Fifty nanograms of genomic DNAwas used for each PCR with primer combinations Kdo-Transf5′–KdoTransf3′, KdoTransf5′–LBa1 or LBa1–KdoTransf3′. Same strategy was used for the isolation of theAtkdtA2 null mutant using the T-DNA primer GABI-T-DNA(5′-CCATTTGGACGTGAATGTAGACAC-3′), AtKDOTF2(5′-GCCTCGTAGACTATCGTGTGG-3′) and AtKDOTR(5′-TTTGACTTGTCGAGCCATTTC-3′).
RNA extraction, cDNA synthesis and RT-PCR analysisWild-type or AtkdtA seedlings grown under light conditions for10–12 days were transferred to soil for growth and used forRNA analysis. Total RNAwere isolated using the TRIzol® Re-agent (Invitrogen) and were treated with DNase RQI(Promega, Charbonnières-les-bains, France) according to themanufacturer's protocol. Two micrograms of total RNA fromplantlets was reverse transcribed into cDNA using oligo(dT)16as a primer, SuperScript™ II RNase H− reverse transcriptase(Invitrogen) in a total volume of 20 μL. The cDNAwas then di-luted 10 times, and 1 μL of the diluted cDNA was used as atemplate for PCR analysis. PCR reactions were performed usingAtKDOTF1 (5′-TCGCTTATCTGGTTTCACGC-3′) and Atkd-tAi3 (5′-ATCTGTTGGCCATGATGTGG-3′) as primers fordetecting KDTA transcripts in the AtkdtA1 mutant, and usingAtkdtAi5 (5′-GCATCTTCATTACATAGAGG-3′) and AtK-DOTR as primers in the AtKdtA2 mutant. After an initialdenaturation step of 5 min at 95°C, the reaction program wasas follows: 30 s at 95°C as a denaturation step, 30 s at 60°Cfor primer annealing, 1 min 20 s at 72°C as an elongation step,for 30 cycles, and a final step of 5 min at 72°C.
625
Characterization of Arabidopsis Kdo transferase
by guest on February 25, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from

Real time quantitative RT-PCRReal time quantitative PCR was performed as to analyze thegene expression for AtKDSA1, AtKDSA2, AtKDSB and AtKD-TA using the following primers, respectively: AtkdsA1i-5′(5′-GAGGTTGGTGACCCAAACGG-3′) and AtkdsA1i-3′(5′-CATGGTGCTTCTTTAAAACGTTCA-3′); AtkdsA2i-5′(5′-CTGCATTTTGCAGGTCCGTAC-3′) and AtkdsA2i-3′(5′-TATCATATCAGGCAACATCAGCTACTTTGA-3′);AtCMPKdoi-5′ (5′-GTTGACACACCCGATGATGTC-3′) andAtCMPKdoi-3′ (5′-CACACACAATGCCACAACTC-3′);AtkdtAi-5′ (5′-GCATCTTCATTACATAGAGG-3′) and Atkd-tAi-3′ (5′-ACTTGTTGGCCATGATGTGG-3′). Reactionmixtures (25 μL) consisted of 12.5 μL of Platinum® quanti-tative PCR Supermix-UDG (Invitrogen) with 1 μL of SYBRGreen I dye (Molecular Probes, Invitrogen, Cergy Pontoise,France) solution (1/3000 dilution in water of the manufac-turer's 10,000× stock solution), 0.2 μM of each primersand cDNA corresponding to 8 ng of total RNA. Cycling con-ditions consisted of an initial uracyl DNA glycosylasetreatment for 2 min and denaturation step of 95°C for5 min as a “hot start”, 40 cycles of 95°C for 30 s and 60°C for 30 s. The PCR reactions and quantifications were car-ried out in the real time PCR detection system iCycler iQ (Bio-Rad, Marne-la-Coquette, France). During PCR amplification,fluorescence emission was monitored. The increase in fluo-rescence emission of the dye used is proportional to theamount of PCR product accumulated that is in turn propor-tional to the starting amount of DNA template. A standardcurve was constructed using templates of known copy num-ber for the target sequence. The copy number of the sampleswas estimated by plotting the threshold cycle (the cycle atwhich fluorescence is considered to be significant above thebackground level and within linear range) against the log ofthe starting copy number (iCycler iQ real time detection sys-tem from Bio-Rad manual). To construct the standard curve,serial dilutions of cloned AtKDSA1, AtKDSA2, AtKDSB andAtKDTA fragments inserted into the pGEM-T Easy vector(Promega) were used. The number of copies in each dilutionwas calculated with the following formula: (number of mo-les) × (6.02 × 1023) = number of copies. All standardsamples were assayed in duplicate wells, and experimentalsamples were assayed in quadruplate wells.
Isolation of Arabidopsis RG-IIOne gram of frozen leaves was suspended in aqueous 80%(v/v) ethanol, heated at 70°C for 15 min and centrifuged at2500 rpm for 5 min. The insoluble residue was treated for4 h at 4°C with 0.1 N NaOH to saponify the methyl and acetylesters. The suspensions were adjusted to pH 5 with 10% (v/v)acetic acid and then treated for 16 h at 30°C with an EPG fromAspergillus niger (30 units, Megazyme, Wicklow, Ireland) andthen with α-amylase (500 units) for 16 h at 25°C. The suspen-sions were centrifuged, and the insoluble residues were washedwith water. The EPG-soluble fractions were dialyzed (1 kDacutoff dialysis tubing) against deionized water and freeze-dried. RG-I and RG-II polysaccharides were purified fromthe EPG-solubilized material by elution from SEC on a Sepha-dex G-75 (2.5 × 90 cm) (Amersham Pharmacia Biotech Inc.,Uppsala, Sweden; 1.6 × 38 cm) column in 50 mM ammo-nium formate as buffer.
Mild acid hydrolysis of RG-IIOne milligram of purified RG-II was hydrolyzed in 200 μL of0.1 M TFA for 16 h at 40°C. For the visualization of the Achain, the sample was then submitted to an additional hydro-lysis at 80°C for 1 h. The solutions were then freeze-driedbefore analysis by ESI-MS-MS.
Gas liquid chromatography analysisThe samples were submitted to a 16-h methanolysis at 80°Cwith 500 μL of dry 2 M methanolic-HCl. After evaporationof the methanol, the methyl glycosides were then converted in-to their trimethylsilyl derivatives and separated by gas liquidchromatography. The gas chromatograph was equipped witha flame ionization detector, a wall-coated open tubular fusedsilica capillary column (length 25 m, i.d. 0.25 mm) with CP-Sil 5 CP as stationary phase and helium as gas vector. The ov-en temperature program was 2 min at 120°C, 10°C/min to 160°C, 1.5°C/min to 220°C and then 20°C/min to 280°C. Thequantification of sugar was done by integration of peaks anddetermination of the corresponding molar values using re-sponse factors established with standard monosaccharides.Gas liquid chromatography coupled to electron impact massspectrometry was performed using an Agilent (Wilmington,DE) GC 6890 chromatograph coupled to an Agilent 5973 massspectrometer. Data collection was performed on HP Chemsta-tion A.03.03. The separation of monosaccharides wasperformed on an Agilent HP5 column (30 m × 0.25 mm ×0.25 mm). Oven temperature was programmed from 60°Cfor 1 min to 140°C at 20°C/min, then a second ramp from140 to 160°C at 5°C/min. Finally, a third ramp was used from160 to 235°C at 2°C/min. This final temperature was main-tained for 3 min. Injector and detector temperatures were 250and 230°C, respectively. The injector was in splitless mode.Helium was used as carrier gas. Mass spectra (MS) were re-corded in the electronic impact (70 eV) mode. The transferline temperature was 250°C with source temperature at 250°C.
Analysis of DMB-labeled side chain CA 14 mM solution of DMB (Sigma-Aldrich, Saint-QuentinFallavier, France) was prepared by dissolving DMB in anaqueous solution of 80 mM β-mercaptoethanol, 40 mM sodi-um hydrosulphite and 2.8 M acetic acid. One milligram of theRG-II fraction was solubilized in 90 μL of deionized water and90 μL of the DMB solution. The mixture was heated at 50°Cfor 2.5 h in the dark. Twenty microliters of the resulting solu-tion was injected in a liquid chromatograph (Kontron, Milan,Italy) equipped with a reverse-phase C18 column (300 ×4.5 mm, C18 monomeric, Vydac, cat. # 238TP54) and SFM-25 fluorescence spectrophotometer (Kontron). Elution of theDMB der ivat ives was performed at a flow rate of0.9 mL min−1 at room temperature using solvent A (acetoni-trile:methanol:water, 4:6:90 [v/v]) and solvent B (acetonitrile:methanol:water, 11:7:82 [v/v]), with an A to B linear gradientfrom 50:50 to 0:100 (v/v) over 40 min. The DMB derivativeswere detected by fluorescence using excitation and emissionwavelengths of 373 and 448 nm, respectively.
Mass spectrometryMALDI-TOF mass spectrum of the disaccharide side chains re-leased by mild acid hydrolysis and coupled to DMB was
626
M Séveno et al.
by guest on February 25, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from

acquired on a Voyager DE-Pro MALDI-TOF instrument (Ap-plied Biosystems, Courtabeuf, France) equipped with a 337 nmnitrogen laser. MS were performed in the reflector delayedextraction mode using 2,5-dihydroxybenzoic acid (Sigma-Aldrich, Saint-Quentin Fallavier, France) as matrix. Thesespectra were recorded in a positive mode using an accelerationvoltage of 20,000 V with a delay time of 100 ns. ESI-MSwas performed on a Q-TRAP mass spectrometer (Applied Bio-systems). Samples in 95% water/5% acetonitrile and 0.02%formic acid (v/v) were infused through Proxeon nanospray cap-illaries (Proxeon Biosystems, Odense, Denmark). The ionsource conditions were adjusted for optimal sample ionization.According to this, infusion needle potential values were adaptedbetween 0.9 and 1.5 kV. Spectra were acquired in a positivemode with a scan rate of 1000 amu s−1 and accumulated untila satisfactory signal to noise ratio had been obtained. Other vol-tages were as recommended by the manufacturer.
Subcellular localization of AtKDTA-GFP proteinThe cDNA of AtKDTA containing the full-length ORF wasPCR amplified from the U17171 clone (from RIKEN cDNAclone RAFL06-11-M15 GenBank accession numberAY065115) with primers 5′-GGGGACAAGTTTGTA-CAAAAAAGCCTTCATGAAGCTCGAGTGTTTG-3′ and5′-GGGGACCACTTTGTACAAGAAAGCTGCGTCTTTG-CAAATGTGATTC-3′. The AtKDTA sequence was theninserted in pGEM-T and sequenced. The cDNA was first in-serted into pDONRTM 207 (Invitrogen). Then, the AtKDTAcDNA was subcloned into pMDC83, which contains theGFP6 cDNA, the dual P35S and the Nos T (ABRC, Ohio StateUniversity, Ohio; Curtis and Grossniklaus 2003) by usingthe Gateway® technology (Invitrogen). The ERD2-CFP con-struct (Saint-Jore et al. 2002), used as a Golgi marker, waskindly given by N. Paris (BPMP, Montpellier, France). TheAtKDTA-GFP expression vector was introduced in the Agro-bacterium strain GV3101 pMP90. Leaves of 5- to 6-week-oldtobacco plants growing at 22°C were infiltrated with trans-formed agrobacteria as described by Brandizzi et al. (2002).
For confocal imaging, the fluorescence emitted by the re-porter proteins was observed 48–72 h after transientexpression. The ERD2-CFP construct used as a Golgi markerwas previously described (Chatre et al. 2005). For mitochon-dria labeling, leaves expressing the AtKDTA-GFP constructwere infiltrated with 100 nM MitoTracker® Orange CMTMRos(Molecular Probes™) in Murashige and Skoog's buffer andplants were then left in the dark for 4 h before observation.For colocalization with chloroplasts in tobacco mesophyllcells, epidermis was removed prior to observation. The confo-cal imaging was carried out on a confocal microscope fromLeica (TCS SP2 AOBS, Wetzlar, Germany) with a ×40 oil im-mersion objective. For imaging the coexpression of ERD2-CFP and AtKDTA-GFP constructs, excitation lines of an argonion laser of 458 nm for CFP and 488 nm for GFP were alter-natively used with line switching mode on the multitracksetting of the microscope. CFP fluorescence was detected be-tween 460 and 485 nm and GFP fluorescence between 509 and602 nm. For imaging both the AtKDTA-GFP fluorescence andMitoTracker® labeling, we used a Zeiss LSM510 Meta laserscanning microscope (Jena, Germany) with a ×63 oil immer-sion objective. To do so, excitation lines of an argon ion
laser of 488 nm for GFP and 543 nm were alternatively used.GFP fluorescence was detected using a 545 nm dichroic beamsplitter and a 505–530 nm bandpass filter, and MitoTracker®
orange fluorescence was detected with a 545 nm dichroic beamsplitter and a 560–615 nm bandpass filter. Image analysis anddeconvolutions were performed with the Huygens RemoteManager (Ponti et al. 2007).
Funding
This research was supported by the Centre National de la Re-cherche Scientifique, the Région Haute-Normandie, the RegionAquitaine and the University of Rouen. F.D. was supported bygrant no. 00-512858 from the Ministère de la Recherche et dela Technologie (France).
Conflict of interest statement
None declared.
Abbreviations
BLAST, basic local alignment search tool; CFP, cyan fluores-cent protein; CMP, cytidine monophosphate; Dha, 3-deoxy-D-lyxo-heptulosonic acid; DMB, 1,2-diamino-4,5-methylenedioxybenzene; EPG, endopolygalacturonase; ESI-MS, electro-spray ionization mass spectrometry; GFP, green fluorescentprotein; Kdo, 3-deoxy-D-manno-octulosonic acid; KDTA,Kdo transferase; LPS, lipopolysaccharide; MALDI-TOF, ma-trix-assisted laser desorption ionization-time of flight; RG-II,rhamnogalacturonan II; RP-HPLC, reverse-phase high-perfor-mance liquid chromatography; RT-PCR, reverse transcriptase-polymerase chain reaction; TFA, trifluoroacetic acid.
Acknowledgements
We thank the cell imaging facilities of Universities de Rouenand Montpellier.
References
Ahn JW, Verma R, Kim M, Lee JY, Kim YK, Bang JW, Reiter WD, Pai HS.2006. Depletion of UDP-D-apiose/UDP-D-xylose synthases results in rham-nogalacturonan-II deficiency, cell wall thickening, and cell death in higherplants. J Biol Chem. 281:13708–13716.
Armstrong MT, Theg SM, Braun N, Wainwright N, Pardy RL, Armstrong PB.2006. Histochemical evidence for lipid A (endotoxin) in eukaryote chloro-plasts. FASEB J. 20:2145–2146.
Anisimova M, Gascuel O. 2006. Approximate likelihood ratio test for branchs:A fast, accurate and powerful alternative. Syst Biol. 55:539–552.
Becker B, Lommerse JPM, Melkonian M, Kamerling JP, Vliegenthart JFG.1995. The structure of an acidic trisaccharide component from a cell wallpolysaccharide preparation of the green alga Tetraselmis striata Butcher.Carbohydr Res. 267:313–321.
Belunis CJ, Clementz T, Carty SM, Raetz CRH. 1995. Inhibition of lipopoly-saccharide biosynthesis and cell growth following inactivation of the KdtAgene in Escherichia coli. J Biol Chem. 270:27646–27652.
Brabetz W, Wolter FP, Brade H. 2000. A cDNA encoding 3-deoxy-D-manno-oct-2-ulosate-8-phosphate synthase of Pisum sativum L. (pea) functionallycomplements a kdsA mutant of the Gram-negative bacterium Salmonella en-terica. Planta. 212:136–143.
Brandizzi F, Frangne N, Marc-Martin S, Hawes C, Neuhaus JM, Paris N. 2002.The destination for single-pass membrane proteins is influenced markedlyby the length of the hydrophobic domain. Plant Cell. 14:1077–1092.
627
Characterization of Arabidopsis Kdo transferase
by guest on February 25, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from

Cantarel BL, Coutinho PM, Rancurel C, Bernard T, Lombard V, Henrissat B.2009. The Carbohydrate-Active EnZymes database (CAZy): An expert re-source for Glycogenomics. Nucleic Acids Res. 37:D233–D238.
Castresana J. 2000. Selection of conserved blocks from multiple alignments fortheir use in phylogenetic analysis. Mol Biol Evol. 17:540–552.
Chatre L, Brandizzi F, Hocquellet A, Hawes C, Moreau P. 2005. Sec22and memb11 are v-SNAREs of the anterograde endoplasmic reticu-lum-Golgi pathway in tobacco leaf epidermal cells. Plant Physiol.139:1244–1254.
Chevenet F, Brun C, Banuls AL, Jacq B, Chisten R. 2006. TreeDyn: Towardsdynamic graphics and annotations for analyses of trees. BMC Bioinfor-matics. 10:439.
Clementz T, Raetz CR. 1991. Agene coding for 3-deoxy-D-manno-octulosonic-acid transferase in Escherichia coli. Identification, mapping, cloning, and se-quencing. J Biol Chem. 266:9687–9696.
Corpet F. 1988. Multiple sequence alignment with hierarchical clustering.Nucleic Acids Res. 16:10881–10890.
Curtis MD, Grossniklaus U. 2003. A gateway cloning vector set for high-throughput functional analysis of genes in planta.PlantPhysiol.133:462–469.
Daskalova SM, Pah AR, Baluch DP, Lopez LC. 2009. The Arabidopsis thali-ana putative sialyltransferase resides in the Golgi apparatus but lacks theability to transfer sialic acid. Plant Biol. 11:284–299.
Delmas F, Petit J, Joubès J, Séveno M, Paccalet T, Hernould M, Lerouge P,Mouras M, Chevalier C. 2003. The gene expression and enzyme activityof plant 3-deoxy-D-manno-2-octulosonic acid-8-phosphate synthase arepreferentially associated with cell division in a cell cycle-dependent manner.Plant Physiol. 133:348–360.
Delmas F, Séveno M, Northey JGB, Hernould M, Lerouge P, Mc Court P, Che-valier C. 2008. The synthesis of the rhamnogalacturonan II component 3-deoxy-D-manno-2-octulosonic acid (Kdo) is required for pollen tube growthand elongation. J Exp Bot. 59:2639–2647.
Dereeper A, Guignon V, Blanc G, Audic S, Buffet S, Chevenet F, DufayardJF, Guindon S, Lefort V, Lescot M, et al. 2008. Phylogeny.fr: Robustphylogenetic analysis for the non-specialist. Nucleic Acids Res. 36:W465–W469.
Dunkley TPJ, Hester S, Shadforth IP, Runions J, Weimar T, Hanton SL, GriffinJL, Bessant C, Brandizzi F, Hawes C, et al. 2006. Mapping the Arabidopsisorganelle proteome. Proc Natl Acad Sci USA. 103:6518–6523.
Edwards K, Johnstone C, Thompson C. 1991. A simple and rapid method forthe preparation of plant genomic DNA for PCR analysis. Nucleic Acids Res.19:1349.
Ekpo P, Nano FE. 2006. Arg276 of GseA, a Chlamydiatrachomatis Kdo trans-ferase, is required for the synthesis of the chlamydial genus-specific epitopein Escherichia coli. FEMS Microbiol Lett. 96:49–53.
Emanuelsson O, Brunak S, von Heijne G, Nielsen H. 2007. Locating pro-teins in the cell using TargetP, SignalP, and related tools. Nat Protoc.2:953–971.
Finn RD, Tate J, Mistry J, Coggill PC, Sammut JS, Hotz HR, Ceric G, ForslundK, Eddy SR, Sonnhammer EL, et al. 2008. The Pfam protein families data-base. Nucleic Acids Res. 36:D281–D288.
Fleisher A, O'Neill MA, Ehwald R. 1999. The pore size of non-graminac-eous plant cell walls is rapidly decreased by borate ester cross-linkingof the pectic polysaccharide rhamnogalacturnan II. Plant Physiol.121:829–838.
Gouet P, Robert X, Courcelle E. 2003. ESPript/ENDscript: Extracting and ren-dering sequence and 3D information from atomic structures of proteins.Nucleic Acids Res. 31:3320–3323.
Guindon S, Gascuel O. 2003. A simple, fast, and accurate algorithm to estimatelarge phylogenies by maximum likelihood. Syst Biol. 52:696–704.
Ishii T, Matsunaga T, Pellerin P, O'Neill MA, Darvill AG, Albersheim P. 1999.The plant cell wall polysaccharide rhamnogalacturonan II self-assembles in-to a covalently cross-linked dimer. J Biol Chem. 274:13098–13104.
Ishii T, Matsunaga T, Hayashi N. 2001. Formation of rhamnogalacturonan II-borate dimer in pectin determines cell wall thickness of pumpkin tissue.Plant Physiol. 126:1698–1705.
Jelakovic S, Schulz GE. 2002. Catalytic mechanism of CMP:2-keto-3-deoxy-manno-octonic acid synthetase as derived from complexes with reactioneduct and product. Biochemistry. 41:1174–1181.
Johnson MA, von Besser K, Zhou Q, Smith E, Aux G, Patton D, Levin JZ,Preuss D. 2004. Arabidopsis hapless mutations define essential gametophyt-ic functions. Genetics. 168:971–982.
Kobayashi M, Matoh H, Azuma J. 1996. Two chains of rhamnogalacturonan IIare cross-linked by borate-diol ester bonds in higher plant cell walls. PlantPhysiol. 110:1017–1020.
Lairson LL, Henrissat B, Davies GJ, Withers SG. 2008. Glycosyltransferases:Structures, functions, and mechanisms. Annu Rev Biochem. 77:521–555.
Matsuura K, Miyagawa I, Kobayashi M, Ohta D, Matoh T. 2003. Arabidopsis3-deoxy-D-manno-oct-2-ulosonate-8-phosphate synthase: cDNA cloningand expression analyses. J Exp Bot. 54:1785–1787.
Meredith TC, Woodward RW. 2006. Characterization of Escherichia coli D-arabinose 5-phosphate isomerase encoded by KpsF: Implication for group2 capsule biosynthesis. Biochem J. 395:427–432.
Münster AK, Eckhardt M, Potvin B, Mühlenhoff M, Stanley P, Gerardy-Schahn R. 1998. Mammalian cytidine 5'-monophosphate N-acetylneurami-nic acid synthetase: A nuclear protein with evolutionarily conservedstructural motifs. Proc Natl Acad Sci USA. 95:9140–9145.
Murashige T, Skoog F. 1962. A revised medium for rapid growth and bio-as-says with tobacco tissue culture. Physiol Plant. 15:473–497.
O'Neill MA, Warrenfeltz D, Kates K, Pellerin P, Doco T, Darvill AG, Alber-sheim P. 1996. Rhamnogalacturonan-II, a pectic polysaccharide in the wallsof growing plant cell, forms a dimer that is covalently cross-linked by aborate ester. J Biol Chem. 271:22923–22930.
O'Neill MA, Eberhard S, Albersheim P, Darvill AG. 2001. Requirement boratecross-linking of the cell wall rhamnogalacturonan II for Arabidopsis growth.Science. 294:846–849.
O'Neill MA, Ishii T, Albersheim P, Darvill AG. 2004. Rhamnogalacturonan II:Structure and function of a borate cross-linked cell wall pectic polysaccha-ride. Annu Rev Plant Biol. 55:109–139.
Pérez S, Mazeau K, Hervé du Penhoat C. 2000. The three-dimensional struc-tures of the pectic polysaccharides. Plant Physiol Biochem. 38:37–55.
Pérez S, Rodriguez-Carvajal MA, Doco T. 2003. A complex plant cell wallpolysaccharide: Rhamnogalacturonan II. A structure in quest of function.Biochimie. 85:109–121.
Pieper U, Eswar N, Webb BM, Eramian D, Kelly L, Barkan DT, Carter H,Mankoo P, Karchin R, Marti-Renom MA, et al. 2009. MODBASE, a data-base of annotated comparative protein structure models and associatedresources. Nucleic Acids Res. 37:D347–D354.
Ponti A, Gulati A, Bäcker V, Scwarb P. 2007. Huygens remote manager: Awebinterface for high-volume batch deconvolution. Imaging Microsc. 9:57–58.
Raetz CRH. 1990. Biochemistry of endotoxins.AnnuRev Biochem. 59:129–170.Raetz RH, Whitfield C. 2002. Lipolysaccharide endotoxins. AnnuRevBiochem.
71:635–700.Rick P. 1987. Lipopolysaccharide biosynthesis. In: Neidhardt FC, editor. Es-
cherichia coli and Salmonella typhimurium: Cellular and MolecularBiology. Washington: ASM.
Royo J, Gomez E, Hueros G. 2000. A maize homologue of the bacterial CMP-Kdo synthetases: Similar pathways operate in plants and bacteria for the ac-tivation of Kdo prior to its incorporation into cellular envelopes. J BiolChem. 275:24993–24999.
Saint-Jore CM, Evins J, Brandizzi F, Batoko H, Moore I, Hawes C. 2002. Re-distribution of membrane proteins between the Golgi apparatus andendoplasmic reticulum in plants is reversible and not dependent on cytoskel-etal networks. Plant J. 29:661–678.
Séveno M, Bardor M, Paccalet T, Gomord V, Lerouge P, Faye L. 2004. Gly-coprotein sialylation in plants? Nat Biotechnol. 22:5–6.
Séveno M, Voxeur A, Rihouey C, Wu A-M, Ishii T, Chevalier C, Ralet M-C,Driouich A, Marchant A, Lerouge P. 2009. Structural characterisation of thepectic polysaccharide rhamnogalacturonan II using an acidic fingerprintingmethodology. Planta. 230:947–957.
Wu J, Patel MA, Sundaram AK, Woodard RW. 2004. Functional and biochem-ical characterization of a recombinant Arabidopsis thaliana 3-deoxy-D-manno-octulosonate 8-phosphate synthase. Biochem J. 381:185–193.
York WS, Darvill AG, McNeil M, Albersheim P. 1985. 3-desoxy-D-manno-2-octulosonic acid (KDO) is a component of rhamnogalacturonan II, a pecticpolysaccharide in the primary cell walls of plants. Carbohydr Res.138:109–126.
628
M Séveno et al.
by guest on February 25, 2016http://glycob.oxfordjournals.org/
Dow
nloaded from
Copyright © 2022 FDOKUMEN







![Molecular Imaging of Murine Intestinal Inflammation With 2-Deoxy-2-[ 18F]Fluoro- d-Glucose and Positron Emission Tomography](https://static.fdokumen.com/doc/165x107/6344fff26cfb3d4064097a1a/molecular-imaging-of-murine-intestinal-inflammation-with-2-deoxy-2-18ffluoro-.jpg)












